
Abstract
We aimed to clarify the seasonal movement, activity range, and habitat use of sika deer (Cervus nippon) in Oze and Okuchichibu areas, including both plateau wetlands and forest ecosystems, Kanto region, central Japan, using GPS collars. We analyzed position data from 56 deer, 37 (5 males and 32 females) in Oze and 19 (6 males and 13 females) in Okuchichibu areas. We divided sika deer into three movement patterns as residents, seasonal migrants, and irregular migrants. The proportion of seasonal migrants was higher in Oze area (75.7%) than that in Okuchichibu (36.8%). Further, we divided seasonal migrants in Oze area into two types, i.e., with and without stopover sites on migration routes between summer and winter habitats, and found that sika deer using stopover sites spend same period in summer habitat, but the duration in winter habitat was shorter than deer without stopover. This is probably due to that they intend to intake energy efficiently by taking time along migration route in accordance with phenology. The overlap rate between wildlife protection area (where sports hunting is prohibited even in hunting season in winter) and the home range of migratory deer was higher in winter than that in summer in Okuchichibu area, and the snow depth in summer habitat was greater than that in winter habitat in Oze area. Therefore, snow depth in Oze area and hunting pressure in Okuchichibu areas should be the important cause for seasonal movement of sika deer.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


1 Introduction
Range expansion of sika deer (Cervus nippon ) in Japan has accelerated in the last few decades. Their distribution areas increased 1.7 times during the 25 years from 1978 to 2003 and 2.7 times during the 40 years from 1978 to 2018 (Ministry of the Environment 2021), respectively. Several factors are considered to have contributed to the expansion of the sika deer distribution. The natural environment includes the decrease in snow cover due to global warming and the planting of Japanese cedar (Cryptomeria japonica), Japanese cypress (Chamaecyparis obtusa var.) in lower elevation, and larch (Larix kaempferi) in the subalpine in wide areas for lumber production by cutting broadleaf forests under the forestation expansion policy that started around 1960. The change in social environment, i.e., the increase of abandoned cultivated farmland due to depopulated and aged human society in the rural areas, is also an important factor (Miura 2007).
In addition, the dramatic decreasing and aging of hunter population since the peak in the 1970s has resulted in the decrease of deer hunting (Kaji et al. 2010). In contrast to the decrease of firearm hunters, trap hunters are increasing (Ueda and Kanzaki 2006). This is because of the fact that more and more farmers start to use traps as a convenient way to capture deer for preventing crop damage. However, most farmers lack knowledge of hunting; it is difficult to expect that farmers should be responsible for population control of sika deer (Ueda and Kanzaki 2006).
On the other hand, the current expansion of deer distribution is considered as the result of the restoration in their former habitats (Kaji et al. 2000), since deer was widely inhabiting the plains areas during the Edo period (1603–1868 CE, Koganezawa 1989; Kaji et al. 2000; Furubayashi and Shinoda 2001). However, in recent years, deer have expanded distribution not only to plains but also to the areas of high elevation (Izumiyama and Mochizuki 2008). By expanding into higher elevation areas where sika deer have not inhabited or inhabited slightly, feeding could have a serious impact on the alpine vegetation and the rare plants in alpine ecosystems, which had been established under the conditions of absence or low density of sika deer (Nagaike 2012).
The seasonal migration of sika deer has been reported to range from a few to tens of kilometers in Hokkaido (Uno and Kaji 2000; Igota et al. 2004) and Kanto regions (Maruyama 1981; Homma 1995; Izumiyama and Mochizuki 2008) in Japan. Most of the migrations are altitudinal movements, moving to higher elevation in summer and lower in winter (Maruyama 1981; Takatsuki et al. 2000; Igota et al. 2004; Izumiyama and Mochizuki 2008), although the opposite movement was also reported in Hokkaido (Igota et al. 2004), the Okuchichibu Mountains (Takii et al. 2012a), and the Tanzawa Mountains (Chap. 12), central Japan, recently.
We collected the position data by fitting GPS collars to sika deer in two specific areas of Kanto region, central Japan. One is the Oze area (including the districts of Oze plateau wetlands and Nikko), where the sika deer distribution has been expanding into wetlands in the recent 30 years and has impacted wetland ecosystem by feeding. The other is the Okuchichibu area (Chichibu-Tama-Kai National Park, the closest national park to Tokyo, and surrounding area). We try to clarify characteristics of the seasonal movement, activity range (home range and core area), and habitat use of sika deer living in both Oze and Okuchichibu areas including both wetland and forest ecosystems, in Kanto region, central Japan.
