
Abstract
During the four Maurit surveys carried out annually from 2007 to 2010, 342 trawl stations were sampled off the Mauritanian coast between 80 and 2000-m depth. Hydroids were captured at 174 stations, totalling 6169 colonies belonging to 63 species and 19 families . Most of the species were Leptothecata and only seven belonged to Anthoathecata . Thirty-two of the 63 species are new records for Mauritania. Plumulariidae showed the highest species richness (8 species), followed by Sertulariidae and Campanulariidae (7 species), Lafoeidae , Haleciidae and Aglopheniidae (6 species). The most abundant were Sertulariidae , followed by Aglaopheniidae, Plumulariidae, Campanulariidae, Halopterididae and Haleciidae . The ability of hydroids to colonize Mauritanian soft bottom s was driven by hydrorhizal modifications for anchoring the colony to the sediment and by epizoism on 11 invertebrate groups. Families whose species developed both strategies and those which were typically epibionts on hydroids were the most frequent and abundant. Most of the species were eurybathic (58 species), with four species recorded only in the coastal realm, 18 exclusively in the deep benthic realm and 41 in both realms. Therefore, overall hydroid diversity was highest in the deep benthic realm . Biogeographical components included two main groups: species with a wide distribution (59%) and species of Atlantic distribution (41%); our results also emphasize the similarity between the Mauritanian hydroid fauna and the Atlantic-Mediterranean region. Only one species, Hydractinia multitentaculata (Millard 1975), has West African distribution. No endemic species were reported.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
- Biodiversity
- Distribution
- Cnidaria
- Hydrozoa
- Anthoathecata
- Leptothecata
- Deep-waters
- Mauritania
- Northwest Africa
Introduction
The hydroid fauna of the Northeastern Atlantic is one of the best known in the world. This knowledge stems mainly from research accomplished throughout the 20th century on European coasts, including the Mediterranean Sea (for bibliographic reviews see Cornelius 1995; Bouillon et al. 2004; Altuna 2007, 2014; Schuchert 2012). Nevertheless, the hydroid fauna of the Northwest African coast is still poorly studied, despite the high zoogeographical interest of this region, which is a border zone between the boreal -temperate and the tropical biota (Van Soest 1993; Briggs 1995; Briggs and Bowen 2012).
Most of the available literature concerns the results of historical oceanographic expeditions: Challenger (Allman 1883, 1888), Prince Albert I of Monaco (Pictet and Bedot 1900; Bedot 1921; Leloup 1940), Travailleur et Talisman (Billard 1906a), Michael Sars (Broch 1913), Valdivia (Stechow 1925) and Atlantide (Vervoort 1959). In addition, study of material collected during the colonial period has contributed to knowledge of littoral hydroids (Billard 1907, 1924, 1927, 1930, 1931, 1934; Ritchie 1907; Broch 1914; Picard 1951; Buchanan 1956, 1957). Some isolated records can also be found in Quelch (1885), Leloup (1937), Vervoort (1946), and Rees and Thursfield (1965).
The best known hydrozoan fauna in the region is that of the Atlantic coast of Morocco , mainly due to the work of Patriti (1970). His paper provided descriptions of species deposited in collections of the Institut Scientifique Chérifien de Rabat (Morocco), and included previous literature records. Additional information exists on deep-water hydroids from the Ibero-Moroccan Gulf (Ramil and Vervoort 1992) and on collections from Dutch CANCAP Expeditions (Medel and Vervoort 1998, 2000; Ansín Agís et al. 2001; Vervoort 2006), which also included material from the Azores , Madeira , Selvagens , Canary and Cape Verde Islands. More recently, three athecate hydroids were reported from Cape Spartel and Agadir (Morocco) by Iazza et al. (2013). Littoral hydroids from the Canary Islands were studied by Izquierdo et al. (1986a, b, 1990), while those from waters of Guinea Bissau were addressed by Gili et al. (1989).
The only works focussing exclusively on hydroids from Mauritania were those of Billard (1906b, 1931), who studied material collected in fishing trawls. As noted above, Medel and Vervoort (1998, 2000), Ansín Agís et al. (2001) and Vervoort (2006) included material collected in Mauritanian waters during the CANCAP III and Tyro-Mauritania II Expeditions. These publications record a combined total of 47 valid species of benthic hydroids recorded off Cape Blanc and the Banc d’Arguin area, mainly north of Cape Timiris, including shallow waters . No data are available on species from waters off the central and southern coasts of Mauritania.
The four Maurit surveys carried out annually from 2007 to 2010 systematically sampled the entire Mauritanian coast, enabling the collection of a significant amount of hydroids . The aim of this chapter is to improve knowledge of the biodiversity , autecology and distribution of the benthic hydroids living on the deep continental shelf and slope off Mauritania.
Materials and Methods
The hydroid species reported here were collected at 174 trawling stations located along the entire Mauritanian coast between 81 and 1835-m depth, during the four Spanish-Mauritanian multidisciplinary Maurit surveys . Most of the stations (145) were sampled with a commercial otter trawl following a random stratified sampling methodology . In addition, 15 stations were also sampled with a 3.5 m horizontal opening beam trawl along five transects perpendicular to the coastline over five bathymetric strata (150, 300, 500, 1000 and 1500 m). Fourteen samples were obtained with a rock dredge over the coral mounds barrier and the canyon borders (for a detailed description of station data, sampling methodology and maps, see Chaps. 1, 4, 8, 13 and 14).
At each station, hydroids were carefully sorted from the total catch and preserved in 70% ethanol . In the laboratory , further studies involved the final sorting, staining and preparation of slides for microscopic observation following a standard procedure (Ansín Agís et al. 2001), as well as species identification based on specialized literature. Data on abundance (number of colonies) and types of substrata were also recorded.
Hydroid diversity and abundance were spatially represented by geostatistical techniques, using ArcGIS krigging with a spherical semivariogram, with 12 neighbors, and the output cell size of 1000 m.
Geographical distribution and bathymetric ranges were established for each species based on an extensive review of the existing hydrozoan literature and of our own data. The biogeographical components employed in the discussion are based on the distribution patterns proposed by Ansín Agís (1998) and Ansín Agís et al. (2001). The bathymetric groups were performed following the schemes proposed by García Carrascosa (1981), Peña Cantero (2004) and Altuna (2007): (A) Coastal realm species, limited exclusively to the continental shelf (80–200 m); (B) Species restricted to the deep benthic realm and absent from the continental shelf (200–2000 m); and (C) species with a wide vertical distribution, including both realms.
We give an updated checklist after our own data and an extensive review of the literature.
