
Abstract
The intercellular communication, commonly called as Quorum Sensing (QS) or auto induction, has been attributed for various coordinative and community phenomena in Gram positive and Gram negative bacteria. The signalling is facilitated by diffusible signal, auto inducers, in response to population of neighbouring bacteria. Consequently, QS influences bacterial phenotype such as the production of antibiotics. Current understanding of how bacteria mediate antibiotic synthesis in the natural environment is limited to classical quorum sensing receptors and ‘orphan’ quorum sensing receptors. The genetic studies and biochemical investigation of carbapenem synthesis in Serratia and Erwinia carotovora have acknowledged a group of nine genes complex in the assembly namely carRABCDEFGH which are responsible for antibiotic assembly. N-(3oxohexanoyl)-L-homoserine lactone (OHHL) is produced as a product of the independent carI gene activates CarR transcription factor. This OHHL reliant transcriptional activation permits the cells to synchronise expression of carbapenem with cell density. An orphan quorum-sensing receptor, discovered as the soil bacterium Burkholderia thailandensis, differs from classical quorum sensing as this receptor does not respond to characteristic quorum sensing signalling partners. The orphan receptor however responds to antibiotics, such as trimethoprim and sulfamethoxazole. Consequently, eliciting the expression of the genes malA-M which is involved in synthesis of the cytotoxic antibiotic malleilactone. This controlling pathway might be vital to sense and compete in mixed communities. The synthesis of antibiotic is very expensive for the bacteria therefore the induction and modus of induction is tightly regulated. This work is an understanding of the current view of quorum sensing and their function in modulating antibiotics synthesis.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
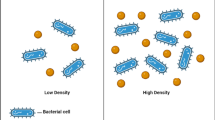
The intercellular communication, commonly called as Quorum Sensing (QS) or auto induction, has been attributed to various coordinative and community phenomena in Gram positive and Gram negative bacteria [1]. The signalling is carried out by diffusible signal, auto inducers, in response to population of neighbouring bacteria. Bacteria guard the expression of specialized gene sets in response to their cell density [2]. The bacteria sense various environmental factors such as the magnitude of their population by sensing the amount of tiny, diffusible, signal molecules termed as auto-inducers as they are produced by self synthesis. These molecules mediate signalling by secretion and reabsorption into the cells by facile diffusion among other processes. These signals thereby accumulate in the extra cellular environment as a function of the growth of the cells. At certain critical concentration which corresponds with a threshold population density, these signals serve as co-inducers to regulate transcription of target genes. The output of which presumably are of asset to the bacteria in the particular habitat. Consequently, QS influences bacterial phenotype such as the production of antibiotics.
The phenomenon of quorum sensing was explained by studying the bioluminescent bacteria Vibrio fischeri. The symbiotic bacteria is commonly found in marine hosts such as fish and cephalopods and is bioluminescent when their number reach threshold population [3]. The study of the bacteria helped decipher the molecular mechanism. The bacteria release its auto inducer, V.fischeri auto-inducer (VAI) during their growth in the marine host’s light organ. Upon reaching the threshold population, Vibrio fischeri auto-inducer activates the transcription of the lux operon using a transcription activator, LuxR [3]. At the necessary concentration of VAI, VAI and LuxR interact and consequently convert LuxR into a function transcriptional activation factor [4]. The functional active LuxR activates the lux operon. LuxI is the first gene on the operon, the protein product of the gene catalyzes the synthesis of VAI [4]. This auto inducer mediated amplification of the auto induction allows increased amount and amplification of the transcription of the gene in the target operon. Thus, the auto inducer increases its production in accordance with the population and bacterial dependent production of desired target gene on the operon.
Quorum sensing has been exhibited to regulate different genes in many bacteria of different genera due to continuous and sustained study. The extensive study has allowed characterization of several genes such as Agrobacterium tumefaciens which regulates conjugal transfer of plasmid by quorum sensing. The conjugal transfer is controlled by using the transcriptional activator TraR and the diffusible molecule AAI [5]. Pseudomonas aeruginosa regulates several gene systems by systems control genes, those include genes needed for the production of rhamnolipids, and extracellular enzymes and toxins [6]. The genes are regulated by two quorum-sensing systems; every system has has its own cognate transcriptional regulator and diffusible signal molecules. Another pseudomonad, P. aureofaciens, monitors production of phenazine antibiotics by quorum-sensing [7]. These antibiotics have been implicated in the disease of wheat suppressive disease. Erwinia carotovora are plant pathogens which guarded the expression of pathogenic traits by quorum sensing systems. [8] Serratia is also mediates quorum sensing which regulates the synthesis of antibiotic carbapenem and pigment prodigiosin [9]. Literature and studies indicate the similar regulatory strategies are employed by the bacteria albeit different target genes are controlled by the auto induction differ in accordance with the bacterial requirements.
