
Abstract
Abiotic stresses like drought, cold, salinity, heat, oxidative stress and the presence of excess levels of heavy metal in the agroecosystems lead to a decrease in the growth and productivity of major crops worldwide. The majority of stresses are connected with each other and results in elevated adverse impacts on the plants as well as other important components of the environment. The intensity of stresses and associated adverse impacts are increasing substantially in the era of climate change that again triggers to produce abnormalities in the crops. To overcome the effects of abiotic stresses, a number of strategies have been investigated, such as developing and cultivate stress-tolerant varieties, use of organic fertilizers, and the application of high yielding varieties. Application of biochar to mitigate the impacts of major abiotic stresses especially drought, salinity, and heavy metal has been found very effective. Amendment of biochar in stress affected agroecosystems improves the soil physicochemical and biological features and thereby enhances the productivity of crops. In this chapter, efforts have been made to discuss about three major stresses, i.e. drought, salinity, and heavy metals, their impacts on soil as well as plant productivity. Further, the efficiency and mechanism of biochar in reducing the impacts of stresses when using as a soil amendment have also been discussed thoroughly.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
14.1 Introduction
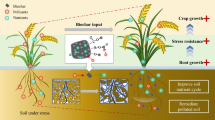
The projected world population by 2050 is from 7 billion (current estimated) to 8.9 billion approximately (Singh et al. 2011), and the food security to this burgeoning number is a paramount concern. The rapid increase in environmental contamination and global climate change is another important threat toward sustainable agriculture. Farmers are under pressure to use excess amounts of chemical fertilizers and pesticides to enhance the agricultural productivity. The application of synthetic fertilizers not only deteriorates the soil health but also enhances the greenhouse gases into the atmosphere as well as contaminates the surface and groundwater. Traditional cropping systems infuse a variety of toxicants to the soil and ultimately enter into the food chain. Abiotic stresses like salinity, acidity, drought, heavy metal, flood, high temperature, freezing, chilling, etc. also damage the agriecosystems, which results in decreased crop yield. These stresses decrease crop productivity by decreasing the photosynthetic pigments, protein denaturation, production of reactive oxygen species (ROS), lipid peroxidation, etc. (Fig. 14.1) (Chodak et al. 2015; Isayenkov and Maathuis 2019; Sallam et al. 2019).

Salinity, drought, and heavy metals are considered as major abiotic stress and reduce a significant portion of agricultural productivity. Plant growth is dependent on the rate and level of photosynthesis, hence environmental stress affecting photosynthesis directly impacts the growth of plant. Salt stress can also influence the synthesis of abscisic acid which when moves to the guard cells, closes the stomata. As a result of the closure of stomata, the rate of photosynthesis declines. The ability of germination and vigor is the most important and primary process in the production of any plant and the adverse impacts of abiotic stress on these parameters are well reported (Bhaskar et al. 2009; Kumar et al. 2009; Singh et al. 2010; Bauddh and Singh 2009, 2011, 2012). Germination being important in the reproduction often influenced by the environment created due to the stress induced by water-scarce conditions (Leila 2007). Water being absorbed by the seed is the first stage in the process of germination. The quantity of water absorbed, mainly dependent on the type, size, and chemical composition of the seed (Rahmani 2006). Generally, soil moisture level lesser than soil’s field capacity is considered favorable for the germination of the seed but in the water-scarce conditions where there is less than desirable water absorption in these seeds are incomplete which can result in diminishing or even stopping the germination process (Leila 2007). Several strategies have been developed to mitigate the impacts of abiotic stresses, especially salinity, drought, and heavy metals. The tools like cultivation of stress-tolerant varieties, application of organic amendments, genetic manipulation, application of microbial inoculants, etc. have been found significantly effective to reduce the adverse impacts of majority of abiotic stresses. Application of biochar has been proven an efficient potential to reduce the adverse impacts of drought, salinity, and heavy metals. Biochar have several other beneficial properties like it retains moisture in soil, balance soil pH, use to make fuel, used in metal remediation from the soil, etc. (Yuan et al. 2019). Several studies have been found that application of biochar to the soil enhances the physicochemical and biological properties of soil and overcome the negative effects created by the stressed conditions.
In the present chapter, the efforts have been done to explore the adverse impacts of three major abiotic stresses, i.e., drought, salinity, and heavy metals. Further, the application of biochar to overcome the impacts of these stresses during the cultivation of crops has also discussed thoroughly.
14.2 Drought, Salinity, and Heavy Metal Stresses
14.2.1 Drought Stress
One of the major concerns of the present time is to cope up with the condition of water scarcity that is arising throughout the world. Water scarcity is among the most catastrophic stress that can severely affect human life both directly and indirectly arising from several other stresses. Drought can be defined as an event of prolonged shortage of water due to insufficient precipitation usually for more than one season (Trenberth et al. 2014). In most of the regions, the phenomenon of drought is a periodic condition and other places can remain in the state of drought for several years. Whenever an area experiences drought, it has a massive effect on life and activities in these areas. Drought also has an impact on hydration of soil, which slowly deteriorates soil quality and also severely affects the animals living in these regions. Some of the climatic factors that are precursors of drought include high wind speeds, high mercury concentrations, etc. Apart from the abovementioned drought, there are other types of drought such as meteorological drought, hydrological drought, and agricultural drought which are described below.
14.2.1.1 Meteorological Drought
It is defined as the drought condition arising due to absence of precipitation in a particular area for a very long time span. However, there is no common consensus among the researchers about a particular value that will define the drought condition like amount of rain or time span of absence of rain. Due to the varying climatic condition, even deserts have drought but their condition is different from the condition of a grassland’s drought.
14.2.1.2 Hydrological Drought
Hydrological drought may be defined as the water deficiency in the surface and subsurface regions, which may affect the supply of water to the water bodies such as freshwater streams, lakes, ponds and most importantly confined water reservoirs and groundwater. Hydrological drought is not a seasonal phenomenon, it occurs and develops over an extended period of time.
14.2.1.3 Agricultural Drought
Agricultural drought is a condition when the water is in such scarce condition that it becomes difficult to sustain the grown crops. This condition usually arises when the expected rainfall is not achieved during essential growth period of crops (this period varies in different crops) and may result in the destruction of crops ranging from mild-to-absolute crop loss. This type of drought can develop very fast if expected rain does not arrive or there is an extensive heat wave, especially during essential growing periods of the crops.
It is a well-known fact that drought condition usually begins with the below-average rainfall in any particular region, although the scarcity of water supply is not the only reason for causing the drought situations. Any kind of activities that deplete the capacity of soil to hold water can be one of the main reasons in the commencement of water scarcity in and around any area. These activities may include over farming of a particular area and extensive deforestation. Both of these activities exploit and deplete the nutrients present in the soil and loosens the soil increasing the soil erosion and destroying the water holding capacity of the soil. This is the prime reason resulting in the desertification of that particular area. On top of that deforestation has been proven to be the reason for reduced rainfall, which adds up to the drought condition already prevailing. When there is a smaller number of trees around any area, vast area of land remains uncovered, which increases water loss directly from the land. All of this can lead to the complete destruction of any kind of forest on its own which further deteriorates the environmental conditions.
14.2.2 Salinity Stress
Salinity is simply the presence of salts such as sodium, magnesium and calcium and bicarbonates in the soil while sodicity refers specifically to the amount of sodium present in the soil. Saline soil may be defined as the soil with an electrical conductivity of at least 4 dS m−1 by measuring the saturated soil extract while sodic soil can be defined as the soil with an exchangeable-sodium percentage (ESP) which is more than 15 (United States Salinity Laboratory Staff 1954). Salinity is of two types; primary salinity and secondary salinity. The salting that results from anthropogenic activities, mainly land development and agriculture is known as secondary salinity. Unrestrained cutting of trees, overgrazing of economic land by animals, and the use of chemical fertilizers are some of the other contributing factors to soil salinity (Lakhdar et al. 2009). Salinity is not bound to rural areas only. Urban salinity occurs generally by the combination of both faulty irrigation-based salinity and due to dry land. Saline soils are mostly dominant in arid and semiarid regions throughout the world, and many productive lands are converted into salt-affected wastelands (Qadir et al. 2000). Globally, the extent of primary salinity is about 955 M ha, whereas the secondary salinization affects 77 M ha, where 58% of the area is affected by irrigation (Metternicht and Zinck 2003). The global estimate of the land affected by salinity is 400 million hectares, which makes up 6% of the total world land area (Arora et al. 2017). The distribution of saline lands is not uniform. The continent of Australia has the highest salinity surface area with 963 million hectares of saline land followed by Asia with about 923 million hectares of saline land (ICARDA 2002). Fifty percent of the arable land is expected to encounter a serious salinity problem by 2050 (Meng et al. 2016).
Classification of salt-affected soil is done on the basis of pH, electrical conductivity (EC), sodium absorption ratio (SAR), and exchangeable sodium percentage (ESP) of the saturated extract (Bohn et al. 2001). Salt-affected soils are thus categorized into saline soil, sodic soil, and saline–sodic soil (Table 14.1).
14.2.3 Heavy Metal Stress
The rampant industrialization and urbanization along with biological and behavioral variations in human, lead to histrionic changes in the environment and climatic patterns. Haggag et al. (2015) discussed feed of devastating population and their food security increased the biotic and abiotic stresses on crop productivity and the environment. Minhas et al. (2017) emphasized on abiotic stresses like salinity, drought, flooding, metal toxicity, nutrient deficiency, high and low temperatures, UV-exposed photo inhibition, air pollution, wind, hail, and gaseous pollutants caused severe damage to the crop productivity and eventually reduced the crop yields. The global per capita food supply and sustainable agricultural food production are the major challenges (Tilman et al. 2002). The technological advancement used an excessive amount of chemical fertilizers such as new cultivars, mineral fertilizers, pesticides, synthetic fertilizers like DDT (1,1′-(2,2,2-trichloroethane-1,1-diyl) bis (4-chlorobenzene)) and polyaromatic hydrocarbons (PAH), etc. these activities increased the concentration of heavy metals in agricultural soil particularly Cd, Pb, As, Zn, Co, Ni, Mn, etc. (Atafar et al. 2010; Chibuike and Obiora 2014). According to Connor et al. (2018), a national soil quality survey in China showed about 19.4% of nations’ agricultural land has been affected by heavy metal contaminations. Heavy metals are naturally present in the soil and geologic as a useful concentration but anthropogenic activities increased the concentration of these elements to amounts that are harmful to plants, animals, and living organisms (Chibuike and Obiora 2014). Plants growing on these heavy metal-containing soils show a reduction in growth, performance, decrease crop yield with quality, and metal accumulations in crops can lead to a potential threat to human health. Chromium toxicity in Bangladesh is increased by the continuous advancement in industrial activities cause cancers to humans through direct contact and other health problems such as respiratory tract and skin problems (Uddin et al. 2017). Some heavy metals are toxic even at low concentrations such as As, Cd, Pb, Cr, and Ni and also carcinogenic by nature (Jan et al. 2015; Mishra and Bharagava 2016 Mishra et al. 2018).
14.3 Adverse Impacts of Drought, Salinity, and Heavy Metal on Soil Quality
14.3.1 Effects of Drought
Drought is one of the worst abiotic stresses which vastly affects the fertility of the soil. It greatly affects the microbial activities, soil respiration, waste decomposition, etc. (Hoogmoed and Sadras 2016; Schmidt et al. 2016). Microbial community and their function in the soil ecosystem plays a very significant role in deciding the fertility and improving the structure of any soil (Wall et al. 2012; Arafat et al. 2017). For example, microbial communities generally mass-produce enzymes that are main reason for 90% of soil organic matter decomposition which in turn helps in C mineralization, this whole process is responsible for the cycling of the nutrients in the soil (Swift et al. 1979). In comparison to fungal properties, the bacterial population is more sensitive to the stress caused due to scarcity of the water and in turn, their reaction adversely impacts the fertility of the soil and soil structure (Bardgett and Wardle 2010).
14.3.1.1 Soil Enzymes
Soil enzymes and the related activities that occur are known to be the indicators of the soil health as they are directly dependent and can give information about microbial and physicochemical status of the soil (Baum et al. 2003). These enzymes are generally synthesized by fungi and bacteria present in the soil and may contain enzymes like ureases, pectinases, phosphatases, and proteases. All these enzymes are very important part and influence soil fertility. Drought is one of the major stresses that can directly and most frequently affect the soil enzymes. Soil enzyme is one of the first affected characteristics during water scarcity thus giving drought the control over nutrient availability and ultimately soil fertility (Sinsabaugh et al. 2013).