Specimens studied were largely deposited in the collections of the laboratory of Zoology of the Faculty of Marine Sciences , University of Vigo (Spain) and of the Oceanographic Centre of Malaga (Marine Fauna Collection, CFM-IEOMA), belonging to the Spanish Institute of Oceanography (IEO).
Results
General Overview
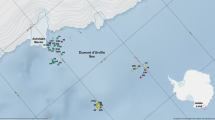
A total of 6169 colonies of hydroids were collected at 174 of the 342 trawl stations carried out during the four Maurit surveys . The frequency did not show high values (50.8%), but the stations where hydroids were sampled were more or less uniformly distributed along the entire Mauritanian coast (Fig. 11.1).

Location of the 179 stations from the four Maurit surveys in which Hydrozoa were collected
We identified 63 species belonging to 19 families . In addition, a total of 529 colonies were only identified to the genus level, due to the absence of reproductive structures and/or to their damaged condition. The current checklist of Mauritanian hydroids includes 81 species and 20 families , the result of combining our data with data extracted from the reviewed literature (Annexes 11.1 and 11.2).
Anthoathecata were only represented by seven species (11%), of which six, Turritopsis dorhnii (Weissmann 1883), Bimeria vestita Wright 1859, Garveia nutans Wright 1859, Eudendrium album Nutting 1898, Eudendrium ramosum (Linnaeus 1758) and Leuckartiara octona (Fleming 1823) are reported here for the first time in Mauritanian waters. The remaining 56 species were Leptothecata , of which 35 are new records from this area (indicated with * in Annexe 11.1). At the family level, Plumulariidae showed the highest species richness (8 species), followed by Sertulariidae and Campanulariidae (7 species), Lafoeidae , Haleciidae and Aglaopheniidae (6 species). Halopterididae , Bougainvilliidae and Tiarannidae recorded intermediate values (4–3 species); the remaining families were only represented by one or two species (Fig. 11.2).

Species richness of the main Hydrozoa families collected in Mauritanian deep waters
Two sertulariid species, Dynamena disticha (Bosc 1802) and Sertularia marginata (Kirchenpauer 1864), also recorded during the surveys, were not considered for further discussion, since they were attached to leaves of Zostera sp, a seagrass absent from the sampled depths and probably fished as floating debris.
Most abundant in the samples were specimens referable to Sertulariidae (1557 colonies), followed by Aglaopheniidae (1278 colonies), Plumulariidae (957 colonies), Campanulariidae (512 colonies), Halopterididae (445 colonies) and Haleciidae (320 colonies) (Fig. 11.3). Sertulariidae also showed the highest occurrence (collected at 53% of the stations where hydroids were recorded), followed by Plumulariidae (49%), Aglaopheniidae (43%), Tiarannidae (40%), Haleciidae (40%) and Campanulariidae (33%) (Fig. 11.4).

Abundance (colonies number) of the main Hydrozoa families collected on the Mauritanian deep shelf and slope

Occurrence (%) of the 18 Hydrozoa families collected on the Mauritanian deep shelf and slope
Species richness was similar within the three sectors considered (north: 20°51′N–19°23′N, centre: 19°23′N–17°40′N and south: 17°40′N–16°04′N) (Ramos et al. 2010), slightly higher in the north (Banc d’Arguin region, 52 species) than in the centre (47 species) and the south (45 species). Thirty-three species were collected along the entire Mauritanian coast, 15 in at least two sectors, and 15 in only one sector (see Annexe 11.1). By station, the species richness varied between 30 and one species and was mostly low: more than 10 species were only recorded at 29 stations, whereas only one or two hydroid species were collected at 77 stations. The distribution of the species richness showed discontinuous latitudinal patterns , peaking off Cape Blanc and on the bottom s adjacent to the submarine canyons off the Banc d’Arguin , north and south Cape Timiris and near the Senegalese border (Fig. 11.5). Moreover, in all these areas the highest figures corresponded to the deep-shelf and upper-slope areas (80–500 m), reaching 1000 m off Northern Cape Timiris and close to the Senegalese border . The lowest values were obtained in some areas off Banc d’Arguin and south of Nouakchott, mainly in deep waters, with the exception of the southernmost area, where intermediate values were obtained between 1000 and 2000 m.

Distribution of the Hydrozoa in Mauritanian waters: species richness (S) (left) and numerical abundances (N, number of colonies) (right)
By station, the abundance of hydroids was mostly low, with only 19 stations recording more than 100 colonies and 93 stations fewer than 10 colonies. The most abundant species were Sertularella gayi gayi (Lamouroux 1821) (1047 colonies) and Aglaophenia lophocarpa Allman 1877 (735 colonies). Another 16 species each accounted for more than 100 colonies (Fig. 11.6). Sertularella gayi gayi showed the highest occurrence value (45%), followed by Streptocaulus dollfussi (Billard 1924), Modeeria rotunda (Quoy & Gaimard 1827), Halecium halecinum (Linnaeus 1758), Nemertesia perrieri (Billard 1901), Clytia paulensis (Vanhöffen 1910) and Racemoramus panicula (G. O. Sars 1874) (20–29%), along with another nine species whose occurrence was higher than 15% (Fig. 11.7). Pictures of some representative hydroid species off Mauritania are shown in Fig. 11.8.

Abundance (N, number of colonies, N > 100) of main Hydrozoa species on the Mauritanian deep shelf and slope

Occurrence (%, >15%) of the main Hydrozoa species on the Mauritanian deep shelf and slope

Photographs of the most representative Hydrozoa species on the Mauritanian deep shelf and slope: 1 Sertularella gayi gayi : colony. 2 S. gayi gayi: colony with gonothecae. 3 Aglaophenia lophocarpa: colony. 4 A. lophocarpa: corbula. 5 A. lophocarpa: hydrocladium. 6 Streptocaulus dollfusi: colony. 7 S. dollfusi: hydrocladium with phylactocarp and gonothecae. 8 Diphasia margareta: colony. 9 Sertularella cylindritheca: colony. 10 Aglaophenia tubulifera: autoepizoism. 11 Nemertesia perrieri : colony. 12 Stegopoma plicatile : colony on Cirripedia . 13 S. plicatile: colony. 14 Modeeria rotunda: colony epibiont on S. gayi gayi. 15 Antennella siliquosa: colony on tube of Polychaeta . 16 Lovenella producta: colony on artificial substratum. 17 L. producta: colony fragment with gonotheca (© Marta Gil)
Distribution of abundances followed the same pattern as species richness , peaking at the same areas and depths (Fig. 11.5). Low abundances recorded in the Banc d’Arguin area must be highlighted, with higher values only at its northern and southern boundaries (Cape Blanc and Cape Timiris, respectively). In addition, at depths of 1500–2000 m, high abundance values were found south of Cape Timiris and intermediate abundances at 17°N.
