
Abstract
The current knowledge about the ammonoid/habitat relation is reviewed and in part newly interpreted. The autecology of ammonoids, such as ontogeny and habitat, based on morphological and geochemical analyses in association with results from modern relatives, forms the foundation for subsequent interpretations. Synecological interactions (predator-prey, infestation) are discussed with reference to sedimentary facies and the corresponding biofacies. Arguments for a possible mode of life and habitat are given based on the modern data on the food/prey habits and predation habitats of ammonoids. The state of the art in scientific investigations on ammonoid life and habitat is summarized, reviewed and in part reinterpreted. Traditional assumptions based on facies analyses are strengthened or contradicted by more recent methods such as morphospace and stable isotope analyses. Not yet tested hypotheses, speculations and mathematical models are tested by comparing the results with new geophysical data.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Ammonoid paleobiology has been a topic of interest for many years and an enormous amount of knowledge and data on extant (Nautilus, Octopus, Sepia, Spirula) and fossil cephalopods (ammonoids, nautiloids, belemnoids, teuthoids) has been compiled in recent decades (Arkell 1957; House and Senior 1981; Wiedmann and Kullmann 1988; House 1993a; Landman et al. 1996a; Payne et al. 1998; Olóriz and Rodríguez–Tovar 1999; Summesberger et al. 2002; Landman et al. 2007a; Tanabe et al. 2010a; Ritterbush et al. 2014). Reconstructing the life history and habitats of extinct groups is one of the classic problems. Ammonoids are an extinct group within the Cephalopoda with an external aragonitic shell (ectocochleate). Various shell morphologies exist, ranging from planispiral to heteromorphic shells with straight, openly coiled, helicospiral shells or even various combinations of these. The shell sculpture varies from smooth (‘Leiostraca’; e.g., Phylloceras) to strongly ribbed (‘Trachyostraca’; e.g., Trachyceras), with tubercles or even long spines (Fig. 18.1).

Main shell sculpture morphogroups in planispirals, from smooth to fine-ribbed ‘Leiostraca’ with Phylloceras and Lytoceras to strongly ribbed ‘Trachyostraca’ with Olcostephanus and Perisphinctes. Schematic sketches not to scale
Ammonoid researchers assumed that the great majority of ammonoids lived close to the epicontinental (epeiric) sea floor, termed either ‘nektobenthic’ or ‘benthopelagic/demersal’ (Westermann 1990, 1996). Westermann (1996) concluded in his work on ammonoid habitats and life that probably more ammonoids were nektic (= nektonic = active swimming) or were members of planktic communities (= planktonic = passively drifting) rather than being only nektobenthic (= demersal = near-bottom swimming). Such general assumptions are nonetheless highly speculative: they are not based on evidence or data sets, yet may represent good ideas and hypotheses.
Knowledge about life cycles, ecology and ontogeny of fossil cephalopods and especially of ammonoids is still poor and often speculative. While ammonoids are frequently found in Paleozoic and Mesozoic marine sediments worldwide from the tropic-subtropics via Boreal/Austral to Arctic/Antarctic zones, information on their habitat and ecology is scarce and imprecise. Their habitat is suggested to be epipelagic, mesopelagic or epibenthic. Ammonoids probably spawned in benthic, demersal or even midwater habitats (Mapes and Nützel 2009) in the neritic to oceanic zone above the shelf areas and upper slopes (Westermann 1996). Females are thought to have laid 100–500000 eggs on the sea floor (r-strategy) or spawned egg masses in the water column. At the latest after hatching, hatchlings become a part of the plankton (e.g., Landman et al. 1983; Tanabe et al. 1993b; Mapes and Nützel 2009; De Baets et al. 2012; epiplankton after Westermann 1996). This strategy is comparable to most oceanic coleoids, except for the K-strategist nautilids and sepiids. Although it is still under debate, female ammonoids returned to primary habitats (home-grounds), spawned and died afterwards, assuming a semelparous strategy (single reproductive event) at least for some taxa (Callomon 1980, Landman et al. 2003; Stephen et al. 2012), as often observed in neritic, extant coleoids (sepiids or loliginids). Ammon oids perhaps undertook vertical diurnal migrations, as is characteristic for many planktic ocean dwellers such as the deep-water squid Spirula (Clarke 1969; Lukeneder et al. 2008, 2010; Doghuzhaeva et al. 2010). Note, however, that a contrasting model arguing for a permanent demersal habitat in adult stages of late Cretaceous ammonoids from Japan has been presented by Moriya et al. (2003) and Moriya (2015).
Since the last detailed reviews on ammonoid habitats and life histories published by Kennedy and Cobban (1976) and Westermann (1996), extensive work has been done on ammonoid autecology as well as on the synecology of ammonoids and Recent relatives. Ammonoids probably started developing from a planktic hatchling stage (Kennedy and Cobban 1976; Shigeta 1993; Mapes and Nützel 2009), subsequently followed by a differentiation in the mode of life as continued planktic or nektonic or nektobenthic (demersal). The earliest juvenile stages were interpreted by Kennedy and Cobban (1976) as mostly being benthic due to their notable substrate affinities while Mapes and Nützel (2009) suggested a planktic life for some Paleozoic forms. Sexually dimorphic pairs (i.e., macroconch females, microconch males) might have been separated into different habitats (e.g., Acanthodiscus: Reboulet 1996; Olcostephanus: Lukeneder and Harzhauser 2003), as is the case in maturity differentiation (i.e., juvenile to adults; Scaphites: Kennedy and Cobban 1976).
While the functional morphology of ammonoids has been discussed starting more than half a century ago (Trueman 1941; Arkell 1957), advanced analyses on buoyancy and shell orientation, for example, started in the 1980s (Saunders and Shapiro 1986; Hewitt and Westermann 1987; Hewitt and Westermann 1988; Jacobs 1992a). Functional morphological research on ammonoids has focused on three major paleobiological questions: 1) buoyancy, 2) mobility, and 3) habitat depth. Although some ambiguities remain, most previous studies (excluding Ebel 1983, 1992) estimated that living ammonoids had a density almost equivalent to that of seawater, making them neutrally buoyant (Hoffmann et al. 2015; Tajika et al. 2015). The neutral buoyancy hypothesis was supported by theoretical morphological considerations on the mode of coiling and life orientation of some Cretaceous heteromorph ammonites (Okamoto 1988a, b, c). Recently, empirical models corroborated Okamoto’s findings (Hoffmann et al. 2015; Tajika et al. 2015; see Westermann 1993a).
A second aspect is swimming and mobility in coiled and chambered ammonoids. It has been shown that shell stability (determined by the distance between the centers of gravity and buoyancy) is influenced by pressure and hence an expression of the water depth in which the animal lived. Shell size and drag coefficient are key factors in estimating swimming ability (Chamberlain 1976; Jacobs 1992a; Jacobs and Chamberlain 1996; Naglik et al. 2015). One of the outcomes of these studies is that a thinner shell shape is more advantageous in a flow at higher Reynolds numbers, while a thicker shape is more advantageous at low Reynolds numbers (Jacobs 1992a). Another aspect of mobility is (diurnal) vertical movement. Living chambered cephalopods (e.g., Nautilus, Sepia, Spirula) have long been considered to move vertically by controlling the amount of cameral liquid using osmotic pressure of the blood vessels (Denton and Gilpin-Brown 1973). The water depth in which efficient pumping using a simple osmotic mechanism can be accomplished is shallower than 240 m (Greenwald et al. 1980). Accordingly, ammonoids from a deep-water habitat might have often ascended to shallow waters to pump out the cameral liquid (Westermann 1989). Direct observations of vertical movements of Nautilus with remote telemetry (Carlson et al. 1984; Ward et al. 1984) support this idea. Greenwald et al. (1982), however, studied the Nautilus siphuncle at the ultrastructural level and documented the presence of structures associated with a hyperosmotic pump. This would enable the animals to discharge cameral liquid in deep water. Consequently, the argument for ammonoid vertical movement by means of osmotic pressure lost some of its power. Emptying and refilling the chambers to control buoyancy has been considered unlikely in Nautilus for some time (Ward 1986a). It has also been considered unlikely for ammonoids, although that is being debated (e.g., Mutvei and Dunca 2007).
Thirdly, the depths that ammonoids might have inhabited were also estimated based on mechanical properties of the shell against ambient hydrostatic pressure. Based on the wide variation in the mechanical strength of shell materials among various Mesozoic ammonoid morphotypes, Westermann (1996) suggested that Mesozoic ammonoids had various habitat depths. Nonetheless, the implosion depth of a Nautilus shell does not represent an actual depth limit of living animals. Direct observations using a remote camera and capture records using baited traps have demonstrated that the optimal habitat depth of Nautilus ranges from 150 to 300 m in Palau and from 300 to 500 m in Fiji (Saunders 1984; Hayasaka et al. 1987), whereas the shell implosion depth of Nautilus is about 800 m (Kanie et al. 1980). Although these analyses in functional morphology provided valuable suggestions and limitations for considering ammonoid ecology, reasonable and unanswered questions remain for future discussion.
Functional morphology describes the relationship of a single individual, group of individuals, or distinct ammonoid species with its environment (e.g., all parameters of seawater). Accordingly, shell features such as composition, thickness, siphuncle strength and shape are extremely important for understanding the ammonoid/habitat relationship. Additional investigations on the life and habitats of Recent genera such as Nautilus, Spirula and Sepia shed light on the paleoecology and lifestyle of fossil cephalopods. These approaches conclude that most morphogroups were poor swimmers. New findings of soft parts (e.g., stomach content, buccal masses) have enhanced the picture of ammonoids, their hydrodynamics and diets (Kruta et al. 2011).
Ammonoids are marine animals; their lives are conditioned by abiotic parameters such as salinity, with most groups inhabiting euryhaline conditions. A few probably lived in superhaline waters, while others could tolerate subhaline or even brackish waters (Zaborski 1982; Kennedy et al. 1998). Temperature is also a crucial parameter (calculated by δ18O values) together with oxygenation (oxygenated to dysoxic). As noted by Westermann (1996), oxygenation and salinity covary in the water column. Water layers (in epeiric seaways) above an oxygen-minimum zone may have had reduced salinity, enabling only euryhaline organisms (certain fishes) to survive and preventing abundant ammonoid populations. Hydrostatic pressure at corresponding water depths are reflected by adapted conchs, septa and siphuncles of ectocochleates, helping to estimate rupture (Chamberlain and Moore 1982) or implosion depths. Biotic changes in trophic conditions (predator-prey relationships) , food supply (diet differs in different water layers), and natural enemies (cephalopods, fishes, marine reptiles) triggered changes in populations and morphotypes. The sum of the abiotic parameters describes and defines the habitat or environment in which ammonoids lived. Biotic factors relate to the environments, also termed as ecological parameters influencing the mode of life.
Numerous papers of the last decades (Scott 1940; Ziegler 1967; Donovan 1985; Batt 1987, 1989; Westermann 1990, 1996 and references therein) have related shell morphotypes and structural features to depth as determined by sedimentary facies and/or submarine topography. The detection of changes in water depth is important to understand habitat changes in ammonoids.
Did ammonoids actively change their depth-dependent habitats during ontogeny, or did similar ammonoid groups (i.e., morphogroups) migrate into different habitats during long-term evolution? Are ammonoids able to adapt their habits to long-term environmental changes or do certain conditions or events result in a dead end in the evolution of certain lineages?
Geological analyses from a sedimentological and lithological view also brought clues to understand ammonoid ecology. Westermann’s (1990, 1996) investigations on ammonoid autecology emphasized the interrelations between sediment and biofacies. Facies changes were triggered by sea-level fluctuations caused by eustasy, tectonics or subsidence. Kennedy and Cobban (1976) assumed that there is no consistent pattern between ammonoid occurrence and facies. Nonetheless, there are morphotypes or taxa that exhibit significant correlations to sedimentary facies, excluding transport by postmortem drift or redeposition (e.g., Scott 1940; Ziegler 1967; Bayer and McGhee 1984; Donovan 1985; Batt 1987, 1989). Maeda and Seilacher (1996) and Westermann (1996) assumed that postmortem sinking occurred mostly in specimens living in greater depths, especially in mesopelagic habitats , as well as in juveniles and small species. Postmortem ascent, followed by surface drift, was thought to occur mostly in shallow habitats and in adults of larger ammonoids, as is known from extant Nautilus, Spirula, and some sepiids. This hypothesis implies that if ammonoids inhabited various depths within the water column (including shallower water), carcasses would have been widely dispersed by postmortem drift. This would be reflected by each morphology showing no coherency to sedimentary facies . Contrary to this expectation, many previous studies on faunal distributions of ammonoid assemblages have shown that spectra of faunal compositions are closely related to lithofacies, as mentioned above. These facies-dependences suggest that ammonoid carcasses were not transported over long distances by postmortem drift, assuming that ammonoids did not inhabit shallower parts of the water column or that they were at greater depths when they died.
Although many studies have assessed ammonoid ecology, the main problem in understanding ammonoid life and habitat is that they are extinct. Investigations on Recent relatives comprising the ectocochleate Nautilus as well as the endocochleate Spirula and Sepia shed some light on the paleoecology and lifestyle of fossil cephalopods. Nonetheless, the systematic distance between extant and extinct cephalopods prevents a direct comparison in anatomical and ethological aspects. Ammonoid soft parts (e.g., arms, eyes) are in most cases unknown except stomach contents and buccal masses. Taxonomic concepts in paleontology can be based only on morphology and not on genetic principles; this hampers determining exact ammonoid lineages and separating points for species or morphotypes.
Besides these morphological and sedimentological considerations, stable isotope data (δ13C , δ18O, 87Sr/86Sr; clumped isotopes) extracted from original shell material (i.e., aragonite) of ammonoids could yield new insights in ammonoid lifestyle, ontogeny and habitats (Jordan and Stahl 1970; Brand 1986; Anderson et al. 1994; Fatherree et al. 1998; Tsujita and Westermann 1998; Cochran et al. 2003, 2010a, b; Moriya et al. 2003; Lukeneder et al. 2010; Dennis et al. 2013; Stevens et al. 2015). If we can neglect vital effects in calcification of ammonoid (Urey et al. 1951; Moriya et al. 2003) and Recent Nautilus (Landman et al. 1994) shells, then the physicochemical properties, especially δ18O , would provide independent and solid evidence for calcification temperature (and salinity). When the ammonoid calcification temperatures are compared with the thermal structure of the water column, we can identify the habitat depth of ammonoids (Anderson et al. 1994; Moriya et al. 2003). Furthermore, a combination of δ13C and δ18O profiles provides information on the life history of ammonoids, such as sexual maturity (Zakharov et al. 2005, 2006, 2011; Lécuyer and Bucher 2006; Lukeneder et al. 2010). The observed seasonal variations in oxygen isotope data of ammonoid shells potentially enables an age estimation for ammonoid maturity. What has been difficult to measure and evaluate in stable isotope data from ammonoid shells is the possible annual migration over hundreds or thousands of kilometers, a phenomenon observed in numerous Recent cephalopods. It is even more complicated when cephalopods die after spawning in habitats that are quite different and distant from those in which they spent most of their lives (Westermann 1996).
Consequently, a key question is how can we compare the circumstances for fossil, extinct ammonoids and Recent, extant cephalopods when ammonoids in most cases show no resemblance to Recent cephalopods. Also, the phylogenetic relationship is likely closer to coleoids than to Nautilus (Jacobs and Landman 1993; Hewitt and Westermann 2003; Warnke et al. 2003; Warnke and Keupp 2005). This fact is a powerful argument for investigating ammonoid paleoecology through the record itself, because modern analogues are not the best. Catastrophic events led to extinctions at the Cretaceous/Paleogene boundary about 66 million years ago, but orders such as Nautilida , Spirulida or members of the Coleoida survived this big mass extinction. Factors such as the differentiation of ecological niches and different spawning strategies may have triggered different histories of extinct and extant cephalopod groups. A difference can be determined between quantity (r) with small but numerous offspring in ‘opportunistic habitats’ (ammonoids) versus quality (K) with bigger but fewer egg capsules in more stable ‘consistent habitats’ (nautilids).
The state of the art is reviewed and summarized, new methods and insights are given resulting in perspectives within this promising field of cephalopod paleontology. The current compilation is based on the inspiring work of Westermann (1996), a pioneer in the field of habitat and life of ammonoids.