14.3.1.2 Microbial Activity
Water un-availability and its stress is one of the primary limiting factor in the survival of microbial community through induced osmotic stress, starvation, and resource competition, thus making the presence of water to be an important factor in the functioning of soil microbial communities and the effect of the stress can be very severe (Griffiths et al. 2003; Sowerby et al. 2005). It has been proven by the scientists from all over the world that soil fertility and its stability is greatly influenced by the microbial community present and they are very sensitive indicators of the environment to stresses like drought (Zornoza et al. 2007). Bacteria are considered to be a major part of any soil microbial community and very important to the fertility of the soil. During the conditions of drought, bacterial community is greatly affected thus limiting the decomposition capacity of the bacteria which may result in a change of structure of soil (Hueso et al. 2012). The availability of water also influences substrate availability and other soil characteristics like its physical characteristics that will affect the population and overall performance (Hueso et al. 2012)
14.3.1.3 Nutrient
Under drought conditions, nutrient uptake through roots as well as transport of nutrients from the roots to shoots is greatly affected mainly due to reduced transpiration rate and the impairment of permeability of membranes (Alam 1999). The drought condition also results in the reduction of the diffusion rate of the nutrients within the soil to the roots (Pinkerton and Simpson 1986).
Nitrogen is one such element which is among the most important mineral for plant growth and is required in subsequent amount as it is a common component of nucleic acid and amino acid. Therefore, the deficiency of nitrogen will immediately cause slow growth in plants. Various researchers have conducted study to establish a relationship between the availability of water and fertilizing property of nitrogen. It has been seen that under water-scarce condition nitrogen fertilizer does not yield good crops and at the same time, increasing water without sufficient supply of nitrogen fertilizer also inhibits growth (Smika et al. 1965). Scientists and researchers have also proven that under drought/water-scarce condition nitrogen-fixing property of leguminous plant is reduced (Streeter 2003). Phosphorus, another important element is a component of phospholipids, dinucleotides, nucleic acids, adenosine triphosphate, and phosphoproteins. Therefore, phosphorus is essential for processes like photosynthesis, transfer and storage of energy, transport of carbohydrates, and regulation of some enzymes. It has been proven that under drought conditions, there are more requirements for adding phosphorus and increased phosphorus uptake into the soil. It is a known fact that under drought or water-scarce condition, phosphorus uptake by plants is reduced (Pinkerton and Simpson 1986), i.e., the transfer and uptake of phosphorus and translocation of phosphorus to the shoots from roots are greatly affected during mildly dry soil conditions (Resnik 1970). In a study by Turner (1985) concluded that in drought-affected areas, phosphorus deficiency is one of the earliest symptoms observed in soil and plants grown in these soils. Therefore, the application of phosphorus fertilizers is required to increase the yield of crops under drought conditions (Garg et al. 2004). Potassium is essential in protein synthesis, photosynthesis, synthesis of glycolytic enzymes, maintaining turgor movements, and also cell expansion (Marschner 1995). Potassium becomes an essential element for the turgor pressure and turgor movement in plants in dry soil conditions. Kuchenbuch et al. (1986) in a study concluded that dry soil reduces the potassium uptake in onion plants. Calcium plays a very important role in maintaining several physiological changes that control growth and other responses to environmental stresses. Some of these physiological changes include solute and water and stomatal closure in a plant. Although calcium uptake decreases under drought conditions same as other elements, but overall calcium uptake by plants is only slightly decreased in comparison to other major elements like nitrogen, phosphorus, potassium, etc. There is not much work done for the effect of drought on elements like magnesium and sulfur. However, studies have shown that drought conditions reduce both magnesium and sulfur uptake (Scherer 2001).
14.3.2 Effect of Salinity on Different Properties of Soil
14.3.2.1 Physical Properties
High level of exchangeable sodium ion can structurally deteriorate the soil that results in the decline of pore volume and also in salt-affected soil it can disrupt the relation between soil–air and soil–water (Rengasamy and Olsson 1991). In sodic soil, slaking, dispersion, and clay swelling are the chief mechanisms that are involved in the breakdown of aggregates as sodicity in the soil affects soil hydraulic conductivity and its infiltration rate (Rengasamy and Sumner 1998). Slaking process causes permanent blockage of the pores even though it is a reversible process while dispersion is an irreversible process that can cause translocation of individual soil particles (Rengasamy and Sumner 1998). This mechanism of soil pore system and its structural stability can be explained by diffuse double layer (DDL) theory, which suggests that the attraction and repulsion between ions in soil particles is a result of intermolecular and electrostatic forces working on them (Quirk 1994). The increase of sodium ion on the exchange site of the clay or soil particles results in the increase of repulsive force which increases the inter-particulate distance turn the breakdown of soil aggregates affecting the soil structure (Oster and Shainberg 2001). Swelling and dispersion in sodium dominated soil bring about changes in the hydraulic properties of the soil.
Low salinity and high sodicity lead to a substantial reduction of hydraulic conductivity and infiltration because of the induced dispersion and swelling (Dikinya et al. 2006). Ensuing slaking and dispersion in the surface layer of sodic soils, a surface crust is formed, which is a thin layer formed after drying. This crust makes the surface soil prone to intense erosion as well as waterlogged conditions (Shainberg et al. 1992).
14.3.2.2 Chemical Properties
Saline and sodic soils have high values of SAR, ESP, EC, and pH. Saline soils normally suffer from deficiencies of NPK while their high pH has an adverse effect on the availability of micronutrients like Al, Fe, Zn, Cu, and Mn (Lakhdar et al. 2009). Salt toxicity, high osmotic pressure as well as soil degradation lead to decline in vegetation growth and decrease in carbon inputs in the soil further deteriorating the physicochemical properties of the soil (Wong et al. 2009). It is also observed that irrigation with sodium-rich water increases the soil sodicity, which results in the decrease of total C and N (Chander et al. 1997). The dispersion of soil aggregates uncovers the confined organic matter, thus increasing the decomposition rate by the soil microbes.
Increase in salinity decreases the mineralization of nitrogen and increases the loss of gaseous NH3 (Paliwal and Gandhi 1976). It was further discovered that in the arid soils with increasing salinity and sodicity even with organic amendment leads to a decrease in the mineralization of C and N (Pathak and Rao 1998). The ability of plants to take up water is reduced by the high ion concentration, primarily Na and Cl ion. This, in turn, affects the plant growth and plant cell (Muneer and Oades 1989).
14.3.2.3 Biological Properties
The changes in soil chemistry negatively affect the soil microbial and biochemical processes, which play a crucial role in maintaining the soil’s ecological functions (Rietz and Haynes 2003). Increasing soil salinity adversely impacts microbial growth and activity, as increased concentration of salt in the soil causes osmotic stress resulting in microbial cell dehydration (Wichern et al. 2006). Other factors, in addition to salt stress, that contributes to the decrease in microbial population and activities in salt-affected soils are Na+ toxicity; nutritional deficiency like Ca2+ deficiency; increase in the level of toxicity of ions such as bicarbonate, carbonate, and chloride; and also significant loss in organic matter resulted by structural degradation (Nelson and Oades 1998). A study was conducted in the arid region of southeast Spain to analyze the effect of salinity on the composition of microbial community in the soil. It was found that the increase in salinity had a negative impact on the microbial community of soil (Garcia et al. 1994). The enzymatic activity, as well as the rate of mineralization of C and N, decreased at high levels of salinity. In a study, the nitrification rate is increased by up to 83% with increase in salinity to 20 dS m−1, which stimulates volatilization of ammonia (Wichern et al. 2006). Soil biochemical and microbial activities are negatively affected by salinity and sodicity induced by irrigation (Rietz and Haynes 2003). Many other studies have also reported that soil microbial biomass and enzyme activities significantly decrease with salinity and sodicity (Tripathi and Srivastava 2006). A shift in the microbial community has been observed with increasing salinity; an adaptive trait to reduce salt stress by lowering the metabolism is noticed (Yuan et al. 2007). Microbial and biochemical activities significantly affect soil ecological function. They play a central role in enhancing the structure of soil as well as in the stabilization of the soil aggregates (Six et al. 2005).
14.3.3 Impact of Heavy Metals
14.3.3.1 Impacts on Soil Physicochemical Quality
Metals exist either as separate entities or in combination with other soil components. Odueze et al. (2017) reported that the soil physicochemical properties affected by heavy metal concentrations present in different layers of soil locations and depth. According to a survey in China by Wang et al. (2018) emphasized on the effects of heavy metal contaminations in agricultural soils concerned to soil physicochemical properties and cultivation age of crop species. Similarly, in Zambia, the physicochemical characteristics of local forest soil were contaminated by the copper mining waste including heavy metals that decreased the pH level (5.05), affect the bulk density (1.24 g/cm3), total organic carbon (2.24%), total nitrogen (0.07%), available P (4.9 mg/kg), K (37.8 mg/kg), Na (49.0 mg/kg), Ca (99.0 mg/kg), and Mg (81.2 mg/kg) (Mutale et al. 2019). As discussed by Obasi et al. (2012), the physicochemical properties of soil are mainly influenced by the mobility and bioavailability of heavy metals. They emphasized that the physicochemical parameters of the soil help to support microbial diversity, metabolic changes, and lastly growth of plant species. The effects of heavy metal on soil physicochemical quality with different levels are shown in Table 14.2.
14.3.3.2 Impact on Soil Biology (Microbial and Enzymatic Activities)
Soil microbes such as bacteria, fungi, algae and protozoa have good metabolic activities for depolymerization and mineralization of organic matter in the forms of N, P, and S. Soil enzyme activities are good indicators of heavy metal toxicity in the soil for bioavailability assessment (Xian et al. 2015). Sobolev and Begonia (2008) reported the effects of Pb on soil microbial community in low (1 ppm) and high (500–2000) concentrations. Kouchou et al. (2017) observed heavy metal contaminations in alkaline soils of the region of Fez (Morocco) and showed adverse impact on actinomycetes and fungi. Atafar et al. (2010) indicated that the microbial biomass in the soil contaminated by Cu, Zn, Pb, and other heavy metals were inhibited drastically. Shi and Ma (2017) observed that Cd contamination affects the microbial activity like respiratory intensity, urease activity, and catalase activity in forest soil and garden soil.
Soil organic matter is the dominant factor contributing to the soil microorganisms and measurements of free soil enzyme activities that can serve as useful indicators of microbial metabolic potential as explained in Table 14.3 (Hagmanna et al. 2015). The research done by Jin et al. (2015) depicted that the heavy metal contaminations adversely affect the soil biological functions, including the size, activity, and diversity, of the soil microbial community and the activity of various enzymes involved in the transformation of C, N, P, and S. Some findings in the review done by Chibuike and Obiora (2014) showed the heavy metal affects the soil microorganism’s quantity, diversity, density, and their activities. The metal toxicity on microorganisms is dependent on several aspects such as pH, soil temperature, organic matter, clay minerals, inorganic anions, and cations, and chemical forms of the metal (Wuana and Okieimen 2011; Chibuike and Obiora 2014). Therefore, the effects of the heavy metals on soil microbial and enzymatic activities are mainly dependent on the soil physiological properties, the stimulus of heavy metals on soil microbial activity, the end product on soil enzyme activity, and the composition of soil microbial community (Khan 2000). Similarly, Khan et al. (2010) found that heavy metals (Pb and Cd) have been an inhibitory impact on soil enzyme activities and as well as microbial community structure. Thus, heavy metals significantly impact on soil enzymes, they restrict the growth and reproduction of microorganisms, reduce the synthesis and metabolism of the microbial enzymes (Chu 2018). The effects of heavy metal on soil microbial diversity may be assessed more comprehensively by using methods such as microbial biomass, C and N mineralization, respiration, other enzymatic activities, etc. (Oijagbe et al. 2019).
14.4 Effects of Drought, Salinity, and Heavy Metals on Growth and Productivity of Crops
14.4.1 Effects of Drought
Drought is a kind of stress that may affect the plants at individual level and even at whole crop level. This extreme stress can cause permanent damage to the plants and has an obvious symptom, e.g., wilting of the leaves. All plants require a particular amount of water for their survival throughout their lifecycle to give better productivity. Plants are well known to possess drought resistance. Other symptoms of drought can be seen by reduction in leaf area, increased leaf drop, reduced leaf growth, sunken stomatal development, yellowing of leaves, and wilting which all lead to the death of the plant (Nakayama et al. 2007).
Water stress inhibits cell enlargement more than cell division. It reduces plant growth by affecting various physiological and biochemical processes, such as photosynthesis, respiration, translocation, ion uptake, carbohydrates, nutrient metabolism, and growth promoters (Jaleel et al. 2008a, b, c, d, e; Farooq et al. 2008).
14.4.1.1 Effect of Drought on Seed Germination
Ability of seed germination and vigor is the most important and primary process in the production of any plant. Germination being important in the reproduction often influenced by environment created due to the stress induced by water-scarce condition (Leila 2007). Water being absorbed by the seed is the first stage in the process of germination. Under drought conditions, osmotic and matric potentials are dramatically decreased which does not allow the required amount of water to reach into plant tissues. The negative effect of drought on plant has been proven by several scientists again and again over time. Zareian (2004) stated that scarcity of water can significantly reduce the germination rate; however, different plant responds differently depending on the stage of germination. Shekari (2000) conducted a study on millet to assess the effect of drought on germination, root and shoot length and concluded that with increasing drought conditions the rate and percent increase in parameters observed reduces considerably. Drought affects many plant growth parameters affecting its overall germination, growth, yield, and production of dry matter.