Types of Substrates
Hydroids collected during Maurit surveys appear to have two different strategies for colonizing soft bottom s off the coast of Mauritania. The first strategy involved morphological adaptations of their hydrorhizae , in which a bundle of interwoven tubules that adhered to muddy or sandy sediments was produced, thereby anchoring colonies to the sea bottom. The second strategy, shared by 50 species, was characterized by use of suitable substrata for fixation of the colonies. It included epizoism on 11 taxa of invertebrates , as well as exploitation of artificial substrata including ghost nets, ropes, fishing lines and plastics.
Thirteen species [Cryptolarella abyssicola (Allman 1888), Halecium beanii (Johnston 1838), Halecium delicatulum Coughtrey 1876, H. halecinum, Lytocarpia myriophyllum (Linnaeus 1758), S. dollfusi, Nemertesia antennina (Linnaeus 1758), Nemertesia perrieri (Billard 1901), Nemertesia ventriculiformis (Marktanner-Turneretscher 1890), Pseudoplumaria marocana (Billard 1930), Diphasia margareta (Allman 1888), S. gayi gayi and Sertularella gayi robusta (Allman 1873)] showed hydrorhizal adaptations to soft bottom s, but only three, C. abyssicola, L. myriophyllum and N. antennina, adopted this strategy exclusively. The remaining species were collected as epizoics on invertebrates , while H. halecinum, S. gayi gayi and S. gayi robusta were also found colonizing artificial substrata . We collected 48 species from different invertebrate groups, of which 29 were exclusively epizoic. Hydrozoa (24%) and Bryozoa (19%) were the most common substrata , followed by Porifera and Polychaeta tubes (11% each), Bivalvia (9%), Cirripedia , Scleractinia , Crustacea , Ascidiacea , Gastropoda and Holothuroidea (5–1%) (Fig. 11.9). Artificial substrata were utilized by 14 species. Of these, only Stegopoma bathyale Vervoort 1966 and Aglaophenia parvula Bale 1882 were limited to such substrates, although several large and dichotomously branched colonies of A. parvula were obtained as detached specimens. Nine species [Acryptolaria conferta conferta (Allman 1877), Bedotella armata (Pictet and Bedot 1900), Cirrholovenia tetranema Kramp 1959, Halecium sessile Norman 1866, Halopteris diaphana (Heller 1868), Nemertesia ramosa (Lamarck 1816), Sertularella mediterranea Hartlaub 1901, Sertularella polyzonias (Linnaeus 1758) and Streptocaulus multiseptatus (Bale 1915)] were collected exclusively as detached colonies. Information about their substrates is therefore unavailable.

Occurrence (%) of Hydrozoa on different types of substrata
The most obvious substrate generalists were Sertularella gayi gayi (nine different substrata), A. lophocarpa, C. paulensis, and S. dollfusi (seven substrata), Halecium sibogae marocanum Billard (1934) and R. panicula (six substrata), Antennella secundaria (Gmelin 1791), H. delicatulum, H. halecinum, Obelia dichotoma (Linnaeus 1758), Stegopoma plicatile (M. Sars 1863) and T. dorhnii (five substrata). Notably, they were among the most abundant and most frequent of the species.
Bathymetric Distribution
Of 63 species collected, four were found exclusively in the coastal realm (6%) and 18 exclusively in the deep benthic realm (29%), with the remaining 41 being recorded from both realms (65%) (Annexe 11.1). In consequence, the hydroid fauna of the shelf is composed of 45 species whereas that of the slope is comprises 59 species (Fig. 11.10).

Bathymetric distribution of the 63 Hydrozoa species collected on the Mauritanian deep shelf and slope (solid lines our data; dotted lines data extracted from the literature)
Bougainvillia muscus (Allman 1863), E. album, Hydractinia multitentaculata (Millard 1975) and N. ramosa were collected only on the deep shelf.
Among the 18 species restricted to the slope, Leukartiara octona (Fleming 1823), Egmundella superba Stechow 1921, C. tetranema, A. conferta conferta, S. multiseptatus, S. mediterranea, B. armata, Plumularia filicula Allman 1877, Kirchenpaueria bonnevieae (Billard 1906), Polyplumaria flabellata (Sars 1873), S. bathyale, Zygophylax levinseni (Saemundsson 1911) and Halecium tenellum Hincks 1861 were recorded on the upper slope at depths between 200 and 800 m. Lovenella producta (G. O. Sars 1874) and H. sessile were found on the middle slope (800–1500 m), with C. abyssicola collected only at 1500 m. Stegopoma plicatile and Acryptolaria conferta minor (Ramil and Vervoort 1992) were distributed along the entire slope. The remaining species occurred over wide bathymetric ranges, encompassing both realms.
Analysis of bathymetric distributions based on our data and those extracted from the literature shows that only 13 of 81 species known from Mauritania are stenobathic , with ranges less than 300 m: B. muscus, Pennaria disticha Goldfuss 1820, Hydractinia marsupialia (Millard 1975), H. multitentaculata, Mitrocomella polydiademata (Romanes 1876), D. disticha, Sertularella ellisii (Deshayes and Milne Edwards 1836), S. marginata, Campanularia volubilis (Linnaeus 1758), Laomedea flexuosa Alder 1857, Laomedea pseudodichotoma Vervoort (1959), Aglaophenia octodonta (Heller 1868) and Clytia hemisphaerica (Linnaeus 1767). Bathymetric ranges of the remaining species exceeded 300 m and, in consequence, are considered eurybathic.
Geographical Distribution
Analysis of geographical distribution patterns of the hydroids collected off Mauritania revealed two main components (Fig. 11.11). The first and most representative group, accounting for 58% of total species, included ones with a wide distribution (cosmopolitan, circumglobal , circumtropical and non-classifiable) . The second group, composed of Atlantic species (Atlantic-Mediterranean , amphi-Atlantic , eastern Atlantic and boreo-Atlantic), accounted for 42% of all species. The most important contingent comprised circumglobal species (32%), followed by cosmopolitan and Atlantic-Mediterranean (both with 17%) and amphi- and (both with 9.5%). The remaining three contingents (boreo-Atlantic, circumtropical and non-classifiable) each represented 5%.

