
Abstract
Wheat (Triticum spp.) is a major staple rabi cereal contributing about 20% calories to the human diet and important cereal for ensuring food and nutritional security in many parts of the world. Burgeoning population and diversification of food habits involving more of wheat-based products have increased the demand for wheat. Therefore, integration of modern crop tools with conventional approaches is necessary to develop the future genotypes that not only will be climate resilient and input responsive but also have higher yield per unit time and space so as to meet the growing demand. It deals with the wheat crop in a holistic manner focussing on a wide range of topics including origin, evolution, taxonomy, genetics, breeding, quality, stresses, high throughput phenotyping, genomic selection, genome editing, speed breeding and new plant breeding techniques (NPBT). This chapter also covers Indian approaches of wheat varietal development and testing under co-ordinated system as a case study, and thus would be a valuable reading material for students, researchers, academicians and policy makers.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
2.1 Introduction
Wheat (Triticum spp.) is a major cereal crop contributing about 20% calories to the diet and is a staple crop of many countries including India. Wheat-based multiple products for the end users have increased the demand for wheat. Apart from major source of starch and energy, it also provides variable amounts of a different components which are essential or beneficial for health including protein, vitamins, minerals, and phytochemicals. Wheat being a major source of cereal dietary fibre, its consumption reduces the risk of cardio-vascular disease, type 2 diabetes, and certain forms of cancer. Compound annual growth rate in production was much higher at national level with 2.07% as compared to 1.35% at global level; attributed to India’s positive growth in productivity (1.23%), followed by crop acreage (0.84%). The level of global and national wheat productivity is broadly similar, although global trends are slightly higher side on year-to-year basis, with the exception of 2000, 2002, 2012 and 2020 production years. It is important to state that in India, we cultivate only spring wheat genotypes with maturity duration of ~140 days as compared to the global scenario that includes a major portion of winter wheats (close to 300 plus days for maturity) and this clearly indicates that our per day productivity is much higher.
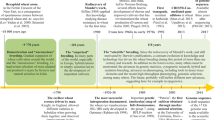
2.1.1 Importance of Crop and Progress Made in the Past 25 Years
The last 25 years of global and national trends on wheat area, production and productivity are analysed and presented in Table 2.1 and Fig. 2.1 (FAOSTAT 2020). In India, wheat area under cultivation increased by 6.35 million hectares (25.38%), i.e. from 25.01 million hectares during 1996 to 31.36 million hectares during 2020. Contrary to the national scenario, global wheat area reduced by 2.73 million hectares (−1.21%), i.e. from 224.58 million hectares to 221.85 million hectares during the same period. India’s wheat production increased from 62.10 million tons during 1996 to a record production of 107.86 million tons during 2020 with a quantum jump of 45.76 million tons. Similar trends at global level were also observed for the wheat production. However, India’s performance with respect to percentage change (73.69%) for production was more than double as compared to the world’s wheat production (34.13%). Our national wheat productivity increased from 2483 kg/ha during 1996 to 3498 kg/ha during 2020. A similar trend was observed for global wheat yield; however, again percentage change was much higher at national level (40.90%) compared to global (33.49%). Overall, during the past 25 years, wheat area increased at national level but it has marginally decreased at global level.

Trend in wheat area, production and productivity for India vis-à-vis the world
2.2 Origin, Evolution and Distribution of Species and Forms: Wild Relatives
Wheat is a classic example for understanding the evolutionary theory of allopolyploid, speciation, adaptation and domestication in plants (Gustafson et al. 2009). Origin and evolutionary pattern of wheat are presented in Fig. 2.2. The cytogenetic and genomic studies indicated that the mutation, polyploidy in the form of amphiploidy and inter-generic hybridisations are the major factors responsible for the evolution of present-day wheat. The wheat species can be broadly classified into three groups, namely diploid, tetraploid and hexaploid, based on the number of chromosome in the reproductive cell, i.e. n = 7, 14, 21, respectively. Cytogenetic studies revealed that four different genomes (ABD and G) representing four different sets of seven chromosome contributed to the origin of tetraploid and hexaploid wheats. The hexaploid wheat consists of two evolutionary lineages. The most known and grown T. aestivum (AABBDD) comprises one lineage while T. zhukovskyi (genome AAAAGG) comprises another lineage.

Origin and evolutionary pattern of wheat. Solid arrows indicate hybridisation followed by chromosome doubling. Dashed arrows indicate domestication or direct selection within a species. Boxes indicate cultivated taxa
The earliest cultivated forms were diploid (genome AA) and believed to be domesticated in south-eastern Turkey (Heun et al. 1997). During the process of domestication major emphasis was given on two traits, i.e. non-brittle rachis or tough rachis and the hull-less grains or free threshing grains. The source of A genome of tetraploid and hexaploid wheats was related to two diploid species, i.e. T. monococcum and T. urartu. Earlier studies by Kihara (1944) indicate that AA genomes were contributed by T. monococcum. However, on the basis of repeated nucleotide sequences in the phylogeny of polyploidy species of wheat, Dvorak et al. (1993) concluded that AA genomes in the tetraploid and hexaploid wheats are more related to AA genomes of T. urartu. The BB genomes of the tetraploid and hexaploid wheats are closely related to Sitopsis section of Aegilops with Ae. speltoides being the closest species. The SS genomes of Ae. speltoides is also closest to the GG genomes of T. timopheevi. The AABBDD genomes are thought to be arisen through hybridisation of T. turgidum with Aegilops tauschii. Petersen et al. (2006) re-sequenced two single copy nuclear genes DMC1 and EF-G isolated from each of the three genomes in hexaploid wheat (BBAADD) and two genomes of the tetraploid wheat (BBAA) using a sophisticated extension of PCR technique, and on the basis of phylogenetic analysis with diploid species, they suggested that the DD genomes of wheat was derived from Ae. taushcii. The D genome contributed significantly to the wheat flour properties that make bread wheat so valuable in bread making (Morris and Sears 1967).
The diploid wheats include two species, viz., T. monococcum and T. urartu. The sterility in the hybrids of monococcum and urartu (Johnson and Dhaliwal 1976) indicate that they are valid species. Reproductive barrier is not essential but one of the important criteria for recognising the species and natural hybridisation between diploid species is a rare phenomenon. The cultivated einkorn T. monococcum ssp. monococcum is the domesticated form of wild einkorn T. monococcum ssp. aegilopoides. The cultivated ssp. of monococcum differs from ssp. aegilopoides in having slightly large kernel and less brittle rachis. It is believed to be domesticated in ‘Fertile Crescent’ of Near East, which encompasses the eastern Mediterranean, south-eastern Turkey, northern and western Iraq and its neighbouring regions of Transcaucasia (Matsuoka 2011). The wild diploid species T. urartu also found in Fertile Crescent though it has never been domesticated. The tetraploid wheat (2n = 4x = 28) originated as a result of natural crossing between T. urartu and Aegilops speltoides. This resulted in origin of both the tetraploid species, namely T. turgidum (AABB genome) and T. timopheevi (AAGG genome).
The wild form of T. turgidum ssp. dicoccoides with brittle spike is reported from the Fertile Crescent. The non-brittle spikes with tough rachis among the wild form were selected by farmers. From there it spread to the other parts of world. Durum wheat is said to be derived from T. dicoccum (Damania 1998). The emmer wheat underwent further diversification in response to the agro-ecological conditions and gave rise to free threshing tetraploid wheats, namely rivet wheat (T. turgidum ssp. turgidum), polish wheat (T. turgidum ssp. polonicum) and khorasan wheat (T. turgidum ssp. turanicum). Interploidy introgression in hybrid swarms is thought to be contributed to further diversification of the turgidum wheat by giving rise to two sub-species T. turgidum ssp. paleocolchicum and T. turgidum ssp. carthlicum. McFadden and Sears (1946) created an artificial spelta (hexaploid wheat) from a hybrid of T. dicoccum and Ae. Tauschii, followed by chromosome doubling with colchicine. Therefore it was inferred that spontaneous hybridisation between emmer wheat (T. dicoccum) and goatgrass produce an early spelta (T. spelta 2n = 6x = 42 genome AABBDD). Later more easily free threshing bread wheat was evolved through natural mutation and selection. Hexaploid T. zhukovskyi originated recently by interspecific hybridisation of cultivated T. timopheevii with cultivated T. monococcum (Dvorak and Luo 2001).
2.3 Taxonomic Position of Genus Triticum
In botanical classification, wheat is a member of the grass family Poaceae (also called Graminae), the subfamily Pooideae and the tribe Triticeae. Beside wheat, many agricultural important crop species like barley, rye, and several forage species belongs to the tribe triticeae. The tribe is mainly distributed in the east Mediterranean and central Asiatic regions. Linnaeus (1753) describes five genera, namely Secale, Triticum, Hordeum, Aegilops and Elymus, in this tribe, whereas Clayton and Renvoize (1986) recognised 18 genera in the tribe triticeae. Later on van Slageren (1994) added Ambylopyrum genus in the tribe. All these genera were classified into two sub-tribes, namely Hordeineae (barley lineage with nine genera (Elymus, Hystrix, Sitanion, Leymus, Psathyrostachys, Hordelymus, Hordeum, Taeniatherum, Crithopsis)) and Triticineae (wheat lineage with nine genera (Agropyron, Eremopyrum, Heteranthellium, Secale, Dasypyrum, Triticum, Ambylopyrum, Aegilops, Henrardia)) (Feldman and Levy 2015). The genus Brachypodium was earlier included in this tribe but later on excluded from this tribe (Hasterok et al. 2004). Bowden (1959) advocated that there is no need to treat Aegilops as a separate genus from Triticum, however van Slageren (1994) considered Aegilops and Triticum are closely related but separate genera.
Several species of genus Secale and Aegilops serve as primary genepool based on crossability with Triticum species, while several species of genera Elymus, Leymus, Psathyrostachys, Dasypyrum are capable of being hybridised with wheat (Wang 2011), and considered as secondary and tertiary genepool of wheat. The taxonomy of genus Triticum is always debatable. In recent years, Mac Key (1966 and 1977) and Dorofeev et al. (1979) illustrated the system of classification of genus Triticum, while van Slageren (1994) revised the classification provided by Mac Key (1966). Goncharov (2011) reviewed the taxonomic history of genus Triticum and differences between various classifications of genus Triticum. van Slageren (1994) classified the genus Triticum into 3 sections and 6 species. These 6 species were further divided into 4 autonym and 13 non-typical subspecies, while Dorofeev et al. (1979) recognised 27 species in the genus Triticum (Table 2.2). van Slageren (1994) considered T. sinskajae a free threshing mutant of T. monococcum, T. jakubzineri a form of T. turgidum, T. militinae a free threshing mutant selected from single specimen of cultivated T. timopheevi, T. petropavlovskyi mutated form of T. polonicum, T. aethiopicum free threshing emmer selected from T. dicoccon, T. isphanicum as a form of T. polonicum due to more standard glume morphology. These were not considered as separate taxa at species or sub-species level by van Slageren (1994).
2.4 Spike Morphology and Pollination System
The wheat inflorescence commonly known as ear and in botany called as terminal distichous compound spike. In spike, the spikelets are arranged on the alternate side of the zig-zag rachis (Fig. 2.3). In a majority of tetraploid and hexaploid species of Triticum, the rachis is tough and resists disarticulation. However, T. dicoccoides and T. dicoccon disarticulate easily when mature. Each spikelet is subtended by two sterile or empty glumes. These glumes are arranged alternately on opposite side of the short central axis called rachilla. In a majority of Triticum species the glumes are shorter than the spikelet. The glumes are glabrous or pubescent. In some of the Triticum species, the glumes are keeled throughout, while in some species lower portion of glume is rounded. The shape of glume apex differs significantly in different species of Triticum. In T. aestivum, each spikelet consists of 3–6 florets attached alternately on rachilla. Each floret has its own lemma and two nerved palea. The tip of lemma may or may not be extended to awn. Based on the length of awn, the wheat is classified as long medium, short or awnless. The wheat flower is hypogynous and simple in structure consisting of whorl of three stamens and a single carpel. The uppermost floret of spikelet is imperfect (Percival 1921).

Wheat inflorescence. (a) Spike of wheat. (b) Spikelet of wheat ×4. (c) Floret of wheat ×8
Wheat is usually self-pollinated species and anthesis starts from the middle third of the spike and proceeds upwards rapidly and downwards little slower. In spikelet, the basal flower is first to open then the secondary florets (de Vries 1971). The receptivity of stigma depends on weather conditions. The anther dehiscence begins with elongation of filament and is completed after the anther is pushed out of floret due to lodicules swelling. Parts of the pollen fall on their own stigma and fertilise the ovary. The wheat pollen is short lived and the viability of pollen depends on the weather conditions. Generally, pollen remains viable upto 30 to 40 min after pollen shedding. The florets close again due to collapse of lodicules. In case ovary remains unfertilised, Okada et al. (2018) reported the second opening of flowers in wheat within a few days of post-anthesis. In the second opening there is a significant enlargement of ovary and forces the lemma end to be apart. Despite cleistogamous flower, less than 1% out crossing is reported in the wheat.
2.5 Gene Banks and Conservation of Genetic Resources
The key to success in any crop breeding programme depends on tapping the genetic variability of existing plant genetic resources in hybridisation programme. The extent of diversity present in the gene bank still remain untapped and may contain many useful traits that can be used in accelerating the rate of genetic gain in wheat breeding. The wheat genetic resources conserved in various gene banks also stored much useful information on passport and evaluation data. The gene bank serves as a focal point for providing information on plant genetic resources. The basic purpose of collating such information is to enable the plant breeders to make more thorough use of plant genetic resources. Significant progress has been made in various modern biotechnology tools such as recombinant DNA technology, cell biology and allied disciplines in recent years. Such tools should be targeted to mine genetic diversity as part of pre-breeding programmes for efficient integration with conventional breeding programme so as to achieve better and faster breeding outcomes.
Conservation of wheat genetic resources is a global concern. The in-situ conservation is one of the way of conservation of biodiversity, especially crop wild relatives. Damania (1996) advocated the need to safeguard the natural ecosystem as it is highly dynamic and understanding it’s components in short notice is unrealistic. Various countries, like Armenia, Russia, Syria, and Turkey, are involved in in-situ conservation of wild relatives of wheat crop (Meilleur and Hodgkin 2004). However, ex-situ conservation of wheat genetic resources had contributed a lot to the improvement of wheat yield, disease resistance and nutritional quality. Around 856,168 germplasm accessions are being conserved in various national and international organisations. Our global partner, CIMMYT has made significant contribution in conserving, improving and distributing wheat germplasm particularly to developing countries. CIMMYT’s wheat programme has a focus on development of new germplasm and their distribution in the form of various international trials and nurseries. In the gene bank of CIMMYT, around 110,281 accessions of wheat are conserved (Table 2.3). These are nearly 13% of total wheat germplasm conserved in various gene banks. The introduction of Norin 10 genes in the CIMMYT programme revolutionised the wheat production in many parts of the world. Pavon, Veery, Bobwhite, Attila and Kauz are some of the CIMMYT lines, which have been used extensively in the breeding programme. The National Small Grain Collection at USA also safeguards the important global collection. Over the years, India and China have also developed sound scientific management for ex situ conservation of wheat genetic resources.
2.6 Molecular Cytogenetics and Breeding
Wheat cytogenetics started with the discovery of chromosome number of durum wheat (2n = 28) by Karl Sax in 1918, followed by the reporting of chromosome number of eight wheat species, viz. T. monococcum (2n = 14), T. dicoccum (2n = 28), T. durum (2n = 28), T. turgidum (2n = 28), T. polonicum (2n = 28), T. spelta (2n = 42), T. compactum (2n = 42) and T. vulgare (2n = 42), with the basic chromosome number of x = 7 by Sakamura in Japan (Sakamura 1918). These chromosomes numbers of identified species were later confirmed by Kihara and Sax in 1919 and 1922. Kihara used genomic analysis to study the evolutionary relationship among different Triticum species as earlier successfully demonstrated in genus Drosera by Rosenberg (1909). Genomic analysis involves the crossing of allopolyploids with their presumed diploid ancestral species to study pairing pattern of their triploid hybrids. In the hybrids, if there is occurrence of basic number of bivalents, it is taken as a sign of genomic homology of the diploid parent with one of the genomes present in allopolyploid species.
Genomic homology becomes confusing when there is presence of higher order of pairing like trivalents, tetravalents and pentavalents, indicating some degree of differentiation of otherwise identical genomes. Kihara (1919) analysed chromosome pairing behaviour in the crosses between tetraploids and hexaploids, and observed 14II and 7I in the pollen mother cells. Similarly, Sax in 1922 observed 7II and 7I in the pollen mother cells of hybrids generated from tetraploid and diploid crosses. Whereas, in crosses of T. vulagre and Ae. cylindrica (CCDD) giving F1 hybrid having 7II + 21I (ABCDD) as well as in reciprocal crosses in the pollen mother cells as earlier reported by Kihara. Based on these studies as well as other studies as depicted in Fig. 2.4, it was concluded that the genus Triticum had three different genomes having 7 chromosomes each and these genomes were designated by Kihara as A, B, C and D. It has been concluded that the hexaploid wheat genomes A and D have been contributed from T. uratru and Ae. Taushii (not from Ae. Caudata and designated as D).

Genomic analysis based on pairing behaviour among diploid and tetra and allohexaploids as described by Kihara (1919)
The exact donor of B genome of present-day hexaploid and diploid bread wheat is still lacking, although many related diploid species exhibited partial homology and chromosome pairing but not to the extent observed for the A and D genome donors. Currently, Ae. speltoides (2n = 14, BB) species considered as the probable B genome donor (Dvorak et al. 1973). However, there are studies against the general acceptance including Kimber (1974), the study suggest that T. speltoides is probably homologous to the G genome of T. timopheevii and B genome donor to T. turgidum or T. aestivum is unrecognised. The C-banding technique helps in the identification of heterochromatin (dark-staining regions) and euchromatin (light-staining regions) on the chromosome axis. Within a homoeologous group, there was no similarity for the C-banding of chromosomes except for fifth group. C-banding comparisons of the diploid species T. monococcum, T. speltoides, and T. tauschii with that of the A, B, and D genomes, respectively, in hexaploid wheat confirmed that T. speltoides could not be the donor of the B genome to wheat and that T. monococcum and T. tauschii are the probable donors of the A and D genomes, respectively (for details see Gill and Kimber 1974).
2.6.1 Structure of Wheat Chromosomes
There are 21 pair of chromosomes identified cytologically having a primary constriction/centromere with secondary constrictions on two pairs of chromosomes indicating sites for ribosomal RNA genes. According to the observations of Sears (1954), the length of chromosomes at metaphase is on average 5.6 μm enclosed in the nucleus having a total volume of 1700 μm3 in which about 36.2 pg of DNA is accommodated. The total DNA length is about 11.2 m with a volume of 35 μm3 and is divided into 42 chromosomes. The DNA is coiled extensively and the first level of compaction is the nucleosome as revealed under electron microscope and by partially digestion of DNA with micrococcal nucleases. A haploid nuclear genome of hexaploid bread wheat has 16.72 × 109 bp (17.325 pg) of DNA, distributed on 42 chromosome 0.682 pg on 1DL to 2.475 pg on 3BL. The DNA contents in the A, B and D sub-genomes are found in the ratio 1.16:1.2:1.0 (Gupta 1991). It has been shown through reassociation kinetics that F75% of this DNA is repetitive (Mitra and Bhatia 1973).
Another 20% of DNA constitutes non-coding unique sequences. In addition, 30% of cytosine residues in this DNA are highly methylated regulating the expression of genes (Moore et al. 1993). Only 1% of this DNA is known to take part in protein synthesis; in this 1% DNA, F1, 000 genes have already been recognised, some of them representing multigene families (Lagudah et al. 2001; McIntosh et al. 1998). With the advancements in banding techniques, Sears (1954) identified individual wheat chromosomes through monatomic analysis and made some morphological observations, although cytologically most chromosomes were indistinguishable. Furthermore, Sears and Sears (1978) expedite the chromosome identification through isolation of marker telocentric chromosomes. Gill and Kimber (1974) identified individual wheat chromosomes, and wheat ideogram was constructed using C bands, which was facilitated by the telocentric chromosomes to recognise individual chromosomes.
2.6.1.1 Cytogenetic Stocks in Wheat
2.6.1.1.1 Wheat Aneuploids
Aneuploids in wheat have been developed by ER Sears, using cytogenetics techniques in the Chinese spring cultivars. These stocks are possible because wheat genome is polyploidy and can tolerate aneuploidy (Sears 1954, 1966a). During the 1930s, ER Sears was able to identify a few haploid plants, resulting from the cross between Chinese Spring with rye, which formed the basis of development of aneuploid stocks. In total, 220 lines were isolated, which comprised the stocks: 21 Monosomics (20″ + 1″); 7 Monoisosomic (20″ + i”); 21 Trisomics (20“+1’“); 38 Nullisomic–tetrasomic (20”+1″“); 4 Monosomic–tetrasomic (20“+1’+1’“‘); 8 Nullisomic (20”); 20 Double monosomic (19“+1’+1’); Ditelosomic (20”+t“), 21; 20 Double-ditelosomic (20”+tS” + tL”) and 41 Ditelo-monotelosomic (20″ + t” + t’). But the maintenance of these stocks is very difficult as it is necessary to characterise the progeny of each aneuploid for ascertaining the chromosome number.
These aneuploidy stocks have a great potential for localisation of genes to specific chromosomes/chromosomal arms (McIntosh 1988). A unique genetic system for production of deletion stocks systematically with variable sized terminal deletions in individual chromosome arms was reported by Endo (1988). He suggested that presence of certain chromosome from Aegilops cylindrica in Chinese spring in the monosomic condition, it induces chromosomal breaks in the gametes lacking A. Cylindrica chromosome and results in generation of various chromosomal aberrations, including deletions. The broken chromosome ends, if not fused to other broken ends, are stabilised by the rapid gain of telomere structure (Werner et al. 1992). Such deletions in plants without the A. cylindrica chromosome are transmitted regularly to the offspring, and they identified 436 deletions by C-banding.
