
Abstract
Head and neck cancer is the sixth most common malignancy worldwide and a major cause of death from cancer, with a median 5-year survival of around 50% (Suh et al. 2014). Over 90% are squamous cell carcinomas arising from the epithelial cells that line the mucosal surfaces of the head and neck. Tumour hypoxia is an important negative prognostic factor for head and neck squamous cell carcinomas (HNSCC), associated with resistance to radiotherapy and decreased overall survival (Bittner and Grosu 2013). Therefore, tumour hypoxia is an important phenomenon in the management of HNSCC and has been the focus of studies to improve treatment response for many years. This chapter aims to discuss some of the key mechanisms affected by tumour hypoxia and review strategies under investigation towards better detection and modulation of tumour hypoxia in order to improve treatment response and survival of head and neck cancer patients.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
- Vascular Endothelial Growth Factor
- Endoplasmic Reticulum Stress
- Unfolded Protein Response
- Hypoxic Tumour
- Formalin Fixed Paraffin Embed
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
3.1 Background
Tumour heterogeneity due to genetic and epigenetic variations within the tumours is an important contributor to phenotypic diversity of tumours and resistance to therapy (Catenacci 2014). In addition, tumour microenvironment influences phenotypic diversity or plasticity within the tumour (Fukumura and Jain 2007). Hypoxia is a fundamental biological phenomenon of solid tumours, which plays an important role in changing tumour microenvironment (Wouters et al. 2002). Hypoxia leads to cellular individuality and is associated with resistance to curative therapies and reduced survival in a number of tumour types including head and neck, pancreatic, cervical and prostate cancers (Buffa et al. 2010; Eustace et al. 2013). Tissue hypoxia occurs when the oxygen level in tissues falls below physiological levels resulting from the inadequate supply of oxygen to meet demand (Hockel and Vaupel 2001). It arises in tumours due to the high metabolic demand of rapidly proliferating cancer cells, and due to disruption in the blood supply. Tumours develop aberrant and chaotic microvasculature, which are structurally immature and leaky, resulting in ineffective oxygen delivery (Vaupel 2004). Fluid accumulation and increased tumour interstitial pressure restricts and compresses intratumoural blood vessels, further reducing the blood supply. Hypoxia leads to increased risk of tumour progression , metastases, resistance to anti-cancer therapies and recurrence due to a wide range of hypoxia-related consequences, resulting in metabolic and genetic changes that promote an aggressive tumour phenotype (Vaupel and Mayer 2007). Hypoxia drives tumour progression by selecting clones that can best adapt to the stress of inadequate perfusion and nutrient deprivation, which then expand and develop into a more malignant phenotype.
Hypoxia has been extensively investigated in HNSCC and its association with poor outcome is well known. However, it is difficult to assess, as intratumoural hypoxia is heterogeneous and dynamic, due to continuing tumour and vessel growth, and constant fluctuations in blood flow. Different biological effects are triggered at different levels of oxygen partial pressure (pO2) in tissues. There is no consensus for hypoxic thresholds in tumours but at pO2 < 25–30 mmHg radiosensitivity progressively decreases (Hockel and Vaupel 2001; Rademakers et al. 2008), and at pO2 < 10–15 mmHg changes in gene expression under the control of the hypoxia regulated transcription factor HIF-1 are demonstrated. At pO2 < 10 mmHg Vaupel et al. (1994) demonstrated intracellular acidosis and adenosine triphosphate (ATP) depletion in murine fibroblasts and this level represents a critical threshold for energy metabolism. There is also decreased protein synthesis and oxygen consumption to increase tolerance to hypoxia. At pO2 < 1 mmHg there is increased apoptosis, reduced oxidative phosphorylation and cells switch to glycolysis (also known as Warburg effect) to maintain adequate energy levels. Lower levels of pO2 of 0.2–1 mmHg lengthen G1 phase of the cell cycle or arrest cells in G1 and anoxia causes immediate cell cycle arrest (Hockel and Vaupel 2001).
Importantly, hypoxia is a transportable property. Studies performed using patient derived xenografts with differing degrees of hypoxia demonstrated that the amount of hypoxia in the xenograft derived from hypoxic tumours had similar oxygen distribution to the patient’s tumour. The hypoxic tumours grew more rapidly and when these human derived xenografts were serially propagated, they maintained the hypoxic feature (Chang et al. 2011). These findings suggest that the genetics of cancer influences the microenvironment within which the tumours are formed, the microenvironment determines tumour behaviour, and together they impact cancer phenotype.
3.2 Classification
Hypoxia can be broadly divided into two basic types: chronic and acute. However, there are no clear cutoff between the two types and there are typically mixed heterogeneous patterns throughout the tumour.
3.2.1 Chronic Hypoxia
Chronic, or diffusion-limited hypoxia , was first described by Thomlinson and Gray in 1955, and is caused by consumption of oxygen by cells close to the blood vessels, leaving an inadequate supply of oxygen and nutrients to the cells further away from the vessels. A distance of approximately 180 μm from vascularised stroma was shown to be the diffusion distance of soluble oxygen (Thomlinson and Gray 1955). It can also be caused by reduced oxygen content in the blood, such as in anaemia, which may be caused by tumour-related factors as well as anti-cancer therapy or carboxyhaemoglobin formation in heavy smokers, and by compromised perfusion of leaky microvessels. Chronic hypoxia usually occurs after several hours to days of hypoxia, long enough to induce changes in gene expression (Bayer et al. 2011).
3.2.2 Acute Hypoxia
Acute hypoxia , otherwise known as cycling, transient, intermittent, or perfusion-limited hypoxia, occurs because of fluctuations in the perfusion of tumour vasculature. This results in time periods of better or worse oxygenation of tumour areas. This type of hypoxia may be ischaemic due to transient occlusion of blood flow by blood clots or tumour cells, or hypoxaemic due to transient reduction in the oxygen content within microvessels (Harris 2002). Acute hypoxia may have greater effects than chronic hypoxia on the development of an invasive phenotype with hypoxia-reoxygenation episodes resulting in genomic instability, accelerated growth and metastases (Rofstad et al. 2007).
3.3 Biological Consequences
Tumour cells exposed to hypoxia activate many signalling pathways, which act in an integrated network affecting common downstream pathways. They work together to ensure adaptation to overcome the lack of oxygen and nutrients. They induce changes such as angiogenesis, glycolysis , inhibition of apoptosis and downregulation of cell adhesion molecules resulting in tumour cell detachment, all of which lead to the development and selection of more aggressive clones of tumour cells.
3.3.1 Metabolism
Glucose is the main source of energy for cells and enters the cell through a family of glucose transporters (GLUT1 to 4). Energy in the form of ATP is generated under aerobic conditions via glycolysis or the tricarboxylic (TCA) cycle. During glycolysis, glucose is metabolised to pyruvate in the cytosol to produce 2 ATPs from each molecule of glucose. Pyruvate enters the mitochondria and is oxidised by pyruvate dehydrogenase (PDH) to produce acetyl coenzyme A (CoA) , which enters the TCA cycle. Acetyl CoA is metabolised through a series of reactions generating nicotinamide adenine dinucleotide hydrogen (NADH) , a reducing agent used by the oxidative phosphorylation pathway in mitochondria. NADH passes electrons derived from the TCA cycle to the electron transport chain (ETC) in mitochondria, and combines with oxygen to produce water and a proton gradient that is used to generate 36 ATPs per glucose. In the mitochondria ETC, electrons transfer steps are catalysed at Complex I (NADH dehydrogenase or ubiquinone oxidoreductase) and III (coenzyme Q or cytochrome c reductase) and is transferred to O2 at Complex IV (cytochrome c oxidase). Each step is coupled with proton translocation across the inner mitochondrial membrane, driving ATP synthesis at Complex V. Electrons can escape the ETC and be captured by O2, forming reactive oxygen species (ROS) and resulting in oxidative stress (Sabharwal and Schumacker 2014).
Under anaerobic conditions the lack of oxygen to act as the final electron acceptor at complex IV in the ETC prevents aerobic metabolism from progressing. Pyruvate is not used in the TCA cycle, but converted to lactate in the cytosol by lactate dehydrogenase (LDH) to regenerate nictotinamide adenine dinucleotide (NAD+) from reduced NADH , which is required by glyceraldehyde-3-phosphate dehydrogenase (GAPDH) for additional cycles of glycolysis. PDH is phosphorylated and inactivated by pyruvate dehydrogenase kinase (PDK) (Lu et al. 2002).
Cancer cells have a high rate of glucose consumption even in the presence of oxygen, known as the Warburg effect or aerobic glycolysis (Warburg et al. 1927). ATP production via glycolysis is much faster but less efficient and cancer cells avidly consume glucose to meet their increased energy and biosynthesis needs. A constant high rate of glycolysis in tumour cells uses up the NAD+ pool and glycolysis cannot be sustained unless NAD+ is regenerated, which is achieved by increasing lactate production.
3.3.2 Regulation of Hypoxia Driven Pathways
The underlying mechanisms that promote hypoxia and influence the level of hypoxia in tumour are currently unclear. There are two fundamentally different concepts for hypoxia driven pathways . One concept describes that hypoxia provides an environment for the selection of mutant cancer cells that tolerate hypoxic conditions (Graeber et al. 1996). Further mutations allow the cell to develop tolerance mechanisms, giving rise to a population of cells resistant to the hypoxic microenvironment. One such mutation is the p53 gene, which has been shown to play an important role in tolerance to hypoxia; when p53 mutant cells were mixed with wild type cells and grown under hypoxic condition, only the p53 mutant cells could be selectively grown (Sermeus and Michiels 2011). The second mechanism is adaptation of normal physiological responses to hypoxia, for example by inducing factors such as hypoxia-inducible factors (HIFs) (Wouters and Koritzinsky 2008). There are a large number of oxygen sensitive enzymes and signalling pathways that are present in all cells, and hypoxia reduces the activity of enzymes that use oxygen as co-factor and leads to changes in cellular signalling.
The best understood pathway is hypoxia-inducible factor (HIF) pathway, which is stabilised due to the inhibition of the PHD (Prolyl Hydroxylase Domain oxygen sensor) enzymes (Rabinowitz 2013). The two other major cellular pathways which have been shown to be influenced by hypoxia include the kinase mammalian target of rapamycin (mTOR) pathway, which is an important moderator of metabolic signals, and the unfolded protein response (UPR), a cellular stress response related to the endoplasmic reticulum (Wouters and Koritzinsky 2008). Although they are activated independently a number of studies suggest that HIF, mTOR and UPR mediated responses to hypoxia act in an integrated fashion, both influencing each other and the common downstream pathways that affect gene expression, metabolism, cell survival, tumourigenesis, tumour growth and therapy resistance.
Below we briefly review the importance of these pathways in relation to hypoxia.
3.3.2.1 Hypoxia-Inducible Factor-1 Signalling Pathway
Tumour cell responses to hypoxia are initiated through activation of the hypoxia-inducible factor (HIF) family of transcription factors. The HIF family comprises the HIF-1α, HIF-1β, HIF-2α, and HIF-3α subunits, the most important of which is HIF-1 (Marxsen et al. 2004) HIF-1 is a heterodimeric protein consisting of an oxygen-regulated α subunit and a constitutively expressed oxygen-independent β subunit. The β subunit of HIF is a constitutive nuclear protein present in normoxic cells.
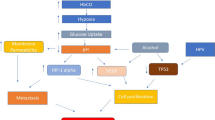
Under conditions of normal oxygen HIF-1α is rapidly degraded by the Von-Hippel-Lindau (VHL) tumour suppressor protein via the ubiquitin-proteasome pathway. The α subunits of HIF are hydroxylated at conserved proline residues P402 and P564 by propyl hydroxylase (PHD), which use oxygen as a co-substrate, resulting in recognition and ubiquitination by the VHL E3 ligase, labelling them for degradation by the proteasome. HIF-1α accumulation is also prevented under normoxia by factor inhibiting HIF-1 (FIH), which hydroxylates the transactivation domain of HIF-1α. This inhibits the binding of CREB binding protein (CBP) and p300 transcription co-factors to the HIF-1 complex (Fig. 3.1) (Semenza 2000).

HIF-1 activation pathway: a Under normoxia HIF-1α is hydroxylated by propyl hydroxylase and is degraded by VHL via the ubiquitin-proteasome pathway. b Under hypoxia propyl hydroxylase and FIH are inhibited and HIF-1α is stabilised, allowing heterodimerisation with HIF-1β and subsequent binding to the hypoxia-responsive elements in target genes. This results in the upregulation of proteins that promote cell survival. HIF-1α can expression can also be upregulated by EGFR and PI3 K signaling pathways. Figure adapted from Rademakers et al. (2008). Reproduced with permission from Elsevier
Hypoxia inhibits hydroxylation of HIF-1α by PHD and FIH, which results in stabilisation and accumulation of HIF-1α. HIF-1α translocates to the nucleus where it heterodimerises with constitutively expressed HIF-1β, and binds to the hypoxia-responsive elements (HREs) in the promotors of target genes. Genes that help overcome hypoxia, such as those involved in cell survival under oxidative stress, erythropoiesis, angiogenesis, glucose metabolism and pH regulation are upregulated, resulting in the upregulation of more than 100 proteins that promote survival and increased aggressiveness of hypoxic tumour cells (Semenza 2000).