Biogeographic components (%) of Mauritanian Hydrozoa . C cosmopolitan, CG circumglobal , CT circumtropical , AM Atlantic-Mediterranean, AA amphi-Atlantic, EA eastern-Atlantic, BA boreo-Atlantic, NC non-classifiable
Discussion
The coast of Mauritania harbours an especially productive marine ecosystem , the result of permanent upwelling conditions off Cape Blanc (Arístegui et al. 2009). In addition, its location on the border between the warm-temperate Lusitanian Province and the Tropical Eastern Atlantic Province (Briggs 1995; Briggs and Bowen 2012) represents a particularly interesting area for biogeographical studies , displaying along the Mauritanian or Senegalese continental shelves the turnover between Mediterranean -Atlantic and tropical West African biotas (Van Soest 1993). However, benthic invertebrates , and especially sessile epifauna , are little known in northwest Africa (Decker et al. 2004), and the southern distributional limits of many well-known North Atlantic species remain uncertain. Moreover, the predominance of soft bottom s on the Mauritanian continental shelf does not encourage study of sessile epifauna (Van Soest 1993). Although hydroids are considered characteristic and abundant representatives of sessile epifauna on hard bottom s in shallow-water communities (Boero 1984; Boero and Fresi 1986; Gili and Hughes 1995; Altuna 2007), their ability to colonize soft bottom s, linked to their capacity to settle on a wide variety of substrates or to anchor directly in muddy or sandy bottom s, has also been pointed out by several authors (Gili et al. 1987, 1989; Calder 1991; Gili and Hughes 1995; Ramil et al. 1997; Peña Cantero 2010; Di Camillo et al. 2013). Results from the intensive sampling programme deployed during the four Maurit surveys highlighted this capacity, as a total of 63 species were collected mainly from muddy bottoms and showed a similar distribution pattern along the entire Mauritanian coast. This diversity is comparable to that obtained by Vervoort (1959), with 60 species collected during the Atlantide Expedition in tropical West Africa , and by Gili et al. (1989), with 50 species obtained during seven exploratory cruises in Guinea Bissau , Namibia and South Africa . Therefore, if we compare the latitudinal range sampled in those surveys, both are much wider than that of the entire Mauritanian coast. Hydroid diversity in Mauritania nevertheless seems to be higher than in more southern tropical areas . However, it is lower than the diversity reported from the Moroccan coast by Patriti (1970) and CANCAP expedition papers (86 species), and from the Ibero-Moroccan Bay by Ramil and Vervoort (1992: 72 species). Moreover, preliminary results from three surveys carried out between 2004 and 2006 in Moroccan and Saharan waters (Maroc surveys) highlighted the existence of rich suspension-feeder communities, and reported a higher hydroid diversity along the Atlantic Moroccan slope, mainly between Gibraltar and Agadir (Ramil, University of Vigo , unpublished data, 2005). Hydroid diversity along the West African coast does not accord with the paradigm of a cline of increasing diversity from poles to tropics in the North Atlantic (Gray 2001). Instead, it coincides with results obtained from the coasts of South Africa and Namibia by Millard (1978) and Gibbons et al. (2010), in which hydroid diversity declines northwards to the tropics. The reverse is known to occur along the east coast of South Africa , with diversity strongly increasing northwards. Low diversity along the west coast of South Africa and Namibia was attributed to the absence of suitable hard substrata (Gibbons et al. 2010). However, our results, which confirm the ability of many hydroid species to colonize soft bottom s, suggests that their diversity can be driven by other factors that clearly influence biotic diversity on the continental margins, such as local hydrological features , bottom heterogeneity and food input (Levin and Sibuet 2012).
The capacity of hydroids , as substratum generalists, to colonize a wide range of substrata enhances their adaptation to different ecological conditions, as already noted by several authors (i.e., Boero 1984; Gili and Hughes 1995; Genzano et al. 2009; Ronowicz et al. 2013). Another strategy, modification of the hydrorhiza for anchoring the colony to muddy or sandy substrata , is also displayed by several genera and families (see Di Camillo et al. 2013) and, together with their substratum generalist character, enhances the diversity of hydroids on soft bottom s along the deep shelf and continental slope . Our results show that the most frequent and therefore most widespread species off Mauritania are those that use both strategies (substratum generalists and hydrorhiza modifiers) , such as S. gayi gayi (the most frequent and also most abundant), S. dollfusi, H. halecinum and some species of Nemertesia. Nevertheless, small epizoic species living on large hydroids , such as M. rotunda, C. paulensis, R. panicula, A. secundaria and Antennella siliquosa (Hincks 1877), were also very frequent and/or abundant. The most colonized invertebrates were hydroids , including auto-epizoism within the Plumularioidea (sensu Millard 1973), and Bryozoa (mainly erect colonies). The importance of large colonies as substrata for other hydroids , reducing both the competition for space and the negative effects of sedimentation , has been highlighted in several works (see Genzano et al. 2009). Moreover, the condition of habitat formers and ecosystem engineers , reported for several invertebrates such as large sponges , octocorals or scleractinians , among others, was also recently applied to L. myriophyllum (Di Camillo et al. 2013) because of their capacity to stabilize sediments, create secondary substrata and increase habitat complexity. On the Mauritanian slope , another 12 large hydroid species with modified hydrorhizae for anchoring in soft-bottom s (see Annexe 11.1) were heavily colonized by hydroids as well as by other epifaunal representatives. In all likelihood, these species play the same role here as those described for L. myriophyllum in the Mediterranean .
As for bathymetric distributions, previous works (Millard 1978; Calder 1996, 1998; Schuchert 2001; Altuna 2007; Ronowicz et al. 2013) have shown that the highest species richness is located in the upper 200 m worldwide, related to the most suitable environment features (hard substrata and food availability among others) (Boero 1984; Calder 1991, 1998), but also influenced, at least in part, by a greater sampling effort deployed in coastal waters (Calder 1998; Altuna 2007). Conversely, in Mauritanian waters where hard bottom s are scarce and the greatest sampling effort was deployed in deep waters, the figures are different: of a total of 63 hydroid species, 58 (92%) were collected along the slope versus 47 (75%) on the shelf, with only four species recorded exclusively on the shelf. By adding data from the literature, mainly from shallow waters in the Banc d’Arguin area, the current checklist of Mauritanian hydroids rises to 81 species (Annexe 11.1 and 11.2) and shows the same general figure. Also remarkable is the eurybathic condition of Mauritanian hydroids , with their depth ranges exceeding 300 m (see Altuna 2007). Only 24 (38.7%) of the 63 species collected during the Maurit surveys had bathymetric ranges narrower than 300 m, but many of them were recorded only once. If we consider the complete checklist and also the data extracted from literature, stenobathic species are reduced to eight (11.6%) (Fig. 11.10, Annexe 11.1).