2.6.1.1.2 Alien Addition and Substitution Lines
Development of alien addition and substitution lines started during the late 1950s by producing amphiploids through crosses between hexaploid wheat and any diploid alien species, whose chromosomes were to be added or substituted. An amphiploid having 2n = 56 was first crossed with hexaploid wheat, giving a heptaploid (2n = 49), which on selfing would produce progenies having different chromosome numbers including monosomic addition lines, which could then be selfed to obtain disomic alien addition lines. Such alien addition lines (including wheat-rye addition lines) were developed in the USA and Canada. These alien addition lines can be crossed with monosomics, and on selfing will give progenies having disomic substitution lines.
The aneuploid stocks offer great benefit as the cytogenetic markers identified on each of the 21 chromosomes will help in identification of each chromosome and chromosome arm. Because of the nulli-tetrasomic lines, Sears (1966b) was able to place the 21 wheat chromosomes into 3 genomes and 7 homeologous groups. For locating genes on to individual chromosomes, monosomic and telosomic have been used (McIntosh et al. 1995; Sears 1966c). These deletion stocks have been used to place the EST markers onto specific deletion bins for precisely map genes of interest (Qi et al. 2004). In addition, the deletion stocks were crucial in relating genetic maps to physical maps of chromosomes, map-based cloning of genes (Simons et al. 2006; Yan et al. 2003), and studying the distribution of genes (Gill et al. 1996) and recombination frequency along the chromosomes (Akhunov et al. 2003). The whole genome sequencing project of wheat has been made possible by the use of the ditelosomic stocks for isolating individual chromosome arm through flow sorting for the construction of arm-specific BAC libraries (Safar et al. 2004).
2.6.1.1.3 Diploidising System
Bread wheat contains three closely related genomes therefore; loss of any chromosome of a homeologous group can be compensated by the homeologous group chromosomes because of which monosomics have been possibly developed. Despite closely related chromosomes, homeologous group chromosomes strictly follow diploid type behaviour and form 21 bivalents. The genetic control of chromosome pairing in wheat is dependent on a series of promoters and suppressing pairing homologues genes (Sears 1976). The strongest effect on pairing is associated with a gene on chromosome 5BL known as Ph1 locus, which prevents homoeologous chromosomes to pair at meiosis. Apart from Ph1 locus, two more suppressors have been identified Ph2 on 3DS and another suppressor on chromosome 3AS. This feature of Ph1 locus was utilised for transfer of desirable genes from alien species to wheat, because in wheat plants deficient for 5B or those carrying a mutation for Ph1 locus (Sears 1977) pairing between wheat chromosomes and the corresponding homoeologous alien chromosomes would occur. This will allow transfer of alien genes on to wheat chromosomes through recombination.
2.6.1.1.4 Alien Introgressions
Development of new plant varieties requires genetic variability, which can be created by crossing the available germplasm in the primary gene pool. Modern plant breeding, although increased crop productivity world-wide, however, it also eroded the genetic variability of the crops (Gill et al. 2011; Hoisington et al. 1999). Alien introgression of different gene(s) for tolerance to biotic/abiotic stress tolerance in wheat is presented in Table 2.4. To introduce new variability, use of secondary and tertiary gene pools is required and is generally referred as ‘wide hybridisation’. Many wheat varieties have been released around the world carrying alien chromosomal introgression from related wild species. About 110 leaf rust (Lr), 86 stem rust (Sr), and 83 stripe rust (Yr) resistance genes have been identified in wheat or wild relatives, most conferring race-specific resistance to these rust pathogens (Bhatta et al. 2019; Cox et al. 1992; Huerta-Espino et al. 2011; Ram et al. 2005; Singh et al. 2015).
Using ionising radiation, Sears (1956) transferred a leaf rust resistance gene from Aegilops umbellulata to chromosome 6B of bread wheat. The classical example of alien introgression T1BL·1RS resulted from the breakage of wheat chromosome 1BL·1BS at the centromere, and the 1BS arm of wheat was replaced by the 1RS arm of rye. This introgressed chromosomal segment carried group of resistance genes to leaf rust (Lr26), stem rust (Sr31), stripe rust (Yr9), powdery mildew (Pm8) (Friebe et al. 1996), along with robust drought-tolerant root system (Sharma et al. 2011). Similarly, powdery mildew resistance gene Pm 21 (T6AL·6VS) was introgressed from Dasyprum villosum (L.) Candargy had other important genes for resistance to wheat curl mite, stripe rust and Fusarium head scab (De Pace et al. 2011). Transfer of Ae. Umbellulata-derived leaf and stripe rust resistance to hexaploid wheat has been demonstrated by Chhuneja et al. (2008) and Bansal et al. (2017). Direct cross between wheat and Ae. kotschyi have been used to develop amphiploids with high grain iron and zinc and flag leaf iron and zinc concentrations as that of the Ae. kotschyi parent (Rawat et al. 2009). CIMMYT had developed about 1300 primary SHW, and have been confirmed to possess valuable traits for better performance under biotic and abiotic stresses, along with yield potential (Mujeeb-Kazi et al. 2008; Yumurtaci 2015). Dyck and Kerber (1970) transferred first leaf rust resistance gene, Lr21, from Ae. squarrosa var. strangulata ‘R.L.5271’ to Canthatch and Thatcher wheat cultivars through direct hybridisation. Lr42 has been one of the most effective Lr genes introduced from Ae. tauschii accession TA2450 for further utilisation in hexaploid wheat breeding (Gill et al. 2019).
2.7 Genetic Studies of Qualitative and Quantitative Traits
Improvement in any crop requires the presence of genetic variation and also the exploitation/manipulation of that variation for developing improved genotypes. Selection is generally practised for selecting desirable plants within the population, which is based only on identification phenotypically superior plants. Phenotype is composed of both heritable and non-heritable variation. The heritable component is governed by the gene(s) present in the genome of the plant, whereas the unexplained variation (environmental influence) affecting the expression of trait is non-heritable. In order to select the plants, understanding of fundamental laws of inheritance and genetics of different traits is necessary. Some characters are governed by one or few genes and are also less influenced by environmental variation and are referred as oligogenic/qualitative traits. These traits produce specific effect, and population can be classified into distinct classes. Some traits are controlled by several genes having small effect are referred to as polygenic/quantitative traits. These traits are largely affected by environmental factors, and population shows continuous variation and cannot be classified into distinct classes as in case of oligogenic traits. The earliest examples of genetics studies in wheat for presence of awns and glume hairiness were reported to be governed by genes, which were monogenically inherited.
However later studies indicated more complex inheritance. Based on chromosome substitution lines genes conditioning earliness were reported to be present on seven chromosomes (Kuspira and Unrau 1957). Similarly, genetics have been worked out by different workers for different wheat traits, like biotic stresses (rusts, powdery mildew, Karnal bunt, Spot blotch, Tan spot, etc.), abiotic stresses (heat, drought, waterlogging, salinity, PHST, etc.), agro-morphological traits, viz. earliness, tillering, lodging, plant height, spike density, 1000-kernel weight, grain number and yield using conventional inheritance studies as well as using cytogenetic stocks to identify chromosomal regions governing these traits. The qualitative inheritance have been reported mainly for biotic stresses, particularly rust and powdery mildew resistance, whereas the quantitative inheritance have been reported for spot blotch, Karnal bunt, Septoria, head scab, heat and drought tolerance and most of the yield contributing traits. Grain yield being a complex trait generally associated with different yield contributing traits, which are also polygenically inherited. Selection for oligogenic traits can be practised early in generation as they are having high heritability. The quantitative traits like grain yield are having low heritability and are also influenced by environmental factors, so selection will be better if practised later in generations as it gives opportunity for combing several component traits to be selected upon. Therefore knowledge of trait genetics is of utmost importance for wheat breeding.
2.8 Breeding Objectives
2.8.1 Grain Yield: The Ultimate Aim
Breeding for higher yield is one of the primary objectives of wheat improvement to meet the ever increasing demand for wheat-based diets across the globe. Breeders of autogenous crops aim to develop inbred varieties through selection and accumulation of desirable alleles with additive effects on desirable traits, including yield. During the initial 3 decades of green revolution, the global wheat yield was around 3% per annum, whereas, the following two decades witnessed the growth of merely 1.4% (FAO 2020). The quantum jump in wheat yields across the globe started in the 1960s, with the continuous effort of Nobel Laureate Norman E. Borlaug with his crossing programme including Norin-10/Brevor cross that introduced the Rht-B1 and Rht-D1 dwarfing alleles, which led to the development and release of input responsive semi-dwarf varieties led to Green Revolution.
Historically, chromosomal translocation of 1RS-1BL between wheat and rye was one of the most landmark introgression till date in wheat improvement, which increased the wheat yield potential and tolerance to biotic and abiotic stresses. This landmark segment is still present in many of the important wheat cultivars, currently cultivated across the globe (Schlegel 1997). The contribution of E.R. Sears deserves a special mention for his great contribution to this field. Development of synthetic wheat is another important strategy by repeating the interspecific crosses that occurred in nature that led to the formation of hexaploid wheat. Different accessions of the hexaploid wheat progenitors T. monococcum, T. turgidum, and Ae. tauschii used for the formation of new genetic constitutions of wheat, greatly increasing the genetic variability of the primary gene pool (Mujeeb-Kazi et al. 2008). Numerous synthetic wheat germplasm pools have been developed by CIMMYT, Mexico.
2.8.1.1 Trait-based Breeding to Increase the Yield Gain
Selection of wheat genotypes based on yield components will help in the cultivar improvement along with yield per se selection, since yield is a complex trait and influenced by several component traits. Introgression of multiple traits for better agronomic and physiological performance into a single variety could help in the genetic progress of grain yield. Some of the yield components that have been successfully utilised for cultivar development with improved grain yield in different wheat breeding programmes are discussed below.
2.8.1.1.1 Early Flowering and Maturity
Development of new high yielding wheat cultivars having early flowering and maturity is one of the important breeding objectives. The main focus of breeding programmes is to develop genotypes, which mature early as an adaptive mechanism for terminal heat and drought stress experiencing environments. Most of the present wheat cultivars incorporated vernalisation and photoperiod insensitive genes to promote early flowering and maturity (Chen et al. 2016). Vernalisation genes Vrn-A1, Vrn-B1, and Vrn-D1 control flowering and maturity in wheat. Wheat workers developed cultivars combining vernalisation to promote early maturity and improve grain yield potential. Photoperiod sensitive genes including Ppd-D1a, Ppd-B1, and Ppd-A1 control photoperiod sensitivity, regulating flowering and maturation times in wheat (Gomez et al. 2014). High yielding with early maturing genotypes suitable for diverse growing conditions have been developed in different breeding programmes across the globe by incorporating the vernalisation, photoperiod, and dwarfing genes (Royo et al. 2018). By articulating wheat phenology, breeding for high yielding and early-maturing wheat genotypes can be achieved due to negative correlations between flowering and yield (Mondal et al. 2016). Such cultivars should have faster growth rates to accumulate sufficient biomass in shorter duration to increase grain yield potential.
2.8.1.1.2 Plant Height
Global wheat productivity has significantly increased due to the development of semi-dwarf wheat cultivars. Dwarfing/height reduction genes like Rht1 (Rht-B1b), Rht2 (Rht-D1b), Rht-D1c, and Rht8 have been extensively utilised for cultivar development by many breeding programmes across the globe (Zhang et al. 2016). Dwarfing genes reduce the coleoptile and internode length resulting in the reduction of plant height, thereby increasing assimilate partitioning to the ear, which results in higher HI and lodging resistance. Although numerous dwarfing genes are available, only a few genes have been extensively utilised for wheat yield improvement (Chen and Hao 2015). Therefore, integration of less explored dwarfing genes, such as Rht4, Rht5, Rht11, Rht12, and Rht24 with the commonly used dwarfing genes (i.e. Rht1, Rht2, Rht8), would further help in improving the yield and lodging resistance (Rebetzke et al. 2012a,b). Some studies including Shearman et al. (2005) suggested that the plant height has reached its theoretical limit of around 70–80 cm in wheat, indicating that little progress will be achieved through further reduction in plant height. Further reduction of plant height will have effect on biomass and yield (Berry et al. 2015). Therefore, strategic breeding that combines both plant height and grain yield to maximise yield potential and lodging resistance has been suggested (Gao et al. 2017).
2.8.1.1.3 Harvest Index
Harvest index (HI) is one of the important physiological traits; it has considerable effect on grain yield. Despite significant improvement in HI, the trait has not been exploited to its full potential, and the trait has remained at approximately 0.55, which is below a theoretical limit of 0.62 (Gaju et al. 2009). Positive and linear relationship was observed between HI with grain yield over time suggesting that HI can improve yield gains even further (Zheng et al. 2011).
2.8.1.1.4 Biomass
Total biomass has a definitive role in wheat grain yield improvement. Physiologically, increase in biomass has been largely attributed to higher photosynthetic rate, stomatal conductance, leaf chlorophyll content and improved radiation-use efficiency (Bustos et al. 2013). It has been suggested that further improvements in grain yield can be achieved by increasing photosynthetic capacity by optimising biomass production, while maintaining lodging resistance (Beche et al. 2014). They suggested that by increasing photosynthetic capacity through optimisation of biomass production, while maintaining lodging resistance will further improve the grain yield.
2.8.1.1.5 Kernel Weight
Grain yield improvement has been associated with increased thousand kernel weight (TKW). TKW is reported nearly linear with moderate to high correlation with grain yield (Gao et al. 2017), suggesting selection of bolder grains could be highly effective for improving wheat yield. As a result, increasing grain weight potential at specific positions within the spikelet has been suggested (Calderini and Reynolds 2000), rather than breeding for higher TKW.
2.8.2 Quality
There are about 46 technological parameters that can be used to judge the quality of wheat grain and its physical and chemical constituents. Among the physical characters, virtuousness, kernel hardness and hectolitre or test weight are the most studied. Among the chemical parameters, starch and proteins are most predominant constituents of wheat grain, which play a vital role in determining its quality. Mainly two approaches are being followed in various breeding programmes across the globe to improve the wheat quality, i.e. testing of fixed lines at F6/F7 generations or testing from F2 onwards. In most of the breeding programmes, testing of fixed lines for various quality parameters is the common practice due to various practical challenges. But for maximum genetic gain selection, easily measurable quality traits like sedimentation value, protein content, iron and zinc are preferable at early generation breeding material. Since age old, the main focus of breeding programmes across the globe is to enhance the productivity per se to feed the increasing the population. After reaching the self-sufficiency of food grains, now breeding for better quality is one of the prime research areas across the globe. Breeding for quality is a tedious, cost intensive and time consuming process, which makes breeding for quality a slow and protracted. Wheat quality has historically been the last to utilise new technology for breeding.
2.8.2.1 Genetic Resources: A Valuable Donor for Quality Improvement
Genetic resources including wild relatives, synthetic wheats and landraces have been reported to contribute to enhance wheat grain quality (Ogbonnaya et al. 2013). Enormous diversity associated with landraces makes them a good source of bread-making related alleles such as wbm (Sanchez-Garcia et al. 2015). Landraces are one of the most important sources of wheat biofortification, as a collection of Sicilian landraces found to contain high levels of micronutrients (Rasheed et al. 2019). Conventional breeding approaches have been successfully used to incorporate several novel alleles for grain zinc content into elite breeding material by crossing high yielding elite wheat lines with Ae. tauschii-based synthetic hexaploid wheats or T. spelta accessions (Velu et al. 2018). Pre-breeding is an important approach in plant breeding and is practiced when desired variations are exhausted in the routine germplasm lines and cultivars. It is an important step wherein one harnesses diversity arising from wild relatives/wild germplasm/unexploited or other unimproved materials. It refers to all activities designed to identify desirable traits and/or genes from unadapted germplasm or materials that cannot be used directly in breeding programme. These traits can be bred to an intermediate set of materials that can be further utilised for producing new varieties for farmers.
Promising sources of high Zn and Fe in wheat are wild emmer (T. dicoccoides), einkorn (T. monococcum), diploid progenitors of hexaploid wheat (such as Aegilops tauschii), T. spelta, T. polonicum, and landraces of T. aestivum. Genetic variability in cultivated hexaploid and tetraploid wheat is low, but wild and primitive wheats are promising genetic resources. Fe and Zn content in wild relatives were 50% higher than in the modern cultivated wheat, and the highest concentrations were upto twice than that of modern cultivars. Among wild wheats, the collections of wild emmer wheat, Triticum turgidum ssp. dicoccoides, showed greater genetic variation with as high as 14 to 190 mg/kg of grain Zn concentrations (Cakmak et al. 2004). In a study consisting of different recombinant substitution lines derived from Triticum dicoccoides, a Gpc-B1 locus has been identified on the short arm of 6B chromosome and that locus affects both wheat grain protein and Zn concentrations (Distelfeld et al. 2007). The grain Fe and Zn concentration of Aegilops kotschyi and A. tauschii showed that the S and D genome species accumulate significantly higher Fe and Zn than the cultivated wheats.
2.8.2.2 Genetic Improvement of Grain Industrial Quality
The definition of wheat quality varies across stakeholders in the wheat chain. The grading system to classify wheat grain into different classes varies considerably among countries. End product appeal and organoleptic properties are the most important attribute of wheat quality for small and marginal farmers with subsistence farming practices as they generally mill and process the wheat grain to feed their families. On the other hand, commercial farmers consider wheat quality characteristics that allow the sale of their grain at the best possible price. Miller’s primary interest is the higher flour/semolina recovery of suitable quality with low milling cost. Therefore, grain size, density, hardness, and roundness are the most important traits for miller’s quality (Edwards et al. 2010). Millers are also interested in the production of suitable flour for processing industry (the ability of a flour or semolina to be processed at minimum cost and to give a uniform product). To produce the flour with the desired characteristics for processing industry, very often millers blend and combine different types of grain differing in quality traits.
Grain hardness, gluten extensibility and strength are the most important for processing industry. Grain hardness and dough visco-elastic properties to be considered while breeding wheat genotypes for industrial grain quality, which includes milling, processing and end-use quality. At global market, generally wheat grains are classified as soft, hard or very hard (durum wheat), sometimes locally as medium hard wheat. Genetically, presence of two small proteins called puroindolines encoded by the Pina-D1 and Pinb-D1genes determines the grain hardness. Presence of both the puroindolines makes the grain texture soft, but if either one is mutated or altered then it makes the grain texture hard. Durum wheat is hard as both the puroindolines are absent due to the lack of the D genome durum wheat species. Grain hardness affects the water absorption, flour particle size, and milling process and partly defines end-use quality like hard grain flour for bread and soft grain flour for cookies and pastries. In contrast, gluten content and composition defines the dough visco-elastic properties including gluten extensibility and elasticity.
Gluten is mainly formed by the glutenins and gliadines, further glutenins are divided into high and low molecular weight. Generally, glutenins are associated with gluten strength or elasticity and gliadins with viscosity and extensibility. The most important genes controlling these proteins are Glu-1, Glu-3 and Gli-2, and it is well known that their different alleles have been associated to gluten quality characteristics. Other less explored but important quality traits such as starch properties and enzymatic activities are in their infancy to utilise them in breeding programmes. Therefore, breeding programmes need to develop a holistic breeding approach, to ensure suitable gluten quality with diverse levels of gluten strength combined with the required extensibility along with medium protein content in hard to semi-hard grain texture genotypes. On the other hand, the overall strategy for durum wheat has been to develop genotypes with suitable quality to be accepted by the pasta-making industry across the globe. Durum wheat genotypes with large kernel size, medium to low gluten strength, and with high yellow pigment content are preferred.
2.8.2.3 Genetic Improvement of Nutritional Quality
Nutritional quality can be defined as the ability of a food to provide enough nutrients for a correct physical and mental development for a healthy life of human beings (Guzman et al. 2019). Recently, development of nutri-dense crops is one of the important research priorities and breeding objectives, particularly for the staple crops including wheat that represent a major proportion of the food and calories in developing and under developed countries. Wheat is a potential source of different micronutrients and other bioactive components, but the levels of some of these are not high enough to meet the daily requirements of people in countries where wheat represents the main source of calories.
2.8.2.4 Biofortification: A Promising Strategy to Contain Malnutrition
Nutrition (protein, vitamins and minerals) deficiency is one of the most important public health issues across the globe, particularly in developing and underdeveloped countries. Biofortification is a process to improve the nutritional value of crop plants through plant breeding, agronomic and transgenic approaches. Consumption of biofortified staple crops will help in the alleviation of malnutrition, thereby it improves the human health condition. Presently, development of nutrient dense staple food crops is one of the prime research areas for the scientific community. Biofortification has been recognised as an economical and sustainable strategy that can be useful as a complementary solution to the problem of malnutrition. Staple crops often exhibit genetic variation in essential nutrient contents, which enable breeders to develop nutrient dense high yielding genotypes through conventional or molecular breeding approaches. Development of nutrient dense crops through conventional/molecular breeding approaches is both economical and does not have any effect on consumer acceptance, unlike transgenic approaches.
With the aim of developing bread wheat cultivars with 40% higher Zn concentration over the current commercial cultivars in the target regions of South Asia, CIMMYT is leading the partnership-based global effort within the HarvestPlus project (Velu et al. 2011). Millions of resource-poor wheat consumers in South Asia and Africa are prone to Zn deficiency, making Zinc to be the primary target nutrient for wheat under the HarvestPlus project. Large genetic variability has been identified for zinc in the wheat genetic resources, which enables to develop high yielding wheat cultivars with elevated levels of zinc through various breeding approaches. Currently, CIMMYT breeding programme mainly focused on transferring genes governing higher Fe and Zn from Triticum dicoccon and Triticum spelta-based synthetics, landraces, and others reported high Zn and Fe sources in to the high yielding elite wheat backgrounds.