Current understanding of how bacteria mediate antibiotic synthesis in the natural environment is limited to classical quorum sensing receptors and ‘orphan’ quorum sensing receptors [6]. The molecular level investigation of carbapenem synthesis in both Serratia and Erwinia carotovora have reported a group of nine genes engaged in the synthesis of the antibiotic (carRABCDEFGH) [9]. N-(3oxohexanoyl)-L-homoserine lactone (OHHL) synthesized by the product of carI gene which activates the transcription factor named as CarR. This transcription factor permits the cells to work in density depended manner in order to express carbapenem. Another quorum sensing receptor also known as orphan receptor has been discovered in the Burkholderia thailandensis which is a soil bacterium [10], differs from classical quorum sensing as this receptor does not respond to common quorum sensing signalling partners. Infact, these receptors respond to selective antibiotics, such as trimethoprim and sulfamethoxazole [10]. Consequently, expression of the genes mal, involved in synthesis of the cytotoxic antibiotic malleilactone is triggered. This regulatory pathway may be important to sense and compete in mixed communities. The synthesis of antibiotic is very expensive for the bacteria therefore the induction and modus of induction is tightly regulated.
The present work explains the classical quorum sensing mechanism as described in various bacteria. The auto inducer and their homologues control the target gene on the operon. The work will focus on the antibiotic synthesis as a result of the quorum sensing by the bacteria. The antibiotic synthesizing gene attributes survival of the population in a mixed bacterial community and also pathogenicity in certain strains of bacteria.
Auto Induction in V. fischeri
The V. fischeri auto inducer (VAI) is 3-oxo-N-(tetrahydro-2- oxo-3-furanyl) hexanamide also known as N-3-(oxohexanoyl) homoserine lactone. The auto induction phenomena of this species help them to distinguish between the plantonic and colonized bacteria within the host and thus induces the luminescences system in the density depended manner only when the bacteria is interacted with the host. At intracellular critical concentration on the order of 10 nM the transcription of the luminescent genes are activated. The bacteria passively diffuse the auto inducer, VAI at low cell densities. On reaching the threshold population VAI accumulates in the bacteria equivalent to the extracellular concentration. In presence of VAI Lux operon transcription activator LuxR is activated. LuxR is encoded by one unit of the lux operon. It is a 250 amino acid protein which in the presence VAI transcribes luminescent genes encoded in the other unit of the cluster. The cluster is divided into two units whose start sites separated by about 150 bp. One unit encodes luxR and the other unit is the operon, luxICDABEG [11]. The LuxR binding site or the lux box has a dyad symmetry is about −40 bp upstream from the start of luxICDABEG transcription. The sequence is required for luxICDABEG activation by LuxR and for positively or negatively auto regulate luxR depending on the VAI and LuxR levels in the cell. As a consequence of the cellular accumulation of VAI and the activation of LuxR luxI gene is transcribed. The luxI gene coded for a 193-amino-acid protein is considered to be an auto inducer synthase as the LuxI catalyses synthesis of VAI leading to its amplification [12]. The other genes in the luxIC- DABEG operon carried out the synthesis of light. lux4 and luxB code for the a and b, subunits of luciferase. luxC, luxD, and luxE encode for components of the fatty acid reductase complex required for synthesis of the aldehyde substrate for luciferase. luxG might code for a flavin mononucleotide reductase that produces reduced flavin mononucleotide as a substrate for luciferase [3]. This genomic cluster shares homologues in other bacteria engaging in quorum sensing and encodes gene required by the bacteria for desired traits.
Auto Inducer
Autoinducer are the small molecules releases in respond to population depended manner. Majority of them are N-acylated derivatives of L-homoserine lactone (acyl- HSLs).The synthesis of these molecules is carried by different organism which varies in the length, the positioning of 3-carbon and the availability of unsaturated bonds within the acyl side chain.The illustration can be well explained by the autoinducers of VAI and Agrobacterium species. The autoinducer of VAI is N (3 oxo hexanoyl) L HSL whereas the N(3 oxo octanoyl) L HSL belongs to the Agrobacterium species. The another example of different nature of AI can be well understood by closely observing the difference in Chromobacterium violaceum which manufactured the N hexanoyl L HSL, while the AI produced by isolates of Rhizobium spp. is N-(3-hydroxy-7-cis-tetradecanoyl)-L-HSL.