These data contrast sharply with the dominance of stenobathic species (66%) in the Bay of Biscay (Altuna 2007), although the importance of eurybathic species in deep waters was pointed out by Calder (1997) in the north-western Atlantic and also by Peña Cantero (2004, 2014) in the Southern Ocean. Nevertheless, if we consider mean values of species richness and the Shannon-Weaver index plotted by 200 m depth strata , the highest values were obtained in the shallowest stratum and declined sharply below the 500 m isobath , concurring with previous results that demonstrate a progressive reduction of diversity below 100 m (Fig. 11.12a). This reduction is more apparent along the continental slope at different depths (250–900 m), depending on the area considered (Millard 1978; Stepanjants 1989; Calder 1996, 1998; Altuna 2007). Abundance also peaks on the deep shelf and falls abruptly to 700 m (Fig. 11.12b). Previous studies on deep hydroids are mostly taxonomic works and abundance data are not reported. Only Gili et al. (1987) reported abundance data for hydroids collected with a commercial trawl on the Catalan continental shelf (NE Spain) between 5 and 870 m, where they collected 2261 colonies in 41 hauls of ca. 2 km2 each.

Specific richness (S, species number) and Shannon-Wiener index (H´) (a) and Abundance (N, number colonies) (b) plotted by 200 m strata
In Maurit surveys , each trawl sampled around 0.1 km2 and we recorded 6169 colonies in 174 hauls. A rough comparison of mean values between our data and those obtained by Gili et al. (1987) clearly reveal differences in hydroid abundances: ca. 28 colonies per km2 on the Catalan coast versus ca. 355 colonies per km2 off Mauritania. The low abundances obtained by Gili et al. (1987) were correlated with the extreme degradation of intensely exploited fishing grounds . Although Mauritanian fishing grounds have also been traditionally exploited by international trawl fleets for more than 50 years (Sobrino and García 1992), operating up to 1000 m depth (FAO 2006, 2012), we have found relatively high values of hydroid abundance . Higher abundances in Mauritania were also reported for other benthic invertebrates compared with those obtained in the North Sea by Duineveld et al. (1993).
Species richness and abundance distributions along the Mauritanian coast (Fig. 11.5) show similar patterns, with highest values recorded on the shelf break and upper slope , mainly around canyons.
Canyons are sources of habitat heterogeneity with high benthic productivity , since they provide suitable areas for recruitment and development of benthic fauna (Ramírez-Llodra et al. 2010; Vetter et al. 2010). Also, their current patterns favour development of suspension-feeder communities (Rowe 1971). The distributional pattern of hydroids suggests that canyons harbour source populations which colonize nearby areas following the source-sink hypothesis , proposed by several authors to explain the maintenance of diversity in non-favourable environments (for review see Rex et al. 2005). On the Mauritanian slope , hydroid density probably allows for successful reproduction, but these populations can be dramatically and periodically disturbed by intense fishing activity. The existence of source populations in the canyons and the generalist nature of many hydroid species allow for a rapid and continuous recolonization of the adjacent slope areas , and could explain the abundance and distribution patterns of hydroids in this area.
Biogeographical analyses of Mauritanian hydroids have been carried out previously only by Ansín Agís et al. (2001), whose work was limited exclusively to the superfamily Plumularioidea. Two main groups were recognized, species limited to the Atlantic (54.5%) and species with a wide distributions (45.5%). Analysis of geographic distribution patterns of all hydroid species collected during the four Maurit surveys highlights the importance of both components, although species with wide distributions prevail (59%) compared with Atlantic species (41%).
The importance of Atlantic-Mediterranean species in Mauritania has already been noted by Ansín Agís et al. (2001) for Plumularioidea and also by Van Soest (1993) for sponges . In our collections , only 17% were Atlantic-Mediterranean species. However, together with other hydroids widely distributed in the northeastern Atlantic (boreo-Atlantic and eastern Atlantic), they account for about 30%. This percentage emphasizes the importance of the systems of currents, and mainly the influence of the Canary Current on the southward dispersion of sessile epifaunal species (Ansín Agís et al. 2001; Van Soest 1993).
Eastern Atlantic species apparently seldom colonize the Americas, so the source of amphi-Atlantic species is taken to be the Western Atlantic Region (Briggs 1995). Notably, the Azores Current , which reaches the continental slope of Morocco and joins the Canary Current (Fernández and Pingree 1996), is considered an extension of the Gulf Stream . This possible link to the western North Atlantic might explain the presence of amphi-Atlantic species in Mauritania. In addition, the Equatorial counter-current also allows eastward dispersion of Western Atlantic species (Briggs 1995). In addition, the Mauritanian Current moves northward in winter to the latitude of Cape Blanc (Stramma and Schott 1999; Pastor et al. 2008), potentially contributing to the amphi-Atlantic contingent in this area.
Noteworthy is the distribution of S. plicatile in the Eastern Atlantic , where it was reported from Arctic and boreal seas (Calder 2012) to the west of Brest, France (Cornelius 1995), and also in West Africa from Mauritania (present work) and Namibia (Gili et al. 1989), but not from the Bay of Biscay (Altuna 2007), Iberian Peninsula (Medel and López-González 1996), Ibero-Moroccan Bay (Ramil and Vervoort 1992) or Macaronesian waters (Vervoort 2006). This distributional pattern suggests a possible origin of West African populations in the Western Atlantic , where S. plicatile is known from Florida (Henry et al. 2008). We also hypothesized the same origin for E. superba, reported from the Western Atlantic (Stechow 1921; Calder 1991) and off the Azores (Leloup 1940 as Campanulina producta), but not from African or European coasts. Finally, we emphasize that only one species, H. multitentaculata, is exclusively distributed along the West African coast , from Natal (South Africa) to Mauritania (Vervoort 2006; present work), which supports the idea that the current system in this area prevents the northward dispersion of benthic tropical species (Arístegui and Cruz 1986; Hernández et al. 2013).
Conclusions
The hydroid fauna of Mauritania is relatively diverse , and similar in richness to other tropical regions of West Africa despite the predominance of soft bottom s in the study area.
Hydroids from submarine canyons probably act as source populations , enabling recolonization of adjacent sea bottom s after fishing impacts and thus maintaining diversity.
The most abundant and frequent species are those that have developed morphological adaptations of their hydrorhizae as well as epizoic strategies , both favouring the colonization of soft bottom s. Small species, living mainly as epibionts on other hydroids , are also frequent and abundant.
Bathymetric distributions reveal higher diversity on the slope (59 species) than on the shelf (45 species), and reflect the eurybathic character of the hydroid fauna of Mauritania.
Biogeographical components are dominated by species with a wide distribution (mainly cosmopolitan and circumglobal ). However, Atlantic species, many of them widely distributed in the Northeastern Atlantic, were also well represented.