2 Morphology, Architecture, Mobility and Stability of Ammonoid Conchs
The most important terms for shell morphology and ontogenetic stages are summarized below. These are needed to understand interpretations on habitat and lifestyle given in the literature and herein. Morphological terms follow Batt (1989, 1991), Westermann (1996), Wright et al. (1996), and Klug et al. (2015a). Westermann (1996) identified two major morphogroups in ammonoids, the planispirals (‘normal coiled’) and the heteromorphs (varying in coiling-axis).
Planispirals include 12 morphogroups (Westermann 1996, Fig. 18.2) with spherocone (planktic), discocone (planktic-nektic), cadicone (planktic), oxycone (nektic), platycone (demersal), planorbicone (planktic-demersal), serpenticone (planktic), ‘Leiostraca’, ‘Trachyostraca’, brevidome, mesodome, and longidome.
Heteromorphs include 7 morphogroups with orthocone (planktic-demersal), cyrtocone (quasiplanktic), gyrocone (quasiplanktic), torticone (‘trochospiral’, planktic), ancylocone (planktic vertical migrants), hamiticone (planktic vertical migrants), and vermicone (planktic drifters; Fig. 18.3).

Basic planispiral ammonoid shell shapes and their principal assumed habitats, adapted from the original figure in Westermann (1996, Fig. 1, p. 611) and modified after Ritterbush et al. (2014, Fig. 3). Distinct morphogroups appear in the hypothetical diagram with overlapping areas (dashed lines). Grey-shaded region was interpreted by Westermann (1996) as being characteristic for a demersal (nektobenthic) life mode

Basic heteromorph ammonoid shell shapes and their principal assumed habitats, adapted from the original figure in Westermann (1996, Fig. 2, p. 612). Distinct morphogroups appear in the hypothetical diagram with overlapping areas (dashed lines)
The basic planispiral and heteromorph shell shapes were shown in ternary diagrams by Westermann (1996). The majority of heteromorphs was suggested to be planktic, mostly vertical migrants (Westermann 1996). Small gyrocone forms were interpreted as pseudoplanktic in floating algal mats, but detailed evidence is still missing. The main morphotypes and hypothesized modes of life were measured and computed on ternary diagrams to estimate the ammonoid ecospace in the water column (Westermann Morphospace Method; Ritterbush and Bottjer 2012; Ritterbush et al. 2014; Fig. 18.2, 18.3).
Klinger (1980) speculated that torticones and ancylocones were demersal or planktic floaters. Additionally, orthocones were suggested to have buoyancy strategies more similar to Sepia or Spirula rather than to Nautilus (Klinger 1980). Numerous authors (e.g., Batt 1989, 1991, 1993; Westermann 1996; Ritterbush and Bottjer 2012) argued that each shell shape or at least morphogroup had its own special habitats (e.g., demersal, planktonic) and motility (e.g., swimming, passive drifting). Hence, shell morphologies have been interpreted as being directly dependent on habitat. Furthermore, habitat and water depth was argued as being reflected in the lithology, geochemistry, and biofacies (assemblages). Such assumptions, however, are often speculative and based on spatial distribution (e.g., Cretaceous Western Interior Seaway; Batt 1989, 1991, 1993). Many workers calculated the syn vivo density of ammonoids based on shell thickness, the volume ratio between phragmocone and body chamber (Fig. 18.4), and the density of supposed soft parts, which commonly is assumed to be equal to that of Nautilus (i.e., neutrally buoyant), for understanding the presumed habitat.

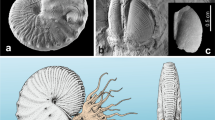
Explanation of dimensions and conch parameters given in the text. a lateral (left) and apertural (right) views of an ammonoid (Kasimlarceltites krystyni, NHMW 2012z0133/0262; adapted from Lukeneder and Lukeneder 2014). b schematic ammonoid-sketch with indicated conch parameters. ah aperture height, bc body chamber, D diameter, d diameter exclusive of last whorl, H whorl height, ph phragmocone, U umbilical width, W whorl breadth. Scale bar: 1 cm
Besides density and buoyancy, the hydrodynamics of ammonoid shells were also examined by many authors. For example, Chamberlain (1980) suggested that a nektobenthic mode of life was predominant among ammonoids. Jacobs et al. (1994) suggested that compressed and depressed morphotypes in Cretaceous scaphitids from the USA had advantages for swimming at higher and lower velocity, respectively. These results are consistent with the fact that compressed morphotypes are found in shallower (more energetic) and depressed morphotypes in deeper (less energetic) sedimentary facies. For more compressed morphotypes, a higher swimming velocity was assumed to be essential for life in shallow waters, whereas the depressed morphotypes are thought to inhabit deeper (offshore) areas (Jacobs et al. 1994).
Landman et al. (2012) examined septal and siphuncle strength (Hewitt 1993; 1996; Tanabe 1979) combined with stable isotope data (Chap. 18.5; Cochran et al. 2010a, b) of Hoploscaphites from the Cenomanian–Maastrichtian of the Western Interior Seaway (USA). They concluded that habitat depth was < 100 m and near the sea floor. The apertural angle excludes a nektobenthic mode of life for these heteromorph morphotypes, at least in adult specimens (Landman et al. 2012, Chap. 18.11). Stable isotope data for the relative habitat depths of Hoploscaphites and Baculites strengthened the supposed shallower environment for Scaphites (Cochran et al. 2010a, b; Landman and Klofak 2012; Henderson and Price 2012). Touch marks on the sea floor attributed to these scaphitids were formed by postmortem bouncing or rolling shells; hence, they do not provide evidence for a demersal lifestyle (Seilacher 1963; Landman and Cobban 2007). Yamada and Wani (2013) measured the thickness ratios of shells (whorl width/diameter) of Scaphites from the lower middle Turonian of Hokkaido (Japan) in order to determine the migration mode: the ratios of Scaphites planus differed significantly between distinct localities. They argued that different populations did not frequently migrate between such areas. Based on hatchling sizes, they concluded that thickness ratios became manifested after hatching due to limited migration within a nektobenthic habitat. That study suggests that scaphitid ammonoids became nektobenthic with limited migration at an earlier stage than previously thought (i.e., not during the transition from normal to heteromorph coiling; Yamada and Wani 2013).
Oxycone ammonoids such as Sphenodiscus from the Western Interior Seaway were used to define migration paths during the Maastrichtian (Ifrim and Stinnesbeck 2010). Sphenodiscids were reported to be characteristic morphotypes for shallow-water environments near the wave base (Batt 1989; Jacobs 1992a; Jacobs and Chamberlain 1996; Ifrim and Stinnesbeck 2010). Ifrim and Stinnesbeck (2010) assumed a connection between near-shore environments and the occurrence of Sphenodiscus, at least for North America.
Many hypotheses about the mode of life of distinct morphogroups are largely speculative. A demersal mode of life (nektobenthic) was proposed for several heteromorph groups by Wiedmann (1973), Vašíček and Wiedmann (1994), and Klinger (1980). The same mode of life was postulated for small heteromorph forms (Karsteniceras) by Lukeneder (2003, 2005) during dysoxic environmental conditions in the Early Cretaceous from the Northern Calcareous Alps (Austria). Ebel (1985, 1990, 1992) suggested a benthic, gastropod-like (see rebuttal comments in Kröger 2001) lifestyle for heteromorphs. Westermann (1990, 1996) summarized the imaginable life habits from active swimming (with varying steerage) to a passively drifting planktic mode, and from horizontal to vertical migratory types. Westermann (1996) separated different strategies within the heteromorphs, a distinct demersal mode of life for cyrtocone forms (e.g., Protancyloceras), a pseudoplanktic one for gyrocone forms (e.g., Crioceratites), and a planktic mode for vermicone ones (e.g., Nipponites; see also Klinger 1980). According to Westermann (1990, 1996), various life modes existed in photic and aphotic zones in the ocean. Stable isotope data confirm this scenario (Lukeneder et al. 2010). Hence, the δ18O values of the forms with strong ribbing such as Hypacanthoplites, Nowakites and Perisphinctes were confirmed to have been inhabitants of the epipelagic zone in the neritic province. Hypacanthoplites (discocone deshayesitid with moderately coarse ribbing) possibly inhabited the photic zone of the uppermost 50–100 m with a planktic to nektobenthic, mobile lifestyle. Nowakites (platycone-discocone pachydiscid with strong ribs) appears to have preferred the transition from the epipelagic to the mesopelagic zone, with a mobile to sluggish mode of life and vertical migrations. Heteromorphs like Baculites potentially preferred a similar water depth (Henderson and Price 2012). In contrast, the much more positive δ18O values of the sub-sphaerical and depressed Cadoceras indicate a habitat in the cooler and deeper mesopelagic zone. Based on this low number of species, the general rule of thumb that strongly ribbed forms dominated above depths of 100 m in the neritic epipelagic zone, whereas smooth forms dominated in the deeper oceanic mesopelagic zone appears to be valid.
A major difference in the modes of life of many ammonoids compared to the modern cephalopods Nautilus, Sepia and Spirula is evident in their adult stage. All extant examples tend to retreat into the deepest environments as adult animals (e.g., Spirula, Sepia) or at least to remain there throughout their post-juvenile phase (e.g., Nautilus). In contrast, all measured ammonoids, except for strongly ornamented Perisphinctes, display a clear tendency to migrate into shallower environments in their latest ontogenetic stage (Chap. 18.5).
Constraints from phylogenetic analyses are needed in the case of arm number, which is mostly given as ten (Mehl, 1984; Jacobs and Landman, 1993; Westermann 1996; Lukeneder 2012; Klug and Lehmann 2015). Moreover, the extent to which the shell was covered by the mantle in different morphogoups remains unclear (Doguzhaeva and Mutvei 1989, 1991, 1992, 1993). Those authors suggested that the shells of the Cretaceous genera Sinzovia, Gaudryceras, and Ptychoceras were semi-internal or even fully covered by mantle. Full coverage in Ptychoceras was rejected by Westermann (1996) due to buoyancy and soft tissue estimations. Shell-wall duplication was interpreted by Birkelund (1981) in Gaudryceras as indication that its juvenile shell was exceptionally covered by the mantle. Additionally, in situ fossil buccal masses are commonly located near the centre of the body chamber, supporting a jaw-function rather than an operculum-function (Schindewolf 1958; Lehmann 1976, 1980, 1988; Kennedy and Cobban 1976; Tanabe 1983; Morton and Nixon 1987; Tanabe and Fukuda 1987; Mapes 1987; Bandel 1988; Nixon 1988, 1996; Seilacher 1993; Westermann 1996; Kennedy et al. 2002; Wippich and Lehmann 2004; Doguzhaeva et al. 2007; Keupp 2007; Landman et al. 2007b, 2010, 2012). As shown by Wani (2007), in situ preservation of buccal masses in the body chamber of ammonoids coincides with the in situ deposition of the ammonoid shell itself (Chamberlain et al. 1981; Wani et al. 2005).
In planispiral ammonoid shells, the static orientation of the aperture (apertural angle) is interrelated with hydrodynamic stability (distance between the centers of buoyancy and mass; Westermann 1996). These centers depend mostly on body chamber length (Fig. 18.4, 18.5), causing differences in soft body volume (i.e., soft body mass). The consistency and density of ammonoid soft bodies are imprecisely known (Hoffmann et al. 2015; Klug and Lehmann 2015). This hampers estimations (Saunders and Swan 1984; Saunders and Shapiro 1986: Shapiro and Saunders 1987; Swan and Saunders 1987) of buoyancy, bathymetry (depth limits), swimming style etc. Assuming neutral buoyancy, Saunders and Swan (1984), Saunders and Shapiro (1986), Shapiro and Saunders (1987), as well as Swan and Saunders (1987) calculated the orientation and stability of Carboniferous goniatitids and prolecanitids. Three body chamber groupings were established in conjunction with their supposed swimming-potential and maneuverability in ammonoids. The criteria were shell shape, aperture orientation, and stability (Westermann 1996; adapted after Saunders and Shapiro 1986), and the groupings were brevidome (< 220° body chamber length), mesodome (220–320°), and longidome (< 320°; Fig. 18.5). Subsequently, the relation between shell thickness ratio (whorl width/diameter) and body chamber length is combined with the apertural orientation. Brevidome, mesodome and longidome ammonoid shell hydrodynamic stability and steerage mechanisms (Fig. 18.5) were discussed by several authors (Davis et al. 1969; Davis 1972; Hengsbach 1978; Chamberlain 1980, 1991; Ward 1981, 1986b; Bayer 1982; Saunders and Swan 1984; Callomon 1985; Saunders and Shapiro 1986; Westermann and Callomon 1988; Checa and Westermann 1989; O’Dor and Wells 1990; Westermann 1990, 1996; Doguzhaeva and Mutvei 1992; Jacobs 1992a; Jacobs and Landman 1993; Dagys and Weitschat 1993; Elmi 1993; Jacobs and Landman 1993, 1996; Jacobs et al. 1994). Heteromorph shell types and their swimming performance were also discussed in numerous papers (Mapes 1979; Ward 1976a; Ward and Westermann 1977; Westermann 1977; Klinger 1980; Okamoto 1984, 1988a, b, c; Batt 1993; Fig. 18.3). A pelagic, quasi-planktic life mode was suggested for Cenomanian openly coiled Allocrioceras by Wippich and Lehmann (2004) conbased on in situ aptychi and stomach contents.

Main classes of body chamber length (angles) in planispiral ammonoids, measured from the aperture. a Brevidome (160–180°), b mesodome (260–300°), and c longidome. (350–400°; adapted from Westermann 1996)
3 Conch Parameters as Proxies for Ecology
Numerous studies drew attention to the relation between the ecology of individual ammonoid taxa and morphological shell parameters (Fig. 18.4) such as rib numbers, diameter (D), siphuncle diameter (sd), siphuncle wall thickness (swt) whorl width-diameter ratios (W/D) or whorl expansion rates (WER; e.g., Kant 1975; Westermann 1996, Nikolaeva 1999; Sarti 1999; Klug 2002, Korn and Klug 2001, 2002, 2012; Korn 2010; Korn et al. 2004; Matsuoka et al. 2010; Ritterbush and Bottjer 2012; Yahama and Wani 2013). Kant (1975) found allometric growth changes in Carboniferous ammonoids. Septal complexity was also investigated biometrically (Kahn and Kant 1975) to gain ideas on ammonoid ontogeny. The same authors showed that measurements on ammonoid sutures can potentially help to estimate growth rate and mode. Ballentine (2007) mathematically analyzed the index of sutural complexity (ISC) used to classify ammonoid shells. He concluded that this index has the same value for suture patterns of very different shapes. Accordingly, it should be combined with other shell characteristics (siphuncle strength). Fernandez-López et al. (1999) demonstrated a relation between ornamentation or conch shape (coiling) and habitat for Tmetoceras (Middle Jurassic, Spain). Evolute morphotypes inhabited basinal areas, whereas involute poplulations were dominant in epicontinental, relatively shallow platforms.
Korn and Klug (2001, 2002, 2012) and Klug (2001, 2002) published conch parameters of Devonian ammonoids (Agoniatitina and Anarcestina) from Morocco, which yielded results on ontogeny, taxonomy, covariation of D, WER, and relations to sea-level fluctuations. Analyses of lithology, grain size and microfacies of the host sediment allowed a correlation with global sea-level curves, although on a weak statistical basis (Klug 2002). Klug (2002) found smaller conchs with lower WER (< 2.0; Anarcestina) in deeper areas and/or trangressive phases and larger morphotypes (Agoniatina; WER > 2.2) with increasing W/D ratios in more shallow areas and/or regressive phases. W/D ratios were also important in detecting sexual dimorphism (Sarti 1999) . Klug (2002) suggested that well-preserved assemblages comprising all growth stages of ammonoids indicate reproduction within the region. Biometric estimations (morphospace) by Neige et al. (1997) on shell parameters including whorl expansion rate, diameter, whorl height and whorl width in Callovian ammonoids from France were used to relate morphology to sea-level changes. A clear trend was detected from subserpenticone (i.e., evolute, depressed, nektopelagic; Fig. 18.2) to suboxycone (i.e., involute, compressed, nektobenthic) morphotypes during a transgression. This indicates a potential usefulness of ammonoids as paleodepth indicators because morphology might change with depth (Navarro et al. 2005). Those authors performed multivariate analyses on Middle Jurassic Cardioceratidae, which made morphospace calculations less intuitive, increasing the reliability of their results and conclusions drawn based on ammonoid habitats. Morphological diversity during immigration phases increased because the animals inhabited distinct niches, a phenomenon probably valid for all ammonoid taxa (see Westermann 1993b).