Orphan Receptor
LuxR proteins are transcription factors that are activated by acyl-homoserine lactone (AHL) signals. Recent studies into the signalling molecules and their receptors have identified conserved LuxR family protein, MalR in B.thailandensis which induces genes independently of AHLs. MalR is needed for expression of genes coding for synthesis of the cytotoxic malleilactone [10]. The mal genes are induced to synthesize MalR which consequently synthesize malleilactone needed few antibiotics, such as trimethoprim. The mechanism needs to be elucidated of the pathways that the antibiotic trimethoprim employs to increase malR expression. At sufficient amounts of malR expression, MalR functions solely of any external signal. It has been reported that the muted biosynthetic genes of malleilactone get activated by MalR which works in a AHLs independent manner.The absence of cognate ligand lends the name orphan receptor to the receptors [6].
LuxR, Transciption Activator
LuxR is an important transcription activator although no homologues have been identified in vitro. It has two domain polypeptide containing GroEL and GroES which folds into an active conformation [1]. Mutational studies and analysis of the LuxR polypeptide identified regions which were responsible for its function. Amino acids at the N terminus mainly residues 10–20 have an inhibitory role which is reversed upon auto inducer accumulation [3]. The amino terminus is responsible for downregulating of the luxR gene but isn’t responsible for the binding to the lux box or activation of luxICDABEG. The mutational studies further showed that removing residues 2–156 results in an increase of an auto inducer-independent luxICDABEG operon transcription [12]. Thus indicates its regulatory function. In addition, the study shows that the C-terminal region, residues 190–210, has a DNA binding function through a helix-turn-helix (HTH) motif and is responsible for transcriptional activation [4]. While the amino-terminal half of the protein has an inhibitory role that is overcome by auto inducer. Single-amino-acid mutations between residues 79 and 127 affected the interactions with VAI. VAI binds to LuxR by a multimer formation by the residues in the region of 116–161 and subsequent binding to the lux boxes [13]. The studies further showed that the residues 230–250 may not be required for DNA binding but they may be needed for activation by making contacts with RNA polymerase.
LuxI
The luxI gene encodes the protein LuxI which amplifies the accumulation and synthesis of VAI in both V. fischeri and E. coli [3]. The function makes a strong case for the gene to encode VAI synthase. It has been shown that The LuxI protein catalyze the production of VAI from S-adenosylmethionine and 3-oxohexanoyl coenzyme A but the amount of 3-oxohexanoyl coenzyme A needed to saturate the enzyme activity are high, suggesting that 3-oxo- hexanoyl acyl carrier protein to be the obligatory substrate for VAI synthase [14]. Production of V. harveyi autoinducer, HAI uses the D isomer of its fatty acid precursor indicating that the acyl moieties of AI are derivative of intermediates lipid biosynthetic pathways [13].
Homologous Systems
Several bacteria contain homologous system of quorum sensing which act through LuxI and LuxR homologues for desired traits. Literature indicates systems for antibiotic synthesis in several bacteria such as Erwinia carotovora.Itis a plant pathogen which colonizes vascular tissues of host plants. The bacteria produces cell wall-degrading exo-enzymes required for virulence [15]. The study of bacteria led to the identification of LuxI homologue ExpI. Quorum sensing by the bacteria E. carotovora is mediated by an auto inducer identical to VAI. VAI induces synthesis of the antibiotic carbapenem in E. carotovora [8]. While no LuxR homolog have been identified and shares sequence similarity with LuxR open reading frame downstream from expI. This indicates it encodes VAI dependent regulator.
The synthesis of antibiotics in Streptomyces spp. by quorum sensingis depend upon the molecule called butyrolactones which is similiar to VAI. A factor from Streptomyces griseus regulates a number of secondary metabolic processes including streptomycin synthesis thus the butyrolactone-mediated regulation seems similar to auto inducer-dependent [16]. It is showed that the LuxR-type proteins activate transcription but the A-factor receptor is a repressor thus is not homologous to LuxI.
Apart the canonical quorum sensing cascade to synthesize the secondary metabolite, the antibiotic has several functions. In certain systems they act as signals while in certain systems their induction deviates from the classical LuxR-LuxI system. Such systems are important indicators of different modalities of quorum sensing and the role of secondary metabolites.