2 Study Area
Oze area including the marshlands of both Ozegahara Plateau and around Lake Ozenuma with the surrounding mountains in Oze district, locate at the elevation around 1400 m on the border of Gunma, Fukushima, and Niigata prefectures, and the mountain forests in Nikko district (Fig. 11.1). Ozegahara Plateau is a marshland developed in the Oze Basin, and Lake Ozenuma is created by the damming of the headwaters of the Tadami River of the Agano River system at the elevation around 1650 m. Water supply is rich due to the heavy snow during winter and abundant rainfall during rainy season, and snow depth reaches around 3 m deep in winter. Oze district has a typical mountain climate, as it is located between the climate types of Sea of Japan and the Pacific. According to the weather station in Hinoemata Village, the closest meteorological observatory to Oze district, the annual precipitation is 1560 mm, and the average temperature is −3.8 °C in January and 20.3 °C in August (30-year average from 1991 to 2020).

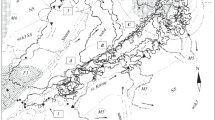
The Kanto region showing the locations of the two study areas of Oze (including Oze and Nikko districts) and Okuchichibu, central Japan (all the background map from the open map source https://maps.gsi.go.jp/development/ichiran.html#english)
The volcanic high moor formed from the damming of a depression by ejecta from volcanic eruptions of the Oze volcanoes. At Ozegahara Plateau, in the lowland along the stream, Asian skunk cabbage (Lysichiton camtschatcensis) and marsh marigold (Caltha palustris) grow, as they prefer the flowing water environment. In places far away from the stream, there are oligotrophic environments in which nutrition is supplied only by the rainwater and specific wetland flora like Hare’s tail cotton grass (Eriophorum vaginatum) and mosses (Sphagnum spp.) grow. The rare and valuable vegetation with many ponds resulted in a beautiful landscape and scenery in Oze distract and was designated as a part of Nikko National Park in 1934 and then was designated as Oze National Park in 2007 independently. The marshlands of Oze area were registered in Ramsar wetlands in 2005.
The occurrence of deer in the marshlands in Oze district was first reported in the 1980–1990s (Naito and Kimura 1998; Maruyama 2000) based on the observation of vegetation damage on altherbosa and Asian skunk cabbage by feeding, trampling, and even digging (Igarashi and Takatsuki 2008). It was believed that those deer inhabited the marshlands of Oze district only during summer due to the deep snow in winter, but there was no information for their winter habitat. Genetic analysis later proved that they originated in Nikko district of Tochigi Prefecture and Katashina Village of Gunma Prefecture (Koganezawa and Fukui 2003). In Nikko district, the deer expanded their range and increased the population size dramatically in 1980s because of decreasing snow, especially at higher elevations (Homma 1995; Li et al. 1996, 2001).
Okuchichibu area is located within the boundary area of five prefectures of Gunma, Saitama, Tokyo, Nagano, and Yamanashi, and the central part is designated as Chichibu-Tama-Kai National Park in 1950 (Fig. 11.1). The main ridge, the core area of Chichibu-Tama-Kai National Park, is lofty and has many deep valleys with steep slopes. The highest peak is Mt. Kita-Okusenjo at 2601 m in elevation, and snow depth reaches 1–2 m at main ridges in winter. The natural forests include deciduous broad-leaved primary forest with Japanese blue beech (Fagus japonica), Japanese beech (Fagus crenata), and Oak (Quercus crispula) in the lower zone around 600–1600 m in elevation and a coniferous forest at the subalpine dominated by Northern Japanese hemlock (Tsuga diversifolia), Maries’ fir (Abies mariesii), and Erman’s birch (Betula ermanii) in the higher area over 1600 m in elevation (Shigyo et al. 2017). Ohashi et al. (2014) reported the impact to natural vegetation caused by sika deer in 1990s first, and in the following decade, the impact expanded rapidly and became serious. In addition, the population increase of sika deer has caused dieback of Maries’ fir and a severe decline of broad-leaved bamboos.
Okuchichibu area is long from east to west. The annual precipitation ranges from 1375 to 1432 mm, and the average temperature ranges from −5.8 to 1.8 °C in January and from 19.5 to 25.5 °C in August (30-year average for the period 1991–2020, Nobeyama weather station in the west and Chichibu weather station in the east part of Okuchichibu area).
3 Capture and Immobilization
We used a tranquilizer gun to capture and immobilize sika deer. We estimated the body weight to confirm the suitability for fitting GPS collar when we encountered sika deer and then injected the anesthetic to suitable deer as quick as possible. For immobilization, a mixture of ketamine and xylazine was used and atropine was added to minimize the side effects.
Under anesthetic condition, we measured body size and fitted a GPS collar. Then we injected suitable atipamezole as an antagonist to help the individual awake smoothly. We were keeping observation until the deer lifted its head and walked away healthily. To avoid the deer to have myopathy by excessive stress, we handled them carefully during capturing and collar fitting process to minimize stress (Suzuki 1999).
