
Abstract
As sessile organisms anchored to their substrate, plants have to develop in such a way that their organs can fulfill essential primary functions, which include photosynthesis, gas exchange and reproduction for shoots, and anchoring as well as water and nutrients uptake for roots. To do so, these organs have to use directional information within their environments as growth guides. Gravity, a constant parameter on Earth, is one of the cues used by most organs to direct growth, a process named gravitropism. Typically, shoots will grow against the gravity vector whereas roots will follow it. Furthermore, lateral organs will grow along shallower vectors relative to gravity, whose obliqueness is dictated by endogenous/hormonal and environmental cues. In this chapter, we review the molecular mechanisms that allow angiosperm organs to use gravity as a growth guide. Gravity-sensing cells named statocytes contain dense starch-filled plastids (amyloplasts) that sediment within their cytoplasm. These cells are located in the columella region of the root cap and in the endodermis that surrounds the vasculature in shoots. Amyloplast sedimentation in these cells promotes a polarization of auxin efflux facilitators to the bottom membrane, creating a downward flow of auxin that results in a lateral gradient across the stimulated organ. Differential auxin accumulation on opposite flanks of the organ results in differential cellular elongation upon transmission to the site of response, a process that is responsible for upward curvature in shoots and downward growth in roots. Lateral organs, on the other hand, respond to similar stimuli by developing weaker lateral auxin gradients, leading to shallower growth angles from gravity. An abundance of research carried out by multiple laboratories around the world has recently led to important new insights into the mechanisms that govern these complex processes and the machinery that fine-tunes them to ultimately yield highly controlled and amazingly complex responses. This chapter attempts to discuss these mechanisms and identify some of the areas in need of further investigation in this important area of plant biology.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Amyloplasts
- Columella cells
- Cyclic nucleotide-gated ion channel
- Gravistimulation
- Gravity set point angle
- Root cap
- Small auxin up RNA (SAUR)
- Statocytes
1 Introduction
As sessile organisms, plants have to direct the growth of their organs to reach out for light, oxygen, carbon dioxide, water and nutrients. Hence, cues required for guidance should be associated, either directly or indirectly, with these environmental gradients in such a way that each organ can better fulfill its primary functions, including gas exchange, photosynthesis and reproduction for shoots, and plant anchoring as well as water and nutrient uptake for roots. One key parameter in the environment that broadly favors organs growth in directions that are compatible with their primary functions is gravity. Indeed, plant organs are equipped with machinery that allows them to detect the direction of gravity and guide their growth relative to it. Named gravitropism, this directional growth response to gravity has received much attention since its recognition two centuries ago (Knight 1806).
Upon germination, seedling primary roots tend to grow vertically downward into the soil, whereas shoots tend to expand in the opposite direction, reaching out for light. Both organs can do this even when the seedlings are exposed to complete darkness, using gravity as a guide. This directional growth response to gravity remains critical throughout the life cycle of a plant. In fact, different plant organs will follow distinct growth vectors relative to gravity. For many species, the primary organs will grow along the gravity vector, as discussed above, whereas lateral organs will emerge from a lower-order organ at a stereotypical angle from it, before curving toward a target vector at a defined angle from the vertical, named gravity set point angle (GSA) (Firn and Digby 1997). Named plagiogravitropism, the latter process may facilitate exploration of the three-dimensional space around a plant, improving its access to essential resources and, consequently, influencing its performance.
In natural environments, resources are often distributed unevenly in the immediate vicinity of a plant. For instance, light will be differently oriented depending on the time of the day, and its intensity and spectrum will vary depending on the density of shading plants in the canopy, cloud cover and period of day and year. Similarly, neighboring soil particles may have varied biophysical properties that confer distinct abilities to retain water, ions, or specific nutrients, thereby creating local gradients in humidity and/or nutrients that may directly influence root growth rate, direction, and/or branching. Hence, the gravity set point for a defined plant organ will be established in coordination with responses to a variety of developmental, environmental, and/or hormonal cues, leading to drastically distinct morphologies under diverse conditions, which may help improve the plant’s ability to cope with a rapidly changing environment (Firn and Digby 1997).
In this chapter, we will introduce the reader to the mechanisms that govern plant responses to gravity, with emphasis on organs that undergo primary growth from apical meristems. Many of the experiments described in this chapter were carried out on Arabidopsis thaliana roots and shoots. This is because most of the recent work uncovering key mechanisms involved in gravity sensing and signal transduction has been carried out using this model dicot organism.
We will first describe the mechanisms that allow shoots and roots to grow and then discuss the mechanisms that govern gravitropism in these organs.
1.1 Cell Division and Elongation Contribute to Plant Organs’ Growth
To grow, plant organs use a combination of cell divisions in apical meristems and cell elongation in subapical regions named the elongation zones, where cells also differentiate into defined cell types (Fig. 5.1). In roots, the apical meristem is covered by a cap, which contributes to environmental sensing as well as meristem protection against mechanical damage during root growth in soil. The root cap is made of large central columella cells as well as lateral cap cells that wrap around the root meristem, and tip cells. Both columella and lateral cap cells are being constantly replaced by new cells originating from the asymmetrical division of lateral and distal initial cells that surround the quiescent center within the promeristem. As we will see later on in this chapter, the central columella cells of the root cap are critical for gravity sensing whereas the lateral cap cells contribute to signal transmission from the gravisensing cap to the graviresponding elongation zone.

Upon reorientation within the gravity field, plant organs respond by developing tip curvatures that involve differential cell elongation between upper and lower flanks of elongation zones. Panels (a) and (b) show the cellular organization and longitudinal patterning of the responding growth zone of dicot Arabidopsis thaliana root (a) and hypocotyl (b). Panels (c–f) show the gravitropic responses of (c) a gravistimulated Arabidopsis thaliana seedling, (d) an Arabidopsis inflorescence stem 0 and 120 min after reorientation, and (e, f) monocot Brachypodium distachyon seedling (e) and stem (f)
Shootward to the tip-organizing quiescent center, additional initials also divide asymmetrically to generate concentric files of cells that will ultimately form the different tissue types that constitute a root, including the lateral root cap, epidermis, cortex, endodermis, pericycle and the rest of the vasculature. Rapid anticlinal cell divisions occur along a defined region of the root tip named meristem. As more divisions occur in that region, those cells that are located more proximally (closer to the shoot) eventually cease division after reaching a defined distance from the root tip. Those interphase cells start expanding along a vector that is parallel to the longitudinal axis of the root. They continue to elongate and differentiate as their distance from the tip increases due to continued division at the meristem. Ultimately, those cells will stop expanding and differentiate into the different cell types listed above. The region where cells elongate is named the elongation zone (EZ). This region is, in fact, complex, being composed of two distinct sub-regions: (1) a distal elongation zone (DEZ) and (2) a central elongation zone (CEZ). The DEZ is made of cells that transit from a state of division to a state of elongation. The region flanking the DEZ on its shootward side is the CEZ. It is made of cells that elongate at rates higher than 30% of their maximal value (Fig. 5.1a) (Ishikawa and Evans 1995). The DEZ is the site where the curvature response to gravistimulation (GS) initiates in roots, as discussed below (Ishikawa and Evans 1995).
Arabidopsis stems have also been used quite extensively in investigations of gravitropism. In young Arabidopsis seedlings, an embryonic stem named the hypocotyl separates the root from the cotyledons and shoot apical meristem. This hypocotyl is made of several concentric layers of cells, including the epidermis, the cortex (2 layers), the endodermis and the stele containing vasculature tissues (Fig. 5.1b). The hypocotyl grows only by cell elongation, at least when etiolated (Gendreau et al. 1997). In fact, the pattern of cell elongation differs significantly between light-grown and dark-exposed hypocotyls. In light-grown seedlings, all epidermal cells elongate continuously during the entire growth period. However, in etiolated seedlings, elongation occurs along a steep acropetal spatial and temporal gradient (Gendreau et al. 1997). In both cases, the gravitropic curvature (Fig. 5.1c) occurs in the region of maximum cellular expansion.
In adult plants, the inflorescence stems are also made of several tissue types arranged in concentric circles, including the epidermis (one layer of cells), the cortex (three layers of cells), the endodermis (one cell layer), and the stele containing the vasculature. Cell divisions occur in the apical meristems, whereas elongation occurs along most of the stem length in young Arabidopsis plants. In older plants cellular elongation is restricted to more distal regions of the stem. The rootward side of older inflorescence stems is formed of mature cells that are surrounded by inextensible walls that contain lignin (Weise et al. 2000). The gravitropic curvature occurs only in the distal region of older mature stems, within their elongation zone (Fig. 5.1d).
In monocots, seedling coleoptiles and shoot pulvini develop strong gravitropic curvatures (Fig. 5.1e). The coleoptiles are seedling leaf sheaths that enclose the primary leaves and protect them as they grow up through the soil. Coleoptiles grow mostly by cell elongation, although some evidence of cell division has also been noted in wheat (Lu et al. 2006). The tip of a coleoptile is important for its growth and gravitropism, as a main source of auxin. The growing coleoptile remains capable of strong gravitropism as long as the developing leaves remain enclosed within it. As soon as the first growing leaf emerges from the tip, the coleoptile loses its ability to develop a gravicurvature (Iino 1995).
In adult monocot plants, shoot gravitropism typically involves the contribution of pulvini, which are short segments of tissue that are apical to the nodes and collectively contribute to bringing a shoot tip that was previously prostrated by wind or rain, back to a more vertical position. In Panicoid species like maize, the pulvini constitute disc-shaped segments of the stem, whereas the pulvini of Festucoid grasses, such as wheat, oat, and barley, are made of a tissue that encircles the leaf sheath immediately apical to the point where it attaches to the node. The cells making up the pulvini of an adult plant typically do not grow any more in the absence of a gravistimulus. However, when monocot stems are being prostrated by wind or rain, cells at the bottom side of several pulvini (two or four in maize) along the stem resume elongation, resulting in local segmental upward curvature (Fig. 5.1f). In maize, each pulvinus can provide a maximum of 30-degree curvature in response to gravistimulation. This process plays an important role in agriculture because it keeps seed away from soil moisture and pathogens, accessible to mechanical harvesting, even after the plants have been prostrated by heavy storms.
In addition to this important economical impact of the gravitropic response in cereals, pulvini have been the target of many investigations on plant gravitropism because their responses are very slow, making it possible to independently investigate the molecular mechanisms that contribute to gravity sensing and/or signal transduction, relative to those involved in the curvature response.
After this brief description of root and shoot elements that display gravitropism in monocot and dicot plants, we will now discuss some of the cellular and molecular mechanisms that drive gravitropism in plants, a process that includes several important steps: (1) gravity sensing and signal transduction, (2) signal transmission, and (3) curvature response.
2 Gravity Sensing and Signal Transduction
2.1 Amyloplast Settling Within Statocytes Contributes to Gravity Sensing
It has long been recognized that cells located at the center of the root cap (the columella cells) and the endodermis tissue surrounding the vasculature in shoots, coleoptiles and pulvini, are well suited for gravity sensing in plants because they contain dense starch-filled plastids that sediment to the bottom upon plant reorientation within the gravity field (Darwin 1880; Haberlandt 1900; Nemec 1900). Those plastids function as statoliths (solid structures/organelles that settle to the bottom of a cell). So, how do we know these cells actually perform a gravity-sensing function in plants?
To illustrate some of the experiments that allowed scientists to answer this question, we will use the root as an example. Indeed, roots have been extensively used to investigate the cellular and molecular mechanisms that govern gravitropism because they physically separate the primary site for gravity sensing (the cap) from the locale of curvature response (the EZ; Fig. 5.1a), facilitating assignment of key molecular mechanisms to distinct phases of a graviresponse.
Several experiments have demonstrated a key role for the root cap in gravity sensing. For example, removing the cap from primary roots by surgical ablation (Barlow 1995), killing specific cap cells with heavy-ion microbeam irradiation (Tanaka et al. 2002), genetically obliterating cap cells by targeted expression of the diphtheria toxin (Tsugeki and Fedoroff 1999) or mutating transcription factor genes that are needed for proper root cap specification (Wang et al. 2005), all resulted in roots that continued to grow, but were unable to develop a gravitropic response. Hence, the root cap is important for root gravitropism. However, does it function in the gravity-sensing phase of the process?
Before explaining some of the experiments that addressed the role of the root cap in gravisensing, we will discuss the concept of gravisensitivity. We will start this discussion by recognizing that plant organs appear to respond to transient gravistimulation by developing curvatures that vary linearly with the logarithm of the dose of stimulation, defined as the product of gravity level multiplied by the duration of stimulation. Known as the reciprocity law, this relationship between curvature angle and dose of stimulation implies that plant organs might be able to sense the force of gravity and use this information to guide their growth. This concept of force sensing has been widely accepted in the field for many years, and researchers have used it to develop methods aimed at evaluating the gravisensitivity of plant organs (although we now know that this is an oversimplification of the process, as discussed later on in this chapter). One of these methods involves the quantification of organ curvature responses to small doses of gravistimulation. On Earth, such experiments involve reorienting the plant within the gravity field (1xg) for short periods of time and quantifying the resulting curvatures. A linear function associating the angles of curvature to the logarithm of stimulation times is then retrofitted to the data and extended to the time axis, intersecting it at a value that can be defined as the minimal gravistimulation time needed to induce a detectable curvature response. This minimal time is named the presentation time, and it is often viewed as a measure of gravisensitivity (Fig. 5.2).

