
Abstract
We analyzed the home range of deer in western Japan. We collected data on 120 GPS-collared deer from Kinki, Shikoku, and Kyushu. The home range size in western Japan was smaller than that of deer in eastern and northern Japan. The percentage of individuals that made seasonal movements in western Japan was 20%. The percentage of seasonal migrants in collared deer varied regionally. The proportion of seasonal migrants was higher in areas with more snow accumulation. As the period during which the snow depth exceeded 50 cm became longer, the proportion of seasonal migrants increased. The migration distance ranged from 1.9 to 14.3 km, which was shorter than those in eastern and northern Japan. Further, individuals that use deciduous broad-leaved forests as their summer habitat tended to migrate seasonally, while deer who used evergreen broad-leaved forests tended to be more sedentary. Vegetation and topography in western Japan differ significantly from those in eastern and northern Japan, and deer behavior is also characteristic and unique to western Japan.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

1 Introduction
In recent years, increase in the population of sika deer has had a significant impact on forest ecosystems, such as decline in understory vegetation (Kabaya 1988; Hasegawa 2000; Ohashi et al. 2007; Tsujino et al. 2007; Murata et al. 2009; Hattori et al. 2010) and soil erosion (Furusawa et al. 2003; Yanagi et al. 2008). Therefore, in 2013, as a drastic control measure, the national government set a goal to halve the population of sika deer estimated in 2011 by 2025. This measure promoted the deer-capturing business. However, since the number of hunters in Japan has decreased, there is a need to develop more efficient capturing methods and conventional practices. Therefore, behavioral characteristic surveys of sika deer were conducted in various national and local governments.
Since the 1980s, surveys on the behavioral characteristics of sika deer have been conducted using very high frequency (VHF) transmitters, mainly in the Kanto and northward (e.g., Maruyama 1981; Furubayashi et al. 1987; Nagata et al. 1994; Yamazaki and Furubayashi 1995; Borkowski and Furubayashi 1998; Sakuragi et al. 2002; Nagata 2005), which replaced GPS collars in the 2000s and used extensively (e.g., Takii et al. 2012a). At that time, the surveys using GPS collars expanded in western Japan as national and local government projects since the increase in the density of sika deer has been having a substantial impact on forest vegetation (Tsujino et al. 2007; Murata et al. 2009; Hattori et al. 2010; Matsui et al. 2011).
Previous surveys reveal two contrasting movement patterns: seasonal migrants in northern and eastern Japan and sedentary individuals in western Japan (Yabe and Takatsuki 2009). It is inferred that in northern and eastern Japan, sika deer migrates to wintering grounds during winter (Miura 1974; Maruyama 1981) due to the presence of snow that restricts their behavior (Takatsuki 1992) and the scarcity of food resources. On the other hand, in western Japan, since there are many evergreen broad-leaved forests and food resources do not change much between summer and winter, there are many sedentary individuals (Yabe and Takatsuki 2009).
Some studies have reported that the snow depth of 50 cm or more restricted sika deer movements and sika deer avoided these areas (Tokita et al. 1981; Takatsuki 1992). In cervids inhabiting high latitudes such as white-tailed deer (Odocoileus virginianus) and roe deer (Capreolus capreolus), their migration patterns from summer to winter habitats are mainly attributed to snow depth (Tierson et al. 1985; Mysterud 1999; Sabine et al. 2002; Brinkman et al. 2005; Ramanzin et al. 2007) and low temperatures (Nelson 1995).
In the case of sika deer, the formation of wintering grounds for seasonal migration has been reported in snowy and high-altitude areas in northern and eastern Japan such as Hokkaido (Uno and Kaji 2000; Igota et al. 2004), Mt. Goyo (Takatsuki et al. 2000), Nikko (Maruyama 1981), Okuchichibu (Takii et al. 2012a), and Kirigamine (Takii et al. 2012b). Although the presence of seasonal migrants in western Japan has also been confirmed, only a few reports are available in limited areas such as Mt. Hyonosen range (Morimitsu et al. 2019), Odaigahara (Maeji et al. 2000), and the central mountains of Kyushu (Yabe et al. 2001). While the seasonal migration in Mt. Hyonosen range and Odaigahara was reported to be caused by snow cover, the movement of males during the breeding season in the mountains of Kyushu has no reference to the effects of snow cover.
Although only a few comprehensive reports on the behavioral characteristics of sika deer in western Japan are available, the number of survey cases conducted by governmental countermeasure projects has increased in recent years. However, in many of these cases, and since the number of individuals surveyed is small, the reports are only fragmentary. Therefore, in this paper, to elucidate the behavioral characteristics of sika deer in western Japan, we have aggregated and analyzed these survey data focusing on the home range area, seasonal migration, and habitat used compared with the previous studies in northern and eastern populations.