We used the GPS collars (GPS Plus and Vertex Plus series, Vectronic Aerospace GmbH, Germany) to track the movement of sika deer. We set a positioning schedule as 1–2-h intervals. The total weight of the GPS collar is about 800 g, and considering animal welfare, we fit the collars to deer ensuring the collar weight is less than 3% of the body weight.
4 Data Analysis
We obtained position data from GPS collars collected from the field after the end of the tracking period. The position data were displayed in the Geographic Information System (GIS), and any position data that were clearly outliers were removed by visual judge (D’Eon and Serrouya 2005). Referring to the plant phenology in the study area, the winter was defined as December through February, spring as March through May, summer as June through August, and autumn as September through November. Only individuals that had more than 30 days and over 100 position data within one season were used for analysis.
We analyzed the movement pattern, activity range (home range and core area), migration route, vegetation use, and the relationship among activity range, migration destination, and land use regulation, i.e., wildlife protection area or hunting permit area, using position data.
We calculated the activity range by ad hoc of Kernel method (Pellerin et al. 2008) of Adehabitat from R package (Calenge 2006) for all sika deer. We defined the 90% of the range as home range and 50% of the range as core area (Worton 1989).
We classified the movement pattern according to the status of overlap of seasonal home ranges and the seasonality of migration of each sika deer. We classified the sika deer with separate summer and winter home ranges and who migrate depending on season regularly as seasonal migrant (migratory) and that migrate not depending on season as irregular migrant. The sika deer with overlapped summer and winter home ranges were categorized as residents (sedentary). Further, we confirmed the home range forming on migration route between summer and winter habitats. We defined the seasonal migratory sika deer with small home range in certain sites on the migration routes and who stay multiple days in the home range as a type of seasonal migratory sika deer with stopover.
The timing and route of migration should have a large impact on the activity range. For further seasonal analyses, we focus on the seasonal variations of four seasons for sedentary sika deer only. And for seasonal migratory sika deer that used separate habitats seasonally, we defined summer period as a period the sika deer stay at the summer habitat and winter period as a period the sika deer remain at the winter habitat. And we analyzed the position data in summer and winter periods separately. We defined the position data on the way between winter and summer periods as migrating position data and the duration as migration periods.
We define the migration distance as the straight line distance between the centers of activity range between the periods of summer and winter. When there were more than one activity ranges formed within summer or winter period for same sika deer, we used the one formed at the beginning of the period for distance calculation.
To determine the characteristics of the seasonal habitat use of each individual, we analyzed the vegetation type of position data, snow depth, and land use regulation in different habitats. For vegetation type, we calculated the percentage of the number of points located in specific vegetation types within the home range of the each sika deer to show the vegetation use, and we collected vegetation data from a High-Resolution Land Use and Land Cover Map published by ALOS/ALOS-2 Science Project and Earth Observation Priority Research: Ecosystem Research Group of EORC (Earth Observation Research Center) and JAXA (Japan Aerospace Exploration Agency). To find the difference in snow depth between summer and winter habitats, we used the deepest record of average data surveyed in each GIS mesh created in 2012 and collected the data from the Digital National Land Information (Ministry of Land, Infrastructure, Transport and Tourism 2010). Further, to find the seasonal home range shift relative to the land use regulation, we calculated the overlap rate between home range and wildlife protection area. And we collected the range of wildlife protection area from the data of Digital National Land Information created in 2015 (Ministry of Land, Infrastructure, Transport and Tourism 2015).
5 Result
5.1 Monitored Individuals
We fit a total of 56 GPS collars to 37 sika deer (5 males and 32 females) in Oze area between May and December from 2013 to 2017 and 19 sika deer (6 males and 13 females) in Okuchichibu area between September and January from 2005 to 2006, 2009, and from 2015 to 2017 (Table 11.1).
5.2 Movement Patterns
We confirmed movement patterns of sedentary and migratory sika deer in both study areas. In Oze area, 28 out of 37 individuals were migratory (Table 11.2, Fig. 11.2). Only one sedentary sika deer was confirmed which was captured and collared in Nikko where the winter habitat of migratory deer is. The proportion of the sedentary and seasonal migratory sika deer accounted for 2.7% and 75.7%, respectively. We cannot define the movement pattern of the remaining eight individuals due to short tracking period.

Home ranges and migration routes of sika deer in Oze area (between Nikko and Oze marshlands), Kanto region, central Japan, from 2013 to 2017. Red indicates seasonal migratory sika deer using stopover sites, blue indicates sika deer not using stopover sites, and orange indicates sedentary sika deer
Further, we classified the migratory sika deer into two types, i.e., migrant with or without stopover sites on the migration routes between summer and winter habitats. Eleven sika deer were seasonal migratory sika deer with temporal stopping sites on migration routes, and 17 were seasonal migratory sika deer without stopover site and migrated directly between summer and winter habitats. The seasonal migratory sika deer with stopover sites stayed for multiple days at each site during migration and showed variable patterns in duration and location of stay depending on individual and year. For example, ID1407 who migrated in spring 2015 from south to north stopped and stayed at five points temporarily on the migrating route to Oze marshlands (Fig. 11.3). ID1410 who migrated in autumn 2015 from north to south stopped and stayed longer at a point on the migrating route to winter habitat.