Vascular endothelial growth factor (VEGF) is upregulated by HIF-1α in hypoxia and is the strongest inducer of angiogenesis, stimulating the proliferation of endothelial cells and the formation of new blood vessels. VEGF also acts as a tumour cell survival factor, inhibiting cell apoptosis by inducing the anti-apoptotic protein Bcl-2 (Ferrara et al. 2003).
Under hypoxia HIF-1α mediates a switch from oxidative to glycolytic metabolism (Lu et al. 2002). HIF-1α activation affects cellular glucose metabolism by inducing the transcription of genes involved in increased glucose uptake, such as the glucose transporters GLUT1 and GLUT3, and stimulation of glycolytic enzymes that breakdown intracellular glucose. HIF-1α induces lactate dehydrogenase A (LDHA), increasing the conversion of pyruvate to lactate. In addition, HIF-1α activates pyruvate dehydrogenase kinase (PDK), which inactivates pyruvate dehydrogenase (PDH), actively shunting pyruvate away from the mitochondria. This reduces flux through the TCA cycle, reducing delivery of NADH to the ETC. Mitochondrial function is therefore reduced, decreasing oxygen demand in hypoxic cells. In addition HIF-1α induces BCL2/adenovirus E1B 19 kDa interacting protein 3 (BNIP3) expression, which triggers mitochondrial autophagy, decreasing mitochondrial mass and oxygen consumption. 18HIF-1α also diverts pyruvate to be phosphorylated by hexokinase to glucose-6-phosphate, which is used in anabolic pathways such as the pentose phosphate pathway, by upregulating hexokinase. This pathway generates the precursors of nucleotides and amino acids required for the rapid tumour cell growth and proliferation. Hypoxia also triggers the generation of reactive oxygen species (ROS) by the ETC, which is released into the cytosol where they inhibit PHDs, resulting in stabilisation of HIF-1α (Chandel et al. 2000). Glycolytic metabolism results in acidosis of the tumour microenvironment due to increased lactate and carbon dioxide levels, which are actively expelled from the tumour cells into the extracellular matrix. HIF-1α upregulates monocarboxylate transporter 4 (MCT4), which removes lactate from the cell, carbonic anhydrase 9 (CA9), which catalyses the conversion of carbon dioxide released during the pentose phosphate pathway to carbonic acid, and sodium-hydrogen exchanger (NHE1), which maintains an alkaline intracellular pH and acidic extracellular pH. Acidosis induces the secretion of matrix-degrading hyaluronidase and metalloproteinases by tumour-associated fibroblasts, creating a tumour microenvironment favourable for invasion and migration (Fang et al. 2008). In addition it reduces anti-tumour immunity, inhibiting the activity of dendritic cells and T cells, and the efficacy of drug uptake by tumours, further driving tumour progression (Balamurugan 2015).
HIF-1α activation has many other downstream effects including the expression of several proteins important in epithelial-mesenchymal transition , preparing tumour cells for invasion and migration, such as SNAIL, SLUG, TWIST (Yang et al. 2008), and inhibition of E-cadherin (Krishnamachary et al. 2006). It has also been implicated in the regulation of cancer stem cells , through activation of NOTCH1, and tumour-mediated immune suppression through attenuation of T cell function (Doedens et al. 2010). HIF-1α is also involved in the activation of tumour-associated inflammatory signalling in conjunction with nuclear factor-kappa B (NF-κB) (Bruning et al. 2012), which activates genes that promote cell proliferation and cell survival, and STAT3, which also works with HIF1α to activate VEGF (Jung et al. 2005). In addition, miRNAs are also activated, with direct binding of HIF-1α to a HRE on the proximal miR-210 promotor (Huang et al. 2009).
Factors other than hypoxia can also activate HIF-1α, such as pyruvate and lactate, indicating a positive feedback mechanism. HIF-1α expression can also be upregulated under conditions of normoxia, such as through activation of the receptors of the tyrosine kinase family including EGFR and HER2/erbB2, activation of the PI3K-AKT-mTOR pathway, and genetic mutations including VHL inactivation, loss of PTEN, p16INK4a and p53 tumour suppressors (Span and Bussink 2015).
3.3.2.2 mTOR Signalling
The adaptive response to hypoxia involves inhibition of energy-intensive cellular processes including protein translation. This effect is mediated in part through a decrease in the kinase activity of mammalian target of rapamycin complex 1 (mTORC1) (Vadysirisack and Ellisen 2012). The mTOR pathway responds to microenvironment signals such as availability of growth factors, energy (ATP) and nutrients including lipids and amino acids. These factors influence upstream kinase signalling pathways that all converge upon mTOR and the outputs of mTOR drive changes in the metabolic rate of the cells (Wouters and Koritzinsky 2008). Activation of mTOR requires oxygenation as one of its inputs and low oxygenation due to hypoxia inhibits signalling through this pathway (Magagnin et al. 2008).
Mild to moderate hypoxia levels suppress mTOR through a HIF-1 independent pathway involving the activation of tuberous sclerosis protein 1(TSC1)-TSC2 complex via adenosine monophosphate-activated protein kinase (AMPK) or via transcriptional regulation of REDD1. Hypoxia reduces the phosphorylation and activation of 4EBP1, an important negative regulator of protein synthesis, by mTOR (Magagnin et al. 2008; Rouschop et al. 2011). mTOR is a principle regulator of autophagy under nutrient depleted conditions, which is regulated by BNIP3 and AMPK during hypoxia. In addition, mTOR interacts with HIF-1α, promoting the rate of HIF-1α mRNA translation. In tumour cells hypoxia has been shown to be less effective at inhibiting mTOR through mechanisms such as constitutive mTOR activity or loss of function of TSC2 or PTEN, the negative regulator of this pathway (Wouters and Koritzinsky 2008).
The shut off of protein synthesis through mTOR inhibition is important for survival of hypoxic condition. Paradoxically mTOR is an oncogene and many known negative regulators of mTOR such as TSC1, TSC2, LKB1 and PTEN signalling have been shown to function as bona fide tumour suppressors, yet hypoxic cells need to shut down this pathway to survive metabolic stress.
3.3.2.3 The Unfolded Protein Response (UPR)
Messenger RNA translation, post-translational modifications and protein folding takes place in the endoplasmic reticulum (ER). The folding of these proteins in ER requires disulphide bind formation which is an oxidation reaction (Nagelkerke et al. 2013).
Under conditions of stress, such as severe hypoxia, misfolding of proteins occur which accumulate in the ER, resulting in ER stress. This activates the unfolded protein response, which aims to relieve this stress and increase cell survival by activating the ER stress sensors such as protein kinase R-like endoplasmic reticulum kinase (PERK). PERK activation results in inhibition of global mRNA translation and protein synthesis to reduce the load of misfolded proteins and lower energy demands. It also upregulates the expression of genes involved in resistance to oxidative stress and amino acid metabolism to promote cell survival, and enhances HIF-1α transcriptional activity. ER-associated degradation and autophagy occur to remove misfolded proteins, further reducing ER stress (Pereira et al. 2014). ER stress also modulates the expression of VEGF, stimulating angiogenesis and promoting cell survival of rapidly growing cells. In addition VEGF signalling can activate the UPR in endothelial cells in the absence of ER stress via mTOR signalling, further promoting angiogenesis (Wang et al. 2012). The UPR is upregulated in tumours and is an important survival response in conditions of prolonged hypoxia, increasing the threshold for apoptosis. Tumours cells with abrogated PERK activity show significantly reduced clonogenic survival and decreased ability to tolerate moderate to extreme hypoxia, with higher levels of apoptosis in hypoxic areas (Bi et al. 2005).
3.3.3 Micro RNAs
Several studies have demonstrated through genome-wide miRNA profiling that miRNA expression in tumour cells is altered in response to hypoxia (Hebert et al. 2007; Hua et al. 2006; Kulshreshtha et al. 2007; Guimbellot et al. 2009; Bruning et al. 2011; Crosby et al. 2009). However due to differences in tumour cell lines, variations in exposure to hypoxia severity and duration, as well as differences in microarray hybridisation platforms resulting in different ranges of miRNAs screened, there has been limited overlap in the patterns of miRNA up- and downregulation. Next generation sequencing techniques have revealed a greater number of previously not identified miRNAs, highlighting the complexity of miRNA response to hypoxia (Camps et al. 2014).
MiR-210, however, has been robustly and consistently induced by hypoxia, in both normal and cancer cells. It has also been shown to be overexpressed in a variety of solid tumours and patient plasma or serum, including HNSCC, where it is generally associated with poor clinical outcome (Wang et al. 2014; Gee et al. 2010). MiR-210 is maintained at low levels in normoxia. Under hypoxic conditions miR-210 is induced independently and directly by HIF-1α through interaction with its promotor sequence, and targets genes involved cell cycle regulation, mitochondrial function, apoptosis, angiogenesis and metastases. MiR-210 promotes stabilisation of HIF-1α and inhibit PHDs preventing the degradation of HIF-1α (Chang et al. 2013).
MiR-210 targets the MYC antagonist MAX network transcriptional repressor (MNT), resulting in MYC activated cell cycle progression, and the cell cycle regulator E2F transcription factor 3 (E2F3), promoting proliferation and inhibiting apoptosis (Zhang et al. 2009). Apoptosis is also inhibited by targeting apoptosis-inducing factor mitochondrion-associated 3 (AIFM3), reducing caspase-dependent apoptosis, and caspase-8-associated protein 2 (CASP8AP2), an integral protein in the cleavage and activation of apoptosis initiator caspase-8 (Kim et al. 2009). MiR-210 has been found to bind to two sites in the 3′UTR of RAD52 mRNA, resulting in degradation. RAD52 is involved in the repair of double stranded DNA breaks and therefore miR-210 suppression of RAD52 increases genetic instability (Crosby et al. 2009). MiR-210 has a repressive effect on the mitochondrial ETC, aiding the metabolic shift to glycolysis in hypoxia. It targets and suppresses several integral components of the mitochondrial ETC such as cytochrome oxidase assembly protein-10 (COX-10), succinate dehydrogenase subunit D (SDHD), compromising electron transport and promoting glycolysis (Puissegur et al. 2011). In addition miR-210 suppresses iron-sulphur cluster scaffold proteins (ISCU), which catalyse the assembly of iron-sulphur clusters that are critical for enzymes in the TCA cycle and for the function of ETC complexes I, II and III (Chan et al. 2009). Angiogenesis is stimulated by the suppression of ephrin-A3 (EFNA3), which stimulates normoxic endothelial cell to form capillary-like structures (Fasanaro et al. 2008). In addition vacuole membrane protein 1 (VMP1), which inhibits cell migration and invasion, is suppressed, inducing a more metastatic phenotype (Ying et al. 2011).
MiR-210 is also reported to have a tumour suppressor role, inducing cell cycle arrest through fibroblast growth factor receptor-like 1 (FGFRL1) (Zuo et al. 2015) and apoptosis through suppression of anti-apoptotic Bcl-2 expression (Chio et al. 2013). As hypoxia can influence both cell death and survival the role of miR-210 is likely to depend on the cell type, extent and duration of exposure to hypoxia, and reflect the adaptation of tumour cells to a hostile environment.
3.4 Clinical Consequences in HNSCC
3.4.1 Resistance to Radiotherapy
Radiosensitvity rapidly decreases when the pO2 in a tumour is less than 25–30 mmHg (Hockel and Vaupel 2001). In 1953 Gray et al. (1953) determined the radiation dose required to achieve the same biological effect is 2.8–3 times higher in the absence of oxygen than presence. When ionising radiation is absorbed in tissue, free radicals are produced as a result of ionisations either directly in the DNA itself, or indirectly in other molecules such as water. Free radicals break chemical bonds and initiate a chain of events that results in DNA damage. Oxygen molecules are able to react with the free radicals and create a stable change in the chemical composition of the DNA damage. Oxygen therefore enhances the damage by “fixing” or making the damage permanent. In the absence of oxygen the target is chemically restored and the damage is repaired. In order for oxygen to consolidate radiation-induced damage it needs to be present at the time of irradiation or within a few milliseconds (Howard-Flanders and Moore 1958). Other hypoxia-induced mechanisms also contribute to radioresistance including glycolytic metabolism and lactate accumulation, which act as antioxidants and scavenge free radicals (Brizel et al. 2001; Sattler et al. 2010).
The changes in hypoxia heterogeneity during a course of fractionated radiotherapy (RT) can affect the response. During the early stages of treatment cell density decreases as the well-oxygenated tumour cells are killed. This results in reduced intratumoural pressure and increased vascular density leading to tumour reoxygenation (Bussink et al. 2000). This reoxygenation does not increase HIF-1α degradation, but the increase in ROS formation stabilises HIF-1α and increases the expression in a hypoxia-independent manner (Harada et al. 2009). This, together with accelerated repopulation that occurs at the later stages of RT, means that the entire cumulative dose of RT needs to be delivered.