Methods used to evaluate plant organ sensitivity to gravistimulation. Panel (a). Seedlings growing on agar-based media in petri dishes are reoriented to the horizontal for short periods of time (2, 5, and 10 mins in this case). After stimulation, the plates are transferred into a clinostat and rotated at a speed of 1–4 rpm for a period of 5 h, thereby constantly modifying the organs’ orientation relative to gravity and avoiding additional gravistimulation (middle section of the drawing). During this period of clinorotation, plant organs will develop a curvature response to the initial stimulus. At the end of this process, the orientations of organ tips are measured (right side of panel a), providing data that relate the resulting tip angle to the dose (period) of gravistimulation. Panel (b). Two mathematical models have been used to represent the relationship between angle of curvature and dose/period of gravistimulation: the hyperbolic model (left) and the logarithmic model (right). The logarithmic model intersects the time axis at a point named the presentation time, which represents the minimal gravistimulation time needed to activate a curvature response. The hyperbolic model, on the other hand, intersects the time axis at the origin. It has been proposed that the slope of the curve at the origin (S = a/b) is another measure of gravisensitivity. The data points shown in these two graphs are identical, deriving from Arabidopsis roots subjected to the protocol described in panel A by Caspar and Pickard (1989), as cited in Perbal et al. (2002)
While this strategy of presentation time determination seems simple at first glance, it is, in reality, complicated by the fact that most plant organs have presentation times that are much shorter than the time needed for the curvature responses to develop. Yet, the plants cannot be returned to the vertical after having been gravistimulated, to allow for curvature development, because this reorientation would constitute a second gravistimulus that would confound the data. To resolve this problem, researchers have used a rotating device, named the clinostat, to randomize the plant orientation within the gravity field while it responds to an initial transient gravistimulus.
Therefore, a typical experiment aimed at evaluating the presentation time of a plant organ involves the following sequence of events (Fig. 5.2). First, short gravistimuli are provided by reorienting the plants within the gravity field. After a defined period of gravistimulation, the plants are positioned on a clinostat, which continuously rotates them along a horizontal axis at a speed of approximately 1–4 revolutions per minute (rpm), over a period of 3–5 h. During this period, the plant organs are not exposed to directional gravistimulation for enough time to reset the gravity signal transduction pathway. Consequently, they develop a curvature that is a direct consequence of the initial short gravistimulus that preceded the clinorotation.
Many researchers have used the presentation time to represent gravisensitivity in plants. However, other investigators have warned that this concept might be misleading. First, earlier experiments had indicated that successive exposures to very short pulses of gravistimulation (much shorter than the presentation time) still allow plant organs to develop curvature responses, implying that such short stimuli are still perceived by the plant. Second, Dr. Perbal and his collaborators (Perbal et al. 2002) noted that the logarithmic model correlating observed angles of curvature to the logarithm of the dose/time of gravistimulation is actually not the best fit to the observed experimental data. In fact, a hyperbolic model better represents those data (Fig. 5.2b). It is quite significant that such a hyperbolic model intersects the X-axis (dose of stimulation) at the origin, invalidating the presentation time/dose concept. Therefore, these authors proposed to use the slope of the hyperbolic curve at the origin to estimate gravisensitivity.
Choosing between these two alternative methods of gravisensitivity determination would require a better understanding of the molecular mechanisms that govern gravity sensing and signal transduction in plants. Short of such information, researchers have adopted both models to evaluate root gravisensitivity in mutants or pharmacologically treated seedlings. In general, these models resulted in similar ranking of gravisensitivities between mutants/treated seedlings and wild-type/control samples (for instance, see (Blancaflor 2013)).
The two methods described above allow estimation of organ gravisensitivity on Earth, exposed to 1xg. However, access to the microgravity environment of the International Space Station allows the design of more direct experiments aimed at measuring plant organ sensitivity to gravity. Under microgravity, fractional g conditions can be created by centrifugation. The minimal centrifugation force needed to trigger an organ tip curvature can be determined. Although very expensive, these experiments have demonstrated that the plant gravisensing machinery is overbuilt, reacting to forces that are well below those encountered under 1 g conditions on Earth (Kiss et al. 2012).
Despite the heavy reliance on presentation time to evaluate gravisensitivity of plant organs, it is important to note that recent studies have suggested that the force sensor model of gravity sensing described above, which is key to the concept of presentation time as a measure of gravisensitivity, may actually not be an adequate or complete representation of the actual mechanism used by plant statocytes to sense gravity. In fact, a mechanism that would sense the inclination of the plant relative to gravity rather than the force of gravity may actually be at play (Chauvet et al. 2016). This inclination sensor model of gravity sensing will be described in the next section.
2.2 The Columella Cells Serve as Statocytes in Roots
The experiments described in the previous section suggest that the root cap contributes to gravity sensing in roots. Which cap cells contribute to this process? As noted above, a simple morphological and cytological analysis of the root cap provides a possible answer to this question. In fact, at the center of the root cap, a group of large cells (the columella cells) appear well suited to serve as statocytes. Devoid of large vacuoles, these cells display a unique organization that suggests a role in gravity sensing. Their nucleus is located in their upper (shootward) half, and their central cytoplasm is depleted of organelles. The endoplasmic reticulum (ER) lines the periphery at the distal side of these cells, as do most other organelles. Importantly, these cells contain large and dense starch-filled plastids (amyloplasts). In most other cell types within the plant, plastids are tightly associated with the actin cytoskeleton network. However, in the columella cells, these organelles are only loosely associated with the cytoskeleton. As a consequence, they do sediment to the bottom while also bouncing around in a saltatory movement that probably derives from transient interactions with a highly dynamic actin cytoskeleton present in these cells as well as with subtending ER membranes (Bérut et al. 2018; Leitz et al. 2009). When a plant is reoriented within the gravity field, columella amyloplasts (also called statoliths) quickly settle to the new bottom side of the cells, following a liquid-like behavior that is dependent upon cellular activity (Bérut et al. 2018). This amyloplast system repositioning within the statocytes triggers a gravity signal transduction pathway that is largely uncharacterized.
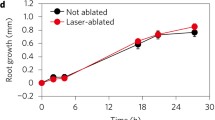
How do we know that the columella cells of the root cap contribute to gravity sensing in plants? To answer this question, Alison Blancaflor and his collaborators evaluated the presentation time (interpreted as a measure of gravisensitivity) of Arabidopsis roots after distinct cells of the root cap had been killed by laser ablation (Blancaflor et al. 1998). In these experiments, wild-type Arabidopsis thaliana seedling roots (Columbia accession) displayed a presentation time of approximately 1.16 min. When cells from layers S1 and S2 of the columella region of their root caps were ablated, the presentation time was increased to 7.13 min, suggesting a decrease in root gravisensitivity relative to control (Blancaflor et al. 1998). A similar alteration was observed when all central columella cells were ablated, suggesting that layers S1 and S2 of columella cells contribute most to gravisensing in roots (Blancaflor et al. 1998). On the other hand, ablating lateral cap or distal tip cells did not affect much the presentation time of treated roots, confirming that gravisensing is mostly performed by a few specialized cells at the center of the root cap – precisely those that contain amyloplasts with the highest sedimentation capability.
Does amyloplast repositioning within the statocytes contribute to gravity sensing? Investigations of gravitropism in starchless and starch-deficient mutants seem to support a role for amyloplast sedimentation in gravity sensing. Indeed, root cap amyloplasts of starchless mutants do not sediment under 1 g because of their lower density in the absence of starch. This phenotype is associated with altered gravitropism, suggesting a role for amyloplast sedimentation in gravity signal transduction (Band et al. 2012; Kim et al. 2011; Kiss et al. 1989, 1996; MacCleery and Kiss 1999). Second, starch-deficient mutants that carry amyloplasts with limited amount of starch do not show evidence of amyloplast sedimentation upon gravistimulation under normal conditions. They also show an altered gravitropic phenotype. However, increased g forces provided by centrifugation can promote a lateral displacement of these starch-deficient amyloplasts, allowing resumption of gravitropism (Fitzelle and Kiss 2001). On the other hand, mutations that affect starch-degrading enzymes, such as starch excess 1 (sex1) in Arabidopsis thaliana, or conditions that result in larger amyloplasts, are associated with greater sensitivity to gravity (Vitha et al. 2007).
Another key experiment that addressed a role for amyloplast settling in gravity signaling relied on the use of high-gradient magnetic fields to laterally displace amyloplasts within the statocytes of vertically oriented seedlings. Being diamagnetic, starch grains can be displaced laterally by application of a local high-gradient magnetic field. The corresponding ponderomotive force is sufficient to displace the statoliths in a direction that is dictated by the geometry of the gradient. Placing paramagnetic particles in proximity of vertical plant organs within a magnetic field creates local high-gradient magnetic fields that are sufficiently large to mobilize amyloplasts within the root cap columella cells, moving them laterally. This lateral displacement of amyloplasts within the statocytes was associated with the development of a tip curvature in the direction dictated by statolith movement (Kuznetsov and Hasenstein 1996). The curvature was not an indirect consequence of exposure to magnetic fields because it did not occur when starchless mutants (whose plastids cannot be displaced by the magnetic fields) were used.
While amyloplast movement within the statocytes is sufficient to trigger a tip curvature, it should still be cautioned that these statoliths are not completely free to sediment. As mentioned above, a dynamic actin-filament network is also present, which transiently interacts with the statoliths, promoting saltatory movements that may fine-tune the gravitropic response (Leitz et al. 2009; Zheng et al. 2014). Interestingly, mutations that affect actin dynamics, such as distorted1, lead to slower kinetics of gravitropism (Zheng et al. 2014). On the other hand, treatments with agents that affect actin-filament dynamics (such as latrunculin B or D) resulted in enhanced kinetics of gravitropism, increased gravisensitivity, and gravitropic signal persistence leading to overshooting the vertical at the end of a response (Hou et al. 2003; Yamamoto et al. 2002).
The experiments described above document a key role for root cap amyloplast sedimentation in gravisensing. However, several experiments have also suggested the existence of a secondary site for gravisensing in roots, localized at the DEZ (Kiss et al. 1999). Indeed, to better characterize the spatiotemporal distribution of gravisensing in responding plant organs, investigators developed a novel device named the rotato, which maintains a specific region of the root tip at a predefined angle from the vertical over time. The rotato is a microscope equipped with a rotating vertical platform that carries petri dishes with growing seedlings in front of the objective (Fig. 5.3a). This platform is equipped with a motor that automatically rotates it to maintain a defined angle between a small, predefined segment of the root tip and the vertical. If the system is set up to maintain the root cap at a defined angle from the vertical over time, the platform will continue to rotate as the root curves, attempting to return the tip to the vertical (Fig. 5.3b). The speed of rotation defines the kinetics of tip curvature.

Rotato is a useful device to study gravitropism. Panel (a) shows two rotato devices working side-by-side in the Wolverton laboratory at Ohio Wesleyan University. Each device is made of a camera located in front of a rotating platform that holds a Petri dish with growing seedlings. An automatic software controls the rotation of the platform to maintain a defined region of the plant organ (root in this case) at a pre-specified angle from the vertical. The software records the speed of stage rotation needed to fulfill this condition. Panel (b) (copied from Wolverton et al. 2002) shows images from a 4-h time-lapse analysis of an Arabidopsis root growing on the rotato system, which was set up to maintain the root tip region at 90° from the vertical. The stage rotates clockwise as the root curves, to maintain the tip at 90°. These two panels were kindly provided by Dr. Chris Wolverton, Ohio Wesleyan University
If the rotato is set up to maintain a subapical region of the root tip at a defined angle from the vertical, the tip is expected to return to the vertical. As soon as the vertical is reached, the root should stop curving, and the tip should resume vertical downward growth without platform rotation (as the gravity set point angle is reached). However, when Wolverton and his collaborators carried out the latter experiment by attempting to keep the DEZ at a constant angle from the vertical, the platform continued to rotate even after the cap had reached the vertical and gone beyond it (Wolverton et al. 2002). This result was surprising and important because it suggested that cells within the root DEZ may also be able to sense gravity. In fact, this conclusion could be corroborated by other observations. For instance, in maize seedlings, decapped roots remain somewhat gravitropic, a response that can be enhanced by disrupting actin filaments or manipulating myosin activity (Mancuso et al. 2006).
These observations are puzzling because the cells in the DEZ of roots do not contain starch-filled plastids, suggesting that a different mechanism of gravity sensing might be at play in these cells. In fact, researchers have postulated that DEZ cells might be able to sense gravity by detecting the overall pressure exerted by the protoplast on its wall. A similar mechanism was previously suggested to explain gravity sensing by the large internodal cells of Chara and rice root tips (Staves et al. 1992, 1997).
2.3 Transduction of the Information Conferred by Amyloplast Movement and/or Position Within the Statocytes
The conclusion of the experiments described in the previous sections of this chapter is that the settling of amyloplasts within the statocytes constitutes a first step in gravity sensing by plants. We will now describe the molecular mechanisms that govern gravity signal transduction in root statocytes.
In roots, the physical separation that exists between sites of gravity sensing and the location of curvature response implies that there is a need for communication between these two regions in order for an adequate response to occur. In other words, the physical information provided by amyloplast settling in the columella cells of the cap has to be converted into a biochemical signal that needs to be transmitted to the DEZ to trigger the resulting curvature. As discussed below, this signal takes the form of a lateral auxin gradient generated across the root cap upon gravistimulation. A first question that comes to mind is: How is amyloplast settling within the statocytes transduced into a repolarization of auxin transport, leading to the formation of a lateral auxin gradient across the cap?
Two models have been suggested to explain the transduction of information provided by amyloplast settling in the statocytes. The first model suggests that sedimenting statoliths press upon sensitive membranes on the inside of the statocytes, triggering the opening of mechanosensitive ion channels, with consequent Ca2+ spikes in the cytoplasm (Sievers et al. 1991). As a second messenger, Ca2+ would trigger a local transduction pathway leading to statocyte repolarization. The second model (named “ligand-receptor hypothesis”) postulates the existence of ligands on the surface of sedimenting statoliths. These ligands would interact with receptors located within sensitive membranes on the side of the statocytes to activate the gravity signaling pathway (Limbach et al. 2005). This model emerged from investigations of gravitropism in single-cell rhizoids from the green algae Chara. Whether it also applies to the statocytes of flowering plant organs remains unknown.