Biometric analyses are useful to detect differences in shell parameters and their ratios. Celtitids from the Carnian of Turkey have quite similar shell morphologies. Taxonomic separation is mainly based on sculptural differences. Several ontogenetic stages could be distinguished by biometric analyses (WER, W/D) in Kasimlarceltites (Lukeneder and Lukeneder 2014). Embryonic to early juvenile stages start with high WER values (2.16–2.17, spherical), which drastically decreases in older juveniles (1.75, strongly depressed early whorls), then increases markedly in mid-age, followed by unstable mid-aged phases of (WER 1.89–1.79), and finally peaks in high values of the preadult to adult stages (2.05–2.17, compressed). This morphological change mirrors a trend from from planktic to nektic or active swimming from the second life-phase on Investigations on the ammonoid shell shape in morphospace of the serpenticone K. krystini pointed to a hypothetical planktic lifestyle of ceratitid ammonoids (calculated by K. Ritterbush 2013, Fig. 18.2 18.6). A mainly planktic lifestyle is assumed based on the calculations for the small ceratitid ammonoid (max. size 33 mm; Lukeneder and Lukeneder (2014).
Despite of the huge amount of data and articles on ammonoid biometry, the question remains whether there is a relation between conch shape and the physical and chemical parameters of the environment (depth, pressure, salinity, temperature, light etc.). Most studies seem to support such a relationship .
4 Ontogenetic Stages in Ammonoids
The terminology of growth stages used herein is based on ontogenetic stages defined by numerous authors (House 1985; Kant 1975; Kullmann and Scheuch 1970, 1972, Westermann 1954, 1958; Landman 1987; Kullmann 1981; Hewitt 1988; Kant and Kullmann 1988; Landman 1988; Bucher et al. 1996; Klug 2001; Korn and Klug 2002, 2007, 2012; Etches et al. 2009; Lukeneder et al. 2010; De Baets et al. 2012, 2013, 2015; Laptikhovsky et al. 2013; Lukeneder and Lukeneder 2014) . The exact definition of the ontogenetic stages is crucial for reconstructing life cycles with coeval changes of habitats.
The ammonitella (= embryonic stage; embryo in Lukeneder et al. 2010) consists of the subelliptical protoconch or initial chamber and three-fourths to one-and-a-quarter whorls with body chamber phragmocone chambers. The ammonitella is usually delimited by the primary (nepionic) constriction (Birkelund 1981; Landman et al. 1996b, Klofak et al. 1999, 2007; Etches et al. 2009). After that stage, a change in ornamentation and shape occured (Currie 1942, 1944; Burnaby 1966, Lehmann 1966; Kulicki 1974, 1979, 1996; Bandel et al. 1982; Ward and Bandel 1987; Landman 1987, 1988, Landman et al. 1996b, 2007a; Maeda 1993; Kulicki and Doguzhaeva 1994; Bucher et al. 1996; Doguzhaeva 2002; Kulicki et al. 2002; Sprey 2002; Korn and Klug 2007; Tanabe et al. 2010b). The corresponding term in nautiloids is nauta (Laptikhovsky et al. 2013). According to Laptikhovsky et al. (2013), the average ammonitella diameter was 0.54–2.6 mm, decreasing in mean size from the Devonian (up to > 5 mm; De Baets et al. 2015) via the Carboniferous with 0.6–1.4 mm to the latest Cretaceous with 0.7–1.0 mm. Laptikhovsky et al. (2013) concluded that seawater temperatures were the key factor provoking historical changes in ammonoid and nautiloid evolution. The authors argued that eggs were larger in species from temperate than from equatorial areas and also during global warming. The negative relationship between egg size and environmental temperatures is a also known as Thorson-Rass rule (Laptikhovsky et al. 2013).
The neanoconch (= neanic stage of Hyatt 1894, hatchling to early juvenile; juvenile in Lukeneder et al. 2010) comprises 2.5–3.3 additional whorls. It grew mainly in height, is often planorbiconic and weakly sculpted with 3–5 mm diameter (Westermann 1958) .
The juvenile phase (= juvenile stage; juvenile to mid-age in Lukeneder et al. 2010) comprises the late juvenile plus adolescent/immature or preadult phase with several additional whorls. These differ from the preceding ones in various growth parameters, e.g., abrupt growth of width and sculpture, roughly to half adult size.
The adult phase (= adult stage or adulthood in Bucher et al. 1996; adult in Lukeneder et al. 2010; maturity) is fully grown and thus mature. The post-juvenile shell has 3 or more whorls, comprising the final body chamber with different coiling, cross section, sculpture, and/or peristome (Davis et al. 1996; Klug et al. 2015b).
As noted by several authors, Recent cephalopods exhibit two principal spawning strategies (Nesis 1987; Boletzky 1987; Hewitt 1988; Westermann 1996; De Baets et al. 2012, 2015). A benthic K-type strategy with few big eggs and a planktic r-type strategy with numerous small eggs. Most ammonoid eggs (Etches et al. 2009; Tajika and Wani 2011; Stephen et al. 2012; Laptikhovsky et al. 2013; Yamada and Wani 2013; De Baets et al. 2012, 2015), as inferred from the ammonitella, resembled those of the small to smallest eggs of present-day cephalopods. Environmental influences such as warm water and good oxygenation enhanced egg growth (Westermann 1996; Laptikhovsky et al. 2013). Accordingly, aerobic conditions were necessary for full development. Most taxa possibly encased their eggs in a light gel, singly or in masses (Westermann 1996; Mapes and Nützel 2009). Alternatively, the female may have carried the eggs, as in Recent pelagic octopods (e.g., Argonauta). Recent Nautilus produces less than 10 eggs of a diameter of c. 20 mm (Tanabe et al. 1993b; Kröger 2005), laying the egg capsule directly on the sea floor (Saunders and Landman 1987, 2010) .
Differing reproductive strategies in nautiloids and ammonoids in early ontogenies are reported by Stephen and Stanton (2002), Kröger (2005) and Klug (2007). Fossil nautiloids produced fewer, larger eggs than ammonoids (Landman et al. 1996b; Chirat 2001; Klug 2001, 2007). A typical ammonoid egg measured 1–2 mm (c. 100 to 500 000 per mother animal; Klug 2001, 2007; Korn and Klug 2007, 2012; De Baets et al. 2012, 2015; Laptikhovsky et al. 2013). In contrast, the hatchling size for Cretaceous nautiloids varies from 9 to 35 mm (Wani et al. 2011).
It may be speculated that the often small ammonoid hatchling size (compared to nautiloids) enabled them to produce more ammonoid offspring (r-strategists, type III survivorship curve). They drifted as plankton in the water column (Klug 2001, 2007; Landman 1985; Landman et al. 1996b) and were part of planktic food webs (Laptikhovsky et al. 2013). Further information is obscured by the lack of knowledge about where ammonoids laid their eggs (Westermann 1996). As noted by Klug (2001) as well as Korn and Klug (2007), floating ammonoid egg masses in the water column (see also Tanabe et al. 1993b, Mapes and Nützel 2009) would change our understanding of ammonoid ecology and habitat preference .
The ecology of hatchlings and early juveniles (neanic stage) of living cephalo- pods is well known (e.g., Boletzky 1974, 1992; Nesis 1987; Westermann 1996; Stephen and Stanton 2002; Korn and Klug 2007; Laptikhovsky et al. 2013). The hatchling stage in ammonoids began at 3–5 mm diameter (Landman 1987; Landman and Waage 1993; Okamoto 1988a, b; Shigeta 1993; Tanabe and Ohtsuka 1985; Tanabe et al. 1981, 1995, 2003; Bucher et al. 1996; De Baets et al. 2015), thus categorizing the hatchlings and neanic stages as microplankton. The early post-hatching interval in at least Cretaceous ammonoids was assumed to be planktic, changing into nektobenthic or nektoplanktic after reaching a diameter of 2–2.5 mm (Shigeta 1993). Westermann (1996) dismissed this based on difficulties in measuring ammonitella volumes. The most common habitat of the neanic stage was probably the deep midwater of hypoxic, epeiric basins, perhaps somewhat above or below the dysaerobic/aerobic interface (e.g., Schindewolf 1959; Landman 1982; Tanabe et al. 1993b, 1995; Westermann 1996) .
Juvenile specimens are usually rare in ammonoid assemblages of the aerobic facies (Westermann 1954, 1996; Kennedy and Cobban 1976; Callomon 1985; Landman 1987). They appear more frequent in anaerobic and dysaerobic black shales (Westermann 1996), being either pelagic or pseudoplanktic or, as postulated by Westermann (1996), developing from planktic, vertical migrants into demersal swimmers.
In adult stages (mature, fully grown), ammonoids were most probably generalists populating almost every single niche in the marine environment environment (Fig. 18.7),, with the ability to migrate between and within several depth zones. Adult stages are mainly detected based on apertural collars (macroconchs, females) or lappets (microconchs, males) reflecting sexual dimorphism, and based on changes in ribbing or on final septal crowding (Callomon 1980; Davis 1972; Elmi 1993; Davis et al. 1996; Lukeneder 2004; Klug et al. 2015b). Ontogenetically controlled migrations mirror long-term habits, whereas foraging and preying would reflect daily behaviors in ammonoids. According to Becker (1993) and Westermann (1996), even brackish waters could be inhabited by at least some ammonoid groups (e.g., some goniatids, Placenticeras, perhaps Dactylioceras). Westermann (1996) considered the lifestyle flexibility of ammonoids versus extant cephalopods as follows: “except for their reduced depth, speed, and, hence, range of migration, ammonoids presumably had similar ecologic diversity” .
Adult modifications such as coarsening sculpture, shell thickening or uncoiling were discussed by numerous authors (Tanabe and Shigeta 1987; Jacobs and Landman, 1993; Jacobs et al. 1994; Jacobs and Chamberlain 1996; Westermann 1996; Davis et al. 1996; Klug et al. 2015b). Based on estimated shape and hydrodynamic properties, a mainly nektobenthic life was suggested for adult ammonoids by Chamberlain (1980). As summarized by Westermann (1996), the main late ontogenetic trends in ammonoid life were (a) from planktic (or demersal vertical migrant) to (b) sluggish nektic or (c) demersal, passive drifters or active swimmers. Late ontogeny was perhaps accompanied by an increased diurnal vertical migration of planktic forms (see Doghuzhaeva et al. 2010), increased speed and acceleration of nektic or demersal forms, from sluggish to mobile, as in many of the smaller microconchs from demersal to nektic .
Post-neanic ammonoids resemble Nautilus in the roughly linear growth rate (Saunders 1984; Hewitt et al. 1994; Bucher et al. 1996). As noted by Westermann (1971, 1977, 1996), maximum growth rates of ammonoid shells probably depended more on chamber growth than on apertural growth. The neanic growth rate may have differed greatly from that of the juvenile, especially if there was a major habitat change, e.g., from planktic to demersal (Westermann 1996). Growth rate might have decreased with increasing habitat depth (e.g., increasing pressure reduced the osmotic emptying rate, decreasing temperature and food supply, thus slowing the metabolism). As speculated by Westermann (1996), the lifespan may have varied from 1 to 2 years for small shallow-water ammonoids (< 50 mm diameter) to 5 to 10 years for most epeiric and inner-neritic ammonoids (see Bucher et al. 1996), to as much as 50 or 100 years (overestimate?) in mesopelagic giants such as large Lytoceras (Stevens 1988). In general, growth rates were higher and size larger in shallow platform dwellers than in basin dwellers (Elmi and Benshili 1987). Ontogenetic stable isotope analyses on baculitid shells from the Campanian from South Dakota (USA) were conducted to assess δ18O data concerning marine paleotemperature. Our understanding of ammonoid growth rate will be enhanced in the future by the analysis of isotope composition, which changes during migration and habitat change. No exact estimates (c. 33 cm/a) were presented by Fatherree et al. (1998) for Baculites, suggesting a quite rapid growth. More detailed ontogenetic analyses of stable isotopes could probably help to solve that problem. Detailed stable isotope studies with the goal of a better reconstruction of seasonal variations will help to estimate at least the duration from embryonic to full growth (Chap. 18.5) .
No evidence has been presented for the exact habitat where ammonoids laid their eggs and where spawning took place. Stable isotope data (Fig. 18.8, 18.9, 18.10, 18.11) are still too imprecise to reconstruct these aspects using protoconchs and ammontellae shell. Specifically, the volumes required for carbonate analyses (e.g., Keil Carbonate Device) are large (3050 µg) and drill bits (30–50 µm) are thicker than ammonitella shell walls (10–20 µm; Kulicki and Doguzhaeva 1994). Large differences in embryonic stages are visible only when comparing different ammonoid groups. This provides no information on where the eggs were stored (i.e., water column or benthic). All ammonoid hatchlings are suggested to have started passively drifting as plankton (Kennedy and Cobban 1976; Tajika and Wani 2011), the duration of that phase being unknown. The mechanisms of dispersal of the hatchlings and neanic stages were likely entirely passive and depended on the presence and strength of marine currents (Westermann 1996). Westermann (1990, 1996) concluded that dispersal distance ranged from tens of kilometers (enclosed epeiric seas) over a few hundred kilometers (shelf seas) up to 1000–2000 km in the open ocean. These early ontogenetic stages likely formed an important constituent of the Mesozoic plankton in shelf seas and oceans (Fig. 18.7, Table. 18.1) .

The serpenticone Upper Triassic (Carnian) ammonoid shell shape and the principal assumed habitats in Kasimlarceltites krystini (calculated by K. Ritterbush 2013). Distinct morphogroups appear in the hypothetical diagram with overlapping areas. For details see Fig. 18.2 herein. A mainly planktic life-style is assumed from the calculations for the small ceratitid ammonoid (max. diameter 33 mm; Lukeneder and Lukeneder 2014). Reconstruction of K. krystini (left, by 7reasons Media Company) and the holotype of K. krystini (right, NHMW 2012z0133/0014; Lukeneder and Lukeneder 2014). Scale bar: 1 cm

Schematic diagram showing the pelagic zone and its horizontal layers, marine environments inhabited by ammonoids. For details and explanations see the text. Ammonoid size and paleogeographic distances not to scale

Lateral view with numbered samples in ontogenetic direction; growth direction with indicated SEM images of aragonitic ultrastructure, × 500 and × 3700. a Jurassic (Callovian) Cadoceras, b Cretaceous (Aptian) Hypacanthoplites and c Cretaceous (Santonian) Nowakites shells. Arrows mark the position of last suture, the beginning of the body chamber. Stable isotope curves (δ18O in black and δ13C in grey) of the corresponding ammonoids Cadoceras, Hypacanthoplites, and Nowakites (adapted from Lukeneder et al. 2010)
Ontogeny in Recent Nautilus, Sepia, Spirula, and Cenozoic Aturia (above). Ontogenetic migrations in the ammonoids Baculites, Cadoceras, Hypacanthoplites, Nowakites, and Perisphinctes (below). Calculated water temperature curves in growth direction, based on δ18O and depth distribution of the cephalopods investigated compared with additional Recent Sepia, Spirula, Nautilus, Cenozoic Aturia, and Mesozoic Perisphinctes and Baculites (all literature data; see text for explanation). Adapted and extended after Lukeneder et al. (2010). Maximal depth range estimates based on siphuncle strength index (SiSI) from Hewitt (1996, black stars) and Westermann (1996, white stars) in corresponding colours to temperature curves

Paleotemperatures estimated based on stable isotope data (δ18O) from the Lower Cretaceous (Albian) of Madagascar. Ammonoid (Eotetragonites, Cleoniceras, Desmoceras, Douvilleiceras), nautiloid (Cymatoceras) and belemnoid data based on data given by Zakharov et al. (2011). Maximal depth range estimations based on Siphuncle Strength Index from Hewitt (1996, black stars) and Westermann (1996, white stars) in corresponding colours to temperature curves (see text for explanation)

Paleotemperature data estimated using stable isotope data (δ18O) from Late Cretaceous (Campanian) ammonoids from Hokkaido, Japan (Moriya et al. 2003). Light blue shading represents the seasonal range of bottom water temperature estimated from oxygen isotope thermometry of co-occurring benthic organisms. Red shading represents annual mean sea surface water temperature estimated from oxygen isotope thermometry of surface-dwelling planktic foraminifera. Maximal depth range estimations based on the Siphuncle Strength Index (SiSI) from Hewitt (1996, black stars) and Westermann (1996, white stars) in corresponding colours to temperature curves (see text for explanation). Black and white lines for distinct specimens. White bars indicate the depth/paletemperature range of distinct genera given in Moriya et al. (2003). Grey bar indicates a Gaudryceras measured by Zakharov et al. (2005) for comparison
Recently the probable ecology of eggs was discussed by Etches et al. (2009), Tajika and Wani (2011), Stephen et al. (2012), and Laptikhovsky et al. (2013). Etches et al. (2009) reported eggs enclosed in egg sacks (see also Lehmann 1966, 1976; Müller 1969) from the Upper Jurassic (Kimmeridgian) of the UK. Some were still attached to ammonoids (possible parentage), e.g., Aulacostephanus and Pectinatites. Etches et al. (2009) suggested a firm substrate where eggs were anchored below the storm wave base. It is controversial whether such egg sacks still contained eggs in the fossil record, and thus whether they were deposited in situ (Lehman 1976). As speculated by Callomon (1980) and Etches et al. (2009), some groups were gregarious, migrating over long distances and depths. The latter interpretation finds some support in the isotope analyses of Lukeneder et al. (2010, Fig. 18.9). In some Mesozic ammonoids, the sexes were perhaps spatially separated as preadults (Davis et al. 1996; Lukeneder and Harzhauser 2003; Lukeneder 2004), met for mating and separated during spawning. As suggested by Callomon (1980), female ammonoids might have returned to their primary habitats (“home-grounds” in Callomon 1980) for oviposition (Lukeneder et al. 2011), spawned and died afterwards (i.e., semelparous strategy; see also Stephen et al. 2012). The overall strategy might have resembled that of Recent neritic coleoids such as sepiids or loliginids (2009) .