14.4.1.2 Effects of Drought on Morphological Characters of Plants
Considering plant physiology, scarcity of water causes serious stress on plant growth, yield, and survival. Ghodsi et al. (1998) studied the effect of induced drought on plant and documented 30–50% decrease in yield due to drought and concluded that plant growth is severely affected as a result of high temperature, low humidity, high sun intensity, and high evapotranspiration. Similarly, Bagheri (2009) stated that high temperature due to high light intensity increases plant respiration, photosynthesis, and enzyme activity. Under drought condition, the increased light intensity from the sun increases the photoreaction processes such as photosynthesis and production of free oxygen radicals that ultimately leads to plant death.
Dryness causes the nutrient content of topsoil to be completely absorbed by the plants, which gets stored in fruits whereas salts and other ions get accumulated into the top soil due to increased drought conditions. The increase in ions and salts around the roots of the plants results in osmotic stress, which ultimately causes ion toxicity and death of plants. Under such conditions, the primary response of plants usually is biophysical responses. These responses after drought conditions include loosening and widening of cell walls, decrease in cell volume, decrease in cell pressure that also affects the potential of cell development and if this potential is reduced then growth rate also decreases (Bagheri 2009).
It has been proven by several researchers throughout the world that drought stress is among the worst stress which turns out to be an important limiting factor during the times of development of plants, especially during initial phases of germination (both expansion and elongation) and establishment (Anjum et al. 2003; Kusaka et al. 2005; Bhatt and Rao 2005; Shao et al. 2008).
Most affected plant crops are usually submerged plant crops such as rice and are considered more susceptible to drought in comparison to other plants. In an experiment with soybean plants, stem length was found to be decreased under water-scarce conditions (Specht et al. 2001). In several studies concerning height of the plants, Wu et al. (2008) concluded that water-scarce conditions reduced the plant height of citrus seedlings by almost 25%. Heuer and Nadler (1995) stated that length of the stem is significantly affected due to drought conditions for potato plant and similar kind of results was observed for Petroselinum crispum (parsley) (Petropoulos et al. 2008), (soybean) (Zhang et al. 2004), Vigna unguiculata (Cowpea) (Manivannan et al. 2007), and Abelmoschus esculentus (Okra) (Sankar et al. 2007, 2008).
Under water-scarce conditions, cell growth and cell expansion are reduced due to extremely low turgor pressure whereas increase in osmotic regulation helps in maintaining cell turgor to help the plant grow under water-scarce conditions in plants such as pearl millet (Shao et al. 2008).
Bhatt and Rao (2005) conducted a study on the plant species Abelmoschus esculentus under dry conditions and concluded that reduction in the cell enlargement was responsible for the reduction of average plant height and also for the leaf area and size reduction. A similar study was conducted for assessing the leaf area change in the Populus plant under water-deficit condition by Wullschleger et al. (2005). It was concluded that water stress reduces the leaf growth thereby reducing leaf area in the plant and since the growth of leaf area is important for adequate photosynthesis and production/yield, the result was an overall reduction in the growth rate. Same results, i.e., reduction in growth was observed for Glycine max (soybean) (Zhang et al. 2004) and many other species (Farooq et al. 2009). The leaf growth was more sensitive to water stress in wheat than in maize (Sacks et al. 1997), Vigna unguiculata (Manivannan et al. 2007), and sunflower (Manivannan et al. 2007, 2008).
14.4.1.3 Effects of Drought on Yield
The main purpose of harvesting any crop in the world is to get a good amount of harvestable yield. Any specie of crop requires a minimum amount of water to sustain and produce a good harvestable yield. Plantings of sunflower with abundant water supply, the yield increase was associated with both an increase in grain number and in individual grain weight (Soriano et al. 2002). Water scarcity primarily affects the dry matter yield and is the most important factor in determining the yield (Petropoulos et al. 2008). Soriano et al. (2002) concluded that in the sunflower harvest water-deficient condition was an important factor in the decrease of the yield and production and that presence of adequate amount of water is important throughout the season from establishment to harvesting period (Soriano et al. 2002). In another study by Edward and Wright (2008) on the wheat plant, they concluded that grain size and number decreased dramatically under drought-induced conditions.
In a similar study on maize, water-deficit stress vastly decreased the final yield, which was attributed to the extent of defoliation caused by water scarcity mainly during the early growth period (Monneveux et al. 2006). Drought conditions decrease seed production and yield in soybean mainly as a response to water-scarce conditions, which generate fewer seeds and pods in an area (Specht et al. 2001). Whereas in a comparitive study between water-stressed soybean seeds and controlled seeds, the yield of stressed seeds was found to be much lesser in comparison to water-controlled plants (Specht et al. 2001).
Tahir and Mehid (2001) conducted a study to know the effect of drought on sunflower plant and found that water-deficit condition greatly reduced the head diameter, dry weight, and harvestable yield per plant per unit area. The study observed a negative correlation between head diameter and fresh root and shoots weight, whereas a positive correlation was observed between dry roots and shoot weight and achene yield per plant per unit area under water-scarce conditions. Water-scarce conditions prevailing for more than 12 days at flowering and grain filling stage was found to be the most damaging one in decreasing the achene yield in sunflower plant (Reddy et al. 2004), in common bean, and green gram (Webber et al. 2006), maize (Monneveux et al. 2006), and Parsley (Petropoulos et al. 2008).
14.4.1.4 Effects of Drought Stress on Pigment Composition
Photosynthetic pigments are important component mainly present in leaves of the plant and phytosynthetic bacteria. Chlorophyll is the most important pigment among several and drying of soil has a great effect on chlorophyll “a” and “b” which in turn affect the health of the plant (Farooq et al. 2009). However, during times of drought, carotenoids play a major role and help plants to withstand adverse effects of drying soil. Some of the changes that occur in the pigments during water-scarce conditions are given below.
14.4.1.4.1 Chlorophylls
Researchers from around the world have proven in a number of experiments that chlorophyll and carotenoid contents are reduced due to the drying of the land (Farooq et al. 2009). The decrease in chlorophyll and carotenoids have been greatly observed in cotton plants (Massacci et al. 2008), sunflower (Kiani et al. 2008), Vaccinium myrtillus (Bilberry) (Tahkokorpi et al. 2007), etc. It has also been proven that the overall foliar photosynthesis rate of plants decreases with the decrease in leaf water potential (Lawlor and Cornic 2002).
Apart from this, there is not a common consensus among researchers whether drought limits photosynthesis through stomatal closing or pigment or metabolic disfigurement (Anjum et al. 2003; Lawson et al. 2003). Some mixed results have also been observed in cases such as that of Ashraf et al. (1994) where they stated that chlorophyll b content was observed to increase in two lines of ladyfinger plants, whereas chlorophyll a was observed to have remained unaffected resulting in the reduction of chlorophyll a:b ratio in water-scarce conditions.
14.4.1.4.2 Carotenoids
Carotenoids are class of photosynthetic pigments mainly composed of yellow, orange, and also red pigments, including carotene that is mainly responsible for giving color to the plants and its parts such as autumn leaves and ripe tomatoes. They also protect chlorophyll pigments from photodamage. These pigments are mainly synthesized by all photosynthetic and some nonphotosynthetic organisms as well (Wilson et al. 2008).
The carotenoid play a more important role during times of drought by playing the protective role of β-carotene through direct quenching of triplet chlorophyll, preventing the production of any singlet oxygen thereby protecting plant tissue from oxidative damage occurring due to water-deficit conditions (Farooq et al. 2009).
14.4.1.5 Effect of Drought on Stomata
Stomata are minute pores usually found in the epidermis of leaves and stems whose main functions are water and gaseous exchange. Stomata are mainly found in the lower side of the leaves where generation of new stomata takes place continuously throughout the growth period of the leaf (Zhao et al. 2015). Active stomatal functioning decreases the rate of transpiration and is among the most important processes that maintain the water balance in the plant tissues. Stomatal functioning/closure is also responsible for reducing cell growth and cell expansion, which affects the overall decrease in the rate of biomass production and yield (Rauf et al. 2015; Nemeskeri et al. 2015). It has been proven by many scientists that stomatal closure is among the primary response that a plant show in case of water scarcity to prevent or reduce water loss due to transpiration (Clauw et al. 2015; Nemeskeri et al. 2015). Closure of stomata is an active mechanism employed by plants to avoid dehydration (Osakabe et al. 2014; Clauw et al. 2015).
14.4.1.6 Drought-Associated Plant Diseases
Dry and hot conditions help the spreading of infections and diseases through pathogens and make plants susceptible to these diseases. Normally some diseases may be less harmful or lethal for the plant depending on the type. Some of the diseases that affect the plant during dry and water-scarce conditions are aflatoxin contamination, Aspergillus ear rot, etc. In general, certain diseases are prevalent while others are less severe or do not occur. With drought conditions, we anticipated yield losses due to soybean cyst nematode (SCN) and charcoal rot, as well as corn grain as a consequence of Aspergillus ear rot, charcoal rot, caused by Macrophomina phaseolina, etc. (Al-Kaisi et al. 2013).
14.4.2 Adverse Impacts of Salinity on Growth and Productivity of Crops
One of the most significant factors that limit the growth and production of crop is salinity. Salt stress generally affects the plants in two ways:
-
1.
Osmotic stress, a high concentration of salt makes extraction of water arduous for roots.
-
2.
Specific ion toxicity where the high levels of salt inside the plant pose harm to it (Munns and Tester 2008; Hussain et al. 2008).
Salinity has a number of damaging effects on physiological processes in plants like increase in the rate of respiration, ion toxicity, change in the metabolism of C and N, mineral distribution, membrane instability and permeability, reduced leaf area, dry mass, stomatal conductance as well as decreased efficiency of photosynthesis leads to the decrease in the productivity of plant (Sudhir and Murthy 2004; Kim et al. 2004; Hayat et al. 2012a, b).
14.4.2.1 Effect of Salt Stress on Plant Growth
Salinity stress has a significant impact on the growth and development of plants (Table 14.4). The effect of salt stress on processes such as germination, vegetative growth, and seedling growth and vigor has an adverse impact that ultimately leads to poor plant growth (Sairam and Tyagi 2004). Osmotic phase inhibits the growth of young and new leaves while the ionic phase accelerates the senescence of mature leaf and causes them to fall off. In soil when there is an excess concentration of salt, not only is the plant growth inhibited but this salt stress even causes death. The connection between the increase in the concentration of sodium chloride and the decrease in the plant length is evident in the result of various studies (Houimli et al. 2008; Rui et al. 2009; Memon et al. 2010). The influence of salinity on plants is such that the number of leaves decreases with an increase in the concentration of salt (Gama et al. 2007; Ha et al. 2008). It is also observed that the leaf area gets negatively affected as well with the increase in the concentration of NaCl (Rui et al. 2009).
14.4.2.2 Effect of Salt Stress on Photosynthesis Process
The inhibition of photosynthesis takes place when high concentrations of Na+ and Cl− are accumulated in the chlorophyll which is a major content for the photosynthesis to take place. This is directly correlated with the health of the plant (Zhang et al. 2008). It was observed that photosynthetic gas exchange parameters are reduced significantly due to salt stress conditions in Solanum melongena (Shaheen et al. 2012). Rate of photosynthesis, transpiration, and stomatal conductance is reduced significantly due to salt stress in Cucumis sativus as well as in Lycopersicon esculentum (Ahmad et al. 2012). It has been noted that the photosynthetic rate decreases with increase in salinity. This takes place due to certain factors (Iyengar and Reddy 1996):
-
1.
Osmotic stress caused by high salt concentration dehydrates the cell membrane, which causes reduction in permeability to CO2.
-
2.
Toxicity caused by Na+ and Cl−. Cl− inhibits N uptake by the roots, hence inhibiting the photosynthetic rate as well (Banuls et al. 1991).
-
3.
The closure of stomata causes reduction in CO2 supply restricting carboxylation reaction as well as the reduction in loss of water through transpiration affecting light-harvesting and energy conservation system (Brugnoli and Björkman 1992).
-
4.
The alteration of the cytoplasmic structure causes a change in enzymatic activity.
-
5.
Salinization enhances the leaf senescence rate and reduces sink activity.
14.4.3 Effect of Heavy Metals
14.4.3.1 Effect of Heavy Metals on Growth and Productivity of Crops
The heavy metals are available for plant uptake as soluble components in the soil solutions (Asati et al. 2016). At trace level some heavy metals like Zn, Ni, Cu, Co, etc. are beneficial for the plant growth. However, high concentrations of these metals in soil become toxic and reduce the growth and productivity of plants. Reduction in growth as a result of changes in physiological and biochemical processes such as decreased growth rate, chlorosis, necrosis, altered stomatal action, leaf rolling, reduced flow rate of cations, decreased water potential, alterations in membrane functions, inhibition of photosynthesis, respiration, altered metabolism, ultimately leads to food insecurity (Ashfaque et al. 2016; Hasanuzzaman et al. 2018). The direct effects of high metal concentration include inhibition of cytoplasmic enzymes and affect cell buildings due to oxidative stress, resulting in cellular damage (Dubey et al. 2018; Syed et al. 2018). Several researchers found that the accumulation of heavy metals in crop plants is mostly concerned with the probability of food contamination (Nazir et al. 2015). Though several heavy metals like Cd, As, Hg, etc. are not essential for plant growth, they are readily taken up and accumulated by plants. Arif et al. (2016) found the presence of heavy metals in both forms, i.e., essential and nonessential in the environment. Essential heavy metals such as magnesium (Mg), iron (Fe), manganese (Mn), and zinc (Zn) play a beneficial role in plant growth and development. A trace level of these beneficial elements improves the plant’s nutritional level and also several mechanisms for the normal growth, productivity, and better yield of plants.