Examples showing migration routes of sika deer using stopover sites in Oze area, Kanto region, central Japan. The brown line shows the spring migration route of ID1407, and the purple line shows the autumn migration route of ID1410. Red circles indicate stopover sites
In Okuchichibu area, 10 out of 19 sika deer were migratory and 9 were sedentary (Table 11.3, Fig. 11.4). Seven out of ten migratory sika deer migrated seasonally between summer and winter habitats, and three migrated irregularly not depending on seasons. The proportion of the sedentary and seasonal migratory sika deer accounted for 47.4% and 36.8%, respectively. And the proportion of sedentary sika deer in Okuchichibu area was higher than that in Oze area (Tables 11.2 and 11.3).

Home ranges and migration routes of sika deer in Okuchichibu area, Kanto region, central Japan, from 2005 to 2017. Blue indicates migratory sika deer and orange indicates resident individuals
5.3 Migration Characteristics
The start of seasonal migration varied depending on the year and study areas. In Oze area, most migrations to summer habitats started from early April to mid-May for seasonal migratory sika deer and arrived from early May to early June (Table 11.4). On the other hand, for sika deer using stopover sites, the migration started from mid-March to mid-April and arrived from early May to early June, showing a tendency of early start and synchronized arrival. When migrating to winter habitats, migratory sika deer who was not using stopover sites started migration from late October to late November and arrived from late October to early December. Sika deer who was using the stopover sites started migration from late September to early November and arrived from late November to early January, showing a late tendency to arrive their winter habitats. The average migration duration required for seasonal migratory sika deer without stopover site was 21.9 days, while it was about twice as long for individuals using stopover sites (41.0 days).
Sika deer in Okuchichibu area migrated to summer habitat starting from late March to mid-May and arrived from late March to late May, showing a 3–18-day duration, which was shorter in migration duration than that of sika deer in Oze area (Table 11.5). For migration to winter habitats, sika deer started in late November to early December and arrived from late November to early December, showing a 3–9-day duration, which was almost same as the opposite direction migration to summer habitat. However, we found one individual collared in 2016 migrated to the winter habitat in August and continually stayed in the winter area.
The migration distance in Oze area was 25.5 ± 7.6 (average ± SD) km for sika deer of seasonal migration without stopover and 30.6 ± 4.0 km for the deer with stopover, respectively (Fig. 11.5). The deer of seasonal migration with stopovers showed longer migration distances than the ones without stopover, and the longest migration distance reached 39.8 km. In Okuchichibu area, the average distance of migration was 8.1 ± 4.6 km which was much shorter than that in Oze area.

Migration distance of seasonal migratory sika deer in both Oze and Okuchichibu areas, Kanto region, central Japan. Error bars mean standard deviation
5.4 Home Range
In Oze area, the home range size of migratory sika deer was 4.4 ± 12.6 (average ± SD) km2 in summer habitat, which was larger than that in winter habitat (3.6 ± 5.9 km2, Table 11.6). The trend of seasonal difference of home range size in Okuchichibu area was same with that in Oze area and showed 6.7 ± 17.0 km2 in summer habitat and 4.1 ± 7.2 km2 in winter habitat.
On the other hand, the sedentary sika deer showed the largest home range in spring as 11.7 km2 (n = 1) in Oze area and 3.7 ± 9.4 km2 in Okuchichibu area, respectively (Table 11.7). The smallest seasonal home range was 6.4 km2 (n = 1) in summer in Oze area and 1.1 ± 1.2 km2 in autumn in Okuchichibu area, respectively.
The overlap rate of seasonal home ranges of migratory sika deer for two consecutive years was high. The lowest overlap rate also reached 88.5% in summer habitat in the Okuchichibu area (Table 11.8). The overlap rate in winter home range showed a higher value comparing with that in summer in both study areas, reaching 94.8% in Oze area and 95.8% in Okuchichibu area, respectively. We also found that sika deer tended to overwinter in almost exactly same area every year.
5.5 Habitat Use
5.5.1 Vegetation Use
In Oze area, the migratory sika deer used marshlands of Ozegahara and surrounding Lake Ozenuma as summer habitat and used forest environment in Nikko district including Ashio Mountains as winter habitat. Migratory sika deer used deciduous broad-leaved forest and deciduous conifer forest throughout years as well as grass cover (Fig. 11.6a). Sedentary sika deer used deciduous conifers mainly throughout tracking period and the utilization rate was higher than 80%. They also showed a relatively high use on deciduous broadleaved forest in early winter (December) and from late winter (February) to late spring (May) (Fig. 11.6b). In general, the migratory sika deer used deciduous broadleaved forest more than the sedentary sika deer throughout the year.