2 Survey Area
Western Japan refers to the region west of Kinki that includes Kinki, Chugoku, Shikoku, and Kyushu (Fig. 14.1). In this survey, data were collected only from three regions: Kinki, Shikoku, and Kyushu.

Study areas in western Japan
There are no mountains higher than 2000 m in western Japan; the highest peak is Mt. Ishizuchi in Shikoku with an elevation of 1982 m. The highest mountain range in Kinki is the Kii Mountains with an elevation of 1800 to 1900 m. The mountains of central Kyushu are the highest with an elevation 1500 to 1800 m, and the only mountain over 1800 m is the Yakushima.
Japan is a long country extending from north to south, ranging from temperate humid to subarctic continental humid climates, and western Japan belongs to the former. The monthly average temperature varies between 5 and 30 °C in Fukuoka, and the hottest days are in August with temperatures exceeding 35 °C at times. Annual precipitation varies between regions from 1000 mm to over 3000 mm. The amount of snowfall tends to be more on the Sea of Japan side and less on the Pacific side. While there are many areas with rainfall exceeding 100 cm in northern and eastern Japan, the rainfall in western Japan varies between 50 and 100 cm in some mountainous areas and 0 to 5 cm on the plains. It is an area of little snowfall and almost no snow cover (Japan Meteorological Agency) (Fig. 14.2).

Maximum snowfall depth in Japan (average values from 1971 to 2000) (Modified from the Japan Climate Map FY2000)(Japan Meteorogical Agency 2012)
While northern and eastern Japan have many deciduous broad-leaved forests dominated by beech (Fagus crenata) and Mongolian oak (Quercus crispula), western Japan has many evergreen broad-leaved forests dominated by Castanopsis spp. and Japanese emperor oak (Quercus dentata), and deciduous broad-leaved forests in high-altitude areas (Fig. 14.3).

Vegetation distribution in Japan. Modified from Japanese Vegetation by Miyawaki (1977)
Another characteristic of western Japan is the large area of plantation forests that accounts for half of the afforested area in Japan. The area of plantation forests is 27% in Hokkaido, 40% in Tohoku, 45% in and Kanto, 39% in Chubu, and 50% in western Japan (51%, 41%, 60%, and 53% in Kinki, Chugoku, Shikoku, and Kyushu, respectively). Japanese cedar (Cryptomeria japonica) and Japanese cypress (Chamaecyparis obtusa) account for 70% or more of the plantation forests in Japan, except for Hokkaido. The proportion of evergreen conifers, such as Japanese cedar and cypress, in the plantation forests is as high as 93%, 94%, and 93% in Kinki, Shikoku, and Kyushu, respectively (Forestry Agency Private Forest Department Planning Division 2018).
3 Methods
3.1 Data Collection
3.1.1 GPS Positioning Data
We collected the positioning data of GPS-collared sika deer in western Japan from the national and local government projects from 2009 to 2019. Inquiring the related organizations in Kinki, Shikoku, and Kyushu, we obtained data on 145 deer in total. The northernmost area was Mt. Ibuki, Shiga Prefecture (35°25′N 136°24′E), and the southernmost was Yakushima, Kagoshima Prefecture (30°13′N 130°30′E). The number of surveyed individuals is shown for each attribution of collected data and survey year (Table 14.1). Among the collected data, those with a tracking period of less than 90 days were excluded from the analysis. Further, 3D data with high GPS positioning accuracy were used for analysis; individuals whose 3D data was less than 1 point per day were excluded from the analysis.
3.1.2 Environmental Data
To analyze the habitat used by sika deer and the factors of its seasonal migration, we used the vegetation and snow depth data. For the vegetation, we used the high-resolution land use/land cover map (resolution 10 m) provided by the Earth Observation Research Center of the Japan Aerospace Exploration Agency (JAXA) (2018). This data classifies the land use into 10 types: water, urban and built-up, rice paddy, crops, grassland, deciduous broad-leaved forest, deciduous conifer forest, evergreen broad-leaved forest, evergreen conifer forest, and bare land.
For the snow cover data, we used the Agro-Meteorological Grid Square Data, NARO (Ohno et al. 2016). This data was created in units of about 1 km2 (reference region mesh) by interpolation based on various meteorological data such as AMeDAS, taking into consideration the altitude.