3.4.2 Resistance to Chemotherapy
Some chemotherapeutic agents, such as etoposide, are dependent on the presence of oxygen for maximum cytotoxic effect (Wozniak et al. 1984). The blood vessels in hypoxic tumours are disordered and leaky, preventing efficient delivery of chemotherapy drugs. Hypoxic cells are distant from blood vessels and therefore not adequately exposed to drugs (Tannock 1998). Alkylating agents, such as cisplatin, and anti-metabolites, such as 5-FU, act during DNA synthesis by damaging DNA and initiating apoptosis. Cellular proliferation and DNA synthesis decreases as a function of distance from blood vessels and hypoxia selects for cells that have lost sensitivity to p53 mediated apoptosis, reducing the cytotoxic effect. Hypoxia upregulates genes involved in drug resistance, such as multidrug resistant 1 (MDR1) leading to the increased expression of P-glycoprotein, which is associated with tumour resistance to chemotherapeutics (Comerford et al. 2002). The acidic tumour microenvironment creates a pH gradient between the tumour cell and extracellular space, inhibiting the accumulation of drugs that are weak bases, such as doxorubicin (Skovsgaard 1977).
3.5 Therapeutic Approaches to Overcome Hypoxia Radioresistance in HNSCC
Hypoxia represents a potential target for therapy. Various strategies have been used to overcome hypoxia in HNSCC, summarised in Table 3.1. A meta-analysis of 32 clinical trials of various strategies of hypoxic modification demonstrated a small but significant improvement in locoregional control (LRC), disease specific control (DSS) and overall survival (OS) in HNSCC treated with primary RT (Overgaard 2011). Hypoxia stratification is required to identify patients at risk of treatment failure but also patients with normoxic tumours, who do not gain benefit from additional treatment, and can avoid additional toxicity.
3.5.1 Improving Haemoglobin (Hb) Levels
Many studies have demonstrated that low haemoglobin level prior to treatment associated with significant reduction in survival and increase in locoregional failure in HNSCC after RT or chemoradiation (Fortin et al. 2008; Lee et al. 1998). Over 1000 patients pooled from two studies demonstrated that high Hb was significantly associated with higher OS and the administration of blood during therapy increased Hb level. However blood transfusions had no impact on LRC, DSS or OS (Hoff et al. 2011). Exogenous erythropoietin administration to stimulate red blood cell production and correct anaemia has been investigated. However this did not improve tumour control or survival as demonstrated in 2 meta-analyses, but suggested an increase in mortality (Bohlius et al. 2009; Lambin et al. 2009).
3.5.2 Increasing Oxygen Availability
Hyperbaric oxygen is the administration of 100% oxygen at higher than normal atmospheric pressure. This elevated pressure results in increased pO2 in tissues and has been delivered in hyperbaric oxygen chambers with RT. This strategy has been shown to significantly improve tumour control and mortality, but also causes an increased rate of severe radiation tissue injury and chance of oxygen toxic seizures during therapy (Bennett et al. 2008).
Carbogen (95% oxygen and 5% carbon dioxide) breathing before RT did not improve tumour control (rubin 1979 carbogen), but combined with accelerated radiotherapy and nicotinamide (ARCON, accelerated radiotherapy with carbogen and nicotinamide) has been shown to result in high LRC in advanced larynx and oropharynx SCC (Kaanders et al. 2002). This combination aims to limit clonogenic repopulation during therapy by reducing the overall duration of RT by delivering more than one fraction a day, decrease diffusion-limited hypoxia with inhalation of carbogen, and decrease perfusion-limited hypoxia by administering nicotinamide, a vasoactive vitamin B6 analogue. A more recent study compared accelerated RT with ARCON in larynx SCC and did not demonstrate a benefit. However, a subgroup of patients had their hypoxic status assessed with pimonidazole IHC, which stains regions of hypoxia, and significant improvement in regional control and disease free survival (DFS) was found in the high pimonidazole staining group treated with ARCON (Janssens et al. 2012).
3.5.3 Radiosensitisers
Cellular sensitivity to RT can be improved by increasing the apparent cellular pO2 levels using radiosensitisers. Nitromidazoles are chemical compounds that mimic the radiosensitising effect of oxygen by inducing and stabilsing free-radical mediated double-stranded DNA breaks. The benefit of the addition of a nitroimidazole compound to RT was carried out in many studies, without clinical benefit (Lee et al. 1995; Van den Bogaert et al. 1995). However these studies used the older generation of the drug which is limited by neurotoxicity. Overgaard (1994) carried out a meta-analysis of over 7000 patients treated with nitroimidazole-based radiosensitisers in various tumour types of unknown hypoxia status and found an improvement in LRC and OS, with the greatest benefit in HNSCC. This was followed by the DAHANCA 5 study which randomized over 400 HNSCC patients to receive RT with nimorazole or RT with placebo. LRC and DSS were significantly higher with the addition of nimorazole (Overgaard et al. 1998). A phase II study investigated accelerated RT with nimorazole and cisplatin in 227 patients with locally advanced HNSCC and recently reported a high 5-year OS of 75% (Bentzen et al. 2015).
3.5.4 Hypoxic Cytotoxins
This group of drugs, of which tirapazamine is the most widely investigated, have a direct cytotoxic effect on hypoxic cells. Tirapazamine (TPZ) is a prodrug that requires one-electron reduction to a radical by intracellular reductases. In normoxia the unpaired electron in the radical is rapidly transferred to molecular oxygen, forming superoxide and regenerating the initial prodrug. In hypoxic conditions the radical accumulates and induces DNA damage, in part due to poisoning of topoisomerase II (Brown 1993).
TPZ combined with cisplatin and RT in HNSCC has shown to be well tolerated in phase I and II studies (Lee et al. 1998; Rischin et al. 2001). Rischin et al. carried out a phase II randomised trial of this combination versus cisplatin and RT and demonstrated a trend towards improved 3-year survival with acceptable toxicity (Rischin et al. 2005). However, further studies have not shown a benefit in survival or quality of life from the addition of TPZ (Rischin et al. 2010). Rischin et al. also carried out a substudy of 45 patients who had their tumour hypoxic status assessed by 18F-fluoromisonidazole, a hypoxic radiotracer, PET scans. Hypoxic patients performed poorly in the absence of TPZ augmentation, whereas the rate of complete response in the hypoxic group receiving TPZ was nearly as high as the rate for normoxicchemoRT patient. Patients with hypoxic tumours were more likely to develop distant metastatic disease compared with normoxic patients (Rischin et al. 2006).
3.5.5 Hypoxia Image-Guided Radiotherapy
Intensity modulated radiotherapy (IMRT) is used in the treatment of HNSCC. The intensity of the radiation beam can be modulated to reduce doses to normal structures without compromising the doses to the tumour. Tumour recurrence after compete response to radiotherapy has been shown to occur predominantly in high radiation dose regions, implying that tumours have radioresistant subvolumes, potentially due to hypoxia, within this region (Hendrickson et al. 2011). IMRT dose painting is a method of assigning different dose levels to structures within the same treatment fraction, resulting in the potential for higher total doses to selected targets (Galvin and De Neve 2007). Dose painting requires functional imaging such as positron emission tomography to direct regions for dose escalation. Intensification of intratumoural subvolumes of hypoxia has been investigated as a strategy to improve outcome in RT planning studies and radiobiological modelling studies only as this requires accurate and stable identification of hypoxia to guide target volume delineation.
3.5.6 Targeted Therapies
3.5.6.1 Targeting Angiogenesis: VEGF Receptors
High VEGF expression levels has been associated with worse overall survival in HNSCC (Kyzas et al. 2005). As tumour vasculature is abnormal, increased VEGF expression and microvascular density does not lead to increased blood flow and oxygen delivery. Targeting VEGF is a strategy to improve oxygenation by normalising the tumour vasculature and thereby increase response to RT. RT can lead to stimulation of angiogenesis through upregulation of VEGR and therefore anti-angiogenic therapy has been tested with RT (Koukourakis et al. 2001). Bevacizumab is a humanised monoclonal antibody against VEGF approved for use in colorectal cancer, and has been investigated in phase II studies concomitant with chemoRT in HNSCC with encouraging results (Fury et al. 2012). It inhibits endothelial cell growth and function, disrupting the formation of capillary networks, and has been shown to enhance RT response in HNSCC xenograft models (Hoang et al. 2012). Anti-angiogenic agents can improve tumour oxygenation by reducing the number of oxygen consuming endothelial and tumour cells, and the number of inefficient blood vessels. This also results in less leaky vessels and decreased interstitial pressure, improving perfusion (Willett et al. 2004).
3.5.6.2 Targeting HIF-1
Targeting HIF-1 is an attractive therapeutic strategy to overcome the effects of hypoxia. Drugs that inhibit HIF-1 activity are already in clinical use, such as the topoisomerase I inhibitor topotecan and mTOR inhibitors which decrease HIF-1 synthesis, the anthracycline doxorubicin which suppresses HIF-1 DNA binding, and the proteasome inhibitor bortezomib, which reduces HIF-1 transactivation (Semenza 2010). Newer drugs have been developed, such as PX-478 that inhibits HIF-1 by inhibiting HIF-1 mRNA translation and HIF-1 dequbiquitination and is being investigated as a phase I trial. In HNSCC tumour models this drug provided significant radiosensitisation of hypoxic cell lines and xenografttumours (Schwartz et al. 2009).
3.6 Methods to Detect Hypoxia in HNSCC
Accurately detecting and quantifying hypoxic tumours is essential in identifying patients who have aggressive, treatment resistant disease and has been one of the limiting factors for translating hypoxia-modification strategies into routine practice.
3.6.1 Clinical Studies Using Eppendorf polarographic Needle Electrodes
Approximately 50–60% of human tumour contain hypoxic and/or anoxic tissue regions that are heterogeneously distributed within the tumour (Vaupel et al. 2007). Many clinical studies, mainly in HNSCC and cervix cancers, using Eppendorf polarographic needle electrode histography have directly demonstrated that tumour hypoxia predicts for decreased local control, increased disease recurrence and reduced overall survival. This technique involves inserting an electrode into multiple sites in a tumour and measuring the pO2 at several points per needle track and data from all tracks form a histogram. In head and neck cancers the pO2 measurements between the primary tumour and metastatic lymph nodes have shown little difference, and nodal measurements have been used to represent the hypoxic status of the patient (Becker et al. 1998).
In head and neck cancer, Gatenby et al. (1988) in 1988 measured the oxygen tension in 31 lymph node metastases from HNSCC patients, and demonstrated a significant relationship between low mean pO2 measurements and poor response to radiotherapy. The volume of tumour containing low oxygen levels was also found to be important. Nordsmark et al. (1996) measured the pretreatment oxygenation status initially in 34 lymph nodes and 1 primary using the Eppendorf polarographic needle electrode histography and evaluated the tumour oxygenation status as the percentage of pO2 values ≤2.5 mmHg (hypoxic fraction 2.5, HF2.5). The median HF2.5 of 15% was used to define hypoxia and these patients had significantly poorer locoregional control (LRC). This was confirmed by the same group in a further 35 patients (Nordsmark and Overgaard 2000). Brizel et al. (1997, 1999) assessed 63 patients for pretreatment tumour oxygenation using the primary site or metastatic lymph node. A preliminary study had shown significantly worse disease free survival (DFS) at 12 months for patients with a median pO2 < 10 mmHg, and this was used to define hypoxic tumours. Whether the measurement was taken at the primary or nodal site did not affect the DFS and hypoxic tumours had significantly worse 3 year LRC, DFS and overall survival (OS), independent of tumour stage. However, a small study of 25 patients did not find an association between pretreatment oxygen levels and LRC or OS (Adam et al. 1999).
Stadler et al. (1999) identified the importance of the hypoxic subvolume, defined as the percentage of pO2 values <5 mmHg multiplied by the total tumour volume. Fifty-nine HNSCC primaries and nodes were assessed and the median pO2 was 16 mmHg, with no difference between the sites. On multivariate analysis the hypoxic subvolume and the HF5 were significant prognostic factors for survival. This was confirmed in a further 125 patients, where hypoxic tumour volume was a strong and independent prognostic factor for survival (Dunst et al. 2003). Rudat et al. (2000) evaluated the repeatability and predictive value of pO2 Eppendorph electrode histography in HNSCC. High variability was seen in patients who had two repeated independent measurement of the same tumour. In 41 patients with follow up data, locally advanced HNSCC the fraction of pO2 ≤ 2.5 mmHg was a significant prognostic factor for survival.
A joint analysis of prospectively collected Eppendorf pO2 measurements from multiple centres was performed, which consisted of 397 patients with HNSCC (Nordsmark et al. 2005). Median tumour pO2 was 9 mmHg and multivariate analysis demonstrated that HP2.5 greater than population median (19%) was associated with poorer OS at 3 years, providing strong evidence that tumour hypoxia has a significant role in HNSCC. Table 3.2 summaries the clinical data from HNSCC.
The use of Eppendorf oxygen electrode histography has been crucial to demonstrate the adverse effects of hypoxia on outcome in the clinical setting. It has the advantage of directly measuring absolute pO2 values and has prognostic potential. However, it has not been incorporated into routine clinical practice as it is an invasive procedure restricted to accessible tumours. It also requires an experiences operator but still exhibits large inter-observer variability (Nozue et al. 1997), and only provides information on the areas sampled. In addition it cannot differentiate between tumour and normal tissues (Vaupel et al. 2007). Other methods that have been investigated are summarised in Table 3.3.