As argued by Tajika and Wani (2011), who studied the Cretaceous ammonoids of Japan, egg masses or ammonitellae (with given protoconch and ammonitellae size ranges) were drifted over long distances at speeds of approx. 110 km within a minimum of 5 days, from spawning places by ocean currents. This is similar to extant coleoids (2009). Tajika and Wani (2011) based their calculations on modern oceanogeography, under the assumption that paleocurrents were similar to modern ocean currents around Hokkaido. Eggs and hatchlings of extant coleoids (e.g., Illex) can be drifted by water currents for 2–3 months, during which they still show the same size range (Haimovici et al. 1998; Tajika and Wani 2011). Laptikhovsky et al. (2013) studied reproduction strategies in the Cretaceous. Based on egg size measurements, they concluded that the evolution of smaller eggs in ammonoids permitted them to occupy new habitats, whereas nautiloids survived despite their inability to migrate and settle new ecological niches. These arguments may also be valid for the ammonoids from the latest Cretaceous (latest Maastrichtian to K/P boundary), which were oceanic to suboceanic (distal neritic), deep epipelagic to mesopelagic nekton and plankton (Ward 1987, 1990a, b; Wiedmann 1988b; Westermann 1996; Fig. 18.6, Table. 18.1). The deep-water character of these last faunas shows low evolutionary rates and a high cosmopolitanism, both characteristic of oceanic taxa (Ward and Signor 1983). The fact that the nautilids survived this event indicates that the main difference might lie in the habitat differences (Westermann 1996) from mostly pelagic Maastrichtian ammonoids to demersal nautilids like Recent Nautilus . An additional cause for the survival of nautiloids was noted by Wani et al (2011): the much larger hatchling size of Cretaceous nautiloids contrasts with those in ammonoids (Ritterbush et al. 2014) .
As noted by Klug et al. (2010), ammonoids presumably began to occupy the water column during the Devonian nekton revolution as nektic organisms, already exhibiting at this early phase various life strategies such as demersal, planktic and nektic (Klug 2001).
Based on Carboniferous anoxic–dysoxic occurrences, Boston and Mapes (1991) speculated that the very young ammonoids might have been benthic (sessile or vagile) and were unable to escape periodic benthic anoxia. In the past decades, a sessile mode of life was rejected for all ontogenetic stages in ammonoids by most ammonoid specialists. Ammonoids of larger growth stages in higher, photic zone depths (Boston and Mapes 1991) were less vulnerable to predators than the very small juveniles or were able to survive brief hypoxic events like living Nautilus (Wells et al. 1992). As noted by Stephens et al. (2012), mass mortalities of ammonoids could occur if they were semelparous (i.e, reproduction mass mortality), or mass mortalities of their ammonitellae or juveniles were mostly driven by environmental catastrophes (e.g., anoxic or toxic events) or reflect taphonomic processes such as stratigraphic condensation or hydrodynamic trapping. Two models for the formation of ammonoid egg masses and the corresponding habitats are possible: egg masses floating in the water column or benthic egg masses. Potential hydrodynamic processes (e.g., accumulation by ocean currents) were not considered by the latter authors .
Abundant embryonic ammonoid assemblages were described from deposits from the USA and Canada (Kansas; Mapes et al. 1992; Tanabe et al. 1993b, 1994a, b; Alberta; Pamenter 1956; Schindewolf 1959; Becker 1993). Complete, and incomplete ammonitellae, rare neanoconchs and small juvenile goniatites (Aristoceras, Vidrioceras) occur in thick layers. Mapes et al. (1992) concluded that egg masses floated above the bottom and were not planktic, or that they were slightly transported from nearby spawning sites. Westermann (1996) suggested a second hypothesis for such egg masses: neanoconchs were floating planktic-quasiplanktic in moderately deep basins, killed from time to time by storms mixing anoxic waters into those areas of the sea; larger juveniles and adults lived higher up in the water column and were unaffected (Westermann 1996). Mapes and Nützel (2009) revised these opinions and suggested floating egg masses and planktic juveniles .
5 Stable Isotopes (δ13C, δ18O)-Implications for Ammonoid Ontogeny and Habitat
Urey (1947), Urey et al. (1951) and Epstein et al. (1953) discovered that oxygen stable isotope ratios of calcium carbonates correlate with the temperature of the water from which they were precipitated and developed a method to determine paleotemperatures (Rexfort and Mutterlose 2006). Since then, many authors have attempted to assess the ancient or present-day seawater temperatures from biologically precipitated calcium carbonates (e.g. Bandel and Hoefs 1975; Wefer 1985; Wefer and Berger 1991). The aragonitic composition of the external shells and septa in ammonoids makes them suitable for isotopic measurements (Jordan and Stahl 1970; Brand 1986; Anderson et al.1994; Moriya et al. 2003; Zakharov et al. 2005, 2006; Lécuyer and Bucher 2006; Lukeneder et al. 2010; Henderson and Price 2012; Stevens et al. 2015), when the aragonite is pristine. Purity of shell material should be tested using modern scanning electron microscopy (SEM), computed tomography (CT) and cathodoluminescence (CL; Lukeneder et al. 2010). The fractionation and isotopic composition in oxygen and carbon isotopes (16O/18O, 12C/13C) refers to a change in stable isotope ratios, reflecting chemical and physical processes (Hoefs 2004) in a cephalopod’s life.
Oxygen isotope composition (δ18O) in carbonate minerals precipitated in water is a function of δ18O of water and precipitated temperature. δ18O of the carbonate reflects the δ18O of the water precipitated under isotopic equilibrium. Therefore, if the δ18O of water can be estimated, the temperature during precipitation can be calculated from δ18O of the carbonate according to the formula
(Grossman and Ku 1986; Goodwin et al. 2003; Kobashi et al. 2003; Lécuyer et al. 2004; Kim et al. 2007, 2010), where δ18Oaragonite is δ18O of carbonate analyzed in VPDB scale, δ18Owater is δ18O of ambient water in VSMOW scale. A shift of one per mill in the oxygen isotope ratio corresponds to a temperature change of approximately 4.3 °C. Lécuyer et al. (2004) assumed that the isotope ratio is related to
(GEOSECS Executive Committee 1987; see also Railsbeck et al. 1989; Geary et al. 1992) where S is the salinity of about 35‰ at a depth of around 300–500 m according to data by Auclair et al. (2004) and Watanabe et al. (2003). Considering that the analytical error in isotope measurements is usually < ± 0.1‰ (practically, error would be slightly > ± 0.1‰), at least 1°C changes in calcification temperature should be detectable and discussable. Biogenic carbonates, however, are sometimes secreted under disequilibrium, which is termed vital effect (Urey et al. 1951). The vital effect is the most confusing bias in the stable isotope records of biologically precipitated carbonates. This effect may be greater in C isotopes than in O isotopes (Wefer and Berger 1991). Only minor or no vital effects are usually observed in the O isotopic compositions of molluscs (Wefer and Berger 1991). In contrast, it is practically impossible to assess the degree of vital effect in ammonoids. Cephalopods (Nautilus, Sepia) precipitate their shells in an O isotopic equilibrium (Landman et al. 1994; Bettencourt and Guerra 1999). One possibility to resolve such problems would be to compare the stable isotope and clumped isotope data to other organisms in the same assemblage.
Carbon isotope composition (δ13C) of synthetic carbonate is predominantly a function of δ13C of dissolved inorganic C and carbonate ion concentration within the solution. However, δ13C in biological carbonate may show a greater vital effect than δ18O , as mentioned above, and it is generally more difficult to assess the specific controls (McConnaughey and Gillikin 2008). Among a variety of biological activities, changes in metabolic rate and incorporation of carbon from food would be major sources of vital effects in molluscs, except for photosymbiotic species (e.g., Jones et al. 1986; Tanaka et al. 1986; Romanek et al. 1987; Henderson and Price 2012). In some cases, abrupt changes of δ13C profiles may be used to identify changes in metabolic rate, such as sexual maturity . As noted by Henderson and Price (2012), studies of Nautilus (Auclair et al. 2004) and the deep-water squid Spirula spirula (Lukeneder et al. 2008; Price et al. 2009) show increasingly depleted carbon values in shell carbonate due to metabolic effects related to changes in rates of growth or food sources.
5.1 Recent Cephalopods-Learning for the Past
δ18O is a proxy for water temperature in aragonitic and calcitic shells such as those of extant (Nautilus, Spirula, Sepia) and ancient cephalopods (ammonoids, nautiloids, belemnoids, teuthoids; Tarutani et al. 1969; Grossmann and Ku 1986; Lukeneder et al. 2008, 2010; Zakharov et al. 2006; Stevens et al. 2015; Fig. 18.8, 18.9, 18.10, 18.11). Food webs and food selection can be examined based on carbon (δ13C) and nitrogen (δ15N ) isotopes (Minagawa and Wada 1984; Cherel and Hobson 2005; Hobson and Cherel 2006). In addition to these techniques, nitrogen isotopes in amino acids are a powerful tool for analysing trophic level in modern species (Chikaraishi et al. 2009; Kashiyama et al. 2010). Consensus exists amongst scientists working with fossil and extant cephalopods on the interpretation of δ18O data. By contrast, δ13C trends are more complicated to interpret in fossil cephalopods and molluscs in general.
Sclerochronologic (i.e., hard-part measurements in chronological order) δ13C and δ18O records from numerous Nautilus species were provided by several authors (Eichler and Ristedt 1966a, b; Cochran et al. 1981; Taylor and Ward 1983; Wefer 1985; Landman et al. 1983, 1994; Auclair et al. 2004; Zakharov et al. 2006). δ13C and δ18O in Sepia cuttlebones were also measured (Longinelli 1966; Longinelli and Nuti 1973; Hewitt and Stait 1988; Bettencourt and Guerra 1999; Hobson and Cherel 2006; Rexfort and Mutterlose 2006; Cherel et al. 2009). Stable isotope composition of Spirula was presented by Lukeneder et al. (2008), Price et al. (2009) and Warnke et al. (2010). Chitinous Octopus beaks were analysed by Cherel and Hobson (2005) for their δ13C and δ15N composition.
Ontogeny-related stable isotope patterns in the shells of Recent cephalopods reveal two main lifecycle types (Fig. 18.9). One is represented by Nautilus and Sepia, which start in warm shallower waters as juveniles and migrate into cooler and deeper waters later in ontogeny. This strategy is contrasted with Spirula, which starts in cold deep waters, subsequently migrates into warmer habitats, and finally inhabits deeper waters in adult stages (Lukeneder et al. 2008). Time averaging of short-term (diurnal) migration does not significantly influence stable isotope trends within the shell. In all taxa, a roughly parallel trend of C isotopes suggests a concomitant change in diet and/or water chemistry due to habitat change .
The comparison of Recent Nautilus, Sepia and Spirula allows quite different modes of life to be deciphered based on stable isotope signatures. Applying these methods to Mesozoic ammonoids sheds light on strategies and environmental requirements of fossil cephalopods. Due to its unusual morphology, Spirula is used as a key genus in paleontological papers that attempt to interpret the mode of life of Mesozoic ammonoids (Lukeneder et al. 2010).
5.2 Spirula
The δ13C , δ18O data reported by Lukeneder et al. (2008) for the deep-water squid Spirula spirula revealed three ontogenetic stages, including a major shift from positive to negative values corresponding to sexual maturation, the initiation of reproduction, and concomitant changes in diet (Fig. 18.9). An embryonic stage with δ18O values at + 3.6 to + 3.1‰ is followed by a steady decrease from + 3.5 to + 3.0‰ to + 1.7 to + 2.0‰ in juvenile stages and an increase to + 2.2 to + 2.6‰ in adults (Lukeneder et al. 2008, see also Price et al. 2009 and Warnke et al. 2010). After hatching at depths > 1000 m (4–6 °C), the squid migrates into shallower, warmer waters at depths of 400–600 m (12–14 °C; Levitus 1994). Subsequently, the animals migrate back into somewhat cooler, deeper habitats at 500–600 m (mean 9.1 °C; Lukeneder et al. 2008), identified for all major oceans. The importance of knowing water temperatures in certain depths is shown by the correlation of known temperatures (Levitus 1994) and stable isotope data of extant molluscs. Little is known about the composition of paleooceanic water stratification (Hay 2008), hence the arrangement of temperature zonation is ambiguous for ancient seas. This makes it much more complicated to obtain reliable conclusions on habitats of fossil organisms than for Recent relatives. As noted by Hay (2008), the circulation of the Cretaceous ocean may have been very different from that of extant conditions, for example by lacking a well-developed pycnocline (e.g., stratification, convection, thermodynamic heat transport) and dominant tropical/subtropical gyres.
5.3 Sepia
Sepia officinalis hatches in warm waters (> 20°C) at depths of < 20 m and descends to greater depths (below thermocline) in accordance with a change in lifestyle from nektobenthic to nektic/nektobenthic (Rexfort and Mutterlose 2006; Lukeneder et al. 2008). Wild-caught specimens displayed δ18O values from − 1.3 to + 3.0‰ during ontogeny, comparable to calcification temperatures of 21 °C in adolescent and 5 °C in adult specimens (i.e., cuttlebones; Fig. 18.9). This ontogenetic trend of migration from warm water into deeper cooler waters was also documented by isotope studies of Longinelli and Nuti (1973) and Bettencourt and Guerra (1999). Compared to Nautilus, sepiids grow much faster, maturing in 1–2 years and dying immediately after reproduction (Packard 1972; Wells 1983; Landman and Cochran 1987, 2010). Contrastingly, Nautilus shows slower growth but lives for several years after maturing (Saunders 1983, 1984). Sepia attains sexual maturity in 200 days (Rexfort and Mutterlose 2006) to 2 years (Bettencourt and Guerra 2001; Challier et al. 2005; Ceriola and Milone 2007), whereas ammonoids might have taken from 5–15 years to mature, as determined by growth line measurements, encrustation patterns and stable isotope data (Kennedy and Cobban 1976; Bucher et al. 1996; Westermann 1996; Fatherree et al. 1998; Lukeneder et al. 2010). Interestingly, sexually dimorphic ammonoids seem to reach maturity at different ages (Kennedy and Cobban 1976), earlier in males and later in females. Estimates were based on whorl number (max. size) with fewer in males and more in females. Stable isotope analyses performed on micro- and macroconchs could help to strengthen or refute these assumptions because δ13C and δ18O data generate details on seasonality and ontogeny .
5.4 Nautilus
δ18O analyses of the shell of various species of Nautilus show more negative values in the embryonic stage and more positive values in the postembryonic stage, indicating migration from warmer shallower into cooler deeper environments during growth (Fig. 18.9; Eichler and Ristedt 1966a, b; Cochran et al. 1981; Taylor and Ward 1983; Landman et al. 1983, 1994; Landman 1988; Oba et al. 1992; Auclair et al. 2004; Zakharov et al. 2006; Kruta et al. 2014). Due to the isochronous timing of the isotope shift from negative to positive values and the decrease in septal spacing and position of the nepionic constriction, this isotope shift is considered to represent hatching of the animal (Cochran et al. 1981; Oba et al. 1992).