Monthly vegetation use (% of vegetation type of the position points) of sika deer of different movement patterns in Oze area, Kanto region, central Japan (a migratory; b sedentary)
In Okuchichibu area, migratory sika deer used deciduous broad-leaved forest more, although the utilization rate was high in evergreen conifers from August to October (Fig. 11.7a). And that rate of broad-leaved deciduous forest showed an opposite trend with that of evergreen conifers. Sedentary deer used mainly evergreen coniferous and deciduous broad-leaved forests, and the utilization rate of evergreen coniferous forest was higher than that of deciduous broad-leaved forests. The utilization rate showed a constant trend throughout the years (Fig. 11.7b). Similar to that in Oze area, the migratory sika deer tented to use deciduous broad-leaved forest more than sedentary sika deer throughout the year.

Monthly vegetation use (% of vegetation type of the position points) of sika deer of different movement patterns in Okuchichibu area, Kanto region, central Japan (a migratory; b sedentary)
5.5.2 The Use of Wildlife Protection Area
In Oze area, seasonal migratory sika deer had a higher overlap rate with wildlife protection area in both summer and winter habitats, but a lower overlap rate was observed during the periods of seasonal migration from March to May and from October to November (Fig. 11.8a). For sedentary sika deer, most of home range was inside wildlife protection areas (Fig. 11.8b).

The monthly overlap rate of home range with wildlife protection area (WPA) of sika deer of different movement patterns in Oze area, Kanto region, central Japan (a migratory; b sedentary)
In Okuchichibu area, migratory sika deer had a relatively low overlap rate with wildlife protection area from April to August. Then that overlap rate accounted for about 10% from September to November and 20–50% from December to March, which showed much higher than that in other months (Fig. 11.9a). On the other hand, sedentary sika deer showed a higher overlap rate with wildlife protection area throughout the years comparing with that in migrants, ranging around 50–60% (Fig. 11.9b).

The monthly overlap rate of home range with wildlife protection area (WPA) of sika deer of different movement patterns in Okuchichibu area, Kanto region, central Japan (a migratory; b sedentary)
5.5.3 Snow Depth
Comparing the maximum snow depths in summer and winter habitats in Oze area, the snow depth of summer habitat in the marshlands of Oze district was 238 (IQR, 210–268) cm, while that of winter habitat in Nikko district was 53.1 (IQR, 47.3–56) cm, indicating a greater snow depth difference between summer and winter habitats (Fig. 11.10). In contrast to Oze area, the snow depth was 27 (IQR, 26–30) cm in summer habitat and was 28.9 (IQR, 28–30) cm in winter habitat in Okuchichibu area, respectively. Snow depth did not show obvious difference between summer and winter habitats like in Oze area (Fig. 11.11).

The snow depth difference between summer (Oze district) and winter (Nikko district) habitats in Oze area, Kanto region, central Japan. The box plots indicate the first quartile and median and third quartile, and (+) indicates the mean value

The snow depth difference during winter between the home ranges of summer and winter habitats in Okuchichibu area, Kanto region, central Japan. The box plots indicate the first quartile and median and third quartile, and (+) indicates the mean value
According to the maximum snow depths of recent 30 normal years, the snow depth in Nikko district, the winter habitat of sika deer in Oze area, is close to the 50 cm limitation for sika deer moving freely (Fig. 11.12). In contrast to Oze area, the snow depth was less than 50 cm at most places in Okuchichibu area (Fig. 11.12).

Distribution of maximum average snow depth during the recent 30 years in the winter habitat in Oze and Okuchichibu areas, Kanto region, central Japan (gradient of blue indicates the variation of snow depth and dark blue the snow depth >50 cm)
6 Discussion
6.1 Characteristics of Migratory Distance
The mean seasonal migration distances in Oze area and Okuchichibu area were 25.5 and 8.1 km, respectively. The maximum migration distance of sika deer in Oze area was 39.8 km. Seasonal migration distances of sika deer have been reported in Hokkaido, ranging from 7.2 to 101.7 km (Igota et al. 2004); in Kanto Mountains, ranging from 2.5 to 31.9 km (Takii et al. 2012a); and in Kirigamine Mountain ranging from 3.0 to 40.3 km (Takii et al. 2012b).