Connor et al. (2018) reported that the pH of paddy soils in southern/eastern China has decreased due to excessive use of fertilizers and consistently increased available heavy metal concentrations and thus, greater contaminant enrichment in crop production. The bioavailability determines contaminant availability to a receptor and the level of risk in land management. Toxic heavy metals are widespread throughout the world and cause acute and chronic toxic effects on plants grown in such contaminated soils (Yadav 2010). For example, Cu is a vital metal for the growth and development of plants; however, it becomes potentially toxic at higher concentrations (Asati et al. 2016). Su et al. (2014) reported when copper content in soil is more than 50 mg kg−1, it affects citrus seedlings and if soil copper content is 200 mg kg−1, wheat suffers from wilting. On the other hand, Zn is the main functional elements to produce plant chlorophyll and also helps in metabolic, growth and development, generation of oxidative properties (Arif et al. 2016; Andresen et al. 2018). A study conducted by Anwaar et al. (2015) indicated that exogenous Si application improved the growth and development of cotton crops suffering from Zn toxicity stress by restricting Zn bioavailability and oxidative damage. The high levels of Hg in agricultural lands may result in Hg toxification in the entire food chain. The Hg always exists in the forms of HgS, Hg2+, Hg°, and methyl Hg. Therefore, the ionic form of Hg is always predominant in agricultural soil. Cr has been reported to cause oxidative stress that involves stimulation of lipid peroxidation in plants resulting in abrupt damage to cell membranes (Hayat et al. 2012a, b). The Pb uptake by plant roots reduced the crucial forms of precipitates with phosphates, sulfates, calcium, and chemicals in the rhizosphere (Fahr et al. 2013). Ni is also toxic to the plants, especially in ionic form like Ni+2 in soil and caused various physicochemical alterations like inhibition in growth and developments, photosynthesis rate, decreased chlorophyll content, and diverse toxicity symptoms such as leaf chlorosis, necrosis, and wilting in different plant species, including rice (Sachan and Lal 2017). Farid et al. (2015) examined the Cd, Pb, and Ni concentrations in soil 0.111 ppm, 0.87–8.97 ppm, and 0.017–1.72 ppm, respectively, at 0–15 cm while 0.88 ppm, 0.43–6.77 ppm, and 0.055–0.852 ppm, respectively, at 15–30 cm. Cd, Pb, and Ni concentrations in the plants ranged 0.00–2.25 ppm, 1.11–5.29 ppm, and 1.51–4.96, respectively, also transferred to the plant tissues ranged 0.00–2.25 ppm 1.11–5.29 ppm, and 1.51–4.96, respectively. Pb and Ni concentrations were found below permissible levels but Cd was found above the permissible levels in plants as well as in groundwater. Considerably, heavy metal toxicity varies from different plant species, concentration, definite metal, chemical forms, and soil solutions and other soil properties (Shah et al. 2010; Ackova 2018). Table 14.5 shows the impacts of heavy metals on different plant productivity and their growth.
14.5 Application of Biochar for Mitigation of Abiotic Stress
14.5.1 Mitigation of Drought Using Biochar
Restoring or improving soil water holding capacity is one of the only few options to improve soil productivity to achieve good crop production during the times of drought. Choosing a better management strategy among numerous others is very important as it can play a great role in restoring soil’s water and nutrient holding capacity (Baronti et al. 2014). A recent advancement has been made in the field of biochar that can effectively achieve the task of improving soil characteristics such as its water holding capacity. Using biochar has been considered one of the most practical and sustainable way to achieve and increase fertility of the soil, including water holding capacity. Several advantages of biochar such as long residence time in the soil large surface area in comparison to other amendments to improve soil quality make it much more preferable than other options throughout the world (Lehmann et al. 2011). Biochar are usually prepared in the absence of oxygen and under high pressure and temperature (above 250 °C), where the end product is a carbonaceous material with very high stability mainly attributed to recalcitrant aromatic structures of biochar molecules (Wardle et al. 2008; Biederman and Harpole 2013). The biochar has been proven to increase the moisture-holding capacity of the soil mainly attributed to its larger surface area and to recalcitrant aromatic structures of biochar molecules which also makes it suitable for improving agronomic performance under various climatic conditions including dry conditions (Biederman and Harpole 2013). It also possesses smaller pore size that allows them to hold capillary water for longer period of time making it suitable for application in drought-affected areas. Biochar are considered to have high potential to change soil characteristics including pH, nutrient retention, and overall fertility; however, the capacity of change varies greatly depending on the physicochemical properties of biochar itself including feedstock material, pyrolysis temperature and pressure, specific surface area, and method of application in the field (Baronti et al. 2014). As mentioned previously, improving soil water holding capacity and moisture retention capacity are mainly attributed to the porosity and large surface area of the biochar and its application in the drought prone areas, there is a lack of common consensus among researchers of a typical type of biochar to be used in these areas as moisture retention capacity of applied biochar mainly depends upon the manufacturing procedure.
The effect of biochar also improves soil quality and makes it suitable for plant growth during various periods including germination to adulthood thereby increasing overall plant growth as it improves the accessibility of roots to soil water and air. The application also improves response of soil to soil moisture, size of aggregate, texture, soil matrix, dynamics, cation retention, and permeability making it a much more preferable option.
14.5.1.1 Application of Biochar with Microorganisms to Mitigate Drought Stress
Egamberdieva et al. (2017) reported an increase in biomass, growth, nodulation, and nutrient uptake (phosphorus and nitrogen) in lupin (Lupinus angustifolius) under water-scarce conditions when biochar is applied with microorganisms in comparison to solely applied microorganism’s inoculation. The study also showed a considerable increase in relative water and chlorophyll content in comparison to the controlled ones. Similarly, Liu et al. (2017) observed that application of biochar with Rhizophagus irregularis decreased the water use efficiency, nutrient uptake, biomass, and overall growth as compared to control in potato. However, in contrary to the above findings, Pressler et al. (2017) concluded that application of wood-derived biochar with microorganisms had no significant effect on soil biota (including protozoa, bacteria, fungi, and nematodes) under low irrigation conditions.
Mickan et al. (2016) observed that inoculation of biochar with Arbuscular mycorrhizal fungi enhances drought tolerance of the plants by improving its physiological mechanisms such as nutrient uptake, and biochemical properties including osmotic adjustment, hormonal activities, and antioxidant systems. However, application of biochar with Arbuscular mycorrhizal fungi to the agricultural soil encouraged the growth of extra-radical hyphae and also increased mycorrhizal colonization of roots. Water potential was observed to be the same with and without biochar application.
Previous studies done by several researchers have proved that the application of biochar with microbial inoculation may prove itself to be far better helpful in mitigating and preventing drought stress in plants. However, there is no common consensus about the effectiveness and type of biochar to be used in such conditions, so there is a vast possibility of work in the near future and discover more advantages of biochar over other applications for drought mitigation.
14.5.2 Mitigation of Salinity Stress Using Biochar
Biochar has the potential to increase plant biomass and crop yield. Recent findings have shown that biochar treatment can increase crop yield by an average of 10% (Jeffery et al. 2011). Studies have also demonstrated that crop yield after several years of the application still show a significant increase after single treatment of biochar (Major et al. 2010). Salinity affects the total land area of about 7% of the world. It has been estimated that 30% of the irrigation land has been adversely affected by salinization (Chaves et al. 2009; Wicke et al. 2011). If no action is taken to prevent land degradation, up to 69% loss in revenue is estimated with time (Munns and Tester 2008). Charcoal and activated charcoal have been utilized in the industry for desalinization processes as it has high capacity to sorb a variety of salts (Bartell and Miller 1923; Zou et al. 2008). Decrease in biomass leads to lower carbon input that further deteriorates the soil. This is a major effect of salinity (Wong et al. 2009).
14.5.2.1 Effects of Biochar on Saline Soil Properties
Improvement of soil physicochemical and biological properties which is related to the removal of sodium such as leaching, absorption ratio as well as electrical conductivity has been observed with the application of biochar, which in turn assists in the reduction of salt stress (Sun et al. 2016; Drake et al. 2016). Soil enzymatic activity in saline soil varies with the rate of application of biochar, soil enzyme type as well as incubation time. The study shows that the soils’ physicochemical properties have improved significantly (Wang et al. 2014; Bhaduri et al. 2016). It has also been observed that the application of biochar (30 g mm−2) in salt-stressed soil though did not affect the pH of the soil, increased the electrical conductivity of the soil (Thomas et al. 2013). Application of furfural biochar in saline soil resulted in the decrease in pH on the other hand it increased the soil organic carbon as well as cation exchange capacity and available phosphorus in soil (Wu et al. 2014). It has been observed that composted biochar lead to the increase in cation exchange capacity and organic matter, while it decreased the exchangeable sodium and also the pH of the soil (Luo et al. 2017). The application of biochar has been observed to improve soil properties, such as increasing soil moisture, the Na binds with the biochar which results in the decrease in root sensitivity to osmotic stress.
14.5.2.2 Effect of Biochar on Plant Growth under Salinity Stress
As discussed in Table 14.6, many researchers have proven that amendment of biochar in fairly saline soils improves growth of plants, biomass as well as the rate of photosynthesis (Akhtar et al. 2015). In the tidal land soils containing high concentrations of soluble salt as well as exchangeable sodium, the application of rice hull biochar increased the growth of maize as well as its biomass (Kim et al. 2016). Application of biochar under saline irrigation (3.6 dS m−1) increased the growth of tomato as well as increased the total biomass (Usman et al. 2016). A study conducted to estimate the effect of biochar derived from Fagus grandifolia on the growth and biomass production of two herbaceous species namely, Prunella vulgaris and Abutilon theophrasti, under saline condition. It was found that the biomass of both plants increased under salt stress. The effect on carbon profit through photosynthesis, water use efficiency, and chlorophyll fluorescence in both species did not have any significant impact under salt stress (Thomas et al. 2013). Amendment of biochar along with suitable microbial inoculation has been reported to improve growth and biomass production of the plant under saline conditions as compared to the control (Nadeem et al. 2013; Fazal and Bano 2016). The results obtained by Akhtar et al. (2015) depict that application of biochar increases leaf area, shoot, and root biomass, which increases with bacterial inoculation. The studies emphasize that the effect of biochar can be enhanced with the application of symbiotic microorganisms.
14.5.3 Mitigation of Heavy Metal Stress by Using Biochar
Several organic fertilizers like compost, vermicompost, manure, organic fertilizers, farmyard manure, etc. have been suggested to restore and improve soil quality and crop productivity (Mas-Carrió et al. 2018). Soil amelioration with biochar is the pre-eminent to promote sustainable agriculture because it has ability to nourish the soil and provide vital nutrients to the plants resulting in improved plant productivity. As described in Table 14.7, the application of biochar has capacity to reduce metal bioavailability and immobilized other contaminants also (Yuan et al. 2019). Similarly, several researchers explained that different types of biochar have potential and efficiency to adsorb the toxic agrochemicals and reduce the bioavailability of heavy metals and their uptake by plants (Abbas et al. 2017). Nie et al. (2018) assessed the bioavailability of Cd, Cu, and Pb and health of soil microbial population in the contaminated soil, by using sugarcane dry pulpy residues-derived biochar and they found that application of biochar enhanced growth of Brassica chinesis L. in terms of root and shoot development, control the bioavailability of metal and also enhanced the microbial population. Similarly, Zheng et al. (2015) investigated the effect of biochar (beanstalk and rice straw) on the bioavailability of Cd in contaminated soil and their accumulation into rice crop and they observed that biochar decreased the phytotoxicity of Cd (35–81 %). Hayyat et al. (2016) found that application of biochar in metal contaminated soil showed a significant decrease in metal toxicity and enhancement in the soil nutrient/fertility, plant growth, crop yield, carbon content, etc. Zheng et al. (2017) examined the effects of biochar (rice straw) on Cd-containing soil and their accumulation in lettuce plant. They observed that the level of Cd reduced by 57 % while increasing biochar rates (0, 6, 12, 18 t ha−1) increased shoot length and yield of lettuce plant. Khan et al. (2013) conducted an experiment to check the effects of biochar (derived from sewage sludge) on the rice plants and they found that the biochar increased the shoot biomass, grain yield, and the bioaccumulation of P and Na, decreased the bioaccumulation of N (excluding in grain) and K and decreased the bioavailability of metals (As, Cr, Co, Cu, Ni, and Pb); however, Cd and Zn concentrations were not affected in the plant parts by the application of biochar. Meng et al. (2018) found that the application of biochar (rice straw and swine manure) decrease the heavy metal bioavailability from contaminated soil in the order Pb > Cu > Zn > Cd. Kim et al. (2015) observed that the rice hull biochar significantly reduced the phytoavailability of metal present in contaminated soil and metal uptake by lettuce plant decreased with the increasing biochar application. Xu et al. (2018) found the adding of macadamia nutshell biochar (5% w/w) to the soil decreased Cd and Pb toxicity and increased total microbial phospholipid fatty acids (PLFAs), microbial respiration rate, biomass carbon and microbial availability.