Initially, there was debate as to whether the more negative δ18O values in Nautilus’ embryonic stage resulted from the precipitation of embryonic shells in isotopic disequilibrium with the ambient water in eggshells (Taylor and Ward 1983). Crocker et al. (1985) showed that the δ18O of the egg water of two Nautilus species is depleted by approximately 1‰ relative to ambient water. However, the eggs analysed by Crocker et al. (1985) did not contain developing embryos, so the fractionation between the egg water and embryonic shell was not evaluated. Landman et al. (1994) finally succeeded to analyze the δ18O of shells of N. belauensis raised under controlled temperatures in the Waikiki Aquarium from embryonic and early postembryonic stages. The shells of both stages were secreted under isotopic equilibrium with the in situ water.
Based on O isotope data, nautilid embryonic development takes place at 22–30 °C and depths of 100–200 m, depending on the species. As inferred from aquarium observations, hatching occurs after about 1 year of development (Landman et al. 1994; Uchiyama and Tanabe 1999); in nature, juveniles then migrate into cooler deeper waters (150–400 m; 14–16 °C). Consequently, Taylor and Ward (1983) defined two ontogenetic stages via stable isotope data, the embryonic stage and the free-swimming stage. The main δ18O shift corresponds with the formation of the 7th to 8th septum, reflecting hatching after the embryonic stage. The embryonic stage has δ18O values below c. –1.0‰ (> 20 °C), the free-swimming stage values above –1.0‰ (< 20 °C). Variations from a δ13C value of –1.3 to a value of + 1.5‰ are correlated with changes from the embryonic to juvenile-adult stages (Taylor and Ward 1983) and the change in habitat (Fig. 18.8).
Nautilus macromphalus was found to reach sexual maturity in c. 2.5–6 years (Martin et al. 1978; Landman and Cochran 2010; Collins and Ward 2010) and 3 years (Zakharov et al. 2006). For Nautilus belauensis, maturity was attained with 15 years (Saunders 1983) and 10 years (Landman and Cochran 1984, 2010). Typical embryonic δ18O values range around –1.07 to –3.0‰. After c. 269 to 362 days (Uchiyama and Tanabe 1999), hatching takes place and a migration into cooler, deeper waters (150–400 m; 14–16 °C) starts. This is documented by increasing δ18O values of + 0.4 to +1.21‰.
This migrational behaviour is characteristic for Nautilidae since at least the Cretaceous because similar δ18O patterns, from more negative to more positive values after hatching, occurred in fossil Eutrephoceras (Landman et al. 1983; Landman 1988). In Eutrephoceras, δ18O shifts occur between septa 2 and 4, with a magnitude of the shift varying from 1.6‰ to 2.9‰. Calcification temperatures at embryonic and postembryonic stages correspond to 22–23 °C and 14–20 °C, respectively. Thereafter, Eutrephoceras descended into deeper waters with c. 14 °C (Landman et al. 1983).
Isotopic analyses on the nautiloid Aturia from the Cenozoic (Eocene, Miocene) of Slovakia indicate a nektobenthic lifestyle with temperatures of 13–16 °C and 14–18 °C for juvenile and adult stages, respectively (Schlögl et al. 2011; Fig. 18.8). Schlögl et al. (2011) suggested that newly hatched juveniles and adults inhabited the same water at c. 240–330 m. The constant habitat depth through ontogeny in Aturia contrasts to the drastic change in Nautilus (Zakharov et al. 2006).
5.5 δ15N-Amino Acids in Nautilus
The δ15N of amino acids incorporated in shell carbonates of wild Nautilus pompilius specimens captured in the Philippines provide new insight into its trophic level (Kashiyama et al. 2010) . Values in bulk δ15N decrease from embryonic to postembryonic stages. Three trophic stages in ontogeny were detected. Trophic levels calculated from compound-specific amino acid δ15N (Chikaraishi et al. 2009) decrease from 12–14‰ (embryonic) to 9–11‰ (post hatching), and finally to 10–12‰ (juvenile to mature). The higher trophic level values in the embryonic versus postembryonic stage is explained by consumption of egg yolk produced by the adult parent; i.e., the offspring is eating parts of their parents. Kashiyama et al. (2010) also applied this method to the Cretaceous (Albian) nautiloid Cymatoceras from Madagascar. In contrast to the stepwise decreasing δ15N values in the extant Nautilus, δ15N values in the fossil nautiloid material gradually increased in ontogeny from 1.2‰ to 3.8‰, probably reflecting a change in diet. The authors, however, did not trust their results because “a potential pitfall of the current method is that we cannot exclude the possibility that organic matter was added to or formed within the samples during postmortem degradation” (Kashiyama et al. 2010). While the applicability of this method to fossil materials might be limited, future new techniques would help to understand the life history of extinct animals .
5.6 Ammonoid Ontogeny-Implications from Stable Isotopes
In early O isotope paleothermometry work on ammonids, the focus was often on understanding marine paleotemperatures (Triassic and Jurassic ammonoid shells, Kaltenegger 1967; Fabricius et al. 1970; Kaltenegger et al. 1971; Jeletzky and Zapfe 1976). For this purpose, belemnoids were also frequently utilized since Lowenstam and Epstein (1954). This method generated insights into ancient ocean water temperatures at different Mesozoic ages (Kaltenegger 1967; Stahl and Jordan 1969; Tourtelot and Rye 1969; Jordan and Stahl 1970; Fabricius et al. 1970; Kaltenegger et al. 1971; Jeletzky and Zapfe 1976; Forester et. al. 1977; Marshall 1981; Landman et al. 1983; Whittaker et al. 1987; Fatherree et al. 1998; Cochran et al. 2003, 2010a, b; Moriya et al. 2003; He et al. 2005; Ifrim and Stinnesbeck 2010; Henderson and Price 2012; Zakharov et al. 2005, 2006, 2001, 2012). Similar investigations were made on the endocochleate belemnites (Lowenstein and Epstein 1954; Spaeth et al. 1971; Ditchfield 1997; Podlaha et al. 1998; Price et al. 2000; Niebuhr and Jochiamski 2002; McArthur et al. 2004; Price and Mutterlose 2004; Dutton et al. 2007; Zakharov et al. 1999, 2005, 2011; Stevens et al. 2015). Additional analyses were performed on nacreous shells of Carboniferous nautiloids from Oklahoma (Seuss et al. 2012).
Most work on isotope paleothermometry involving ammonoids utilized only the shells (outer shell, septa), with more recent efforts using calcitic aptychi and rhyncholites (Kruta et al. 2014) to discuss paleooceanography or ammonoid ecology. If we assume that ammonoids were mobile organisms living in the water column, one must be cautious when discussing isotope results based on ammonoid shells. When δ18O data of a single ammonoid individual show a sinusoidal ontogenetic pattern, we have two potential explanations for it: (1) the temperature of the inhabited water fluctuated seasonally or (2) the analyzed individual successively migrated into warmer/cooler water masses during ontogeny.
A pioneering work on ammonoid isotope paleothermometry (Stahl and Jordan 1969) used this approach to discuss ammonoid ecology, especially growth rate. Since then, the number of ontogenetic stable isotope analyses on ammonoid shells has increased. Ontogenetic sampling is especially useful for paleotemperature estimations because δ18O data of single cephalopod shells can range over 2‰, spanning a temperature range of almost 8–10 °C (Fig. 18.9, 18.10, 18.11). Single point measurements (Kaltenegger 1967; Fabricius et al. 1970; Kaltenegger et al. 1971; Jeletzky and Zapfe 1976) will only snapshot ocean water temperatures at a specific point of development (Lukeneder et al. 2010).
Life habitat throughout ontogeny was reconstructed for several ammonoids by Lukeneder et al. (2010). Entire ammonoid specimens were measured in the spiral direction (embryonic to adult aperture) to gain information on ecology and habitat preferences. δ18O data from a Jurassic Cadoceras and Cretaceous Hypacanthoplites and Nowakites were chosen due to the primary aragonitic shell preservation (Fig. 18.8), and compared with Recent Nautilus , Spirula, and Sepia as well as Cenozoic Aturia, which possess equivalent or comparable hard parts (Fig. 18.9). The most suitable approach to assess diagenetic processes and the value of shell alteration (in all molluscs) is to check the purity of the shell material (i.e., aragonite; see methods listed above).
The δ13C values revealed three ontogenetic stages in Cadoceras and Haypacanthoplites, including two major shifts from positive to negative and from negative to positive values (Fig. 18.8). These probably correspond to sexual maturation, the start of reproduction, and concomitant changes in diet (Lukeneder et al. 2010). δ13C and δ18O records both suggest separated phases corresponding to ontogenetically controlled vertical migrations within the water column. The values mark three to four phases in ontogeny: embryonic, juvenile, mid-aged (preadult) and adult.
The δ18O values of juvenile Hypacanthoplites shells increase from juvenile –2.29‰ to an adult value of –2.80‰, with a mid-aged minimum of –3.11‰ (Fig. 18.8). This reflects an ontogenetic migration from warm shallow marine environments to even warmer environments (27–28 °C; Fig. 18.9). The δ18O values of juvenile Cadoceras shells decrease from juvenile (–0.93‰) to adult (+ 0.39‰), with a juvenile minimum of –1.35‰ and a mid-aged minimum of + 0.75‰ (Fig. 18.8). This points to a shallow, c. 21 °C warm marine habitat of juvenile Cadoceras. Later, the mid-aged animal preferred slightly cooler and deeper environments (12–16 °C). Finally, the adult Cadoceras migrated back to slightly warmer and shallower environments (c. 17 °C; Fig. 18.9).
The resulting maximum temperature range to which the ammonoids were exposed during ontogeny is about 10 °C in Cadoceras and 6 °C in Hypacanthoplites. Compared to recent seawater temperatures (Levitus 1994; NOAA 2014), this suggests a bathymetric range of 50–700 m in Cadoceras and of only 60 m in Hypacanthoplites. With respect to the probably unstratified ocean system in the Jurassic and Cretaceous (Hay 2008), the observed data might point to even larger ranges.
From the isotope results, ontogenetic strategies for the morphologic groups ‘Leiostraca’ and ‘Trachyostraca’ were postulated by Lukeneder et al. (2010). The wcw-type (warm–cool–warm type) of Cadoceras resembles the patterns in Nautilus and Sepia, starting (a) as early juvenile in shallow waters migrating (b) as juvenile to mid-aged into deeper environments and (c) as adult back to shallow and thus warm waters. The cww-type (cool–warm–warmer type) of Hypacanthoplites, represented in modern cephalopods by Spirula (cwc-type), migrates (a) as a juvenile from deeper environments into (b) shallower ones and then (c) as an adult back into deeper habitats. The latter trend is slightly modified in Hypacanthoplites: it experiences a higher temperature plateau as an adult. During this adult phase from 28 to 29 °C (e.g., late mid-aged to adult), a slightly decreasing temperature trend occurs with age (28.9 °C to aperture with 27.5 °C; Fig. 18.9).
Data on Perisphinctes were published by Lécuyer and Bucher (2006), although they did not cover the earliest ontogenetic stages (Fig. 18.9). A threefold post-embryonic development is signaled by the O isotope values: a shallow and warm late juvenile to mid-aged phase at c. 23–24 °C is contrasted by slightly cooler and deeper juvenile and adult stages at 20–21 °C (Lukeneder et al. 2010). This pattern is similar to that of Spirula, although its deep mesopelagic habitat differs from the epipelagic environment of Perisphinctes. The C pattern of Perisphinctes, however, has little in common with Spirula, but is strongly reminiscent of that of Nautilus: very low embryonic and early juvenile stages and a considerable shift towards positive values thereafter. As this shift is not reflected in the δ18O values, the δ13C shift was probably induced by a major change in diet or the onset of sexual maturity rather than a marked habitat change. The animals probably changed their prey preference after attaining a certain size (Lukeneder et al. 2010). The impulse for the ontogenetic migrations in both groups (i.e., ‘Leiostraca’ and ‘Trachyostraca’) may parallel that in modern cephalopods. Firstly, the change in diets and, secondly, the mating–spawning phase in mid-aged to adult ammonoids.
As summarized by Lukeneder et al. (2010) and reviewed herein, stable isotope data on ammonoids (mostly single point measurements) are available for the Triassic Arcestes, Carnites, Sagenites and Rhacophyllites, the Jurassic Amaltheus, Cadoceras, Leioceras, Parkinsonia, Perisphinctes, Phlyseogrammoceras, Quenstedtoceras and Staufenia and the Cretaceous Acanthoceras, Acanthoscaphites, Anagaudryceras, Baculites, Canadoceras, Chimbuites, Cleoniceras, Demesites, Desmoceras, Didymoceras, Discoscaphites, Douvilleiceras, Eotetragonites, Euomphaloceras, Eupachydiscus, Exiteloceras, Gaudryceras, Hauericeras, Hoploscaphites, Hypacanthoplites, Hypophylloceras, Hypoturrilites, Jeletzkytes, Kossmaticeras, Menuites, Mesopuzosia, Nowakites, Otohoplites, Oxytropidoceras, Pachydiscus, Phyllopachyceras, Polyptychoceras, Scalarites, Scaphites, Sciponoceras, Simbirskites, Sphenodiscus, Tetragonites, Tragodesmoceras, Turrilites and Yokoyamaoceras.
Cochran et al. (2003) measured Maastrichtian cephalopods (e.g., Eutrephoceras, Jeletzkytes) from the Western Interior Sea in North America to define various depositional settings. δ18O values of the fossils show decreases from the marine (ca. 35‰) to brackish (ca. 20‰) biofacies consistent with increasing temperatures (ca. 13 to 23 °C) or, if temperatures were relatively constant, to a decrease in the δ18O of the water in which the shell formed (Cochran et al. 2003). The latter interpretation is consistent with less-than-fully marine salinities in the nearshore biofacies, but changes in both the temperature and the isotopic composition of the water may have occurred in this environment. Stahl and Jordan (1969) and Jordan and Stahl (1970) found a seasonal variation of about 8–9 °C in the Callovian based on Quenstedtoceras and Staufenia. Similarly, Lécuyer and Bucher (2006) analyzed Jurassic Perisphinctes from Madagascar to obtain seasonal surface water temperatures of the southern hemisphere. Zakharov et al. (2005, 2011) suggested seasonal temperature fluctuations of 1.7 °C in Anagaudryceras during the Coniacian from Russia and in Albian ammonoid shells (e.g., Cleoniceras, Douvilleiceras) from Madagascar. Numerous values in Zakharov et al. (2011) may be uncertain because the percent aragonite (e.g., 88%) is lower, presumably due to diagenesis. Stable isotope data from the Albian of Madagascan belemnoids (e.g., Parahibolites, Tetrabelus), ammonoids (e.g., Eotetragonites, Cleoniceras, Desmoceras, Douvilleiceras) and nautiloid shells (e.g., Cymatoceras) given by Zakharov et al. (2011) were interpreted as seasonal signals. Seasonality is probably over-interpreted in that study because the measurements were not performed ontogenetically (insufficient number of samples), only encompassed small parts of the shell (Zakharov et al. 2005) and the spacing between sampling points was probably too wide. These data more likely reflect habitat changes during ontogeny as suggested by Lukeneder et al. (2010), and not seasonal variations as assumed by Zakharov et al. (2011). Nevertheless, the interpretation of paleotemperature of the Albian Madagascan sea (tropical–subtropical zone) appears to be correct, as are habitat inferences from upper mesopelagic to lower epipelagic for the ammonoids (Fig. 18.7, 18.10, Table 18.1). Eotetragonites might have lived within a paleotemperature range of 13.3–16.4 °C, Cleoniceras of 16.4–19.4 °C, Desmoceras of 15.5–21.1 °C, and Douvilleiceras of 20.2–21.6 °C. The only nautilod, Cymatoceras, hints at epi– to mesopelgic water layers of 16.4–17.2 °C (only 2 samples in Zakharov et al. 2011).