Erwinia carotova
Erwinia carotova is a plant pathogen which releases exo-enzymes to degrade plant cell wall, it also releases beta- lactam antibiotic to compete against other bacterial population in a mixed community [17]. The release is cell number is dependent, which enables the pathogen to mount a sustained and successful attack on the host. The synthesis of the antibiotic is regulated by quorum sensing wherein the antibiotic synthesis follows the LuxR system [18]. The auto induction mediates beta-lactam (carbapenem) synthesis on intracellular accumulation of auto inducer N-(3-oxohexanoyl)-L-homoserine lactone (3-oxo-C6-HSL) [9]. The accumulation of HSL intracellularly mediates the amplification by LuxI homologue CarI. CarI mediates synthesis of 3-oxo-C6-HSL consequently the HSL binds to LuxR homologue CarR. Studies show unequivocal evidence of the LuxR superfamily protein being receptor of HSL ligands [8]. LuxR homologue, CarR attains its active form on HSL binding. This binding activates the CarR transcription activator mediated activation of car genes [17]. The binding of HSL activates the CarR to its active protein form due to its intrinsic tendency to multimerize. The activation induces the transcriptional activation of genes [8].
Serratia also produces beta-lactam antibiotic, 1-carbapen-2-em-3-carboxylic acid [18]. Extensive study into the genetics of the antibiotic synthesis has identified a cluster of nine genes (barABCDEFGH) [9]. Functions have been assigned to all the genes in the cluster except carH. The gene cluster encodes the enzymes involved in the beta lactam biosynthesis, a novel beta-lactam resistance machinery and also a positive regulator of the carbapenem gene. carR in the gene cluster encodes CarR which is LuxR family homologue [18]. Thus lactone auto inducer N-(3-oxohexanoyl)-L-homoserine lactone (OHHL) synthesized by the unlinked gene product CarI activates CarR which in turn activates the transcription of the remaining genes in the cluster. Thus the auto inducer dependent activation coordinates the expression of carbapenem with cell density and therefore called quorum sensing. The production of antibiotic is regulated by protein Rap in Serratia and Hor in Erwinia [9]. The regulators directly bind to the DNA and activate the genes in the stimulon. Despite, the key similarities the bacterial systems are not identical. The bacteria induce the antibiotic production depending on the growth phase in Erwinia while it parallel to the growth in Serratia. Further unlike Erwinia, Serratia can also induce carbapenem synthesis in response to certain pheromones.
Pseudomonas aeruginosa System, Antibiotic as Terminal QS Signal
Pseudomonas aeruginosa is a gram negative bacteria which regulates gene expression by quorum sensing. Extensive studies have identified two systems controlled by their transcription activators LasR and RhIR. These systems respond to auto inducer AHLs 3-oxo-dodecanoyl-homoserine lactone (3O-C12-HSL) and butanoyl-homoserine lactone (C4-HSL) respectively [6]. The two systems induce gene expression by accumulating the auto inducers in the cells. The accumulation and binding of respective ligands to transcription activators activates a molecular switch to active protein. The transcription activator mediates transcription of lasI and rhII genes and other genes in the operon. The proteins LasI and RhII amplify AHLs 3-oxo-dodecanoyl-homoserine lactone (3O-C12-HSL) and butanoyl-homoserine lactone (C4-HSL) synthesis respectively [11]. These systems are LuxR system homologues but recent studies into the bacteria revealed a third system responsible for antibiotic synthesis [2]. The signal is found to be a part of the P.aeruginosa quorum sensing through a quinolone 2-heptyl-3-hydroxy-4-quinolone (PQS). PQS production is regulated by quorum sensing and is similar to Pyo compounds [11]. These compounds are identified as antibiotics belonging to the 4-quinolones family. The quorum sensing mechanism of PQS is different from classical systems or LuxR systems because it is not an AHL. Further the genes responsible for the synthesis of PQS are not LuxI homologues. None-the-less, they are qualified as a quorum sensing signal as the signals are small diffusible compounds which are cell density dependent and accumulates in the cell. [7] The accumulation triggers transcriptional response of specific genes thus PQS are defined in QS systems. The PQS system does not have canonical quorum sensing homologues but the signal and response system is in temporal cascade. AHLs are released in the early phase while PQS is released in the late exponential phase as the terminal signal in the temporal cascade [11]. P. aeruginosa positively regulates several virulent factors, monooxygenase and the proton driven efflux pumps of resistance nodulation cell division transporter mexGHI-opmD operon through MvfR [19]. These responses are mediated by PQS through PqsE. In addition to the virulent factors, PQS signalling produces phenazines in its response. Phenazines are heterocyclic, redox active compounds which are toxic to other prokaryotes and eukaryotes [19]. Phenazines are also released from cells at late exponential phase. Thus in the three signal system phenazine is the terminal product [7]. The quorum sensing cascade comprises of three signal which are interdependent and function in a growth stage manner. LasI and LasR synthesize AHLs 3O-C12-HSL and butanoylhomoserine lactone (C4-HSL) respectively. These signals bind to their transcription activators LasR and RhIR and in turn induce the gene expression [7]. This results in the synthesis and secretion of PQS, required for the production of phenazines. The terminal phenazine signal activates the transcription factor SoxR and induces the expression of mexGHI-opmD operon and virulent factor PA2274 [19]. The mechanism of activation of SoxR remains to be elucidated but it is found to be active in strictly anaerobic condition. Thus, the phenazine is a necessary signalling molecule to upregulate the expression of a set of gene. The genes are involved in the efflux and redox action; in addition to that they are also important in iron acquisition. MexGHI-opmD and PA2274 are controlled by the phenazine signal as well.