3.2 Analysis
3.2.1 Home Range
The home range was estimated using two methods: minimum convex polygon (MCP) and fixed kernel (Worton 1989). For both methods, the “adehabitat” package for the R software (ver.4.0.3) was used. The home range was estimated by 90% home range of MCP and fixed kernel (hereafter referred to as “MCP90HR” and “FK90HR”, respectively) using all data during the follow-up period. For the core area, 50% home range was calculated using fixed kernel (hereafter referred to as the “core area”). The estimated home range was converted into a “shape file,” and the home range was analyzed using ArcGIS (ver.10.5.1). For the individuals for whom multiple core areas were formed due to seasonal migration or the like, estimation was performed by dividing the period into before and after migration.
The statistical difference in home range areas between sexes was examined by a Mann-Whitney U in Kinki and Kyushu and a Welch t-test in Shikoku. The Steel-Dwass test was used to examine the difference of home range areas between three regions. The significant level was set as 0.05.
3.2.2 Determination of the Presence or Absence of Seasonal Migration
To identify the individuals that have migrated, the core area was estimated for the entire survey period, and the ones in which multiple core areas were formed were extracted. In addition, the positioning data of each individual was classified per month, and the presence or absence of migration was confirmed from the monthly position point distribution. Further, the core areas were estimated for the period before and after the migration; individuals with no overlapping core areas (before and after migration) were regarded as seasonally migrating. The distance between the centers of gravity of the estimated core areas before and after the migration was calculated by ArcGIS.
3.2.3 Habitat Use
The snow depth of the center of gravity of the MCP90HR of the individuals for whom the tracking period included winter were extracted and classified into 50 cm of snow depth classes. The vegetation from the GPS data points was extracted using the high-resolution land use/land cover map (resolution 10 m) and analyzed. A Pearson’s chi-squared test was used to compare the vegetation used by sedentary deer with migratory individuals.
4 Results
4.1 Information of Analyzed Individuals
Among the data collected from the GPS-collared sika deer (n = 145), three deer had less than one point of positioning data per day. The number of deer with a tracking period of more than 90 days used in the analysis was 120 individuals (with male:female ratios of 8:37, 3:3, and 21:48 in Kinki, Shikoku, and Kyushu, respectively). The total individuals with a tracking period of 180 days or more were 69 (male:female ratios of 6:23, 3:1, and 9:27 in Kinki, Shikoku, and Kyushu, respectively). Only 32 individuals with a tracking period of 300 days or more were found (male:female ratios of 4:17, 3:1, and 0:7 in Kinki, Shikoku, and Kyushu, respectively) (Table 14.2). Since long-term surveys have not been conducted in Shikoku and Kyushu, only a few individuals had a tracking period of 300 days or more.
4.2 Home Range Size
Table 14.3 shows the average home range and core areas of sika deer in Kinki, Shikoku, and Kyushu. The home range size of individuals with a tracking period of 90 days or more was the largest for males in Kinki with 5.90 km2 in MCP90HR and 3.84 km2 in FK90HR. The home range of females was also the largest in Kinki, with 2.09 km2 and in MCP90HR. Males had a larger core area size (FK50) than females in Kyushu.
Fig. 14.4 shows the home range size by region and by sex. Comparing the home range size between the sexes by region, there was a significant difference between Kinki and Kyushu in both MCP90HR and FK90HR, and the home range area was larger for males in both regions (MCP90HR; Kinki: W = 254, p < 0.001, Kyushu: W = 681, p < 0.05, FK90HR; Kinki: W = 204, p < 0.05, Kyushu: W = 694, p < 0.05). There was a significant difference between the core area of males and females in Kyushu, with a broader core area for males (W = 774, p < 0.05).

Home range size and core area estimated using the minimum convex polygon and fixed kernel for 90% of the home range (MCP90HR and KF90HR, respectively) analyzed for individuals with a tracking period of 90 days or longer
Comparing the regions by sex, there was a significant difference between the males in Kinki and Kyushu in MCP90HR, where Kinki showed a larger home range (t = 2.830, p < 0.05). The same was true for females; Kinki had a larger home range than Kyushu (t = 2.810, p < 0.05). In FK90HR, while there was no regional difference in males, there was a significant difference between the two regions in females, with a larger home range in Kinki (t = 3.383, p < 0.01). In the core area, while there was no regional difference in males, there was a significant difference in females, with a larger core area in Kinki (t = 2.651, p < 0.05).
4.3 Percentage of Seasonally Migrating Individuals
The percentages of seasonally migrating individuals by region were 37% and 10% in Kinki and Kyushu (Table 14.4), respectively, and no migrating individuals in Shikoku. Males migrated at a higher rate than females. Of the 120 GPS-collared sika deer, 80% exhibited sedentary behavior, and 20% exhibited seasonal migration.
In Kinki, the males migrated 1.9 to 5.9 km, with an average of 3.6 km (n = 6), while the females migrated 2.1 to 9.2 km, with an average of 4.3 km (n = 11). In Kyushu, the males migrated 3.6 to 4.0 km, with an average of 3.8 km (n = 5), while females migrated 2.1 to 14.3 km with an average of 7.0 km (n = 2).