As pointed out above, the first model postulates a contribution of mechanosensitive ion channels to gravity signaling within the statocytes. Unfortunately, the channels responsible for this process have not been identified. However, pharmacological studies using drugs that inhibit the opening of ion channels, chelate Ca2+, or inhibit Ca2+ sensors, such as calmodulins, calmodulin-like proteins, and/or Ca2+/calmodulin-dependent protein kinases, strongly affected plant gravitropism, suggesting a role for Ca2+ in gravitropic signaling (Lu and Feldman 1997; Sinclair and Trewavas 1997). Unfortunately, gravity-induced changes in cytosolic Ca2+ levels within the statocytes have not been documented. For instance, investigators have used a transgenic AEQUORIN Ca2+-reporter system to analyze possible changes in cytosolic Ca2+ levels early in response to gravistimulation. This system involves expressing a protein named AEQUORIN in transgenic plants. When present in the cytoplasm, this protein can be altered to emit photons in a Ca2+-dependent manner by simply adding a luminophore named coelenterazine to the medium. This compound is taken up by the plant and accumulates in the cytoplasm of exposed cells. Expressed AEQUORIN binds to available coelenterazine within the cell, forming a complex that emits light in a Ca2+-dependent manner. Using this system, investigators demonstrated the existence of biphasic spikes in cytosolic Ca2+ within seconds of a gravistimulus (Plieth and Trewavas 2002; Toyota et al. 2008). Yet, these Ca2+ spikes derived only from hypocotyls and petioles, not roots, and they could not be assigned to specific cell types because the signal was too small to allow cell-specific mapping (Toyota et al. 2008). In fact, we now know that Ca2+ contributes to plant cell responses to auxin ((Monshausen et al. 2011); see below). Whether it also contributes to gravity signal transduction in the statocytes remains unclear.
A possible involvement of Ca2+ in gravity signal transduction has also been suggested based on observations of changes in inositol 1,4,5-trisphosphate (InsP3) levels in oat coleoptiles and Arabidopsis inflorescence stems upon gravistimulation (Perera et al. 2006). As a component of the phosphoinositide-signaling pathway, InsP3 is a signaling molecule that has been implicated in the regulation of cytosolic Ca2+ levels and vesicle trafficking in both animal and plant systems (Munnik and Nielsen 2011; Perera et al. 2006). Interestingly, overexpression of human inositol trisphosphatase, which hydrolyzes InsP3, in Arabidopsis roots, stems and hypocotyls, caused altered gravitropism (Perera et al. 2006). Similarly, inhibiting the synthesis of InsP3 with a phospholipase C inhibitor (U73122) led to altered root gravitropism (Andreeva et al. 2010), whereas mutations affecting the Arabidopsis INOSITOL POLYPHOSPHATE 5-PHOSPHATASE 13 gene enhanced gravitropism while reducing endomembrane trafficking (Wang et al. 2009). Hence, InsP3 and/or other molecules of the phosphoinositide-signaling pathway may play a role in gravity signaling (Perera et al. 2006). Whether this InsP3 contribution to gravity signaling implies a role for Ca2+ in this process remains unclear, though, as the ability of InsP3 to gate the opening of Ca2+ channels at intracellular locations is not as obvious in plants as it is in animals (Munnik and Nielsen 2011).
The first models described above assume that the plant gravisensing machinery is a force sensor. This assumption was initially based on the fact that the gravitropic response of plant organs appears to follow the reciprocity law (the curvatures resulting from transient gravistimulation vary linearly with the logarithm of the dose of gravistimulation, as discussed above). However, recent, clever experiments carried out by Hugo Chauvet and collaborators cast doubts on this assumption. Growing wheat seedlings in growth chambers carried by a rotating table, these authors were able to show that the developing coleoptiles respond to continuous gravistimulation within a large range of effective gravity forces (0.5xgearth to 2gearth) with similar angles of curvature. In other words, under their experimental conditions, the coleoptiles did not follow the reciprocity law. On the other hand, the curvature response developed by these coleoptiles followed the so-called sine rule, which postulates that the curvature response to gravistimulation is proportional to the sine of the angle of stimulation (the inclination of the plant). The authors interpreted their results by suggesting that the gravity sensing machinery in plants functions as an inclination/position sensor rather than a force sensor (Fig. 5.4). The inclination sensor would be sensitive to the position of the bulk of amyloplasts within the statocytes, rather than responding to the force exerted by sedimenting amyloplasts (or the entire protoplast) on sensitive membranes. Under their model, plant organs subjected to transient gravistimulation (such as those subjected to a presentation-time assay) would follow the reciprocity law simply because these experiments involve transient stimuli that are sufficiently short to only allow incomplete repositioning of the amyloplasts within the statocytes under regular 1xgearth conditions. Consequently, under such conditions, higher doses of g would promote a faster sedimentation of the plastids to the bottom of the cells, allowing a stronger graviresponse. In experiments that involve continuous gravistimulation, amyloplasts are allowed to fully settle at the bottom of the statocytes, allowing for a full response to develop. Therefore, the presentation time experiment is a better setup to evaluate effectiveness of amyloplast sedimentation than it is to estimate organ gravisensitivity. Similar results and conclusions were obtained when these experiments were repeated with seedlings of a wide range of plant species, including representatives of Asterids, Rosids, and Commelids. Although roots were not tested in these experiments, the shoots of these diverse plant groups developed gravitropic responses that obeyed the sine law and were independent of gravity intensity. Therefore, the shoots of these diverse species may also use a mechanism of gravisensing that involves an inclination sensor system (Bérut et al. 2018; Chauvet et al. 2016).

Two models have been proposed to explain gravisensing by plant statocytes. In the inclination sensor model (panel a), the position of sedimenting amyloplasts within the statocytes determines the polarity of auxin transport. In the statocyte illustrated here, amyloplasts occupy a larger surface on the right side of the cell after reorientation (right drawing) relative to the vertically oriented cell (left). The area of peripheral ER covered by sedimenting amyloplasts is represented in yellow. This model predicts that the curvature response to gravistimulation will not depend upon the pressure level. It also predicts that very small levels of inclination can trigger a curvature response. In the pressure sensor model (panel b), the force exerted by sedimenting amyloplasts on sensitive membranes (red arrows) triggers the opening of mechanosensitive ion channels, activating a transduction pathway that leads to statocyte polarization. This model predicts that a curvature response to gravistimulation will depend upon the pressure level. In these drawings, the peripheral ER is represented in blue and yellow, whereas amyloplasts are represented by gray circles. N Nucleus
The ligand-receptor model discussed earlier in this section would function as an inclination sensor mechanism, as would other models that postulate functional interactions between the group of sedimented amyloplasts and the vesicle trafficking machinery that is critical for proper location of auxin transporters (Pouliquen et al. 2017).
It is interesting to note here that the inclination sensor hypothesis is, in fact, compatible with the ability of plant organs to respond to very low inclinations from the vertical while not overreacting to the vibrations created by wind, rain, or other temporary environmental perturbations (Pouliquen et al. 2017).
In summary, two main models have been suggested to explain gravity sensing by amyloplast sedimentation in the statocytes: (1) the force sensor model suggests that amyloplasts settling on side membranes of a statocyte, or the entire weight of the protoplast on its cell wall, may trigger a transduction pathway within these cells, possibly by promoting the opening of mechanosensitive ion channels that remain to be characterized, and (2) the recently proposed inclination sensor model, which postulates that the location of the amyloplast system within the statocytes, rather than the pressure exerted by individual amyloplasts or the overall protoplast, is the main response trigger. While most investigators have focused many years of research on attempting to resolve the molecular machinery that makes up a force sensor in plant statocytes, the clever experiments carried out by Moulia and collaborators suggest a distinct mechanism responding to organ inclination rather than force. Recognition of this possibility is reshaping our view of the process, and it is likely to catalyze new exciting research to identify the molecular mechanisms that contribute to gravisensing in plants.
In conclusion, we do not know the identity of the gravitropic receptors that function to activate the gravitropic signal transduction pathway in the statocytes in response to gravistimulation. However, we do know that this pathway leads to an asymmetric redistribution of auxin across the root tip, ultimately responsible for differential cell elongation between upper and lower flanks at a distal side of the elongation zone. We will now review some of the molecular mechanisms that lead to gravity-induced lateral polarization of the statocytes. However, before we do so, we will provide a brief description of the molecular mechanisms that govern auxin synthesis, transport and response in plants.
2.4 How Is Auxin Synthesized and Transported Within the Plant, and How Do Plant Cells Respond to It?
Auxin is a hormone that contributes to many facets of plant growth and development regulation as well as responses to the environment. Before discussing its contribution to gravitropism, we will first describe some of the molecular mechanisms that specifically contribute to auxin transport and response.
Auxin is mostly synthesized in young shoot tissues, using a combination of tryptophan-dependent and tryptophan-independent pathways (Zhang and Peer 2017). From there, it is transported to other regions of the plant where it regulates a variety of cellular processes including cell division, elongation, differentiation and death. It is also transported through the vasculature into the root tip, where it adds to a pool of locally synthesized auxin and accumulates in the quiescent center and upper layers of the columella cells. From this maximum center at the root tip, auxin is redistributed to more peripheral tissues and then transported back toward the root meristem and elongation zone, where it regulates cell division, inhibits elongation, and modulates cell differentiation (Brumos et al. 2018; Ding and Friml 2010; Mironova et al. 2010). Auxin transport is a highly regulated process that follows cell files. In each transporting cell within a file, auxin import facilitators of the AUX1/LAX family contribute to auxin uptake from the apoplast, helped along by free auxin diffusion across the plasma membrane. The latter process is possible because indole-3-acetic acid (IAA, the most common natural auxin in plants) is a weak acid, and the acidic condition of the apoplast (pH~5.6) facilitates its protonation, a process that is needed for free diffusion across the plasma membrane. On the other hand, plant cell cytoplasm has a neutral pH, resulting in the ionization of almost all auxin molecules within the cell. Ionized auxin cannot cross membranes. Therefore, auxin efflux facilitators of the PIN family, along with P-glycoprotein-type transporters (such as AtPGP1 and AtPGP19 in Arabidopsis; (Geisler et al. 2005; Noh et al. 2001)), are needed to export it away from the cell interior. Interestingly, the PIN proteins are often distributed asymmetrically within the plasma membrane, accumulating at one side of the transporting cells. Therefore, the polarity of auxin transport through cell files is dictated by the polar localization of the PIN auxin efflux facilitators in transporting cells (Fig. 5.5).

In roots, gravistimulation promotes the formation of a lateral auxin gradient across the cap, which is responsible for a tip curvature upon transmission to the elongation zone. Panel (a) shows the flow of auxin in vertical (left) and gravistimulated seedlings (middle and right drawings), whereas panel (b) shows amyloplast sedimentation and PIN protein relocalization within the statocytes at different phases of a graviresponse. The reverse fountain model of auxin transport is shown in the left drawing of panel A. Mainly synthesized in young shoot tissues, auxin is transported through the vasculature into the root tip where it accumulates at the quiescent center and upper tiers of the columella region of the cap. It is then redistributed laterally to more peripheral tissues of the lateral cap, using the PIN3 and PIN7 auxin efflux carriers. From the lateral cap, auxin is transported back toward the elongation zone through lateral cap and epidermal cells. Auxin transport through cell files involves AUX1 influx carriers and polarized PIN2 efflux transporters. PIN2 is also expressed in the cortical cells of the root distal elongation zone, where it localizes to the inner and rootward membranes, contributing to a reflux of peripheral auxin toward the provasculature. Upon gravistimulation (middle panel), amyloplast sedimentation within the statocytes triggers a gravity signal transduction pathway that leads to a polarization of the PIN3 and PIN7 proteins to the lower side of the cells. Consequently, auxin is transported preferentially to the lower flank of the cap, and the resulting gradient is transferred toward the elongation zone where it triggers a downward curvature. Increased levels of auxin on the lower side of the root also result in increased retention of the PIN2 auxin efflux carrier within the plasma membrane relative to cells on the topside, contributing to auxin gradient maintenance. When the graviresponding root tip reaches an angle of 50° from the vertical (right panel), amyloplasts have settled back to their original position within the statocytes, and PIN3 and PIN7 redistribute more or less symmetrically on all sides of the statocytes. Consequently, the lateral auxin gradient dissipates, and the remaining curvature toward the vertical proceeds in the absence of an auxin gradient. In these drawings, the direction of auxin transport is represented by red arrows, whose widths represent auxin flow intensities. C represents a cortical cell whereas E represents the epidermis. Other symbols are defined in the legend provided underneath the figure. This figure is a modification from Figures 2 and 3 in Su et al. (2017)
Auxin influx and efflux carriers are encoded by rather large gene families in plants, with each gene within a family displaying specific expression patterns and protein localizations. In Arabidopsis roots, the PIN1, PIN3, PIN4, and PIN7 proteins contribute to auxin transport toward the tip (“rootward” transport) through provasculature cell files. By contrast, the PIN2 protein contributes to auxin transport from cap to elongation zone (“shootward” transport) within peripheral tissues. The PIN2 protein localizes on the shootward-facing side of the lateral cap and epidermal cells of the elongation zone, moving auxin in a shootward direction (away from the root tip toward the shoot). Additionally, PIN2 is also expressed in the cortical cells of the elongation zone, where it localizes on the inner and rootward-facing side of the cells, thereby refluxing auxin from the shootward peripheral stream back to the central rootward flow (directed toward the root tip; Fig. 5.5) (Adamowski and Friml 2015).
