
Abstract
Arbuscular mycorrhizal fungi (AMF) are ubiquitous, soil-borne, endophytic, obligate biotrophs that colonize the roots of most terrestrial plants to establish mutually beneficial relationships. The colonization of host roots by AMF improves the access of plant roots to nutrients, mainly phosphorus (P) and nitrogen (N). In return, host plants provide the colonizing fungus with carbon (C)-rich photosynthates such as sugar and lipids. This nutrient exchange strongly impacts plant and microbial ecosystems by influencing plant fitness, core soil processes, and the C cycle. Crucially, both mutualism and ecosystem services largely rely on the bidirectional flow of resources. The AMF symbiosis not only provides plants with nutrients but also supplies water and enhances their resistance to environmental and biotic challenges. The physiological and molecular processes by which AMF aids plants in nutrient foraging and stress resilience have been extensively studied. Recent work has established the framework for nutrient exchange and regulatory interactions between host plants and AMF, but the mechanisms underlying AMF-mediated host plant tolerance to stress still remain unclear. AMF are of potential applications in crop production, forestry management, bioremediation, and ecological restoration as biofertilizers and bioprotectors. In the first chapter, we introduce AMF, plant-AMF symbioses and their characteristic features, functions, and potential applications.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1.1 Introduction
Arbuscular mycorrhizal fungi (AMF) are ubiquitous, soil-borne, endophytic, obligate biotrophs that colonize the roots of 70–90% of terrestrial plants in various soil types and environmental conditions to establish mutually beneficial relationships (Branco et al. 2022; Shi et al. 2023). The evolutionary trajectory of AMF has been intrinsically tied to the history of plant evolution with the appearance of the first terrestrial plant around 400 MYA ago (Salvioli di Fossalunga and Novero 2019). Taxonomically, AMF belong to the phylum Glomeromycota (Giovannini et al. 2020). The name AMF signifies the peculiar relationship between plant roots and mycorrhizal fungi and its key features, wherein “Arbuscule” stands for a special treelike fungal (hyphal) structure in the inner cortex of the root that is used for resource-exchange between “plant root” and “AMF,” whereas “mycorrhiza” derived from two words “myco,” meaning fungus, and “rhiza,” meaning “root” (Shi et al. 2023).
In plant-AMF symbiosis, AMF benefit from the plant-supplied carbon (C) substances such as sugars and lipids derived from photosynthesis (Jiang et al. 2017), while the AMF provide the plant host with mineral nutrients, particularly phosphorus (P) and nitrogen (N) (Wipf et al. 2019). This nutrient exchange strongly impacts plant and microbial ecosystems by influencing plant fitness, core soil processes, and the C cycle (Diagne et al. 2020; Giovannini et al. 2020). AMF assist the roots of the host plant with their extensive hyphal network in absorbing water and nutrients from faraway soils, thus decreasing the requirement for chemical fertilizers and irrigation (Kakouridis et al. 2022). AMF are found in different kinds of environments including severely disturbed ecosystems containing soils polluted with excessive salt, xenobiotics, and heavy metals (Boorboori and Zhang 2022; Branco et al. 2022; Ahammed et al. 2023).
The symbiosis between plant roots and AMF hyphae results in the recruitment of specialized microorganisms into the mycorrhizosphere (Giovannini et al. 2020; Phour et al. 2020). AMF stimulate the mycorrhizosphere microbiome and help plants develop, take up nutrients, and resist abiotic and biotic hazards (Philippot et al. 2013; Zhang et al. 2022). By facilitating nutrient uptake, AMF aid plant development and stress tolerance including tolerance to drought stress (Begum et al. 2019), salinity (Cui et al. 2022; Li et al. 2022), nutrient deficiency (Shi et al. 2021), or heavy metal stress (Dhalaria et al. 2020; Alam et al. 2019). The physiological, biochemical, microbial, and molecular processes by which AMF aid plants in nutrient foraging and stress resilience have been extensively studied (Salvioli di Fossalunga and Novero 2019). Recent work has established the framework for nutrient exchange and regulatory interactions between host plants and AMF, but mechanisms underlying AMF-mediated host plant tolerance to stress still remain unclear (Shi et al. 2023). AMF have potential applications in crop production, forestry management, bioremediation, and ecological restoration as biofertilizers and bioprotectors (Phour et al. 2020; Boorboori and Zhang 2022; Zhu et al. 2022). In this chapter, we provide a brief introduction to AMF, their characteristic features, functions, and potential applications in crop production and environmental management.
1.2 Arbuscular Mycorrhizal Symbiosis in Higher Plants: Characteristic Features
An estimated 250,000 plant species develop mycorrhizal symbiosis with around 50,000 fungal species of four major mycorrhizal types, such as arbuscular mycorrhizal, ectomycorrhizal, ericoid mycorrhizal, and orchid mycorrhizal fungi (van der Heijden et al. 2015). About 70–90% of land plant species develop arbuscular mycorrhizal symbiosis (AMS) with the obligate symbiotic fungi of the phylum Glomeromycota (Shi et al. 2023). This includes the vast majority of herbs, grasses, and trees, which act as hosts of AMF in AMS. Importantly, AMF establish symbiotic relationships with around 90% of agricultural plants, encompassing a wide variety of agricultural crops such as cereals, vegetables, and horticultural plants (Diagne et al. 2020). Non-mycorrhizal plants with regard to food crops are limited to genera and species that fall under the families Brassicaceae and Chenopodiaceae (Giovannini et al. 2020).
Although species of host plants are found across the phylogeny of terrestrial plants (van der Heijden et al. 2015), only approximately 345 species of AMF from the phylum Glomeromycota were documented on the basis of DNA barcoding (http://www.amf-phylogeny.com/, accessed on 30 June 2023), indicating that AMF are distinguished by limited diversity in species and largely lacking host specificity as opposed to a huge diversity of host plants (Shi et al. 2023). However, a substantial degree of variation exists within the AMF species itself (intraspecific diversity). The significant intraspecific variation among AMF may lead to considerable functional differences such as extraradical hyphal expansion, spore formation, root colonization, nutrient and water fluxes, as well as varying mycorrhizal growth responses. It was proposed that the concurrent evolution of plant and fungal communities might lead to a high level of genetic diversity among AMF isolates (Wipf et al. 2019).