PQS on the other hand, controls the expression of the phz operon which synthesize phenazine. It has been shown that the production of phenazine connects expression of mexGHI-opmD and PA2274 to PQS. This explains the phenazines position as a terminal signal molecule in QS circuitry. SoxR controls the six genes which includes monooxygenase (PA2274) and two transporters, i.e. the RND efflux pump MexGHI-OpmD and the MFS transporter PA3718. PA2774 which is defined as monooxygenase act similar ActVA Orf6 of Streptomyces coelicolor, thus oxygenates actinorhodin. Actinorhodin is an aromatic polypeptide and similar to structure of phenazines [19]. The conversion of PCA to PYA is carried out by PhzS This enzyme is also involve in monooxygenase as well. Thus it clears that PA2274 might recognize phenazines, permits to work against the monooxygenase in either function as an enzymatic or act as a competitor. The PhzS might also works as a chaperone which enables the protection of the cellular environment from the toxic phenazines, or behave as receptors which sense phenazines, thereby affecting gene expression. MexGHI-OpmD is one of 10 potential RND pumps in P. aeruginosa [11]. It has indicated that the efflux pump is needed for the excretion of a PQS precursor. Further, it is also indicated that MexGHI-OpmD is implicit in the secretion of a toxic PQS precursor and also in the efficient to and fro of phenazines [11]
Phenazine and quinolones have been classically considered only as antibiotics due to their antimicrobial activities [7]. Recent studies add an additional dimension wherein quinolone and the phenazine PYO can function as intercellular indicators thus as a ‘secondary’ metabolites which maybe of significant importance adapting the cellular response to a particular physiological state [7]. The quorum sensing network is intricately designed to optimize the desired effect for a population. One sees such intricacy in phenazine production initiated under limited oxygen conditions. In such conditions when the phenazines are synthesized, they respond with oxidized species such as ferric oxide and consequently facilitate the acquisition of iron. Hence phenazine reins the countenance of genes involved in iron attainment and redox homeostasis, and genes that modulate the self-processing (mexGHI-opmD) [6] Fig. 1.

Model of the network in P. aeruginosa. The quorum sensing system is a growth stage dependent flow of three signalling molecules wherein the LasI and LasR manufacture the AHLs 3O-C12-HSL and butanoylhomoserine lactone (C4-HSL) respectively, during the exponential phase. Consequently, interacting with transcriptional activators, LasR and RhlR respectively induce the gene expression. Which in turn potentiates the synthesis and release of PQS? PQS is mandatory for the production of phenazines and the phenazine PYO triggers the transcription factor SoxR. Thus increasing the manifestation of mexGHI-opmD and PA2274. (Source: Molecular Microbiology Volume 61, Issue 5, pages 1308–1321, 25 JUL 2006 DOI: https://doi.org/10.1111/j.1365-2958.2006.05306.x)
Thus, this bacterial system proves that the antibiotic synthesis is not just a secondary metabolite which is an important for the population survival in a mixed community or for successful host invasion, can also function as signalling during the quorum sensing cascade. Further, it can coordinate multiple responses which transcend their antimicrobial activities.
Burkholderia thailandensis, Burkholderia pseudomallei and Burkholderia mallei, Orphan Receptors
Burkholderia thailandensis, Burkholderia pseudomallei and Burkholderia mallei are associated bacteria referred in beta-proteo-bacteria category [20]. They are related with sequence identity which spans several genomic regions. Despite their genomic identity B. mallei is a mammalian pathogen which obligatory chooses solipeds as host but the bacteria can infect human. B. pseudomallei are prevalent human infecting bacteria which are commonly found in soil and water. B.thailandensis is also a mammalian infecting bacteria endemic to central America. These bacteria are pathogenic in nature and quorum sensing has been known to contribute to its virulence. These bacteria have been studied for multiple quorum sensing systems. These systems are classical activated by self-produced extracellular signal that are population density dependent.