The molecular mechanisms that contribute to cellular responses to auxin have been well investigated, involving a combination of gene expression-dependent and gene expression-independent processes. A fast response to auxin involves the direct activation of ion channels (Fig. 5.6a). In roots, auxin promotes a depolarization of the responding cell, associated with an alkalinization of the apoplast, which leads to increased cell wall rigidity, decreased cell elongation and slower root growth (Cosgrove 2000; Monshausen et al. 2011). This process is dependent upon auxin uptake by the cell, and its use by a cytoplasmic pool of an SCF-based ubiquitylation complex that contains the TIR1/AFB auxin receptor. This process leads to the activation of a cyclic nucleotide-gated ion channel (CNGC14). Opening of this channel results in increased levels of cytoplasmic Ca2+, leading to the activation of a plasma membrane H+/OH− conductance, with concomitant alkalinization of the apoplast (Monshausen et al. 2011; Mullen et al. 1998). This alkalinization leads to increased cell wall rigidity, decreased cell elongation, and lower root growth rate (Cosgrove 2000; Dindas et al. 2018; Monshausen et al. 2011).

Plant cellular responses to auxin involve expression-dependent and expression-independent processes. Panel (a) represents an Arabidopsis root cell responding to auxin (orange circle). Auxin penetrates the cell by free diffusion through the plasma membrane or through AUX1-like transporters. Once in the cytoplasm, auxin activates the opening of a cyclic nucleotide-gated ion channel (CNGC14), allowing a pulse in cytoplasmic Ca2+. This triggers a pathway that leads to activation of a H+/OH- antiporter, responsible for alkalinization of the apoplast and inhibition of cell expansion. Some auxin molecules can also enter the nucleus, where they favor the formation of a complex between the SCFTIR1 ubiquitylation enzyme and Aux/IAA target proteins. Without auxin, Aux/IAA interacts with auxin response transcription factors (ARFs), inhibiting their function. In the presence of auxin, the Aux/IAA proteins are ubiquitylated by SCFTIR1 and targeted to the proteasome where they get degraded. As a consequence, the ARF transcription factors are free to modulate the expression of multiple auxin response genes. Panel (b) shows transgenic Brachypodium distachyon roots expressing a DR5p:RFP auxin-activity reporter (left) or a dII-Venus auxin-level reporter (right). In both cases, the top picture shows the fluorescent signals displayed by the root, whereas the bottom picture is an overlay of the fluorescent signals with a bright-field image of the root. The red-fluorescent protein (RFP) reporter shown on the left is expressed under the control of the synthetic DR5 promoter, which carries several copies of an auxin-responsive transcriptional enhancer targeted by auxin response factors. Its transcription is modulated by the expression-dependent pathway described under panel A. The dII-Venus reporter (right), on the other hand, is expressed ubiquitously in the plant. It produces a dII-Venus fluorescent protein carrying a dII motif that allows its recognition by SCFTIR1 in the presence of auxin. Consequently, increased levels of auxin result in increased polyubiquitylation of dII-Venus, a modification that targets it to degradation by the proteasome. Therefore, increased levels of auxin within the cell lead to lower reporter signals. Please note that DR5:RFP expression is highest around the quiescent center and columella region of the cap, whereas the dII-Venus reporter is mostly visible in peripheral tissues. These two transgenic lines were provided by Devin O’Connor, University of Cambridge, UK
In gene expression-dependent responses, intracellular auxin interacts with the same SCFTIR1/AFB complex, allowing its interaction with and ubiquitylation of a group of AUX/IAA proteins, targeting them to degradation by the proteasome (Fig. 5.6a). Because these AUX/IAA proteins serve as inhibitors of AUXIN RESPONSE FACTORS (transcription factors that regulate the expression of specific subgroups of target genes), auxin allows ARF-dependent regulation of target gene expression (Dharmasiri et al. 2005; Kepinski and Leyser 2005; Lau et al. 2011). Participation of this system in root gravitropism is evidenced by the altered gravitropism phenotype displayed by Arabidopsis tir1 mutants (Dharmasiri et al. 2005; Kepinski and Leyser 2005; Lau et al. 2011).
Plants contain many genes that encode AUX/IAA (29 in Arabidopsis, for instance) and ARF proteins (23 in Arabidopsis). Each cell type expresses specific subsets of AUX/IAA and ARF genes. Furthermore, different AUX/IAA proteins display distinct binding preferences for different groups of ARF proteins. This combinatorial system leads to cell-specific regulation of gene expression in response to auxin. Some of the auxin-responsive genes expressed in the epidermis were shown to encode proteins that contribute to cell wall remodeling (Swarup et al. 2005). Others were shown to encode proteins that modulate auxin conjugation, thereby contributing to feedback regulation of the pathway (Zhang and Peer 2017). The SMALL AUXIN UP RNA (SAUR) genes, which are among the fastest auxin responders, were shown to encode small proteins that inhibit PP2C.D phosphatases, thereby activating plasma membrane proton ATPases and modulating cell expansion (Ren and Gray 2015).
Interestingly, the discovery of those auxin response pathways in plants led to the development of two complementary transgenic auxin reporter systems that allowed detection of auxin gradients generated across the root tip upon gravistimulation (Fig. 5.6b). The first reporter is a fusion between a synthetic promoter (DR5) that carries several copies of an auxin response element (enhancer elements recognized by ARF transcription factors) and the open reading frame of a gene that encodes a reporter protein (fluorescent protein such as GFP or VENUS or β-glucuronidase, a bacterial enzyme that converts a colorless soluble compound named X-glu into a blue insoluble precipitate that stains the cells expressing it). When plants are transformed with such a reporter construct, the levels of reporter transcripts (hence, the level of reporter protein) increase when auxin levels increase or when expressing cells become more sensitive to auxin.
The second type of auxin reporter involves a sensor protein (typically the fluorescent protein VENUS) engineered to carry a motif (named dII) that is recognized by the SCFTIR1/AFB ubiquitylation complex in the presence of auxin. This transgenic reporter is expressed in plants under the control of a ubiquitous promoter. When expressing cells are exposed to increased levels of auxin, the dII-VENUS protein is targeted by the SCFTIR1/AFB complex, which ubiquitylates it and targets it to degradation by the proteasome. Therefore, with the DR5 reporter system, increased auxin levels (or auxin sensitivity) lead to increased reporter expression, whereas the dII-VENUS reporter is degraded in the presence of auxin. The latter construct is a more direct sensor of auxin levels than the former.
When transgenic plants expressing a DR5-GFP reporter are subjected to gravistimulation (reorientation within the gravity field), their roots quickly develop a lateral gradient of reporter expression across the root cap, with increased expression on the lower flank. This gradient then progresses along the root tip toward the elongation zone. When such gravistimulated plants express dII-VENUS instead of DR5-GFP, the fluorescence signal decreases in cells at the bottom flank of the cap. The corresponding fluorescent-signal gradient also propagates toward the elongation zone over time, again reflecting formation of a lateral auxin gradient across the root cap upon gravistimulation (Fig. 5.6). In the next few sections, we will summarize the molecular mechanisms that contribute to gravity-induced formation of lateral auxin gradients across gravistimulated root tips.
2.5 Gravistimulation Promotes a Relocalization of Auxin Efflux Facilitators in the Statocytes
Critical to gravitropic regulation is a root cap-specific lateral auxin redistribution stream that connects the auxin maximum at the center of the root tip (quiescent center and upper columella cells) to its peripheral shootward stream. The PIN3 and PIN7 proteins, which are expressed in overlapping domains within the columella region of the root cap, play key roles in this lateral redistribution. In fact, AtPIN3 is expressed in the upper two tiers of columella cells, whereas AtPIN7 is expressed in tiers 2 and 3 (Fig. 5.1) (Friml et al. 2002; Kleine-Vehn et al. 2010; Wang et al. 2015). The PIN3 and PIN7 proteins are distributed uniformly within the plasma membrane on all sides of the statocytes in vertically oriented roots, allowing symmetrical auxin redistribution to the lateral cap cells.
Upon plant reorientation within the gravity field, the PIN3 and PIN7 proteins quickly relocalize to the lower side of the statocytes, thereby generating a downward stream of auxin across the cap, with accumulation in its lower side (Fig. 5.5) (Friml et al. 2002; Kleine-Vehn et al. 2010). This process appears to be mediated by a transcytotic mechanism that involves endocytosis of PIN3/7-carrying vesicles from the plasma membrane and their recycling toward the lower membrane of the cells. It is regulated by PIN protein phosphorylation and is dependent upon several factors that are known to contribute to vesicle trafficking within plant cells such as small GTPases of the ADP-ribosylation factor (ADP-RF) type, associated with GDP/GTP exchange factors (GEFs) of the GNOM type (Ganguly et al. 2012; Kleine-Vehn et al. 2010). Brefeldin A, a pharmacological agent of fungal origin that inhibits the GNOM-dependent step of vesicular trafficking, also affects gravitropism, supporting a role for vesicular trafficking in this response.
The regulatory molecules that contribute to the modulation of PIN3/7 trafficking toward the lower membrane upon gravistimulation remain poorly characterized. In fact, genetic approaches have been carried out to identify some of the contributing factors. The corresponding screens involved seeking mutations that specifically affect gravitropism while having no effects on phototropism and/or organs growth responses to exogenous auxin or auxin transport inhibitors. The rationale for such screening criteria is as follows. Because both gravitropism and phototropism involve the formation of an auxin gradient across stimulated organs, mutations that affect both gravi- and phototropism are more likely to affect auxin transport and/or response. Mutations that specifically affect gravitropism, on the other hand, are more likely to affect early (and specific) steps of gravity sensing and/or signal transduction. On the other hand, mutations that affect both gravitropism and organs’ growth responses to exogenous auxin and/or polar auxin transport inhibitors are more likely to affect the later phases of auxin transport and/or curvature response.
When mutations fulfilling the criteria defined above are found, contribution of the corresponding genes to early steps of gravity sensing and/or signal transduction in the statocytes can be verified by demonstrating a lack of PIN3/7 relocalization in the statocytes upon gravistimulation and an absence of lateral auxin gradient across gravistimulated root tips in mutant seedlings.
Using this approach, researchers were able to isolate several mutations that affect gravity sensing and/or early steps of gravity signal transduction in the root statocytes. The first Arabidopsis mutations found to alter at the same time root gravitropism, PIN3 relocalization and lateral auxin gradient formation upon gravistimulation, affected two genes that encode paralogous proteins named ALTERED RESPONSE TO GRAVITY 1 (ARG1) and ARG1-LIKE2 (ARL2). These mutations affected both root and hypocotyl gravitropism without altering phototropism. arg1 and arl2 mutant seedlings displayed wild-type root growth responses to auxin and auxin transport inhibitors, and their statocytes contained starch-filled amyloplasts that sedimented like wild type (Boonsirichai et al. 2003; Harrison and Masson 2008). The ARG1 and ARL2 genes were found to encode peripheral membrane proteins that associate with the plasma membrane, ER, Golgi and endosome, thereby probably regulating vesicular trafficking, a process needed for PIN3/7 protein relocalization in the statocytes upon gravistimulation (Boonsirichai et al. 2003; Harrison and Masson 2008).
One interesting feature of the arg1 and arl2 mutant seedlings is that they display only partial defects in root and hypocotyl gravitropism. Therefore, arg1 (or arl2) plants can be used as sensitized lines to isolate novel mutations that either enhance or suppress their gravitropic responses. Such genetic modifiers of arg1 (or arl2) would likely also contribute to early steps of gravity sensing and/or signal transduction. A secondary screen for genetic enhancers of arg1 was carried out, identifying plants with enhanced gravitropism defects relative to arg1. modifier of arg1-1 (mar1-1) carried a missense mutation in TOC75, a gene that encodes the channel component of plastidic TRANSLOCON ON THE OUTER CHLOROPLAST MEMBRANE (TOC) complex, which mediates the import of cytoplasmic proteins through the outer membrane of plastids. A second modifier of arg1, named mar2-1, was also isolated, carrying a missense mutation in TOC132, which encodes another component of the same TOC complex (Stanga et al. 2009). These two mar mutations did not obliterate TOC’s function as protein importer. In fact, mutant root cap amyloplasts accumulated starch like wild type, and they sedimented at wild-type rates upon gravistimulation. These data suggested a role for amyloplasts in gravity signal transduction that goes beyond their ability to sediment as statoliths (Stanga et al. 2009; Strohm et al. 2014). A differential proteomic analysis comparing wild-type and toc132 mutant roots identified candidate gravity signal transducers, whose functions remain to be characterized (Strohm et al. 2014).
Genetic investigations of gravity sensing and signal transduction have not been limited to Arabidopsis thaliana. In fact, work done in the legume model Medicago truncatula uncovered a mutation that leads to upward-oriented roots that grow out of the soil. Named negative gravitropic response of roots (ngr), this mutation was found to affect a gene that encodes a plant-specific protein of unknown function (Ge and Chen 2016). Interestingly, this protein shares similarity with LAZY1, a protein known to contribute to gravitropism in rice, maize, and Arabidopsis (Dong et al. 2013; Li et al. 2007; Yoshihara et al. 2013). In Arabidopsis, six genes with spatially distinct expression patterns encode LAZY1-like proteins. Phenotypic analysis of higher-order mutants revealed key contributions played by distinct members of this gene family to root and shoot branch angles as well as seedling primary organs gravitropism (Yoshihara and Spalding 2017; Taniguchi et al. 2017). Importantly, the reversed gravitropic response displayed by some higher-order Atlazy mutants relative to wild type was associated with reversed asymmetric distribution of PIN3 in gravistimulated statocytes and a reverse lateral gradient of auxin (Taniguchi et al. 2017; Yoshihara and Spalding 2017). Starch content and amyloplast sedimentation were not affected in analyzed mutants, indicating that the NGR/LAZY genes contribute to a step of gravity sensing and/or signal transduction that follows amyloplast sedimentation. In one of the triple mutants, the reversed gravitropism phenotype could be rescued by expression of a wild-type LAZY1 transgene specifically in the statocytes, demonstrating a statocyte-specific contribution of the gene to gravitropism. Taken together, these exciting results position the LAZY/NGR proteins at an important step of the transduction pathway that is needed for proper interpretation of the gravity vector by the statocytes (Ge and Chen 2016).