6.2 Characteristics of Home Range
In both Oze and Okuchichibu areas, the size of the home range of seasonal migratory sika deer showed a tendency to be smaller in winter than in summer. The same trend has been reported in sika deer of Kanto Mountains (Takii et al. 2012a). The smaller winter home range is coincidence with the behavior for the ungulates in northern hemisphere to reduce food intake, activity, and movement within their limited home ranges during winter (Georgii 1980; Clutton-Brock and Harvey 1978; Georgii and Schröder 1983). In addition, the home range of migratory sika deer was larger than that of sedentary ones. Such a trend has also been reported for red deer (Kropil et al. 2015). In the case in Oze area, sika deer seems to take some time in forming stable home range after arriving at new habitat; the process may have resulted in a relatively larger home range for migratory sika deer. Further, migratory sika deer showed a high overlap rate in home ranges annually, especially in winter. The relatively higher overlap rate in winter may have resulted from the limited habitat available in winter due to heavy snow cover.
6.3 Factors of Seasonal Migration
In Oze area, 28 out of 29 and, in Okuchichibu area, 7 out of 19 were seasonal migratory sika deer. In terms of percentage, 96.6% in Oze area and 36.8% in Okuchichibu area were seasonal migratory sika deer and showed a higher proportion in Oze area than that in Okuchichibu area. The winter range of deer is determined not only by altitude but also by the distribution of snow depth in the habitat (Schmidt 1993; Takii et al. 2012a), and the determining factor for seasonal migration of deer is avoidance of deep snow cover that limits food abundance and daily movement in winter (Maruyama 1981; Uno and Kaji 2000; Igota et al. 2004). In particular, deer in Honshu of Japan have been reported to avoid areas where snow depth exceeds 50 cm (Maruyama 1981; Takatsuki 1992). The summer habitat of seasonal migratory sika deer in Oze area is located in marshlands of Oze district where the snow depth exceeds 2 m in winter. The heavy sown cover makes it difficult for sika deer to move as well as to obtain food. One sedentary deer in Oze area was collared in Nikko district, which is a part of the winter habitat of migratory sika deer in Oze area; this is why it can spend the winter without migrating. Therefore, deep snow cover was considered to be a main factor to drive seasonal migration in Oze area.
In contrast to deep snow in Oze area, the snow depth at summer habitat was about 30 cm in winter in Okuchichibu area, which was not significantly different from that at winter habitat. Therefore, it is unlikely that seasonal migration occurred for avoiding snow cover. The sedentary sika deer use about half of the home range in wildlife protection area throughout the year. On the other hand, seasonal migratory sika deer were mostly out of the protected area during summer, but the proportion using the protected area increased from December to March.
In Japan, the sports hunting starts from November 15 and finishes at February 15 of the following year (lasts until March 15 for deer and wild boar hunting in our study areas) outside wildlife protection areas. The higher overlap rate of home ranges with protected areas from December to March than that in other periods in Okuchichibu area may suggest that seasonal migratory sika deer tend to use protected areas as a refuge mainly in winter during the hunting season, for avoiding hunting risk. The seasonal change in using protected areas also suggests that the behavior of sika deer may change due to the effects of the hunting policy. Kamei (2010) and Takii et al. (2012a) reported the same trend for seasonal migration of sika deer in other areas.
In addition, deer harvest has been intensified due to the deer increase. Deer culling is conducted as permit hunting outside of the hunting period and is carried out both inside and outside of wildlife protection areas. The pressure of irregular deer culling in both timing and place may make the habitat use to become diversified and uncertain.
6.4 Characteristics of Migration with Stopover on Migrating Route
We confirmed two types of seasonal migration of sika deer in Oze area, i.e., seasonal migration with or without stopover sites on the migration route between summer and winter habitats. Both types had same summer and winter habitats, but the duration of the migration for the sika deer with stopover sites was longer than that of without stopover sites.
Seasonal migratory sika deer using stopover sites tended to arrive at their winter habitats later than those deer without stopover sites, although all of sika deer start migration at similar time to winter habitats. In contrast, in the case of migration to the summer habitat, they started migration about 1 month earlier than the individuals without stopover sites, but they arrived at summer habitat at same time. In other words, all migratory sika deer spent almost same duration in summer habitats, but the sika deer with stopover sites stayed shorter period in winter habitats than those deer without stopover sites.
Ashio Mountains of Nikko district is a winter habitat of the sika deer of Oze area, and the deer density became high for a long time before (Koganezawa et al. 2013). The high density of winter habitat will result in decrease in food resources and mass mortality during winter (Kaji et al. 1988, 2004; Takii 2013). The behavior for avoiding competition was confirmed when the density of deer becomes higher and higher in winter habitat (Mysterud et al. 2011). We considered that sika deer in Oze area may also avoid competition by shortening the period in winter habitat.