These bacteria produce acyl-HSL as their auto inducers. The signals are produced by acyl- synthases belonging to the LuxI superfamily. The acyl-HSL binds to LuxR superfamily proteins and active the transcription factor the LuxR homolog reacts effectively with the HSL synthesized by the equivalent LuxI homolog as their nature varies depending on different LuxI homologs. Therefore the gene for the homologs are present often on adjacent locus of the chromosome and are said to be cognate pair. The homologs in these bacteria are different genes in each and each bacterium has multiple lux homologs for multiple quorum sensing systems. The homolog genes are known as bma in B. mallei, bps in B.pseudomallei and bta in B.thailandensis [6]. These three have highly preserved R1-I1 genes, R3-I3 genes and possess two homolog of orphan luxR. The luxR homolog is R4 and R5. B.pseudomallei and B. thailandensis also have added quorum sensing genes called R2-I2 is absent in B.mallei. Despite this the bacteria have sequence similarity in the amino acid of non-orphan receptors R1-I1, with octanoyl-HSL as their cognate signal. The system has been implicated in siderophore synthesis, phospholipase C manufacture and oxidative stress reaction. The BmaR3-BmaI3 in B.mallei responds to N-3-hydroxy-octanoyl-HSL and BtaI3 of B. thailandensis in addition, responds to the same HSL [21]. 120 kb DNA region of BtaR2-BtaI3 in B.thailandensis is preserved in B.pseudomallei but lacking in B.mallei due to genomic size reduction by deletion during evolution. Similar phenomenon is seen with quorum sensing system associated with R2-I2 containing DNA element. Literature indicates that Bta system generates HSL and responds to the signal by production of antibiotic which would be active against various gram positive bacteria. Bta system produces 3OHC8-HSL and 3OHC10-HSL but BtaI2 is a LuxI homologs in B.thailandensis, further BtaR2 responds to both signals. Thus the HSLs are synthesized by cognate HSL synthase BtaI2 and BtaR2 responds to both the signal [20]. The BtaR2-BtaI2 system of quorum-sensing modulates two of the gene clusters that are present on the 120-kb element. The element is additionally present in B. pseudomallei but is lacking from B. mallei. This120-kb element is considered an island of quorum-sensing [21].
The B. thailandensis mal gene cluster expression can be triggered by certain antibiotics. Studies show that sufficient levels of malR expression are enough to stimulate mal gene manifestation. Further, studies indicate that MalR is an orphan receptor with no cognate HSL [20]. Other than the absence of ligand the MalR is similar to LuxR. It activates lux like sequence for antibiotic mallei-lactone synthesis. The MalR expression for malR gene is activated by some antibiotics in the environment. Antibiotics that triggered mal gene expression retard the development of B.thailandensis. Thus, development and mal activation are inversely correlated [21]. Nonetheless, certain antibiotics slowed development but did not distress mal gene stimulation. Thus, manifestation of some antibiotics stimulates a precise cellular reaction pathway, consequently triggers expression of malR.
The mal genes encode for production of mallei-lactone. The molecule is a cytotoxin that additionally has antibiotic function against many Gram-positive bacteria. The mallei-lactone-biosynthetic gene clusters are conserved and MalR are identical across these three species. Trimethoprim and sulfamethoxazole are two antibiotics which activate mallei-lactone production in B. thailandensis by MalR mediated expression [20]. Thus the BtaR2-BtaI2 reins the synthesis of the ancillary metabolite along with antibiotic function against several of the gram-positive bacteria. The antibiotic production gives microbes a competitive edge in their environment over added antibiotic sensitive microbes in similar environment. Apart from the classical LuxR-LuxI system, orphan receptor activation by antibiotic also produces antibiotics which are important for population survival.
Summary
Quorum-sensing systems are widespread among the bacterial populations. Quorum sensing is facilitated by diffusible minor molecules released by the bacteria till it reaches a threshold population density. On reaching the critical density the molecules accumulate intracellularly to match the external levels. These molecules are recognized by self- receptors and elicit a response by activating a cascade of events which help in survival of population [17]. The system involves the auto inducers which are produced by participants of the LuxI family of synthases. The LuxR protein family members act as related auto inducer reliant transcription factors. The LuxI synthesizes the auto inducer which consequently binds to LuxR [16]. The binding activates the transcription factor and mediates the expression of desired trait. The genes beleaguered by the members of LuxR family have an 18- to 20-base inverted repeat in the promoter regions which is conserved through species [22]. These essentials are known as lux box-like sequences and function to serve as obligatory sites for the transcription factors. Such classical system is exists in several bacteria and has been studied to control the synthesis of antibiotics. Such system is present in Serrati and Erwinia which release Beta- lactam [23].
An emergent view is that antibiotics actually assist intercellular signalling roles as opposed to munitions against competitors. Such a system is seen in phenazine antibiotic released by Pseudomonas aeruginosa. The system uses the antibiotic not only as an important secondary metabolite but also as signalling molecule in the cascade of quorum sensing.