14.6 Conclusions
Along with other conventional and biotechnological interventions, the application of organic manures may also be used to overcome the adverse impacts of abiotic stresses. The application of biochar as an organic amendment has proved to mitigate from drought, salinity and heavy metals. Application of biochar is an environment friendly and economically viable. Its application improves soil physicochemical and biological properties that help the plants to fight with adverse conditions of the environment. It has been found that biochar enhances water holding capacity, increase the nodulation, nutrient uptake, microbial diversity in the soil and thereby increases the growth and productivity of the plants. In the same way, biochar has potential, and efficiency to adsorbs the toxic agrochemicals and reduces their bioavailability, especially heavy metals and ultimately reduces their uptake by plants.
References
Abbas T, Rizwan M, Ali S, Zia-ur-Rehman M, Qayyum MF, Abbas F, Hannan F, Rinklebe J, Ok YS (2017) Effect of biochar on cadmium bioavailability and uptake in wheat (Triticum aestivum L.) grown in a soil with aged contamination. Ecotoxicol Environ Saf 140:37–47
Abbas AH, Sulaiman WR, Jaafar MZ, Aja AA (2018) Micelle formation of aerosol-OT surfactants in sea water salinity. Arab J Sci Eng 43:2515–2519
Ackova DG (2018) Heavy metals and their general toxicity on plants. Plant Sci 5(1):14–18
Ahmad M, Lee SS, Yang JE, Ro HM, Lee YH, Ok YS (2012) Effects of soil dilution and amendments (mussel shell, cow bone, and biochar) on Pb availability and phytotoxicity in military shooting range soil. Ecotoxicol Environ Saf 79:225–231
Akhtar SS, Andersen MN, Naveed M, Zahir ZA, Liu F (2015) Interactive effect of biochar and plant growth-promoting bacterial endophytes on ameliorating salinity stress in maize. Funct Plant Biol 42(8):770–781
Alaboudi KA, Ahmed B, Brodie G (2019) Effect of biochar on Pb, Cd and Cr availability and maize growth in artificial contaminated soil. Ann Agric Sci 4(1):95–102
Alam SM (1999) Nutrient uptake by plants under stress conditions. Handbook of plant and crop stress 19(2):285–313
AL-Amoudi OA, Rashed AA (2012) Effect of nutrient cations to improving salinity-tolerance responses in Sorghum bicolor L. Life Sci 50:77–81
Alharby HF, Al-Zahrani HS, Hakeem KR, Iqbal M (2019) Identification of physiological and biochemical markers for salt (NaCl) stress in the seedlings of mung bean [Vigna radiata (L.) Wilczek] genotypes. Saudi J Biol Sci 26(5):1053–1060
Al-Kaisi MM, Elmore RW, Guzman JG, Hanna HM, Hart CE, Helmers MJ, Hodgson EW, Lenssen AW, Mallarino AP, Robertson AE, Sawyer JE (2013) Drought impact on crop production and the soil environment: 2012 experiences from Iowa. J Soil Water Conserv 68(1):19A–24A
Amini S, Ghadiri H, Chen C, Marschner P (2015) Salt-affected soils, reclamation, carbon dynamics, and biochar: a review. J Soils Sediments 16(3):939–953
Andresen E, Peiter E, Kupper H (2018) Trace metal metabolism in plants. J Exp Bot 69:909–954
Anjum F, Yaseen M, Rasul E, Wahid A, Anjum S (2003) Water stress in barley (Hordeum vulgare L.) II. Effect on chemical composition and chlorophyll contents. Pak J Agric Sci 40:45–49
Anwaar SA, Ali S, Ali S, Ishaque W, Farid M, Farooq MA, Najeeb U, Abbas F, Sharif M (2015) Silicon (Si) alleviates cotton (Gossypium hirsutum L.) from zinc (Zn) toxicity stress by limiting Zn uptake and oxidative damage. Environ Sci Pollut Res Int 5:3441–3450
Arafat Y, Wei X, Jiang Y, Chen T, Saqib H, Lin S, Lin W (2017) Spatial distribution patterns of root-associated bacterial communities mediated by root exudates in different aged ratooning tea monoculture systems. Int J Mol Sci 18(8):1727
Arif N, Yadav V, Singh S, Singh S, Ahmad P, Mishra RK, Sharma S, Tripathi DK, Dubey NK, Chauhan DK (2016) Influence of high and low levels of plant-beneficial heavy metal ions on plant growth and development. Chemosphere 139:604–608
Arora N, Patel A, Sharma M, Mehtani J, Pruthi PA, Pruthi V, Poluri KM (2017) Insights into the enhanced lipid production characteristics of a fresh water microalga under high salinity conditions. Ind Eng Chem Res 56(25):7413–7421
Asati A, Pichhode M, Nikhil K (2016) Effect of heavy metals on plants: an overview. Int J Innov Eng Res Manag 5(3):56–66
Ashfaque F, Inam A, Sahay S, Iqbal S (2016) Influence of heavy metal toxicity on plant growth, metabolism and its alleviation by phytoremediation—a promising technology. J Agric Ecol Res Int 6(2):1–19
Ashraf MY, Azmi AR, Khan AH, Ala SA (1994) Effect of water stress on total phenols, peroxidase activity and chlorophyll content in wheat (Triticum aestivum L.) genotypes under soil water deficits. Acta Physiol Plant 16:185–191
Atafar Z, Mesdaghinia A, Nouri J, Homaee M, Yunesian M, Ahmadimoghaddam M, Mahvi AH (2010) Effect of fertilizer application on soil heavy metal concentration. Environ Monit Assess 160:83–89
Bagheri A (2009) Effects of drought stress on chlorophyll, proline and rates of photosynthesis and respiration and activity of superoxide dismutase and peroxidase in millet (Panicum milenaceum L.). National conference on water scarcity and drought management in agriculture. Islamic Azad University Arsanjan. p. 16
Banuls J, Legaz F, Primo-Millo E (1991) Salinity-calcium interactions on growth and ionic concentration of citrus plants. Plant Soil 133(1):39–46
Bardgett RD, Wardle DA (2010) Aboveground-belowground linkages: biotic interactions, ecosystem processes, and global change. Oxford University Press, Oxford
Baronti S, Vaccari FP, Miglietta F, Calzolari C, Lugato E, Orlandini S, Pini R, Zulian C, Genesio L (2014) Impact of biochar application on plant water relations in Vitis vinifera (L.). Eur J Agron 53:38–44
Bartell FE, Miller EJ (1923) Adsorption by activated sugar charcoal. II1, 2. J Am Chem Soc 45(5):1106–1115
Bauddh K, Singh RP (2009) Genotypic differences in cadmium (Cd) toxicity in Indian mustard (Brassica juncea L.). Pollut Res 28(4):699–704
Bauddh K, Singh RP (2011) Differential toxicity of cadmium to mustard (Brassica juncia L.) genotypes under higher metal levels. J Environ Biol 32(3):355–362
Bauddh K, Singh RP (2012) Growth, tolerance efficiency and phytoremediation potential of Ricinus communis (L.) and Brassica juncea (L.) in salinity and drought affected cadmium contaminated soil. Ecotoxicol Environ Saf 85:13–22
Bauddh K, Kumar A, Srivastava S, Singh RP, Tripathi RD (2015) A study on the effect of cadmium on the antioxidative defense system and alteration in different functional groups in castor bean and Indian mustard. Arch Agron Soil Sci 62(6):877–891
Bauddh K, Singh K, Singh RP (2016) Ricinus communis L. a value added crop for remediation of cadmium contaminated soil. Bull Environ Contam Toxicol 96(2):265–269
Baum C, Leinweber P, Schlichting A (2003) Effects of chemical conditions in re-wetted peats on temporal variation in microbial biomass and acid phosphatase activity within the growing season. Appl Soil Ecol 22(2):167–174
Bhaduri D, Saha A, Desai D, Meena HN (2016) Restoration of carbon and microbial activity in salt-induced soil by application of peanut shell biochar during short-term incubation study. Chemosphere 148:86–98
Bhaskar P, Bauddh K, Singh RP (2009) Differential response of two high yielding cultivars of Indian mustard (Brassica juncea L.) to NaCl salinity during seed germination and early seedling growth. J Ecophysiol Occup Health 9:137–144
Bhatt RM, Rao NS (2005) Influence of pod load on response of okra to water stress. Indian J Plant Physiol 10(1):54
Biederman LA, Harpole WS (2013) Biochar and its effects on plant productivity and nutrient cycling: a meta-analysis. GCB Bioenergy 5(2):202–214
Bohn HL, McNeal BL, O’Connor GA (2001) Soil chemistry, 3rd edn. Wiley, New York
Břendová K, Tlustoš P, Száková J (2015) Biochar immobilizes cadmium and zinc and improves phytoextraction potential of willow plants on extremely contaminated soil. Plant Soil Environ 61(7):303–308
Brugnoli E, Björkman O (1992) Growth of cotton under continuous salinity stress: influence on allocation pattern, stomatal and non-stomatal components of photosynthesis and dissipation of excess light energy. Planta 187(3):335–347
Chander K, Goyal S, Mundra MC, Kapoor KK (1997) Organic matter, microbial biomass and enzyme activity of soils under different crop rotations in the tropics. Biol Fertil Soils 24(3):306–310
Chaves MM, Flexas J, Pinheiro C (2009) Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Ann Bot 103:551–560
Chibuike GU, Obiora SC (2014) Heavy metal polluted soils: effect on plants and bioremediation methods. Appl Environ Soil Sci 752708:1–12
Chodak M, Gołębiewski M, Morawska-Płoskonka J, Kuduk K, Niklińska M (2015) Soil chemical properties affect the reaction of forest soil bacteria to drought and rewetting stress. Ann Microbiol 65:1627–1637
Chu D (2018) Effects of heavy metals on soil microbial community. Earth Environ Sci 113:1–5
Clauw P, Coppens F, De Beuf K, Dhondt S, Van Daele T, Maleux K, Storme V, Clement L, Gonzalez N, Inzé D (2015) Leaf responses to mild drought stress in natural variants of Arabidopsis. Plant Physiol 167(3):800–816
Connor DO, Peng T, Zhang J, Tsang DCW, Alessi DS, Shen Z, Bolan NS, Hou D (2018) Biochar application for the remediation of heavy metal polluted land: a review of in situ field trials. Sci Total Environ 619–620:815–826
Dikinya O, Hinz C, Aylmore G (2006) Dispersion and re-deposition of fine particles and their effects on saturated hydraulic conductivity. Soil Res 44(1):47–56
Drake JA, Cavagnaro TR, Cunningham SC, Jackson WR, Patti AF (2016) Does biochar improve establishment of tree seedlings in saline sodic soils? Land Degrad Dev 27(1):52–59
Dubey S, Shri M, Gupta A, Rani V, Chakrabarty D (2018) Toxicity and detoxifcation of heavy metals during plant growth and metabolism. Environ Chem Lett 16(4):1169–1192
Edward D, Wright D (2008) The effects of winter water-logging and summer drought on the growth and yield of winter wheat (Triticum aestivum L.). Eur J Agron 28:234–244
Egamberdieva D, Reckling M, Wirth S (2017) Biochar-based Bradyrhizobium inoculum improves growth of lupin (Lupinus angustifolius L.) under drought stress. Eur J Soil Biol 78:38–42
Eze OC, Tukura BW, Atolaiye BO, Opaluwa OD (2018) Assessment of some physicochemical parameters of soil and heavy metals in vegetables cultivated on irrigated sites along the bank of Mpape River in FCT, Abuja, Nigeria. J Environ Sci Toxicol Food Technol 12(5):28–38
Fahr M, Laplaze L, Bendaou N, Hocher V, Mzibri ME, Bogusz D, Smouni A (2013) Effect of lead on root growth. Front Plant Sci 4:175. https://doi.org/10.3389/fpls.2013.00175
Farhangi-Abriz S, Torabian S (2018a) Biochar improved nodulation and nitrogen metabolism of soybean under salt stress. Symbiosis 74(3):215–223
Farhangi-Abriz S, Torabian S (2018b) Effect of biochar on growth and ion contents of bean plant under saline condition. Environ Sci Pollut R 25(12):11556–11564
Farid G, Sarwar N, Saifullah AA, Ghafoor A, Rehman M (2015) Heavy metals (Cd, Ni and Pb) contamination of soils, plants and waters in Madina town of Faisalabad metropolitan and preparation of GIS based maps. Adv Crop Sci Techn 4(1):1–7
Farooq M, Basra SM, Wahid A, Cheema ZA, Cheema MA, Khaliq A (2008) Physiological role of exogenously applied glycinebetaine to improve drought tolerance in fine grain aromatic rice (Oryza sativa L.). J Agron Crop Sci 194(5):325–333
Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SM (2009) Plant drought stress: effects, mechanisms and management. In: Lichtfouse E, Navarrete M, Debaeke P, Véronique S, Alberola C (eds) Sustainable agriculture. Springer, Dordrecht, pp 153–188
Fazal A, Bano A (2016) Role of plant growth-promoting rhizobacteria (PGPR), biochar, and chemical fertilizer under salinity stress. Commun Soil Sci Plant Anal 47(17):1985–1993
Gama PB, Inanaga S, Tanaka K, Nakazawa R (2007) Physiological response of common bean (Phaseolus vulgaris L.) seedlings to salinity stress. Afr J Biotechnol 6(2):421–426
Ganesh SK, Sundaramoorthy P (2018) Copper and zinc induced changes in soybean (Glycine max (L.) Merr.). Innov Agri 1(1):9–12
Garcia C, Hernandez T, Costa F (1994) Microbial activity in soils under Mediterranean environmental conditions. Soil Biol Biochem 26(9):1185–1191
Garg BK, Burman U, Kathju S (2004) The influence of phosphorus nutrition on the physiological response of moth bean genotypes to drought. J Plant Nutr Soil Sci 167(4):503–508
Ghodsi M, Nazeri M, Zarea-Fizabady A (1998) The reaction of new cultivars and elite lines of spring wheat into drought stress. In Collection of abstract articles of 5th Iranian agronomy and plant breeding conference, Karaj, Iran. p. 252
Griffiths RI, Whiteley AS, O’Donnell AG, Bailey MJ (2003) Physiological and community responses of established grassland bacterial populations to water stress. Appl Environ Microbiol 69:6961–6968
Ha E, Ikhajiagba B, Bamidele JF, Ogic-Odia E (2008) Salinity effects on young healthy seedling of Kyllingia peruviana collected from escravos, Delta state. Glob J Environ Res 2(2):74–88
Haggag WM, Abouziena HF, Abd-El-Kreem F, El Habbasha S (2015) Agriculture biotechnology for management of multiple biotic and abiotic environmental stress in crops. J Chem Pharm Res 7(10):882–889
Hagmanna DF, Goodeya NM, Mathieua C, Evans J, Aronsonc MFJ, Gallagher F, Krumins JA (2015) Effect of metal contamination on microbial enzymatic activity in soil. Soil Biol Biochem 91:291–297
Hasanuzzaman M, Nahar K, Rahman A, Mahmud JA, Alharby HF, Fujita M (2018) Exogenous glutathione attenuates lead-induced oxidative stress in wheat by improving antioxidant defense and physiological mechanisms. J Plant Interact 13(1):203–212
Hayat S, Khalique G, Irfan M, Wani AS, Tripathi BN, Ahmad A (2012a) Physiological changes induced by chromium stress in plants: an overview. Protoplasma 249(3):599–561
Hayat S, Yadav S, Wani AS, Irfan M, Alyemini MN, Ahmad A (2012b) Impact of sodium nitroprusside on nitrate reductase, proline content, and antioxidant system in tomato under salinity stress. Hortic Environ Biotechnol 53(5):362–367
Hayyat A, Javed M, Rasheed I, Ali S, Shahid MJ, Rizwan M (2016) Role of biochar in remediating heavy metals in soil. Phytoremediation 3:420–437
Heuer B, Nadler A (1995) Growth and development of potatoes under salinity and water deficit. Aust J Agric Res 46(7):1477–1486
Hniličková H, Hnilička F, Orsák M, Hejnák V (2019) Effect of salt stress on growth, electrolyte leakage, Na+ and K+ content in selected plant species. Plant Soil Environ 65(2):90–96
Hoogmoed M, Sadras VO (2016) The importance of water-soluble carbohydrates in the theoretical framework for nitrogen dilution in shoot biomass of wheat. Field Crop Res 193:196–200
Houimli SI, Denden M, El Hadj SB (2008) Induction of salt tolerance in pepper (Capsicum annuum) by 24-epibrassinolide. Eur Asian J Bio Sci 2:83–90
Hu XF, Jiang Y, Shu Y, Hu X, Liu L, Luo F (2014) Effects of mining wastewater discharges on heavy metal pollution and soil enzyme activity of the paddy fields. J Geochem Explor 147:139–150
Hueso S, García C, Hernández T (2012) Severe drought conditions modify the microbial community structure, size and activity in amended and unamended soils. Soil Biol Biochem 50:167–173
Hussain TM, Hazara M, Sultan Z, Saleh BK, Gopal GR (2008) Recent advances in salt stress biology a review. Biotechnol Mol Biol Res 3(1):8–13
ICARDA (2002) Marginal quality water: a solution to fresh water scarcity. International Center for Agricultural Research in the Dry Areas, Aleppo, Syria
Isayenkov SV, Maathuis FJM (2019) Plant salinity stress: many unanswered questions remain. Front Plant Sci 10:80. https://doi.org/10.3389/fpls.2019.00080
Iyengar ERR, Reddy MP (1996) Photosynthesis in highly salt tolerant plants. In: Pessaraki M (ed) Handbook of photosynthesis. Marcel Dekker, New York, pp 897–909
Jaleel CA, Gopi R, Manivannan P, Gomathinayagam M, Sridharan R, Panneerselvam R (2008a) Antioxidant potential and indole alkaloid profile variations with water deficits along different parts of two varieties of Catharanthus roseus. Colloids Surf B Biointerfaces 62(2):312–318
Jaleel CA, Gopi R, Panneerselvam R (2008b) Growth and photosynthetic pigments responses of two varieties of Catharanthus roseus to triadimefon treatment. C R Biol 331(4):272–277
Jaleel CA, Gopi R, Sankar B, Gomathinayagam M, Panneerselvam R (2008c) Differential responses in water use efficiency in two varieties of Catharanthus roseus under drought stress. C R Biol 331(1):42–47
Jaleel CA, Manivannan P, Lakshmanan GM, Gomathinayagam M, Panneerselvam R (2008d) Alterations in morphological parameters and photosynthetic pigment responses of Catharanthus roseus under soil water deficits. Colloid Surface B Biointerfaces 61(2):298–303
Jaleel CA, Sankar B, Murali PV, Gomathinayagam M, Lakshmanan GM, Panneerselvam R (2008e) Water deficit stress effects on reactive oxygen metabolism in Catharanthus roseus; impacts on ajmalicine accumulation. Colloid Surface B Biointerfaces 62(1):105–111
Jan AT, Azam M, Siddiqui K, Ali A, Choi I, Mohd A, Haq R (2015) Heavy metals and human health, Mechanistic insight into toxicity and counter defense system of antioxidants. Int J Mol Sci 16(12):29592–29630
Jatav HS, Singh SK, Singh Y, Kumar O (2016) Biochar and sewage sludge application increases yield and micronutrient uptake in rice (Oryza sativa L.). Commun Soil Sci Plan 50(14):1732–1742
Jeffery S, Verheijen FGA, van der Velde M, Bastos AC (2011) A quantitative review of the effects of biochar applications to soils on crop productivity using meta-analysis. Agric Ecosyst Environ 144:175–187
Jin Z, Li Z, Li Q, Hu Q, Yang R, Tang H, Li M, Huang B, Zhang J, Li G (2015) Canonical correspondence analysis of soil heavy metal pollution, microflora and enzyme activities in the Pb–Zn mine tailing dam collapse area of Sidi village, SW China. Environ Earth Sci 73(1):267–274
John R, Ahmad P, Gadgila K, Sharma K (2009) Heavy metal toxicity: effect on plant growth, biochemical parameters and metal accumulation by Brassica juncea L. Int J Plant Prod 3(3):65–67
Kanwal S, Ilyas N, Shabir S, Saeed M, Gul R, Zahoor M, Batool N, Mazhar R (2018) Application of biochar in mitigation of negative effects of salinity stress in wheat (Triticum aestivum L.). J Plant Nutr 41(4):526–538
Karami N, Clemente R, Moreno-Jiménez E, Lepp NW, Beesley L (2011) Efficiency of green waste compost and biochar soil amendments for reducing lead and copper mobility and uptake to ryegrass. J Hazard Mater 191(1-3):41–48
Khan M (2000) Effect of metals contamination on soil microbial diversity, enzymatic activity, organic matter decomposition and nitrogen mineralization (a review). Pak J Biol Sci 3(11):1950–1956
Khan S, Hesham AEL, Qiao M, Rehman S, He JZ (2010) Effects of Cd and Pb on soil microbial community structure and activities. Environ Sci Pollut Res 17(2):288–296
Khan S, Chao C, Muhammad W, Arp HPH, Zhu YG (2013) Sewage sludge biochar influence upon rice (Oryza sativa L) yield, metal bioaccumulation and greenhouse gas emissions from acidic paddy soil. Environ Sci Technol 47(15):8624–8632
Kiani SP, Maury P, Sarrafi A, Grieu P (2008) QTL analysis of chlorophyll fluorescence parameters in sunflower (Helianthus annuus L.) under well-watered and water-stressed conditions. Plant Sci 175:565–573
Kim DI, Matsuyama Y, Nagasoe S, Yamaguchi M, Yoon YH, Oshima Y, Imada N, Honjo T (2004) Effects of temperature, salinity and irradiance on the growth of the harmful red tide dinoflagellate Cochlodinium polykrikoides Margalef (Dinophyceae). J Plankton Res 26(1):61–66
Kim HS, Kim KR, Kim HJ, Yoon JH, Yang JE, Ok YS, Owens G, Kim KH (2015) Effect of biochar on heavy metal immobilization and uptake by lettuce (Lactuca sativa L.) in agricultural soil. Environ Earth Sci 74:1249–1259
Kim BH, Ramanan R, Kang Z, Cho DH, Oh HM, Kim HS (2016) Chlorella sorokiniana HS1, a novel freshwater green algal strain, grows and hyper accumulates lipid droplets in seawater salinity. Biomass Bioenergy 85:300–305
Kouchou A, Rais N, Elsass F, Duplay J, Fahli N, Ghachtouli NEL (2017) Effects of long-term heavy metals contamination on soil microbial characteristics in calcareous agricultural lands (Saiss plain, North Morocco). J Mater Environ Sci 8(2):691–695
Kuchenbuch R, Claassen N, Jungk A (1986) Potassium availability in relation to soil moisture. Plant Soil 95(2):233–243
Kumar N, Bauddh K, Singh R, Anand K, Barman SC, Singh DP (2009) Phytotoxicity of trace metals (Cu and Cd) to gram (Cicer aerientinum) and mung (Phasiolus mungo). J Ecophysiol Occup Health 9(2):59–65
Kusaka M, Ohta M, Fujimura T (2005) Contribution of inorganic components to osmotic adjustment and leaf folding for drought tolerance in pearl millet. Physiol Plant 125(4):474–489
Lakhdar A, Rabhi M, Ghnaya T, Montemurro F, Jedidi N, Abdelly C (2009) Effectiveness of compost use in salt-affected soil. J Hazard Mater 171(1-3):29–37
Lashari MS, Liu Y, Li L, Pan W, Fu J, Pan G, Zheng J, Zheng J, Zhang X, Yu X (2013) Effects of amendment of biochar-manure compost in conjunction with pyroligneous solution on soil quality and wheat yield of a salt-stressed cropland from Central China Great Plain. Field Crop Res 144:113–118
Lawlor DW, Cornic G (2002) Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ 25:275–294
Lawson T, Oxborough K, Morison JI, Baker NR (2003) The responses of guard and mesophyll cell photosynthesis to CO2, O2, light and water stress in a range of species are similar. J Exp Bot 54:1743–1752
Lehmann J, Rillig MC, Thies J, Masiello CA, Hockaday WC, Crowley D (2011) Biochar effects on soil biota–a review. Soil Biol Biochem 43(9):1812–1836
Leila R (2007) Response of Tunisian autochthonous peral millet to drought stress induced by polyethylene glycol 6000. Afr J Biotechnol 6:1102–1105
Liu C, Liu F, Ravnskov S, Rubæk GH, Sun Z, Andersen MN (2017) Impact of wood biochar and its interactions with mycorrhizal fungi, phosphorus fertilization and irrigation strategies on potato growth. J Agron Crop Sci 203:131–145
Lu HP, Li ZA, Gascó G, Méndez A, Shen Y, Paz-Ferreiro J (2018) Use of magnetic biochars for the immobilization of heavy metals in a multi-contaminated soil. Sci Total Environ 622-623:892–899
Luo JY, Zhang S, Peng J, Zhu XZ, Lv LM, Wang CY, Li CH, Zhou ZG, Cui JJ (2017) Effects of soil salinity on the expression of Bt toxin (Cry1Ac) and the control efficiency of Helicoverpa armigera in field-grown transgenic Bt cotton. PLoS One 12(1):0170379
Major J, Rondon M, Molina D, Riha SJ, Lehmann J (2010) Maize yield and nutrition after 4 years of doing biochar application to a Colombian savanna oxisol. Plant Soil 333:117–128
Manivannan P, Jaleel CA, Kishorekumar A, Sankar B, Somasundaram R, Sridharan R, Panneerselvam R (2007) Changes in antioxidant metabolism of Vigna unguiculata (L.) Walp. by propiconazole under water deficit stress. Colloids Surf B Biointerfaces 57(1):69–74