Despite the considerable diversity observed in AMF, limited research has been conducted on their functional diversity, aiming to identify and select the most suitable isolates for agricultural applications (Edlinger et al. 2022; Powell and Rillig 2018). The majority of commercially available inocula are formulated using Rhizoglomus irregulare (previously named Rhizophagus irregularis, previously classified as Glomus intraradices) and Funneliformis mosseae (previously named Glomus mosseae), which are common symbiotic organisms with a broad range of host plants and are extensively distributed in various soil types and climatic regions worldwide (Giovannini et al. 2020).
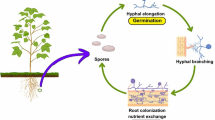
Arbuscular mycorrhizal fungi live in dual habits, one is the root, from where they obtain organic C and transfer nutrients, and the other is the soil from where they acquire nutrients, mainly N and P (van der Heijden et al. 2015; Yang et al. 2022; Shi et al. 2021). AMF primarily forage for nutrients derived from plants and saprotrophic microorganisms (Shi et al. 2023). While soil fungal communities are very abundant, AMF make up just a tiny fraction of their abundance (<0.5%) (Fierer 2017). The initiation of AMS involves a communication process between host plants and AMF during the pre-contact phase (López-Ráez and Pozo 2013; Zhou et al. 2019). This interaction progresses to the infection phase and ultimately leads to the development of fully formed arbuscule within the inner root cortical cells of the host plant, facilitating nutrient exchange. Before the development of appressoria, the roots of host plants release strigolactones (SLs, a group of plant hormones commonly known for controlling bud outgrowth) into the rhizosphere in order to induce the germination of fungal spores and hyphal branching of AMF, thereby facilitating the establishment of physical contact between AMF and roots (Zhou et al. 2019). Notably, SL biosynthesis elevates under P-deprived conditions. The regulation of P uptake in host plants is governed by transcription factors involved in the phosphate starvation response (Ho-Plágaro and García-Garrido 2022). In particular, a group of the MYB-CC family of transcription factors, phosphate starvation responses (PHRs), serve as key regulators of AMS (Shi et al. 2021; Das et al. 2022). This implies that the primary driving force behind the establishment of AMS is the host plants’ need for phosphate (Wang et al. 2022). AMF release specific substances known as mycorrhizal factors, which include lipo-chitooligosaccharides (Myc-LCOs) as well as short-chain chitin oligomers CO4 and CO5 (Maillet et al. 2011; Phour et al. 2020). These substances serve as signals that are detected by host plants (Genre et al. 2013). Following the mutual exchange of respective signals between AMF and host plants, the hyphae of AMF exhibit growth toward the roots of the host plant, utilizing the SL gradients as a navigational cue within the rhizosphere (Kretzschmar et al. 2012). Upon contact with the epidermis of root cells, the hyphae proceed to extensively branch out, resulting in the formation of a structure known as an appressorium, which is also commonly referred to as a hyphopodium (Fig. 1.1). Subsequently, the hyphae of AMF originating from the appressoria proceed to infiltrate the epidermis of the host plant root and the cells of the cortex. The hyphal infiltration is facilitated by the pre-penetration apparatus (PPA), a complex consisting of cytoskeletal and endoplasmic reticulum components, which guides the migration of the hyphae in a manner directed by the nucleus (Genre et al. 2013; Bonfante and Genre 2010). After traversing the epidermis and outer root cortical cells, the AMF hyphae proceed to expand intercellularly along the longitudinal direction of the root and subsequently infiltrate the inner cortex (Shi et al. 2023). The hyphae of AMF undergo multiple instances of dichotomous branching, leading to the development of arbuscules, which are treelike structures found within the root cells of the cortex (Bonfante and Genre 2010). Arbuscules are enveloped by a layer of plant membranes known as the periarbuscular membrane (PAM). Arbuscules serve as major sites of resource exchange between the AMF and host plants and are the functional hub of such mutually beneficial relationships (Salvioli di Fossalunga and Novero 2019). The arbuscules exhibit a remarkable degree of dynamism, characterized by a relatively short life span lasting only a few days prior to their eventual collapse (Giovannini et al. 2020). The regulation of arbuscule degeneration is affected by both the AMF and host plants, indicating the existence of molecular communication between the cortex and AMF hyphae prior to the degeneration of arbuscules over time (Shi et al. 2023).

Plant root colonization by arbuscular mycorrhizal fungi (AMF). Created with BioRender.com.
The life cycle of AMF is dependent on the existence of host plants due to their obligate biotrophic nature (Ahammed et al. 2023). It commences with an initial phase characterized by the absence of symbiotic interactions. During this phase, spores undergo germination in response to various physical stimuli, including the availability of moisture, optimal temperature, and pH. This germination process leads to the formation of hyphae, which have a short life span (Giovannini et al. 2020). Subsequently, a sequence of events occurs, commencing with the physical interaction between the AMF hyphae and the root. This interaction as mentioned above leads to the penetration of the root cells by the AMF hyphae, ultimately resulting in the formation of arbuscules. This marks the initiation of the symbiotic phase. There are two distinct forms of root colonization: (1) Arum type and (2) Paris type (Tominaga et al. 2022). In the Arum type, the AMF pass on intercellularly through the cells of the cortex, resulting in the development of terminal arbuscules on intracellular hyphal branches (Smith and Smith 1997). The Paris type of root colonization is characterized by the direct growth of the fungus from one root cell to another root cell within the cortex. This growth pattern gives rise to intracellular hyphal coils and intercalary arbuscules (i.e., the arbusculate coil), which are distributed along the coils (Tominaga et al. 2019). Nevertheless, the majority of the available data concerning AMF come from studies conducted on the Arum-type root colonization, which is extensively found in both agricultural and natural ecosystems (Smith and Smith 1997). In addition to the formation of arbuscules, numerous species of AMF exhibit the ability to generate intraradical vesicles. The vesicles are specialized storage structures resembling fungal spores and containing lipid reserves (Giovannini et al. 2020). Upon receiving C from the host plant root, the fungal symbiont can grow outside of the root system, establishing itself in the surrounding soil. This colonization enables the absorption of mineral nutrients, which can then be delivered to the host plant. Additionally, AMF interact with microbes present in the rhizosphere and soil and are capable of colonizing the roots of neighboring plants, even those belonging to different species, genera, and families than their original host plants (Zhang et al. 2022). Furthermore, AMF are capable of transferring nutrients from one host plant to another (van der Heijden et al. 2015). The life cycle of AMF is completed through the production of asexual spores by the extraradical mycelium, which plays a key role in sustaining a high level of mycorrhizal potential in the soil (Giovannini et al. 2020). This, in turn, contributes to the overall biological fertility of the soil.