Synthetic hexaploids and other donor parents with significantly higher Fe and Zn concentrations were used as donor parents for a limited-backcross breeding approach onto adapted CIMMYT wheat parents (Velu et al. 2011). Energy-dispersive X-ray fluorescence spectrometry is used for rapid estimation of grain iron and zinc centration in biofortification programmes (Paltridge et al. 2012). Five biofortified wheat varieties have been released in different countries with elevated levels of Zn (Bari Gom 33 in Bangladesh, Zinc Shakti (Chitra), WB02 and HPBW-01 in India, and Zincol 2016 in Pakistan (Velu et al. 2015). In India, a project entitled Consortia for Research Platform (CRP) in ‘Biofortification in wheat for nutritional security’ is undergoing under the umbrella of ICAR.
2.8.3 Biotic Stresses
2.8.3.1 Rusts
Historically, the rust diseases have been one of the major biotic production constraints for wheat production. Globally, yellow rust (Puccinia striiformis f. sp. tritici), stem rust (Puccinia graminis f. sp. tritici), and leaf rust (Puccinia triticina) are the most damaging diseases of wheat (Roelfs et al. 1992). Historically, yellow rust has caused and is presently causing significant and severe losses on susceptible wheat cultivars worldwide. Moreover, detection of the widely virulent race Ug99 in Uganda in 1998 challenged the misconception that stem rust was a conquered disease. Now, upto 90% of world’s wheat cultivars are considered stem rust susceptible (Singh et al. 2006), and the disease is threatening 20% of the world’s wheat in Central and North Africa, the Middle East and Asia.
Leaf rust of wheat, caused by the fungus Puccini tritici, can cause heavy yield losses in wheat. Symptoms are small, round-to-oval, raised, orange-red, dusty pustules that are scattered mostly on the upper surface of the leaves and leaf sheaths of infected plants. Leaf rust frequently starts on the lower leaves and gradually progresses up the plant to the flag leaf. As the season progresses, the pustules become more and more numerous until more of the total leaf area is destroyed. Such severely infected leaves usually shrivel and die prematurely. Leaf rust decreases the yield, grain quality and forage value, and in most wheat producing regions the use of susceptible cultivars has resulted in yield losses of 10%–70%.
Stripe rust caused by Puccinia striiformis f. sp. Tritici continues to be a dominant factor, limiting yield potential in wheat. Pustules coalesce to produce long yellow stripes between veins of the leaf and sheath and so are also known as yellow rust. Small yellow, linear lesions occur on floral bracts. These pustules are filled with spores of the fungus. The stem rust disease caused by fungus Puccinia graminis f. sp. Tritici, appears as elongate blister like pustules, or uredinia, most frequently on the leaf sheaths of the wheat plant, but also on stem tissues, leaves, glumes and awns. Stem rust pustules on leaves develop mostly on the lower side, but may penetrate and make limited sporulation on the upper side. As infected plants mature, uredinia change into telia, altering colour from red into dark brown to black, thus the disease is also called black rust. Severe infection of stems interrupts nutrient flow to the developing heads, resulting in shrivelled grains and stems weakened by rust infection are prone to lodging (Roelfs et al. 1992). Severe wheat yield losses due to stem rust ranged from 9 to 33% in Scandinavia in 1951 and 5–20% in eastern and central Europe in 1932 (Zadoks 1961).
2.8.3.2 Powdery Mildew
Powdery mildew of wheat is caused by an obligate, biotrophic ascomycetous fungus Blumeria graminis sp. Tritici during the late winter and early spring. Powdery mildew is a wind-borne disease favoured by the presence of disease in the preceding season. Disease infection can start during early crop growth when conditions are cool and wet. As the temperature rises and the humidity falls, the incidence and severity tend to diminish. The disease is preferred by mild temperatures (10–22 °C), and 100% relative humidity (RH) favours the conidium germination. Prolonged cloudy weather fastens the disease development. During winter, spores survive in the host tissue after infection and may come from earlier infections within the field or from fields farther away. The disease is most common in dense early sown crops with high nitrogen fertility and rapid plant growth. Cultivation of disease-resistant cultivars/varieties is an efficient method for commercial breeding and disease control by the introgression of resistance genes which enhance the durability of the variety. Host resistance is more likely to be durable when two or more resistance genes are pyramided in a single wheat variety. Information about the genetic diversity and distribution of Pm genes in a set of wheat varieties is required for the pyramiding of resistance genes. Until now, nearly 73 Pm genes/alleles have been identified in 60 loci from common wheat and its wild relatives.
2.8.3.3 Loose Smut, Karnal Bunt, Common Bunt and Tan Spot
Loose smut (Ustilago tritici) is a disease that affects wheat all over the world. This seed borne fungus survives from crop to crop in the embryo and is disseminated with the grain, making it spread easily but difficult to control. The fungus is protected within the seed and grows with the growing point of the wheat plant, therefore not only is it protected from being cleaned from the seed, it is also protected from contact (protectant) fungicides. Karnal bunt (KB) is a fungal disease caused by Tilletia indica (Mitra). The incidence of KB varies considerably from year to year, depending on the variability in favourable weather during heading stage of wheat crop. The importance of KB lies in the fact that the disease is prevalent only in a few countries around the world, and the pathogen being seed, soil and airborne is difficult to manage once introduced in an area. Therefore, wheat importing countries have imposed strict quarantine measures and insist on zero tolerance limits on shipment of wheat from KB prone regions.
Common bunt (CB) caused by Tilletia caries, is the world’s most destructive wheat disease. Instead of spikes filled with healthy wheat seeds present at crop maturity, the fungus produces kernels filled with bunt balls of spores that possess a fishy odour. The bunt spores then adhere to healthy wheat seeds during harvest. Tan spot is a foliar spotting disease caused by the fungus Pyrenophora tritici repentis also known as yellow spot or yellow leaf blotch and occurs in all the major wheat-growing areas worldwide. Intensified wheat production, changes in cultural practices involving adoption of conservation agriculture practices, including shifts from conventional tillage and stubble burning to reduced or zero tillage practices with residue retention, and shorter crop rotations contributed to the development of tan spot in epidemic proportions worldwide.
2.8.3.4 Head Blight/Head Scab
Fusarium head blight (FHB), also called ear blight or head scab, is caused by Fusarium spp. and is one of the most destructive diseases of wheat (Triticum aestivum). FHB occurs primarily in warm and humid climatic conditions during the flowering stage. The most common species causing FHB is Fusarium graminearum (sexual stage Gibberella zeae). This fungus is also associated with stalk rot of corn. Another Fusarium species that causes FHB is Fusarium culmorum.
2.8.3.5 Septoria Blotch
Septoria of wheat is a disease complex caused by three pathogens: Mycosphaerella graminicola, Phaeosphaeria nodorum and P. avenaria triticae. Nowadays the name for the disease caused by Mycosphaerella graminicola usually is referred to as Septoria tritici blotch (STB) or simply Septoria blotch, while that caused by P. avenaria and P. nodorum is called Stagonospora blotch. Losses to Stagonospora blotch caused by S. nodorum typically are much lower, but can reach 15% or more (King et al. 1983). Both pathogens can reduce grain test weight in addition to yield, and also reduce the quality of the grain produced. Losses to STB can range from 30 to 50% during severe epidemics (Eyal and Levy 1987) but typically are much lower. Epidemics of STB are most severe in areas with extended periods of cool, wet weather, particularly northern North America, northern Europe and areas with a Mediterranean climate such as North Africa, South Africa, parts of South America and western North America.
2.8.3.6 Viral Diseases
The economically most important wheat infecting viruses are the soil-borne viruses either belonging to the genus Furovirus (family Potyviridae), i.e. soil-borne wheat mosaic virus (SBWMV) and soil-borne cereal mosaic virus (SBCMV), or belonging to the genus Bymovirus, i.e. wheat spindle streak mosaic virus (WSSMV) and wheat yellow mosaic virus (WYMV). All these viruses are transmitted by the soil-borne plasmodiophorid Polymyxa graminis Ledingham.
2.8.3.7 Nematode Disease
Plant parasitic nematodes are recognised as one of the major constraints in wheat production with nearly 17 important nematode species, the majority of them belonging to three genera, namely Heterodera, Pratylenchus and Meloidogyne.
2.8.3.8 Wheat Blast: A New Threat
Wheat blast, caused by Magnaporthe oryzae pathotype triticum (MoT), was discovered in Parana state of Brazil in 1985, and since then spreading to an area of about 3.0 mha, causing losses of 10–100% depending on years, genotypes, planting date and environment. Later the wheat blast disease reported in central and south low-lying areas of Santa Cruz region of Bolivia, south and south-east Paraguay, and north east Argentina. Resistance genes for wheat blast disease are presented in Table 2.5. Most recently it was observed in Bangladesh in 2016, eight districts with upto 25–30% yield loss. Wheat blast is a seed and airborne disease, till date resistance sources are identified but there was limited knowledge on its genetics available. MoT population is very diverse and exhibits many pathotypes that could cross-infect different hosts and overcome resistance. Fungicide schemes are partially effective under low to medium WB pressure. Pathogen has the ability to develop fungicide resistance. The wheat blast pathogen an attack any aerial parts of wheat plant but disease is seen mainly on spikes. The pathogen spread is favoured by warm and humid weather due to rainy days and temperature of 18–25 °C, during flowering followed by hot, sunny and humid days. Minimum temperature for the infection is 10 °C and maximum is 32 °C with optimum between 25 and 30 °C.
India has proactively been involved in stopping the spread of wheat blast further from the neighbouring country Bangladesh. After the Bangladesh outbreak, India, the world’s second largest wheat producer, all of sudden became vulnerable to WB although as of the cropping season pertaining to the year 2020, this deadly disease has been successfully averted to gain an entry into the country (Goddard et al. 2020). The major wheat growing regions of North Eastern Plain Zone (NEPZ) and the Central Zone (CZ) could probably be vulnerable based on pathogen climate requirements. Even the country’s main wheat-producing region, i.e. North Western Plain Zone (NWPZ), is also vulnerable to blast winters and becomes humid and warm (Cardoso et al. 2008). The Government of India (GOI) enforced wheat holidays in Nadia and Murshidabad districts of West Bengal and created wheat-free zones of 5 kilometres from the Bangladesh border. ICAR-Indian Institute of Wheat and Barley Research conducts regular survey and surveillance for monitoring and mitigating the blast threat. Blast-resistant varieties like DBW 187, HD 3249 and DBW 352 were recommended to grow in the disease-prone areas of NEPZ.
2.8.4 Abiotic Stress
Abiotic factors are one of the major yield-limiting factors for crop plants including wheat. Temperature extremes, drought, flooding, salinity and heavy metal stress, among others, affect the growth and yield of crop plants. Abiotic stresses at a given growth phase are likely to affect the organs development and so yield components set at that phase, leading to a reduction in yield potential. Drought and heat are the two major abiotic factors limiting the wheat crop productivity; the problem is further complexed by climate change. Wheat is particularly vulnerable to high temperatures and every single degree raise in average temperature during reproductive period may lead to significant yield losses (Yu et al. 2014).
Rising of every 1–2 °C temperature reduces the time taken for grain filling and also affects the survivability of the productive tillers around 15.38%, which ultimately affects the grain yield (53.57%) (Nahar et al. 2010). Even a short spell of heat waves during grain filling may result in substantial grain yield loss (Mason et al. 2010). Although frost injury to wheat is relatively less compared to heat stress situations, mountainous and sub-mountainous region are experiencing cold or frost injury due to sudden temperature fall. Sodicity or salinity is another abiotic stress which significantly affects crop growth and yield. Globally one-fifth of the irrigated land and 2% of the dry land agriculture is affected by salt stress. In India, about 6.73 mha land is salt affected, out of which a 3.77 mha is sodic while the remaining 2.96 mha is under salinity affected (Mythili and Goedecke 2016).
Another major and highly unpredictable abiotic stress is lodging. Inadequate root anchorage, poor stem structure and strength, and adverse weather disturbances like excess wind velocity, rain, hailstorm along with topography, soil type, crop management practices and disease collectively may result in lodging (Mulsanti et al. 2018). It is a global problem as many parts of the world have been recorded with significant percent of yield reduction caused due to crop lodging. Along with these, other abiotic stresses like pre-harvest sprouting and water logging are further exacerbating the yield loss in wheat (Abhinandan et al. 2018).
Breeding for abiotic stress prone environments has been the major focus area from decades, but the advancements are far below the expectation due to its complex nature. Advancement in developing stress tolerant germplasm relies heavily on the efficient breeding programmes and phenotyping approaches. Phenotyping includes identification, induction and categorisation of desired target environment, stress management and complete characterisation of experimental material. Phenotyping is mainly required to understand the complexity of genotype-phenotype interaction and to accelerate plant breeding through deeper understanding of plant phenology and physiology. Recent scenario of the agricultural research strongly favours the adoption of a ‘trait-based’ crop improvement approach for increasing productivity under changing climatic conditions. Identification and selection of right traits allow the plants to uptake more resources under stress condition and also to use them more efficiently. Physiological traits linked to abiotic stress adaptation are the best available opportunities for genetic improvement of wheat, as they involve a combination of favourable alleles (Reynolds et al. 2009).
2.8.4.1 Role of Phytohormones Under Abiotic Stress
Hormones play an important role in plants adaptation to adverse environmental conditions. Cross-talk in hormone signalling reflects plant ability to integrate different inputs and respond appropriately. There are six main groups of hormones, namely auxin, cytokinin(CK), gibberellic acid (GA), abscisic acid(ABA), ethylene and brassinosteroids. Among all plant hormones ABA is most critical and hence termed as ‘stress hormone’. Stress-induced senescence and abscission are the characteristic features of ABA. Under water-deficit conditions, ABA-modified root architecture contributes for the development of deeper root system along with enhancing hydraulic conductivity of plant and maintenance of cell turgor, which will finally contribute towards desiccation tolerance. Other hormones such as auxin, ethylene and cytokinins (CKs), may alter the effect and biosynthesis of ABA.
Under water and temperature stress, ethylene can regulate root growth and development by limiting organ expansion. A significant positive correlation was observed between rate of grain filling and ABA content. The higher grain ABA concentration might result from autosynthesis within the grain and partly by the translocation from leaves and roots during soil drying. ABA increased the endogenous content of proline under drought conditions. CK plays a supportive role during water deficit conditions by stimulating osmotic adjustment. Brassinosteroids increase the tolerance to high temperature in wheat leaves and brome grass. The tolerance in plants to high temperature due to application of brassinosteroids is associated with induction of de novo polypeptide (heat shock protein) synthesis. In a dwarf wheat variety, high temperature-induced decrease in cytokinin content was found to be responsible for reduced kernel filling and its dry weight.
2.8.4.2 Wheat Improvement for Waterlogging Tolerance
Frequent occurrence of climatic extremes such as heavy rainfalls, reduction in freshwater availability and saline water intrusion close to the coastal area adversely affect agricultural production worldwide. Waterlogging adversely affects bread wheat production in about 4.5 million hectares in irrigated soils of the Indo-Gangetic Plains of Northern India and some other parts of the country. Under high soil salinity, leaf area index, leaf area, maturity duration and dry matter accumulation in spikes are also reduced. The combined salt and waterlogging stresses significantly reduce wheat yield by reduction in effective tiller number grain weight, length of spike and spikelets number, and also show more adverse effect than salt alone stress in case of compacted soils. The soil where water stands on the soil surface for a prolonged period of time or the available water fraction in the soil surface layer is at least 20% higher than the field water capacity, is defined as waterlogged soil. Tolerance to waterlogging by plants defined as the capability to maintain high rates of growth coupled with greater source to sink accumulation and eventually higher grain yield under adverse climatic conditions. Due to the increased frequency of extreme climate events, waterlogging has become an important constraint to crop production globally.
2.8.4.3 Pre-harvest Sprouting Tolerance in Wheat
Among abiotic stresses, pre-harvest sprouting (PHS) is a major concern for wheat cultivation in eastern and far-eastern parts of the country due to untimely rains around maturity time. The PHS in wheat is characterised by premature germination of kernels in a mature spike prior to harvest (usually under wet and humid conditions) due to early breakage of seed dormancy usually the result of moist weather conditions that persist after physiological maturity. It causes yield loss due to decrease in thousand grain weight and also affects end product quality.
Besides, wheat flour made from the sprouted wheat loses its thickening power and bread baked from sprouted wheat grain shows smaller volume and a compact interior. This decrease in quality is mainly due to early α-amylase activity, which can be characterised by Hagberg falling number. Either too low or high seed dormancy is undesirable, allowing pre-harvest sprouting after seed maturity or delaying germination after seed sowing. Therefore, development of wheat genotypes with a balanced degree of seed dormancy is needed to grow wheat in such areas where temperature and moisture during grain development adversely affects the expression of dormancy and pre-harvest sprouting resistance. PHS tolerance is a complex trait and its genetics need to be dissected using modern methods of QTL analysis.
2.8.5 Heterosis and Hybrid Development
Hybrid wheat is considered to be one of the possibilities for increasing wheat yield potential. Despite over 45 years of research primarily on development of hybrid wheat through cytoplasmic genetic male sterility-fertility restoration system, only limited success could be achieved. The reasons for its limited success are limited heterotic advantage; their lack of agronomic quality or diverse resistance advantage, high cost of seed production and fixing in polyploid with the result no advantage over pure lines was observed. Few reports indicate that hybrid cultivars have also been released through the use of chemical hybridising agents (CHA) in some countries. Though the prospects of exploiting of hybrid vigour in wheat were recognised as early as 1962 and work on hybrid wheat started the world over including India, due to the advent of high-yielding varieties utilising Rht gene(s) the emphasis was shifted. In the post Green Revolution era, it was felt that the northwestern plains of the country comprising Punjab, Haryana, western Uttar Pradesh, plains of Jammu and Uttarakhand and northern Rajasthan, which were considered as the seat of the Green revolution and contributed most to the wheat basket, have reached a sort of saturation level. To keep the productivity growth rate in tune to the future demand, it is needed to explore new innovative approaches to break the yield barriers and make wheat cultivation more remunerative. In this context, exploiting hybrid vigour at commercial level through development of hybrid wheat is considered promising that offer a significant means of overcoming food shortages because of yield heterosis.
Although the heterosis was reported in wheat in the early periods of the twentieth century, the discovery of an effective cytoplasmic male sterility (CMS) and pollen fertility restoration systems in wheat in 1951 by Kihara opened up new avenues for commercial hybrid seed production. After search of the male sterility system in wheat, the efforts were made in India to explore the basic facts for development of hybrid wheat. The levels of heterosis were explored through hand pollination, and reports have provided ample evidence of significant and positive heterobeltiosis (heterosis over better parent) for yield ranging from 0 to 100% in wheat, but most of these results are based on space planted and small plot trials. Harvest index was noticed as an important indicator of source to sink relationship, and therefore, the possibility of increasing yield potential through better harvest index was advocated during the 1990s. Under the field conditions, the minimum accepted standard heterosis for yield was established at 20% level for commercial exploitation of a hybrid and studies indicated significant standard heterosis of more than 20% for yield and yield traits under drill sown conditions.
As floral biology of wheat is key factor in order to understand the outcrossing behaviour, the extent of natural outcrossing up to 1.82% in cultivated varieties of wheat was observed. It was reported that wheat has mixed chasmogamous/cleistogamous type of flowering and autogamous/allogamous mode of pollination. Important floral traits that influence outcrossing in wheat are stigma size, anther size, anther extrusion, pollen number and pollen viability. The stigma length of wheat genotypes has been noted to be 1.84–3.75 mm, whereas anther length was observed from 2.87 mm to 5.07 mm. Anther extrusion has been observed from 8.7 to 87.4%. Phenotypic differences among wheat cultivars for days to heading, anthesis, anther size, pollen grain size, pollen viability, duration of floral opening and openness of florets were also observed, and it was suggested that the selection for long anthers, high rate of anther extrusion and more openness of florets may be effective in promoting natural cross pollination.
The pollen viability in wheat ranged from 83.4 to 98.4%. The largest separation angle between the glumes of the first two florets of spikelet was found to be 11.50–35.80, and it was observed that wheat florets get closed within 8.7–40.3 min of floral opening. All the viable pollen grains that have access to the stigma surface germinate as soon as they come in contact with the stigma. There is a positive association between anther size and the quantity of pollen produced /anther. A significant association of anther length with stigma length and anther extrusion with duration of floral opening was observed, and it was suggested that the selection for the traits that promote outcrossing may result in the genotypes with more open pollination ability and these may be utilised as parents to improve yielding ability through enhanced heterozygosity.
2.8.5.1 Male Sterility and Fertility Restoration
Male sterility and fertility restoration are two most important components in cytoplasmic genetic male sterility system. A number of studies have been conducted for utilisation of cytoplasmic male sterility system for hybrid production in wheat. In the investigations on basic aspects of male sterility system, the effect of timopheevii cytoplasm was reported on level of hybrid vigour, and it was observed that restoration of male sterility in the F1 was genetically complex, incomplete and affected by genetic background. Investigations on genetic male sterility were also carried out, and a specific type of male sterility ‘S738’ caused by the interaction of three recessive genes was reported for the first time that had an additive effect occurring in F2 population of a composite cross received from Mexico. In this system, degree of male sterility was dependent upon ms gene number (the lesser the number, the lesser is the expression of male sterility). Thus, the genotypes having one, two and three ms genes exhibited 36–49, 47–68 and 73–97% male sterility, respectively. This expression was found to be influenced by environment and by modifying genes. The application of genetic male sterility in recurrent selection schemes was also advocated.
2.8.5.2 The CMS System
Systematic investigations revealed that T. timopheevii and T. araraticum cytoplasm (G type) induced complete male sterility and there are genes restoring fertility for G type cytoplasm. At Indian Agricultural Research Institute (IARI), New Delhi, ten CMS lines, namely Lok-1KMS 9A, 2009KMS 9A, 2038 A, 2046KMS 9A, 2041 KMS 9A, 2022 A, 2042 A, 2046 KMS A, 2019KMS 9A and 2160A having cytoplasm from T. timopheevii, T. araraticum, T. zhukovskyi, Aegilops speltoides and Ae. caudata, have been developed. Three cytoplasmic male sterile lines, namely 2041 KMS 9A, 2046 KMS 9A and 2338 KMS 20A carrying T. araraticum cytoplasm, were characterised. Cytoplasmic male sterile lines carrying cytoplasm from Aegilops umbellulata, Ae. comosa, Ae. caudata and Ae. speltoides have also been produced.