Manivannan P, Jaleel CA, Somasundaram R, Panneerselvam R (2008) Osmoregulation and antioxidant metabolism in drought-stressed Helianthus annuus under triadimefon drenching. C R Biol 331(6):418–425
Marschner H (1995) Mineral nutrition of higher plants. Academic, London, p 889
Mas-Carrió E, Dini-Andreote F, Brossi MJDL, Salles JF, Olff H (2018) Organic amendment under increasing agricultural intensification: effects on soil bacterial communities and plant productivity. Front Microbiol 9:2612
Massacci A, Nabiev SM, Pietrosanti L, Nematov SK, Chernikova TN, Thor K, Leipner J (2008) Response of the photosynthetic apparatus of cotton (Gossypium hirsutum) to the onset of drought stress under field conditions studied by gas-exchange analysis and chlorophyll fluorescence imaging. Plant Physiol Biochem 46(2):189–195
Memon SA, Hou X, Wang LJ (2010) Morphlogical analysis of salt stress response of Pak Choi. Electron J Environ Agric Food Chem 9(1):203–207
Meng L, Li W, Zhang S, Wu C, Jiang W, Sha C (2016) Effect of different extra carbon sources on nitrogen loss control and the change of bacterial populations in sewage sludge composting. Ecol Eng 94:238–243
Meng J, Tao M, Wang L, Liu X, Xu J (2018) Changes in heavy metal bioavailability and speciation from a Pb-Zn mining soil amended with biochars from co-pyrolysis of rice straw and swine manure. Sci Total Environ 633:300–307
Metternicht GI, Zinck JA (2003) Remote sensing of soil salinity: potentials and constraints. Remote Sens Environ 85(1):1–20
Mickan BS, Abbott LK, Stefanova K, Solaiman ZM (2016) Interactions between biochar and mycorrhizal fungi in a water-stressed agricultural soil. Mycorrhiza 26:565–574
Minhas PS, Rane J, Pasala RK (2017) Abiotic stresses in agriculture: an overview. In: Minhas P, Rane J, Pasala R (eds) Abiotic stress management for resilient agriculture. Springer, Singapore
Mishra S, Bharagava RN (2016) Toxic and geno-toxic effects of hexavalent chromium in environment and its bioremediation strategies. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev 34(1):1–32
Mishra S, Bharagava RN, More N, Yadav A, Zainith S, Mani S, Chowdhary P (2018) Heavy metal contamination: an alarming threat to environment and human health. In: Sobti R, Arora N, Kothari R (eds) Environmental biotechnology: for sustainable future. Springer, Singapore, pp 103–125
Monneveux P, Sanchez C, Beck D, Edmeades GO (2006) Drought tolerance improvement in tropical maize source populations: evidence of progress. Crop Sci 46:180–191
Muneer M, Oades JM (1989) The role of Ca-organic interactions in soil aggregate stability. II. Field studies with 14C-labeled straw, CaCO3 and CaSO42.H2O. Soil Res 27(2):401–409
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59:651–681
Mutale I, Franco IB, Jewette M (2019) Corporate sustainability performance: an approach to effective sustainable community development or not? A case study of the Luanshya copper mine in Zambia. Sustainability 11(20):5775
Nadeem SM, Zahir ZA, Naveed M, Nawaz S (2013) Mitigation of salinity-induced negative impact on the growth and yield of wheat by plant growth-promoting rhizobacteria in naturally saline conditions. Ann Microbiol 63(1):225–232
Nakayama NO, Saneoka HI, Moghaieb RE, Premachandra GS, Fujita KO (2007) Response of growth, photosynthetic gas exchange, translocation of 13C-labelled photosynthate and N accumulation in two soybean (Glycine max L. Merrill) cultivars to drought stress. Int J Agric Biol 9(5):669–674
Nazir R, Khan M, Masab M, Rehman HU, Rauf NU, Shahab S, Ameer N, Sajed M, Ullah M, Rafeeq M, Shaheen Z (2015) Accumulation of heavy metals (Ni, Cu, Cd, Cr, Pb, Zn, Fe) in the soil, water and plants and analysis of physico-chemical parameters of soil and water collected from Tanda Dam Kohat. J Pharm Sci Res 7(3):89–97
Nelson PN, Oades JM (1998) Organic matter, sodicity and soil structure. In: Sumner ME, Naidu RI (eds) Sodic soils, distribution, processes, management and environmental consequences. Oxford University Press, New York, pp 51–75
Nemeskeri E, Molnar K, Vigh R, Nagy J, Dobos A (2015) Relationships between stomatal behaviour, spectral traits and water use and productivity of green peas (Pisum sativum L.) in dry seasons. Acta Physiol Plant 37:1–16
Nguyen D, Rieu I, Mariani C, van Dam NM (2016) How plants handle multiple stresses: hormonal interactions underlying responses to abiotic stress and insect herbivory. Plant Mol Biol 91(6):727–740
Nie C, Yang X, Niazi NK, Xu X, Wen Y, Rinklebei J, Ok YS, Xu S, Wang H (2018) Impact of sugarcane bagasse-derived biochar on heavy metal availability and microbial activity: a field study. Chemosphere 200:274–282
Obasi NA, Akubugwo EI, Ugbogu OC, Chinyere GC (2012) Heavy metals bioavailability and phyto-accumulation potentials of selected plants on burrow-pit dumpsites in Aba and Ntigha Dumpsite in Isiala Ngwa of Abia State, Nigeria. Nigerian J Biochem Mol Biol 27(1):27–45
Odueze JC, Nwakonobi TU, Itodo IN (2017) Influence of physico-chemical characteristics of soils on heavy metal contamination in Makurdi, Benue state. J Environ Sci Toxicol Food Technol 11(5):84–92
Oijagbe IJ, Abubakar BY, Edogbanya PRO, Suleiman MO, Olorunmola JB (2019) Effects of heavy metals on soil microbial biomass carbon. J Biol Med 4(1):30–32
OlabimpeIyabo O, Oluwafemi JR, Grace AM, Oladele O (2017) Heavy metals and some physicochemical parameters in soil of major domestic dumpsites in Akure township, ondo state, South-Western Nigeria. Int J Eng Res Manag Technol 4(10):26–32
Olayinka OO, Akande OO, Bamgbose K, Adetunji MT (2017) Physicochemical characteristics and heavy metal levels in soil samples obtained from selected anthropogenic sites in Abeokuta, Nigeria. J Appl Sci Environ Manag 21(5):883–891
Osakabe Y, Osakabe K, Shinozaki K, Tran LSP (2014) Response of plants to water stress. Front Plant Sci 5:1–8
Oster JD, Shainberg I (2001) Soil responses to sodicity and salinity: challenges and opportunities. Soil Res 39(6):1219–1224
Paliwal KV, Gandhi AP (1976) Effect of salinity, SAR, Ca: Mg ratio in irrigation water, and soil texture on the predictability of exchangeable sodium percentage. Soil Sci 122(2):85–90
Pathak H, Rao DLN (1998) Carbon and nitrogen mineralization from added organic matter in saline and alkali soils. Soil Biol Biochem 30(6):695–702
Petropoulos SA, Daferera D, Polissiou MG, Passam HC (2008) The effect of water deficit stress on the growth, yield and composition of essential oils of parsley. Sci Hortic 115(4):393–407
Pinkerton A, Simpson JR (1986) Interactions of surface drying and subsurface nutrients affecting plant growth on acidic soil profiles from an old pasture. Aust J Exp Agric 26(6):681–689
Pressler Y, Foster EJ, Moore JC, Cotrufo MF (2017) Coupled biochar amendment and limited irrigation strategies do not affect a degraded soil food web in a maize agroecosystem, compared to the native grassland. GCB Bioenergy 9(8):1344–1355
Qadir A, Sharif M, Ziad M (2000) Homotheties of cylindrically symmetric static manifolds and their global extension. Classical Quant Gravity 17(2):345
Quirk J (1994) Interparticle forces: a basis for the interpretation of soil physical behavior. Adv Agron 53:121–183
Rahimi G, Kolahchi Z, Charkhabi A (2017) Uptake and translocation of some heavy metals by rice crop (Oryza sativa) in paddy soils. Agriculture (Poľnohospodárstvo) 63(4):163–175
Rahman M, Sabir AA, Mukta JA, Khan MMA, Mohi-Ud-Din M, Miah MG, Islam MT (2018) Plant probiotic bacteria Bacillus and Paraburkholderia improve growth, yield and content of antioxidants in strawberry fruit. Sci Rep 8:2504–2510
Rahmani N (2006) Effect of irrigation and nitrogen application on the quantity and quality of medicinal plant Marigold (Calendula officinalis L.). Master Thesis Agronomy, University of Takestan Branch.
Rauf S, Al-Khayri JM, Zaharieva M, Monneveux P, Khalil F (2015) Breeding strategies to enhance drought tolerance in crops. In: Al-Khayri JM et al (eds) Advances in plant breeding strategies; agronomic, abiotic and biotic stress traits. Springer, Berlin, pp 1–70
Reddy AR, Chaitanya KV, Vivekanandan M (2004) Drought induced responses of photosynthesis and antioxidant metabolism in higher plants. J Plant Physiol 161:1189–1202
Rengasamy P, Olsson KA (1991) Sodicity and soil structure. Soil Res 29(6):935–952
Rengasamy P, Sumner ME (1998) Processes involved in sodic behaviour. In: Sumner ME, Naidu R (eds) Sodic soils: distribution management and environmental consequences. Oxford University Press, New York, pp 35–50
Resnik ME (1970) Effect of mannitol and polyethylene glycol on phosphorus uptake by maize plants. Ann Bot 34(3):497–504
Rietz DN, Haynes RJ (2003) Effects of irrigation-induced salinity and sodicity on soil microbial activity. Soil Biol Biochem 35(6):845–854
Rui L, Sun W, Chao M, Ji C, Wang M, Ye B (2009) Leaf anatomical changes of Bruguiera gymnorrhiza seedlings under salt stress. J Trop Subtropical Bot 17(2):169–175
Sachan P, Lal N (2017) An overview of nickel (Ni2+) essentiality, toxicity and tolerance strategies in plants. Asian J Biol 2(4):1–15
Sacks MM, Silk WK, Burman P (1997) Effect of water stress on cortical cell division rates within the apical meristem of primary roots of maize. Plant Physiol 114(2):519–527
Sairam RK, Tyagi A (2004) Physiology and molecular biology of salinity stress tolerance in plants. Curr Sci 10:407–421
Sallam A, Alqudah AM, Dawood MFA, Baenziger PS, Börne A (2019) Drought stress tolerance in wheat and barley: advances in physiology, breeding and genetics research. Int J Mol Sci 20(13):31–37
Salman SA, Zeid SAM, Seleem E-MM, Abdel-Hafiz MA (2019) Soil characterization and heavy metal pollution assessment in Orabi farms, El Obour, Egypt. Bull Nat Res Cent 43:42. https://doi.org/10.1186/s42269-019-0082-1
Sankar B, Jaleel CA, Manivannan P, Kishorekumar A, Somasundaram R, Panneerselvam R (2007) Effect of paclobutrazol on water stress amelioration through antioxidants and free radical scavenging enzymes in Arachis hypogaea L. Colloids Surf B Biointerfaces 60(2):229–235
Sankar B, Jaleel CA, Manivannan P, Kishorekumar A, Somasundaram R, Panneerselvam R (2008) Relative efficacy of water use in five varieties of Abelmoschus esculentus (L.) Moench. Under water-limited conditions. Colloids Surf B Biointerfaces 62(1):125–129
Scherer HW (2001) Sulphur in crop production. Eur J Agron 14(2):81–111
Schmidt R, Kunkowska AB, Schippers JH (2016) Role of reactive oxygen species during cell expansion in leaves. Plant Physiol 172(4):2098–2106
Shah FUR, Ahmad N, Masood KR, Peralta-Videa JR, Ahmad FD (2010) Heavy metal toxicity in plants. In: Ashraf M, Ozturk M, Ahmad M (eds) Plant adaptation and phytoremediation. Springer, Dordrecht
Shaheen HL, Shahbaz M, Ullah I, Iqbal MZ (2012) Morpho-physiological responses of cotton (Gossypium hirsutum) to salt stress. Int J Agric Biol 14(6):980–984
Shainberg I, Levy GJ, Rengasamy P, Frenkel H (1992) Aggregate stability and seal formation as affected by drops’ impact energy and soil amendments. Soil Sci 154(2):113–119
Shao HB, Chu LY, Shao MA, Jaleel CA, Hong-Mei M (2008) Higher plant antioxidants and redox signaling under environmental stresses. C R Biol 331(6):433–441
Shekari F (2000) Effect of drought stress on phenology, water relations, growth, yield and quality canola, doctorate thesis in the field of Agriculture, University of Tabriz, p. 180.