The existing body of literature suggests that plants have the ability to regulate the extent of root colonization by AMF based on their needs for specific nutrients particularly P and N (Shi et al. 2021). It is widely postulated that the nutrients transported to the cortical cells of the root are responsible for initiating a signaling mechanism that regulates the release of C-rich photoassimilates such as sugars to the partner AMF (López-Ráez and Pozo 2013). The underlying reasons for employing this particular approach rely on the premise that the symbiotic fungal partner (AMF) that is incapable of providing substantial quantities of essential nutrients (e.g., Pi and N) would only be able to obtain limited amounts of C from the apoplast of roots (Wipf et al. 2019).
Plants colonized with AMF (AM plants) employ two distinct pathways for nutrient acquisition, namely, the direct pathway and the mycorrhizal pathway (Salvioli di Fossalunga and Novero 2019). In the first pathway, nutrient uptake takes place directly through the epidermis of root cells and root hairs, while in the latter pathway, nutrient acquisition is facilitated by the AMF hyphae, which enable the transport of nutrients that are acquired from the surrounding soil to the root cells (Ma et al. 2021; van der Heijden et al. 2015). The choice of AM plants between the direct pathway and mycorrhizal pathway is contingent upon the level of phosphorus availability in the rhizosphere (Zhang et al. 2021b). The hyphae of AMF exhibit the ability to extend their growth beyond the zone of phosphorus (Pi, inorganic P) depletion, thereby enabling them to access Pi resources that are otherwise unavailable to plant roots (Fig. 1.2).

Mycorrhizal pathways of nutrient acquisition and resource exchange in arbuscules. Created with BioRender.com.
1.3 Functions of AMS in Plant Growth, Development, and Stress Tolerance
1.3.1 Plant Growth, Development, and Nutrient Acquisition
The growth and reproductive development of plants were found to be significantly influenced by the nutrient availability in the soil and the presence of AMF (Wang et al. 2022). The plant root colonization by AMF enhances the photosynthetic rate and elevates the levels of chlorophyll concentrations and biomass accumulation of the partner plants (Chen et al. 2017b). A recent study found that AMS had a direct positive impact on the growth and nutrient content of cherry tomato plants (Wang et al. 2022). The study confirmed that AMS played a role in promoting flower and seed formation in cherry tomato plants, and the positive outcomes of AMS are thought to be primarily due to the improved nutrient status of the host plants. For instance, phosphorus plays a vital role in the growth, development, and survival of plants; however, its availability to plants is limited in common soil types (Zhang et al. 2021b). The presence of AMS has been shown to enhance the efficiency of phosphate uptake by the host plant (Liao et al. 2022; Ma et al. 2021). Plants allocate approximately 10–20% of their photosynthetic carbon to AMF. In reciprocation, AMF are known to provide ~90% of the phosphorus required by plants (van der Heijden et al. 2015). AMF possess the ability to enter nutrient-rich areas of soil located beyond the rhizosphere through their broad extraradical hyphal network, thereby enabling them to acquire P that would otherwise be inaccessible to plants. It is worth mentioning that extraradical hyphae are capable of releasing low-molecular-weight organic acids, which have the ability to dissolve phosphorus from sources that are typically not accessible to biological processes (Andrino et al. 2021). AMF acquire phosphate (Pi) from the immediate surroundings in soil by means of phosphate transporters (PHT1), which are specifically located on the extraradical hyphae (Wipf et al. 2019).
AMF can potentially contribute around 33% of the root protein nitrogen (Govindarajulu et al. 2005). The uptake of nitrogen is facilitated by a range of transport systems, which include the transport of inorganic nitrogen in the forms of nitrate (NO3–) and ammonium (NH4+), as well as the transport of organic nitrogen in the forms of peptides and amino acids (Wipf et al. 2019). AMF primarily acquire inorganic N from the soil by utilizing ammonium transporters (AMTs), and they exhibit a preference for NH4+ over NO3– as a nitrogen source (Hui et al. 2022). This preference arises from the fact that NO3– is to be converted to NH4+ before it can be assimilated into organic compounds, a process that requires energy expenditure (Shi et al. 2023). Nitrogen transport through the mycorrhizal pathway is significantly impacted by AMTs (Breuillin-Sessoms et al. 2015). Nevertheless, the mutualistic transport of NH4+ may not be a necessary prerequisite for the formation of AMS or the development of arbuscules. Furthermore, it has been observed that apart from NH4+, there is a symbiotic pathway for NO3– uptake. Nitrate is acquired through an uptake mechanism that relies on energy expenditure and is facilitated by specialized transporters belonging to the large family of nitrate and peptide transporters, namely, NPF (NRT1/PTR), NRT2, and NRT3 families (Wipf et al. 2019).
1.3.2 Abiotic and Biotic Stress Tolerance
Plants in natural environments face a multitude of challenges, encompassing both biotic factors, such as pathogenic fungi, bacteria, nematodes, and insect pests, and abiotic factors, including salinity, drought, and toxic metal contamination (Sanchez-Bermudez et al. 2022; Li and Ahammed 2023). Crucially, AMF have the potential to significantly augment the resistance of the plant partner to various forms of stress (Fig. 1.3). AMS has been shown to increase the acquisition of mineral nutrients in plants in both non-stressful and stressful environments (Chen et al. 2017b). However, the impact of AMS on nutrient uptake is particularly significant under conditions of stress (Diagne et al. 2020). Moreover, plant roots colonized by AMF have the ability to absorb more water from the rhizosphere, and AMF have the potential to ameliorate drought stress (Begum et al. 2019; Moradtalab et al. 2019; Puschel et al. 2020). It was found that the water transported by AMF Rhizophagus intraradices through the air gap constituted approximately 34.6% of the total water transpired by the host plant Avena barbata, commonly known as wild oat (Kakouridis et al. 2022). This study presents empirical evidence demonstrating that AMF can function as extensions of the root system within the water movement continuum encompassing the soil, plant, and air. The enhanced uptake of mineral nutrients and increased water content in AM plants can potentially be associated with the indirect effect of AMF in mitigating stress (Hajiboland et al. 2019; Puschel et al. 2020; Diagne et al. 2020).

Effects of plant root colonization by arbuscular mycorrhizal fungi (AMF) on plant growth, development, and stress tolerance. Created with BioRender.com.