3.6.2 Exogenous Markers of Hypoxia
The most commonly used exogenous markers of hypoxia are 2-nitroimidazole compounds such as pimonidazole and pentafluorinatedetanidazole (EF5) (Evans et al. 2000; Raleigh et al. 1998). They are irreversibly bioreduced by cellular nitroreductases and bind to thiol-containing proteins in viable hypoxic cells, forming stable adducts (Varghese et al. 1976). These adducts can then be detected by IHC staining with specific antibodies. IHC staining can quantify hypoxia by visually or digitally estimating the fraction of stained cells and provides hypoxic measurements with high spatial resolution. Binding is commonly seen adjacent to regions of necrosis and at a distance from blood vessels, in keeping with diffusion-limited hypoxia (Evans et al. 2000).
2-nitroimidazoles bind to tissue at a pO2 level of <10 mmHg and are more sensitive at lower pO2 levels than microelectrodes (Raleigh et al. 1999). The amount of bound marker is dependent on oxygen, the accumulation rate of individual hypoxic cells and intact nitroreductase enzymes in viable hypoxic cells (Ljungkvist et al. 2007). Differences in the activity of the enzymes can affect level detected and the accumulation rate of pimonidazole is dependent on pH (Dennis et al. 1985). The main disadvantage to exogenous markers of hypoxia is that they have to be administered intravenously prior to tissue biopsy or surgical resection, to allow fixation of the adducts that form in hypoxic regions. Pimonidazole binding has shown good correlation with Eppendorf electrode measurements in mouse models (Raleigh et al. 1999), but not in the clinical setting in cervix cancers (Nordsmark et al. 2003). Neither pimonidazole nor Eppendorf electrode pO2 were not prognostic for outcome in 127 cervix cancer patients (Nordsmark et al. 2006).
The prognostic value of 2-nitroimidazoles has been shown in HNSCC. High pimonidazole binding in 43 biopsies was significantly associated with LRC and DFS at 2 years. This association was lost in the subgroup treated with ARCON, suggesting a predictive role for pimonidazole (Kaanders et al. 2002). Pretreatment hypoxia using EF5 binding was investigated in 22 HNSCC patients and severe hypoxia, defined as maximum 30% EF binding which approximated to a pO2 of 0.76 mmHg, was associated with shorter event free survival at 3 years (Evans et al. 2007).
3.6.3 Endogenous Markers of Hypoxia
Endogenous markers are genes or gene products that are specifically upregulated under hypoxic conditions and can be measured on tumour specimens. HIF-1α, GLUT1 and 3, CA9 and osteopontin are detected by antibody staining and immunohistochemistry and can be assessed on formalin fixed paraffin embedded (FFPE) archival material, allowing correlation with outcomes. However the techniques of tissue processing and staining analysis have not been validated between different laboratories, limiting comparisons and clinical applications. Samples are usually from small biopsy specimens and therefore not representative of the hypoxia heterogeneity of entire tumour. In addition their expression may be affected by the type of hypoxia, for example HIF-1α is induced and repressed rapidly and reflect acute changes in hypoxia, whereas CA9 is slow to accumulate (Sobhanifar et al. 2005). In general, studies using these markers show conflicting results, and the association between these markers and oxygen status of tissue is weak.
3.6.3.1 HIF-1α
HIF-1α is overexpressed in a wide variety of tumours. In HNSCC many studies have demonstrated an association with poor survival and poor response to chemoRT (Aebersold et al. 2001; Koukourakis et al. 2002), and has been found to be an independent prognostic factor in HNSCC. However there are also studies that show no association with prognosis (Kyzas et al. 2005; Wachters et al. 2013) and in one study of in surgically resected cohort HNSCC, HIF-1α expression was associated with significantly better DFS and OS (Beasley et al. 2002).
3.6.3.2 Glucose Transporters (GLUT)
These glucose transporters are present in normal tissue but upregulated in tumours due to increased glucose requirements in anaerobic glycolysis (Rademakers et al. 2011). In HNSCC GLUT-1 has been associated with poor treatment outcome in multiple retrospective series (Mineta et al. 2002; Oliver et al. 2004). In 58 patients treated with ARCON GLUT-1 was associated with an increased rate of distant metastasis and worse OS but GLUT-3 was associated with better LRC (Jonathan et al. 2006). Co-expression of HIF-1α and GLTU-1 significantly correlated with an increased risk of tumour-related death (Eckert et al. 2011).
3.6.3.3 Carbonic Anhydrase 9 (CA9)
CA9 is a transmembrane enzyme and a downstream target of HIF-1. It catalyses the reversible hydration of carbon dioxide to carbonic acid and is involved in pH regulation. Elevated expression has been demonstrated at pO2 < 20 mmHg (Wykoff et al. 2000). Overexpression of CA9 has been found in different types of cancer including HNSCC. Kaanders et al. (2002) showed that the expression of CA9 in HNSCC demonstrated some overlap in the distribution pattern as pimonidazole, with increasing signal at greater distance from blood vessels, but overall the correlation was found to be weak and not associate with outcome. Greater CA9 expression was seen at shorter distances from the vessels, suggesting that upregulation may occur at pO2 levels higher than that required for pimonidazole (Kaanders et al. 2002). CA9 shows conflicting results as a prognostic marker. Strong CA9 expression was found to be related to poor complete response rate after treatment with chemoRT (Koukourakis et al. 2001), but positivity was associated with better LRC and freedom from distant metastases in 58 patients treated with ARCON (Jonathan et al. 2006), or no association with LRC (Nordsmark et al. 2007). In the DAHANCA 5 study patients 320 samples were available for CA9 staining and the expression of CA9 did not correlate with any of the tumour or patient characteristics, was not a prognostic marker and did not correlate with nimorazole treatment (Eriksen et al. 2007).
3.6.3.4 Osteopontin (OPN ) in Tissue and Plasma
Osteopontin (OPN ) is an extracellular matrix-associated integrin-binding glycoprotein protein induced by hypoxia, initially identified in non-collagenous bone matrix. Binding of OPN to cell surface receptors on tumour cells activates integrins and matrix metalloprotein signalling pathways, increasing the risk of tumour invasion and migration (Chien et al. 2009). It is also involved in angiogenesis, promotes cell survival through PI3K/AKT and JAK/STAS3 signalling and regulation of NF-κB. It is upregulated through AKT activation, independent of HIF-1 under hypoxia (Ahmed et al. 2011). High OPN levels in both tissue and plasma have been correlated with low tumour pO2 in HNSCC and outcome (Nordsmark et al. 2007). A retrospective study of the DAHANCA 5 trial patients demonstrated that high OPN plasma levels was associated with higher locoregional tumour failure and disease specific mortality (Overgaard et al. 2005). Patients with high OPN levels who received nimorazole had better outcomes compared with patients with high OPN levels who did not receive nimorazole. No effect was seen in the low/intermediate OPN group, suggesting that high concentrations of OPN could predict clinically relevant hypoxia and identify patients who may benefit from hypoxic modification. Lim et al. (2012) investigated the predictive potential of OPN in patients who received tirapazamine but did not find that high levels were associated with poor prognosis and found no interaction between OPN and treatment. The difference in results compared with the DAHANCA study may be due to differences in treatment protocols, but also lack of standardisation for OPN thresholds between the studies.
3.6.3.5 Gene Expression Signatures of Hypoxia
As hypoxia is influenced many biological pathways, a single marker is incapable of adequately describing this complex heterogeneous response. To improve hypoxia specificity combining several markers in gene expression signatures of hypoxia have been investigated. Different methods have been used to derive clinically applicable hypoxic gene signatures and are summarised in Table 3.4.
In vitro derived gene sets have been described by culturing tumour cell lines under normoxic and hypoxic conditions, then assessing the differences in gene expression. Koong et al. (2000) used FaDu HNSCC and SiHa cervical SCC cell lines to identify 9 genes exhibiting greater than 3-fold induction under hypoxic conditions when using the expression of VEGF as a cut off point for assessing hypoxia-induced genes. An 84 upregulated gene signature was generated by culturing normal and transformed keratinocytes at various different oxygen levels, which could be grouped into 6 functional groups, including as metabolism/transport, angiogenesis, tissue remodelling, apoptosis, proliferation/differentiation and gene expression (Denko et al. 2003).
Chi et al. (2006) evaluated the gene expression in response to hypoxia in several primary cell lines in vitro using cDNA microarrays and found a wide variation in the response between cell and tissue types. However, a 253 gene signature was identified which were concordant with gene expression data from a distinct subset of renal tumours, allowing classification by hierarchical clustering into 2 groups with high or low expression of the hypoxia response genes. The high expression group consisted of clear cell renal carcinomas, which typically have loss of function al VHL proteins, and the low group were other histological subtypes or normal samples. Similar classification grouped breast cancers into ductal adenocarcinomas (high expression) and fibroadenomas or normal samples (low expression). In addition, the gene signature was a strong predictor of clinical outcome in independent breast and ovarian cancer datasets.
Human mammary epithelial cells were culture under hypoxic conditions at early and late time points to assess the time dependency of hypoxia-regulated gene expression (Seigneuric et al. 2007). The early response gene signature was characterised by genes related to growth, apoptosis, insulin and oestrogen receptor signalling, whereas the late response was characterised by genes involved in angiogenesis, glucose transport, proliferation, metastasis and apoptosis, and were similar to the genes identified by Chi et al. The early but not late signature was prognostic in univariate analysis, but not maintained in multivariate analysis in a breast cancer dataset. Sorensen et al. (2010) exposed 4 head and neck cell lines to different oxygen concentrations at normal or low pH, and analysed the gene expression analysed to identify genes upregulated by hypoxia, independent of pH. Hypoxia induced pH independent genes were selected if they were common in 3 of the 4 head and neck cell lines, resulting in a 27 gene signature.
Winter et al. (2007) developed a hypoxia metagene from 59 HNSCC fresh frozen samples by clustering around the mRNA expression of 10 well-known hypoxia-regulated genes, such as CA9, GLUT1 and VEGF. Strongly correlated upregulated genes appearing in >50% of clusters defined a signature comprising of 99 genes, which was found to be an independent prognostic factor for RFS in an independent head and neck cancer and breast cancer dataset. The same group derived a common hypoxia metagene consisting of 51 genes by selecting genes that were consistently co-expressed with previously validated hypoxia-regulated genes (Buffa et al. 2010). By applying more training sets and co-expression networks a reduced metagene had prognostic significance in 4 independent datasets of breast, lung and head and neck cancers, and outperformed larger published signatures. The top 26 genes from this signature were retrospectively assessed in FFPE samples from laryngeal and bladder cancer patients treated with ARCON or CON (carbogen and nicotinamide) respectively (Eustace et al. 2013). Tumours were categorised into high and low hypoxia groups, and laryngeal tumours in the high hypoxia group showed greater benefit from ARCON than the low hypoxia group. The hypoxia signature did not predict benefit from CON in bladder cancer.
In contrast to previous studies on hypoxia gene expression, Toustrup et al. (2011) developed a classifier based on the hypoxic status of tumours. The hypoxia induced pH independent gene profile developed by Sorensen et al. was validated in vivo in a xenograft study using 18F-FAZA as an exogenous hypoxia radiotracer in autoradiographic studies. Hypoxic, non-hypoxic and mixed heterogeneous tumour areas defined by 8F-FAZA positive and negative regions were demarcated and dissected. All the genes investigated were significantly upregulated in hypoxic tumour areas compared with non-hypoxic areas, and all but 3 were upregulated in samples from mixed heterogenous versus non-hypoxic areas. To identify the most informative genes, a training set of 58 hypoxia-evaluated HNSCC FFPE biopsies were analysed for gene expression. The oxygenation status of these tumours had previously been evaluated in accordance to the relative number of oxygen electrode measurements less than 2.5 mmHg in their metastatic cervical lymph nodes. A 15-gene expression classifier was generated containing 15 of the in vitro identified hypoxia-responsive genes, which could best discriminate between ‘more’ and ‘less’ hypoxic human HNSCCs. This was evaluated in an independent data set, where patients with HNSCC were randomised to receive either hypoxic modification with nimorazole or placebo concomitant with radiotherapy. Patients with ‘more’ hypoxic tumours defined by the classifier had significantly higher cumulative incidence of locoregional failure at 5 years compared with those with less hypoxic tumours. Within the ‘more’ hypoxic group, patients treated with nimorazole had significantly reduced incidence of failure compared with placebo, whereas in the ‘less’ hypoxic group there was no significant difference in outcome, suggesting the classifier potentially has predictive value. Further subgroup analysis demonstrated that this benefit was only found in HPV-negative ‘more’ hypoxic tumours and not HPV-positive, whose outcome was unaffected by hypoxic modification (Toustrup et al. 2012). The same frequencies of ‘more’ and ‘less’ hypoxic tumours were in HPV-negative and positive groups as assessed by the classifier. More recently the classifier was used to assess 55 patients recruited in a randomised trial accelerated RT with or without nimorazole (Hassan Metwally et al. 2015). RT alone in 16 patients with ‘more’ hypoxic tumours was associated with higher locoregionaltumour failure.