In this survey, 7 out of 11 migratory males moved to their winter habitat around November, and the rest 4 moved during summer and autumn months. One, in the Kinki, out of four males moved to an area 5 km far from its normal habitat in May, stayed there for a month, and then returned to its normal habitat. And one male in the Kinki and two in the Kyushu moved to the unusual area in the period between September and November.
On the other hand, all collared migratory females moved from summer habitat to winter habitat between November and January. None of them moved at different times of the year as in the case of males.
4.4 The Effect of Snow Depth on Percentage of Seasonally Migrating Individuals
The snow depth of the center of gravity of MCP90HR and the numbers of sedentary and migrating individuals are shown in Table 14.5. In the presence of 0–50 cm snow, 22.2% of males and 1.7% of females migrated. At a snow depth of 50–100 cm, 28.6% of males and 33.3% of females migrated. On the contrary, in areas with heavy snowfall (snow depth of more than 100 cm), 100% of males and 75% of females were migratory, showing high percentage of migratory sika deer.
A previous study suggested that sika deer tended to avoid areas with more than 50 cm of snowfall for 10 days or more (Tokita et al. 1981). Table 14.6 shows the percentage of migratory sika deer, according to the three classes with more than 50 cm of snow: less than 10 days, 10 to 19 days, and 20 days or more. When the snow depth was 50 cm or more for less than 10 days, the percentage of migratory deer was 23.8% and 3.2% for males and females, respectively. When the duration of snow was for 20 days or more, 71.4% and 70.0% of males and females migrated, respectively. No migration was recorded between 10 and 19 days.
Further, the number of days on which the snow depth was 50 cm or more was compared with the home range size of MCP90HR (Fig. 14.5). The median home range (MCP90HR) for the class where the snow depth was 50 cm or more for less than 10 days was 0.42 km2, and for 20 days or more, the median was 3.37 km2, a significantly larger home range (W = 313, p < 0.001) than the former class.

Relationship between the number of days with a snow depth of 50 cm or more and MCP90HR area of sika deer in Kinki, Shikoku, and Kyushu regions
4.5 Habitat Use
The vegetation types from the GPS positions were compared separately for sedentary and migratory individuals (Fig. 14.6). The sedentary individuals most often use evergreen coniferous forests compared to seasonal migrants in Kinki (with 48.3% and 51.3%, and 24.8% and 18.2% for males and females, respectively), showing a higher utilization rate (males: χ2 = 874, p < 0.001, females: χ2 = 9624, p < 0.001). Seasonal migrants utilized the deciduous broad-leaved forests more often than sedentary individuals (59.0% and 73.4%, and 42.3% and 32.2% for males and females, respectively). The utilization of the deciduous broad-leaved forests by seasonal migrants was therefore higher than that of sedentary individuals (males: χ2 = 365, p < 0.001, females: χ2 = 14,475, p < 0.001).

Comparison of vegetation used by sedentary migrants of sika deer in Kinki and Kyushu regions. The percentages indicate the percentage of points of the vegetation type at the positioning point
In Kyushu, evergreen coniferous forests were most often used by sedentary individuals compared to seasonal migrants (with 48.2% and 45.4%, and 32.0% and 35.7% for males and females, respectively) showing a lower utilization rate than sedentary individuals (males: χ2 = 1448, p < 0.001, females: χ2 = 507, p < 0.001). In males, there was no significant difference between the seasonal migrants and sedentary individuals for vegetation types other than evergreen coniferous forests. The evergreen broad-leaved forest utilization rate by females was 29.0% and 21.8% for sedentary individuals and seasonal migrants (χ2 = 343, p < 0.001), respectively. The utilization rate of deciduous broad-leaved forests was 22.4% and 37.8% for sedentary individuals and seasonal migrants (χ2 = 1792, p < 0.001), respectively.
The monthly variation in the vegetation used by individuals with a tracking period of 300 days or more is shown in Fig. 14.7. Sedentary individuals did not show any significant change in vegetation type used throughout the year, regardless of the region or sex. However, grasslands tended to be used more frequently from April to October. Seasonally migrating males use grasslands more often between June and July. On the other hand, there was an apparent seasonal change in the vegetation type used by seasonally migrating females. While the utilization rate of deciduous broad-leaved forests was high during summer, evergreen coniferous and broad-leaved forests were mostly preferred during the winter.