However gene(s) restoring fertility in these cytoplasms have not yet been identified. No apparent adverse effects on morphology by T. araraticum and Ae. speltoides have been noticed. About eighty CMS lines including those of T. araraticum, T. timopheevii, Ae. speltoides, Ae. kotschyii and Ae. Variabilis were backcrossed with respective maintainers for diversification. Two exotic genetic stocks registered as PWR 4099 and PWR 4101 indicated complete fertility restoration in T. timopheevii-based CMS lines. Genetic studies revealed that there are duplicate dominant genes involved in restoring complete fertility in T. araraticum cytoplasm and that one out of two genes restored fertility with greater degree than the other. In order to enhance variability among alloplasmic lines, agronomically superior genotypes maintaining male sterility have been utilised. Although there is no significant result for heterosis for yield in totality, few hybrids showed heterosis for yield components, viz. spikelet number, spike length and tillers/plant.
2.8.5.3 Genetic Male Sterility
Although genetic male sterility has been reported in wheat, there is no report available showing development of any hybrid wheat. The reason could be the non-availability of appropriate kind of genetic male sterility system. A number of male sterile plants were isolated from crosses involving Selection 212 and other hexaploid wheats including Selection 82. These were isolated from F3 progenies derived from a cross involving Selection 212 and HD 2009. In Selection 82, male sterility is caused by the modification of anthers into fully fertile ovaries. This modification of anther into ovaries takes place in 85 to 90% of the florets per spike. Breeding behaviour of genetic male sterile plants was studied for 8 generations, which showed consistency of hpg-mst trait. During this period, selfed hpg-mst spikes produced 6.06 seeds per spike, which was much lower than Selection 212 (40.4 seeds per spike). In terms of sterility, hpg-mst plants exhibited 85 to 100% male sterility and 100% female fertility. The inheritance of hpg-mst traits was studied by crossing with cultivar Kundan. All the Fl hybrids produced 45 seeds per spike amounting to full fertility, indicating the dominance of fertility trait over the sterility.
A segregation ratio of 3 fertile: 1 sterile plants in the F2 generation clearly revealed the monogenic recessive control of hpg-mst trait. The hpg-mst system has few advantages as the hpg-mst plants produce 10–12% seeds (selfed or left open); therefore, it is most economical to maintain it in homozygous condition and it enhances the chances of out crossing (female fertility) because of the presence of multiple ovaries in the florets of hpg-mst plants. Selection 82 was used for developing wheat hybrids involving ten different genotypes. In one combination where pollinator was Agra Local, 53 hybrid plants produced 3240 grams of grain yield in 3 m2 area. On the other hand, it has disadvantages also as hpg-mst seeds in crossing block may give rise to 10–12% male sterile plants that lower the production per unit area. The genotype possessing hpg-mst are slightly tall (about 110 cm), and therefore, not suited to favourable environments. However, diversification of genetic male sterility system to dwarf genotypes is needed to exploit the system to its fullest extent.
2.8.5.4 Chemical Hybridising Agents
The chemicals were used as a tool to create male sterility in wheat; and more than 40 chemicals have been patented as potential chemical hybridising agents (CHAs) world over. In the 1970s, the use of CHAs as pollen suppressant began to get serious thought, and maleic hydrazide (MH) was identified as potential chemical in 1960 at IARI, New Delhi. The technique of inducing male gametic sterility looked promising as it does not require restorer parents and large quantity of seed can be produced using CHA. Some of the growth regulators like ethrel and herbicide daltapon, which expectedly have strong phytotoxic response, were also tried. Results indicated that these chemicals induced partial male sterility.
The successful utilisation of anilates as chemical hybridising agents in rice inspired the synthesis and screening of some oxanilates and inalonani lates at premeiotic stage on few varieties during 1997. Systematic investigation helped in identification of most effective fluoro-anilates inducing male sterility without having any adverse effect on different growth and yield parameters including female fertility. In the 1990s, comparative studies were made to study the effect of etherel and MH, two most widely used CHAs, and it was observed that the etherel was more effective towards reduction in seed set. The appropriate stage for higher efficacy of CHA was reported from spike length of 7–8 mm to early boot stage. Further, the late sown crop was found more responsive to CHA than normal sown crop. Deficiency of copper was also reported to induce male sterility in wheat. In the private sector, the Maharashtra Hybrid Seed Company (MAHYCO) in collaboration with Monsanto of USA is the only company, which has commercialised hybrid wheat based on CMS approach and has released three varieties Pratham 7050, Pratham 7070 and New gold for the Central and Eastern zone of India.
2.8.5.5 Bottlenecks in Research
The insufficient levels of heterosis, low seed multiplication rate and complexity of the hybridisation systems were explored as major limiting factors for hybrid wheat development. The self-pollinated nature of wheat with occasional outcrossing of usually less than 1% makes the selection for floral characters, which enable sufficient cross fertilisation like more open flowering habit, duration of flower opening, improved anther extrusion in the male parent and stigma receptivity in the female parent more crucial for successful development of hybrids. These traits need to be investigated properly to identify the parents that can be put under conversion to male sterility system and fertility restoration.
The discovery of an effective cytoplasmic male sterility and pollen fertility restoration systems in wheat using Ae. caudata cytoplasm opened up new avenues for commercial hybrid seed production, but the stability of male sterility across the locations is another bottleneck in the direction of development of hybrids, which restricts the hybrid lines to the location specificity. T. timopheevii seems to be the most suitable one for commercial production of hybrid seed. The inclusion of yield potential in the bread wheat is also an important issue. As wheat in natural polyploid (allohexaploid), the transfer of donor traits from related species takes in more negative traits than the positive components. This needs strengthening of the pre- breeding activities for improving parental lines. The economics of hybrid seed production is of major concern for successful hybrid technology. The contributing factors such as the plant population, male and female row ratios, plant spacings and input managements should be optimised for getting maximum hybrid seed at lower costs.
2.9 Breeding Approaches: Conventional and Molecular Including Use of Genomic Tools
Bread wheat is one of the most important food crops improved by mankind over the past 8000 to 10,000 years when the species first arose in the world. The wheat domestication coincides with the beginning of agriculture, and since then it has been constantly under selection by humans sometimes with accidental and also applying intentional but empirical selective pressures. With the advancement in scientific breeding methods, the focus of breeding programmes on yield potential, resistance/tolerance to biotic and abiotic stresses and quality have been prioritised. Additionally efforts have been made to introgress new variability from alien species which otherwise was not available in the primary gene pool. With the development of molecular genetics, tools like molecular markers, genome sequencing, better understanding of the complex traits are also progressing. New approaches like transgenic, gene editing, speed breeding and high-throughput phenotyping are emerging and have shown promise to improve and enhance the efficiency in crop breeding.
2.9.1 Conventional Breeding Methods
2.9.1.1 Mass Selection
From a variable population, seeds are collected from phenotypically desirable individuals in a population, and next generation is planted from the selected mixed seed. Mass selection has been practised by local conservators/farming communities to improve old varieties/land races. A variant of this method is progeny selection, in which best plants are harvested separately and their progenies are grown separately and compared for their performance. The better performing progenies are selected and the rest are rejected based not only on phenotypic selection but also on progeny performance.
2.9.1.2 Pure-Line Selection
From a genetically variable population, several phenotypically superior plants are selected and individual plant progenies are grown and evaluated over a period of several years. The homozygous progenies are bulk harvested separately, and yield trials are undertaken to ascertain the yield performance of selected progenies. Any progeny performing superior to varieties under cultivation is then released as a new ‘pure-line’ variety.
2.9.1.3 Hybridisation-Based Methods
Breeding methods employing hybridisation have the objective of combining superior gene/gene combinations from diverse parents to produce homozygous progenies in self-pollinated crops. Genes, however, are always in the company of other genes in a collection called a genotype. Different methods like pedigree, bulk and backcross breeding methods are in practice.
2.9.1.3.1 Pedigree Method
Pedigree method involves crossing of two genotypes differing in a few traits but possessing traits, which are absent, in each one of them followed by selfing and selection in successive segregating generations. The unique feature of this method is that the record is maintained for parent-progeny relationship in all the segregating generations until homozygosity is achieved. From every F2 plant individual, F3 progeny is planted separately and a few plants are selected based on desirable features to constitute the next generation followed up to F5–6. At this stage, each phenotypically superior progeny is harvested in bulk to obtain large seed for further evaluation in yield trials. These lines are evaluated for yield, disease resistance, quality features, etc.
2.9.1.3.2 Bulk Method
Bulk method of breeding does not require the record keeping of parent-progeny as in case of pedigree method and also in handling of segregating generations. In bulk method of breeding, the F2 onwards seed is harvested in bulk and used for raising the next generation, and both natural selection and artificial selection are practised. After several cycles of selfing, single plants are selected and evaluated just like in the pedigree method.
2.9.1.3.3 Back Cross Method
Backcross method is generally applied in situations when an outstanding variety becomes susceptible to major diseases or it lacks some important character. In this method the outstanding variety lacking few traits is crossed with donor having that trait followed by four to six backcrosses with the outstanding variety to recover recurrent parent genome. At the end of backcrossing cycles, selfing is done to recover the homozygous progenies having the target trait in the recurrent parent genome.
2.9.1.3.4 Hybrid Breeding
Hybrids are generally referred to a specific cross between two good combiners in a particular cross and the superior performance of hybrids over parents is referred to as hybrid vigour (a separate section in this chapter provides further details).
2.9.2 Non-Conventional Breeding Methods
Till the 1980s, genetic enhancement of crop plants was primarily based on conventional plant breeding approaches. Although conventional breeding has continued to be the breeder’s choice, faster genetic gain is hampered particularly for complex traits (Tuberosa 2012). Selection of desirable traits by indirect selection through closely linked molecular markers was conceived as an alternative to solve the limitations of conventional breeding (Collard and Mackill 2008). Since the 1990s, marker assisted selection (MAS) has been used in plant breeding programmes through tagging of the major genes, which enabled in the development of many varieties in different crop plants. Initially, both public and private sector organisations funded large number of marker assisted backcross breeding (MABB) projects across the globe; later the focus was shifted towards only foreground selection due to faster varietal replacement ratio. Later, marker assisted recurrent selection (MARS) being used as an effective tool to accumulate the alleles for the trait of interest (Rai et al. 2018). Nevertheless, MAS (Collard and Mackill 2008; Servin et al. 2004) and MARS (Crossa et al. 2010) are still the breeder’s choice to rectify the drawbacks associated with widely adapted cultivars through gene pyramiding and further incorporate novel gene(s) into desirable parents. Recently, genomic selection (GS) and speed breeding have emerged as the most promising breeding strategies to accelerate genetic gain in crop plants. GS has a clear-cut advantage over pedigree breeding and MAS to enhance genetic gains for complex traits (Crossa et al. 2017). Integrated approach of genomic selection and speed breeding could fast-track gene bank mining for rapid genetic gain (Li et al. 2018).
2.10 Precise and High Throughput Phenotyping Protocols
2.10.1 Physiological Techniques for Abiotic Stresses
2.10.1.1 Normalised Difference Vegetation Index (NDVI)
Canopy health in terms of canopy greenness indicating canopy growth or early vigour is a crucial characteristic of a high yielding cultivar and therefore is largely used as a proxy trait for abiotic stress tolerance. Canopy greenness is measured by green seeker or NDVI (Normalised Difference Vegetation Index), which is also used to estimate the nitrogen levels in many agricultural crops (Lawal et al. 2018). Several studies indicated significant linear relationship between NDVI values and seedling vigour, growth rate and senescence patterns in wheat (Lopes and Reynolds 2012).
2.10.1.2 Canopy Temperature (CT)
The genotypes with low canopy temperature are able to regulate stomatal functioning efficiently along with extraction of water from deeper layer of soil. Plant canopy emit long wave infrared radiation, which are sensed by infrared thermometer and displayed as temperature. Several studies demonstrated CT as a promising trait for screening large population under low moisture with multiple complementary traits like, deep root, stomatal conductance and finally better water use (Balota et al. 2008). However, CT is affected by various factors such as soil moisture, solar radiation, wind speed, temperature and relative humidity. Water stress was imposed between tillering and anthesis stages in wheat (Rashid et al. 1999), and mean canopy temperatures were used as a selection trait. These CT variations among genotypes were significantly correlated with yield.
2.10.1.3 Water Use Efficiency (WUE)
In general, efficient user is expected to be a higher producer, and hence higher water use efficiency is a preferred trait. Schulz et al. (2021) pursued WUE as a crucial factor for determining the final yield as it is directly related to the stomatal evaporative rate and the biomass produced. However, management practices also play an important role for WUE but at plant level, stomata are the key players. WUE is highly dependent on the carboxylation pathway, and compared to C3 and C4 plants CAM plants have a higher WUE because of unique carboxylation pattern (Hatfield and Dold 2019). WUE can be used as a key trait to screen large segregating population and significantly correlated with lower stomatal conductance. The importance of selecting crop cultivars with higher WUE which can produce higher or at par yield under limited or reduced irrigation has been previously reported (Meena et al. 2019).
There is a necessity for designing specialised experiments aimed at identification and development of germplasm with true genetic ability with increased WUE. Physiological trait-based identification of drought adaptive genotypes have been earlier reported by Fletcher et al. (2018) and Nakhforoosh et al. (2016). These studies had a few limitations in the form of screening a very few genotypes initially and that too under pot culture conditions, such results are often difficult to translate to field conditions (Meena et al. 2019). Few planned wheat breeding efforts by screening large set of germplasm lines leading to identification of cultivars with enhanced WUE at global level were reported earlier (Meena et al. 2019).
2.10.1.4 Transpiration Efficiency (TE)
The balance between transpiration (H2O) and gas exchange (CO2) helps to maintain optimum leaf temperature and photo-respiratory activity for better productivity, and hence TE used as an alternate trait for WUE (Farooq et al. 2009; Pietragalla and Vega 2012). So, under moisture deficit condition genotypes that reduce water loss through efficient transpiration should be identified and selected. TE is an important representative selection trait for light interception, leaf transpiration, gas exchange and photosynthetic ability (Steduto et al. 2007). However, it is also directly or indirectly dependent on the carboxylation efficiency of genotype (Ludlow and Muchow 1990) and higher for C4 plants (maize and sorghum), compared to C3 plants (wheat, oats, cotton). The cereal crops have higher TE than the legumes due to higher energy requirements of N fixation. For wheat, wide variations in TE has been found and correlated well with carbon isotope discrimination. Transpiration efficiency has a significantly negative association with carbon isotope discrimination (CID). The relationship between CID and TE was confirmed in many C3 crops (Richards et al. 2010). Rebetzke et al. (2002) concluded that low 13C/12C discrimination along with low stomatal conductance helps wheat to improve water use efficiently to improve harvest index.
2.10.1.5 Stomatal Conductance (SC)
For an active soil–plant–atmosphere continuum, stomata are the key controller. Under the variable soil–atmosphere moisture, stomatal alterations help to maintain plant internal water status by restricting the CO2: H2O movement between leaf and atmosphere. The significance of SC as an early response to water stress has been documented extensively for drought conditions (Damour et al. 2010; Ennahli and Earl 2005). Many studies have found a significant positive correlation between SC and crop yield, but still for field selection SC is not given much importance in breeding programmes (Roche 2015). Changhai et al. (2010) showed that the transpiration efficiency was more affected than the photosynthetic efficiency in wheat under drought stress. The major role to maintain transpiration efficiency was played by stomatal conductance and more precisely by stomatal pores. In a drought experiment, Liu et al. (2005) highlighted a significant reduction in stomatal conductance in water-stressed potato plants at tuber initiation and tuber bulking stages after 48 h and 24 h of withholding of irrigation, respectively. Studies have shown that partially stomatal closure is an adaptive strategy under drought stress to reduce SC and loss of water (Zait et al. 2019). However, the exact mechanism of stomatal closure is not well understood, but root hydraulic conductance and abscisic acid (a signalling molecule) are reported as the main determinant of SC. Under well irrigated conditions, higher stomatal conductance will maximise photosynthetic rates and hence the yield, but when the water is limiting then sustained SC accompany drought avoidance in plants to maintain adequate yield.
2.10.1.6 Relative Water Content (RWC)
RWC is a regularly used parameter to identify the variation in plant water status among genotypes and to quantify the extent of dehydration under abiotic stress (Guo et al. 2010). The RWC value ranges between 95 and 98% for a tolerant genotype or under normal conditions and ~ 40% under severe stress condition.
2.10.1.7 Leaf Chlorophyll Content
The crop canopy greenness contributed mainly by the photosynthetic pigment; chlorophyll is another trait of importance to screen germplasm for heat tolerance in germplasm lines. The chlorophyll pigment reflects only the green fraction of the light after absorbing all other colour fractions and hence it is green in colour. The canopy greenness is directly related to photosynthetic efficiency of the plants. The chlorophyll content of the leaf can be estimated by a destructive lab-based DMSO: acetone extraction method and by using an instrument called chlorophyll meter, which is non-destructive and optical method. The measurement by optical method using different types of chlorophyll meters is found to be more relevant than DMSO method under field conditions (Dwyer et al. 1991). The chlorophyll content measured through chlorophyll meters is in the form of an index called chlorophyll content index (CCI). The CCI ranges from 0 to 99.9, and with the increase in the level of heat stress the CCI decreases and CCI of healthy plant ranges from 40 to 60. As optical method is based on leaf reflectance, it is influenced by time of day in terms of light (Mamrutha et al. 2017). Care should be taken to measure chlorophyll content at uniform time and in specific leaf across the genotypes under field (Mamrutha et al. 2017).
2.10.1.8 Canopy Greenness/Stay Green Habit
Prolonged maintenance of canopy greenness also referred to as stay-green nature is a physiological adaptation mechanism by plants under heat stress and drought environments. Lim et al. (2007) described stay greenness as ‘leaf senescence is characterized initially by structural changes in the chloroplast, followed by a controlled vacuolar collapse, and a final loss of integrity of plasma membrane and disruption of cellular homeostasis’. Stay green trait in tolerant genotypes helps in withstanding chlorophyll loss and maintains photosynthesis levels under high temperature stress. Association of stay green habit with sustained yield levels under heat stress has been earlier reported, and QTL regions regulating this have been identified (Vijayalakshmi et al. 2010). There are mainly two types of stay green types. One is productive type, where in the stay green plant parts actually contribute for sink/grain filling. Another is cosmetic stay green type, where in greenness in these plants will not contribute for grain filling. Hence, identification of true and productive stay green types are also a challenge and can be done by considering other traits like water soluble carbohydrates in stem, peduncle, etc.
The canopy greenness can be measured by an instrument known as normalized difference vegetation index (NDVI) sensor. Spectral reflectance-based NDVI values are highly correlated with yield under temperature stress (Lopes and Reynolds 2012). NDVI values range from 0 to 1. Zero represents no greenness and one represents maximum greenness (Mamrutha et al. 2017). Stay green habit can also be measured by other instruments such as canopy analyser (Licor) or porometer, which measures leaf area index and green area index (GAI). Many other techniques like the digital photography of the canopy can also be taken from same height from the ground level, and pictures can be analysed with different softwares (Adobe photoshop CS3 extended or later version) to assess the early ground cover (Mullan and Reynolds 2010).
2.10.1.9 Earliness Per Se in Wheat
Earliness (earliness per se) in wheat is an adaptation strategy characterised by early heading, followed by early maturity of genotypes under high temperature stress environments. Earliness helps genotypes to complete the essential plant growth stages, such as seed setting and grain filling under favourable temperatures, thereby avoiding the occurrence of terminal/late heat stress. Mondal et al. (2013) reported that the early heading entries performed well in areas affected from terminal heat stress as earliness helps them to escape high temperatures during grain filling stages. In addition to helping them escape the terminal heat stress, earliness also resulted in achieving >10% higher yield compared to the local check varieties under high temperature stress environments. High grain filling rate in early maturing genotypes was also reported to be promoting heat stress tolerance in durum wheat (Al-Karaki 2012). Tewolde et al. (2006) reported that earliness helped cultivars adapt to high temperature stress as they had longer post-heading period resulting in longer grain filling duration. Therefore, earliness was also suggested as a key trait in breeding for high temperature stress tolerance (Joshi et al. 2007).
2.10.1.10 Photosynthetic Efficiency
The differential rate of photosynthesis expressed as photosynthetic efficiency is again a very essential component trait contributing to tolerance under high temperature stress. Stable photosynthetic rates over longer duration in heat tolerant genotypes contributed to higher grain weight, higher harvest index under stress showing the positive association of rate of photosynthesis with yield parameters under heat (Al-Khatib and Paulsen 1990). Looking at the major role played by photosynthesis in determining yield under heat stress, it is also pertinent to have phenotyping techniques to help breeders to select for genotypes with higher photosynthetic efficiency. The relative photosynthetic efficiency can be indirectly predicted using the chlorophyll content index, however there are instruments available which can measure the photosynthesis exactly. Infra-red gas analyser (IRGA) is used to measure the photosynthesis on a real time basis when stress period is available or stress is imposed under experimental conditions. IRGA measures the amount of CO2 fixed during photosynthesis by estimating the difference in amount of CO2 pumped in and moving out of closed leaf chamber (Nataraja and Jacob 1999).