Burkholderia thailandensis has three sets of luxI-luxR-type genes namely, btaI1-btaR1, btaI2-btaR2, and btaI3-btaR3 [23]. The luxR homologs is related to its equivalent luxI homolog, similarly luxI-luxR-type gene sets in several other bacteria. This species have two added luxR homologs, btaR4 (now called malR or btaR5). Members of LuxR family (like MalR and BtaR5) are called orphans due the absence of cognate ligand associated with LuxI [7]. These systems synthesize mallei-lactone as the antibiotic.
The synthesis of antibiotics is important for the bacteria to mount cell population dependent invasion into host, compete in mixed communities of bacteria and activate cascade needed for population survival [22]. These bacterial models of antibiotic synthesis cover the classical models and the present view of structural and functional significance in the pathways. The present work describes the quorum sensing system present in bacteria and the contributing element of the cascade. The homologues of the cascade and their function are elaborated. Further the secondary metabolite i.e., antibiotic is synthesized by several bacteria. This chapter also covered the three models of antibiotic induction using model bacteria. Wherein, Erwinia and Serratia mediated quorum sensing by classical LuxR LuxI system. Pseudomonad displayed the quorum sensing cascade where the phenazine was the desired antibiotic but its synthesis was also a terminal signal and was an integral part in the system. Finally the proteobacteria genera mediate quorum sensing by classical system as well as novel orphan receptor, which regulates the synthesis of antibiotic in response to the environmental antibiotic such as trimethoprim. Hereby, explaining the biosynthetic pathways and their cognate contributing factors.
References
Bassler, B. L. (1999). How bacteria talk to each other: regulation of gene expression by quorum sensing. Current Opinion in Microbiology, 2(6), 582–587. https://doi.org/10.1016/S1369-5274(99)00025-9.
Cha, C., Gao, P., Chen, Y.-C., Shaw, P. D., & Farrand, S. K. (1998). Production of acyl-homoserine lactone quorum-sensing signals by gram-negative plant-associated bacteria. Molecular Plant-Microbe Interactions, 11(11), 1119–1129. https://doi.org/10.1094/MPMI.1998.11.11.1119.
Fuqua, W. C., & Winans, S. C. (1994). Peter Greenberg2 AE. MINIREVIEW quorum sensing in bacteria: The luxR-luxI family of cell density-responsive transcriptional regulatorst. Journal of Bacteriology, 176(2), 269–275. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC205046/pdf/jbacter00020-0013.pdf. Accessed 1 Jan 2018.
Cha, C., Gao, P., Chen, Y.-C., Shaw, P. D., & Farrand, S. K. (1998). Production of acyl-homoserine lactone quorum-sensing signals by gram-negative plant-associated bacteria. Molecular Plant-Microbe Interactions, 11(11), 1119–1129. https://apsjournals.apsnet.org/doi/pdf/10.1094/MPMI.1998.11.11.1119. Accessed 1 Jan 2018.
Raffa, R. B., Iannuzzo, J. R., Levine, D. R., et al. (2005). Bacterial communication (‘quorum sensing’) via ligands and receptors: A novel pharmacologic target for the design of antibiotic drugs. The Journal of Pharmacology and Experimental Therapeutics, 312(2), 417–423. https://doi.org/10.1124/jpet.104.075150.
Oinuma, K., & Greenberg, E. P. (2011). Acyl-homoserine lactone binding to and stability of the orphan pseudomonas aeruginosa quorum-sensing signal receptor QscR. Journal of Bacteriology, 193(2), 421–428. https://doi.org/10.1128/JB.01041-10.
Dietrich, L. E. P., Price-whelan, A., Petersen, A., Whiteley, M., & Newman, D. K. (2006). The phenazine pyocyanin is a terminal signalling factor in the quorum sensing network of Pseudomonas aeruginosa. Molecular Microbiology, 61(5), 1308–1321. https://doi.org/10.1111/j.1365-2958.2006.05306.x.
Welch, M., Todd, D. E., Whitehead, N. A., Mcgowan, S. J., Bycroft, B. W., & Salmond, G. P. C. (2000). N -acyl homoserine lactone binding to the CarR receptor determines quorum-sensing specificity in Erwinia. EMBO Journal, 19(4), 631–641.
Thomson, N. R., Crow, M. A., Mcgowan, S. J., Cox, A., & Salmond, G. P. C. (2000). Biosynthesis of carbapenem antibiotic and prodigiosin pigment in Serratia is under quorum sensing control. Molecular Microbiology, 36, 539–556.