Monthly vegetation use by sika deer by sedentary and migratory with a tracking period of 300 days or more in Kinki and Kyushu regions. The percentages indicate the percentage of points of the vegetation type at the positioning point
5 Discussion
5.1 Home Range Size
The home range size is influenced by various factors such as the quality and quantity of food resources, density of sika deer (Takatsuki 1992), and disturbance caused by human activities such as hunting (Kamei et al. 2010). The presence or absence of seasonal migration due to snow cover, which restricts the behavior of sika deer, strongly affects the size of their home range (Takatsuki 1992; Igota et al. 2004; Takii et al. 2012a). In the results of this analysis, males showed a larger home range size and core area size than females. In general, males of sika deer are known to have a larger home range than females (Maeji et al. 2000), and the same result was obtained in this study. The percentage of seasonal migrants varied from region to region. Further, the proportion of seasonal migrants was higher in areas with more snow, and as the period during which the snow depth exceeded 50 cm became longer, the proportion of seasonal migrants increased, which affected the home range size. This result is similar to the results from other regions such as Hokkaido (Igota et al. 2004), Nikko (Maruyama 1981), Kirigamine (Takii et al. 2012b), and Mt. Hyonosen (Morimitsu et al. 2019), where snow cover is reported as one of the main causes for seasonal migration.
In terms of the size of the core area, the summer home range was 0.19 to 6.02 km2 in Hokkaido (northern Japan) (Igota et al. 2004), 0.03 to 0.4 km2 in Okuchichibu (eastern Japan) (Takii et al. 2012a), and 0.1 to 0.6 km2 in Aichi Prefecture (eastern Japan) (Kamata et al. 2020). The size of the core area of sika deer in western Japan was 0.1 to 0.2 km2, which is relatively small. It is presumed that in Japan as a whole, the core area size is larger at high latitudes.
In western Japan, the use of evergreen coniferous and broad-leaved forests was high, especially in Kyushu; the evergreen broad-leaved forests were used extensively throughout the year. The size of the core area was smaller in Kyushu than Kink, where evergreen broad-leaved forests were intensively used due to the quantity and quality of food resources. However, the home range size may be affected not only by the food resources but also by the body size of the deer, topography, surrounding human activities, and deer density. Since these factors were not evaluated in this analysis, further research is necessary.
5.2 Percentage of Seasonal Migrants
The percentage of seasonal migrants was only 38% (17 out of 45 individuals) in Kinki and 10% (7 out of 69 individuals) in Kyushu, compared to 76% in Hokkaido (Igota et al. 2004), 100% (6 of 6 individuals) in Okuchichibu (Takii et al. 2012a), and 65% (15 out of 23 individuals) in Kirigamine (Takii et al. 2012b). Compared to these regions, the proportion of sika deer migrants in western Japan is low. In this survey, the distance traveled was between 1.9 and 14.3 km for all individuals, compared to 7.2–101.7 km in Hokkaido (Igota et al. 2004), 3.0–40.3 km in Kirigamine (Takii et al. 2012b), and 2.5–31.9 km in Okuchichibu (Takii et al. 2012a). Therefore, the migration distance in western Japan is shorter than those in other regions.
In the Kinki region, the movements of individuals that underwent seasonal migration in winter showed that they did not stay in their winter habitat all the way until spring but that they tried to return to their summer habitat, moved, but then turned back to their winter habitat again (Fig. 14.8). In northern and eastern Japan, deer tend to migrate directly to the wintering ground and remain there until spring. However, the seasonal migrants in western Japan tried to return to their summer home range when the deer was at the wintering ground. From this behavior, we can infer that the winter habitat of the sika deer in western Japan is not a fixed one but rather serves as a temporary shelter from the snow.

Example of migration routes of two deer (Mt. Ibuki, Shiga Prefecture) ID6 and ID9 are adult females. The land cover is shown as DBF for deciduous broad-leaved forest, DCF for deciduous conifer forest, EBF for evergreen broad-leaved forest, and ECF for evergreen conifer forest
5.3 Causes of Seasonal Migration
We confirmed that most of the migratory males migrated in winter, while some migrated in autumn and summer. The reason for the individuals migrating in winter can be attributed to the scarcity of food due to snowfall and defoliation, which was clear from the relationship of snow depth and number of days of snow. The breeding season of the sika deer is in the fall, from mid-September to late November (Yamauchi et al. 1997, 1999; Yokoyama et al. 2003), and individuals that migrated in the fall are considered to have migrated for breeding purposes. Another survey in the central mountains of Kyushu (Yabe et al. 2001) also reported that males migrated in the breeding season; therefore, we can confirm that the individuals that migrated in the autumn do so to breed. The vegetation at the summer habitat in Shiga Prefecture was grasslands, and many grazing marks have been confirmed (Wildlife Management Office 2011). Therefore, migration during summer may be due to the search of better-quality food resources.