References
Allman GJ (1883) Report on the Hydroida dredged by H.M.S. Challenger during the years 1873–76. Part I Plumularidae. Rep Sci Res Voy HMS Challenger. Zool 7(20):1–55
Allman GJ (1888) Report on the Hydroida dredged by H.M.S. Challenger during the years 1873–76. Part II The Tubularinae, Corymorphinae, Campanularinae, Sertularinae and Thalamorphora. Rep Sci Res Voy HMS Challenger. Zool 23(70):i–lxix
Altuna A (2007) Bathymetric distribution patterns and biodiversity of benthic Medusozoa (Cnidaria) in the Bay of Biscay (north-eastern Atlantic). J Mar Biol Assoc UK 87:681–694
Altuna A (2014) Bibliografía de los cnidarios (Cnidaria) de la Península Ibérica e Islas Baleares. http://www.faunaiberica.es/faunaib/altuna8.pdf. Accessed 20 Oct 2014
Ansín Agís JA (1998) Plumularoidea (Cnidaria, Hydrozoa) recolectados por las expediciones holandesas “CANCAP” en el Atlántico nor-oriental. Ph.D. Dissertation, University of Vigo
Ansín Agís J, Ramil F, Vervoort W (2001) Atlantic Leptolida (Hydrozoa, Cnidaria) of the families Aglaopheniidae, Halopterididae, Kirchenpaueriidae and Plumulariidae collected during the CANCAP and Mauritania-II expeditions of the National Museum of Natural History, Leiden, the Netherlands. Zool Verh Leiden 333:1–268
Arístegui J, Cruz T (1986) Consideraciones biogeográficas sobre el orden Cheilostomata (Ectoprocta) en Canarias. Vieraea 16:161–171
Arístegui J, Barton ED, Álvarez-Salgado XA, Santos AMP, Figueiras FG, Kifani S, Hernández-León S, Mason E, Machú E, Demarcq H (2009) Sub-regional ecosystem -variability in the Canary Current upwelling. Prog Oceanogr 83(1–4):33–48
Bedot M (1921) Hydroïdes provenant des campagnes des yachts Hirondelle et Princesse-Alice (1887 à 1912). I. Plumularidae. Res Camp Scient Albert Ier Prince Monaco 60:1–73
Billard A (1906a) Hydroïdes. In: Expéditions scientifiques du ‘Travailleur’ et du ‘Talisman’ 1906, Paris, pp 153–243
Billard A (1906b) Hydroïdes. In: Mission des pêcheries de la côte occidental d’Afrique, III. Act Soc Linn Bordeaux 61:173–180
Billard A (1907) Hydroïdes de Madagascar et du Sud-Est de l’Afrique. Arch Zool Exp Gen 4me sér 7(8):335–396
Billard A (1924) Note sur une espèce nouvelle de Plumularide des côtes du Maroc. (Cladocarpus dollfusi). Bull Soc Zool Fr 49:87–89
Billard A (1927) Hydrozoa I. Hydrozoa benthonica (excl Hydrocorallidae) In: Monod T (ed) Contribution à l’étude de la faune du Cameroun. Faune Col françaises 1(6):467–474
Billard A (1930) Note sur deux espèces d’hydroïdes de la côte Atlantique du Maroc. Bull Soc Sci Nat Maroc 10:79–80
Billard A (1931) Hydroïdes de Mauritanie. Bull Mus natn Hist nat 2me sér 3(7):673–678
Billard A (1934) Notes sur quelques hydroïdes du Maroc. Bull Soc Zool Fr 59:227–231
Boero F (1984) The ecology of marine hydroids and effects of environmental factors: A review. Mar Ecol 5(2):93–118
Boero F, Fresi E (1986) Zonation and evolution of a rocky bottom hydroid community. Mar Ecol 7(2):123–150
Bouillon J, Medel MD, Pagès F, Gili JM, Boero F, Gravili C (2004) Fauna of the mediterranean Hydrozoa. Sci Mar 68(2):5–438
Briggs JC (1995) Global biogeography. Developments in palaeontology and stratigraphy, vol 14. Elsevier Science, Amsterdam
Briggs JC, Bowen W (2012) A realignment of marine biogeographic provinces with particular reference to fish distributions. J Biogeogr 39:12–30
Broch H (1913) Hydroida from the “Michael Sars” North Atlantic Deep-Sea Expedition 1910. Rep Sci Res ‘Michael Sars’ N Atl Deep-Sea Exped Zool 3(1):1–18
Broch H (1914) Hydrozoa benthonica. Beitr Kennt Meeresfauna Westafr 1:19–50
Buchanan JB (1956) Contributions to the hydroid fauna of the Cameroons. Rev Zool Bot Afr 53(3–4):276–280
Buchanan JB (1957) The hydroid fauna of the Gold coast. Rev Zool Bot Afr 56(3–4):349–372
Calder DR (1991) Abundance and distribution of hydroids in a mangrove ecosystem at Twin cays, Belize, Central America. In: Williams RB, Cornelius PFS, Hughes RG, Robson EA (eds) Coelentare biology: Recent research on Cnidaria and Ctenophora. In: Proceedings of the fifth international conference on coelenterate biology, 1989. Dordrecht, Boston, London, pp 221–228
Calder DR (1996) Hydroids (Cnidaria: Hydrozoa) recorded from depths exceeding 3000 m in the abyssal western North Atlantic. Can J Zool 74:1721–1726
Calder DR (1997) Synopsis of hydroids from 1000 m and deeper in the western North Atlantic. In: den Hartog JC (ed) Proceedings of the 6th international conference on coelenterate—biology, The Leenwenhorst, Noordwijkerhout, July 1995. Coelenterate Biology, Leiden, National Natuurhistorisch Museum, pp 85–90
Calder DR (1998) Hydroid diversity and species composition along a gradient from shallow waters to deep sea around Bermuda. Deep-Sea Res PT I 45:1843–1860
Calder DR (2012) On a collection of hydroids (Cnidaria, Hydrozoa, Hydroidolina) from the west coast of Sweden, with a checklist of species from the region. Zootaxa 3171:1–77
Cornelius PFS (1995) North-West European thecate hydroids and their medusae. Part 1. In: Barnes RSK, Crothers JH (eds) Synopses of the British Fauna (New Series), vol 50, pp 1–347
Decker C, Griffiths C, Prochazka K, Ras C, Whitfield A (eds) (2004) Marine biodiversity in Sub-Saharan Africa: the Known and the unknown. In: Proceedings of the Marine biodiversity in Sub-Saharan Africa: The known and the unknown, Cape town, 23–26 Sept 2003
Di Camillo CG, Boero F, Gravili C, Previati M, Torsani F, Cerrano C (2013) Distribution, ecology and morphology of Lytocarpia myriophyllum (Cnidaria: Hydrozoa), a Mediterranean Sea habitat former to protect. Biodivers Conserv 22:773–789