2.10.1.11 Cell Membrane Thermal Stability
Under high temperature conditions, the cell membrane becomes weak and tends to rupture, leading to leakage of electrolytes. Membrane thermal stability is being repeatedly used as a measure of electrolyte diffusion resulting from heat induced cell membrane leakage. Increased level of electrolyte leachates diffused from cells is measured here. Heat tolerant genotypes are identified by measuring electrical conductivity as an index to indirectly measure membrane thermal stability (Blum and Ebercon 1981). Greater amount of electrical conductivity said to be indicating better heat-stress tolerance (Saadalla et al. 1990). Presence of high genetic heritability of membrane stability in wheat was seen to be an advantage for its use in breeding for heat tolerance (Fokar et al. 1998).
2.10.1.12 Root Studies
Based on the hypothesis that under water-stress conditions root biomass contributes significantly towards higher yields, Jain et al. (2014) conducted an experiment to identify the most stable wheat variety under water-deficit environment for root dry matter and root volume. They also calculated the stress tolerance index (STI), which indicates the tolerance to moisture stress. Tomar et al. (2016) studied a set of 158 wheat genotypes after screening in polyvinyl chloride (PVC) pipes for root architecture traits. The visible evaluation of root images using WinRhizo root scanner of HW2004 (water stress tolerant) indicated compact root system with longer depth, while HD2877 (water stress sensitive) exhibited higher horizontal root spread and less depth at reproductive stage. The importance of water is much more in the crops requiring higher water input, like rice and semi-irrigated aerobic cultivation of rice is recommended as a water saver strategy.
2.10.2 Other Screening Methods
2.10.2.1 Index-Based Field Screening
For large scale screening in open field conditions, we do not have a standardised methodology as we don’t have any hold over the indirect water sources available like rain, dew, etc. Hence, we could only screen the same if there is no rain fall during the crop period. Under such scenario we use physiological indices like Drought Sensitivity Index (DSI). The formula for DSI calculation is as given below:
where YD is the grain yield for each genotype under drought condition, Yi is the grain yield for each genotype under irrigated condition, XD is the mean of genotypes grain yield under drought condition, Xi is the mean of genotypes grain yield under irrigated condition, DSI less than 1 is desirable for water moisture stress-tolerant genotypes. The lower value of DSI represents better tolerance under water stress.
2.10.2.2 High Throughput Screening Platforms
Under controlled conditions screening could be done with high-throughput phenotyping (HTP). High-throughput phenotyping is a remote sensing technology, which may meet the requirements for the phenotyping of large number of genotypes grown in plots in less time. Use of active and passive spectral sensing systems for drought and high yield was described by Becker and Schmidhalter (2017) is also a high-throughput technology used to evaluate the drought tolerance of winter wheat. Wheat crop was grown under drought conditions for 2 years, to estimate the moisture stress tolerance of 20 wheat cultivars by using high-throughput measurements. Thermometric measurements showed a strong linear relationship to drought-related parameters (RLWC and CID of leaf and grain) and grain yield under drought stress, and demonstrated a significantly high suitability for high-throughput measurements. Additionally, four spectral reflectance sensors, including a hyperspectral passive sensor, an active flash sensor, the Crop Circle and the GreenSeeker were utilised to evaluate drought stress related destructive and non-destructive morpho-physiological characteristics. The experimental results emphasised that precision phenotyping supported the incorporation of plant traits in breeding programmes for effective phenotyping to screen drought-tolerant genotypes.
In addition to using spectral reflectance measurements, novel facilities such as temperature-controlled phenotyping facility (TCPF) has been employed to precisely phenotype genotypes for abiotic stresses such as heat (Sharma et al. 2018, 2019) (Fig. 2.5). In a study involving 75 genotypes from a recombinant inbred line population, they screened against heat stress using TCPF, the authors reported that greater precision in differentiating high-temperature responses in the TCPF was evident from the repeatability in terms of growth, physiology and productivity.

Temperature-controlled phenotyping facility (TCPF) at ICAR-IIWBR, Karnal
2.11 Emerging Challenges at National and International Level
Wheat is one of the widely grown staple food crops for feeding the global human population. Suitability of wheat for making infinite and diverse end products has made it a popular cereal over other crops. As per the WHO estimates, the world would need almost 60% higher wheat by 2050 from its current production level of 732 million tons in 2018–19 (FAO 2021); more specifically South Asia and Sub-Saharan Africa are expected to double their wheat production to meet the then population load (Fig. 2.6). The survey report of FAO also estimates that North African, Middle East’s, Sub-Saharan Africa’s, Indonesia’s, Philippines and Brazilian wheat imports would soar by 2050 due to population growth rate and wheat consumption per capita growth rate. The world’s significant wheat exporters, viz., USA, Canada, Australia, the Black Sea Region, Europe and Argentina, are expected to see minimal, or even negative, population growth towards 2050. In contrast, population growth will be strongest in the countries of the tropic and subtropical regions where little wheat is grown. It is believed that, even without projecting large imports by China, the world wheat trade will likely double by 2050, to 240 MMT or more.

Comparative wheat yield (t/ha)—India vis-à-vis other countries during 2019
Thus, realising that wheat already accounts for one-third of all global grain traded. Such a large expansion of trade will have major implications for all segments of the industry, including buyers, shippers, handlers, and especially the producers in those countries that will supply the increased exports, including the USA. To meet this demand, developing countries should increase their wheat production by 77%, and more than 80% of demand should come from vertical expansion. The production target is not very high, however, it has to be achieved when productivity growth (genetic gain per annum) in wheat is hovering around 0.9%. Besides, there is an urgent need for enhancing productivity through agronomic (water, nutrients, weed management, etc.), genetic and physiological interventions along with resource conservation technologies.
Providing an adequate supply of food, however, seems to be achieved at present for the current global population, sustaining this to future will be very challenging in view of steadily increasing population, increasing purchasing power and continuous diminishing of available fertile cultivable land and water for agriculture. The challenge is expected to make even more difficult by the projected changes in climate, particularly higher temperatures and changes in rainfall distribution and amount (Parry et al. 2010). Food supply will need to grow by 2–3% each year to meet the projected demand; but in the past decade the yields of the major cereals, rice, maize and wheat have increased at less than half this rate. Apart from that, a majority of the population in the developing countries is facing the challenge of malnutrition because of the consumption of cereals-based diets for meeting their energy demand. HarvestPlus have initiated bio-fortification programme prioritising different nutrients for different countries in different crops. Zinc is target micronutrient for which bio-fortification programme on wheat is underway in India.
The AICRP on Wheat and Barley is successful in releasing the bio-fortified varieties for cultivation in different zones of the country. WB 02 is the first bio-fortified variety released for cultivation in the country having a fair amount of grain iron and zinc contents (Chatrath et al. 2018). The climate predictions by the Intergovernmental Panel on Climate Change (IPCC) indicated that the mean atmospheric temperatures are expected to increase between 1.8 and 5.8 °C by the end of this century (IPCC 2007). The increase in frequency of hot days and greater variability in temperatures in the future is also predicted as an effect of climate change (Pittock et al. 2003; Team et al. 2014). Important crops like maize and wheat are more prone to be affected as they produce less grain at temperatures above 30 °C. The impacts of climate change on food systems are expected to be widespread, complex, geographically and temporally variable, and profoundly influenced by the socio-economic conditions (Vermeulen et al. 2012). The projected rise in temperature of 0.5 °C to 1.2 °C will be the major cause of grain yield reduction in most areas of South Asia. Higher temperatures are likely to affect around seven million hectares of wheat area in developing countries and around 36 million hectares in temperate wheat production countries. The Asia-Pacific region is likely to face the worst impacts on cereal crop yields. Loss in yields of wheat, rice and maize are estimated in the vicinity of 50, 17 and 6% respectively by 2050 (IFPRI 2009). This yield loss will threaten the food security of at least 1.6 billion people in South Asia. Warmer temperatures resulted in an annual wheat yield reduction to the tune of 19 million tons, amounting to a monetary loss of $2.6 billion between 1981 and 2002 (Lobell and Field 2007).
In India, it has been predicted that with every rise in 1 °C temperature, the wheat production will decline by 4–six million tons (Ramadas et al. 2019). Approximately, three million ha wheat area in north eastern and north western plain zones is exposed to terminal/reproductive heat stress (Gupta et al. 2013). Another report by Joshi et al. (2007) stated that around 13.5 million ha wheat area in India is vulnerable to heat stress. Temperatures above 34 °C in northern Indian plains leading to significant yield loss was reported (Lobell et al. 2012). India is considered to be the second largest producer of wheat. The Northern Indian states such as Uttar Pradesh, Punjab and Haryana are some of the major wheat producing states, where the crop is more vulnerable at a 1 °C rise in temperature resulting in reduction wheat yield. In South Asia, higher night temperatures during February and March impacted subsequent wheat production and would likely to endanger food security (Janjua et al. 2010; Paymard et al. 2019; de Lima et al. 2020). High temperature stress affects wheat crop at germination, early establishment stages, dry matter partitioning, reproductive organ development and reproductive processes leading to decrease germination, poor seedling emergence leading to abnormal seedlings, poor vigour, reduced overall growth of developing seedlings, decreased seed set and low grain number (Kumar et al. 2021; Prasad and Djanaguiraman 2014; Sehgal et al. 2018).
Climate change also affects the cropping patterns and crop rotations and influences the emergence of new biotypes/pathoypes. The outbreak of Ug99 stem rust causing major upheaval in Ug99, as it has overcome the resistance in most wheat cultivars. An estimated 80–90% of all global wheat cultivars growing in farmer’s fields are now susceptible to Ug99 or variants (Joshi et al. 2008). Similarly, Yr9 virulence first reported in East Africa and then migrated to South Asia through Middle East and West Asia over 10 years and caused heavy yield losses, and this virulence was reported in 1996, in North Western India (Nayar et al. 1996). Now, for the first time, Bangladesh reported wheat blast in early 2016, a deadly disease which otherwise confined to Latin America. Now the wheat blast disease was also reported from Zambia (Tembo et al. 2020). This disease results in complete crop loss in case of severe infection as it attacks the rachis and may completely hamper the grain filling. There are a few reported sources of resistance, which confers moderate level of resistance to the wheat crop (Urashima et al. 2004 and 2005). In China, sharp eye spot disease caused by Rhizoctonia cerealis is an emerging problem in high intensity cropping systems, and during 2003, this disease was also reported from Egypt (Hammouda 2003). Similarly, the prevalent pathogens and pests may evolve due to selection pressure posed by resistance gene deployment as well as due to climate change.
With the introduction and adoption of semi-dwarfing high yielding varieties of wheat for cultivation during and after green revolution, the soil fertility status have been depleted. Micronutrient deficiency and organic carbon content have gone down. There is an urgent need to address this issue by following addition of green manuring crops in the cropping system, residue retention and farm yard manure. Further the use of high doses of fertilisers and excess use of irrigation led to the depletion and deterioration of soil health and water. Half of the applied fertilisers are lost in one way or the other. There is an urgent need to breed for efficient genotypes, which are both efficient in utilisation of fertiliser and also give optimum yield under limited or reduced irrigation. With the adoption of conservation agriculture practices in high intensive cropping systems, the foliar diseases of wheat like septorea, tan spot and Fusarium head blight are more frequently affecting the crops due to the hemi-biotrophic nature of pathogens (Sharma and Duveiller 2003).
In the humid subtropics of South Asia, there is evidence of stress conditions, which favour foliar blight (Dubin and Bimb 1994). Factors such as minimum tillage or surface seeding, irrigation, late planting or low soil fertility may be responsible for higher foliar blight severity in the wheat-based cropping systems of the Indo-Gangetic plains. Keeping in view the present and future challenges, researchers need to look for new variability in cultivated as well as secondary and tertiary pools and keep on continuously churning that into the breeding programmes to develop climate resilient, nutrient rich and efficient genotypes in terms of nutrient, water and radiation use.
2.12 Breeding Progress and Varietal Development
2.12.1 Conventional Breeding
In India three wheat species, namely T. aestivum (bread wheat), T. durum (Kathia or Macaroni wheat) and T. dicoccum (Khapli or Emmer wheat), are commercially cultivated in different parts of the country. The wheat growing farmers have played a significant role in preserving enormous variability in blend of land races, comprising of variable grain traits and morphological characteristics. The systematic wheat improvement work started with the establishment of the Imperial Agricultural Research Institute at Pusa in Bihar by Howard and Howard in 1905. Land races were subjected to pure-line selection for developing better yielding NP Series of wheat varieties. Among these, special mention can be made of NP 4, which won several national and international awards for its grain quality. In the next phase, the wheat improvement was carried out through recombination breeding involving selected pure lines in hybridisation process at number of government agricultural colleges came up at Kanpur, Lyallpur, Pune and Sabour. Also, the contemporary wheat researchers took up massive hybridisation work at Shimla, Powarkheda, Niphad and some other places, which led to develop large number of improved wheat varieties bearing initials like NP, K, C, Pb, Pbc, AO, Hyb, RS, Niphad, Kenphad, etc.
Later the focus shifted towards resistance breeding and in this pursuit, success was achieved in the development of a classical variety NP 809, the first Indian wheat variety resistant to all the three rust and loose smut diseases. Later on NP 824 was the first variety developed for the good management conditions. Among large number of varieties developed during this period prior to 1965, the notable varieties like Pbc 518, Pbc 591, NP 52, NP 165, K 13, K 46, K 65, K 68, Pbc 228, C 273, C 281, C 519, Niphad 4, AO 113, AO 115, Hyb 23, Hyb 38, RS 31–1, Kenphad 28, etc. played important role in augmenting wheat production to some extent. During 1947, the wheat production of the country was 5.6 million tons, which was increased to the extent of 12.3 million tons in 1965 but was far below to meet the demand which was mainly due to the tall growing habit as well as proneness to lodging under high fertility conditions. A famine-like situation was predicted by Paddok brothers in India by the year of 1975. In fact it was a do or die situation, and there was no immediate solution in sight to bail out the country from such precarious situation. At this juncture when no other approach proved fruitful, a dwarf Korean wheat land race ‘Daruma’ showed a ray of hope. The Japanese researchers developed the well-known ‘Norin-10’ dwarf wheat genotype by crossing with Daruma.
Later on after the Second World War was over, the Norin-10 was picked up by U.S. biologist SD Salmon. In USA., Orville Vogel recombined the dwarfing trait of Norin-10 in the winter wheat background and successfully developed the first high yielding dwarf winter wheat variety known as ‘Gains’ in the early 1950s. It was Norman Ernest Borlaug who for the first time in the latter half of the 1950s transferred the Norin-10 dwarfing genes (Rht1, Rht2) in to the spring wheat background, while working in Mexico. The Indian wheat scientists took note of these dwarf Mexican wheat in the international nurseries received from Borlaug and grown at Indian Agricultural Research Institute (IARI), New Delhi, during 1961–62. From this point onwards what had happened in the field of wheat research in India is a history. It was a turning point for achieving a spectacular change in the production and productivity of wheat in the country. A food deficit country not only became self-sufficient in wheat production but also joined the elite group of wheat exporting countries in the world. Due to the coordinated efforts made by the multidisciplinary team of scientists with the progressive support of able administrators and hard work of farmers under the banner of the All India Coordinated Wheat Improvement Project (AICWIP) initiated by the Indian Council of Agricultural Research (ICAR) in 1965.
It could be a sheer coincidence that the advent of dwarf wheats and initiation of AICWIP took place simultaneously in India, in 1965. Considering the encouraging results obtained from the preliminary trials on dwarf wheats, the Indian Scientists visited Mexico to take on the spot stock of large number of dwarf wheat strains being grown there. In the beginning, the seed of four varieties, namely Lerma Rojo 64A, Sonora 63, Sonora 64 and Mayo 64 along with 613 advance generation progenies exhibiting segregation for rust resistance, plant height, maturity duration, grain attributes and phenomenon of grain shattering were introduced. After conducting the multilocational field trials, Lerma Rojo 64A and Sonora 64 were released for the first time as dwarf wheats for commercial cultivation in the country. However, the consumers did not like them because of their red grains and poor Chapati making quality.
For seeking answer to this drawback, the segregating generations of 613 progenies introduced from Mexico were subjected to rigorous selection by the breeders working at IARI, New Delhi; Punjab Agricultural University (PAU), Ludhiana; Govind Ballabh Pant University of Agriculture & Technology (GBPUA&T), Pantnagar; Government Agriculture College, Kanpur and Chaudhary Charan Singh Hisar Agricultural University (CCSHAU), Hisar. Number of amber-seeded genotypes exhibiting rust resistance, semi-dwarf plant type and appropriate maturity duration were developed at these centres for yield evaluations. Based on their yield performance in multilocational coordinated trials and National demonstrations, four amber-seeded improvement dwarf varieties, namely Kalyansona, Sonalika, Safed Lerma and Chhoti Lerma were released in 1967. The commercial success of these varieties acted like a catalyst, which not only brought revolution in the wheat production but also encouraged the breeders to work with more vigour and dedication. Very soon, Kalyansona and Sonalika became most popular varieties among the farmers and both these varieties occupied larger area in the country.
The area under wheat has increased from 12.84 million hectares in 1966–1967 to 20.92 million hectares in 1976–1977, but the production jumped tremendously from 11.4 million tons to a record level of 29 million tons during the corresponding period. In this way, an era of ‘Green Revolution’ was actually ushered in India. A massive hybridisation programme was initiated by Indian breeders, involving the dwarf wheat germplasm received from the International Centre for Maize and Wheat Research (CIMMYT), Mexico and indigenous cultivars/land races. This strategy of breeding led to develop better wheat varieties year after year to suit to varying production conditions of different wheat growing zones of the country. The wheat production progressively scaled new heights year after year. The phenomenon increase in area, production and productivity established new to newer records (Fig. 2.7).

Trend of area, production and productivity of wheat in India
The AICRP on Wheat and Barley have been successful in releasing more than 480 high yielding wheat varieties, which are notified for the different agro-ecological regions of the country. Of which, 311 wheat varieties have been released and notified by the CVRC, while 169 wheat varieties released by the SVRC. So far, 405 bread wheat varieties, 64 durum wheat, and 7 dicoccum wheat besides 4 triticale varieties have been notified. Among these, the land mark varieties like Kalyansona, Sonalika, C 306, WL 711, UP 262, WH 147, HD 2189, HD 2009, Lok 1, HUW 234, HD 2329, VL 616, HD 2285, GW 496, HI 8498, GW 322, WH 542, PBW 343, UP 2338, HD 2733, DBW 17, HD 2967, HD 3086 and DBW 187, etc. dominated and occupied larger area for seasons of wider adaptability, high yield potential, disease resistance, better grain quality, etc. At present, good choice of improved varieties is available for farmers for growing under different production conditions (Table 2.6).
Two new bread wheat varieties namely, DBW 187 and DBW 303 have been released recently for cultivation in the North Western Plains zones, the country having high yield potential and are adoptable for early sown high productive environments (Table 2.7). The extent of genetic gain in grain yield achieved in the Indo-Gangetic plains can be seen from the yield potential of wheat varieties, which rose from 33.7 quintal per hectare in 1965 to 61.3 quintal per hectare in 2020 (Fig. 2.8). The released varieties have been identified for making various products like bread, biscuit, chapatis, pasta products, etc. (Table 2.8).

Landmark varieties of wheat in India
In addition to improved varieties, 247 wheat genetic stocks were registered by the Plant Germplasm Registration Committee of NBPGR, New Delhi. Of them, 140 genetic stocks are for biotic stresses, 42 for quality components and 23 for abiotic stresses. These genetic stocks are continuously shared with the researchers in the country for use in their breeding programmes. A single genotype HD 2160 has played important role in the development of as many as 18 improved wheat varieties (HD 2987, HD 2967, HD 2501, HD 2428, HD 2402, HD 2327, HD 2307, HD 2281, DL 788-2, PBW 54, PBW 120, PBW 154, PBW 175, PBW 222, K 9465, K 8962, K 8434 and Raj 1972). In the same context, a single cross number 8156 performed in the 1950s at Mexico proved most productive, giving rise to well-known varieties like Kalyansona, Mexi-Pak, Super X, etc., which led the foundation of wheat revolution in the developing countries of Indian sub-continent.
India is producing enough wheat, and now the country is exporting wheat to several countries. Besides production, breeding programmes are focusing to develop nutrient rich wheat for consumption to mitigate wide spread micronutrient malnutrition. WB 02 is the first biofortified bread wheat variety released for cultivation in North India, which is rich in zinc (42.0 ppm) and iron (40.0 ppm) developed by the ICAR-Indian Institute of Wheat and Barley Research, Karnal, Haryana. Besides this a few other wheat varieties like HPBW 01 (iron 40.0 ppm and zinc 40.6 ppm), Pusa Tejas (HI 8759) durum wheat variety (12%protein, 42.1 ppm iron and 42.8 ppm zinc), Pusa Ujala (HI 1605) (high protein 13%, iron 43 ppm and zinc 35 ppm), MACS 4028 (d) (14.7% protein 46.1 ppm iron and 40.3 ppm zinc) have been released during the last 5 years. The Indian wheat breeding programme has made tremendous achievements in terms of developing high yielding varieties and making India not only self-sufficient but also exporter of wheat.
2.12.2 Genomics Assisted Breeding
At national level, a few varieties developed using marker-assisted selection have been released for cultivation. PBW723 (Unnat PBW343) is the first variety using modified marker assisted back cross breeding (MABB) and released at the national level having five resistant genes introgressed into it. Based on APR against individual pathotypes, PBW723 possesses resistance against all predominant pathotypes of yellow and brown rusts. PBW723 also has enhanced resistance to Karnal bunt compared to recipient variety PBW343. Post release, the variety PBW723 has made its way to farmer’s field and is being grown in Punjab at an average yield of 55–60 qtls/ha. More than 11,000 quintals seed have been produced since the last 2 years (Sharma et al. 2021).
Another variety, Unnat PBW550 possesses gene Yr15 in PBW550 background and provides complete foliage resistance to rusts. Gene Lr57/Yr40 has been introgressed in DBW17 background and the variety PBW771 has been released and recommended for cultivation under late sown conditions of Indo Gangetic Plains. Similarly, another cultivar, PBW752 having Yr10 gene has been released and recommended for NWPZ for late sown irrigated conditions. PBW757, a short duration cultivar released by PAU for cultivation under very late sown conditions having Yr36 gene in PBW550 background. A spectrum of wheat varieties having one or more resistant genes are available for cultivation under almost all target environments of the region and are the outcomes of systematic resistance breeding efforts.