Truong, T. T., Seyedsayamdost, M., Greenberg, E. P., & Chandler, J. R. (2015). A Burkholderia thailandensis acyl-homoserine lactone-independent orphan luxR homolog that activates production of the cytotoxin. Journal of Bacteriology, 197(21), 3456–3462. https://doi.org/10.1128/JB.00425-15.Editor.
Zhang, Z., & Pierson, L. S., III. (2001). A second quorum-sensing system regulates cell surface properties but not phenazine antibiotic production in Pseudomonas aureofaciens. Applied and Environmental Microbiology, 67(9), 4305–4315. https://doi.org/10.1128/AEM.67.9.4305.
Schaefer, A. M. Y. L., Hanzelka, B. L., & Eberhard, A. (1996). Quorum sensing in Vibrio fischeri: Probing autoinducer-luxR interactions with autoinducer analogs. Journal of Bacteriology, 178(10), 2897–2901. Downloaded from http://jb.asm.org/ on January 31, 2018 by NATIONAL INSTITUTE OF IMMUNOLOGY (NII).
Vendeville, A., Winzer, K., Heurlier, K., Tang. C. M., & Hardie, K. R. (2005). Making “sense ” of metabolism: Autoinducer ≡ 2, LuxS and pathogenic bacteria. Nature Reviews Microbiology, 3(5):383–396. https://doi.org/10.1038/nrmicro1146.
Schaefer, A. M. Y. L., Hanzelka, B. L., & Cronan, J. E. (1996). Generation of cell-to-cell signals in quorum sensing: Acyl homoserine lactone synthase activity of a purified Vibrio fischeri LuxI protein. Proceedings of the National Academy of Sciences of the United States of America, 93(18), 9505–9509.
McGowan, S., Sebaihia, M., Jones, S., et al. (1995). Carbapenem antibiotic production in Erwinia carotovora is regulated by CarR, a homologue of the LuxR transcriptional activator. Microbiology, 141(3), 541–550. https://doi.org/10.1099/13500872-141-3-541.
Williams, P. (2007). SGM special lecture quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2018:3923–3938. https://doi.org/10.1099/mic.0.2007/012856-0.
Gristwood, T., Fineran, P. C., Everson, L., Williamson, N. R., & Salmond, G. P. (2009). The PhoBR two-component system regulates antibiotic biosynthesis in Serratia in response to phosphate. BMC Microbiology, 9, 112. https://doi.org/10.1186/1471-2180-9-112.
Thomson, N. R., Crow, M. A., McGowan, S. J., Cox, A., & Salmond, G. P. C. (2002). Biosynthesis of carbapenem antibiotic and prodigiosin pigment in Serratia is under quorum sensing control. Molecular Microbiology, 36(3), 539–556. https://doi.org/10.1046/j.1365-2958.2000.01872.x.
Veselova, M. A., Klein, S., Bass, I. A., et al. (2008). Quorum sensing systems of regulation, synthesis of phenazine antibiotics, and antifungal activity in rhizospheric bacterium pseudomonas chlororaphis 449. Russian Journal of Genetics, 44(12), 1400–1408. https://doi.org/10.1134/S102279540812003X.
Duerkop, B. A., Varga, J., Chandler, J. R., et al. (2009). Quorum-sensing control of antibiotic synthesis in Burkholderia thailandensis. Journal of Bacteriology, 191(12), 3909–3918. https://doi.org/10.1128/JB.00200-09.
Truong, T. T., Seyedsayamdost, M., Greenberg, E. P., & Chandler, J. R. (2015). A Burkholderia thailandensis acyl-homoserine lactone-independent orphan luxR homolog that activates production of the cytotoxin malleilactone. Journal of Bacteriology, 197(21), 3456–3462. https://doi.org/10.1128/JB.00425-15.
de Kievit, T. R., & Iglewski, B. H. (2000). Bacterial quorum sensing in pathogenic relationships. Infection and Immunity, 68(9), 4839–4849. https://doi.org/10.1128/IAI.68.9.4839-4849.2000.
Duerkop, B. A., Varga, J., Chandler, J. R., et al. (2009). Quorum-sensing control of antibiotic synthesis in Burkholderia thailandensis. Journal of Bacteriology, 191(12), 3909–3918. https://doi.org/10.1128/JB.00200-09.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Mohan, K.V., Sahu, P. (2018). Quorum Sensing in Microbes and their Function in Modulating Antibiotic Synthesis. In: Pallaval Veera Bramhachari (eds) Implication of Quorum Sensing System in Biofilm Formation and Virulence. Springer, Singapore. https://doi.org/10.1007/978-981-13-2429-1_11
Download citation
DOI: https://doi.org/10.1007/978-981-13-2429-1_11
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-13-2428-4
Online ISBN: 978-981-13-2429-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)