Meanwhile seasonally migrating females only migrated in winter. Thus, sika deer in western Japan migrated during winter due to scarcity of food resources, while males also migrated to access better forage and breeding areas in summer and autumn.
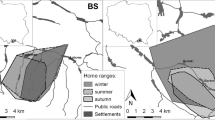
Considering western Japan as a whole, seasonal migrants often utilized deciduous broad-leaved forests. Particularly, the females in Kinki used deciduous broad-leaved forests from summer to autumn and migrated to evergreen coniferous or evergreen broad-leaved forests in winter. On the other hand, sedentary individuals used deciduous broad-leaved forests in low proportion and evergreen broad-leaved forests and evergreen coniferous forests in high proportion. Fig. 14.9 shows the home range of Mt. Ibuki in Kinki, where many seasonal migrants were present, while the home range of Yakushima, characterized by sedentary individuals, is superimposed on the vegetation map. In Mt. Ibuki, where there were many seasonal migrants, the primary vegetation in the home range was deciduous broad-leaved forests, whereas in Yakushima, where there were many sedentary individuals, the entire home range was covered with evergreen broad-leaved forests, and the home range was significantly small.

Comparison of the sika deer home range on Yakushima Island, Kagoshima Prefecture, and Mt. Ibuki, Shiga Prefecture. (Left: Yakushima 30°21′N 130°23′E, Right: Mt. Ibuki 35°24′N 136°25′E). The scales of the left and right figures are almost the same
The individual that traveled the maximum distance was an adult female in the central mountains of Kyushu. She initiated migration in October, moved southward over 2 days, and stayed there until January of the following year when the collar was recovered (Fig. 14.10). In her summer habitat, the most utilized vegetation was deciduous broad-leaved forests. On the other hand, in her winter habitat, deciduous broad-leaved forests were less used, but evergreen coniferous forests and evergreen broad-leaved forests were mainly used. From these observations, individuals that utilize deciduous broad-leaved forests in summer migrate to evergreen broad-leaved and evergreen coniferous forests as deciduous broad-leaved forests defoliate and snow limits the understory food resources, the forage quantity decrease during the winter. Hence, sika deer migrated to evergreen forests to access food resources. As the snow deepens, evergreen forests can also be used as a shelter, which explains why evergreen forests are used more in winter. On the other hand, individuals inhabiting evergreen forests are more sedentary, because the seasonal changes in the food resources are less.

Vegetation used by the seasonal migrants in the summer and winter habitat in Kyushu
5.4 Behavioral Characteristics of the Sika Deer in Western Japan
The sika deer in western Japan had a smaller home range and core area than those in northern and eastern regions, and the individuals that migrate seasonally were those from areas with heavy snow. Further, individuals that use deciduous broad-leaved forests as their summer habitat tended to migrate seasonally, while the individuals that used evergreen broad-leaved forests tended to be more sedentary. In this study, although it became clear that the proportion of migratory deer increased in areas with heavy snow, other factors that affect seasonal migration, such as topography, deer density, and human activities, could not be determined.
Japan is a country that stretches long from north to south, with a complex topography and diverse natural environment. Besides, the population density of sika deer and human activities vary from region to region, so their behavioral patterns may differ depending on the anthropogenic impact on the environment. Vegetation and topography in western Japan differ significantly from those in eastern and northern Japan, and deer behavior is also thought to have characteristics unique to western Japan. There are few reports on the behavioral characteristics of Japanese deer in western Japan, and further research is needed.