In conclusion, genetic investigations of root gravitropism have identified a number of loci that contribute to the transduction of information provided by amyloplast settling into a transcytotic process that results in a relocalization of auxin efflux facilitators PIN3 and PIN7 to the lower membranes of the statocytes, thereby leading to the formation of a lateral auxin gradient across the cap, and ultimately a curvature response at the DEZ. It should however be cautioned that the pathway may be a little more complicated. Indeed, the pin3 and pin7 knockout mutants, as well as the pin3 pin7 double mutants, still display significant root curvature responses to gravistimulation. This implies that other unknown auxin transporters may also contribute to gradient formation across the root tip and to gravitropism.
Another important point that should be raised here is that auxin transport across the root cap may be facilitated by other physiological changes that occur in the root cap statocytes in response to gravistimulation. For instance, gravistimulated statocytes have been shown to undergo a rapid alkalinization of their cytoplasm, accompanied by an acidification of the apoplast (Fasano et al. 2001). This process is ARG1-dependent, and it may result from an activation of plasma membrane proton pumps and/or vacuolar ATPases. It is critical for full responsiveness to gravity, possibly by decreasing the fraction of ionized auxin in the apoplast and facilitating its mobility through membranes and/or transporters (Fasano et al. 2001).
3 Auxin Gradient Propagation from the Root Cap to the DEZ
The mechanisms discussed above allow establishment of a lateral auxin gradient across the root cap upon gravistimulation. Yet, the initial phases of curvature response occur at the distal side of the EZ. Therefore, auxin transport machinery has to move this auxin gradient from root tip to EZ, maintaining it along the road. Which mechanisms contribute to this complex process?
The lateral root cap, epidermis and cortex all contribute to the shootward flow of auxin in roots. In Arabidopsis, these cells take up auxin using mostly the AUX1 influx carrier, whereas PIN2, along with members of the p-glycoprotein family, contribute to its export from the transporting cells. The polar localization of PIN2 within these transporting cells dictates shootward transport in lateral cap and epidermal cells and rootward reflux in the cortical cells at the distal side of the elongation zone (Blilou et al. 2005) (Fig. 5.5). Both flows of auxin (shootward in epidermal and lateral cap cells and rootward in cortical cells) are critical for an efficient gravitropic response (Blilou et al. 2005). Interestingly, reversible phosphorylation of PIN2 plays a key role in its localization within transporting cells, with the serine/threonine protein kinases PINOID (PID) and PID-like WAG1 and WAG2 kinases contributing to its phosphorylation, whereas type-IIA protein phosphatase complexes (PP2A) contribute to its dephosphorylation. When phosphorylated, PIN2 localizes at the rootward side of the transporting cells, whereas it associates with the shootward side of the cell when dephosphorylated (Barbosa et al. 2014; Dhonukshe et al. 2010).
The shootward transport of auxin from root cap to elongation zone is also subject to feedback regulatory mechanisms that assure lateral gradient maintenance during its transfer toward the EZ. Indeed, as emphasized earlier in this chapter, auxin has been shown to promote the maintenance of PIN proteins within the plasma membrane of transporting cells (Abas et al. 2006). As a consequence, auxin accumulation on the bottom side of a gravistimulated root tip leads to increased auxin transport potential on that side and lower transport potential on the upper side. Therefore, the gravity-induced lateral auxin gradient that was generated across the root cap becomes increasingly pronounced as it progresses toward the elongation zone (Abas et al. 2006; Li and Xue 2007; Lin et al. 2012). This process is exacerbated by increased auxin-dependent production of small signaling peptides GOLVEN1 (GLV1) and GLV2 by cells on the lower side of the stimulated roots, triggering a response pathway that also favors PIN2 association with the plasma membrane (Whitford et al. 2012).
On the other hand, the increased apoplast alkalinization that occurs on the lower side of a graviresponding root as a consequence of increased auxin levels is responsible for decreasing the fraction of protonated IAA molecules in the apoplast, thereby decreasing the rate of free diffusion through the plasma membrane of transporting cells. Consequently, the auxin influx carrier AUX1 is needed for adequate shootward auxin transport and root gravitropism (Dharmasiri et al. 2006; Monshausen et al. 2011). In fact, the AUX1 gene is expressed broadly in the root tip, including the provasculature, root cap, and epidermal cells. However, restricting its expression to the lateral cap and epidermal cells of the root meristem and EZ is sufficient to rescue the altered root gravitropism phenotype displayed by aux1 mutant seedlings (Dharmasiri et al. 2006; Swarup et al. 2005). The latter observation is important because it demonstrates that the contribution of AUX1 to root gravitropism requires its expression only within the peripheral tissues of the root tip, where shootward auxin transport occurs (Dharmasiri et al. 2006; Swarup et al. 2005).
From the preceding discussion, it appears that Arabidopsis PIN2 plays a key role in root gravitropism, transporting the gravity-induced auxin gradient from the root cap toward the elongation zone, where it regulates differential cellular elongation and curvature. It is therefore quite surprising that Arabidopsis pin2 mutant roots retain some gravitropic capability (Baldwin et al. 2013). This implies that PIN2 function may be redundant with other transporters. In agreement with this contention, P-glycoprotein-type transporters, which use ATP hydrolysis to carry specific molecules through membranes (auxin in this case), may fulfill this redundant auxin transport function. Indeed, Arabidopsis AtPGP1 and AtPGP19 genes are also expressed in the root EZ, and functional studies in heterologous systems (plant protoplasts, yeast and mammalian cells) have demonstrated their ability to transport auxin (Geisler et al. 2005; Yang and Murphy 2009). Furthermore, pgp19 single mutants and pgp1 pgp19 double mutants exhibited reduced basipetal auxin transport (Lewis et al. 2007). Surprisingly, these mutants displayed an enhancement of gravitropism and phototropism. This phenotype is, in fact, a consequence of these genes being expressed more proximally (shootward) than PIN2 in the root tip. Hence, pgp1 pgp19 mutant roots develop a stronger auxin gradient across the DEZ relative to wild type, allowing for enhancement of the curvature response (Noh et al. 2001; Rojas-Pierce et al. 2007).
4 Root Curvature Response to Gravistimulation
4.1 The Gravitropic Curvature Involves Differential Cell Elongation Between Opposite Root Tip Flanks
Upon transmission to the EZ, the gravity-induced lateral auxin gradient promotes differential cell elongation between upper and lower flanks of the gravistimulated root, leading to initiation of a curvature within 10–15 min of reorientation. Which mechanisms contribute to gravitropic curvature development?
As mentioned above, gravistimulation promotes a lateral movement of auxin across the root cap, with accumulation at the bottom half. This gradient is then transmitted to the EZ. There, higher auxin level on the lower side of the gravistimulated root leads to an inhibition of cell elongation, whereas lower auxin level on the upper side leads to increased elongation. As a consequence, a downward curvature develops.
Increased auxin levels on the upper half of gravistimulated roots leads to the activation of a plasma membrane H+/OH− conductance, which results in alkalinization of the apoplast, as described in Sect. 5.2 (D part) of this chapter (Monshausen et al. 2011; Mullen et al. 1998). The resulting increase in cell wall rigidity leads to decreased rate of cellular elongation on the lower side of the root. The upper flank, on the other hand, is exposed to lower auxin levels, resulting in increased cell wall acidity. Lower wall pH is known to promote the breakage of intermolecular cross-links between wall polymers by expansins and xyloglucan endotransglucosylases/hydrolases (XTHs), favoring cellular elongation. Consequently, increased cell elongation on the topside and decreased expansion at the bottom results in a downward curvature (Cosgrove 2000; Monshausen et al. 2011).
In addition to these direct effects of auxin via its SCFTIR1/AFB receptor on cellular expansion, other signaling molecules have also been implicated in the curvature response to gravistimulation. For instance, nitric oxide (NO) was found to accumulate on the lower side of gravistimulated roots in response to auxin accumulation, where it inhibits auxin transport and modulates auxin signaling through S-nitrosylation of TIR1 (Terrile et al. 2012). Similarly, reactive oxygen species (ROS) have been shown to accumulate at the bottom side of roots in an auxin-dependent manner, where they contribute to the regulation of gravicurvature (Krieger et al. 2016).
The epidermis is believed to be the main driver of root tip curvature. Indeed, expressing an auxin response repressor (axr3-1) in epidermal cells of the elongation zone is sufficient to obliterate the gravitropic response, whereas expressing it in different cell types within the elongation zone has little impact (Swarup et al. 2005).
4.2 How Does a Root Know It Has Curved Enough?
As discussed earlier in this chapter, the availability of in situ auxin sensors (DR5-GFP; dII-VENUS) and fluorescent protein-PIN fusion reporters in Arabidopsis has allowed careful spatiotemporal investigations of PIN3/7 relocalization to the bottom membrane of statocytes and auxin gradient formation across the root tip upon gravistimulation, this in relation with the timing of amyloplast sedimentation in the statocytes and root curvature response. Such experiments have demonstrated that the auxin gradient generated across the root disappears when the tip reaches an approximate angle of 50° from vertical. At this point, the statoliths have returned to the distal side of the statocytes as a consequence of cellular morphology, and the PIN3/7 proteins have returned to a symmetrical distribution on all sides of the statocytes (Band et al. 2012). This suggests that auxin asymmetry during a gravitropic response is susceptible to a tipping-point mechanism that is triggered when the tip reaches a 50° angle from vertical. At that point, the curvature has to proceed in the absence of a lateral auxin gradient, until the tip reaches the vertical. Unfortunately, the molecular mechanisms that contribute to this second auxin gradient-independent phase of gravicurvature and its termination when the tip reaches the vertical remain unexplained (Band et al. 2012).
5 The Gravity Set Point Angle of Lateral Roots Differs from That of Primary Roots
While the primary roots of most plants tend to grow orthogravitropically (parallel to gravity), the lateral roots that develop from pericycle initials within the primary root will tend to grow either diagravitropically (horizontally) or plagiogravitropically (obliquely), allowing for better soil exploration for water and nutrients. The environmental conditions can alter the angle of lateral root growth from the vertical, favoring either steeper angles (allow better adaptation to drought, for instance) or shallower growth (when there is a need for better exploitation of surface resources such as phosphate) (Bai et al. 2013). What do we know of the mechanisms that allow lateral roots to grow at a different GSA from the vertical?
In Arabidopsis, lateral roots emerge perpendicularly from the primary root and then progressively acquire plagiogravitropism as starch accumulates in the statocytes, and the EZ becomes established (Guyomarch et al. 2012; Kiss et al. 2002; Rosquete et al. 2013). These roots curve toward an initial GSA, which is rather shallow, and then straighten up and grow along this vector for some time. Subsequently, these laterals may start curving again, leaning toward positive orthogravitropism (vertically downward). The initial plagiogravitropic phase of lateral root growth may result from an auxin-dependent antigravitropic offset mechanism that opposes gravitropism to regulate the distribution of auxin levels and response between opposing sides (Roychoudhry et al. 2013).
Regulation of auxin transport in lateral root statocytes seems responsible for the regulation of positive orthogravitropism (Fig. 5.7). Early after emergence, only PIN3 is expressed in the columella cells of the Arabidopsis lateral root cap. This PIN3 protein is quickly redistributed asymmetrically toward the bottom side of the statocytes, yielding a lateral gradient of auxin that triggers downward curvature. When the young lateral root reaches its first GSA plateau, PIN3 expression decreases, and PIN4 and PIN7 are activated to very low expression levels. At this stage, the overall level of PIN expression in the statocytes is low, and the PIN proteins are symmetrically distributed in the statocytes, allowing the laterals to continue growing straight along the GSA. Subsequently, the PIN4 and PIN7 genes increase their expression, and the corresponding proteins redistribute to the bottom side of the statocytes, again creating a lateral auxin gradient that is responsible for a new phase of downward curvature (Roychoudhry et al. 2013). It is tempting to speculate that the regulatory system discussed above may constitute a target for developmental and environmental signals to dictate whether a root system will be radially expanded or organized axially (Rosquete et al. 2013). Interestingly, the LAZY proteins described in Sect. 3.5. of this chapter may play an important role in this process (Taniguchi et al. 2017).

Lateral root gravity set point angle (GSA) correlates with decreased auxin flow at the tip. In gravistimulated primary Arabidopsis thaliana roots, polar localization of PIN3 and PIN7 auxin efflux facilitators in the statocytes leads to the formation of a strong lateral auxin gradient across the cap, which is responsible for root tip curvature upon transmission toward the distal side of the elongation zone. These primary roots will tend to grow vertically downward (orthogravitropism). In young lateral roots (center drawing), PIN3 is expressed at lower levels in the root cap, decreasing over time. Consequently, the lateral gradient of auxin that develops across the cap remains mild. This leads to shallower GSA relative to primary roots (plagiogravitropism). In older lateral roots, PIN3 expression ceases and is replaced by stronger expression of the PIN4 and PIN7 genes in the statocytes, leading to a stronger lateral auxin gradient across the tip relative to your laterals. As a consequence, the older lateral root will curve back to a steeper angle. (Roychoudhry et al. 2013)
6 How Do Shoots Respond to a Reorientation Relative to Gravity?
In dicots, shoot and hypocotyl gravitropism also involves a differential elongation between top and bottom sides, typically leading to upward curvatures. Unlike roots, stems do not show evidence of a physical separation between sites of gravity sensing and curvature response. Instead, the curvature occurs along the entire length of the EZ, and gravity-sensing statocytes occupy the entire region, forming the endodermal layer or starch sheath parenchyma that surrounds the vasculature. This implies that the curvature response to gravistimulation will typically follow more complex kinetics in shoots. As an example, Arabidopsis inflorescence stems are characterized by different rates of elongation along their length. Upon gravistimulation, the apical region will initially curve faster than the basal (rootward) region. When this apical region reaches the vertical, the basal region is still curving, implying that the tip will soon overshoot the vertical. As a consequence, the apical segment will sense an opposite gravistimulus and will start curving in the opposite direction. This back-and-forth oscillation may occur several times before the stem eventually reaches its final posture. This complex behavior is, in fact, compatible with the existence of gravity-sensing cells along the entire length of the EZ in shoots, along with differential rates of cellular elongation between upper and lower segments, and an added mechanism of autostraightening (Bastien et al. 2013; Fukaki et al. 1996; Morita 2010).