The colonization of plant roots by AMF can alleviate the detrimental effects associated with drought and salinity through enhancements in nutrient absorption, reduction of oxidative stress, augmentation of osmotic adjustment, and alterations in stress responses (Shi et al. 2023). AMF colonization assists host plants in maintaining appropriate ion homeostasis when subjected to salinity stress (Chen et al. 2017a; Porcel et al. 2016). Studies also indicate that the presence of AMS has a beneficial impact on host plants subjected to flooding stress (Diagne et al. 2020). AMF have a positive impact on plant performance in conditions of heavy metal stress (Boorboori and Zhang 2022).
In plants, abiotic stress often coincides with oxidative stress, as a result of the excessive production of reactive oxygen species (ROS) causing elevated levels of ion leakage and lipid peroxidation (Sanchez-Bermudez et al. 2022). The colonization of AMF has been observed to mitigate oxidative stress in AM plants (Diagne et al. 2020). This is achieved through the upregulation in the activity and transcripts of antioxidant enzymes, such as superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and glutathione reductase (GR) in AM plants (Zhu et al. 2022). The modulation of enzyme activity leads to a reduction in the accumulation of ROS and a decrease in oxidative damage to lipids and other biomolecules. In addition, the colonization of AMF has been found to have a significant impact on the accumulation of osmoregulatory substances such as soluble sugars and free proline in AM plants when subject to stress (Yooyongwech et al. 2016; Pasbani et al. 2020).
Symbiotic associations with AMF provide beneficial effects to host plants in mitigating biotic stress as well, particularly in the context of pathogen infections (Zhang et al. 2021a). The beneficial impacts of AMS in mitigating pathogen infections can be ascribed to several factors, including enhancements in plant nutrient availability, alterations in root exudate compositions, influences on the rhizosphere microbiome, improvement of plant resistance (both local and systemic resistance), and priming of defense responses (Wu et al. 2021). Enhanced resistance of AM plants is achieved through the direct activation of genes (pathogenesis-related protein PR1) that are involved in the host’s defense responses against pathogen infections (Fiorilli et al. 2018). After being infected by pathogens, the roots that are colonized by AMF exhibit the deposition of callose in the spaces between cells (Sanmartin et al. 2020). Additionally, they form structures resembling papillae at the locations where fungal pathogens penetrate, effectively limiting the growth of pathogens both within and between cells (Shi et al. 2023). The pre-inoculation of tomato plants with AMF leads to a significantly enhanced defense response compared to control plants that are not colonized by AMF when infected with Alternaria solani (Song et al. 2015). AMF can assist host plants in mounting a systemic resistance against pathogens (Fiorilli et al. 2018).
1.3.3 Soil Fertility, Nutrient Cycling, and Ecosystem Services
The AMF-plant symbiosis is of great significance in the carbon cycle of soil ecosystems (Fierer 2017; van der Heijden et al. 2015). Soil organic matter (SOM) represents the most substantial C reservoir on land, and its composition is influenced by many factors such as plant biomass, primary productivity, and the transfer of photosynthetic C to underground biomass (Phour et al. 2020). The primary role played by AMF in soil is to serve as a carbon sink for plant-derived carbon, which is subsequently allocated to AMF hyphae and other microorganisms present in the soil (Giovannini et al. 2020; Ossowicki et al. 2021; Hermans et al. 2023). AMF and the diverse soil microbial communities play a decisive role in the release or intake of carbon from the soil through processes such as respiration, decomposition, mineralization, and immobilization (Parihar et al. 2020; Philippot et al. 2013). Consequently, the absorption of nutrients by AMF and subsequent transfer to the host plant can enhance the assimilation of C by plants, resulting in a greater supply of C to mycorrhizal fungi. This establishes a positive feedback loop involving host plants, AMF, and carbon. A substantial portion of this carbon is subsequently deposited in the soil in the form of glomalin-related soil protein (GRSP) and fungal chitin (Genre et al. 2013). Glomalin is a glycoprotein synthesized by AMF that exhibits a significant degree of homology with heat shock protein 60 (Gao et al. 2019). There exists a strong positive correlation between GRSP and soil aggregate stability, as GRSP represents an important constituent of soil organic matter. In fact, GRSP alone constitutes approximately 30–40% of the total organic carbon present in undisturbed soils (Parihar et al. 2020). Thus, AMF-released glomalin exhibits potential functionality within soil ecosystems, encompassing the facilitation of soil organic carbon storage, enhancement of soil aggregate structure, augmentation of plant resistance, and mitigation of plant metal toxicity (Riaz et al. 2021; Ahammed et al. 2023; Gao et al. 2019). Moreover, it should be noted that the extraradical hyphae of AMF make up a significant amount of the microbial biomass in the soil. These hyphae account for approximately 54–900 kg ha-1 of SOM (Parihar et al. 2020). Additionally, the presence of extraradical mycelia and GRSP is crucial in the development of soil aggregates, which are crucial in safeguarding organic carbon against decomposition (Parihar et al. 2020).
In addition to mycorrhizae, various fungi and bacteria residing in the rhizosphere of host plants are drawn to and consume the rhizo-deposits, thereby enhancing plant growth, nutrient absorption, and stress resilience (Zhang et al. 2022; Philippot et al. 2013; Hermans et al. 2023). Bacteria that contribute positively to the ecological health, establishment, and functions of AMS are referred to as mycorrhizae helper bacteria (MHBs) (Sangwan and Prasanna 2022). MHBs exert their promotional effects through the production of volatile compounds that stimulate growth and hydrolytic enzymes. Additionally, MHBs enhance plant nutrient uptake by AMF (Sangwan and Prasanna 2022). Previous studies have reported that MHBs exhibit an affinity for the external surface of mycorrhizal fungi, thereby augmenting mycorrhizal symbiotic interactions. This is exemplified by their ability to enhance nutrient uptake (Sangwan and Prasanna 2022). Plant growth-promoting rhizobacteria (PGBs) are currently recognized as MHBs, exhibiting promising prospects for practical implementation in the fields of agriculture and forestry (Giovannini et al. 2020; Shi et al. 2023).