Taxa such as Phlyseogrammoceras from the Toarcian of Germany display variations of 3–4 °C (Jordan and Stahl 1970), indicating considerable ontogentic differences in lifestyles. Several ‘ranges’, however, are attributed to variations in calcite-aragonite ratios and thus reflect rather diagenetic effects (e.g., Pliensbachian Amaltheus in Jordan and Stahl 1970). The relatively high content of secondary calcite of up to 15% (mean 5–9%) within the original aragonite of the analyzed Jurassic ammonoids (Jordan and Stahl 1970) shows such secondary alteration and replacement processes. Original, unaltered aragonite material (i.e., 100% pristine primary aragonite) should be used in stable isotope analyses. A case in point are the baculitids, which have repeatedly been used to decipher ontogenetic changes of these Cretaceous heteromorphs (Tourtelot and Rye 1969; Forester et al. 1977; Whittaker et al. 1987; Fatherree et al. 1998; Fig. 18.9).
The Late Cretaceous heteromorph Polyptychoceras pseudogaltinum from Hokkaido was analyzed by Okamoto and Shibata (1997). The authors concluded, based on buoyancy and mode of growth investigations, either a demersal mode of life almost touching the sea bottom or a nekto–planktic mode in the water column. A demersal lifestyle was proposed by Moriya et al. (2003) for several taxa from the Upper Cretaceous of the northwestern Pacific (Fig. 18.11). The relatively short shell-sections utilized for that study, however, did not include a full record of the ontogenetic shifts and potential migrations. It remains to be determined whether data from Moriya et al. (2003) on the wide range of δ18O values in Hypophylloceras (i.e., Hs1 to Hs2) or in Polyptychoceras (i.e., Pp1 to Pp2) reflect slight differences in stratigraphy and variations in lithology (i.e., concretions, Moriya et al. 2003) or instead mark different ontogenetic stages reflecting different habitats. Stable isotope (δ13C, δ18O) ratios show that depth distributions changed within ammonoid ontogeny and the values differ in various morphological groups (e.g. ‘Leiostraca’ vs. ‘Trachyostraca’; Lukeneder et al. 2010; see Stevens et al. 2015).
The long-term (i.e., lifespan) and large shifts in δ18O observed in modern Nautilus and Mesozoic species (Lukeneder et al. 2010) can be explained as major habitat change during ontogeny (see for Cretaceous pelagic Simbirskites Stevens et al. 2015). A crucial feature in O isotope trends is the knowledge about thermal structures with special issues in epeiric seas (e.g., Western Interior Seaway; Wright 1987) of the water column at the time of formation of the ammonoid shells. It is difficult to determine the exact habitat depth of an individual ammonoid based on isotope paleothermometry. For modern Nautilus, habitat depth of embryos and adult individuals have been measured based on δ18O data (Eichler and Ristedt 1966a, b; Cochran et al. 1981; Taylor and Ward 1983; Landman et al. 1983, 1994; Landman 1988; Oba et al. 1992; Moriya et al. 2003; Auclair et al. 2004; Lukeneder et al. 2010). Similarly, if we know the local seasonal variation of ancient sea surface temperatures, which is comparable to the ontogenetic variation of isotope temperature of an analyzed ammonoid individual, we can infer habitat depth (Moriya et al. 2003).
Anderson et al. (1994) presented a substantial data set on the thermal structure of the water column and isotope thermometry on ammonoids to identify the depth habitats of Jurassic Kosmoceras. Unfortunately, instead of sampling through ontogeny, they used shell fragments embedded in mudstone. Accordingly, their results were expressed as the general distribution of calcification temperature of Kosmoceras. Nonetheless, their results clearly indicate that calcification temperatures of Kosmoceras were significantly warmer than those of benthic bivalves, and slightly cooler than surface-dwelling vertebrates, pointing to a near-surface habitat of Kosmoceras.
Moriya et al. (2003; see also Moriya 2015) also determined the thermal structure of the water column and the calculated calcification temperature of numerous Campanian ammonoids from Japan. They used this information to infer the depth habitat of each species in the adult stage (Fig. 18.11). The paleotemperature range and calcification temperature data were from benthic and planktonic foraminifera, bivalves, gastropods and ammonoids. Within the ammonoids, the authors utilized 15 discrete individuals of 9 species and 4 superfamilies, which include discocones, oxycones, planorbicones and hamitocones. However, Moriya et al. (2003) provided no data on embryonic and juvenile stages. Nonetheless, the calcification temperatures of all analyzed adult individuals (adult body chamber parts) are comparable to the bottom water temperatures derived from isotope thermometry of co-occurring benthic foraminifera, bivalves and gastropods. Moriya et al. (2003) concluded that, regardless of morphology, all species were nektobenthic (demersal).
As shown from δ18O values, estimated paleotemperatures (Fig. 18.11) and the subsequently calculated paleodepths of Campanian ammonoids (i.e., all morphogroups and ammonoid families from Japan), Tetragonites, Damesites, Hauericeras and Polyptychoceras inhabited shallower waters, compared to literature data (Moriya et al. 2003). By correlating recently obtained isotope data from Japanese material with literature on ammonoid δ18O data (Moriya et al. 2003; Lukeneder et al. 2010) and with estimated habitat depths (Westermann 1996), I developed a new approach for the reinterpretation and determination of ammonoid/habitat relations. Recent δ18O data from the literature (Moriya et al. 2003; Zakharov et al. 2005; Lukeneder et al. 2010) were compared with other published data mainly based on assumptions concerning morphology, siphuncle strength, shell wall thickness or facies dependency (Hewitt 1996; Westermann 1996). The earlier data were then re-evaluated (Fig. 18.9, 18.10, 18.11). Applying this method to all taxa measured in Moriya et al. (2003) yields a more detailed range of 70–350 m (Fig. 18.11), exclusively from the lower to middle epipelagic, with a clear dominance in the lower epipelagic areas. The exception are the Phylloceratina (Hypophylloceras): they also inhabited the upper mesopelagic, ranging up to the middle epipelagic from 100–300 m. This contrasts with much greater depths given in the literature (down to 600 m; Westermann 1996). Accordingly, the ranges of estimated paleodepths and habitat boundaries of Cretaceous ammonoids from Japan (Moriya et al. 2003) are narrower than assumed based on earlier data (Hewitt 1996; Westermann 1996). Nonetheless, a shallowing gradient is evident from smooth to fine ribbed types (i.e., ‘Leiostraca’) of Phylloceratina (Hypophylloceras, Phyllopachyceras) and Lytoceratina (Tetragonites, Gaudryceras), over stronger ribbed types (i.e., ‘Trachoystraca’) of the Ammonitina (Eupachydiscus, Yokoyamaoceras) to heteromorph Ancyloceratina (Polyptychoceras). At least Polyptychoceras (i.e., Pp1 to Pp2 in Moriya et al. 2003) appears to have migrated in ontogeny from lower epipelagic at 200 m to higher epipelagic zones at 60 m (Fig. 18.11). Japanese Gaudryceras is suggested to have inhabited depths of 80–210 m, similar to estimated 100–180 m for Gaudryceras from eastern Russia given by Zakharov et al. (2005; only last whorl; Fig. 18.11; see also Zakharov et al. 2006). Note that this would follow the hypothesis given in Lukeneder et al. (2010) suggesting that numerous groups migrate within the water column during ontogeny. This is in contrast with Moriya et al. (2003; also pers. comm. K. Moriya 2013), who concluded that their Late Cretaceous ammonoids from Japan did not migrate during ontogeny, rather being nektobenthic (demersal) for their entire lives. One important distinction between the arguments in Moriya et al. (2003) and proposals of Lukeneder et al. (2010) and additional data (herein) is the duration of suggested migrations. Diurnal migrations are not ruled out by Lukeneder et al. (2010) and Moriya et al. (2003). In fact, they are likely. Moriya et al. (2003) argued for a constant long-term mean depth (e.g., average depth over months to years) that does not change in ontogeny. This argument is not necessarily unequivocal because only few individuals have been sampled to date. Including the data from the scheme outlined by Lukeneder et al. (2010) for ammonoid and extant cephalopod habitats, combined with data from Hewitt (1996) and Westermann (1996), indicates that a migration of at least a few ammonoid species from Japan is possible or even likely in the Late Cretaceous sea (Fig. 18.11). New geochemical results indicate that classic assumptions about the habitat depth of each ammonoid morphotype based on functional morphology should be reassessed or partly revised based on physicochemical evidence.
Possible habitat depths of ammonoids, according to literature data and more recent stable oxygen isotope data (δ18O), range from approx. 20–500 m (Fig. 18.9, 18.10, 18.11). Marcinowski and Wiedmann (1988) and Wiedmann (1988a) already stated that bathymetry and climate (i.e., temperature) are the main motors and controlling factors for ammonoid distribution. Depth data (implosion depth) estimated from siphuncle and shell wall strengths (Tanabe 1979; Hewitt 1993, 1996; Westermann 1971, 1975a, b, 1996) are reliable for ammonoid paleobathymetry. Earlier data based on such estimations were demonstrated as being correct or at least valuable in most cases (i.e., by comparing with δ18O-paleotemperature data). Nonetheless, several values given in the literature seem to be imprecise.
Lukeneder et al. (2010) described a bathymetric range of 50–700 m for Cadoceras starting in the lower epipelagic and migrating deeper during ontogeny to the upper mesopelagic realm. With a water-depth range from 10–60 m in Hypacanthoplites, ontogeny starts in deeper epipelgic layers, followed by shallower epipelagic stages. Nowakites is interpreted to have lived from 50–300 m, implying migrations from deeper mesopelagic layers to shallower epipelagic habitats (Fig. 18.9). The maximal depth range given for Baculites (Cochran et al. 2010a, b; Landman and Klofak 2012, Henderson and Price 2012) could be specified to be relatively shallow from 50–100 m in the epipelagic zone. Persiphinctes apparently inhabited almost the same middle epipelagic zone (50–100 m), but migrated to deeper areas as adults (comparable to early juveniles).
Stable isotope data from Albian ammonoids of Madagascar (e.g., Eotetragonites, Cleoniceras, Desmoceras, Douvilleiceras) were given by Zakharov et al. (2011). The ontogenetic rows (as indicated by shell diameters) enable an interpretation on paleotemperature and habitat: tropical-subtropical, upper mesopelagic to lower epipelagic (Fig. 18.7, 18.10) with a depth range from 70–400 m. Zakharov et al. (2006) concluded that belemnites underwent ontogenetic changes, such as in Nautilus, but argued against an ontogenetic migration in ammonoids. Price et al. (2011) described a habitat differentiation within different belemnite groups from the Early Cretaceous based on δ18O data combined with Mg/Ca ratios (Dutton et al. 2007; see Stevens et al. 2015). Alberti et al. (2011) measured 61 belemnite rostra and reconstructed a paleotemperature model based on the δ18O data from the Middle to Upper Jurassic of western India. The authors proposed a migration at least during spawning times from cooler areas into warmer strata. The conclusions drawn by Zakharov et al. (2006) are contrasted by their own figured distribution patterns of Cretaceous ammonoids and by their assumptions that large pachydiscids (Canadoceras, Menuites, Eupachydiscus) were able to migrate to colder waters during ontogeny (shown herein and in Lukeneder et al. 2010). The numerous ammonoid data given by Zakharov et al. (2006) point to a range of at least 7 °C, which, in their opinion, is explainable rather by seasonal fluctuations than ontogenetic trends. That range, however, is too wide for seasonal temperature variations at 100–200 m depth (for temp. see Levitus 94); hence, migration during ontogeny is indicated for most Albian ammonoids. Consideration must also be given to the paleogeographic position of Madagascar during the Albian at middle latitudes of 30–40°S (Stampfli and Borel 2002; Zakharov et al. 2006; Lukeneder et al. 2013).
The Lytoceratina (Eotetragonites) appear to be the only inhabitants of the upper mesopelagic layers (200–400 m; Fig. 18.10). Contrastingly, Westermann (1996) suggested a shallower epipelagic range (100–180 m) based on siphuncle index estimates. Albian members of the Ammonitina lived within the lower epipelagic zone. The finely ribbed forms Cleoniceras and Desmoceras are a case in point: the former ranges from 100–200 m (max. 250 m in Westermann 1996) with an ontogenetic trend from deeper waters, into shallower epipelagic layers and back (cwc–type), the latter from 70–250 m (min. 180 m in Westermann 1996). The ontogenetic trend in Desmoceras shows a major habitat change from higher epipelagic waters into the upper mesopelagic waters (wc–type), i.e. much deeper in adult stages. Strongly ribbed Ancyloceratina like Douvilleiceras are relatively shallow-water inhabitants of the middle epipelagic at depths from 70–90 m, migrating into slightly shallower waters during ontogeny (Fig. 18.10). The Albian nautiloid Camytoceras inhabited depths around 180–200 m (not ontogenetically measured), equivalent to modern Nautilus. Scott (1940) was among the first to present estimations on comparable Albian faunas (e.g., Desmoceras, Douvilleiceras, Hypacanthoplites) from Texas. He suggested that ammonoids dominated in the infraneritic zone (20–100 fathoms in Scott 1940, = 37–183 m). Depth was estimated mostly based on morphological shell features and facies: 36.6–183 m in Hypacanthoplites, 146–183 m in Douvilleiceras, 183–200 m in Desmoceras. Recent estimates from isotope data show much shallower depth values than already given in the historic literature for all members: 10–60 m for Hypacanthoplites, 70–90 m for Douvilleiceras and 70–250 m for Desmoceras (Fig. 18.9, 18.10).
δ18O data are available from Cenomanian ammonoids of Australia (e.g., Euompaloceras, Acanthoceras; Henderson and Price 2012). The reconstructed paleotemperatures ranged up to 34 °C in accordance with the subtropical paleolatitude and the thermal maximum of the northern Australian area during the early Late Cretaceous (Henderson and Price 2012). Similar isotope data from benthic gastropods and bivalves as well as the straight-shelled baculitid Sciponoceras show calculated paleotemperatures of 21 °C, assuming a nektobenthic mode of life. Heteromorph (helically coiled) ammonoids such as Hypoturrilites and Turrilites with δ18O from – 2.9‰ to – 0.8‰ were interpreted to inhabit bottom- and midwater depths, excluding surface waters (Henderson and Price 2012; Batt 1989). Henderson and Price (2012) argued that the relatively positive δ13C values (0.0‰ to 2.8‰) of the Australian helical ammonoids are typical of normal marine waters, but may also point to a sluggish mode of life and slow metabolic rate. More negative δ18O data were derived (Henderson and Price 2012) from the planispiral, strongly ornamented Acanthoceras and Euomphaloceras (– 4.9‰ to – 1.6‰). These shell morphologies suggested pelagic and nektobenthic life habits (Stilwell and Henderson 2002). Henderson and Price (2012) interpreted these ammonoids as growing in near-surface waters based on the most negative δ18O values. This conflicts with the interpretation of a deep marine nektobenthic habit and with data from Campanian planispiral taxa recorded by Moriya et al. (2003), as is also challenged herein (see above discussion of the interpretation of data in Moriya et al. 2003). Based on δ18O paleotemperature records relative to associated benthos, those authors considered a taxonomically diverse assemblage of planispiral forms to be nektobenthic. Note that the Campanian forms in Moriya et al. (2003) differ from those in Henderson and Price (2012) in morphology as they are, except Eupachydiscus, smooth to lightly ornamented and lack ventral tubercules. Based on δ18O values for ammonoids, the general rule of thumb that ‘Trachyostraca’ dominated above 200 m in the neritic epipelagic zone whereas ‘Leiostraca’ dominated in the lowermost epipelgic and deeper mesopelagic zones appears valid.
6 Morphological Assessment of Bathymetry in Ammonoids
Ecological conclusions drawn on habitat, bathymetry and life history of ammonoids are mostly speculative. Hewitt (1993) stated that ammonoids and nautiloids are directly adapted to a maximum water depth, in either pelagic or benthic habitats. A maximal depth of approximately 240 m was given for ammonoids (Denton 1974; Hewitt 1993) based on the maximal depth for cameral fluid osmosis .
Information about the paleobiolgy of ammonoid assemblages can be gained in several ways. The first is a synoptic examination of the literature on ammonoid ecology dealing with morphology–habitat relations (Ziegler 1963, 1967; Kauffmann 1977; Donovan 1985; Batt 1989, 1993; Jacobs et al. 1994; Westermann 1996; Neige et al. 1997; Navarro et al. 2005). A second approach is to analyze the relation between the ammonoid fauna and the embedding facies (Chap. 18.7; literature reviewed by Westermann 1996). An additional possibility is to regard conch parameters (shell morphology, siphuncle strength, etc.) as being directly linked to and hence reflecting primary habitat conditions (depth, temperature etc.) as displayed by morphotypes and hypothesized life modes (Hewitt 1993; Westermann 1996; Klug 2002; Mutvei and Dunca 2007; Ritterbush and Bottjer 2012) .