Gravity sensing by shoot endodermal statocytes also involves sedimenting amyloplasts. However, the endodermal cells of shoots differ from the columella cells of the root cap by the existence of a large central vacuole that pushes all organelles to the cell periphery. Consequently, amyloplasts have to traverse transvacuolar strands during their sedimentation. This implies that vacuolar integrity and biogenesis are critical for normal gravitropism. We know this because many of the mutations that have been identified in Arabidopsis for their impact on shoot gravitropism were shown to affect either endodermal cell fate specification (sgr1/scr and sgr7/shr, for instance) or vacuolar biogenesis and function (sgr2, sgr3, sgr4, sgr8, for instance).
The SHOOT GRAVITROPIC RESPONSE 7/SHORTROOT (SGR7/SHR) and SHOOT GRAVITROPISM 1/SCARECROW (SGR1/SCR) transcription factors are critical for endodermis specification. Indeed, sgr1/scr and sgr7/shr mutant plants lack a fully differentiated endodermal layer with sedimenting amyloplasts. These developmental phenotypes are accompanied by an inability for mutant shoots and hypocotyls (but not roots) to respond to gravistimulation (Fukaki et al. 1998). The sgr2, sgr3, sgr4 and sgr8 mutations, on the other hand, display altered vacuolar phenotypes. In fact, the proteins encoded by the SGR3, SGR4 and SGR8 genes appear to contribute to vesicular trafficking between Golgi and vacuole, providing an environment that is favorable to amyloplast sedimentation upon gravistimulation (Silady et al. 2007; Yano et al. 2003; Zheng et al. 1999). SGR2, on the other hand, encodes a putative phospholipase that also localizes to vacuolar membranes, possibly modifying their composition and their biophysical properties, thereby interfering with amyloplast sedimentation (Kato et al. 2002; Morita et al. 2002).
That the gravitropic defect associated with these mutations is a consequence of their negative impact on amyloplast sedimentation in endodermal statocytes was supported by centrifugation experiments, which demonstrated a concomitant rescue by higher g forces (provided by centrifugation) of amyloplast sedimentation and gravitropism for sgr2, sgr9 and pgm (Toyota et al. 2013). The authors summarized the results of their experiments by indicting that “Arabidopsis shoots have a gravisensing mechanism that linearly converts the number of amyloplasts that settle to the ‘bottom’ of the cell into gravitropic signals” (Toyota et al. 2013).
So, amyloplast sedimentation within the endodermal statocytes of shoots leads to the activation of a gravity signal transduction pathway that promotes an upward curvature. What are the mechanisms that contribute to this gravity transduction pathway? In fact, as already discussed for roots, the mechanisms that transduce the information derived from amyloplast sedimentation into a biochemical signal that is responsible for the curvature remain poorly understood. However, we do know that this pathway leads to the development of a lateral auxin gradient across the stem, with auxin accumulation on the lower flank. As for roots, we know this because auxin-level or auxin-activity reporters (such as DR5-GFP) demonstrated asymmetric activation on opposite flanks of the shoot upon gravistimulation, consistent with increased auxin accumulation on the lower side. Because auxin promotes cell elongation in shoots, this gravity-induced auxin gradient leads to an upward curvature.
Research involving Arabidopsis thaliana hypocotyls and inflorescence stems led to a better characterization of the molecular mechanisms that control establishment of this auxin gradient across gravistimulated shoots (Fig. 5.8). Indeed, seedling reorientation within the gravity field was shown to promote a relocalization of the PIN3 auxin efflux facilitator to the lower membrane of the statocytes, with accumulation in the inner membrane of upper-half endodermal cells and outer membrane of lower half cells. This repolarization of PIN3 is consistent with a lateral downward transport of auxin from the upper to lower flanks of the stem, leading to an upward curvature (Fig. 5.8). As for root statocytes, gravity-induced PIN3 polarization in the endodermal cells requires a GNOM-dependent endocytotic recycling pathway which is modulated by PINOID-dependent protein phosphorylation (Rakusová et al. 2011).

PIN3 polarization in endodermal cells surrounding the vasculature contributes to auxin gradient formation and gravitropism in hypocotyls. In vertically oriented hypocotyls, the PIN3 protein is localized symmetrically at the plasma membrane of endodermal cells surrounding the vasculature. As a consequence, auxin is distributed equally on all sides of the hypocotyl, and the organ can grow straight up (left). Upon seedling reorientation within the gravity field, the PIN3 protein relocalizes to the lower membrane of endodermal statocytes, leading to lateral auxin gradient formation and upward curvature (2 h). Over time (18 h), auxin accumulation on the lower half of the hypocotyl leads to an inner lateralization of PIN3 in the lower endodermal cells, resulting in a reflux of excessive auxin from the lower half into the vasculature. As a consequence, the gradient dissipates and the differential growth responsible for increasing curvature ends. (Rakusová et al. 2016)
In experiments carried out with Arabidopsis hypocotyls, the gravitropic curvature initiated within 2 h, and it proceeded quickly during the initial phases of the response. However, after approximately 18 h, the rate of curvature diminished to eventually taper off. Interestingly, the termination of this curvature followed a second change in PIN3 protein distribution within endodermal cells at the lower side of the responding hypocotyl, with relocalization from outer to inner membrane (Fig. 5.8). Amazingly, this inner lateralization of the PIN3 protein was shown to be a consequence of auxin accumulation in the endodermal cells (Rakusová et al. 2016). This relocalization results in a back flow of auxin from the lower flank of the responding hypocotyl into the vasculature. As a consequence, the auxin gradient initially created across the stimulated hypocotyl progressively dissipates, eventually disappearing. With the auxin gradient gone, the curvature ceases, and the hypocotyl tip resumes straight upward growth (Rakusová et al. 2016).
The amazing conclusion from the studies described above is that the auxin gradient generated across the hypocotyl upon gravistimulation by PIN3 polarization triggers both an upward curvature and a subsequent condition that is favorable to PIN3 inner lateralization on the lower, auxin exposed, flank of the hypocotyl, leading to gradient dissipation and termination of the response. This elegant analysis provided a simple explanation to the mechanism involved in the termination of shoot gravitropism. Interestingly, mutations or pharmacological treatments leading to alterations in this auxin-dependent inner lateralization of the PIN3 protein in shoot endodermal cells lead to overshooting the gravitropic response, as anticipated by the model.
In monocots, seedling coleoptiles and shoot pulvini develop strong gravitropic responses. Coleoptile gravitropism has received much attention over the years. Immediately after germination, a monocot coleoptile grows mostly by cell expansion for a few hours, enclosing the first developing leaves. It develops strong gravitropic responses, accompanied by auxin accumulation along the lower flank, as long as the leaves remain enclosed. As soon as the first growing leaves emerge from the tip, the coleoptile loses its ability to develop a gravicurvature. As discussed in Sect. 3.3 of this chapter, experiments using wheat coleoptiles allowed Chauvet and collaborators to suggest an inclination/position sensor mechanism of gravity sensing by plant statocytes (Chauvet et al. 2016).
In adult monocot plants, shoot gravitropism typically involves the contribution of pulvini, which are short segments of tissue that are apical to the nodes and collectively contribute to bringing a shoot tip back up after previous prostration by wind or rain. In Panicoid species like maize, the pulvini constitute disc-shaped segments of the stem, whereas the pulvini of Festucoid grass species, such as wheat, oat, and barley, are made of a tissue that encircles the leaf sheath immediately apical to the point where it attaches to the node. When monocot stems are gravistimulated, such as upon prostration by heavy wind or rain, cells at the bottom side of 3–4 pulvini along the stem resume cell elongation on the lower flank, resulting in local segmental upward curvature (Fig. 5.9). In maize, each pulvinus can provide a maximum of 30° curvature in response to gravistimulation. This process plays an important role in agriculture because it keeps seed away from soil moisture and pathogens after prostration in heavy storms. Another reason for which pulvinus gravitropism has been heavily investigated in monocot plants is that it takes a long time for a pulvinus to develop a curvature response to gravistimulation. Therefore, this system can be used quite efficiently to independently investigate the molecular mechanisms that contribute to gravity sensing and/or signal transduction, relative to those involved in the curvature response.

The gravitropic response of a Brachypodium distachyon stem involves localized curvatures at several successive leaf pulvini. Panel (a) shows vertically oriented (left) and gravistimulated (right) stems (with leaves cut off at their bases to allow better observation of the pulvini). Panels (b and c) show individual pulvini from vertical control (b) and gravistimulated stems (c). Please note that gravistimulation promotes cell elongation on the bottom side of the pulvini (left side in panel c) relative to control or the upper side
Similar to the other shoot systems described above, gravity sensing in cereal pulvini seems to involve the sedimentation of starch-filled plastids within the starch sheath cells that surround the vasculature. Upon gravistimulation, an auxin gradient also forms across stimulated pulvini, with accumulation at the bottom flank. This gradient leads to increased cellular expansion on the lower side, hence upward curvature. However, before this auxin gradient can form, a number of very fast physiological changes also occur, which may contribute to gravity signal transduction. First, an increase in the levels of InsP3 was documented on the bottom flank of gravistimulated pulvini (Perera et al. 1999, 2001). In maize pulvini, this change occurred already within 10 s of gravistimulation, and it was followed by fluctuations between upper and lower sides over a period of 30 min. Subsequently, a stable increase in the levels of InsP3 was observed in the lower flank of the stimulated pulvini over a period of 3–7 h. The first signs of gravicurvature appeared about 8 h after the onset of gravistimulation (Perera et al. 2001). As previously discussed in this chapter, these changes in InsP3 levels upon gravistimulation may contribute to Ca2+ signaling, although gravity-induced Ca2+ changes have not been observed in pulvinus statocytes.
Investigators have also demonstrated the existence of fast changes in the levels of reactive oxygen species, including H2O2, in pulvinus statocytes upon gravistimulation, with initial changes occurring in proximity of the sedimenting amyloplasts within 1 min and expanding throughout the cytoplasm within 30 min of a gravistimulus (Clore 2013). Subsequently, more H2O2 was found on the lower half than on the upper side, possibly contributing directional information for upward bending. This is consistent with a parallel accumulation of a cytoplasmic aconitase/iron regulatory protein 1 (IRP1), which may function as a redox sensor (Clore 2013). The role of reactive oxygen species in gravity signaling remains uncertain. However, reactive oxygen species have been suggested to function both before and after auxin redistribution in gravistimulated maize pulvini (Clore et al. 2008).
In addition to the changes discussed above, pulvinus statocytes also display fast changes in cytosolic pH in response to gravistimulation. A significant alkalinization of the cytoplasm was observed at the bottom side, near the sedimenting amyloplasts, after 30 min of gravistimulation, and it was accompanied by a slight acidification at the sides of the same cells (Johannes et al. 2001). These cytosolic pH changes were suggested to contribute to gravity signaling.
Protein kinases were also implicated in pulvinus responses to gravistimulation. Indeed, MAP kinase activities were shown to first fluctuate in gravistimulated maize pulvini starting 75 min into continuous gravistimulation and followed by a stabilization of the response, with increased activity on the topside after 2 h. Inhibition of MAP kinase activity using the U0106 inhibitor led to alterations in the gravitropic response, suggesting a contribution in gravity signal transduction (Clore et al. 2003). It has been suggested that this sustained increase in MAP kinase activity on the upper section of pulvini may contribute to sustained inhibition of growth on that side.
7 Conclusion
In this chapter, we tried to summarize the current state of our understanding of the molecular mechanisms that govern gravitropism in plants undergoing primary growth. We described seminal experiments that allowed mapping regions within plant organs that contribute to gravity sensing and/or curvature response and discussed several models that attempt to explain gravity sensing by statocytes. We also pointed out recent experiments that suggest a mechanism allowing detection of organ inclination rather than gravity force. We discussed how activation of a signal transduction pathway within the statocytes triggers a change in polar distribution of auxin transporters, resulting in a lateral transport of auxin toward the bottom side of the stimulated organ. A variety of regulatory mechanisms that contribute to the propagation of the resulting auxin gradient from the site of sensing to the site of response, and its maintenance during transport, were also reported, as were the molecular mechanisms that contribute to cellular responses and organ curvature. We described some of the mechanisms that lead to termination of curvature at the end of a response.
Lateral plant organs were reported to grow at distinct angles from the vertical relative to the primary organs they originated from and to be able to modify that angle in response to a variety of environmental and endogenous parameters. Overall, these responses allow a plant to develop a general architecture that allows efficient exploration for acquisition of the resources it needs to sustain growth, development and reproduction.
Yet, despite the tremendous progress recently made toward a better understanding of plant gravitropism, many questions remain unanswered. For instance, the receptors involved in converting information derived from amyloplast sedimentation and/or position within the statocytes into a biochemical signal have not been identified, and the secondary messengers that contribute to this response have not been characterized. The secondary mechanism of gravity sensing known to function in the DEZ of roots remains ill-defined, and functionally redundant auxin transporters contributing to gravity signal transduction and auxin gradient transmission toward the DEZ have not been characterized. Similarly, the mechanisms that modulate root curvature termination at the end of a gravity response remain unknown. Finally, the molecular mechanisms that govern differential cell elongation in response to gravistimulation remain poorly understood, in part because the key regulators of cell wall loosening and/or other aspects of anisotropic cell expansion are functionally redundant.