1.4 Applications of AMF in Crop Production, Crop Protection, and Bioremediation
Despite the incomplete understanding of the mechanistic underpinnings of chemical signaling and regulation in the responses of plants during the AMS, it is of great importance to exploit this signaling process for the purposes of enhancing crop protection and increasing yields (López-Ráez and Pozo 2013; Ho-Plágaro and García-Garrido 2022). The mutualistic association between AMF and plants has the potential to significantly enhance crop productivity and promote the long-term stability of ecosystems (Diagne et al. 2020). AMF not only exhibit nutritional benefits, such as enhanced water and nutrient absorption, but also possess non-nutritional advantages, including soil aggregate stabilization, erosion prevention, and mitigation of biotic and abiotic stressors (Parihar et al. 2020). Given the ongoing elucidation of the nature and chemistry of the major molecules implicated in the establishment of AMS in the subterranean environment, there exists significant potential for the manipulation of their production to enhance AMS and its associated advantages, thereby offering promising prospects for the advancement of sustainable agriculture (Phour et al. 2020).
Notably, plant tolerance to heavy metals such as chromium (Cr) is improved by the direct influence of AMF on the transformation and stabilization of Cr, as well as the symbiotic relationship between AMF and plants, which indirectly enhances nutrient acquisition and physiological regulation in plants (Ahammed et al. 2023; Riaz et al. 2021). Studies provide clear evidence that AMF have the potential to not only enhance crop production in heavy metal-contaminated soils but also improve the effectiveness of phytoremediation (Boorboori and Zhang 2022). For AMF-assisted phytoremediation, it is imperative to explore AMF isolates that exhibit a notable capacity for tolerating heavy metals, as well as plant species that demonstrate both an increased growth rate and the ability to accumulate substantial biomass (Dhalaria et al. 2020; Ahammed et al. 2023).
AMF have been employed as biofertilizers to augment plant growth and yields in agricultural practices, albeit their utilization remains relatively limited in comparison to conventional approaches (Gelvez-Pardo et al. 2023). The positive and beneficial effects of the symbiotic relationship on the plant host are contingent upon the specific combination of plant and AMF (Chen et al. 2017b). Hence, the crucial aspect lies in the careful selection of suitable AMF isolates that are well adapted to the specific local conditions and possess optimal functional characteristics for the particular plant species under consideration (Gianinazzi et al. 2010; Giovannini et al. 2020). Gaining a comprehensive understanding of the mechanisms that govern colonization, symbiosis efficiency, and competence is of utmost importance in order to maximize the benefits derived from mycorrhizal associations (Wipf et al. 2019). The comprehensive understanding of the molecules implicated in the interaction between host plants and AMF will undoubtedly enhance the effective application of “mycorrhizal technology” in agricultural systems (Gelvez-Pardo et al. 2023). There exists a debate regarding the suitability of AMF in agricultural systems, primarily stemming from the potential counteractive impact of excessive chemical fertilizer application, a prevalent practice in numerous countries, on the beneficial outcomes of AMF on crop growth (Giovannini et al. 2020). Nevertheless, AMF do possess pivotal roles in augmenting the efficacy and durability of agroecosystems (Gianinazzi et al. 2010; Piliarová et al. 2019; Powell and Rillig 2018).
1.5 Conclusions
In natural ecosystems, terrestrial plants establish symbiotic associations with mycorrhizal fungi. Subterranean symbionts AMF are of paramount importance in terrestrial ecosystems due to their crucial role in the regulation of nutrients and the carbon cycle, as well as their influence on soil structure and the multifaceted functions of ecosystems. AMF are integral components of earth ecosystems and exert significant influence on global C and nutrient cycles. AMF play a vital role in the provision of nitrogen and phosphorus to plants. Numerous vascular plant species rely on these symbiotic associations with AMF for their growth and overall survival.
The fundamental requirement for any AMF isolate intended for agricultural application is a strong capacity for root colonization. This ability is crucial as it enables the isolate to effectively compete with native AMF, which are known to be highly competitive. AMF offer mineral nutrient provision and protective benefits to their host plants in the face of biotic and abiotic stressors, suggesting their potential application as biofertilizers within the context of sustainable agricultural practices.
The role of bacterial communities in the mycorrhizosphere as a potential mediator of the root colonization capacity of AMF has been well established in scientific literature. The symbiotic relationship between host plants, AMF symbionts, and related bacteria exhibits advantageous emergent characteristics that can be effectively harnessed in the context of sustainable agriculture. The colonization capacity, efficiency, and stress tolerance of AMF exhibit significant variation among different isolates. The extensive inter- and intraspecific diversity of AMF can be effectively utilized by carefully selecting AMF inocula based on their colonization capacity and efficiency. These characteristics are influenced by the genotypes of both fungi and host plants, as well as various environmental factors.
Biotechnological tools encompass processes of biological importance that leverage the chemistry of living organisms to advance the development of novel and enhanced methods for the production of conventional products all while upholding the integrity of the natural environment. AMS as a biotechnological tool plays a crucial role in shaping plant growth and facilitating stress adaptation. The effective management and assessment of ecosystem services offered by AMF is imperative for enhancing plant production both in terms of quality and quantity within the framework of agriculture that relies on the limited use of synthetic chemical inputs. To effectively manage AMF in ecological engineering and crop production and in selecting plant species that enhance the benefits of AMF, it is crucial to comprehend the intricate mechanisms that govern the development and functions of AMS.