More recently further investigation into the time dependent gene expression changes in response to hypoxia was carried out using prostate, colon and breast cancer cell lines exposed to hypoxia at 8 different time points between 0 and 24 h (Starmans et al. 2012). Seven different signatures consisting of induced genes with distinct temporal profiles and 2 general hypoxia signatures were generated, but none were prognostic in a large breast cancer cohort. In contrast previously published in vivo derived signatures showed clear prognostic power, suggesting the importance of the tumour microenvironment in the response to hypoxia.
Classification of hypoxia gene expression has the potential to represent prognostic and predictive markers in cancer and has been demonstrated retrospectively to be applicable to unselected FFPE biopsy samples, and therefore translatable into clinical practice. However, the 2 smaller gene signatures (15 and 26 gene signatures) which have demonstrated predictive value, have only 4 genes that overlap, suggesting that this method alone may not identify all hypoxic tumours. Prospective validation in clinical studies is required before further conclusions can be made.
3.6.4 Magnetic Resonance Imaging (MRI)
3.6.4.1 Dynamic Contrast-Enhanced MRI (DCE MRI)
In this technique fast repeated images are acquired before, during and after the rapid administration of a small hydrophilic gadolinium-based contrast agent, which diffuses through blood vessel walls and distributes into the extracellular space. The change in signal intensity reflects tumour perfusion, vessel permeability and the volume of extracellular space (Turkbey et al. 2010). It does not correlate directly with tissue pO2 levels but a small study in HNSCC found positive correlation between imaging parameters of poor perfusion and pimonidazole staining (Newbold et al. 2009), and with hypoxia defined by 18F-fluoromisonidazole PET uptake in nodal metastases (Jansen et al. 2010).
3.6.4.2 Blood Oxygen Level-Dependent MRI (BOLD MRI)
This is an indirect measure of visualising pO2 in blood vessels and surrounding tissue based on the paramagnetic properties of deoxyhaemoglobin, which is related to tissue oxygenation. It does not provide quantitative information on oxygen concentration and the signal can be influenced by many factors such as blood flow and pH. However, studies have shown correlation between this technique and polargraphic electrode pO2 measurements and pimonidazole staining in prostate cancer, suggesting BOLD MRI may provide complementary information related to tissue oxygenation (Chopra et al. 2009).
3.6.5 Positron Emission Tomography (PET)
PET is a molecular imaging technique that can visualise and quantify tumours and their microenvironment. Different biological tumour characteristics can be imaged depending on the radiotracer used. Several hypoxia-specific PET radiotracers have been developed and extensively investigated as it is a potentially useful non-invasive technique for identification, quantification of hypoxia and repeated measurements after intervention. The metabolic activity demonstrated by the commonly used radiotracer 18Fluorine-fluororodexoxyglucose (18F-FDG) is indirectly related to the proliferative activity and oxygenation status of tumours, but cannot reliably distinguish hypoxic tumours. A good hypoxia tracer should be able to detect pO2 levels that are clinically relevant to therapy, and be able to distinguish between normoxia, hypoxia, anoxia and necrosis. The molecule should be small and lipophilic making it highly membrane permeable leading to rapid uptake into cells and rapid clearance from normoxic cells to allow a high tumour to background contrast. It should also have a good dosimetry profile with simple radiolabelling and production, and low radiation dose to the patient. Two main classes of hypoxia imaging radiotracers are in available in the clinical trial setting: one based on the nitroimidazole compounds and one based on a complex of copper with diacetyl-bis (N4-methylsemicarbazone) (ATSM) ligands. A meta-analysis of published hypoxia imaging studies, which included PET and other imaging modalities of hypoxia, showed a uniform tendency for poor response to radiotherapy in tumours showing a higher uptake despite the heterogeneity of the image acquisition, data analysis and treatments (Horsman et al. 2012).
3.6.5.1 F-Fluoromisonidazole (18F-FMISO )
18F-FMISO is a derivative of the nitroimidazole group of compounds and is the most widely studied hypoxic radiotracer. It is moderately lipophilic and enters cells by passive diffusion across the cell membrane. The nitro group of the imidazole ring structure (R-NO2) is reduced by intracellular reductases to R-NO2 −, which is reoxidised in the presence of oxygen and the tracer can flow back into the extracellular compartment. In conditions of hypoxia R-NO2 − can be further reduced with progressive production of R-NH2 compounds that bind covalently to intracellular molecules and the tracer becomes trapped. This occurs at pO2 < 10 mmHg and therefore 18F-FMISO detects clinically relevant hypoxia. The process requires viable cells with functional nitroreductases and the tracer does not accumulate in necrotic cells (Prekeges et al. 1991).
3.6.5.2 Clinical Studies of 18F-FMISO in HNSCC
Rajendran et al. (2006) reported the largest study 18F-FMISO, which involved 73 patients with HNSCC. Uptake was seen in 85% of patients and tumour to blood ratio and hypoxic volume showed a trend to be an independent prognostic measure. Rischin et al. (2006) demonstrated that pretreatment 18F-FMISO was effective in determining hypoxic regions and tirapazamine was effective in patients with hypoxic tumours as assessed by 18F-FMISO. Another group found that an standardised uptake value (SUV) > 2 and tumour to muscle ratio (TMR) > 1.6 at 4 h after injection were associated with disease recurrence after RT (Eschmann et al. 2005). The same group also investigated the changes in 18F-FMISO uptake during RT by performing a scan pretreatment and repeating at 30 Gy. The mean SUV and TMR significantly decreased during RT, indicating RT-induced reoxygenation. Lee et al. (2009) also performed a baseline and midtreatment scan and found 90% of 20 patients had hypoxia before treatment and only 2 had detectable hypoxia on their midtreatment scan, which did not correlate with outcome. Zips et al. (2012) carried out a baseline scan and 3 further scans during RT. The imaging parameters from scans at week 1 and 2 of RT strongly correlated with progression free survival, suggesting the prognostic value of imaging hypoxia at the start of RT rather than at baseline. Smaller studies have showed variable results, with 18F-FMISO showing borderline significance for stratifying patients into treatment outcome groups in 12 patients, and high SUVmax but not TMR correlating with poor DSS after RT or surgery in 17 patients (Kikuchi et al. 2011).
18F-FMISO is not a universal hypoxia tracer in cancer. It has shown minimal activity in pancreatic cancer patients (Segard et al. 2013) and high background uptake in the normal rectum in patients with colorectal cancer, as well as tracer diffusion through the bowel wall, making images difficult to interpret (Roels et al. 2008). The low uptake of 18F-FMISO in target tissue and slow clearance of unbound 18F-FMISO from normoxic areas results in images of poor contrast. The relatively short half-life of 110 min hampers late imaging that could enhance contrast between hypoxic and normoxic tissues. This has led to the development of other tracers with improved pharmacokinetics.
3.6.5.3 18F-Fluoroazomycin Arabinofuranoside (18F-FAZA)
This second generation 2-nitroimidazole is more hydrophilic than 18F-FMISO and therefore has faster clearance from normal tissues and higher tumour to background ratios. Souvatzoglou et al. (2007) carried out an 11 patient pilot study in HNSCC and reported a higher contrast with non-target tissues compared to 18F-FMISO, with an average TMR of 2.0 at 2 h post injection. Mortensen et al. (2012) performed static 18F-FAZA imaging in 40 oropharynx SCC patients. A hypoxic volume could be identified in 25 patients, with a median TMR of 1.5. The distribution of hypoxia among HPV positive and negative tumours was not significantly different and there was a significant difference in DFS in patients with non-hypoxic tumours compared with hypoxic tumours. Tumours were ranked according to the expression of genes included in a 15-gene hypoxia classifier but no correlation was found between the hypoxic status as assessed by the classifier and 18F-FAZA imaging, despite the gene profile being validated using 18F-FAZA autoradiography in human tumourxenografts (Toustrup et al. 2011).
3.6.5.4 Copper(II)-Diacetyl-Bis (N4-Methylthiosemicarbazone) (Cu-ATSM )
Cu-ATSM was developed to overcome some of the limitations of 18F-FMISO imaging. It is an uncharged lipophilic molecule of low molecular weight and high membrane permeability. It has fast tumour uptake and clearance form normoxic tissues, allowing rapid imaging after injection and higher hypoxic-to-normoxic contrast. It is also simpler to synthesise and radiolabel compared with 18F-FMISO, but the longer half-life results in a higher radiation dose to patients (Lewis et al. 2001).
The hypoxia selectivity of Cu-ATSM is thought to be due to the bioreduction of Cu(II)-ATSM to Cu(I)-ATSM, which occurs in both normoxic and hypoxic cells, generating unstable anionic Cu(I)-ATSM. Cu(I) slowly dissociates from the ATSM ligand in cells with low oxygen concentration, becoming irreversibly trapped within the cell due to its negative charge. In the presence of oxygen Cu(I)-ATSM oxidised back to neutral Cu(II)-ATSM and diffuses out of the cell. The hypoxia selectivity therefore is a competition between dissociation and reoxidation of the reduced copper complexes, which is dependent on the oxygen concentration (Dearling et al. 2002). The precise cellular retention mechanism of Cu-ATSM is still unclear and uptake and retention has been associated with a number of other factors, such as over-reduced states caused by mitochondrial dysfunction (Yoshii et al. 2012), the expression of multidrug resistance protein (Liu et al. 2009) and Cu metabolism itself (Hueting et al. 2014). In addition preclinical studies have suggested hypoxia specificity may be dependent on tumour type (O’Donoghue et al. 2005; Yuan et al. 2006) and therefore concerns have been raised regarding the use of Cu-ATSM as a hypoxia radiotracer. However, small clinical studies in a number of tumour types have demonstrated correlation between uptake and poor prognosis, warranting further investigation.
3.6.5.5 Clinical Studies of Cu-ATSM in HNSCC
In 2001 Chao et al. (2001) used 60Cu-ATSM in a radiotherapy planning study to demonstrate the feasibility of Cu-ATSM-guided IMRT to overcome hypoxia tumour resistance. Minagawa et al. (2011) assessed tumour hypoxia using 62Cu-ATSM as a predictor of response in 17 patients. After 2 years of follow up SUVmax but not TMR was significantly different in patients with or without residual/recurrent tumour, and a SUVmax > 5 was a possible cut off threshold for poor outcome. There was no significant difference in 18F-FDG between patients with and without residual/recurrence tumour. Sato et al. (2014) also evaluated the prognostic potential of 62Cu-ATSM in 25 patients with head and neck cancer. 62Cu-ATSM SUVmax > 3.6 was associated significantly worse progression free survival (PFS), and a TMR > 3.2 was associated with significantly worse progression free survival and cause specific survival (CSS). 18F-FDG uptake parameters did not distinguish patient with good and poor outcome. 64Cu-ATSM in head and neck cancers was used to assess the efficacy of pretherapy 64Cu-ATSM scanning as a prognostic factor of response to therapy in 11 patients (Grassi et al. 2014). Patients received a 64Cu-ATSM and 18F-FDG PET scan before commencing treatment and response was determined by an 18F-FDG PET 3 months after therapy. The sensitivity and specificity of 64Cu-ATSM SUVmax in predicting complete response to therapy were 100 and 50% respectively.
3.7 Conclusions
Tumour hypoxia remains one of the major causes of treatment failure in head and neck cancer. Hypoxia promotes a more aggressive tumour phenotype and is considered an important therapeutic target. Current methods of hypoxia identification are inefficient and therefore it is difficult to accurately assess hypoxia in tumours; accurate detection of hypoxia is important to allow stratification of patients for tailored treatment, and to reduce unnecessary side effects of intensive treatment modalities for less hypoxic tumours. In recent years, research in this field has substantially increased our understanding of the molecular pathways that contribute to tumour hypoxia and key pathways such as HIF, mTOR, UPR, and many more upstream and downstream biological targets of tumour hypoxia have been identified. Several of hypoxia-regulated genes are under investigation for drug development. In addition, a number of mRNA and microRNA signatures of tumour hypoxia have been published. So far, the clinical applications of such biomarkers have been limited due to inconsistencies and heterogeneity between studies. Approaches exploiting tumour hypoxia both for development of anti-cancer drugs that are activated in a hypoxic environment and therapeutic approaches that more directly target tumour hypoxia are under development.