References
Borkowski J, Furubayashi K (1998) Home range size and habitat use in radio-collared female sika deer at high altitudes in the Tanzawa Moutains Japan. Ann Zool Fenn 35:181–186
Brinkman TJ, DePerno CS, Jenks JA, Haroldson BS (2005) Movement of female white-tailed deer: effects of climate and intensive row-crop agriculture. J Wildl Manag 69:1099–1111
Earth Observation Research Center of the Japan Aerospace Exploration Agency (2018) https://www.eorc.jaxa.jp/ALOS/lulc/lulc_jindex.htm#3. Accessed 10 Aug 2020
Forestry Agency Private Forest Department Planning Division (2018) Current status of forest resources, https://www.rinya.maff.go.jp/j/keikaku/genkyou/h29/index.html. Accessed 15 Dec 2020
Furubayashi K, Yamazaki K, Mori Y, Higashi H (1987) Home range of young sika stag. J Jpn For Soc Kanto 39:155–156. (in Japanese with English title)
Furusawa H, Miyanishi H, Kaneko S, Hino T (2003) Movement of soil and litter on the floor on a temperate mixed forest with an impoverished understory grazed by deer (Cervus nippon). J Jpn For Soc 85(4):318–325. Springer, Tokyo
Hasegawa J (2000) Changes of vegetation by sika deer grazing on Mt. Nikko-Shirane, Central Japan. J Phytogeogr Taxon 48:47–57. (in Japanese with English title)
Hattori T, Tochimot D, Minamiyama N, Hashimoto Y, Fujiki D, Ishida H (2010) Influence of feeding pressure by sika deer (Cervus nippon) on the primeval lucidophyllous forest in Kawanaka, Aya, Miyazaki Prefecture. Veg Sci 27:35–42. (in Japanese with English title)
Igota H, Sakuragi M, Uno H, Kaji K, Kaneko M, Akamatsu R, Maekawa K (2004) Seasonal migration patterns of female sika deer in eastern Hokkaido, Japan. Ecol Res 19:169–178
Japan Meteorogical Agency (2012) Japan climate:. https://www.jma.go.jp/jma/kishou/know/kisetsu_riyou/tenkou/Average_Climate_Japan.html. Accessed 22 Dec 2020
Kabaya H (1988) Changes of undergrowth of a broadleaved evergreen forest in relation to grazing effects by sika deer. Bull Tokyo Univ For 78:67–82. (in Japanese with English title)
Kamata A, Ishida A, Eguchi N (2020) Home range and habitat use of Sika deer around betwixt mountains and grassland in Aichi Prefecture. Chubu For Res 68:35–40. (in Japanese with English title)
Kamei T, Takeda K, Izumiyama S, Ohshima K (2010) The effect of hunting on the behavior and habitat utilization of sika deer (Cervus nippon). Mammal Study 35:235–241
Maeji I, Kurosaki T, Yokoyama S, Shibata E (2000) Home range of sika deer (Cervus nippon) on Mt. Ohdaigahara, central Japan. Nagoya Univ For Sci 19:1–10
Maruyama N (1981) A study of the seasonal movements and aggregation patterns of sika deer. Bull Fac Agric Tokyo Univ Agric Technol 23:1–85. (in Japanese with English title)
Matsui K, Horii A, Yanagi T, Morino S, Imamura A, Koda R, Tsujino R, Yumoto T, Takada K (2011) The status of forest vegetation and sika deer in the Zenki region, Mt Ohmine. Jpn J Conserv Ecol 16:111–119. (in Japanese with English title)
Miura S (1974) On the seasonal movements of sika deer population in Mt. Hinokiboramaru. J Mammal Soc Jpn 6(2):51–66. (in Japanese with English title)
Miyawaki A (1977) Vegetation of Japan compared with other region of world. Gakken, Tokyo, p 535
Morimitsu Y, Fujiki D, Saita E (2019) Analysis of seasonal migratory behavior patterns among GPS-collared sika deer (Cervus nippon) on Mt. Hyonosen, Hyogo prefecture, Japan. Hyogo Wildl Monogr 11:58–67. (in Japanese with English title)
Murata I, Inoue S, Yabe T, Kabemura Y, Kaji K, Kubota K, Mabuchi T, Shiiba Y, Utsumi Y (2009) Sika deer density and vegetation changes for 37 years in Shiiba Research Forest. Bull Kyushu Univ For 90:13–24. (in Japanese with English title)
Mysterud A (1999) Seasonal migration pattern and home range of roe deer (Capreolus capreolus) in an altitudinal gradient in southern Norway. J Zool 247:479–486
Nagata K (2005) Home range characteristic of sika deer in Fudakake of Tanzawa Moutains. Mammal Study 45(1):25–33. (in Japanese with English title)
Nagata K, Furubayashi K, Fujikami F, Makino S, Yamane M, Minagawa Y, Ishii T (1994) Sexual segregation on sika deer. J Jpn For Soc 105:551–554. (in Japanese with English title)
Nelson ME (1995) Winter range arrival and departure of white-tailed deer in northeastern Minnesota. Can J Zool 73:1069–1076
Ohashi H, Hoshino Y, Oono K (2007) Long-term changes in the species composition of plant communities caused by the population growth of sika deer (Cervus nippon) in Okutama, Tokyo. Veg Sci 24:123–151. (in Japanese with English title)
Ohno H, Sasaki K, Ohara G, Nakazono K (2016) Development of grid square air temperature and precipitation data compiled from observed, forecasted, and climatic normal data. Climate Biosph 16:71–79. (in Japanese with English title)