Duineveld GCA, Lavalaye MSS, Van Noort GJ (1993) The trawl fauna of the Mauritanian shelf (Northwest Africa): density, species composition and biomass. In: Wolff WJ, Van Der Land J, Nieuhuis PH, de Wilde PAWJ (eds) Ecological studies in the coastal waters of Mauritania: proceedings of a symposium held at Leiden, The Netherlands, March 1991. Hydrobiologia, vol 258, pp 165–174
FAO (2006) Report of the FAO/CECAF working group on the assessment of demersal resources—subgroup North; Saly, Senegal, 14–23 Sept 2004. FAO CECAF/ECAF Ser 06/68. FAO, Rome
FAO (2012) Report of the FAO/CECAF working group on the assessment of demersal resources—subgroup North. Agadir, Morocco, 8–17 Feb 2010. FAO CECAF/ECAF series 11/72. FAO, Rome
Fernández E, Pingree RD (1996) Coupling between physical and biological fields in the North Atlantic subtropical front southeast of the Azores. Deep-Sea Res PT I 43(9):1369–1393
García Carrascosa AM (1981) Hidrozoos tecados (Hydrozoa Calyptoblastea) de las costas mediterráneas españolas: faunística, ecología, bionomia bentica y biogeografía. Ph.D. Dissertation University of Valencia
Genzano GN, Giberto D, Schejter L, Bremec C, Meretta P (2009) Hydroid assemblages from the Southwestern Atlantic Ocean (34–42°S). Mar Ecol 30:33–46
Gibbons MJ, Buecher E, Thibault-Botha D, Helm RR (2010) Patterns in marine hydrozoan richness and biogeography around southern Africa: implications of life cycle strategy. J Biogeogr 37:606–616
Gili JM, Hughes RG (1995) The ecology of marine benthic hydroids. Oceanogr Mar Biol 33:351–426
Gili JM, Ros JD, Pagés F (1987) Types of bottoms and benthic cnidarian from the trawling grounds (Littoral and Bathyal) off Catalonia (NE Spain). Vie Milieu 37:85–98
Gili JM, Vervoort W, Pagés F (1989) Hydroids from the West African coast: Guinea Bissau, Namibia and South Africa. Sci Mar 53(1):67–112
Gray JS (2001) Marine diversity: the paradigms in patterns of species richness examined. Sci Mar 65:41–56
Henry LA, Nizinski MS, Ross SW (2008) Occurrence and biogeography of hydroids (Cnidaria: Hydrozoa) from deep-water coral habitats off the southeastern United States. Deep-Sea Res PT I 55(6):788–800
Hernández JC, Clemente S, Tuya F, Pérez-Ruzafa A, Sangil C, Moro-Abad L, Bacallado-Aránega JJ (2013) Echinoderms of the Canary Islands, Spain. In: Alvarado JJ, Solís-Marín FA (eds) Echinoderm research and diversity in Latin America. Springer, Berlin, Heidelberg, pp 471–510
Iazza B, González-Duarte MM, Moukrim A, Martínez CM (2013) First report of the marine hydroids Eudendrium glomeratum, E. merulum and Garveia grisea (Cnidaria: Hydrozoa) from the Moroccan Atlantic coast. Mar Biodivers Rec 6:1–6
Izquierdo MS, García-Corrales P, Bacallado JJ (1986a) Contribución al conocimiento de los hidrozoos caliptoblástidos del Archipiélago Canario. Parte I: Haleciidae, Lafoeidae, Campanulariidae y Syntheciidae. Bol Inst Esp Oceanogr 3(1):81–94
Izquierdo MS, García-Corrales P, Bacallado JJ (1986b) Contribución al conocimiento de los hidrozoos caliptoblástidos del Archipiélago Canario. Parte II: Plumulariidae. Bol Inst Esp Oceanogr 3(2):49–66
Izquierdo MS, García-Corrales P, Bacallado JJ, Vervoort W (1990) Contribución al conocimiento de los hidrozoos caliptoblástidos del Archipiélago Canario. Parte III: Sertulariidae. Bol Inst Esp Oceanogr 6(2):29–40
Leloup E (1937) Hydroidea, Siphonophora, Ceriantharia. I. Hydropolypes. In: Résultats scientifiques des croisières du navire-école belge “Mercator”. Mém Mus R Hist Nat Belg 2me sér 9:91–121
Leloup E (1940) Hydropolypes provenant des croisières du Prince Albert Ier de Monaco. Rès Camp Scient Albert Ier de Monaco 140:1–38
Levin LA, Sibuet M (2012) Understanding continental margin biodiversity: a new imperative. Annu Rev Mar Sci 4:79–112
Medel MD, López-González PJ (1996) Updated catalogue of hydrozoans of the Iberian Peninsula and Balearic Islands, with remarks on zoogeography and affinities. In: Piraino S, Boero F, Bouillon J, Cornelius PFS, Gili JM (eds) Advances in hydrozoan biology. Sci Mar 60(1):183–209
Medel MD, Vervoort W (1998) Atlantic Thyroscyphidae and Sertulariidae (Hydrozoa, Cnidaria) collected during the CANCAP and Mauritania-II expeditions of the National Museum of Natural History, Leiden, The Netherlands. Zool Verh Leiden 320:1–85
Medel MD, Vervoort W (2000) Atlantic Haleciidae and Campanulariidae (Hydrozoa, Cnidaria) collected during the CANCAP and Mauritania-II expeditions of the National Museum of Natural History, Leiden, The Netherlands. Zool Verh Leiden 330:1–68
Millard NAH (1973) Auto-epizoism in South African hydroids. In: Proceedings of the second international symposium on Cnidaria. Publ Seto Mar Biol Lab 20:23–34
Millard NAH (1978) The geographical distribution of southern African hydroids. Ann S Afr Mus 74(6):159–200
Pastor MV, Pelegrí JL, Hernández-Guerra A, Font J, Salat J, Emelianov M (2008) Water and nutrient fluxes off Northwest Africa. Cont Shelf Res 28:915–936
Patriti G (1970) Catalogue des cnidaries et cténaires des côtes Atlantiques marocaines. Trav Inst Scient Cherifien (Zool) 35:1–149
Peña Cantero AL (2004) How rich is the deep-sea Antarctic benthic hydroid fauna? Polar Biol 27:767–774
Peña Cantero AL (2010) Benthic hydroids (Cnidaria: Hydrozoa) from Peter I Island (Southern Ocean, Antarctica). Polar Biol 33:761–773
Peña Cantero AL (2014) Chapter 5.6. Benthic hydroids (Cnidaria, Hydrozoa) In: De Broyer C, Koubbi P, Griffiths HJ, Raymond B, d’Udekem d’Acoz C, Van de Putte A, Danis B, David B, Grant S, Gutt J, Held C, Hosie G, Huettmann F, Post A, Ropert-Coudert Y (eds) Biogeographic Atlas of the Southern Ocean. Scientific Committee on Antarctic Research, Cambridge, pp 103–106