References
Adam MF, Gabalski EC, Bloch DA et al (1999) Tissue oxygen distribution in head and neck cancer patients. Head Neck 21:146–153
Aebersold DM, Burri P, Beer KT et al (2001) Expression of hypoxia-inducible factor-1alpha: a novel predictive and prognostic parameter in the radiotherapy of oropharyngeal cancer. Cancer Res 61:2911–2916
Ahmed M, Behera R, Chakraborty G et al (2011) Osteopontin: a potentially important therapeutic target in cancer. Expert Opin Ther Targets 15:1113–1126
Balamurugan K (2015) HIF-1 at the crossroads of hypoxia, inflammation, and cancer. Int J Cancer J Int du Cancer
Bayer C, Shi K, Astner ST, Maftei CA, Vaupel P (2011) Acute versus chronic hypoxia: why a simplified classification is simply not enough. Int J Radiat Oncol Biol Phys 80:965–968
Beasley NJ, Leek R, Alam M et al (2002) Hypoxia-inducible factors HIF-1alpha and HIF-2alpha in head and neck cancer: relationship to tumor biology and treatment outcome in surgically resected patients. Cancer Res 62:2493–2497
Becker A, Hansgen G, Bloching M, Weigel C, Lautenschlager C, Dunst J (1998) Oxygenation of squamous cell carcinoma of the head and neck: comparison of primary tumors, neck node metastases, and normal tissue. Int J Radiat Oncol Biol Phys 42:35–41
Bennett M, Feldmeier J, Smee R, Milross C (2008) Hyperbaric oxygenation for tumour sensitisation to radiotherapy: a systematic review of randomised controlled trials. Cancer Treat Rev 34:577–591
Bentzen J, Toustrup K, Eriksen JG, Primdahl H, Andersen LJ, Overgaard J (2015) Locally advanced head and neck cancer treated with accelerated radiotherapy, the hypoxic modifier nimorazole and weekly cisplatin. Results from the DAHANCA 18 phase II study. Acta Oncol 1–7
Bi M, Naczki C, Koritzinsky M et al (2005) ER stress-regulated translation increases tolerance to extreme hypoxia and promotes tumor growth. EMBO J 24:3470–3481
Bittner MI, Grosu AL (2013) Hypoxia in head and neck tumors: characteristics and development during therapy. Front Oncol 3:223
Bohlius J, Schmidlin K, Brillant C et al (2009) Recombinant human erythropoiesis-stimulating agents and mortality in patients with cancer: a meta-analysis of randomised trials. Lancet 373:1532–1542
Brizel DM, Sibley GS, Prosnitz LR, Scher RL, Dewhirst MW (1997) Tumor hypoxia adversely affects the prognosis of carcinoma of the head and neck. Int J Radiat Oncol Biol Phys 38:285–289
Brizel DM, Dodge RK, Clough RW, Dewhirst MW (1999) Oxygenation of head and neck cancer: changes during radiotherapy and impact on treatment outcome. Radiother Oncol J Eur Soc Ther Radiol Oncol 53:113–117
Brizel DM, Schroeder T, Scher RL et al (2001) Elevated tumor lactate concentrations predict for an increased risk of metastases in head-and-neck cancer. Int J Radiat Oncol Biol Phys 51:349–353
Brown JM (1993) SR 4233 (tirapazamine): a new anticancer drug exploiting hypoxia in solid tumours. Br J Cancer 67:1163–1170
Bruning U, Cerone L, Neufeld Z et al (2011) MicroRNA-155 promotes resolution of hypoxia-inducible factor 1alpha activity during prolonged hypoxia. Mol Cell Biol 31:4087–4096
Bruning U, Fitzpatrick SF, Frank T, Birtwistle M, Taylor CT, Cheong A (2012) NFkappaB and HIF display synergistic behaviour during hypoxic inflammation. Cell Mol Life Sci CMLS 69:1319–1329
Buffa FM, Harris AL, West CM, Miller CJ (2010) Large meta-analysis of multiple cancers reveals a common, compact and highly prognostic hypoxia metagene. Br J Cancer 102:428–435
Bussink J, Kaanders JH, Rijken PF, Raleigh JA, Van der Kogel AJ (2000) Changes in blood perfusion and hypoxia after irradiation of a human squamous cell carcinoma xenograft tumor line. Radiat Res 153:398–404
Camps C, Saini HK, Mole DR et al (2014) Integrated analysis of microRNA and mRNA expression and association with HIF binding reveals the complexity of microRNA expression regulation under hypoxia. Mol Cancer 13:28
Catenacci DV (2014) Next-generation clinical trials: novel strategies to address the challenge of tumor molecular heterogeneity. Mol Oncol
Chan SY, Zhang YY, Hemann C, Mahoney CE, Zweier JL, Loscalzo J (2009) MicroRNA-210 controls mitochondrial metabolism during hypoxia by repressing the iron-sulfur cluster assembly proteins ISCU1/2. Cell Metab 10:273–284
Chandel NS, McClintock DS, Feliciano CE et al (2000) Reactive oxygen species generated at mitochondrial complex III stabilize hypoxia-inducible factor-1alpha during hypoxia: a mechanism of O2 sensing. J Biol Chem 275:25130–25138
Chang Q, Jurisica I, Do T, Hedley DW (2011) Hypoxia predicts aggressive growth and spontaneous metastasis formation from orthotopically grown primary xenografts of human pancreatic cancer. Cancer Res 71:3110–3120
Chang W, Lee CY, Park JH et al (2013) Survival of hypoxic human mesenchymal stem cells is enhanced by a positive feedback loop involving miR-210 and hypoxia-inducible factor 1. J Veterinary Sci 14:69–76
Chao KS, Bosch WR, Mutic S et al (2001) A novel approach to overcome hypoxic tumor resistance: Cu-ATSM-guided intensity-modulated radiation therapy. Int J Radiat Oncol Biol Phys 49:1171–1182
Chi JT, Wang Z, Nuyten DS et al (2006) Gene expression programs in response to hypoxia: cell type specificity and prognostic significance in human cancers. PLoS Med 3:e47
Chien CY, Su CY, Chuang HC et al (2009) Comprehensive study on the prognostic role of osteopontin expression in oral squamous cell carcinoma. Oral Oncol 45:798–802
Chio CC, Lin JW, Cheng HA et al (2013) MicroRNA-210 targets antiapoptotic Bcl-2 expression and mediates hypoxia-induced apoptosis of neuroblastoma cells. Arch Toxicol 87:459–468
Chopra S, Foltz WD, Milosevic MF et al (2009) Comparing oxygen-sensitive MRI (BOLD R2*) with oxygen electrode measurements: a pilot study in men with prostate cancer. Int J Radiat Biol 85:805–813
Comerford KM, Wallace TJ, Karhausen J, Louis NA, Montalto MC, Colgan SP (2002) Hypoxia-inducible factor-1-dependent regulation of the multidrug resistance (MDR1) gene. Cancer Res 62:3387–3394
Crosby ME, Kulshreshtha R, Ivan M, Glazer PM (2009) MicroRNA regulation of DNA repair gene expression in hypoxic stress. Cancer Res 69:1221–1229
Dearling JL, Lewis JS, Mullen GE, Welch MJ, Blower PJ (2002) Copper bis(thiosemicarbazone) complexes as hypoxia imaging agents: structure-activity relationships. J Biol Inorg Chem JBIC Publ Soc Biol Inorg Chem 7:249–259
Denko NC, Fontana LA, Hudson KM et al (2003) Investigating hypoxic tumor physiology through gene expression patterns. Oncogene 22:5907–5914
Dennis MF, Stratford MR, Wardman P, Watts ME (1985) Cellular uptake of misonidazole and analogues with acidic or basic functions. Int J Radiat Biol Related Stud Phys Chem Med 47:629–643
Doedens AL, Stockmann C, Rubinstein MP et al (2010) Macrophage expression of hypoxia-inducible factor-1 alpha suppresses T-cell function and promotes tumor progression. Cancer Res 70:7465–7475
Dunst J, Stadler P, Becker A et al (2003) Tumor volume and tumor hypoxia in head and neck cancers. The amount of the hypoxic volume is important. Strahlentherapie und Onkologie: Organ der Deutschen Rontgengesellschaft [et al]179:521–526
Eckert AW, Lautner MH, Schutze A, Taubert H, Schubert J, Bilkenroth U (2011) Coexpression of hypoxia-inducible factor-1alpha and glucose transporter-1 is associated with poor prognosis in oral squamous cell carcinoma patients. Histopathology 58:1136–1147
Eriksen JG, Overgaard J, Danish H (2007) Neck cancer study G. Lack of prognostic and predictive value of CA IX in radiotherapy of squamous cell carcinoma of the head and neck with known modifiable hypoxia: an evaluation of the DAHANCA 5 study. Radiother Oncol J Eur Soc Ther Radiol Oncol 83:383–388
Eschmann SM, Paulsen F, Reimold M et al (2005) Prognostic impact of hypoxia imaging with 18F-misonidazole PET in non-small cell lung cancer and head and neck cancer before radiotherapy. J Nucl Med Official Publ Soc Nucl Med 46:253–260
Eustace A, Mani N, Span PN et al (2013a) A 26-gene hypoxia signature predicts benefit from hypoxia-modifying therapy in laryngeal cancer but not bladder cancer. Clin Cancer Res: Official J Am Assoc Cancer Res 19:4879–4888
Eustace A, Mani N, Span PN et al (2013b) A 26-gene hypoxia signature predicts benefit from hypoxia-modifying therapy in laryngeal cancer but not bladder cancer. Clin Cancer Res Official J Am Assoc Cancer Res 19:4879–4888
Evans SM, Hahn S, Pook DR et al (2000) Detection of hypoxia in human squamous cell carcinoma by EF5 binding. Cancer Res 60:2018–2024
Evans SM, Du KL, Chalian AA et al (2007) Patterns and levels of hypoxia in head and neck squamous cell carcinomas and their relationship to patient outcome. Int J Radiat Oncol Biol Phys 69:1024–1031
Fang JS, Gillies RD, Gatenby RA (2008) Adaptation to hypoxia and acidosis in carcinogenesis and tumor progression. Semin Cancer Biol 18:330–337
Fasanaro P, D’Alessandra Y, Di Stefano V et al (2008) MicroRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand Ephrin-A3. J Biol Chem 283:15878–15883
Ferrara N, Gerber HP, LeCouter J (2003) The biology of VEGF and its receptors. Nat Med 9:669–676
Fortin A, Wang CS, Vigneault E (2008) Effect of pretreatment anemia on treatment outcome of concurrent radiochemotherapy in patients with head and neck cancer. Int J Radiat Oncol Biol Phys 72:255–260
Fukumura D, Jain RK (2007) Tumor microenvironment abnormalities: causes, consequences, and strategies to normalize. J Cell Biochem 101:937–949
Fury MG, Lee NY, Sherman E et al (2012) A phase 2 study of bevacizumab with cisplatin plus intensity-modulated radiation therapy for stage III/IVB head and neck squamous cell cancer. Cancer 118:5008–5014
Galvin JM, De Neve W (2007) Intensity modulating and other radiation therapy devices for dose painting. J Clin Oncol Official J Am Soc Clin Oncol 25:924–930
Gatenby RA, Kessler HB, Rosenblum JS et al (1988) Oxygen distribution in squamous cell carcinoma metastases and its relationship to outcome of radiation therapy. Int J Radiat Oncol Biol Phys 14:831–838
Gee HE, Camps C, Buffa FM et al (2010) hsa-mir-210 is a marker of tumor hypoxia and a prognostic factor in head and neck cancer. Cancer 116:2148–2158
Graeber TG, Osmanian C, Jacks T et al (1996) Hypoxia-mediated selection of cells with diminished apoptotic potential in solid tumours. Nature 379:88–91
Grassi I, Nanni C, Cicoria G et al (2014) Usefulness of 64Cu-ATSM in head and neck cancer: a preliminary prospective study. Clin Nucl Med 39:e59–e63
Gray LH, Conger AD, Ebert M, Hornsey S, Scott OC (1953) The concentration of oxygen dissolved in tissues at the time of irradiation as a factor in radiotherapy. Br J Radiol 26:638–648
Guimbellot JS, Erickson SW, Mehta T et al (2009) Correlation of microRNA levels during hypoxia with predicted target mRNAs through genome-wide microarray analysis. BMC Med Genomics 2:15
Harada H, Itasaka S, Kizaka-Kondoh S et al (2009) The Akt/mTOR pathway assures the synthesis of HIF-1alpha protein in a glucose- and reoxygenation-dependent manner in irradiated tumors. J Biol Chem 284:5332–5342
Harris AL (2002) Hypoxia–a key regulatory factor in tumour growth. Nat Rev Cancer 2:38–47
Hassan Metwally MA, Ali R, Kuddu M et al (2015) IAEA-HypoX. A randomized multicenter study of the hypoxic radiosensitizer nimorazole concomitant with accelerated radiotherapy in head and neck squamous cell carcinoma. Radiother Oncol J Eur Soc Ther Radiol Oncol
Hebert C, Norris K, Scheper MA, Nikitakis N, Sauk JJ (2007) High mobility group A2 is a target for miRNA-98 in head and neck squamous cell carcinoma. Mol Cancer 6:5
Hendrickson K, Phillips M, Smith W, Peterson L, Krohn K, Rajendran J (2011) Hypoxia imaging with [F-18] FMISO-PET in head and neck cancer: potential for guiding intensity modulated radiation therapy in overcoming hypoxia-induced treatment resistance. Radiother Oncol J Eur Soc Ther Radiol Oncol 101:369–375