Yet, we anticipate major progress in our understanding of the mechanisms that govern plant gravitropism in the next few years. Indeed, novel tools are available to answer the remaining questions. Available growth resources in the microgravity environment provided by the International Space Station should allow a better characterization of the mechanisms involved in gravity sensing. Furthermore, the revolution in genome editing driven by the development of CRISPR/Cas9 technologies will be instrumental at identifying and functionally characterizing key genes involved in the different phases of gravitropism (Jiang et al. 2014). Novel biological sensors allow detection of more signaling molecules previously implicated in gravity signaling (Ca2+, H+, InsP3, NO and ROS) (Costa et al. 2013; Hou et al. 2011). Coupled with the development of better real-time imaging approaches and computerized image analysis routines, these sensors should allow the development of better spatiotemporal maps of signal evolution along plant organs during gravitropic responses. Furthermore, system biology approaches relying on forward and reverse genetics, genome-wide association studies, and transcriptomic, proteomic, and metabolomic approaches should allow the identification of novel gravity signal transducers. Finally, the development of mathematical models that attempt to explain quantitative aspects of the gravitropic response by integrating some of its contributing factors should provide a holistic view of the process (Band et al. 2012).
While our description of gravitropism has focused on plant organs that undergo primary growth (driven by apical meristems), it is important to understand that plant organs that undergo secondary growth, such as the woody stems of trees, can also change their orientation relative to gravity by developing reaction wood that provides a force to reorient upward. This process plays important roles in tree architecture, tree posture, and stem reorientation after prostration by heavy storms. Graviperception by angiosperm woody stems leads to the formation of tension wood on their upper side, which creates a tensile force that pulls it upward. On the other hand, a prostrated gymnosperm stem will develop compression wood on its bottom side, which generates a compressive force that pushes it upward. These reactions of prostrated woody stems are important because they modulate the architecture of trees and also contribute to plant survival. Furthermore, the reaction wood they generate alters the market value of lumber, thereby leading to important economical impact in forestry. The molecular mechanisms that govern these tree responses have also received some attention from researchers in the last few years. Unfortunately, space constraints do not allow us to further explore these fascinating processes. For further information, we would like to refer interested readers to a recently published review of the process (Groover 2016).
References
Abas L, Benjamins R, Malenica N, Paciorek T, Wirniewska J, Moulinier-Anzola J, Sieberer T, Friml J, Luschnig C (2006) Intracellular trafficking and proteolysis of the Arabidopsis auxin-efflux facilitator PIN2 are involved in root gravitropism. Nat Cell Biol 8:249–256
Adamowski M, Friml J (2015) PIN-dependent auxin transport: action, regulation, and evolution. Plant Cell 27:20–32
Andreeva Z, Barton D, Armour W, Li M, Liao L, McKellar H, Pethybridge K, Marc J (2010) Inhibition of phospholipase C disrupts cytoskeletal organization and gravitropic growth in Arabidopsis roots. Planta 232:1263–1279
Bai H, Murali B, Barker K, Wolverton C (2013) Low phosphate alters lateral root setpoint angle and gravitropism. Am J Bot 100:175–182
Baldwin K, Strohm A, Masson P (2013) Gravity sensing and signal transduction in vascular plant primary roots. Am J Bot 100:126–142
Band L, Wells D, Larrieu A, Sun J, Middleton A, French A, Brunoud G, Sato E, Wilson M, Péret B et al (2012) Root gravitropism is regulated by a transient lateral auxin gradient controlled by a tipping-point mechanism. Proc Natl Acad Sci U S A 109:4668–4673
Barbosa I, Zourelidou M, Willige B, Weller B, Schwechheimer C (2014) D6 PROTEIN KINASE activates auxin transport-dependent growth and PIN-FORMED phosphorylation at the plasma membrane. Dev Cell 29:674–685
Barlow P (1995) Gravity perception in plants – a multiplicity of systems derived by evolution. Plant Cell Environ 18:951–962
Bastien R, Bohr T, Moulia B, Douady S (2013) Unifying model of shoot gravitropism reveals proprioception as a central feature of posture control in plants. Proc Natl Acad Sci USA 110:755–760
Bérut A, Chauvet H, Legué V, Moulia B, Pouliquen O, Forterre Y (2018) Gravisensors in plant cells behave like an active granular liquid. Proc Natl Acad Sci USA 115:5123–5128
Blancaflor E (2013) Regulation of plant gravity sensing and signaling by the actin cytoskeleton. Am J Bot:1–10
Blancaflor E, Fasano J, Gilroy S (1998) Mapping the functional roles of cap cells in the response of Arabidopsis primary roots to gravity. Plant Physiol 116:213–222
Blilou I, Xu J, Wildwater M, Willemsen V, Paponov I, Friml J, Heidstra R, Aida M, Palme K, Scheres B (2005) The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 433:39–44
Boonsirichai K, Sedbrook J, Chen R, Gilroy S, Masson P (2003) ARG1 is a peripheral membrane protein that modulates gravity-induced cytoplasmic alkalinization and lateral auxin transport in plant statocytes. Plant Cell 15:2612–2625
Brumos J, Robles L, Yun J, Vu T, Jackson S, Alonso J, Stepanova A (2018) Local auxin biosynthesis is a key regulator of plant development. Dev Cell 47:306–318
Caspar T, Pickard B (1989) Gravitropism in a starchless mutant of Arabidopsis: implications for the starch-statolith theory of gravity sensing. Planta 177:185–197
Chauvet H, Pouliquen O, Forterre Y, Legué V, Moulia B (2016) Inclination, not force, is sensed by plants during shoot gravitropism. Sci Rep 6:35431
Clore A (2013) Cereal grass pulvini: Agronomically significant models for studying gravitropism signaling and tissue polarity. Am J Bot 100:101–110
Clore A, Turner W, Morse A, Whetten R (2003) Changes in mitogen-activated protein kinase activity occur in the maize pulvinus in response to gravistimulation and are important for the bending response. Plant Cell Environ 26:991–1001
Clore A, Doore S, Tinnirello S (2008) Increased levels of reactive oxygen species and expression of a cytoplasmic aconitase/iron regulatory protein 1 homolog during the early response of maize pulvini to gravistimulation. Plant Cell Environ 31:144–158
Cosgrove D (2000) Loosening of plant cell walls by expansins. Nature 407:321–326
Costa A, Candeo A, Fieramonti L, Valentini G, Bassi A (2013) Calcium dynamics in root cells of Arabidopsis thaliana visualized with selective plane illumination. PLoS One 8:e75646
Darwin C (1880) The power of movement in plants. John Murray, London
Dharmasiri N, Dharmasiri S, Estelle M (2005) The F-box protein TIR1 is an auxin receptor. Nature 435:441–445
Dharmasiri S, Swarup R, Mockaitis K, Dharmasiri N, Singh S, Kowalchyk M, Marchant A, Mills S, Sandberg G, Bennett M et al (2006) AXR4 is required for localization of the auxin influx facilitator AUX1. Science 312:1218–1220
Dhonukshe P, Huang F, Galvan-Ampudia C, Mähönen A, Kleine-Vehn J, Xu J, Quint A, Prasad K, Friml J, Scheres B et al (2010) Plasma membrane-bound AGC3 kinases phosphorylate PIN auxin carriers at TPRXS(N/S) motifs to direct apical PIN recycling. Development 137:3245–3255
Dindas J, Scherzer S, Roelfsema M, von Meyer K, Müller H, Al-Rasheid K, Palme K, Dietrich P, Becker D, Bennett M et al (2018) AUX1-mediated root hair auxin influx governs SCFTIR1/AFB-type Ca2+ signaling. Nat Commun 9:1174
Ding Z, Friml J (2010) Auxin regulates distal stem cell differentiation in Arabidopsis roots. Proc Natl Acad Sci USA 107:12046–12051
Dong Z, Jiang C, Chen X, Zhang T, Ding L, Song W, Luo H, Lai J, Liu R, Chen H et al (2013) Maize LAZY1 mediates shoot gravitropism and inflorescence development through regulating auxin transport, auxin signaling and light response. Plant Physiol 163:1306–1322
Fasano J, Swanson S, Blancaflor E, Dowd P, Kao T, Gilroy S (2001) Changes in root cap pH are required for the gravity response of the Arabidopsis root. Plant Cell 13:907–921
Firn R, Digby J (1997) Solving the puzzle of gravitropism – has a lost piece been found? Planta 203:S159–S163
Fitzelle K, Kiss J (2001) Restoration of gravitropic sensitivity in starch-deficient mutants of Arabidopsis by hypergravity. J Exp Bot 52:265–275
Friml J, Wisniewska J, Benkova E, Mendgen K, Palme K (2002) Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 415:806–809
Fukaki H, Fujisawa H, Tasaka M (1996) Gravitropic response of inflorescence stems in Arabidopsis thaliana. Plant Physiol 110:933–943
Fukaki H, Wysocka-Diller J, Kato T, Fujisawa H, Benfey P, Tasaka M (1998) Genetic evidence that the endodermis is essential for shoot gravitropism in Arabidopsis thaliana. Plant J 14:425–430
Ganguly A, Lee S-H, Cho H-T (2012) Functional Identification of the phosphorylation sites of Arabidopsis PIN-FORMED3 for Its subcellular localization and biological role. Plant J 71:810–823
Ge L, Chen R (2016) Negative gravitropism in plant roots. Nat Plants 2:16155
Geisler M, Blakeslee J, Bouchard R, Lee O, Vincenzetti V, Bandyopadhyay A, Titapiwatanakun B, Peer W, Bailly A, Richards E et al (2005) Cellular efflux of auxin catalyzed by the Arabidopsis MDR/PGP transporter AtPGP1. Plant J 44:179–194
Gendreau E, Traas J, Desnos T, Grandjean O, Caboche M et al (1997) Cellular basis of hypocotyl growth in Arabidopsis thaliana. Plant Physiol 114:295–305
Groover A (2016) Gravitropisms and reaction woods of forest trees – evolution, functions and mechanisms. New Phytol 211:790–802
Guyomarch S, Léran S, Auzon-Cape M, Perrine-Walker F, Lucas M, Laplaze L (2012) Early development and gravitropic response of lateral roots in Arabidopsis thaliana. Philos Trans R Soc Lond B Biol Sci 367:1509–1516
Haberlandt G (1900) Ueber die perzeption des geotropischen reizes. Ber Dtsch Bot Ges 18:261–272
Harrison B, Masson P (2008) ARL2, ARG1 and PIN3 define a gravity signal transduction pathway in root statocytes. Plant J 53:380–392
Hou G, Mohamalawari D, Blancaflor E (2003) Enhanced gravitropism of roots with a disrupted cap actin cytoskeleton. Plant Physiol 131:1360–1373
Hou B-H, Takanaga H, Grossmann G, Chen L-Q, Qu X-Q, Jones A, Lalonde S, Schweissgut O, Wiechert W, Frommer W (2011) Optical sensors for monitoring dynamic changes of intracellular metabolite levels in mammalian cells. Nat Protoc 6:1818–1833
Iino M (1995) Gravitropism and phototropism of maize coleoptiles: evaluation of the Cholodny-Went theory through effects of auxin application and decapitation. Plant Cell Physiol 36:361–367
Ishikawa H, Evans M (1995) Specialized zones of development in roots. Plant Physiol 109:725–727
Jiang W, Yang B, Weeks D (2014) Efficient CRISPR/Cas9-mediated gene editing in Arabidopsis thaliana and inheritance of modified genes in the T2 and T3 generations. PLoS One 9:e99225
Johannes E, Collings D, Rink J, Allen N (2001) Cytoplasmic pH dynamics in maize pulvinal cells induced by gravity vector changes. Plant Physiol 127:119–130
Kato T, Morita M, Fukaki H, Yamauchi Y, Uehara M, Niihama M, Tasaka M (2002) SGR2, a phospholipase-like protein, and ZIG/SGR4, a SNARE, are involved in the shoot gravitropism of Arabidopsis. Plant Cell 14:33–46
Kepinski S, Leyser O (2005) The Arabidopsis F-box protein TIR1 is an auxin receptor. Nature 435:446–451
Kim K, Shin J, Lee S, Kweon H, Maloof J, Choi G (2011) Phytochromes inhibit hypocotyl negative gravitropism by regulating the development of endodermal amyloplasts through phytochrome-interacting factors. Proc Natl Acad Sci USA 108:1729–1744
Kiss J, Hertel R, Sack F (1989) Amyloplasts are necessary for full gravitropic sensitivity in roots of Arabidopsis thaliana. Planta 177:198–206
Kiss J, Wright J, Caspar T (1996) Gravitropism in roots of intermediate-starch mutants of Arabidopsis. Physiol Plant 97:237–244
Kiss J, Edelmann R, Wood P (1999) Gravitropism of hypocotyls of wild-type and starch-deficient Arabidopsis seedlings in spaceflight studies. Planta 209:96–103
Kiss J, Miller K, Ogden L, Roth K (2002) Phototropism and gravitropism in lateral roots of Arabidopsis. Plant Cell Physiol 43:35–43