References
Ahammed GJ, Shamsy R, Liu A, Chen S (2023) Arbuscular mycorrhizal fungi-induced tolerance to chromium stress in plants. Environ Pollut. https://doi.org/10.1016/j.envpol.2023.121597
Alam MZ, Hoque MA, Ahammed GJ, Carpenter-Boggs L (2019) Arbuscular mycorrhizal fungi reduce arsenic uptake and improve plant growth in Lens culinaris. PLoS ONE 14(5):e0211441. https://doi.org/10.1371/journal.pone.0211441
Andrino A, Guggenberger G, Kernchen S, Mikutta R, Sauheitl L, Boy J (2021) Production of organic acids by arbuscular mycorrhizal fungi and their contribution in the mobilization of phosphorus bound to iron oxides. Front Plant Sci 12:661842. https://doi.org/10.3389/fpls.2021.661842
Begum N, Ahanger MA, Su Y, Lei Y, Mustafa NSA, Ahmad P, Zhang L (2019) Improved drought tolerance by amf inoculation in maize (Zea mays) involves physiological and biochemical implications. Plants 8(12):579. https://doi.org/10.3390/plants8120579
Bonfante P, Genre A (2010) Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis. Nat Commun 1:48. https://doi.org/10.1038/ncomms1046
Boorboori MR, Zhang HY (2022) Arbuscular mycorrhizal fungi are an influential factor in improving the phytoremediation of arsenic, cadmium, lead, and chromium. J Fungi 8(2):176. https://doi.org/10.3390/jof8020176
Branco S, Schauster A, Liao HL, Ruytinx J (2022) Mechanisms of stress tolerance and their effects on the ecology and evolution of mycorrhizal fungi. New Phytol 235(6):2158–2175. https://doi.org/10.1111/nph.18308
Breuillin-Sessoms F, Floss DS, Gomez SK, Pumplin N, Ding Y, Levesque-Tremblay V, Noar RD, Daniels DA, Bravo A, Eaglesham JB, Benedito VA, Udvardi MK, Harrison MJ (2015) Suppression of arbuscule degeneration in Medicago truncatula phosphate transporter4 mutants is dependent on the ammonium transporter 2 family protein AMT2;3. Plant Cell 27(4):1352–1366. https://doi.org/10.1105/tpc.114.131144
Chen J, Zhang H, Zhang X, Tang M (2017a) Arbuscular mycorrhizal symbiosis alleviates salt stress in black locust through improved photosynthesis, water status, and K(+)/Na(+) homeostasis. Front Plant Sci 8:1739. https://doi.org/10.3389/fpls.2017.01739
Chen S, Zhao H, Zou C, Li Y, Chen Y, Wang Z, Jiang Y, Liu A, Zhao P, Wang M, Ahammed GJ (2017b) Combined inoculation with multiple arbuscular mycorrhizal fungi improves growth, nutrient uptake and photosynthesis in cucumber seedlings. Front Microbiol 8:2516. https://doi.org/10.3389/fmicb.2017.02516
Cui X, Jia B, Diao F, Li X, Xu J, Zhang Z, Li FY, Guo W (2022) Transcriptomic analysis reveals the molecular mechanisms of arbuscular mycorrhizal fungi and nitrilotriacetic acid on Suaeda salsa tolerance to combined stress of cadmium and salt. Process Saf Environ Prot 160:210–220. https://doi.org/10.1016/j.psep.2022.02.019
Das D, Paries M, Hobecker K, Gigl M, Dawid C, Lam HM, Zhang J, Chen M, Gutjahr C (2022) Phosphate starvation response transcription factors enable arbuscular mycorrhiza symbiosis. Nat Commun 13(1):477. https://doi.org/10.1038/s41467-022-27976-8
Dhalaria R, Kumar D, Kumar H, Nepovimova E, Kuča K, Torequl Islam M, Verma R (2020) Arbuscular mycorrhizal fungi as potential agents in ameliorating heavy metal stress in plants. Agronomy 10(6):815. https://doi.org/10.3390/agronomy10060815
Diagne N, Ngom M, Djighaly PI, Fall D, Hocher V, Svistoonoff S (2020) Roles of arbuscular mycorrhizal fungi on plant growth and performance: importance in biotic and abiotic stressed regulation. Diversity 12(10):370. https://doi.org/10.3390/d12100370
Edlinger A, Garland G, Hartman K, Banerjee S, Degrune F, García-Palacios P, Hallin S, Valzano-Held A, Herzog C, Jansa J, Kost E, Maestre FT, Pescador DS, Philippot L, Rillig MC, Romdhane S, Saghaï A, Spor A, Frossard E, van der Heijden MGA (2022) Agricultural management and pesticide use reduce the functioning of beneficial plant symbionts. Nat Ecol Evol 6(8):1145–1154. https://doi.org/10.1038/s41559-022-01799-8
Fierer N (2017) Embracing the unknown: disentangling the complexities of the soil microbiome. Nat Rev Microbiol 15(10):579–590. https://doi.org/10.1038/nrmicro.2017.87
Fiorilli V, Vannini C, Ortolani F, Garcia-Seco D, Chiapello M, Novero M, Domingo G, Terzi V, Morcia C, Bagnaresi P, Moulin L, Bracale M, Bonfante P (2018) Omics approaches revealed how arbuscular mycorrhizal symbiosis enhances yield and resistance to leaf pathogen in wheat. Sci Rep 8(1):9625. https://doi.org/10.1038/s41598-018-27622-8
Gao WQ, Wang P, Wu QS (2019) Functions and application of glomalin-related soil proteins: a review. Sains Malaysiana 48(1):111–119. https://doi.org/10.17576/jsm-2019-4801-13
Gelvez-Pardo I, Lobo-Berbesi L, Santos-Díaz A (2023) Biological efficacy of plant growth–promoting bacteria and arbuscular mycorrhizae fungi: assessments in laboratory and greenhouse conditions. Curr Protocols 3(4):e732. https://doi.org/10.1002/cpz1.732
Genre A, Chabaud M, Balzergue C, Puech-Pagès V, Novero M, Rey T, Fournier J, Rochange S, Bécard G, Bonfante P, Barker DG (2013) Short-chain chitin oligomers from arbuscular mycorrhizal fungi trigger nuclear Ca2+ spiking in Medicago truncatula roots and their production is enhanced by strigolactone. New Phytol 198(1):190–202. https://doi.org/10.1111/nph.12146
Gianinazzi S, Gollotte A, Binet MN, van Tuinen D, Redecker D, Wipf D (2010) Agroecology: the key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza 20(8):519–530. https://doi.org/10.1007/s00572-010-0333-3
Giovannini L, Palla M, Agnolucci M, Avio L, Sbrana C, Turrini A, Giovannetti M (2020) Arbuscular mycorrhizal fungi and associated microbiota as plant biostimulants: research strategies for the selection of the best performing inocula. Agronomy 10(1):106. https://doi.org/10.3390/agronomy10010106