2.13 Modernisation of Crop Improvement Programme
Several new tools and techniques have been constantly invented and are being used to facilitate breeding of new improved crop varieties including wheat. Although the field of New Plant Breeding Techniques (NPBTs) is young, it reveals a great potential and offers several advantages over conventional breeding techniques.
2.13.1 New Plant Breeding Techniques
2.13.1.1 Targeted Mutagenesis: ODM, ZFN, MGN, TALEN
Targeted mutagenesis aims in the creation of small mutations in the plant DNA at the pre-determined specific sites, and sometimes it is also known as ‘site-specific mutagenesis’. Conventionally, plant cells are exposed to chemical or physical mutagens to obtain random mutations. Whereas, in the targeted mutagenesis minor mutations occur at pre-decided sites, usually to inactivate a target gene of interest or to restore the function of a mutated gene. However, precise knowledge of the targeted gene is an essential pre-requisite in targeted mutagenesis compared to conventional mutagenesis. Several targeted mutagenesis techniques developed in the past decade can be employed in plants, including ZFN (Zinc Finger Nuclease) techniques, ODM (Oligonucleotide Directed Mutagenesis), MGN (Meganuclease) techniques and TALEN (Transcriptional Activator like Effector—Nuclease) technique.
ODM is mainly based on the use of oligonucleotides for the induction of targeted mutations in the plant genome. Approximately, 20–100 long oligonucleotides chemically synthesised in order to share homology with the target sequence except nucleotides to be modified in the host genome. ZFNs are custom-designed proteins to cut at specific DNA sequences. They consist of a ‘zinc finger’ domain (recognising specific DNA sequences in the genome of the plant) and a nuclease that cuts double-stranded DNA. ZFN techniques in plant breeding are the novel tools for the introduction of site-specific mutations in the plant genome or the site-specific integration of genes (Osakabe et al. 2010). MGNs are very specific restriction enzymes that recognise 12–30 base pairs of DNA sequences and create a double strand break (DSB) that activates repair mechanisms and DNA recombination. MGNs are a very broad group of proteins expressed by several different organisms. Among them, the family group of LAGLIDAGD MGNs is commonly used for targeted mutagenesis. TALENs are artificial restriction enzymes that are custom-designed to cut at specific DNA sequences like ZFN.
2.13.1.2 Techniques Resulting in ‘Negative Segregants’: Reverse Breeding, RdDM
This group of techniques also called ‘transgenic construct-driven breeding techniques’ (Lusser et al. 2012) has a common feature of transgenesis only in an intermediate step of the breeding process. The transgene used is subsequently eliminated by crossing and selection and is therefore not present in the final products, and for this reason it is called ‘negative (for the absence of the transgene) segregants’.
2.13.1.3 Variants of Plant Transformation Techniques: Cisgenesis and Intragenesis
As against transgenesis which can be used to insert genes from any organism, both eukaryotic and prokaryotic, into plant genomes, cisgenesis and intragenesis are terms recently created by scientists to describe the restriction of transgenesis to DNA fragments from the species itself or from a cross-compatible species. In the case of cisgenesis, the inserted genes, associated introns and regulatory elements are contiguous and unchanged. In the case of intragenesis, the inserted DNA can be a new combination of DNA fragments from the species itself or from a cross-compatible species (Rommens 2007; Schouten and Jacobsen 2008).
2.13.2 Speed Breeding: Faster and Better Phenotyping
Speed breeding is a recent technique that involves the manipulation of environmental conditions under which crop genotypes are grown, aiming to accelerate flowering and seed set, to advance to the next breeding generation as quickly as possible. Speed breeding (SB) techniques have now been developed for wheat by accelerating plant growth, flowering, seed set and maturation using supplemental lighting under controlled growth conditions. Consequently, the generation time of crop plants is reduced significantly as compared to the field or normal glasshouse conditions. The method saves breeding time and resources through rapid generation advancement. Watson et al. (2018) could get upto 6 generations of spring wheat (Triticum aestivum), durum wheat (T. durum) in a year with supplemental lighting using LED lights in environment controlled growth chambers and single seed descent (SSD) method, compared to 2–3 under glasshouse conditions and one generation under field conditions. As a result, speed breeding offers opportunities to rapidly develop homozygous and stable genotypes, and to facilitate rapid generation advancement, resulting in accelerated development and release of new cultivars (Watson et al. 2018). Also, speed breeding technology fits well with MAS and high-throughput phenotyping methodologies for multiple trait selection.
2.13.3 Genome Editing
The second generation genome editing tool CRISPR (clustered regularly interspaced short pal-indromic repeats)/associated nuclease Cas9 (CRISPR/Cas9) system is gaining momentum over the ZFN and TALEN genome editing strategies (Upadhyay et al. 2013). Just like other nucleases, CRISPR/Cas9 involves RNA-guided Cas9 nuclease from bacteria or archaea to generate targeted double-stranded breaks that are repaired by NHEJ or HDR for efficient genome editing in eukaryotes (Horvath and Barrangou 2010). Genome editing technologies can accelerate wheat breeding by allowing the introduction of precise and predictable modifications directly in an elite varietal background. Recently, the International Wheat Genome Sequencing Consortium (IWGSC; https://www.wheatgenome.org) released the fully annotated high quality reference genome of bread wheat variety ‘Chinese Spring’. This will provide more novel target genes responsive to various biotic, abiotic, quality and agronomical traits improvement through CRISPR/Cas9 system. CRISPR technology is useful both in precise enhancing the activity of positive-regulator genes and in eliminating the negative-regulator genes that affect the trait of interest.
However, there are only few reports available for validation of CRISPR technique in wheat compared to other crops like rice. Most of these genes are targeted by wheat researchers to address the major abiotic and biotic stresses, along with improving agronomic traits in wheat. The first demonstrations of the CRISPR/Cas9 system in wheat were used to knockout TaMLO locus (Shan et al. 2013), TaPDS and TaINOX (Upadhyay et al. 2013). In subsequent research, simultaneous knockout of the three TaMLO homoeoalleles has been established to confer resistance to powdery mildew in bread wheat (Wang et al. 2014). Shan et al. (2014) also validated the CRISPR/Cas9 system by targeting TaLOX2 by expressing the sgRNA under the transcriptional control of TaU6 promoter in wheat. Recently, Wang et al. (2019) showed multiplexed genome editing through CRISPR in hexaploid wheat by targeting three different genes, viz. TaGW2, TaLpx-1 and TaMLO.
2.13.4 Genomic Selection: Rapid Genetic Gain
Recently, GS and speed breeding have emerged as the most promising breeding strategies to accelerate genetic gain in crop plants. An integrated approach of GS and speed breeding could fast-track gene bank mining for rapid genetic gain in crop plants (Li et al. 2018). GS could be a promising strategy to accelerate genetic gain per unit time and cost, especially for traits governed by small and cumulative effect genes. However, the optimal integration of GS in active breeding programmes faces several challenges. Nevertheless, GS has a clear-cut advantage over other breeding techniques to enhance genetic gains for complex traits (Crossa et al. 2017). During the last one decade number of empirical GS studies reported in different crop plants, highest number of GS studies reported in wheat, followed by maize and rice, which together make upto 75% of all the studies (Krishnappa et al. 2021). Various studies suggest that GS is becoming a substantial component of modern crop breeding programmes due to rapid increase of genetic gain (Gorjanc et al. 2018). But there are very limited reports on the actual impact of GS on realised performance improvement (Voss-Fels et al. 2019) in different breeding programmes. Maize is an exception; drought-tolerant high-yielding commercial maize referred to as ‘AQUAmax’ hybrids were developed by the private sector through precision phenotyping and crop growth models in genomic prediction frameworks in the USA (Cooper et al. 2014).
2.14 Status Varietal Development and Maintenance Breeding
During the recent decades, India has observed a remarkable advancement in agricultural production and productivity owing to development of high yielding varieties by the National Agricultural Research System. The strides made in varietal development programme would not have been possible without concurrent advancement of institutional system for crop breeding research and quality seed production. Crop improvement research was initiated at various Indian Council of Agricultural Research (ICAR) institutes and State Agricultural Universities (SAU’s) and further strengthened with initiation of All India Coordinated Improvement Projects during the 1960s. The system of varietal release, farm verification trials and maintenance breeding are very well developed and standardised in wheat. There are precise guidelines for conducting All-India Coordinated Trials in a uniform way, which ensures quality breeding material/potential entries are promoted further for release. Once the variety is identified, seeds of the variety are to be deposited with the NBPGR for conservation in gene bank. After obtaining the acknowledgment with IC No. from the NBPGR, the release and notification proposal of the variety/hybrid needs to be submitted to the Central Sub-Committee on Crop Standards, Notification and Release of Varieties.
The AICWIP was initiated during 1965 at the Indian Agricultural Research Institute (IARI), New Delhi, and under this project several high-yielding wheat varieties were developed. Varieties developed by AICWIP were quickly adopted by farmers due to their high yield potential and wider adaptability (Singh et al. 2019). During 2017, the erstwhile AICWIP project was reconstituted as the All India Coordinated Research Project (AICRP) on Wheat and Barley under the ICAR-Indian Institute of Wheat and Barley Research (ICAR-IIWBR) at Karnal (Haryana). ICAR-IIWBR through its network of cooperating centres engaged in coordinating the multidisciplinary and multi-location testing of varieties across the different ecosystems for enhancing and sustaining the wheat production.
Breeder seed indents are reflections of demand of variety at national level and extent of adoption of particular variety by the farmers. As breeder seeds are produced by the ICAR institutes/SAUs and further supplied to the various indenting agencies, viz. National Seed Corporation, State Seed Corporations, State Department of Agriculture, Private Seed Companies for multiplication into foundation and certified seed. During the past 5 years, among various wheat varieties, HD- 2967 developed by the ICAR-IARI, New Delhi, remained as a top indented variety; however, during 2021-22, HD-3086 released during 2014 by the ICAR-IARI, New Delhi, took over the top slot. Further, two latest varieties, viz. DBW- 187 (2019) and DBW-222 (2020) developed by the ICAR-IIWBR, Karnal, rapidly adopted by the farmers and demand for breeder seed production has increased within a short span of time to 1617.4 q and 506.3 q during 2021–22, respectively. All the below mentioned varieties (Table 2.9) are outcome of the concentrated efforts of scientist engaged in the varietal development programme through (AICRP) on Wheat and Barley.
The new varieties developed by the plant breeder needs to be genetically pure, uniform and free from any seed borne disease. The purity of basic seed/breeder seed is the most critical aspect, which determines the success of entire seed multiplication chain. Genetically impure seed may lead to high cost of rogueing and eventually may lead to even rejection of foundation and certified seed plots. In order to maintain highest genetic purity, utmost care needs to be taken while production of nucleus and breeder seed under the strict supervision of concerned breeder (Chakarabrty and Sharma 2018).
2.14.1 Maintenance Breeding
2.14.1.1 Selection of Earhead
In a plot of advanced generation seed multiplication at second year of AVT Trial, more than 350 years may be selected randomly based upon diagnostic characters of variety. In case of released varieties, ear heads may be collected from uniform seed multiplication filed. These selected ears are threshed separately and are examined for their colour, shape and size, and ear heads which are not true to type are rejected.
2.14.1.2 Nucleus Seed Multiplication: Stage 1 (Ear to Row Method)
Seeds of selected ear heads are sown in three-meter row (six) for each variety, hence known as ear to row method. The individual rows are periodically examined throughout the growing season. All the segregating and lines containing off types are rejected. Ear rows which show the typical characters of variety and show uniformity are harvested and threshed individually. The harvested seed if bulked and used for planting of breeder seed plot, then called as nucleus seed. Around 300–500 ear to rows are grown, considering the demand of breeder seed of particular variety. Generally, five-meter isolation distance is maintained to avoid any contamination from other source.
2.14.1.3 Nucleus Seed Multiplication: Stage II (Ear to Row Progeny Plot)
In case of varieties having very large area under cultivation, breeder seed requirement is higher. Therefore, in such cases, another cycle of nucleus seed production is followed, which involves sowing of seeds of each selected ear row separately. The plots sown from seed of each selected ear row is known as ‘Ear to Row Progeny Plot’. These plots are examined for the essential diagnostic characters of the varieties at different stages as in the nucleus seed multiplication stage. In this method, six-meter length of rows are sown in six rows having spacing of 20 cm each, further these plots are isolated with minimum of five-meter length for any other varieties to avoid contamination in NSS-II.
2.14.1.4 Breeder Seed Production
Breeder seed is produced as per the allocation of indents by the Department of Agriculture Cooperation and Farmers Welfare, Government of India to the concerned institute. The breeder seed is produced from the nucleus seed (stage I or II) under the supervision of a qualified plant breeder. The isolation of three meter is recommended from other wheat variety and needs to ensure that the breeder seed plot is 150 meter isolated from loose smut infected wheat plots. At the time of sowing, care must be taken to keep one row blank after every eight rows for easy inspection and roguing. All plants which are not typical of the variety are considered as offtypes, and it is very essential to rogue out these offtypes from breeder seed plots to maintain purity of foundation and certified seeds. Breeder seed plots are monitored by team of experts consist of breeder of the variety, the concerned Project Director or his/her nominee, representative of the National Seed Corporation or seed certification agency and based on the monitoring report of team (BSP III), breeder seed is harvested and made available to the varied indenting agencies for further multiplication.
Varietal improvement/development programme is the backbone of food security of India. Wheat improvement programme over the years have developed varied high yielding, multiple stress tolerant and bio-fortified varieties catering the needs of farmers. Further, vigorous efforts are being made to make available quality seeds of such varieties at farmer’s doorstep, which is reflected in terms of higher varietal replacement rate (74.0% for varieties which are less than 10 years old) and seed replacement rate (40.30%) in wheat in India.
2.15 Coordinated System of Testing
In India, the systematic wheat research started about 100 years ago after joining of Sir Howards as the Imperial Botanist at Pusa (Bihar) in 1905. Later on, after establishment of the Indian Council of Agricultural Research (ICAR) in 1935, it became the main funding agency and promoter of wheat research in India. An important milestone in this process was the establishment of the All India Coordinated Wheat Improvement Project (AICWIP) in the year 1965, by the ICAR. Then AICWIP was elevated to the status of the Directorate of Wheat Research (DWR) in the year 1978, and in 1991, it moved from IARI, New Delhi, to its present location at Karnal, along with two regional stations (Flowerdale, Shimla and Dalang Maidan). In 2014, it became an institute, ICAR-Indian Institute of Wheat and Barley Research. In India, wheat is grown on an area of about 30 million hectares, and the cultivation extends from 90N (Palni hills) to above 350N (Srinagar valley of J & K), thus the wheat crop is exposed to a wide range of agro-climatic changes such as humidity, temperature, photoperiod during crop season, soil types, altitudes, latitudes and cropping systems. From wheat research and coordination point of view and based on land use planning, the country is divided into following five major zones: (1) Northern Hills Zone (NHZ), (2) North Western Plains Zone (NWPZ), (3) North Eastern Plains Zone (NEPZ), (4) Central Zone (CZ) and (5) Peninsular Zone (PZ) as described below Table 2.10.
Through coordinated research efforts, many high yielding wheat varieties suited to different agro-ecological conditions and growing situations have been released. These genotypes were very successfully helped in increasing the wheat production from a mere 12.5 million tons in 1964 to 108.75 million tons during 2020–2021. The wheat crop in India is menaced by a number of diseases. The survey and surveillance activity has helped to monitor the dynamics of important wheat diseases, particularly the three rust diseases. Through this mechanism, the occurrence/evolution of new pathotypes is made known before crossing the threshold limit of disease infestation, in the meantime, the genetic resistance is created against the new virulence in form of resistant varieties. Large number of donors lines carrying ‘R-genes’ conferring resistance against different rust races have been identified for utilisation in breeding programmes. The coordinated research through the AICRP on Wheat and Barley caters to the needs of cooperating centres by streamlining the research efforts and facilitating the evaluation and screening of the breeding materials. Every year national and international nurseries are supplied to different centres across various wheat growing zones, with an aim of screening the lines at hotspots and assessing the resistances across locations and environments.
Multi-disciplinary approach of variety testing in AICRP on Wheat and Barley: The Crop Improvement Division is primarily involved in coordination activities of the AICRP on Wheat and Barley, wherein the multilocation evaluation of wheat varietal trials and nurseries are undertaken. In addition to breeding work, pathological, agronomical and quality programmes also support in variety of development and testing approach of wheat in India. The wheat coordinated varietal evaluation programme entails a huge multilocation testing programme, which is undertaken with the cooperation of 29 funded and 95 voluntary centres spread across 5 wheat growing zones in the country.
Initially, the system of varietal evaluation was confined to the specific zones including initial varietal trials and advance varietal trials for different production conditions. Under this system flow material from one zone to another zone was not done, and the adaptability of genotypes was limited to specific zones. Realising this problem and widening the testing environments and to have free flow material of material across zones, the system of testing was re-structured with the incorporation of National Initial Varietal Trials (NIVTs) and their details along with production conditions and zones are presented below Table 2.11.
However, in NHZ separate zone specific Initial Variety Trials (IVTs) in place of NIVTs are conducted. This way, the Indian wheat programme is unique regarding the multi-location testing of new genotypes through different trials. The procedure of evaluation system was re-structured in such a way that the materials from different centres are pooled and tested at different levels, namely station trials, national initial varietal trials (NIVT) and advance varietal trials (AVTs) to sort out superior germplasm with respect to yield, disease resistance and quality in the following manner. After 1 year of testing in IVTs, deserving genotypes are promoted to AVTs in NHZ at zonal level. However, in case of remaining four zones, deserving genotypes come from NIVTs for advance testing in each of four AVTs of NWPZ, NEPZ, CZ and PZ (Table 2.12 and Fig. 2.9).

Flow chart showing varietal testing system in AICRP on wheat and Barley
The impact of wheat varieties is immense in Indian agriculture and helped country to increase ten-fold wheat production from 9.5 million tons in 1963–64 to 108.75 million tons in 2021–21. Thus, it has contributed in un-precedential growth in wheat production could have been possible because of continuous replacement of old varieties with high yielding improved varieties (Table 2.4) as a result of concerted efforts of wheat researchers and a systematic and effective seed replacement mechanism. In nut shell, this system is very unique and un-parallel that has been very effective for development and deployment of high yielding, rust resistant and end product specific quality wheat meet food and nutritional security of the country. The importance of this system of evaluation in different trials and nurseries in India’s wheat improvement programme is reflected in the effective management of Wheat and Barley rusts in India through the deployment of diverse rust resistant wheat varieties based on the pathotype distribution in different areas.
2.16 Conclusions
Breeding programmes in India have made significant progress, and achievements are reflected in terms of record wheat production of 108.75 million tons during 2021 as compared to 12.3 million tons in 1965. Deployment of high yielding, resistant varieties through the use of both cultivated and wild germplasm have led to significant improvement in yield potential, and breeding programme have been able to contain the losses caused by biotic and abiotic stresses. Nutrient rich varieties have been developed and released to minimise the micronutrient malnutrition. Varieties having specific traits which are required for industrial products like Chapatti, bread, biscuit, pasta, macaroni, noodles, etc. have been released. The wheat crop is also facing challenges of abiotic stresses particularly heat and drought. Although several heat tolerant varieties have been released but needs further improvements in the traits to breed better climate resilience. The natural resources like land and water are shrinking due to urbanisation and depletion of ground water due to excessive use. Varieties having high nutrient, water and radiation use efficiency, and productivity needs to be targeted so that same or higher production levels can be achieved with minimum use of these resources. The varietal replacement rate with the deployment of new varieties has been improved in the recent decade in comparison to the past. Many high yielding and disease resistant varieties have been released for different zones of the country but the life span of varieties is usually considered to be 3–5 years due to the breakdown of deployed resistance. The recent advances in genomics like GWAS, genomic selection, speed breeding and phenotyping platforms need to be adopted in the breeding programmes for better understanding and dissecting of complex traits. These technologies can assist the conventional breeding approaches to breed high yielding, nutrient rich and climate resilient varieties with a higher precision. Besides record production the country achieved, there is a continuous need to sustain and further improve the productivity in India to feed the growing population.