Sawyer and Kauffman (2011) reported that the quality of food at stopover sites on the way of seasonal migration was higher than that in winter habitat of mule deer. We hypothesize that sika deer using stopover sites in Oze area may also be for ingesting food efficiently by staying at stopover sites to shorten staying period in winter habitat where food is relatively scarce. In temperate areas, vertical migration from winter habitats in lower elevation to summer habitats in higher elevation is a common migration pattern (Mysterud et al. 2001), and migration allows large herbivores to have more continuous access to nutritious, newly emerged food (Fryxell and Sinclair 1988; Hebblewhite et al. 2008; Mysterud 2013). In other words, seasonal migration responds to changes in phenology and provides constant access to young, high-quality forages, which in turn influences the timing of seasonal migration (Mysterud et al. 2001). The possibility that migration using stopover sites occurs in accordance with the phenology transitions was considered. This is the first observation of sika deer using stopover sites during migration in Oze area in Japan, and the relationship between the use of stopover sites and the environmental conditions including food abundant and quality needs to be verified in the future.
References
Calenge C (2006) The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals. Ecol Model 197:516–519
Clutton-Brock TH, Harvey PH (1978) Mammals, resources and reproductive strategies. Nature 273:191–195
D’Eon RG, Serrouya R (2005) Mule deer seasonal movements and multiscale resource selection using global positioning system radio telemetry. J Mammal 86:736–744
Fryxell JM, Sinclair ARE (1988) Causes and consequences of migration by large herbivores. Trends Ecol Evol 3:237–241
Furubayashi K, Shinoda Y (2001) Distribution of sika deer (Cervus nippon) around Edo in Edo era. Wildl Conserv Jpn 7:1–24. (in Japanese)
Georgii B (1980) Home range patterns of female red deer (Cervus elaphus) in the Alps. Oecologia 47:278–285
Georgii B, Schröder W (1983) Home range and activity patterns of male red deer (Cervus elaphus) in the Alps. Oecologia 58:238–248
Hebblewhite M, Merrill E, McDermid G (2008) A multi-scale test of forage maturation hypothesis in a partially migratory ungulate population. Ecol Monogr 78:141–166
Homma K (1995) Analysis of migration patterns and habitat selection of Japanese deer in the Oku-Nikko and Ashio areas. Master thesis, Joetsu University of Education, p 60. (in Japanese)
Igarashi T, Takatsuki S (2008) Effects of defoliation and digging caused by sika deer on the Oze mires of central Japan. Biosph Conserv Nat Wildl Hum 9:9–16
Igota H, Sakuragi M, Uno H, Kaji K, Kaneko M, Akamatsu R, Maekawa K (2004) Seasonal migration patterns of female sika deer in eastern Hokkaido, Japan. Ecol Res 19:169–178
Izumiyama S, Mochizuki T (2008) Seasonal range use of sika deer which inhabits the sub-alpine zone in the Southern Japan Alps. Bull Shinshu Univ AFC 6:25–32. (in Japanese)
Kahji K, Koizumi T, Ohtaishi N (1988) Effects of resource limitation on the physical and reproductive condition of sika deer on Nakanoshima Island, Hokkaido. Acta Theriol 33:187–208
Kaji K, Miyaki M, Saitoh T, Ono S, Kaneko M (2000) Spatial distribution of an expanding sika deer population on Hokkaido Island, Japan. Wildl Soc Bull 28:699–707
Kaji K, Okada H, Yamanaka M, Matsuda H, Yabe T (2004) Irruption of a colonizing sika deer population. J Wildl Manag 68:889–899
Kaji K, Saitoh T, Uno H, Matsuda H, Yamamura K (2010) Adaptive management of sika deer populations in Hokkaido, Japan: theory and practice. Popul Ecol 52:373–387
Kamei T, Takeda K, Izumiyama S, Ohshima K (2010) The effect of hunting on the behavior and habitat utilization of sika deer (Cervus nippon). Mamm Study 35:235–241
Koganezawa M (1989) The distribution of deer and wild boar in the Edo period as seen in the “Teppo Documents”. Bull Tochigi Pref Mus 6:65–80. (in Japanese)
Koganezawa M, Fukui E (2003) Genetic structure of the local population of Nikko and Tone deer. Where do the deer of Oze come from? Wildl Forum 8:75–76. (in Japanese)
Koganezawa M, Matsuda N, Maruyama T (2013) Increase and density control of sika deer in the Nikko Wildlife Protection Area. Water Sci 334:11–25. (in Japanese)
Kropil R, Smolko P, Garaj P (2015) Home range and migration patterns of male red deer (Cervus elaphus) in Western Carpathians. Eur J Wildl Res 61:63–72
Li Y, Maruyama N, Koganezawa M, Kanzaki N (1996) Expansion of wintering grounds of Japanese deer in Nikko. Relationship between population growth and global warming. Wildl Conserv Jpn 2:23–35. (in Japanese)