Hoang T, Huang S, Armstrong E, Eickhoff JC, Harari PM (2012) Enhancement of radiation response with bevacizumab. J Exp Clin Cancer Res CR 31:37
Hockel M, Vaupel P (2001) Tumor hypoxia: definitions and current clinical, biologic, and molecular aspects. J Nat Cancer Inst 93:266–276
Hoff CM, Hansen HS, Overgaard M et al (2011) The importance of haemoglobin level and effect of transfusion in HNSCC patients treated with radiotherapy–results from the randomized DAHANCA 5 study. Radiother Oncol J Eur Soc Ther Radiol Oncol 98:28–33
Horsman MR, Mortensen LS, Petersen JB, Busk M, Overgaard J (2012) Imaging hypoxia to improve radiotherapy outcome. Nat Rev Clin Oncol 9:674–687
Howard-Flanders P, Moore D (1958) The time interval after pulsed irradiation within which injury to bacteria can be modified by dissolved oxygen. I. A search for an effect of oxygen 0.02 second after pulsed irradiation. Radiat Res 9:422–437
Hua Z, Lv Q, Ye W et al (2006) MiRNA-directed regulation of VEGF and other angiogenic factors under hypoxia. PLoS ONE 1:e116
Huang X, Ding L, Bennewith KL et al (2009) Hypoxia-inducible mir-210 regulates normoxic gene expression involved in tumor initiation. Mol Cell 35:856–867
Hueting R, Kersemans V, Cornelissen B et al (2014) A comparison of the behavior of (64)Cu-acetate and (64)Cu-ATSM in vitro and in vivo. J Nucl Med Official Publ Soc Nucl Med 55:128–134
Jansen JF, Schoder H, Lee NY et al (2010) Noninvasive assessment of tumor microenvironment using dynamic contrast-enhanced magnetic resonance imaging and 18F-fluoromisonidazole positron emission tomography imaging in neck nodal metastases. Int J Radiat Oncol Biol Phys 77:1403–1410
Janssens GO, Rademakers SE, Terhaard CH et al (2012) Accelerated radiotherapy with carbogen and nicotinamide for laryngeal cancer: results of a phase III randomized trial. J Clin Oncol Official J Am Soc Clin Oncol 30:1777–1783
Jonathan RA, Wijffels KI, Peeters W et al (2006) The prognostic value of endogenous hypoxia-related markers for head and neck squamous cell carcinomas treated with ARCON. Radiother Oncol J Eur Soc Ther Radiol Oncol 79:288–297
Jung JE, Lee HG, Cho IH et al (2005) STAT3 is a potential modulator of HIF-1-mediated VEGF expression in human renal carcinoma cells. FASEB J Official Publ Fed Am Soc Exp Biol 19:1296–1298
Kaanders JH, Pop LA, Marres HA et al (2002a) ARCON: experience in 215 patients with advanced head-and-neck cancer. Int J Radiat Oncol Biol Phys 52:769–778
Kaanders JH, Wijffels KI, Marres HA et al (2002b) Pimonidazole binding and tumor vascularity predict for treatment outcome in head and neck cancer. Cancer Res 62:7066–7074
Kikuchi M, Yamane T, Shinohara S et al (2011) 18F-fluoromisonidazole positron emission tomography before treatment is a predictor of radiotherapy outcome and survival prognosis in patients with head and neck squamous cell carcinoma. Ann Nucl Med 25:625–633
Kim HW, Haider HK, Jiang S, Ashraf M (2009) Ischemic preconditioning augments survival of stem cells via miR-210 expression by targeting caspase-8-associated protein 2. J Biol Chem 284:33161–33168
Koong AC, Denko NC, Hudson KM et al (2000) Candidate genes for the hypoxic tumor phenotype. Cancer Res 60:883–887
Koukourakis MI, Giatromanolaki A, Sivridis E et al (2001a) Squamous cell head and neck cancer: evidence of angiogenic regeneration during radiotherapy. Anticancer Res 21:4301–4309
Koukourakis MI, Giatromanolaki A, Sivridis E et al (2001b) Hypoxia-regulated carbonic anhydrase-9 (CA9) relates to poor vascularization and resistance of squamous cell head and neck cancer to chemoradiotherapy. Clin Cancer Res Official J Am Assoc Cancer Res 7:3399–3403
Koukourakis MI, Giatromanolaki A, Sivridis E et al (2002) Hypoxia-inducible factor (HIF1A and HIF2A), angiogenesis, and chemoradiotherapy outcome of squamous cell head-and-neck cancer. Int J Radiat Oncol Biol Phys 53:1192–1202
Krishnamachary B, Zagzag D, Nagasawa H et al (2006) Hypoxia-inducible factor-1-dependent repression of E-cadherin in von Hippel-Lindau tumor suppressor-null renal cell carcinoma mediated by TCF3, ZFHX1A, and ZFHX1B. Cancer Res 66:2725–2731
Kulshreshtha R, Ferracin M, Wojcik SE et al (2007) A microRNA signature of hypoxia. Mol Cell Biol 27:1859–1867
Kyzas PA, Cunha IW, Ioannidis JP (2005a) Prognostic significance of vascular endothelial growth factor immunohistochemical expression in head and neck squamous cell carcinoma: a meta-analysis. Clin Cancer Res Official J Am Assoc Cancer Res 11:1434–1440
Kyzas PA, Stefanou D, Batistatou A, Agnantis NJ (2005b) Hypoxia-induced tumor angiogenic pathway in head and neck cancer: an in vivo study. Cancer Lett 225:297–304
Lambin P, Ramaekers BL, van Mastrigt GA et al (2009) Erythropoietin as an adjuvant treatment with (chemo) radiation therapy for head and neck cancer. The Cochrane database Syst Rev CD006158
Lee DJ, Cosmatos D, Marcial VA et al (1995) Results of an RTOG phase III trial (RTOG 85-27) comparing radiotherapy plus etanidazole with radiotherapy alone for locally advanced head and neck carcinomas. Int J Radiat Oncol Biol Phys 32:567–576
Lee WR, Berkey B, Marcial V et al (1998a) Anemia is associated with decreased survival and increased locoregional failure in patients with locally advanced head and neck carcinoma: a secondary analysis of RTOG 85-27. Int J Radiat Oncol Biol Phys 42:1069–1075
Lee DJ, Trotti A, Spencer S et al (1998b) Concurrent tirapazamine and radiotherapy for advanced head and neck carcinomas: a phase II study. Int J Radiat Oncol Biol Phys 42:811–815
Lee N, Nehmeh S, Schoder H et al (2009) Prospective trial incorporating pre-/mid-treatment [18F]-misonidazole positron emission tomography for head-and-neck cancer patients undergoing concurrent chemoradiotherapy. Int J Radiat Oncol Biol Phys 75:101–108
Lewis JS, Sharp TL, Laforest R, Fujibayashi Y, Welch MJ (2001) Tumor uptake of copper-diacetyl-bis(N(4)-methylthiosemicarbazone): effect of changes in tissue oxygenation. J Nucl Med Official Publ Soc Nucl Med 42:655–661
Lim AM, Rischin D, Fisher R et al (2012) Prognostic significance of plasma osteopontin in patients with locoregionally advanced head and neck squamous cell carcinoma treated on TROG 02.02 phase III trial. Clin Cancer Res Official J Am Assoc Cancer Res 18:301–307
Liu J, Hajibeigi A, Ren G et al (2009) Retention of the radiotracers 64Cu-ATSM and 64Cu-PTSM in human and murine tumors is influenced by MDR1 protein expression. J Nucl Med Official Publ Soc Nucl Med 50:1332–1339
Ljungkvist AS, Bussink J, Kaanders JH, van der Kogel AJ (2007) Dynamics of tumor hypoxia measured with bioreductive hypoxic cell markers. Radiat Res 167:127–145
Lu H, Forbes RA, Verma A (2002) Hypoxia-inducible factor 1 activation by aerobic glycolysis implicates the Warburg effect in carcinogenesis. J Biol Chem 277:23111–23115
Magagnin MG, van den Beucken T, Sergeant K et al (2008) The mTOR target 4E-BP1 contributes to differential protein expression during normoxia and hypoxia through changes in mRNA translation efficiency. Proteomics 8:1019–1028
Marxsen JH, Stengel P, Doege K et al (2004) Hypoxia-inducible factor-1 (HIF-1) promotes its degradation by induction of HIF-alpha-prolyl-4-hydroxylases. Biochem J 381:761–767
Minagawa Y, Shizukuishi K, Koike I et al (2011) Assessment of tumor hypoxia by 62Cu-ATSM PET/CT as a predictor of response in head and neck cancer: a pilot study. Ann Nucl Med 25:339–345
Mineta H, Miura K, Takebayashi S et al (2002) Prognostic value of glucose transporter 1 expression in patients with hypopharyngeal carcinoma. Anticancer Res 22:3489–3494
Mortensen LS, Johansen J, Kallehauge J et al (2012) FAZA PET/CT hypoxia imaging in patients with squamous cell carcinoma of the head and neck treated with radiotherapy: results from the DAHANCA 24 trial. Radiother Oncol J Eur Soc Ther Radiol Oncol 105:14–20
Nagelkerke A, Bussink J, Mujcic H et al (2013) Hypoxia stimulates migration of breast cancer cells via the PERK/ATF4/LAMP3-arm of the unfolded protein response. Breast Cancer Res BCR 15:R2
Newbold K, Castellano I, Charles-Edwards E et al (2009) An exploratory study into the role of dynamic contrast-enhanced magnetic resonance imaging or perfusion computed tomography for detection of intratumoral hypoxia in head-and-neck cancer. Int J Radiat Oncol Biol Phys 74:29–37
Nordsmark M, Overgaard J (2000) A confirmatory prognostic study on oxygenation status and loco-regional control in advanced head and neck squamous cell carcinoma treated by radiation therapy. Radiother Oncol J Eur Soc Ther Radiol Oncol 57:39–43
Nordsmark M, Overgaard M, Overgaard J (1996) Pretreatment oxygenation predicts radiation response in advanced squamous cell carcinoma of the head and neck. Radiother Oncol J Eur Soc Ther Radiol Oncol 41:31–39
Nordsmark M, Loncaster J, Aquino-Parsons C et al (2003) Measurements of hypoxia using pimonidazole and polarographic oxygen-sensitive electrodes in human cervix carcinomas. Radiother Oncol J Eur Soc Ther Radiol Oncol 67:35–44
Nordsmark M, Bentzen SM, Rudat V et al (2005) Prognostic value of tumor oxygenation in 397 head and neck tumors after primary radiation therapy. An international multi-center study. Radiother Oncol J Eur Soc Ther Radiol Oncol 77:18–24
Nordsmark M, Loncaster J, Aquino-Parsons C et al (2006) The prognostic value of pimonidazole and tumour pO2 in human cervix carcinomas after radiation therapy: a prospective international multi-center study. Radiother Oncol J Eur Soc Ther Radiol Oncol 80:123–131
Nordsmark M, Eriksen JG, Gebski V, Alsner J, Horsman MR, Overgaard J (2007) Differential risk assessments from five hypoxia specific assays: the basis for biologically adapted individualized radiotherapy in advanced head and neck cancer patients. Radiother Oncol J Eur Soc Ther Radiol Oncol 83:389–397
Nozue M, Lee I, Yuan F et al (1997) Interlaboratory variation in oxygen tension measurement by Eppendorf “Histograph” and comparison with hypoxic marker. J Surgical Oncol 66:30–38
O’Donoghue JA, Zanzonico P, Pugachev A et al (2005) Assessment of regional tumor hypoxia using 18F-fluoromisonidazole and 64Cu(II)-diacetyl-bis(N4-methylthiosemicarbazone) positron emission tomography: comparative study featuring microPET imaging, Po2 probe measurement, autoradiography, and fluorescent microscopy in the R3327-AT and FaDu rat tumor models. Int J Radiat Oncol Biol Phys 61:1493–1502
Oliver RJ, Woodwards RT, Sloan P, Thakker NS, Stratford IJ, Airley RE (2004) Prognostic value of facilitative glucose transporter Glut-1 in oral squamous cell carcinomas treated by surgical resection; results of EORTC translational research fund studies. Eur J Cancer 40:503–507
Overgaard J (1994) Clinical evaluation of nitroimidazoles as modifiers of hypoxia in solid tumors. Oncol Res 6:509–518
Overgaard J (2011) Hypoxic modification of radiotherapy in squamous cell carcinoma of the head and neck–a systematic review and meta-analysis. Radiother Oncol J Eur Soc Ther Radiol Oncol 100:22–32
Overgaard J, Hansen HS, Overgaard M et al (1998) A randomized double-blind phase III study of nimorazole as a hypoxic radiosensitizer of primary radiotherapy in supraglottic larynx and pharynx carcinoma. Results of the Danish Head and Neck Cancer Study (DAHANCA) Protocol 5-85. Radiother Oncol J Eur Soc Ther Radiol Oncol 46:135–146
Overgaard J, Eriksen JG, Nordsmark M, Alsner J, Horsman MR (2005) Plasma osteopontin, hypoxia, and response to the hypoxia sensitiser nimorazole in radiotherapy of head and neck cancer: results from the DAHANCA 5 randomised double-blind placebo-controlled trial. Lancet Oncol 6:757–764
Pereira ER, Frudd K, Awad W, Hendershot LM (2014) Endoplasmic reticulum (ER) stress and hypoxia response pathways interact to potentiate hypoxia-inducible factor 1 (HIF-1) transcriptional activity on targets like vascular endothelial growth factor (VEGF). J Biol Chem 289:3352–3364