References
Abhinandan K, Skori L, Stanic M et al (2018) Abiotic stress signaling in wheat—an inclusive overview of hormonal interactions during abiotic stress responses in wheat. Front Plant Sci 9:734. https://doi.org/10.3389/fpls.2018.00734
Akhunov ED, Goodyear AW, Geng S et al (2003) The organization and rate of evolution of wheat genomes are correlated with recombination rates along chromosome arms. Genome Res 13:753–763. https://doi.org/10.1101/gr.808603
Anh VL, Anh NT, Tagle AG, Vy TTP, Inoue Y, Takumi S, Chuma I, Tosa Y (2015) Rmg8, a new gene for resistance to Triticum isolates of Pyricularia oryzae in hexaploid wheat. Phytopathology 105:1568–1572
Al-Karaki GN (2012) Phenological development-yield relationships in durum wheat cultivars under late-season high-temperature stress in a semiarid environment. ISRN Agron 2012:1–7
Al-Khatib K, Paulsen GM (1990) Photosynthesis and productivity during high-temperature stress of wheat genotypes from major world regions. Crop Sci 30(5):1127
Balota M, Payne WA, Evett SR (2008) Morphological and physiological traits associated with canopy temperature depression in three closely related wheat lines. Crop Sci 48(5):1897–1910
Bansal M, Kaur S, Dhaliwal HS et al (2017) Mapping of Aegilops umbellulata -derived leaf rust and stripe rust resistance loci in wheat. Plant Pathol 66:38–44. https://doi.org/10.1111/ppa.12549
Beche E, Benin D, Da Silva CL et al (2014) Genetic gain in yield and changes associated with physiological traits in Brazilian wheat during the 20th century. Eur J Agron 61:49–59. https://doi.org/10.1016/j.eja.2014.08.005
Becker E, Schmidhalter U (2017) Evaluation of yield and drought using active and passive spectral sensing systems at the reproductive stage in wheat. Front Plant Sci 8:379
Berry PM, Kendall S, Rutterford Z, Orford S, Griffiths S (2015) Historical analysis of the effects of breeding on the height of winter wheat (Triticum aestivum) and consequences for lodging. Euphytica 203:375–383. https://doi.org/10.1007/s10681-014-1286-y
Bhatta M, Morgounov A, Belamkar V et al (2019) Genome-wide association study for multiple biotic stress resistance in synthetic hexaploid wheat. Int J Mol Sci 20:3667–3682
Blum A, Ebercon A (1981) Cell membrane stability as a measure of drought and heat tolerance in wheat 1. Crop Sci 21(1):43–47
Bowden WM (1959) The taxonomy and nomenclature of the wheats, barleys, and ryes and their wild relatives. Can J Bot 37:657–684
Bustos DV, Hasan AK, Reynolds MP et al (2013) Combining high grain number and weight through a DH-population to improve grain yield potential of wheat in high-yielding environments. Field Crop Res 145:106–115. https://doi.org/10.1016/j.fcr.2013.01.015
Cakmak I, Torun A, Millet E et al (2004) Triticum dicoccoides: an important genetic resource for increasing zinc and iron concentration in modern cultivated wheat. Soil Sci Plant Nutr 50:1047–1054
Calderini DF, Reynolds MP (2000) Changes in grain weight as a consequence of de-graining treatments at pre- and post-anthesis in synthetic hexaploid lines of wheat (Triticum durum × T). Funct Plant Biol 27:183–191. https://doi.org/10.1071/PP99066
Cardoso CAA, Reis EM, Moreira EN (2008) Development of a warning system for wheat blast caused by Pyriculariagrisea. Summa Phytopathol 34:216–221
Guzman C, Ammar K, Govindan V (2019) Genetic improvement of wheat grain quality at CIMMYT. Front Agric Sci Eng 6(3):265–272
Changhai S, Baodi D, Yunzhou Q et al (2010) Physiological regulation of high transpiration efficiency in winter wheat under drought conditions. Plant Soil Environ 56(7):340–347
Chatrath R, Tiwari V, Gupta V et al (2018) WB 2: a high yielding bread wheat variety for irrigated timely sown conditions of North Western Plains zone of India. Wheat Barley Res 10(1):40–44
Chen H, Moakhar NP, Iqbal M (2016) Genetic variation for flowering time and height reducing genes and important traits in western Canadian spring wheat. Euphytica 208:377–390. https://doi.org/10.1007/s10681-015-1615-9
Chen X, Hao MD (2015) Low contribution of photosynthesis and water-use efficiency to improvement of grain yield in Chinese wheat. Photosynthetica 53:519–526. https://doi.org/10.1007/s11099-015-0147-9
Chhuneja P, Kaur S, Goel RK et al (2008) Transfer of leaf rust and stripe rust resistance from Aegilops umbellulata Zhuk. To bread wheat (Triticum aestivum L.). Genet Resour Crop Evol 55:849–859. https://doi.org/10.1007/s10722-007-9289-3
Clayton WD, Renvoize SA (1986) Genera graminum. Bull. Add. Ser, Kew, p 13
Collard BC, Mackill DJ (2008) Marker-assisted selection: an approach for precision plant breeding in the twenty-first century. Philos Trans R Soc B 363:557–572. https://doi.org/10.1098/rstb.2007.2170
Colmer TD, Flowers TJ, Munns R (2006) Use of wild relatives to improve salt tolerance in wheat. J Exp Bot 57(5):1059–1078. https://doi.org/10.1093/jxb/erj124
Cooper M, Gho C, Leafgren R et al (2014) Breeding drought-tolerant maize hybrids for the US corn-belt. Discovery to product. J Exp Bot 65(21):6191–6204. https://doi.org/10.1093/jxb/eru064
Cox TS, Raupp WJ, Wilson DL et al (1992) Resistance to foliar diseases in a collection of Triticum tauschii germplasm. Plant Dis 76:1061–1064. https://doi.org/10.1094/pd-76-1061
Crossa J, de los Campos G, Perez P et al (2010) Prediction of genetic values of quantitative traits in plant breeding using pedigree and molecular markers. Genetics 186:713–724. https://doi.org/10.1534/genetics.110.118521
Crossa J, Perez-Rodriguez P, Cuevas J et al (2017) Genomic selection in plant breeding: methods, models, and perspectives. Trends Plant Sci 22(11):961–975. https://doi.org/10.1016/j.tplants.2017.08.011
Cruz CD, Magarey RD, Christie DN, Fowler GA, Fernandes JM, Bockus WW, Valent B, Stack JP (2016) Climate suitability for Magnaporthe oryzae Triticum pathotype in the United States. Plant Dis 100:1979–1987
Cumagun CJR, Anh VL, Vy TTP, Inoue Y, Asano H, Hyon GS, Chuma I, Tosa Y (2014) Identification of a hidden resistance gene in tetraploid wheat using laboratory strains of Pyricularia oryzae produced by backcrossing. Phytopathology 104:634–640
Damania A (1996) Biodiversity conservation: a review of options complementary to standard ex situ methods. Plant Genetic Res Newsl 107(3):1–18
Damania AB (1998) Diversity of major cultivated plants domesticated in the near east. In: Damania AB, Valkoun J, Willcox G et al (eds) The origins of agriculture and crop domestication. Proceedings of the Harlan Symposium. ICARDA, Aleppo, pp 51–64
Damour G, Simonneau T, Cochard H et al (2010) An overview of models of stomatal conductance at the leaf level. Plant Cell Environ 33(9):1419–1438
de Lima CZ, Buzan JR, Moore FC et al (2020) Heat stress on agricultural workers exacerbates crop impacts of climate change. Environ Res Lett 16(4):044020
De Pace C et al (2011) In: Kole C (ed) Dasypyrum. Wild crop relatives: genomic and breeding resources, cereals. Springer, Berlin
de Vries AP (1971) Flowering biology of wheat, particularly in view of hybrid seed production—a review. Euphytica 20:152–170
Distelfeld A, Cakmak I, Peleg Z (2007) Multiple QTl-effects of wheat Gpc-B1 locus on grain protein and micronutrient concentrations. Plant Physiol 129:635–643
Dorofeev VF, Filatenko AA, Migushova EF (1979) Wheat. In: DorofeevVF IL, Korovina ON (eds) Flora of cultivated plants. Leningrad, Russia, vol vol. 1
Dubin HJ, Bimb HP (1994) Studies of soil borne diseases and foliar blights of wheat at the National Wheat Research Experiment Station, Bhairahawa, Nepal. In: Wheat special report no 36. CIMMYT, Mexico
Dvorak J, Harvey BL, Coulman BE (1973) The use of nitrous oxide for producing eupolyploids and aneuploids in wheat and barley. Can J Genet Cytol 15:205–214
Dvorak J, Luo MC (2001) Evolution of free-threshing and hulled forms of Triticum aestivum: old problems and new tools. In: Caligari PDS, Brandham PE (eds) Wheat taxonomy: the legacy of John Percival. Academic, London, pp p127–p136
Dvorak J, Diterlizzi P, Zhang HB et al (1993) The evolution of polyploidy wheats: identification of the a genome donor species. Genome 36:21–31
Dwyer L, Tollenaar M, Houwing L (1991) A nondestructive method to monitor leaf greenness in corn. Can J Plant Sci 71(2):505–509
Dyck PL, Kerber ER (1970) Inheritance in hexaploid wheat of adult-plant leaf rust resistance derived from Aegilops squarrosa. Can J Genet Cytol 12:175–180. https://doi.org/10.1139/G70-025
Edwards MA, Osborne BG, Henry RJ (2010) Puroindoline genotype, starch granule size distribution and milling quality of wheat. J Cereal Sci 52(2):314–320
Endo TR (1988) Induction of chromosomal structural changes by a chromosome of Aegilops cylendrica L. in common wheat. J Hered 79:366–370
Ennahli S, Earl HJ (2005) Physiological limitations to photosynthetic carbon assimilation in cotton under water stress. Crop Sci 45(6):2374–2382
Eyal Z, Levy E (1987) Variations in pathogenicity patterns of Mycosphaerella graminicola within Triticum spp. in Israel. Euphytica 36:237–250
Farooq M, Wahid A, Kobayashi N et al (2009) Plant drought stress: effects, mechanisms and management. Sust Agric 4:153–188
Feldman M, Levy AA (2015) Origin and evolution of wheat and related triticeae species. In: Molnar M, Ceoloni C, Dolezel J (eds) Alien introgression in wheat. Springer International Publishing, Switzerland, pp 21–72
Fletcher A, Christopher J, Hunter M et al (2018) A low-cost method to rapidly and accurately screen for transpiration efficiency in wheat. Plant Methods 14(1):77
Fokar M, Nguyen H, Blum A (1998) Heat tolerance in spring wheat. I. Genetic variability and heritability of cellular thermotolerance. Euphytica 104(1):1–8
Friebe B, Jiang J, Raupp WJ et al (1996) Characterization of wheat-alien translocations conferring resistance to diseases and pests: current status. Euphytica 91:59–87
Gaju O, Reynolds MP, Sparkes DL et al (2009) Relationships between large-spike phenotype, grain number, and yield potential in spring wheat. Crop Sci 49:961–973. https://doi.org/10.2135/cropsci2008.05.0285
Gao F, Ma D, Yin G et al (2017) Genetic progress in grain yield and physiological traits in Chinese wheat cultivars of southern Yellow and Huai Valley since 1950. Crop Sci 57:760–773. https://doi.org/10.2135/cropsci2016.05.0362
Gill BS, Friebe BR, White FF (2011) Alien introgressions represent a rich source of genes for crop improvement. Proc Natl Acad Sci U S A 108:7657–7658
Gill BS, Kimber G (1974) Giemsa C-banding and the evolution of wheat. Proc Natl Acad Sci U S A 71:4086–4090
Gill HS, Li C, Sidhu JS et al (2019) Fine mapping of the wheat leaf rust resistance gene Lr42. Int J Mol Sci 20:2445–2457. https://doi.org/10.3390/ijms20102445
Gill KS, Gill BS, Endo TR et al (1996) Identification and high density mapping of gene-rich regions in chromosome group 5 of wheat. Genetics 143:1001–1012
Goddard R, Steed A, Chinoy C et al (2020) Dissecting the genetic basis of wheat blast resistance in the Brazilian wheat cultivar BR 18-Terena. BMC Plant Biol 20(1). https://doi.org/10.1186/s12870-020-02592-0
Gomez D, Vanzetti L, Helguera M et al (2014) Effect of Vrn-1, Ppd-1 genes and earliness per se on heading time in Argentinean bread wheat cultivars. Field Crop Res 158:73–81. https://doi.org/10.1016/j.fcr.2013.12.023
Goncharov NP (2011) Genus Triticum L. taxonomy: the present and the future. Plant Syst Evol 295:1–11. https://doi.org/10.1007/s00606-011-0480-9
Gorjanc G, Gaynor RC, Hickey JM (2018) Optimal cross selection for long-term genetic gain in two-part programs with rapid recurrent genomic selection. Theor Appl Genet. https://doi.org/10.1007/s00122-018-3125-3
Guo J, Yang Y, Wang G et al (2010) Ecophysiological responses of Abies fabri seedlings to drought stress and nitrogen supply. Physiol Plant 139(4):335–347
Gupta PK (1991) Cytogenetics of wheat and its close wild relatives—Triticum and Aegilops. In: Gupta PK, Tsuchiya T (eds) Chromosome engineering in plants: genetics, breeding, evolution. Elsevier, Amsterdam
Gupta NK, Agarwal S, Agarwal VP (2013) Effect of short-term heat stress on growth, physiology and antioxidative defence system in wheat seedlings. Acta Physiol Plant 35(6):1837–1842
Gustafson P, Raskina O, Ma X et al (2009) Wheat evolution, domestication, and improvement. In: Carver BF (ed) Wheat: science and trade. Wiley, Danvers, pp 5–30
Hammouda AM (2003) First report of sharp eyespot of wheat in Egypt. Plant Dis 87(5):598
Hasterok R, Draper J, Jenkins J (2004) Laying the cytotaxonomic foundations of new model grass, Brachypodium distachyon (L.) Beauv. Chromosome Res 12:397–403
Hatfield JL, Dold C (2019) Water-use efficiency: advances and challenges in a changing climate. Front Plant Sci 10:103
Helguera M, Khan IA, Kolmer J, Lijavetzky D, Zhong-qi L, Dubcovsky J (2003) PCR assays for the Lr37-Yr17-Sr38 cluster of rust resistance genes and their use to develop isogenic hard red spring wheat lines. Crop Sci 43:1839–1847. https://doi.org/10.2135/cropsci2003.1839
Heun M, Pregal-Schafer R, Klawan D et al (1997) Site of einkorn wheat domestication identified by DNA finger printing. Science 278:131–1314
Hoisington D, Khairallah M, Reeves T et al (1999) Plant genetic resources: what can they contribute toward increased crop productivity? Proc Natl Acad Sci U S A 96:5937–5943. https://doi.org/10.1073/pnas.96.11.5937
Horvath P, Barrangou R (2010) CRISPR/Cas, the immune system of bacteria and archaea. Science 327:167–170. https://doi.org/10.1126/science.1179555
Huerta-Espino J, Singh RP, Germán S (2011) Global status of wheat leaf rust caused by Puccinia triticina. Euphytica 179:143–160. https://doi.org/10.1007/s10681-011-0361-x
IFPRI (2009) Impact of climate change on agriculture–factsheet on Asia. IFPRI
IPCC (Intergovernmental Panel on Climate Change) (2007) Intergovernmental panel on climate change fourth assessment report: climate change 2007. Synthesis report. World Meteorological Organization, Geneva
Jain N, Singh G, Yadav R (2014) Root trait characteristics and genotypic response in wheat under different water regimes. Cereal Res Commun 42(3):426–438
Janjua PZ, Samad G, Khan NUM (2010) Impact of climate change on wheat production: a case study of Pakistan. Pak Dev Rev 1:799–822
Johnson BL, Dhaliwal HS (1976) Reproductive isolation of Triticum boeticum and Triticum urartu and the origin of the tetraploid wheats. Am J Bot 63:1088–1094
Joshi A, Mishra B, Chatrath R et al (2007) Wheat improvement in India: present status, emerging challenges and future prospects. Euphytica 157(3):431–446
Joshi AK, Mishra B, Prashar M et al (2008) Ug99 race of stem rust pathogen: challenges and current status of research to sustain wheat production in India. Indian J Genet 68(3):231–241
Kerber ER, Dyck PL (1973) Inheritance of stem rust resistance transferred from diploid wheat (Triticum monocooum) to tetraploid and exaploid wheat and chromosome location of the gene involved. Can J Genet Cytol 15:397–409
Kihara H (1919) Uber cytologische Studien bei einigen Getreidearten. Mit. I. Spezes-bastard des Weizen und weizenroggen-bastarde. Bot Mag 33:17–38
Kihara H (1944) Discovery of the DD-analyser, one of the ancestors of Triticum vulgare. Agric Hortic 19:13–14
Kimber G (1974) A reassessment of the origin of the polyploid wheats. Genetics 78(1):487–492
King JE, Cook RJ, Melville SC (1983) A review of Septoria diseases of wheat and barley. Ann Appl Biol 103:345–373
Knott DR, Anderson RG (1956) The inheritance of rust resistance I. The inheritance of stem rust resistance in ten varieties of common wheat. Can J Agric Sci 36:174–195
Kolmer J, Long D, Hughes M (2010) Physiologic specialization of Puccinia triticina on wheat in the United States in 2008. Plant Dis 94:775–780. https://doi.org/10.1094/pdis-94-6-0775
Krishnappa G, Savadi S, Tyagi BS (2021) Integrated genomic selection for rapid improvement of crops. Genomics 113:1070–1086. https://doi.org/10.1016/j.ygeno.2021.02.007
Kumar P, Gupta V, Singh G et al (2021) Assessment of terminal heat tolerance based on agro-morphological and stress selection indices in wheat. Cereal Res Commun 49:217–226. https://doi.org/10.1007/s42976-020-00112-2
Kuspira J, Unrau J (1957) Genetic analysis of certain characters in common wheat using whole chromosome substitution lines. Can J Plant Sci 37:300–326
Lagudah E, Dubcovsky J, Powell W (2001) Wheat genomics. Plant Physiol Biochem 39:1–10
Lawal BA, Raji IA, Egedegbe O et al (2018) Prediction of nitrogen application in maize based on the normalised difference vegetation index (NDVI). Asian J Soil Sci Plant Nutr 1:1–13
Li H, Rasheed A, Hickey LT et al (2018) Fast-forwarding genetic gain. Trends Plant Sci 23(3):184–186. https://doi.org/10.1016/j.tplants.2017.12.008
Lim PO, Kim HJ, Gil Nam H (2007) Leaf senescence. Annu Rev Plant Biol 58:115–136
Linnaeus C (1753) Species plantarum. Printed for the Ray Society, London
Liu F, Jensen CR, Shahanzari A et al (2005) ABA regulated stomatal control and photosynthetic water use efficiency of potato (Solanum tuberosum L.) during progressive soil drying. Plant Sci 168(3):831–836
Lobell DB, Field CB (2007) Global scale climate–crop yield relationships and the impacts of recent warming. Environ Res Lett 2(1):014002
Lobell DB, Sibley A, Ivan Ortiz-Monasterio J (2012) Extreme heat effects on wheat senescence in India. Nat Climate Change 2(3):186–189
Lopes MS, Reynolds MP (2012) Stay-green in spring wheat can be determined by spectral reflectance measurements (normalized difference vegetation index) independently from phenology. J Exp Bot 63(10):3789–3798
Ludlow M, Muchow R (1990) A critical evaluation of traits for improving crop yields in water-limited environments. Adv Agron 43:107–153
Lusser M, Parisi C, Plan D (2012) Deployment of new biotechnologies in plant breeding. Nat Biotechnol 30:231–239
Mac Key J (1966) Species relationship in Triticum. In: Proceedings of 2nd international wheat genetics symposium Hereditas Suppl, vol vol. 2, pp 237–276
Mac Key J (1977) Diccocoidea Flaksb. Of wheat, its phylogeny, diversification and subdivision. In: Proceedings of symposium on extended availability of wheat genetics resources, Bari, pp 5–46
Maia N (1967) Obtention de blés tendres résistants au piétin-verse (Cercosporella herpotrichoides) par croisements interspécifiques. C R Acad Sci Fr 53:149–154
Mamrutha HM, Sharma D, Kumar KS (2017) Influence of diurnal irradiance variation on chlorophyll values in wheat: a comparative study using different chlorophyll meters. Nat Acad Sci Lett 40(3):221–224
Mason RE, Mondal S, Beecher F (2010) QTL associated with heat susceptibility index in wheat (Triticum aestivum L.) under short-term reproductive stage heat stress. Euphytica 174:423–436. https://doi.org/10.1007/s10681-010-0151-x
Matsuoka Y (2011) Evolution of polyploid triticum wheats under cultivation: the role of domestication, natural hybridization and allopolyploid speciation in their diversification. Pl Cell Physiol 52:750–764
McFadden ES (1930) A successful transfer of emmer characters to vulgare wheat. J Am Soc Agron 22:1020–1034. https://doi.org/10.2134/agronj1930.00021962002200120005x
McFadden ES, Sears ER (1946) The origin of Triticum spelta and its free-threshing hexaploid relatives. J Hered 37:107–116. https://doi.org/10.1093/oxfordjournals.jhered.a105594
Mcintosh RA (1988) Catalogue of gene symbols for wheat. In: Miller TE, Koebner RMD (eds) Proceedings of the seventh international wheat genetics symposium, Cambridge, England, 13–19 July 1988. Institute of Plant Science Research, Cambridge, Engfand, pp 1225–1323
McIntosh RA et al (1984) Cytogenetical studies in wheat. XIII. Sr35—a third gene from Triticum monococcum for resistance to Puccinia graminis tritici. Z Pflanzenzucht 92:1–14
McIntosh RA, Gyarfas J (1971) Triticum timopheevi as a source of resistance to wheat stem rust. Z Pflanzenzucht 66:240–248
McIntosh RA, Hart GE, Devos KM et al (1998) Catalogue of gene symbols for wheat. In: Slinkard AE (ed) Proc 9th intern wheat genet Symp, Saskatoon, Canada (2–7 august, 1998), vol vol. 5. University Extension Press, Saskatchewan, pp 1–236. 1998
McIntosh RA, Wellings CR, Park RF (1995) Wheat rusts. An atlas of resistance genes. CSIRO Publishing, Melbourne
Meena RP, Karnam V, Rinki et al (2019) Identification of water use efficient wheat genotypes with high yield for regions of depleting water resources in India. Agril Water Manag 223:105709
Meilleur B, Hodgkin T (2004) In situ conservation of crop wild relatives: status and trends. Biodivers Conserv 13:663–684