Li Y, Maruyama N, Koganezawa M (2001) Factors explaining the extension of the sika deer’s range in Nikko, Japan. Biosph Conserv Nat Wildl Hum 3:55–69
Maruyama N (1981) A study of the seasonal movements and aggregation patterns of sika deer. Bull Fac Agric Tokyo Univ Agric Technol 23:1–85. (in Japanese with English title)
Maruyama N (2000) Proposal on the deer problem in Oze: from deer population management to natural ecosystem protection. Wildl Forum 5:73–84. (in Japanese)
Ministry of Environment (2021) Report on the population estimation and habitat survey of Japanese deer and wild boar in 2020, p 371. (in Japanese)
Ministry of Land, Infrastructure, Transport and Tourism (2010) Digital national land information. https://nlftp.mlit.go.jp/ksj/index.html. Accessed 31 Dec 2020
Ministry of Land, Infrastructure, Transport and Tourism (2015) Digital national land information. https://nlftp.mlit.go.jp/ksj/index.html. Accessed 31 Dec 2020
Miura S (2007) Comments on the specified wildlife conservation and management plans for sika deer in Japan. Mamm Sci 47:81–83. (in Japanese)
Mysterud A (2013) Ungulate migration, plant phenology, and large carnivores: the times they are a-changin’. Ecology 94:1257–1261
Mysterud A, Langvatn R, Yoccoz NG, Stenseth NCH (2001) Plant phenology, migration and geographical variation in body weight of a large herbivore: the effect of variable topography. J Anim Ecol 70:915–923
Mysterud A, Loe LE, Zimmermann B, Bischof R, Veiberg V, Meisingset E (2011) Partial migration in expanding red deer populations at northern latitudes—a role for density dependence? Oikos 120:1817–1825
Nagaike T (2012) Effects of browsing by sika deer (Cervus nippon) on subalpine vegetation at Mt. Kita, central Japan. Ecol Res 27:467–473
Naito T, Kimura Y (1998) Sika deer at Oze. Scientific researches of the Oze area in central Japan. Oze Sci Res Gr:725–739
Ohashi H, Hoshino Y, Nakayama C, Okumura T, Otsu C (2014) Evaluating the conservation emergency of plant communities by vulnerability to the sika deer overabundance and endangered species. J Jpn Soc Reveg Technol 39:512–520. (in Japanese)
Pellerin M, Saïd S, Gaillard JM (2008) Roe deer Capreolus home-range sizes estimated from VHF and GPS data. Wildl Biol 14:101–110
Sawyer H, Kauffman MJ (2011) Stopover ecology of a migratory ungulate. J Anim Ecol 80:1078–1087
Schmidt K (1993) Winter ecology of nonmigratory red deer. Oecologia 95:226–233
Shigyo N, Umeki K, Ohashi H, Kawada K, Hirao T (2017) Phylogenetic constraints to soil properties determine elevational diversity gradients of forest understory vegetation. Plant Ecol 218:821–834
Suzuki M (1999) A review on capture myopathy. Mamm Sci 39:1–8. (in Japanese)
Takatsuki S (1992) Foot morphology and distribution of sika deer in relation to snow depth. Ecol Res 7:19–23
Takatsuki S, Suzuki K, Higashi H (2000) Seasonal elevational movements of sika deer on Mt. Goyo, northern Japan. Mamm Study 25:107–117
Takii A (2013) Seasonal migration of sika deer in mountainous region of central Honshu. Doctor’s thesis, p 101
Takii A, Izumiyama S, Mochizuki T, Okumura T, Sato S (2012a) Seasonal migration of sika deer in the Oku-Chichibu Mountains, central Japan. Mamm Study 37:127–137
Takii A, Izumiyama S, Taguchi M (2012b) Partial migration and effects of climate on migratory movements of sika deer in Kirigamine Highland, central Japan. Mamm Study 37:331–340
Ueda G, Kanzaki N (2006) New hunters’ activities and attitudes in Shimane Prefecture. Wildl Conserv Jpn 10:9–19. (in Japanese)
Uno H, Kaji K (2000) Seasonal movements of female sika deer in eastern Hokkaido, Japan. Mamm Study 25:49–57
Worton BJ (1989) Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70:164–168
Acknowledgments
We sincerely thank the Ministry of the Environment and Bureau of Waterworks Tokyo Metropolitan Government for understanding and permitting the use of data collected in the project investigations. For field works, we thank many employees/researchers of Wildlife Management Office, Inc. For GPS data analysis, we thank N. Mitsui, A. Mitsuhashi, and S. Kanno. We also thank N. Yokoyama and S. Hamasaki for taking time to discuss data analysis and share references.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Okumura, T., Namba, Y., Jiang, ZW., Yamada, Y., Sugiura, Y. (2022). Movement Characteristics and Habitat Use of Sika Deer in Kanto Region, Central Japan. In: Kaji, K., Uno, H., Iijima, H. (eds) Sika Deer: Life History Plasticity and Management. Ecological Research Monographs. Springer, Singapore. https://doi.org/10.1007/978-981-16-9554-4_11
Download citation
DOI: https://doi.org/10.1007/978-981-16-9554-4_11
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-16-9553-7
Online ISBN: 978-981-16-9554-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)