Picard J (1951) Hydraires littoraux du Sénégal récoltés par H. Sourie aux environs de Dakar. Bull Inst fr Afr Noire 13(1):109–115
Pictet C, Bedot M (1900) Hydraires provenant des campagnes de l’ “Hirondelle” (1886–1888). Res Camp Scient Albert Ier Prince Monaco 18:1–59
Quelch JJ (1885) On some deep-sea and shallow water Hydrozoa. Ann Mag Nat Hist 16:1–20
Ramil F, Vervoort W (1992) Report on the Hydroida collected by the “BALGIM” expedition in and around the Strait of Gibraltar. Zool Verh Leiden 227:3–262
Ramil F, Ansín Agís J, Fernández Pulpeiro E (1997) Soft-bottom hydroids (Cnidaria: Hydrozoa) collected in the Ría de Vigo (NW Spain). Zool Verh Leiden 323:181–208
Ramírez-Llodra E, Company JB, Sardá F, Rotllant G (2010) Megabenthic diversity patterns and community structure of the Blanes submarine canyon and adjacent slope in the Northwestern Mediterranean: a human overprint? Mar Ecol 31:167–182
Ramos A, Alcalá C, Fernández F, Fernández-Peralta L, González-Porto M, López V, Moya JA, Pascual P, Presas C, Puerto MA, Ramil F, Salmerón F, Sanz JL, Rey J, Viscasillas L, Abed JO, Baye SO, Ciré BA, Mohamed BO, Samba AO, Valy YO (2010) Estudio de los ecosistemas de la plataforma y margen continental de Mauritania. Informe de resultados de la campaña ‘Maurit-0911’. Report IEO-IMROP, Vigo
Rees WJ, Thursfield S (1965) The hydroid collections of James Ritchie. Proc R Soc Edinb (B) 69(1–2):34–220
Rex MA, McClain CR, Johnson NA, Etter RJ, Allen JA, Bouchet P, Warén A (2005) A source-sink hypothesis for abyssal biodiversity. Am Nat 165(2):163–178
Ritchie J (1907) On collections of the Cape Verde Islands marine fauna, made by Cyril Crossland of St Andrews University, July to September 1904. The hydroids. Proc Zool Soc Lond 1907:488–514
Ronowicz M, Wlodarska-Kowalczuk M, Kukliríski P (2013) Depth- and substrate-related pattems of species richness and distribution of hydroids (Cnidaria, Hydrozoa) in Arctic coastal waters (Svalbard). Mar Ecol 34(Suppl 1):165–176
Rowe GT (1971) Observations on bottom currents and epibenthic populations in Hatteras submarine canyon. Deep-Sea Res 18:569–581
Schuchert P (2001) Hydroids of Greenland and Iceland (Cnidaria, Hydrozoa) Meddelelser om Grønland, Bioscience 53:1–184
Schuchert P (2012) North-West European Athecate hydroids and their Medusae. In: Crothers JH and Hayward PJ (eds) Synopses of the British Fauna (New Series) 59:1–364
Sobrino I, García T (1992) Análisis y descripción de las pesquerías de crustáceos decápodos en aguas de la República Islámica de Mauritania durante el periodo 1987–1990. Inf Téc Inst Esp Oceanogr 112:1–38
Stechow E (1921) Über Hydroiden der Deutschen Tiefsee-Expedition, nebst Bemerkungen über einige andre Formen. Zool Anz 56(5–6):97–119
Stechow E (1925) Hydroiden der Deutschen Tiefsee-Expedition. Wiss Ergebn dt Tiefsee-Exped ‘Valdivia’ 1898–1899, 17:383–546
Stepanjants SD (1989) Hydrozoa of the Eurasian Artic Seas. In: Herman Y (ed) The Artic Seas. Climatology, oceanography, geology and biology. Van Nostrand Reinhold, New York, pp 397–430
Stramma L, Schott F (1999) The mean flow field of the tropical Atlantic Ocean. Deep-Sea Res PT II 46:279–303
Van Soest RWM (1993) Distribution of sponges on the Mauritanian continental shelf. In: Wolff WJ, Van der Land J, Nienhuis PH, Wilde PAW (eds) Ecological studies in the Coastal waters of Mauritania. Hydrobiologia 258:95–106
Vervoort W (1946) Exotic hydroids in the collections of the Rijksmuseum van Natuurlijke Historie and the Zoological Museum at Amsterdam. Zool Med Leiden 26(1–4):287–351
Vervoort W (1959) The Hydroida of the tropical west coast of Africa. Atlantide Report Scient Res Danish Exped coasts trop W Afr 1945–1946, 5:211–325
Vervoort W (2006) Leptolida (Cnidaria: Hydrozoa) collected during the CANCAP and Mauritania-II expeditions of the National Hystory, Leiden, the Netherlands [Anthoathecata, various families of Leptothecata and addenda]. Zool Med Leiden 80:181–318
Vetter EW, Smith CR, De Leo FC (2010) Hawaiian hotspots: enhanced megafaunal abundance and diversity in submarine canyons on the oceanic islands of Hawaii. Mar Ecol 31:183–199
Acknowledgements
The authors wish to express their sincere gratitude to their friend and colleague, geologist Luis Miguel Agudo, for preparing the maps. We also acknowledge Uxía Tenreiro and Alexia Luiña, the friendly librarians of Oceanographic Centre of Vigo, whose effectiveness allows us to always have all the scientific literature on hydrozoans necessary for our taxonomic research.
Our sincere thanks to the referees, Dr Dale R. Calder from the Department of Natural History of Royal Ontario Museum (Canada), and Dr Alvaro Peña-Cantero from the University of Valencia (Spain) for his insightful comments that have greatly improved this manuscript.
This work was undertaken within the framework of the EcoAfrik project and has been partially funded by the MAVA Fondation pour la Nature (MAVA contract 12/87 AO C4/2012). This is ECOAFRIK publication number 12.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Annexes
Annexes
Rights and permissions
Copyright information
© 2017 Springer Science+Business Media B.V
About this chapter
Cite this chapter
Gil, M., Ramil, F. (2017). Hydrozoans from Mauritanian Deep-Waters. In: Ramos, A., Ramil, F., Sanz, J. (eds) Deep-Sea Ecosystems Off Mauritania. Springer, Dordrecht. https://doi.org/10.1007/978-94-024-1023-5_11
Download citation
DOI: https://doi.org/10.1007/978-94-024-1023-5_11
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-024-1021-1
Online ISBN: 978-94-024-1023-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)