The main structural elements increasing the strength of an ammonoid shell against hydrostatic pressure and predators are the shell wall and the septa (form and spacing). Their stremgth is thus a main proxy for the mode of life (Westermann 1971; Denton 1974; Ebel 1983; Saunders and Swan 1984; Saunders and Shapiro 1986; Shapiro and Saunders 1987; Swan and Saunders 1987; Hewitt 1993; Shigeta 1993). According to Westermann (1996), decreasing shell thickness correlates with an increase in body chamber length, a change in aperture orientation (more horizontally) and a reduced hydrostatic stability. Nonetheless, Westermann (1996) stated that the siphuncle strength (SiSI) is a more reliable proxy than shell thickness for estimating depth and bathymetry. The septa are truncated by the siphuncle from the first, embryonic chamber to the last septum at the border to the body chamber (Westermann 1958, 1996; Bayer 1977; Jacobs 1992b; Hewitt and Westermann 1987; Korn 1992) .
External shell features such as shell strength (thickness) and whorl cross section are crucial for the hydrostatic stability of ammonoid shells in the water column. The most pressure-resistant shells have circular to semicircular sections, followed by elliptical and ovate ones (Westermann 1958, 1975b, 1996; Ward and Westermann 1985; Jacobs 1992b; Hewitt and Westermann 1987). In contrast, shallow-water cephalopods required thick shells mainly for protection from predators (Westermann 1977; Bond and Saunders 1989). Mathematical and statistical estimations of shell thickness and internal features (see below) that draw conclusions about implosion depth based on the maximal pressure resistance point of an ammonoid shell remain theoretical and provide no direct proof. This calls for additional methods (e.g., stable isotope data, assemblage analyses).
Internal shell features such as septa (thickness, spacing), septal necks (length, diameter, orientation), siphuncles (thickness, diameter) and sutures (complexity) were often used to infer habitat depth (Westermann 1996; Oloriz and Palmqvist 1995; Mutvei and Dunca 2007; Klug and Hoffmann 2015). Strength calculations were based on septal flutes (Hewitt 1996). The septal flute strength index (SFSI, function of maximum septal thickness, minimum whorl radius, and tensile strength; Hewitt and Westermann 1987, 1997; Wang and Westermann 1993; Lewy 2002), combined with statistical methods, help to estimate habitat depth limits (Hewitt and Westermann 1988; 1990a). The relation between septal shape, suture complexity (shape and amplitude; Checa and Garcia–Ruiz 1996), septal spacing and the resulting assumptions about habitat depth (grade of resistance againts seawater pressure) were discussed in several articles (Kahn and Kant 1975; Westermann 1958, 1971, 1975b, 1996; Westermann and Ward 1980; Ward and Westermann 1985; Doghuzaeva 1988; Batt 1989; Hirano et al. 1990; Jacobs 1992b; Boyajian and Lutz 1992; Olóriz and Palmqvist 1995; Saunders 1995; Hewitt and Westermann 1997, 2003; Checa 2003; Hassan et al. 2002; Lewy 2002, 2003) .
Olóriz and Palmqvist (1995) questioned whether sutural complexity is a real indicator for habitat depth or whether it is only an artifact in ammonoid ecology. The underlying question is whether sutures are adaptive or non-adaptive elements. They concluded, based on Jurassic ammonoids, that suture complexity is related more to shell coiling and whorl sections than to bathymetry (Olóriz et al. 1999). Ward and Westermann (1985) favoured siphuncle strength as reliable function for paleodepth estimations.
Concomitant changes in habitat and shell structure reflect life cycles and changes in ecology (Westermann 1996). Changes in siphuncle strength (SiSI) and shell/septa thickness were reported to be reliable proxies for habitat depth estmates (Hewitt and Westermann 1988; Hewitt 1996; Westermann 1996). High pumping rates would have been limited to ammonoids with large siphuncular radii that are found only among shallow-water forms with small SiSI, such as the Upper Cretaceous large ammonoid Placenticeras (Westermann 1996) . Shell growth was accelerated in shallow water, resulting in thinner, more economic, and lighter septa (Westermann 1996). The relative septal thickness, the major variable relating to septal strength (Hewitt and Westermann 1988; Hewitt 1993), was therefore either ecologically controlled or evolved rapidly in the new habitat. Both are good evidence for autochthonous populations. Thus, the exceptionally large difference between the septal and siphuncular strengths is indicative of the rapid habitat change from the usual oceano–mesopelagic to the new epeiro–epipelagic habitat (Westermann 1996; see Fig. 18.7). Hewitt (1993) suggested hatching in epipelagic zones and dispersal in deeper mesopelagic areas after hatching; this was based on septal strength estimation, assuming that the last septum is the weakest part against hydrostatic pressure in ammonoids. Fossil nautiloids were believed to have hatched at shallower benthic sites (Hewitt 1993). The author noted that assumptions on epibenthonic or epipelagic habitats could not be made by the estimated water pressure based on septal strength. Sutural complication has been used in bathymetry, assuming that it improves shell strength against ambient pressure and, hence, varies with habitat depth (Westermann 1996).
Contrastingly, Daniel et al. (1997) demonstrated that there was no positive effect of septal complexity on habitat depth, and even suggested a shallower position for ammonoids with complex sutures. They concluded that selective pressures from predators and buoyancy control were the determining factors: depth did not cause increasing septa complexity. Assumptions based on computed models, simulated and estimated from hypothetical chambers and septal shapes, and values filled in formulae, do not sufficiently mirror the real life of ammonoids and should only be used in combination with other bathymetric indicators (e.g., foraminifera, facies, ammonoid assemblage, stable isotopes ) to avoid overestimations .
Westermann (1996) referred to an additional internal shell component believed to influence the possible maximal habitat depth: the septal neck (Doguzhaeva 1988; Tanabe et al. 1993a; Hewitt 1996; Tanabe and Landman 1996). Estimations on bathymetry were calculated by correlating the length of septal necks and the siphuncular strength index (SiSI; Westermann 1971, 1973, 1975b, 1982; Hewitt and Westermann 1988). The congruent ontogenetic reduction of SiSI and septal flute strength indicates that connecting rings weaken during ammonoid ontogeny (Westermann 1971, 1996). Mutvei and Dunca (2007) rejected the siphuncle strength index as an appropriate tool for paleobathymtric estimations; they noted that the index should only be used as an indicator for relative differences in distinct ammonoids, in combination with connecting ring permeability data.
As noted by Mutvei (1975), Tanabe (1977), Westermann (1971, 1996), Hewitt and Westermann (1988), Geraghty and Westermann (1994), Hewitt (1996) and Ballentine (2007), the siphuncle strength and connecting ring ultrastructure (Mutvei and Dunca 2007) can directly be connected to implosion depth for ammonoids. Doghuzhaeva et al. (2010) noted that conchs with a small siphuncle diameter and long septal necks, as in most phylloceratids and lytoceratids, could resist hydrostatic pressure and thus inhabit deeper waters . The main parameters are the siphuncle diameter (sd) and the siphuncle wall thickness (swt), yielding an estimated siphuncle strength index (SiSI). Westermann (1971) incorporated these parameters in the formula SiSI = h/r × 100 (h siphuncle wall thickness, r siphuncle radius), adapted by Hewitt (1996) to
(d′ wall thickness, r inner radius). An adapted formula,
is applied to Late Triassic ceratitids from Turkey (swt = siphuncle wall thickness; Lukeneder and Lukeneder 2014). The resulting SiSI values for Kasimlarceltites are 10–12.6, which, following Geraghty and Westermann (1994), are equivalent to a maximal depth of 200–252 m. Westermann (1971) also noted that ammonoids with ceratitic or pseudoceratitic sutures like Kasimlarceltites probably inhabited even shallower waters. This would tentatively strengthen the habitat suggestions as epipelagic at shallow, upper mid ramp for Kasimlarceltites as discussed above (Lukeneder et al. 2012; Lukeneder and Lukeneder 2014; Lukeneder and Mayrhofer 2014; see Fig. 18.7) .
7 Allochthonous Versus Autochthonous Ammonoid Assemblages
Based on the considerations in Westermann (1996), this chapter discusses the relation of ammonoid associations and biofacies using Paleozoic to Recent case studies. All scientists working on ammonoid assemblages, regardless of the time slice, have to deal with the same problem: recognizing whether ammonoid shells are autochthonous or allochthonous. Postmortem drift could have been controlled by sea surface currents (wind) or by bottom currents (density differences). Paleobiostratinomy deals with the period from the ammonoid’s death to its final deposition on the sea floor before burial. The main feature causing postmortem drift in cephalopod shells is the presence of a gas-filled phragmocone.
Postmortem drift of empty cephalopod shells can be observed in Recent Nautilus (Kennedy and Cobban 1976; Saunders and Landman 1987; Ward 1987; Reyment 2008). Hamada (1964, 1984) and Toriyama et al. (1964) report similar shell concentrations for modern Nautilus. Despite Reyment’s (1958) doubts, that empty shells of fossil cephalopods floated like those of Spirula (mostly washed on shore) and modern Nautilus, a ‘surface-drift-theory’ for the postmortem drifts of the Cenozoic nautilid Aturia shells is favoured (Lukeneder and Harzhauser 2002). The studies of Hamada (1964, 1984) and Toriyama et al. (1964) on the necroplanktic features of recent Nautilus shells provide evidence for postmortem driftings of 3000 km and floats of more than 1 year, which are reflected in bioerosion and epifaunal settlement. Similarly, Saunders and Spinosa (1979) recorded a postmortem drift of a Nautilus shell over a distance of 1000 km (between Palau and Mindanao) in a maximum of 138 days, an average of approximately 7 km per day. The maximum drifting distance (so far) involved a Nautilus specimen that drifted ashore after 11 years (Ishii 1981). Postmortem drift behaviour and mechanisms for Recent cephalopods such as Nautilus and Spirula were summarized by Reboulet et al. (2005) and Reyment (2008).
Sedimentological and paleoecological studies (Lukeneder and Harzhauser 2002) on a Miocene section from Austria revealed significant shell accumulations of the nautilid Aturia aturi. Each of these allochthonous occurrences containing abundant nautiloid shells within a littoral to shallow sublittoral mollusc fauna reflected current-induced, postmortem drifts of the otherwise rare cephalopod. Taphonomic features (including broken shells with broken phragmocones, broken body chambers and fragmented specimens without body chamber) strengthen the idea of prolonged transport. This is comparable to abundant Nautilus washed ashore after stormy weather (own observations Susanne Mayrhofer, Andaman and Nocobar Islands, Indian Ocean, August 2006). As noted by Hamada (1964), ca. 80% of the Nautilus shells washed ashore had lost their body chamber.
According to Ward (1987), nautiloids are never abundant in Cenozoic strata but are not rare either. However, the Austrian occurrences provide evidence that specimens of Aturia can be abundant in special cases and may even form shell concentrations in environments distinctly different from their habitat. The described shell accumulations are restricted to littoral to shallow sublittoral environments along the coast of the Paratethys. Since the rocky shores and shallow embayments cannot be expected to have been a suitable habitat for a nectonic, deep-water dwelling cephalopod, the shell accumulations are linked to current-induced, postmortem drifts of Aturia shells (Lukeneder and Harzhauser 2002).
Such open seasurface drift-routes are associated with currents, whereas nearshore drifts are mainly induced by wind. Although the actualistic ecological comparison of Nautilus with the fossil Aturia is complicated by differences in shell shape and suture lines , which may reflect adaptation to different habits (see Ward 1980, 1987), the general drift-behaviour is considered to be identical. This hypothesis is also strongly supported by the observations of Kobayashi (1954), who related the various Cenozoic, nearshore occurrences of Aturia in Japan with the proto-Kuroshiwo current. As shown by Kobayashi (1954) by bottle-float experiments, it is not possible to reconstruct the necroplanktic history of shells. During one of his experiments, one bottle arrived after 10 months, whereas a second bottle reached the same locality after 2 months. This difference is explained by the complicated interplay of oceanic currents and countercurrents, tidal currents and winds. Correspondingly, the unknown interplay of these factors, resulting in the formation of shell accumulations, prevent a detailed reconstruction of paleocurrent-induced drift routes.
Similar to the Austrian shell accumulations (Lukeneder and Harzhauser 2002), most of the Japanese findings were classified by Kobayashi (1954) as embayment type occurrences, representing necroplanktic floats. He also stated that the animals did not inhabit these embayments, but does conclude that the natural habitat was probably not very far from these localities. This might also be valid for the Austrian aturiids. The usually excellent preservation of the shell surfaces (as seen on plaster moulds) indicates a short floating period. Epifauna as described by Hamada (1964, 1984) on Nautilus shells and by Seilacher (1960) on ammonoid shells, being characteristic for long floatings, is completely missing on the material from Obermarkersdorf and Unternalb. Similarly, heavy exfoliation or abrasion can be excluded: even delicate growth lines are well preserved. Thus, the shells are interpreted to have been transported over a rather short distance from the adjacent Molasse Basin. According to Teichert and Matsumoto (1987), the endogastric position of the siphuncle in Aturia indicates adaptations to deep rather than shallow-water environments, pointing at the deep Miocene Molasse Basin as its habitat. The shallow embayments along the coast acted as traps for postmortem drifts (Lukeneder and Harzhauser 2002).
The transgression, which affected the coasts in that region during the Miocene (Roetzel et al. 1999) resulted in a rapid sediment covering of the Aturia-bearing strata and probably explains the preservation of the littoral to sublittoral taphocoenoses. Moreover, the slightly heterochronous (i.e., up to 1000 years) deposition of the two shell accumulations seems to be linked to this transgression. Hence, the reported mass-occurrences of Aturia are always bound to littoral environments, whereas the sublittoral fauna of the distal section Unternalb bears only a single specimen. This specimen consists solely of the early parts of the phragmocone and displays some breakage of the shell surface. As demonstrated by Toriyama et al. (1964), Recent Nautilus floats easily if the camerate portion of the shell is in a good state of preservation, even when the body chamber is broken. By contrast, during floating experiments, the Nautilus shells sank rather abruptly if the phragmocone was broken. The single fragmented Aturia from Unternalb therefore had little drifting capacity. This Aturia occurrence coincides with a thin coquina of the archaeogastropod Diloma (Paroxystele) amedei. Kroh and Harzhauser (1999) interpreted this layer to represent an allochthonous occurrence of shells, which were transported into the shallow basin from an adjoining littoral environment during a storm. This nautilid shell was probably damaged during this high-energy event and then transported from the coast to the outer bay. The energetic conditions along the coast of the investigated area of the Bohemian Massif resulted in a fair percentage of breakage of the body chambers, but the good preservation of shell surfaces shows that the fragmentation usually did not affect the phragmocones.
Nautiloid accumulations are well known in the literature, but are interpreted in different ways. Cymatoceras accumulations from Cretaceous deposits of Russia (Shimansky 1975) were related to turbidites. Aturia accumulations from the Miocene of western USA (Moore 1984) and from the Miocene of Slovakia were considered to be deposited autochthonously (Schlögl et al. 2011), Aturia from the Eocene of Antarctica (Zinsmeister 1987) to be deposited in high-energy environments (e.g., shoreline). Hercoglossa from the Paleogene of Argentina are interpreted by Casadío and Concheyro (1992) to be shore-drifted specimens. Grunert et al. (2010) reported an Aturia accumulation from the Miocene as being triggered by several consecutive biostratinomic mechanisms. The authors suggested a deep-water environment as its habitat, followed by transport to the shoreline and redeposition in the dysoxic-anoxic basin by storm events (> 100 m depth). Schlögl et al. (2011) interpreted the accumulation of Aturia shells (juvenile–adult) as being living and hence deposited autochthonously in dysoxic environments in 240–330 m depth (Chap. 18.5). These authors excluded transport and drift based on the presence of related jaws and the shell diameter range (7–38 mm). An exceptional storm accumulation of Early Cretaceous nautiloids was reported by Cichowolski et al. (2011) from Argentina. This occurrence was deposited at deeper environments near mid-ramp, accumulated by storm-induced drifts, trapping on sea floor and probably reflecting gregareous lifestyle. Nonetheless, recent Nautilus can be accumulated in deep water areas, as reported by Roux (1990 in Maeda et al. 2003), Roux et al. (1991) and Mapes et al. (2010a, b).