Shi W, Ma X (2017) Effects of heavy metal Cd pollution on microbial activities in soil. Ann Agric Environ Med 24(4):722–725
Singh RP, Bauddh K, Sainger M, Sainger PA, Singh JS, Jaiwal PK (2010) Nitrogen use efficiency in higher plants under drought, high temperature, salinity and heavy metal contaminations. In: Jain V, Kumar PA (eds) Nitrogen use efficiency in higher plants. New India Publishing Agency (India) Pvt. Ltd., New Delhi, pp 99–123
Singh JS, Pandey VC, Singh DP (2011) Efficient soil microorganisms: a new dimension for sustainable agriculture and environmental development. Agric Ecosyst Environ 140:339–353
Sinsabaugh RL, Manzoni S, Moorhead DL, Richter A (2013) Carbon use efficiency of microbial communities: stoichiometry, methodology and modelling. Ecol Lett 16(7):930–939
Six D, Fily M, Blarel L, Goloub P (2005) First aerosol optical thickness measurements at dome C (East Antarctica), summer season 2003–2004. Atmos Environ 39(28):5041–5050
Smika DE, Haas HJ, Power JF (1965) Effects of moisture and nitrogen fertilizer on growth and water use by native grass 1. Agron J 57(5):483–486
Sobolev D, Begonia MF (2008) Effects of heavy metal contamination upon soil microbes: lead-induced changes in general and denitrifying microbial communities as evidenced by molecular markers. Int J Environ Res Public Health 5(5):450–456
Soriano MA, Villalobos FJ, Federes E (2002) Stress timing effects on sunflower harvest index. In: Villalobos FJ, Testi L (eds) VII Congress of the European Society for Agronomy. Sevilla, Consejería de Agricultural Pesca, Spain JA, pp 141–142
Sowerby A, Emmett B, Beier C, Tietema A, Penuelas J, Estiarte M, Freeman C (2005) Microbial community changes in heathland soil communities along a geographical gradient: interaction with climate change manipulations. Soil Biol Biochem 37(10):1805–1813
Specht JE, Chase K, Macrander M, Graef GL, Chung J, Markwell JP, Germann M, Orf JH, Lark KG (2001) Soybean response to water. A QTL analysis of drought tolerance. Crop Sci 41:493–509
Streeter JG (2003) Effects of drought on nitrogen fixation in soybean root nodules. Plant Cell Environ 26(8):1199–1204
Su C, Jiang L , Zhang W (2014) A review on heavy metal contamination in the soil worldwide: situation, impact and remediation techniques. Environ Skep Critic 3(2):24–27
Sudhir P, Murthy SD (2004) Effects of salt stress on basic processes of photosynthesis. Photosynthetica 42(2):481–486
Sun L, Wei P, Pu W, Wang B, Wu Y, Tan T (2016) The oil recovery enhancement by nitrogen foam in high-temperature and high-salinity environments. J Pet Sci Eng 147:485–494
Swift MJ, Heal OW, Anderson JM, Anderson JM (1979) Decomposition in terrestrial ecosystems. University of California Press, Berkeley
Syed R, Kapoor D, Bhat AA (2018) Heavy metal toxicity in plants: a review. Plant Arch 18(2):1229–1238
Tahir MHN, Mehid SS (2001) Evaluation of open pollinated sunflower (Helianthus annuus L.) populations under water stress and normal conditions. Int J Agric Biol 3:236–238
Tahkokorpi M, Taulavuori K, Laine K, Taulavuori E (2007) After effects of drought-related winter stress in previous and current year stems of Vaccinium myrtillus L. Environ Exp Bot 61:85–93
Thomas S, Anand A, Chinnusamy V, Dahuja A, Basu S (2013) Magnetopriming circumvents the effect of salinity stress on germination in chickpea seeds. Acta Physiol Plant 35(12):3401–3411
Tilman D, Cassman KG, Matson PA, Naylor R, Polasky S (2002) Agricultural sustainability and intensive production practices. Nature 8(418):671–677
Tiwari KK, Singh NK, Rai UN (2013) Chromium phytotoxicity in radish (Raphanus sativus): effects on metabolism and nutrient uptake. Bull Environ Contam Toxicol 91:339–344
Trenberth KE, Dai A, Van Der Schrier G, Jones PD, Barichivich J, Briffa KR, Sheffield J (2014) Global warming and changes in drought. Nat Clim Chang 4(1):17–22
Tripathi VN, Srivastava S (2006) Ni2+-uptake in Pseudomonas putida strain S4: a possible role of Mg2+-uptake pump. J Biosci 31(1):61–67
Turner LB (1985) Changes in the phosphorus content of Capsicum annuum leaves during water stress. J Plant Physiol 121(5):429–439
Uddin SR, Farid M, Saeed R, Ali S, Md R, Md TH, SAH B (2017) Citric acid enhanced the antioxidant defense system and chromium uptake by Lemna minor L. grown in hydroponics under Cr stress. Environ Sci Pollut Res 24(21):17669–17678
United States Salinity Laboratory Staff (1954) Diagnosis and improvement of saline and alkali soils, Agriculture Handbook No. 60. United States Department of Agriculture, Washington, DC
Usman AR, Al-Wabel MI, Abdulaziz AH, Mahmoud WA, El-Naggar AH, Ahmad M, Abdulelah AF, Abdulrasoul AO (2016) Conocarpus biochar induces changes in soil nutrient availability and tomato growth under saline irrigation. Pedosphere 26(1):27–38
Wall DH, Ritz K, Six J, Strong DR, van der Putten WH (2012) Soil ecology and ecosystem services. Oxford University Press, Oxford
Wang YP, Li QB, Shi JY, Lin Q, Chen XC, Wu W, Chen YX (2008) Assessment of microbial activity and bacterial community composition in the rhizosphere of a copper accumulator and a non-accumulator. Soil Biol Biochem 40(5):1167–1177
Wang X, Yuan B, Chen Y, Li X, Ren Y (2014) Integration of micro-filtration into osmotic membrane bioreactors to prevent salinity build-up. Bioresour Technol 167:116–123
Wang S, Xu Y, Norbu N, Wang Z (2018) Remediation of biochar on heavy metal polluted soils. IOP Conference Series: Earth and Environmental Science 108(4):042113. IOP Publishing
Wardle DA, Nilsson MC, Zackrisson O (2008) Fire-derived charcoal causes loss of forest humus. Science 320(5876):629–632
Webber M, Barnett J, Finlayson B, Wang M (2006) Pricing China’s irrigation water (working paper). School of Anthropology, Geography and Environmental Studies, The University of Melbourne, VIC, Australia
Wichern J, Wichern F, Joergensen RG (2006) Impact of salinity on soil microbial communities and the decomposition of maize in acidic soils. Geoderma 137(1-2):100–108
Wicke B, Smeets E, Dornburg V, Vashev B, Gaiser T, Turkenburg W, Faaij A (2011) The global technical and economic potential of bioenergy from salt affected soils. Energy Environ Sci 4:2669–2681
Wilson A, Punginelli C, Gall A, Bonetti C, Alexandre M, Routaboul JM, Kerfeld CA, Van Grondelle R, Robert B, Kennis JT, Kirilovsky D (2008) A photoactive carotenoid protein acting as light intensity sensor. PNAS 105(33):12075–12080
Wong VN, Dalal RC, Greene RS (2009) Carbon dynamics of sodic and saline soils following gypsum and organic material additions: a laboratory incubation. Appl Soil Ecol 41(1):29–40
Wu QS, Xia RX, Zou YN (2008) Improved soil structure and citrus growth after inoculation with three arbuscular mycorrhizal fungi under drought stress. Eur J Soil Biol 44(1):122–128
Wu W, Mhaimeed AS, Al-Shafie WM, Ziadat F, Dhehibi B, Nangia V, De Pauw E (2014) Mapping soil salinity changes using remote sensing in Central Iraq. Geoderma Reg 2:21–31
Wuana RA, Okieimen FE (2011) Heavy metals in contaminated soils: a review of sources, chemistry, risks and best available strategies for remediation. Int Sch Res Notices Ecol 402647:1–20
Wullschleger SD, Yin TM, DiFazio SP, Tschaplinski TJ, Gunter LE, Davis MF, Tuskan GA (2005) Phenotypic variation in growth and biomass distribution for two advanced-generation pedigrees of hybrid poplar. Can J For Res 35(8):1779–1789
Xian Y, Wang M, Chen W (2015) Quantitative assessment on soil enzyme activities of heavy metal contaminated soils with various soil properties. Chemosphere 139:604–608
Xiao X, Wang M, Zhu H, Guo Z, Han X, Zeng P (2017) Response of soil microbial activities and microbial community structure to vanadium stress. Ecotoxicol Environ Saf 142:200–206
Xu Y, Seshadri B, Sarkar B, Wang H, Rumpel C, Sparks D, Farrell M, Hall T, Yang X, Bolan N (2018) Biochar modulates heavy metal toxicity and improves microbial carbon use efficiency in soil. Sci Total Environ 621:148–159
Yadav SK (2010) Heavy metals toxicity in plants: an overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. Soil Biol Biochem 76(2):67–179
Yadav G, Srivastava PK, Singh VP, Prasad SM (2014) Light intensity alters the extent of arsenic toxicity in Helianthus annuus L. seedlings. Biol Trace Elem Res 158:410–421
Yuan B, Shen X, Li X, Xu C, Wang S (2007) Mitogen-activated protein kinase OsMPK6 negatively regulates rice disease resistance to bacterial pathogens. Planta 226(4):953–960
Yuan P, Wang J, Pan Y, Shen B, Wu C (2019) Review of biochar for the management of contaminated soil: preparation, application and prospect. Sci Total Environ 659:473–490
Zareian J (2004) Effects of drought on the different stages of growth, and growth traits, varieties of winter canola, MS Thesis Agronomy, Faculty of Agriculture, Islamic Azad University of Khorasgan
Zeid IM, Ghazi SM, Shedeed ZA, Nabawy DM (2018) Biochar mitigates cadmium stress on alfalfa seeds during germination. Int J Prog Sci Technol 6:251–261
Zhang H, Wang X, Wang S (2004) A study on stomatal traits of Platanus acerifolia under urban stress. J Fudan Univ 43:651–656
Zhang H, Kim MS, Sun Y, Dowd SE, Shi H, Paré PW (2008) Soil bacteria confer plant salt tolerance by tissue-specific regulation of the sodium transporter HKT1. Mol Plant-Microbe Interact 21(6):737–744
Zhang FP, Li CF, Tong LG, Yue LX, Li P, Ciren YC, Cao CG (2010) Response of microbial characteristics to heavy metal pollution of mining soils in Central Tibet, China. Appl Soil Ecol 45(3):144–151
Zhao W, Sun Y, Kjelgren R, Liu X (2015) Response of stomatal density and bound gas exchange in leaves of maize to soil water deficit. Acta Physiol Plant 37(1):1704. https://doi.org/10.1007/s11738-014-1704-8
Zheng R, Chen Z, Cai C, Tie B, Liu X, Reid BJ, Huang Q, Lei M, Sun G, Baltrėnaitė E (2015) Mitigating heavy metal accumulation into rice (Oryza sativa L.) using biochar amendment a field experiment in Hunan China. Environ Sci Pollut Res 22:11097–11108
Zheng R, Sun G, Li C, Reid BJ, Xie Z, Zhang B, Wang Q (2017) Mitigating cadmium accumulation in greenhouse lettuce production using biochar. Environ Sci Pollut Res 24(7):6532–6542
Zornoza R, Mataix-Solera J, Guerrero C, Arcenegui V, García-Orenes F, Mataix-Beneyto J, Morugán A (2007) Evaluation of soil quality using multiple lineal regression based on physical, chemical and biochemical properties. Sci Total Environ 378(1-2):233–237
Zou L, Morris G, Qi D (2008) Using activated carbon electrode in electrosorptive deionisation of brackish water. Desalination 225(1–3):329–340
Acknowledgments
Kuldeep Bauddh is thankful to the Science and Engineering Research Board (SERB) for the award of Research Grant (EEQ/2017/000476). Author Abhishek Guldhe is thankful to the Department of Biotechnology, Govt. of India for the Ramalingaswami Fellowship.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Kumari, K. et al. (2020). Biochar Amendment in Agricultural Soil for Mitigation of Abiotic Stress. In: Bauddh, K., Kumar, S., Singh, R., Korstad, J. (eds) Ecological and Practical Applications for Sustainable Agriculture. Springer, Singapore. https://doi.org/10.1007/978-981-15-3372-3_14
Download citation
DOI: https://doi.org/10.1007/978-981-15-3372-3_14
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-15-3371-6
Online ISBN: 978-981-15-3372-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)