Prekeges JL, Rasey JS, Grunbaum Z, Krohn KH (1991) Reduction of fluoromisonidazole, a new imaging agent for hypoxia. Biochem Pharmacol 42:2387–2395
Puissegur MP, Mazure NM, Bertero T et al (2011) miR-210 is overexpressed in late stages of lung cancer and mediates mitochondrial alterations associated with modulation of HIF-1 activity. Cell Death Differ 18:465–478
Rabinowitz MH (2013) Inhibition of hypoxia-inducible factor prolyl hydroxylase domain oxygen sensors: tricking the body into mounting orchestrated survival and repair responses. J Med Chem 56:9369–9402
Rademakers SE, Span PN, Kaanders JH, Sweep FC, van der Kogel AJ, Bussink J (2008) Molecular aspects of tumour hypoxia. Mol Oncol 2:41–53
Rademakers SE, Lok J, van der Kogel AJ, Bussink J, Kaanders JH (2011) Metabolic markers in relation to hypoxia; staining patterns and colocalization of pimonidazole, HIF-1alpha, CAIX, LDH-5, GLUT-1, MCT1 and MCT4. BMC Cancer 11:167
Rajendran JG, Schwartz DL, O’Sullivan J et al (2006) Tumor hypoxia imaging with [F-18] fluoromisonidazole positron emission tomography in head and neck cancer. Clin Cancer Res Official J Am Assoc Cancer Res 12:5435–5441
Raleigh JA, Calkins-Adams DP, Rinker LH et al (1998) Hypoxia and vascular endothelial growth factor expression in human squamous cell carcinomas using pimonidazole as a hypoxia marker. Cancer Res 58:3765–3768
Raleigh JA, Chou SC, Arteel GE, Horsman MR (1999) Comparisons among pimonidazole binding, oxygen electrode measurements, and radiation response in C3H mouse tumors. Radiat Res 151:580–589
Rischin D, Peters L, Hicks R et al (2001) Phase I trial of concurrent tirapazamine, cisplatin, and radiotherapy in patients with advanced head and neck cancer. J Clin Oncol Official J Am Soc Clin Oncol 19:535–542
Rischin D, Peters L, Fisher R et al (2005) Tirapazamine, cisplatin, and radiation versus fluorouracil, cisplatin, and radiation in patients with locally advanced head and neck cancer: a randomized phase II trial of the Trans-Tasman Radiation Oncology Group (TROG 98.02). J Clin Oncol Official J Am Soc Clin Oncol 23:79–87
Rischin D, Hicks RJ, Fisher R et al (2006) Prognostic significance of [18F]-misonidazole positron emission tomography-detected tumor hypoxia in patients with advanced head and neck cancer randomly assigned to chemoradiation with or without tirapazamine: a substudy of Trans-Tasman radiation oncology group study 98.02. J Clin Oncol Official J Am Soc Clin Oncol 24:2098–2104
Rischin D, Narayan K, Oza AM et al (2010) Phase 1 study of tirapazamine in combination with radiation and weekly cisplatin in patients with locally advanced cervical cancer. Int J Gynecol Cancer Official J Int Gynecol Cancer Soc 20:827–833
Roels S, Slagmolen P, Nuyts J et al (2008) Biological image-guided radiotherapy in rectal cancer: is there a role for FMISO or FLT, next to FDG? Acta Oncol 47:1237–1248
Rofstad EK, Galappathi K, Mathiesen B, Ruud EB (2007) Fluctuating and diffusion-limited hypoxia in hypoxia-induced metastasis. Clin Cancer Res Official J Am Assoc Cancer Res 13:1971–1978
Rouschop KM, Dubois L, Schaaf MB et al (2011) Deregulation of cap-dependent mRNA translation increases tumour radiosensitivity through reduction of the hypoxic fraction. Radiother Oncol J Eur Soc Ther Radiol Oncol 99:385–391
Rudat V, Vanselow B, Wollensack P et al (2000) Repeatability and prognostic impact of the pretreatment pO(2) histography in patients with advanced head and neck cancer. Radiother Oncol J Eur Soc Ther Radiol Oncol 57:31–37
Sabharwal SS, Schumacker PT (2014) Mitochondrial ROS in cancer: initiators, amplifiers or an Achilles’ heel? Nat Rev Cancer 14:709–721
Sato Y, Tsujikawa T, Oh M et al (2014) Assessing tumor hypoxia in head and neck cancer by PET with (6)(2)Cu-diacetyl-bis(N(4)-methylthiosemicarbazone). Clin Nucl Med 39:1027–1032
Sattler UG, Meyer SS, Quennet V et al (2010) Glycolytic metabolism and tumour response to fractionated irradiation. Radiother Oncol J Eur Soc Ther Radiol Oncol 94:102–109
Schwartz DL, Powis G, Thitai-Kumar A et al (2009) The selective hypoxia inducible factor-1 inhibitor PX-478 provides in vivo radiosensitization through tumor stromal effects. Mol Cancer Ther 8:947–958
Segard T, Robins PD, Yusoff IF et al (2013) Detection of hypoxia with 18F-fluoromisonidazole (18F-FMISO) PET/CT in suspected or proven pancreatic cancer. Clin Nucl Med 38:1–6
Seigneuric R, Starmans MH, Fung G et al (2007) Impact of supervised gene signatures of early hypoxia on patient survival. Radiother Oncol J Eur Soc Ther Radiol Oncol 83:374–382
Semenza GL (2000) Expression of hypoxia-inducible factor 1: mechanisms and consequences. Biochem Pharmacol 59:47–53
Semenza GL (2010) Defining the role of hypoxia-inducible factor 1 in cancer biology and therapeutics. Oncogene 29:625–634
Sermeus A, Michiels C (2011) Reciprocal influence of the p53 and the hypoxic pathways. Cell Death Dis 2:e164
Skovsgaard T (1977) Transport and binding of daunorubicin, adriamycin, and rubidazone in Ehrlich ascites tumour cells. Biochem Pharmacol 26:215–222
Sobhanifar S, Aquino-Parsons C, Stanbridge EJ, Olive P (2005) Reduced expression of hypoxia-inducible factor-1alpha in perinecrotic regions of solid tumors. Cancer Res 65:7259–7266
Sorensen BS, Toustrup K, Horsman MR, Overgaard J, Alsner J (2010) Identifying pH independent hypoxia induced genes in human squamous cell carcinomas in vitro. Acta Oncol 49:895–905
Souvatzoglou M, Grosu AL, Roper B et al (2007) Tumour hypoxia imaging with [18F]FAZA PET in head and neck cancer patients: a pilot study. Eur J Nucl Med Mol Imaging 34:1566–1575
Span PN, Bussink J (2015) Biology of hypoxia. Semin Nucl Med 45:101–109
Stadler P, Becker A, Feldmann HJ et al (1999) Influence of the hypoxic subvolume on the survival of patients with head and neck cancer. Int J Radiat Oncol Biol Phys 44:749–754
Starmans MH, Chu KC, Haider S et al (2012) The prognostic value of temporal in vitro and in vivo derived hypoxia gene-expression signatures in breast cancer. Radiother Oncol J Eur Soc Ther Radiol Oncol 102:436–443
Suh Y, Amelio I, Guerrero Urbano T, Tavassoli M (2014) Clinical update on cancer: molecular oncology of head and neck cancer. Cell Death Dis 5:e1018
Tannock IF (1998) Conventional cancer therapy: promise broken or promise delayed? Lancet 351(Suppl 2):SII9–16
Thomlinson RH, Gray LH (1955) The histological structure of some human lung cancers and the possible implications for radiotherapy. Br J Cancer 9:539–549
Toustrup K, Sorensen BS, Nordsmark M et al (2011) Development of a hypoxia gene expression classifier with predictive impact for hypoxic modification of radiotherapy in head and neck cancer. Cancer Res 71:5923–5931
Toustrup K, Sorensen BS, Lassen P, Wiuf C, Alsner J, Overgaard J (2012) Gene expression classifier predicts for hypoxic modification of radiotherapy with nimorazole in squamous cell carcinomas of the head and neck. Radiother Oncol J Eur Soc Ther Radiol Oncol 102:122–129
Turkbey B, Thomasson D, Pang Y, Bernardo M, Choyke PL (2010) The role of dynamic contrast-enhanced MRI in cancer diagnosis and treatment. Diagn Interv Radiol 16:186–192
Vadysirisack DD, Ellisen LW (2012) mTOR activity under hypoxia. Methods Mol Biol 821:45–58
Van den Bogaert W, van der Schueren E, Horiot JC et al (1995) The EORTC randomized trial on three fractions per day and misonidazole (trial no. 22811) in advanced head and neck cancer: long-term results and side effects. Radiother Oncol J Eur Soc Ther Radiol Oncol 35:91–99
Varghese AJ, Gulyas S, Mohindra JK (1976) Hypoxia-dependent reduction of 1-(2-nitro-1-imidazolyl)-3-methoxy-2-propanol by Chinese hamster ovary cells and KHT tumor cells in vitro and in vivo. Cancer Res 36:3761–3765
Vaupel P (2004) Tumor microenvironmental physiology and its implications for radiation oncology. Semin Radiat Oncol 14:198–206
Vaupel P, Mayer A (2007) Hypoxia in cancer: significance and impact on clinical outcome. Cancer Metastasis Rev 26:225–239
Vaupel P, Schaefer C, Okunieff P (1994) Intracellular acidosis in murine fibrosarcomas coincides with ATP depletion, hypoxia, and high levels of lactate and total Pi. NMR Biomed 7:128–136
Vaupel P, Hockel M, Mayer A (2007) Detection and characterization of tumor hypoxia using pO2 histography. Antioxid Redox Signaling 9:1221–1235
Wachters JE, Schrijvers ML, Slagter-Menkema L et al (2013) Prognostic significance of HIF-1a, CA-IX, and OPN in T1-T2 laryngeal carcinoma treated with radiotherapy. The Laryngoscope 123:2154–2160
Wang Y, Alam GN, Ning Y et al (2012) The unfolded protein response induces the angiogenic switch in human tumor cells through the PERK/ATF4 pathway. Cancer Res 72:5396–5406
Wang J, Zhao J, Shi M et al (2014) Elevated expression of miR-210 predicts poor survival of cancer patients: a systematic review and meta-analysis. PLoS ONE 9:e89223
Warburg O, Wind F, Negelein E (1927) The metabolism of tumors in the body. J Gen Physiol 8:519–530
Willett CG, Boucher Y, di Tomaso E et al (2004) Direct evidence that the VEGF-specific antibody bevacizumab has antivascular effects in human rectal cancer. Nat Med 10:145–147
Winter SC, Buffa FM, Silva P et al (2007) Relation of a hypoxia metagene derived from head and neck cancer to prognosis of multiple cancers. Cancer Res 67:3441–3449
Wouters BG, Koritzinsky M (2008) Hypoxia signalling through mTOR and the unfolded protein response in cancer. Nat Rev Cancer 8:851–864
Wouters BG, Weppler SA, Koritzinsky M et al (2002) Hypoxia as a target for combined modality treatments. Eur J Cancer 38:240–257
Wozniak AJ, Glisson BS, Hande KR, Ross WE (1984) Inhibition of etoposide-induced DNA damage and cytotoxicity in L1210 cells by dehydrogenase inhibitors and other agents. Cancer Res 44:626–632
Wykoff CC, Beasley NJ, Watson PH et al (2000) Hypoxia-inducible expression of tumor-associated carbonic anhydrases. Cancer Res 60:7075–7083
Yang MH, Wu MZ, Chiou SH et al (2008) Direct regulation of TWIST by HIF-1alpha promotes metastasis. Nat Cell Biol 10:295–305
Ying Q, Liang L, Guo W et al (2011) Hypoxia-inducible microRNA-210 augments the metastatic potential of tumor cells by targeting vacuole membrane protein 1 in hepatocellular carcinoma. Hepatology 54:2064–2075
Yoshii Y, Yoneda M, Ikawa M et al (2012) Radiolabeled Cu-ATSM as a novel indicator of overreduced intracellular state due to mitochondrial dysfunction: studies with mitochondrial DNA-less rho0 cells and cybrids carrying MELAS mitochondrial DNA mutation. Nucl Med Biol 39:177–185
Yuan H, Schroeder T, Bowsher JE, Hedlund LW, Wong T, Dewhirst MW (2006) Intertumoral differences in hypoxia selectivity of the PET imaging agent 64Cu(II)-diacetyl-bis(N4-methylthiosemicarbazone). J Nucl Med Official Publ Soc Nucl Med 47:989–998
Zhang Z, Sun H, Dai H et al (2009) MicroRNA miR-210 modulates cellular response to hypoxia through the MYC antagonist MNT. Cell Cycle 8:2756–2768
Zips D, Zophel K, Abolmaali N et al (2012) Exploratory prospective trial of hypoxia-specific PET imaging during radiochemotherapy in patients with locally advanced head-and-neck cancer. Radiother Oncol J Eur Soc Ther Radiol Oncol 105:21–28
Zuo J, Wen M, Lei M, Peng X, Yang X, Liu Z (2015) MiR-210 links hypoxia with cell proliferation regulation in human Laryngocarcinoma cancer. J Cell Biochem 116:1039–1049
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Tavassoli, M., Suh, Ye. (2017). Hypoxia in Head and Neck Cancer. In: Warnakulasuriya, S., Khan, Z. (eds) Squamous cell Carcinoma. Springer, Dordrecht. https://doi.org/10.1007/978-94-024-1084-6_3
Download citation
DOI: https://doi.org/10.1007/978-94-024-1084-6_3
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-024-1083-9
Online ISBN: 978-94-024-1084-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)