Kiss J, Millar K, Edelmann R (2012) Phototropism of Arabidopsis thaliana in microgravity and fractional gravity on the International Space Station. Planta 236:635–645
Kleine-Vehn J, Ding Z, Jones A, Tasaka M, Morita M, Friml J (2010) Gravity-induced PIN transcytosis for polarization of auxin fluxes in gravity-sensing root cells. Proc Natl Acad Sci USA 107:22344–22349
Knight T (1806) On the direction of the radicle and germen during the vegetation of seeds. Philos Trans R Soc 99:108–120
Krieger G, Shkolnik D, Miller G, Fromm H (2016) Reactive oxygen species tune root tropic responses. Plant Physiol 172:1209–1220
Kuznetsov O, Hasenstein K (1996) Intracellular magnetophoresis of amyloplasts and induction of root curvature. Planta 198:87–94
Lau S, Smet I, Kolb M, Meinhardt H, Jürgens G (2011) Auxin triggers a genetic switch. Nat Cell Biol 13:611–615
Leitz G, Kang B-H, Schoenwaelder M, Staehelin L (2009) Statolith sedimentation kinetics and force transduction to the cortical endoplasmic reticulum in gravity-sensing Arabidopsis columella cells. Plant Cell 21:843–860
Lewis D, Miller N, Splitt B, Wu G, Spalding E (2007) Separating the roles of acropetal and basipetal auxin transport on gravitropism with mutations in two Arabidopsis multidrug resistance-like ABC transporter genes. Plant Cell 19:1838–1850
Li G, Xue H (2007) Arabidopsis PLD{zeta}2 regulates vesicle trafficking and is required for auxin response. Plant Cell 19:281–295
Li P, Wang Y, Qian Q, Fu Z, Wang M, Zeng D, Li B, Wang X, Li JJ (2007) LAZY1 controls rice shoot gravitropism through regulating polar auxin transport. Cell Res 17:402–410
Limbach C, Hauslage J, Schäfer C, Braun M (2005) How to activate a plant gravireceptor. Early mechanisms of gravity sensing studied in Characean rhizoids during parabolic flights. Plant Physiol 139:1030–1040
Lin D, Nagawa S, Chen J, Cao L, Chen X, Xu T, Li H, Dhonukshe P, Yamamuro C, Friml J et al (2012) A ROP GTPase-dependent auxin signaling pathway regulates the subcellular distribution of PIN2 in Arabidopsis roots. Curr Biol 22:1319–1325
Lu Y-T, Feldman L (1997) Light-regulated root gravitropism: a role for, and characterization of, a calcium/calmodulin-dependent protein kinase homolog. Planta 203:S91–S97
Lu C, Yuan S, Li J (2006) Contribution of both cell elongation and cell division to wheat coleoptile elongation. Belg J Bot 139:167–172
MacCleery S, Kiss J (1999) Plastid sedimentation kinetics in roots of wild-type and starch-deficient mutants of Arabidopsis. Plant Physiol 120:183–192
Mancuso S, Barlow P, Volkmann D, Baluska F (2006) Actin turnover-mediated gravity response in maize root apices. Gravitropism of decapped roots implicates gravisensing outside of the root cap. Plant Signal Behav 1:52–58
Mironova V, Omelyanchuk N, Yosiphon G, Fadeev S, Kolchanov N, Mjolsness E, Likhoshvai V (2010) A plausible mechanism for auxin patterning along the developing root. BMC Syst Biol 4:98
Monshausen G, Miller N, Murphy A, Gilroy S (2011) Dynamics of auxin-dependent Ca2+ and pH signaling in root growth revealed by integrating high-resolution imaging with automated computer vision-based analysis. Plant J 65:309–318
Morita M (2010) Directional gravity sensing in gravitropism. Annu Rev Plant Biol 61:705–720
Morita M, Kato T, Nagafusa K, Saito C, Ueda T, Nakano A, Tasaka M (2002) Involvement of the vacuoles of the endodermis in the early process of shoot gravitropism in Arabidopsis. Plant Cell 14:47–56
Mullen J, Ishikawa H, Evans M (1998) Analysis of changes in relative elemental growth rate patterns in the elongation zone of Arabidopsis roots upon gravistimulation. Planta 206:598–603
Munnik T, Nielsen E (2011) Green light for polyphosphoinositide signals in plants. Curr Opin Plant Biol 14:489–497
Nemec B (1900) Ueber die art der wahrnehmung des schwekraftreizes bei den pflanzen. Ber Dtsch Bot Ges 18:241–245
Noh B, Murphy A, Spalding E (2001) Multidrug resistance-like genes of Arabidopsis required for auxin transport and auxin-mediated development. Plant Cell 13:2441–2454
Perbal G, Jeune B, Lefranc A, Carnero-Diaz E, Driss-Ecole D (2002) The dose-response curve of the gravitropic reaction: a re-analysis. Physiol Plant 114:336–342
Perera I, Heilmann I, Boss W (1999) Transient and sustained increases in inositol-1,4,5-trisphosphate precede the differential growth response in gravistimulated maize pulvini. Proc Natl Acad Sci USA 96:5838–5843
Perera I, Heilmann I, Chang S, Boss W, Kaufman P (2001) A role for inositol 1,4,5-trisphosphate in gravitropic signaling and the retention of cold-perceived gravistimulation of oat shoot pulvini. Plant Physiol 125:1499–1507
Perera I, Hung C-Y, Brady S, Muday G, Boss W (2006) A universal role for inositol 1,4,5-trisphosphate-mediated signaling in plant gravitropism. Plant Physiol 140:746–760
Plieth C, Trewavas A (2002) Reorientation of seedlings in the Earth’s gravitational field induces cytosolic calcium transients. Plant Physiol 129:786–796
Pouliquen O, Forterre Y, Bérut A, Chauvet H, Bizet F, Legué V, Moulia B (2017) A new scenario for gravity detection in plants: the position sensor hypothesis. Phys Biol 14:035005
Rakusová H, Gallego-Bartolomé J, Vanstraelen M, Robert H, Alabadí D, Blázquez M, Benková E, Friml J (2011) Polarization of PIN3-dependent auxin transport for hypocotyl gravitropic response in Arabidopsis thaliana. Plant J 67:817–826
Rakusová H, Abbas M, Han H, Song S, Robert H, Friml J (2016) Termination of shoot gravitropic responses by auxin feedback on PIN3 polarity. Curr Biol 26:3026–3032
Ren H, Gray W (2015) SAUR proteins as effectors of hormonal and environmental signals in plant growth. Mol Plant 8:1153–1164
Rojas-Pierce M, Titapiwatanakun B, Sohn E, Fang F, Larive C, Blakeslee J, Cheng Y, Cutler S, Peer W, Murphy A et al (2007) Arabidopsis P-glycoprotein19 participates in the inhibition of gravitropism by gravacin. Chem Biol 14:1366–1376
Rosquete M, von Wangenheim D, Marhavý P, Barbez E, Stelzer E, Benková E, Maizel A, Kleine-Vehn J (2013) An auxin transport mechanism restricts positive orthogravitropism in lateral roots. Curr Biol 23:817–822
Roychoudhry S, Del Bianco M, Kieffer M, Kepinski S (2013) Auxin controls gravitropic setpoint angle in higher plant lateral branches. Curr Biol 23:1497–1504
Sievers A, Buchen B, Volkmann D, Hejnowicz Z (1991) Role of the cytoskeleton in gravity perception. In: Lloyd C (ed) The cytoskeletal basis of plant growth and form. Academic Press, London, pp 169–182
Silady R, Ehrhardt D, Jackson K, Faulkner C, Oparka K, Somerville C (2007) The GRV2/RME-8 protein of Arabidopsis functions in the late endocytic pathway and is required for vacuolar membrane flow. Plant J 53:29–41
Sinclair W, Trewavas A (1997) Calcium in gravitropism: a re-examination. Planta 203:S85–S90
Stanga J, Boonsirichai K, Sedbrook J, Otegui M, Masson P (2009) A role for the TOC complex in Arabidopsis root gravitropism. Plant Physiol 149:1896–1905
Staves M, Wayne R, Leopold A (1992) Hydrostatic pressure mimics gravitational pressure in characean cells. Protoplasma 168:141–152
Staves M, Wayne R, Leopold A (1997) The effect of external medium on the gravitropic curvature of rice (Oryza sativa, Poaceae) roots. Am J Bot 84:1522–1529
Strohm A, Barrett-Wilt G, Masson P (2014) A functional TOC complex contributes to gravity signal transduction in Arabidopsis. Front Plant Sci 5:148
Su S-H, Gibbs N, Jancewicz A, Masson P (2017) Review: molecular mechanisms of root gravitropism. Curr Biol 27:R964–R972
Swarup R, Kramer E, Perry P, Knox K, Leyser H, Haseloff J, Beemster G, Bhalerao R, Bennett M (2005) Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal. Nat Cell Biol 7:1057–1065
Tanaka A, Kobayashi Y, Hase Y, Watanabe H (2002) Positional effect of cell inactivation on root gravitropism using heavy-ion microbeams. J Exp Bot 53:683–687
Taniguchi M, Furutani M, Nishimura T, Nakamura M, Fushita T, Iijima K, Baba K, Tanaka H, Toyota M, Tasaka M et al (2017) The Arabidopsis LAZY1 family plays a key role in gravity signaling within statocytes and in branch angle control of roots and shoots. Plant Cell 29:1984–1999
Terrile M, París R, Calderón-Villalobos L, Iglesias M, Lamattina L, Estelle M, Casalongué C (2012) Nitric oxide influences auxin signaling through S-nitrosylation of the Arabidopsis TRANSPORT INHIBITOR RESPONSE 1 auxin receptor. Plant J 70:492–500
Toyota M, Furuichi T, Tatsumi H, Sokabe M (2008) Cytoplasmic calcium increases in response to changes in the gravity vector in hypocotyls and petioles of Arabidopsis seedlings. Plant Physiol 146:505–514
Toyota M, Ikeda N, Sawai-Toyota S, Kato T, Gilroy S, Tasaka M, Morita M (2013) Amyloplast displacement is necessary for gravisensing in Arabidopsis shoots as revealed by a centrifuge microscope. Plant J 76:648–660
Tsugeki R, Fedoroff N (1999) Genetic ablation of root cap cells in Arabidopsis. Proc Natl Acad Sci USA 96:12941–12946
Vitha S, Yang M, Sack F, Kiss J (2007) Gravitropism in starch-excess mutant of Arabidopsis thaliana. Am J Bot 94:590–598
Wang J-W, Wang L-J, Mao Y-B, Cai W-J, Xue H-W, Chen X-Y (2005) Control of root cap formation by microRNA-targeted auxin response factors in Arabidopsis. Plant Cell 17:2204–2216
Wang Y, Lin W, Chen X, Xue H (2009) The role of Arabidopsis 5PTase13 in root gravitropism through modulation of vesicle trafficking. Cell Res 19:1191–1204
Wang H-Z, Yang K-Z, Zou J-J, Zhu L-L, Xie Z, Morita M, Tasaka M, Friml J, Grotewold E, Beeckman T et al (2015) Transcriptional regulation of PIN genes by FOUR LIPS and MYB88 during Arabidopsis root gravitropism. Nat Commun 6:8822
Weise S, Kuznetsov O, Hasenstein K, Kiss J (2000) Curvature in Arabidopsis inflorescence stems is limited to the region of amyloplast displacement. Plant Cell Physiol 41:702–709
Whitford R, Fernandez A, Tejos R, Pérez A, Kleine-Vehn J, Vanneste S, Drozdzecki A, Leitner J, Abas L, Aerts M et al (2012) GOLVEN secretory peptides regulate auxin carrier turnover during plant gravitropic responses. Dev Cell 22:678–685
Wolverton C, Mullen J, Ishikawa H, Evans M (2002) Root gravitropism in response to a signal originating outside of the cap. Planta 215:153–157
Yamamoto K, Pyke K, Kiss J (2002) Reduced gravitropism in inflorescence stems and hypocotyls, but not in roots, of Arabidopsis mutants with large plastids. Physiol Plant 114:627–636
Yang H, Murphy A (2009) Functional expression and characterization of Arabidopsis ABCB, AUX 1 and PIN auxin transporters in Schizosaccharomyces pombe. Plant J 59:179–191
Yano D, Sato M, Saito C, Sato M, Morita M, Tasaka M (2003) A SNARE complex containing SGR3/AtVAM3 and ZIG/VTI11 in gravity-sensing cells is important for Arabidopsis shoot gravitropism. Proc Natl Acad Sci USA 100:8589–8594
Yoshihara T, Spalding E (2017) LAZY genes mediate the effects of gravity on auxin gradients and plant architecture. Plant Physiol 175:959–969
Yoshihara T, Spalding E, Iino M (2013) AtLAZY1 is a signaling component required for gravitropism of the Arabidopsis thaliana inflorescence. Plant J 74:267–279
Zhang J, Peer W (2017) Auxin homeostasis: the DAO of catabolism. J Exp Bot 68:3145–3154
Zheng H, van Mollard G, Kovaleva V, Stevens T, Raikhel N (1999) The plant vesicle-associated SNARE AtVTI1a likely mediates vesicle transport from the trans-Golgi network to the prevacuolar compartment. Mol Biol Cell 10:2251–2264
Zheng Z, Zou J, Li H, Xue S, Wang Y, Le J (2014) Microrheological insights into the dynamics of amyloplasts in root gravity-sensing cells. Mol Plant 8:660–663
Acknowledgments
This chapter was made possible by grants from the National Aeronautics and Space Administration (#NNX14AT23G and #80NSSC18K1481), the University of Wisconsin-Madison College of Agriculture and Life Sciences HATCH program (#WIS10338), and a Fall Competition Award from the Office of Vice Chancellor for Research and Graduate Education from the University of Wisconsin-Madison, to PHM. We thank Dr. Chris Wolverton (Ohio Wesleyan University) for providing information on and images of the rotato device.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Su, SH., Masson, P.H. (2019). Gravitropism of Plant Organs Undergoing Primary Growth. In: Sopory, S. (eds) Sensory Biology of Plants. Springer, Singapore. https://doi.org/10.1007/978-981-13-8922-1_5
Download citation
DOI: https://doi.org/10.1007/978-981-13-8922-1_5
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-13-8921-4
Online ISBN: 978-981-13-8922-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)