Mitra R, Bhatia CR (1973) Repeated and non-repeated nucleotide sequences in diploid and polyploid wheat species. Heredity 31:251–262
Mondal S, Singh RP, Crossa J et al (2013) Earliness in wheat: a key to adaptation under terminal and continual high temperature stress in South Asia. Field Crop Res 151:19–26
Mondal S, Rutkoski JE, Velu G, Singh PK, Crespo-Herrera LA, Guzmán C, Bhavani S, Lan C, He X, Singh RP (2016) Harnessing diversity in wheat to enhance grain yield, climate resilience, disease and insect pest resistance and nutrition through conventional and modern breeding approaches. Front Plant Sci 7:991. https://www.frontiersin.org/article/10.3389/fpls.2016.00991. https://doi.org/10.3389/fpls.2016.00991
Moore G, Gale MD, Kurata N, Flavell RW (1993) Molecular analysis of small grain cereal genomes: current status and prospects. Biotechnology 11:584–589
Morris R, Sears ER (1967) The cytogenetics of wheat and its relatives. In: Quisenberry KS, Reitz LP (eds) Wheat and wheat improvement. Am Soc Agron Monographs, Madison, WI, pp 18–97
Mujeeb-Kazi A, Gul A, Farooq M et al (2008) Rebirth of synthetic hexaploids with global implications for wheat improvement. Aust J Agr Res 59:391–398. https://doi.org/10.1071/AR07226
Mullan DJ, Reynolds MP (2010) Quantifying genetic effects of ground cover on soil water evaporation using digital imaging. Funct Plant Biol 37(8):703–712
Mulsanti IW, Yamamoto T, Ueda T et al (2018) Finding the superior allele of japonica-type for increasing stem lodging resistance in indica rice varieties using chromosome segment substitution lines. Rice 11:25
Mythili G, Goedecke J (2016) Economics of land degradation in India. In: Nkonya E, Mirzabaev A, von Braun J (eds) Economics of land degradation and improvement- a global assessment for sustainable development. Springer, Cham, pp 431–469
Nahar K, Ahamed KU, Fujita M (2010) Phenological variation and its relation with yield in several wheat (Triticum aestivum L.) cultivars under normal and late sowing mediated heat stress condition. Not Sci Biol 2(3):51–56
Nakhforoosh A, Bodewein T, Fiorani F (2016) Identification of water use strategies at early growth stages in durum wheat from shoot phenotyping and physiological measurements. Front Plant Sci 7:1155. https://doi.org/10.3389/fpls.2016.01155/full
Nataraja K, Jacob J (1999) Clonal differences in photosynthesis in Hevea brasiliensis Müll. Arg Photosynthetica 36(1–2):89
Nayar SK, Prashar M, Bhardwaj SC (1996) Occurence of Yr9 virulence of yellow rust and its management in Indian context. In: Proc 2nd intern. crop science congress 17–24 November, 1996, New Delhi, p 127
Nga N, Hau VT, Tosa Y (2009) Identification of genes for resistance to a Digitaria isolate of Magnaporthe grisea in common wheat cultivars. Genome 52:801–809. https://doi.org/10.1139/G09-054
Ogbonnaya FC, Abdalla O, Mujeeb-Kazi A (2013) Synthetic hexaploids: harnessing species of the primary gene pool for wheat improvement. Plant Breed Rev 37:35–122
Okada T, Jayasinghe JEAM, Nansamba M et al (2018) Unfertilized ovary pushes wheat flower open for cross-pollination. J Exp Bot 69:399–412. https://doi.org/10.1093/jxb/erx410
Osakabe K, Osakabe Y, Toki S (2010) Site-directed mutagenesis in Arabidopsis using custom- designed zinc finger nucleases. Proc Natl Acad Sci U S A 107:12034–12039
Paltridge NG, Milham PJ, Ortiz-Monasterio JI et al (2012) Energy-dispersive X-ray fluorescence spectrometry as a tool for zinc, iron and selenium analysis in whole grain wheat. Plant Soil 361:261–269. https://doi.org/10.1007/s11104-012-1423-0
Parry MA, Hawkesford MJ et al (2010) Genetic approaches to reduce greenhouse gas emissions: increasing carbon capture and decreasing environmental impact. Clim Change Crop Prod 1:139–150
Paymard P, Yaghoubi F, Nouri M et al (2019) Projecting climate change impacts on rainfed wheat yield, water demand, and water use efficiency in Northeast Iran. Theor Appl Climatol 138(3–4):1361–1373
Percival J (1921) The wheat plant. Duckworth, London
Petersen G, Seberg O, Yde M et al (2006) Phylogenetic relationships of Triticum and Aegilops and evidence for the origin of the A, B, and D genomes of common wheat (Triticum aestivum). Mol Phylogenet Evol 39:70–82
Pietragalla J, Vega AM (2012) Normalized difference vegetation index. In: Physiological breeding II: a field guide to wheat phenotyping. CIMMYT, Mexico, pp 37–40
Pittock B, Arthington A, Booth T (2003) Climate change: an Australian guide to the science and potential impacts. Australian Greenhouse Office
Prasad PVV, Djanaguiraman M (2014) Response of floret fertility and individual grain weight of wheat to high temperature stress: sensitive stages and thresholds for temperature and duration. Funct Plant Biol 41(12):1261
Qi LL, EchalierB CS et al (2004) A chromosome bin map of 16 000 expressed sequence tag loci and distribution of genes among the three genomes of polyploid wheat. Genetics 168:701–712. https://doi.org/10.1534/genetics.104.034868
Rai N, Amasiddha B, Prashant Kumar KC et al (2018) Marker assisted backcross breeding (MABB) for improvement of drought tolerance in bread wheat (Triticum aestivum L. emThell). Plant Breed 137:514–552. https://doi.org/10.1111/pbr.12605
Ram RB, Singh SS, Ahamed ML et al (2005) Genetic analysis for adult plant resistance to leaf rust in synthetic hexaploid wheats derived from Triticum turgidum (AABB) x T. tauschii (DD). Indian Phytopathol 58:149–152
Ramadas S, Kiran Kumar TM, Pratap Singh G (2019) Wheat production in India: trends and prospects. In: Recent advances in grain crops research. IntechOpen. https://www.intechopen.com/online-first/wheat-production-in-india-trends-and-prospects
Rasheed A, Jin H, Xiao Y et al (2019) Allelic effects and variations for key bread-making quality genes in bread wheat using high-throughput molecular markers. J Cereal Sci 85:305–309
Rashid A, Stark J, Tanveer A et al (1999) Use of canopy temperature measurements as a screening tool for drought tolerance in spring wheat. J Agron Crop Sci 182(4):231–238
Rawat N, Tiwari VK, Neelam K (2009) Development and characterization of Triticum aestivum-Aegilops kotschyi amphiploids with high grain iron and zinc contents. Plant Genet Resour Charact Util 7:271–280
Reader SM, Miller TE (1991) The introduction into bread wheat of a major gene for resistance to powdery mildew from wild emmer wheat. Euphytica 53:57–60
Rebetzke G, Condon AG, Richards R et al (2002) Selection for reduced carbon isotope discrimination increases aerial biomass and grain yield of rainfed bread wheat. Crop Sci 42(3):739–745
Rebetzke GJ, Bonnett DG, Ellis MH (2012b) Combining gibberellic acid-sensitive and insensitive dwarfing genes in breeding of higher-yielding, sesqui-dwarf wheats. Field Crop Res 127:17–25. https://doi.org/10.1016/j.fcr.2011.11.003
Rebetzke GJ, Ellis MH, Bonnett DG (2012a) Height reduction and agronomic performance for selected gibberellin-responsive dwarfing genes in bread wheat (Triticum aestivum L.). Field Crop Res 126:87–96. https://doi.org/10.1016/j.fcr.2011.09.022
Reynolds M, Manes Y, Izanloo A (2009) Phenotyping approaches for physiological breeding and gene discovery in wheat. Ann Appl Biol 155:309–320
Richards RA, Rebetzke GJ, Watt M et al (2010) Breeding for improved water productivity in temperate cereals: phenotyping, quantitative trait loci, markers and the selection environment. Funct Plant Biol 37(2):85–97
Roche D (2015) Stomatal conductance is essential for higher yield potential of C3 crops. Crit Rev Plant Sci 34(4):429–453
Roelfs AP, Singh RP, Saari EE (1992) Rust diseases of wheat: concepts and methods of disease management. CIMMYT, Mexico, D.F
Rommens CM (2007) Intragenic crop improvement: combining the benefits of traditional breeding and genetic engineering. J Agric Food Chem 55:4281–4288
Rosenberg O (1909) Cytologische und morphologische Studien an Drosera longgoliu X rotudifolia, vol vol. 43. Kgl Svensk Vet Hand, pp 3–64
Royo C, Ammar K, Alfaro C (2018) Effect of Ppd-1 photoperiod sensitivity genes on dry matter production and allocation in durum wheat. Field Crop Res 221:358–367. https://doi.org/10.1016/j.fcr.2017.06.005
Rouse MN, Jin Y (2011) Stem rust resistance in A-genome diploid relatives of wheat. Plant Dis 95:941–944
Chakarabrty SK, Sharma SP (2018) Breeder seed production in self-pollinated crops published in. In: Techniques in seed science and technology by, 3rd edn. Brillion Publishing, New Delhi, pp 1–21
Saadalla M, Shanahan J, Quick J (1990) Heat tolerance in winter wheat: I. hardening and genetic effects on membrane thermostability. Crop Sci 30(6):1243–1247
Safar J, Bartos J, Janda J et al (2004) Dissecting large and complex genomes: flow sorting and BAC cloning of individual chromosomes from bread wheat. Plant J 39:960–968. https://doi.org/10.1111/j.1365-313X.2004.02179.x
Sakamura T (1918) Kurze Mitteilung uber die Chromosomenzahlen und die Verwandtschaftsverhaltnisse der Triticum-Arten. Bot Mag 32:151–154
Sanchez-Garcia M, Alvaro F, Peremarti A (2015) Changes in bread-making quality attributes of bread wheat varieties cultivated in Spain during the 20th century. Eur J Agron 63:79–88
Sax K (1918) The behavior of the chromosomes in fertilization. Genetics 3(4):309–327
Sax K (1922) Sterility in wheat hybrids. II. Chromosome behavior in partially sterile hybrids. Genetics 7:513–552
Schlegel R (1997) Current list of wheats with rye introgression of homoelogous group 1, 2nd update. Wheat Inf Serv 84:64–69
Schouten HJ, Jacobsen E (2008) Cisgenesis and intragenesis, sisters in innovative plant breeding. Trends Plant Sci 13:260–261
Schulz P, Piepenburg K, Lintermann R (2021) Improving plant drought tolerance and growth under water limitation through combinatorial engineering of signalling networks. Plant Biotechnol J 19(1):74
Sears E (1956) The transfer of leaf rust resistance from Aegilops umbellulata to wheat. Brookhaven Symp Biol 9:1–22
Sears ER (1954) The aneuploids of common wheat, vol vol. 572. Univ Mo Agric Exp Stat Bull, pp 1–58
Sears ER (1966a) Nullisomic–tetrasomic combinations in hexaploid wheat. In: Lewis DR (ed) Chromosome manipulation and plant genetics. Oliver and Boyd, London, pp 29–47
Sears ER (1966b) Nullisomic-tetrasomic combinations in hexaploid wheat. In: Riley R, Lewis KR (eds) Chromosome manipulations and plant genetics. 10th International botanical congress. Plenum, New York, pp 29–45
Sears ER (1966c) Chromosome mapping with the aid of telocentrics. Hereditas 2:370–381
Sears ER (1976) Genetic control of chromosome pairing in wheat. Annu Rev Genet 10:31–51. https://doi.org/10.1146/annurev.ge.10.120176.000335
Sears ER (1977) An induced mutant with homoeologous pairing in common wheat. Can J Genet Cytol 19:585–593
Sears ER, Sears LMS (1978) The telocentric chromosomes of common wheat. In: Proc. 5th inter. wheat genet. symp, New Delhi, vol 1, pp 389–407
Sehgal A, Sita K, Siddique KHM et al (2018) Drought or/and heat-stress effects on seed filling in food crops: impacts on functional biochemistry, seed yields, and nutritional quality. Front Plant Sci. https://doi.org/10.3389/fpls.2018.01705/full
Servin B, Martin OC, Mezard M et al (2004) Toward a theory of marker-assisted gene pyramiding. Genetics 168:513–523. https://doi.org/10.1534/genetics.103.023358
Shan Q, Wang Y, Li J et al (2013) Targeted genome modification of crop plants using a CRISPR-Cas system. Nat Biotechnol 31:686–688. https://doi.org/10.1038/nbt.2650
Shan Q, Wang Y, Li J et al (2014) Genome editing in rice and wheat using the CRISPR/Cas system. Nat Protoc 9:2395–2410. https://doi.org/10.1038/nprot.2014.157
Sharma RC, Duveiller E (2003) Effect of stress on Helminthosporium leaf blight in wheat. In: Friesen JBRTL, Ali S (eds) Proc. 4th int. wheat tan spot and spot blotch workshop. North Dakota State University, Fargo, pp 140–144
Sharma S, Xu S, Ehdaie B et al (2011) Dissection of QTL effects for root traits using a chromosome arm-specific mapping population in bread wheat. Theor Appl Genet 122:759–769. https://doi.org/10.1007/s00122-010-1484-5
Sharma D, Tiwari R, Gupta VK et al (2018) Genotype and ambient temperature during growth can determine the quality of starch from wheat. J Cereal Sci 79:240–246
Sharma D, Chandra Pandey G, Mamrutha HM et al (2019) Genotype–phenotype relationships for high-temperature tolerance: an integrated method for minimizing phenotyping constraints in wheat. Crop Sci 59(5):1973–1982
Sharma P, Mehta G, Shefali, Singh SK, Muthusamy SK, Singh GP (2021) Development and validation of heat-responsive candidate gene and mirna gene based ssr markers to analysis genetic diversity in wheat for heat tolerance breeding. Mol Biol Rep 48(1):381–393. https://doi.org/10.1007/s11033-020-06059-1
Shearman VJ, Sylvester-Bradley R, Scott RK, Foulkes MJ (2005) Physiological processes associated with wheat yield progress in the UK. Crop Sci 45:175–185. https://doi.org/10.2135/cropsci2005.0175a
Sheen SJ, Snyder LA (1964) Studies on the inheritance of resistance to six stem rust cultures using chromosome substitution lines of a Marquis wheat selection. Can J Genet Cytol 6:74–82
Simons KJ, Fellers JP, Trick HN et al (2006) Molecular characterization of the major wheat domestication gene Q. Genetics 172:547–555. https://doi.org/10.1534/genetics.105.044727
Singh GP, Sendhil R, Jasrotia P (2019) AICRP on wheat and barley: salient achievements and future directions. Indian J Fert 15(4):80–90
Singh RP, Hodson DP, Jin Y et al (2006) Current status, likely migration and strategies tomitigate the threat to wheat production from race Ug99 (TTKS) of stem rust pathogen. CAB Rev Perspect Agric Veter Sci Nutr Nat Resour 1(54):1–13
Singh RP, AylifHodson DP, Jin Y et al (2015) Emergence and spread of new races of wheat stem rust fungus: continued threat to food security and prospects of genetic control. Phytopathology 105:872–884. https://doi.org/10.1094/PHYTO-01-15-0030-FI
Steduto P, Hsiao TC, Fereres E (2007) On the conservative behavior of biomass water productivity. Irrig Sci 25(3):189–207
Tagle AG, Chuma I, Tosa Y (2015) Rmg7, a new gene for resistance to Triticum isolates of Pyricularia oryzae identified in tetraploid wheat. Phytopathology 105:495–499
Takabayashi N, Tosa Y, Oh HS, Mayama S (2002) A gene-for-gene relationship underlying the species-specific parasitism of Avena/Triticum isolates of Magnaporthe grisea on wheat cultivars. Phytopathology 92(11):1182–1188
Team CW, Pachauri R, Meyer L (2014) IPCC, 2014: climate change 2014: synthesis report. In: Contribution of working groups I II III to fifth assess rep intergov panel clim chang IPCC, Geneva, Switz, p 151
Tembo B, Mulenga RM, Sichilima S et al (2020) Detection and characterization of fungus (Magnaporthe oryzae pathotype Triticum) causing wheat blast disease on rain-fed grown wheat (Triticum aestivum L.) in Zambia. PLoS One 15(9):e0238724. https://doi.org/10.1371/journal.pone.0238724
Tewolde H, Fernandez CJ, Erickson CA (2006) Wheat cultivars adapted to post-heading high temperature stress. J Agron Crop Sci 192(2):111–120
Tomar RSS, Tiwari S, Naik BK (2016) Molecular and morpho-agronomical characterization of root architecture at seedling and reproductive stages for drought tolerance in wheat. PLoS One 11(6):e0156528
Tuberosa R (2012) Phenotyping for drought tolerance of crops in the genomics era. Front Physiol 3:347. https://doi.org/10.3389/fphys.2012.00347
Upadhyay SK, Kumar J, Alok A et al (2013) RNA-guided genome editing for target gene mutations in wheat. G3 (Bethesda) 3:2233–2238. https://doi.org/10.1534/g3.113.008847
Urashima AS, Galbieri R, Stabili A (2005) DNA finger printing ad sexual characterization revealed two distinct populations of Magnaporthe grisea in wheat blast from Brazil. Czech J Genet Plant Breed 41:238–245
Urashima AS, Lavorenti NA, Goulart ACP et al (2004) Resistance spectra of wheat cultivars and virulence diversity of Magnaporthe grisea isolates in Brazil. Fitopatol Bras 29:511–518
van Slageren MW (1994) Wild wheats: a monograph of Aegilops L. In: And Ambylopyrum (Jaub and Spach) Eig (Poaceae). ICARDA and Wageningen Agricultural University, Netherlands
Velu G, Singh R, Huerta-Espino J (2011) Breeding for enhanced zinc and iron concentration in CIMMYT spring wheat germplasm. Czech J Genet Plant Breed 47:S174–S177
Velu G, Singh RP, Balasubramaniam A (2015) Reaching out to farmers with high zinc wheat varieties through public-private partnerships-an experience from eastern-gangetic plains of India. Adv Food Technol Nutr Sci 1:73–75
Velu G, Singh RP, Crespo-Herrera L (2018) Genetic dissection of grain zinc concentration in spring wheat for mainstreaming biofortification in CIMMYT wheat breeding. Sci Rep 8:13526. https://doi.org/10.1038/s41598-018-31951-z
Vermeulen SJ, Campbell BM, Ingram JS (2012) Climate change and food systems. Ann Rev Environ Res 37:195–222
Vijayalakshmi K, Fritz AK, Paulsen GM (2010) Modeling and mapping QTL for senescence-related traits in winter wheat under high temperature. Mol Breed 26(2):163–175
Voss-Fels KP, Cooper M, Hayes BJ (2019) Accelerating crop genetic gains with genomic selection. Theor Appl Genet 132:669–686. https://doi.org/10.1007/s00122-018-3270-8
Vy TTP, Hyon G-S, Nga NTT, Inoue Y, Chuma I, Tosa Y (2014) Genetic analysis of host–pathogen incompatibility between Lolium isolates of Pyricularia oryzae and wheat. J Gen Plant Pathol 80:59–65. https://doi.org/10.1007/s10327-013-0478-y
Wang RRC (2011) Agropyron and Psathyrostachys. In: Kole C (ed) Wild crop relatives: genomic & breeding resources, cereals. Springer-Verlag, Berlin Heidelberg, pp 77–108
Wang Y, Cheng X, Shan Q (2014) Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat Biotechnol 32:947–951. https://doi.org/10.1038/nbt.2969
Wang W, Pan Q, Tian B, He F, Chen Y, Bai G, Akhunova A, Trick HN, Akhunov E (2019) Gene editing of the wheat homologs of TONNEAU1-recruiting motif encoding gene affects grain shape and weight in wheat. Plant J 100:251–264. https://doi.org/10.1111/tpj.14440
Watson A, Ghosh S, Williams MJ (2018) Speed breeding is a powerful tool to accelerate crop research and breeding. Nat Plants 4(1):23–29
Werner JE, Kota RS, Gui BS et al (1992) Distribution of telomerlc repeats and their role in the healing of broken chromosome ends in wheat. Genome 35:844–848
Williamson VM, Thomas V, Ferris H, Dubcovsky J (2013) An Aegilops ventricosa translocation confers resistance against root-knot nematodes to common wheat. Crop Sci 53:1412–1418. https://doi.org/10.2135/cropsci2012.12.0681
Yan L, Loukoianov A, Tranquilli G (2003) Positional cloning of the wheat vernalization gene VRN1. Proc Natl Acad Sci U S A 100:6263–6268. https://doi.org/10.1073/pnas.0937399100
Yu Q, Li L, Luo Q et al (2014) Year patterns of climate impact on wheat yields. Int J Climatol 34:518–528
Yumurtaci A (2015) Utilization of wild relatives of wheat, barley, maize and oat in developing abiotic and biotic stress tolerant new varieties. Emirates J Food Agric 27:1–23. https://doi.org/10.9755/ejfa.v27i1.17852
Zadoks JC (1961) Epidemiology of wheat rusts in Europe. In: FAO plant protection bulletin, vol vol. 13, pp 97–108
Zait Y, Shtein I, Schwartz A (2019) Long-term acclimation to drought, salinity and temperature in the thermophilic tree Ziziphus spina-christi: revealing different tradeoffs between mesophyll and stomatal conductance. Tree Physiol 39(5):701–716
Zhang Y, Xu W, Wang W et al (2016) Progress in genetic improvement of grain yield and related physiological traits of Chinese wheat in Henan Province. Field Crop Res 199:117–128. https://doi.org/10.1016/j.fcr.2016.09.022
Zhan SW, Mayama S, Tosa Y (2008) Identification of two genes for resistance to Triticum isolates of Magnaporthe oryzae in wheat. Genome 51(3):216–221. https://doi.org/10.1139/G07-094
Zheng TC, Zhang XK, Yina GH et al (2011) Genetic gains in grain yield, net photosynthesisand stomatal conductance achieved in Henan Province of China between 1981 and 2008. Field Crop Res 122:225–233. https://doi.org/10.1016/j.fcr.2011.03.015
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive licence to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Krishnappa, G. et al. (2022). Wheat Breeding. In: Yadava, D.K., Dikshit, H.K., Mishra, G.P., Tripathi, S. (eds) Fundamentals of Field Crop Breeding. Springer, Singapore. https://doi.org/10.1007/978-981-16-9257-4_2
Download citation
DOI: https://doi.org/10.1007/978-981-16-9257-4_2
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-16-9256-7
Online ISBN: 978-981-16-9257-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)