Ramanzin M, Sturaro E, Zanon D (2007) Seasonal migration and home range of roe deer (Capreolus capreolus) in the Italian eastern Alps. Can J Zool 85:280–289
Sabine DL, Morrison SF, Whitlaw HA, Ballard WB, Forbes GJ, Bowman J (2002) Migration behavior of white-tailed deer under varying winter climate regimes in New Brunswick. J Wildl Manag 66:718–728
Sakuragi M, Igota H, Uno H, Kaji K, Kaneko M, Akamatsu R, Maekawa K (2002) Comparison of diurnal and 24-hour sampling of habitat use by female sika deer. Mammal Study 27:101–105
Takatsuki S (1992) Foot morphology and distribution of sika deer in relation to snow depth. Ecol Res 7:19–23
Takatsuki S, Suzuki K, Higashi H (2000) Seasonal elevational movements of sika deer on Mt. Goyo, northern Japan. Mammal Study 25:107–117
Takii A, Izumiyama S, Taguchi M (2012a) Partial migration and effects of climate on migratory movements of sika deer in Kirigamine Highland, central Japan. Mammal Study 37:331–340
Takii A, Izumiyama S, Mochizuki T, Okumura T, Sato S (2012b) Seasonal migration of sika deer in the Oku-Chichibu Mountains, Central Japan. Mammal Study 37:127–137
Tierson WC, Mattfeld GF, Sage RW, Behrend DF (1985) Seasonal movements and home ranges of white-tailed deer in the Adirondacks. J Wildl Manag 49:760–769
Tokita T, Maruyama N, Ito T, Furubayashi K, Abe H (1981) Factors affecting the geographical distribution of sika deer. In: Report on animal distributions-mammal (the second basic survey on natural environment conservation). Japan Wildlife Research Center, Tokyo, pp 38–68. (in Japanese with English summary)
Tsujino R, Mtsui K, Ushimaru A, Seo A, Kawase D, Uchihashi H, Suzuki K, Takahashi J, Yumoto T, Takemon Y (2007) Invasion of the Mizorogaike Wetland by sika deer, and their effects on vegetation. Jpn J Conserv Ecol 12:20–27. (in Japanese with English title)
Uno H, Kaji K (2000) Seasonal movements of female sika deer in eastern Hokkaido, Japan. Mammal Study 25:49–57
Wildlife Management Office (2011) Survey on the effects of sika deer on vegetation in Higashiomi City. In: Higashiohmi city report, p 44. (in Japanese)
Worton BJ (1989) Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70:164–168
Yabe T, Takatsuki S (2009) Migratory and sedentary behavior patterns of sika deer in Honshu and Kyushu, Japan. In: McCullough DR, Takatsuki S, Kaji K (eds) Sika deer. Springer, Tokyo, pp 273–283
Yabe T, Koizumi T, Endo A, Seki S, Miura Y (2001) Home range of sika deer in the central mountains in Kyushu. For For Kyushu 54:131–132. (in Japanese with English title)
Yamauchi K, Hamasaki S, Takeuchi U, Mori Y (1997) Assessment of reproductive status of sika deer by fecal steroid analysis. J Reprod Dev 43(3):221–226
Yamauchi K, Hamasaki S, Takeuchi U, Mori Y (1999) Application of enzyme immunoassay to fecal steroid analysis in sika deer (Cervus nippon). J Reprod Dev 45(6):429–434
Yamazaki K, Furubayashi K (1995) A record on dispersal of a young sika stag in Western Tanzawa, Central Japan. J Jpn For Soc 77(4):305–313. (in Japanese with English title)
Yanagi Y, Takada M, Miyashita T (2008) Changes in the physical properties of forest soils in the Boso Peninsula due to sika deer revealed by surveys. Jpn J Conserv Ecol 13:65–74. (in Japanese with English title)
Yokoyama M, Sakata H, Hamasaki S, Mitani M, Tanaka T (2003) Condition assessment of sika deer population in Hyogo prefecture: characteristics of reproduction, food habits and nutritional condition. Hum Nat 14:21–31. (in Japanese with English title)
Acknowledgments
We would like to thank the Forestry Agency’s Kinki-Chugoku Regional Forest Office and Kyushu Regional Forest Office, the Ministry of the Environment’s Chugoku-Shikoku Regional Environment Office, and the prefectures of Shiga, Mie, Tokushima, and Fukuoka for generously providing GPS data.
We would also like to thank our colleagues Shin Kanno and Yu Iwata for taking time out of their work to help us analyze the GPS data. We are very grateful to Akiko Takii of Shinshu University for her advice on reviewing the text.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Yokoyama, N., Hamasaki, S., Mitsuhashi, A. (2022). Movement Patterns of Sika Deer in Western Japan. In: Kaji, K., Uno, H., Iijima, H. (eds) Sika Deer: Life History Plasticity and Management. Ecological Research Monographs. Springer, Singapore. https://doi.org/10.1007/978-981-16-9554-4_14
Download citation
DOI: https://doi.org/10.1007/978-981-16-9554-4_14
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-16-9553-7
Online ISBN: 978-981-16-9554-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)