Govindarajulu M, Pfeffer PE, Jin H, Abubaker J, Douds DD, Allen JW, Bücking H, Lammers PJ, Shachar-Hill Y (2005) Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 435(7043):819–823. https://doi.org/10.1038/nature03610
Hajiboland R, Joudmand A, Aliasgharzad N, Tolrá R, Poschenrieder C (2019) Arbuscular mycorrhizal fungi alleviate low-temperature stress and increase freezing resistance as a substitute for acclimation treatment in barley. Crop Pasture Sci 70(3):218–233
Hermans SM, Lear G, Case BS, Buckley HL (2023) The soil microbiome: an essential, but neglected, component of regenerative agroecosystems. Science 26(2):106028. https://doi.org/10.1016/j.isci.2023.106028
Ho-Plágaro T, García-Garrido JM (2022) Molecular regulation of arbuscular mycorrhizal symbiosis. Int J Mol Sci 23(11):5960. https://doi.org/10.3390/ijms23115960
Hui J, An X, Li Z, Neuhäuser B, Ludewig U, Wu X, Schulze WX, Chen F, Feng G, Lambers H, Zhang F, Yuan L (2022) The mycorrhiza-specific ammonium transporter ZmAMT3;1 mediates mycorrhiza-dependent nitrogen uptake in maize roots. Plant Cell 34(10):4066–4087. https://doi.org/10.1093/plcell/koac225
Jiang Y, Wang W, Xie Q, Liu N, Liu L, Wang D, Zhang X, Yang C, Chen X, Tang D, Wang E (2017) Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 356(6343):1172–1175. https://doi.org/10.1126/science.aam9970
Kakouridis A, Hagen JA, Kan MP, Mambelli S, Feldman LJ, Herman DJ, Weber PK, Pett-Ridge J, Firestone MK (2022) Routes to roots: direct evidence of water transport by arbuscular mycorrhizal fungi to host plants. New Phytol. https://doi.org/10.1111/nph.18281
Kretzschmar T, Kohlen W, Sasse J, Borghi L, Schlegel M, Bachelier JB, Reinhardt D, Bours R, Bouwmeester HJ, Martinoia E (2012) A petunia ABC protein controls strigolactone-dependent symbiotic signalling and branching. Nature 483(7389):341–344. https://doi.org/10.1038/nature10873
Li Z, Ahammed GJ (2023) Salicylic acid and jasmonic acid in elevated CO2-induced plant defense response to pathogens. J Plant Physiol. https://doi.org/10.1016/j.jplph.2023.154019
Li X, Zhang Z, Luo J, Cui X, Xu J, Hou Y, Hao B, Guo W (2022) Arbuscular mycorrhizal fungi and nitrilotriacetic acid regulated Suaeda salsa growth in Cd-contaminated saline soil by driving rhizosphere bacterial assemblages. Environ Exp Bot 193:104669. https://doi.org/10.1016/j.envexpbot.2021.104669
Liao D, Sun C, Liang H, Wang Y, Bian X, Dong C, Niu X, Yang M, Xu G, Chen A, Wu S (2022) SlSPX1-SlPHR complexes mediate the suppression of arbuscular mycorrhizal symbiosis by phosphate repletion in tomato. Plant Cell 34(10):4045–4065. https://doi.org/10.1093/plcell/koac212
López-Ráez JA, Pozo MJ (2013) Chemical signalling in the arbuscular mycorrhizal symbiosis: biotechnological applications. In: Symbiotic endophytes. Soil biology. Springer, Cham, pp 215–232. https://doi.org/10.1007/978-3-642-39317-4_11
Ma X, Li X, Ludewig U (2021) Arbuscular mycorrhizal colonization outcompetes root hairs in maize under low phosphorus availability. Ann Bot 127(1):155–166. https://doi.org/10.1093/aob/mcaa159
Maillet F, Poinsot V, André O, Puech-Pagès V, Haouy A, Gueunier M, Cromer L, Giraudet D, Formey D, Niebel A, Martinez EA, Driguez H, Bécard G, Dénarié J (2011) Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature 469(7328):58–63. https://doi.org/10.1038/nature09622
Moradtalab N, Hajiboland R, Aliasgharzad N, Hartmann TE, Neumann G (2019) Silicon and the association with an arbuscular-mycorrhizal fungus (Rhizophagus clarus) mitigate the adverse effects of drought stress on strawberry. Agronomy 9(1):41
Ossowicki A, Raaijmakers JM, Garbeva P (2021) Disentangling soil microbiome functions by perturbation. Environ Microbiol Rep 13(5):582–590. https://doi.org/10.1111/1758-2229.12989
Parihar M, Rakshit A, Meena VS, Gupta VK, Rana K, Choudhary M, Tiwari G, Mishra PK, Pattanayak A, Bisht JK, Jatav SS, Khati P, Jatav HS (2020) The potential of arbuscular mycorrhizal fungi in C cycling: a review. Arch Microbiol 202(7):1581–1596. https://doi.org/10.1007/s00203-020-01915-x
Pasbani B, Salimi A, Aliasgharzad N, Hajiboland R (2020) Colonization with arbuscular mycorrhizal fungi mitigates cold stress through improvement of antioxidant defense and accumulation of protecting molecules in eggplants. Sci Hortic 272:109575
Philippot L, Raaijmakers JM, Lemanceau P, van der Putten WH (2013) Going back to the roots: the microbial ecology of the rhizosphere. Nat Rev Microbiol 11(11):789–799. https://doi.org/10.1038/nrmicro3109
Phour M, Sehrawat A, Sindhu SS, Glick BR (2020) Interkingdom signaling in plant-rhizomicrobiome interactions for sustainable agriculture. Microbiol Res 241:126589. https://doi.org/10.1016/j.micres.2020.126589
Piliarová M, Ondreičková K, Hudcovicová M, Mihálik D, Kraic J (2019) Arbuscular mycorrhizal fungi – their life and function in ecosystem. Agriculture 65(1):3–15. https://doi.org/10.2478/agri-2019-0001
Porcel R, Aroca R, Azcon R, Ruiz-Lozano JM (2016) Regulation of cation transporter genes by the arbuscular mycorrhizal symbiosis in rice plants subjected to salinity suggests improved salt tolerance due to reduced Na(+) root-to-shoot distribution. Mycorrhiza 26(7):673–684. https://doi.org/10.1007/s00572-016-0704-5
Powell JR, Rillig MC (2018) Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol 220(4):1059–1075. https://doi.org/10.1111/nph.15119
Puschel D, Bitterlich M, Rydlova J, Jansa J (2020) Facilitation of plant water uptake by an arbuscular mycorrhizal fungus: a Gordian knot of roots and hyphae. Mycorrhiza 30(2-3):299–313. https://doi.org/10.1007/s00572-020-00949-9
Riaz M, Kamran M, Fang Y, Wang Q, Cao H, Yang G, Deng L, Wang Y, Zhou Y, Anastopoulos I, Wang X (2021) Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: a critical review. J Hazard Mater 402:123919. https://doi.org/10.1016/j.jhazmat.2020.123919
Salvioli di Fossalunga A, Novero M (2019) To trade in the field: the molecular determinants of arbuscular mycorrhiza nutrient exchange. Chem Biol Technol Agric 6(1):12. https://doi.org/10.1186/s40538-019-0150-7
Sanchez-Bermudez M, Del Pozo JC, Pernas M (2022) Effects of combined abiotic stresses related to climate change on root growth in crops. Front Plant Sci 13:918537. https://doi.org/10.3389/fpls.2022.918537
Sangwan S, Prasanna R (2022) Mycorrhizae helper bacteria: unlocking their potential as bioenhancers of plant-arbuscular mycorrhizal fungal associations. Microb Ecol 84(1):1–10. https://doi.org/10.1007/s00248-021-01831-7
Sanmartin N, Pastor V, Pastor-Fernandez J, Flors V, Pozo MJ, Sanchez-Bel P (2020) Role and mechanisms of callose priming in mycorrhiza-induced resistance. J Exp Bot 71(9):2769–2781. https://doi.org/10.1093/jxb/eraa030
Shi J, Zhao B, Zheng S, Zhang X, Wang X, Dong W, Xie Q, Wang G, Xiao Y, Chen F, Yu N, Wang E (2021) A phosphate starvation response-centered network regulates mycorrhizal symbiosis. Cell 184(22):5527–5540. https://doi.org/10.1016/j.cell.2021.09.030
Shi J, Wang X, Wang E (2023) Mycorrhizal symbiosis in plant growth and stress adaptation: from genes to ecosystems. Annu Rev Plant Biol 74(1):569–607. https://doi.org/10.1146/annurev-arplant-061722-090342
Smith FA, Smith SE (1997) Structural diversity in (vesicular)–arbuscular mycorrhizal symbioses. New Phytol 137(3):373–388. https://doi.org/10.1046/j.1469-8137.1997.00848.x
Song Y, Chen D, Lu K, Sun Z, Zeng R (2015) Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front Plant Sci 6:786. https://doi.org/10.3389/fpls.2015.00786
Tominaga T, Miura C, Takeda N, Kanno Y, Takemura Y, Seo M, Yamato M, Kaminaka H (2019) Gibberellin promotes fungal entry and colonization during Paris-type arbuscular mycorrhizal symbiosis in eustoma grandiflorum. Plant Cell Physiol 61(3):565–575. https://doi.org/10.1093/pcp/pcz222
Tominaga T, Yao L, Saito H, Kaminaka H (2022) Conserved and diverse transcriptional reprogramming triggered by the establishment of symbioses in tomato roots forming arum-type and Paris-type arbuscular mycorrhizae. Plants 11(6):747. https://doi.org/10.3390/plants11060747
van der Heijden MGA, Martin FM, Selosse MA, Sanders IR (2015) Mycorrhizal ecology and evolution: the past, the present, and the future. New Phytol 205(4):1406–1423. https://doi.org/10.1111/nph.13288
Wang L, Chen X, Du Y, Zhang D, Tang Z (2022) Nutrients regulate the effects of arbuscular mycorrhizal fungi on the growth and reproduction of cherry tomato. Front Microbiol 13:843010. https://doi.org/10.3389/fmicb.2022.843010
Wipf D, Krajinski F, van Tuinen D, Recorbet G, Courty PE (2019) Trading on the arbuscular mycorrhiza market: from arbuscules to common mycorrhizal networks. New Phytol 223(3):1127–1142. https://doi.org/10.1111/nph.15775
Wu M, Yan Y, Wang Y, Mao Q, Fu Y, Peng X, Yang Z, Ren J, Liu A, Chen S, Ahammed GJ (2021) Arbuscular mycorrhizal fungi for vegetable (VT) enhance resistance to Rhizoctonia solani in watermelon by alleviating oxidative stress. Biol Control 152:104433. https://doi.org/10.1016/j.biocontrol.2020.104433
Yang H, Fang C, Li Y, Wu Y, Fransson P, Rillig MC, Zhai S, Xie J, Tong Z, Zhang Q, Sheteiwy MS, Li F, Weih M (2022) Temporal complementarity between roots and mycorrhizal fungi drives wheat nitrogen use efficiency. New Phytol. https://doi.org/10.1111/nph.18419
Yooyongwech S, Samphumphuang T, Tisarum R, Theerawitaya C, Cha-um S (2016) Arbuscular mycorrhizal fungi (AMF) improved water deficit tolerance in two different sweet potato genotypes involves osmotic adjustments via soluble sugar and free proline. Sci Hortic 198:107–117. https://doi.org/10.1016/j.scienta.2015.11.002
Zhang C, He J, Dai H, Wang G, Zhang X, Wang C, Shi J, Chen X, Wang D, Wang E (2021a) Discriminating symbiosis and immunity signals by receptor competition in rice. Proc Natl Acad Sci 118(16):e2023738118. https://doi.org/10.1073/pnas.2023738118
Zhang L, Chu Q, Zhou J, Rengel Z, Feng G (2021b) Soil phosphorus availability determines the preference for direct or mycorrhizal phosphorus uptake pathway in maize. Geoderma 403:115261. https://doi.org/10.1016/j.geoderma.2021.115261
Zhang L, Zhou J, George TS, Limpens E, Feng G (2022) Arbuscular mycorrhizal fungi conducting the hyphosphere bacterial orchestra. Trends Plant Sci 27(4):402–411. https://doi.org/10.1016/j.tplants.2021.10.008
Zhou Y, Ge S, Jin L, Yao K, Wang Y, Wu X, Zhou J, Xia X, Shi K, Foyer CH, Yu J (2019) A novel CO2-responsive systemic signaling pathway controlling plant mycorrhizal symbiosis. New Phytol 224(1):106–116. https://doi.org/10.1111/nph.15917
Zhu B, Gao T, Zhang D, Ding K, Li C, Ma F (2022) Functions of arbuscular mycorrhizal fungi in horticultural crops. Sci Hortic 303:111219. https://doi.org/10.1016/j.scienta.2022.111219
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2024 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Ahammed, G.J., Hajiboland, R. (2024). Introduction to Arbuscular Mycorrhizal Fungi and Higher Plant Symbiosis: Characteristic Features, Functions, and Applications. In: Ahammed, G.J., Hajiboland, R. (eds) Arbuscular Mycorrhizal Fungi and Higher Plants. Springer, Singapore. https://doi.org/10.1007/978-981-99-8220-2_1
Download citation
DOI: https://doi.org/10.1007/978-981-99-8220-2_1
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-99-8219-6
Online ISBN: 978-981-99-8220-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)