Drifted nautiloids and ammonoids can change the relative proportions of ammonid assemblages (Stevens 1997). Nonetheless, in practice the effect of postmortem drift of ammonoid shells, becoming nekroplanktic, is thought to be minimal (Chamberlain et al. 1981; Cecca 1992; Reboulet 2001; Westermann 1990, 1996; Chirat 2000). For detailed factors in the distribution of fossil cephalopods and exact floating orientations, see Reyment (1958, 1973, 1980; 2008).
The main and probably the starting point of ammonoid biostratinomy is why, how and when they sank. For details on waterlogging, implosion, buoyancy of empty ammonoid shells, pressure of the cameral gas, surfacing to never surfacing and drifting behaviour of ammonoids, see Maeda and Seilacher (1996) and Wani and Grupta (2015). As noted by Chamberlain et al. (1981) for Nautilus, postmortem drift is enhanced when predators or scavengers remove the soft body. This considerably increases buoyancy and the shell drifts easily. In contrast, when shells become negatively buoyant at certain depths (critical at 50 m in Nautilus), they sink. The critical depth of surfacing and dispersal patterns were discussed by Maeda and Seilacher (1996). The depth limit, at which ammonoid shells float up to the surface, marks a boundary in the water column at a specific hydrostatic pressure. Below, ammonoid shells would not have surfaced, above, they would have drifted.
Sinking history can also be detected by analyzing the encrusting community and the distribution of the latter on the shell (Reyment 1973, 2008; Seilacher 1982). The path of an ammonoid shell after the animal died is controlled by several environmental factors such as depth at death, the initial buoyancy of the empty conch, the morphological and physiological parameters (e.g., morphology, size, weight) of the conch, and the rate of seawater influx into the phragmocone chambers (Kennedy and Cobban 1976; Chamberlain and Weaver 1978; Chamberlain et al. 1981; Olóriz et al. 1996; Chirat 2000; Reboulet et al. 2003, 2005).
The cephalopod/aptychi ratio provides a useful criterion to evaluate the postmortem transport of ammonoids (Olóriz et al. 1996; Lukeneder and Tanabe 2002; Reboulet et al. 2003; Doguzhaeva et al. 2007; Schlögl et al. 2011). Accumulated ammonoid shells can be affected by resedimentation processes generating fragmentation (sensu Fernández–López 1991, 1997; Fernández–López and Meléndez 1994, 1995; 2004; Olóriz et al. 1996; Fernández-López et al. 1999; Landman and Klofak 2012). Resedimentation does not necessarily imply significant lateral transport. The described mechanisms for postmortem drift and the biostratinomic processes in fossil cephalopods show the complexity and the difficulties for a detailed assignment of a distinct ammonoid assemblage to a specific habitat or water depth.
8 Ammonoid Associations and Facies
Ammonoids have been used extensively by cephalopod researchers to correlate shell morphologies with environmental conditions (habitats), and distinct morphological groups were assigned to certain sea level changes, reflected by the lithology and facies (Ziegler 1963, 1967; Tanabe 1979; Enay 1980; Donovan 1985; Marchand and Thierry 1986, Marchand 1992, Hantzpergue 1995, Westermann 1996, Neige et al. 1997; Klug 2002). Proxies for estimations related to shell morphology and the development of morphospace are ornament strength and whorl cross-section (e.g., circular, subtriangular, elliptic; depressed vs compressed etc.).
Abel (1916) was the first to show a correspondence and interaction between the environment and the newly evolving morphologies of cephalopods. That seminal paper on the paleobiology of cephalopods underlined that cephalopod evolution is closely related with changing environmental conditions. Evolution moulds the genetic programming of cephalopods and the potential for adaptation (Jacobs et al. 1994; Young et al. 1998; Yacobucci 1999). Adaptation is one major motor for evolution, a situation recognized by Abel (1916) when he erected the new field paleobiology. Paleobiology shows how important the animal–environment interaction is for promoting evolution. Do ammonoids speciate profusely because internal factors enhance variability and reproductive success? Or do ammonoids respond passively to environmental changes and therefore react after changes of the environment? Problems associated with these major issues in cephalopod research have been highlighted by diverse papers expressing different points of view. The adoption of new habits interacts with long-lasting morphological change and therefore appears as a new evolutionary trend. Evolutionary trends show the main directions and pathways, but are only descriptive mirrors for more important processes that more cephalopod workers should recognize (House and Senior 1981; House 1993b). Spectacular evolutionary radiations mostly took place when the environment changed drastically. New forms evolved due to environmental changes or due to adaptions to the preferred habitat. The adaptive strategy is reflected in the change of morphology in the fossil record and the embedding fauna.
There are still numerous open questions on the ammonoid–facies relation. The confusion in the literature calls for clarifying that ammonoids cannot be dependent on a facies: they can merely show dependency on primary environments. Environments are characterized by several parameters such as depth, light, pressure, oxygen, salinity and others. All these factors together led to the formation of a certain sediment, yielding a special lithology or facies. The appearance–disappearance or presence–absence of ammonoid taxa within different facies was interpreted to be related to specific life habits and habitats (Kennedy and Cobban 1976; Westermann 1996). Kennedy and Cobban (1976) noted the wide range of ammonoid-facies relations from little relation indicating nektic to planktic habits over strong relation reflecting a benthic mode to almost no relation in many taxa. No relation, or independence, is indicated when a certain ammonoid taxon appears in every single facies, contrasted by high relation when a taxon appears in a single facies only. As noted by Kennedy and Cobban (1976), relative abundances can hint at different habitat–ammonoid relations, if postmortem drift can be excluded.
An extensive review of the relation of ammonid assemblages to the corresponding facies was given in Westermann (1996). As stated by Kennedy and Cobban (1976) in their paper on Late Cretaceous ammonoids (N America), no clear patterns of an ammonoid-facies relationship were found (Reyment 1958; Ziegler 1967). Kennedy and Cobban (1976) referred to another problem. It involves the assumption by several authors (e.g., Böse 1928; Sornay 1955; Collignon 1963, 1964) that micromorph ammonoids (tiny or dwarfed forms) are exclusively linked to shaly, offshore deep-water facies in the Cretaceous. As noted by the latter authors and observations below, such pyritized small ammonoids are often the nuclei (inner, juvenile whorls), hence synonymous forms of large specimens from shallower areas. As argued by Keupp (1997), highly morphologically variable species refute the paradigm of Westermann (1996) that explicitly requires a dependency of shell morphology on the mode of life. The Bajocian Sonninia apparently shows 64 different morphologies (from oxycone to serpenticone) in a single layer, all living in the same habitat (Keupp 1997). Keupp (1997) also quoted the apertural injuries in Dactylioceras inflicted by benthic crustaceans on the sea floor (Liassic, Germany). Dactylioceras was assumed by Westermann (1996) to be a planktic form, hence should not have been attacked by benthic organisms. Polymorphism from a single horizon was shown by Wilmsen and Mosavinia (2011) for Schloenbachia from the Cenomanian. The morphotypes were distributed within a gradient from shallow-water areas (proximal) to deeper environments (distal). Strongly sculptured morphotypes dominate the shallow environments; deep-water areas were dominated by compressed, weakly ornamented morphotypes (Wilmsen and Mosavinia 2011) causally induced by water velocity and predation pressure.
Less attention was given in the literature to the ontogenetic variation within one species or even within a single specimen. Such information, however, is crucial for understanding the life and habitat changes in ammonoids. New methods and data on stable isotope analyses (Chap. 18.5.) have shifted our knowledge on paleodepths, habitats, and lifestyles summarized in Westermann (1996).
As evident in the literature (Hewitt and Westermann 1987; Batt 1991) there are also different points of view about the complex relation between suture complexity and habitat and its proxies such as depth (i.e., pressure, water density). Commonly used terms such as “probably planktic”, “presumably pelagic” and “may have lived” show the insecurity in asigning habitats or lifestyles.
The same applies to the connection at various systematic levels to particular facies. Westermann (1996) still separated ammonoid assemblages and biofacies types from the typical environments such as the ocean floor, the ocean margin, the hypoxic cratonic seas, the epeiric basins and the epeiric reef slopes and carbonate platforms. He summarized Paleozoic and Mesozoic fossil assemblages with the intention of showing a relation between ammonoid association and facies (e.g., litho- and biofacies). He was also the first to figure dioramas and possible scenarios of ammonoid habitats, summarized (for adult stages) related to the corresponding facies (tab. 1, p 684–686 in Westermann 1996). However, most of these scenarios are speculative and need testing with geochemical and stable isotope data.
A crucial fact is that ammonoids lived in the water column above the sediment in which they finally became embedded, although postmortem transport is possible. In most cases, it remains unknown, hence highly speculative, in which depth the ammonoid lived. Siphuncle strength index (SiSI) and suture amplitude index (SAI) could be used in combination with stable isotope shell analyses (Fig. 18.8, 18.9, 18.10, 18.11). Nonetheless, Westermann (1996) correctly assumed that ammonoids were probably capable of inhabiting most parts of the sea.
In ocean floor ammonoid assemblages (inclusive aptychi), Westermann (1996) included associations from the Jurassic to Cretaceous of the North American Basin and the Central Pacific (Jansa et al. 1979; Renz 1972, 1973, 1978, 1979). In fact, these occurrences merely show that ammonoids lived in the water column from the bottom to the surface.
8.1 Sculpture and Bathymetry
Morphological parameters (e.g., ornamentation, conch geometry) are meant to reflect the lifestyle of separated morphogroups such as the ‘Leiostraca’ (i.e., smooth shells in deeper water; Fig. 18.1, Table 18.1) and the ‘Trachyostraca’ (i.e., strong ribbing in shallow water; Westermann 1996; Lukeneder et al. 2010). Characteristic ‘Leiostracans’ are Lytoceras, Phylloceras, Desmoceras or Haploceras, comparable to recent Nautilus. Typical ‘Trachyostracans’ are Trachyceras, Stephanoceras or Deshayesites. The ratio between the ‘Leiostraca’ and the ‘Trachyostraca’ within an assemblage (deep water with up to 90% ‘Leiostraca’, platform environments with 0–10% ‘Leiostraca’) is often used to conclude on the inhabited paleoenvironments and water depths. The ratio was given for Asian examples from the Cretaceous (Hokkaido, Japan; Obata and Futakami 1977; Tanabe et al. 1978; Tanabe 1979; Matsumoto et al. 1981; Westermann 1996; Kawabe 2003) and the Triassic (Lukeneder and Lukeneder 2014). North American examples were given for the Cretaceous in the Western Interior (USA) by Scott (1940) and Batt (1989). Examples from Europe characterized by that ratio were mentioned by Kennedy (1971) for the Late Cretaceous of England, and by Lukeneder and Reháková (2004) for the Early Cretaceous of Austria. Assumptions on ammonoid assemblages were mostly drawn from the percentages of their constituents (Marchand 1984, 1992; Company 1987; Reboulet et al. 1992; Cecca et al. 1993; Reboulet 1996; Reboulet and Atrops 1997; Cecca 1998) and from absolute abundance variations (Reboulet et al. 2000, 2003).
Especially deep water areas (e.g., deep structural rises, basins and the distal shelves to upper slopes) from the Triassic to the Cretaceous were characterized by the presence/absence rate of smooth ‘Leiostraca’ (Oloriz 1976; Geczy 1982, 1984; Sandoval 1983; Clari et al. 1984; Bruna and Martire 1985; Galacz and Horvath 1985; Fourcade et al. 1991; Oloriz et al. 1991; Cecca 1992; Westermann 1996). Accordingly, the proportion of ‘Leiostraca’ among the ammonoids increases with depth (Westermann 1996).
Cecca (1992) assumed, based on Tithonian ammonoid associations, the ‘Leiostraca’/‘Trachyostraca’ ratio and the suture complexity of the Mediterranean, that most ammonoids were demersal swimmers (neritic nektobenthos, epipelagic; e.g., Aspidoceras, Simoceras, Pseudolissoceras) at depths from about 100–200 m. Additionally, pelagic benthic habitats (i.e., mesopelagic; e.g., Haploceras, Ptychophylloceras, Protetragonites) or intermediate habitats (Cecca 1992) were considered for other ammonoids, showing the wide range of presumed habitats. Late Cretaceous ammonoid assemblages from Hokkaido (Japan) show a gradient (‘Leiostraca’ vs. ‘Trachyostraca’) correlating to a neritic (above 200 m) to oceanic (below 200 m) gradient (Tanabe 1977, 1979; Westermann 1996). The baculitids were suggested to be planktic, hamiticones as epipelagic, scapihitids as epipelagic (at least juveniles), desmoceratids mostly as mesopelagic, phylloceratids as nektic and lytoceratids as planktic (Tanabe 1977, 1979; Fig. 18.7). ‘Leiostraca’ (desmoceratids only 1% in shallow areas) vs. ‘Trachyostraca’ (hoplitids up to 99% in shallow areas) show the same ratio in the Albian from Poland (Marcinowski and Wiedmann 1985). Shallow-water hoplitids were suggested to be demersal and desmoceratids to be pelagic with planktic heteromorphs. Westermann (1996) doubted the benthic lifestyle suggested for these heteromorphs by Marcinowski and Wiedmann (1985). According to Westermann (1996), the planktic lifestyle was also supported by the synonymy with planktic morphogroups (e.g., morphogroup 16; see Fig. 18.2) given by Batt (1989). The literature shows that the morphotype groups given in Batt (1989) and Westermann (1996) were often cited without being questioned. This led to numerous interpretations without testing. Analyzing the data and figures given in Batt (1989) reveals that a real morphotype–facies relation is rare (e.g., Calycoceras, Sciponoceras) or absent in most genera (e.g., Watinoceras, Collignoniceras). This is why Batt (1989) designated the habitats as “possible”. Nonetheless, stronger ribbing appears to dominate in shallower areas. A change in environmental conditions (e.g., oxygenation), as given by Batt (1993) and thus an ecologically dependent change in ammonoid assemblage composition, is more likely. This hampers different groups from inhabiting distinct zones (e.g., near sea floor; Monnet 2009, Chap. 18.8.3). Other forms might be able to live above such hostile conditions due to a different mode of life.
Numerous authors (Ziegler 1967; Kennedy and Cobban 1976; Westermann 1990, 1996; Cecca 1992; Fernandez-Lopez and Melendez 1996) have drawn attention to the ecological differences between these two major groups of ammonoids: the streamlined, smooth-shelled ‘Leiostraca’ and the strongly ornamented ‘Trachyostraca’ (Fig. 18.1). The quantitative composition of the faunal assemblages is important both for paleoecologic and paleobiogeographic studies. The fluctuation in relative abundance and diversity of the various morphotypes reflects environmental changes related to sea-level changes (Reboulet and Atrops 1995; Reboulet 1996, Reboulet et al. 2005).
The ‘Leiostraca’ have been declared to indicate pelagic to deep-shelf habitats (ca. 250–300 m depth; Westermann 1990, 1996; Cecca et al. 1990; Mouterde and Elmi 1991). In contrast, the ‘Trachyostraca’ are generally considered to have inhabited neritic shallow-water environments (ca. 30–100 m depth; Westermann 1990; 1996). Kennedy and Cobban (1976) described local abundances of ‘Leiostraca’ in shallow-water deposits, uncover the weakly ornamented ‘Leiostraca’ as not exclusively deep-water inhabitants. Proportions of ‘Leiostraca’ vs. ‘Trachyostraca’ were often used as indicators for habitat depth (Ziegler 1967). Early Cretaceous ammonoid assemblages of Austria (Lukeneder and Reháková 2004) are represented by means of spectra that illustrate the proportions of ‘Leiostraca’ and ‘Trachyostraca’. Ziegler (1967) proposed that the dominance of ‘Leiostraca’ at a specific stratigraphic level was indicative of deep-water conditions (ca. 300–400 m depth). However, as cautioned by Kennedy and Cobban (1976), ‘off–shore’ rather than ‘deep-water’ per se may be the more appropriate appellation. Westermann (1990, 1996) and Cecca (1992) regarded ‘Leiostraca’ as pelagic, capable of occupying deep-water habitats, in contrast to the neritic shallow-water ‘Trachyostraca’. Fernandez-Lopez and Melendez (1996) have correlated the relative abundance of ‘Leiostraca’ with changes accompanying eustatic rises in sea-level (see also Gygi 1986).
These percentages may be interpreted as indicating either the presence of offshore (pelagic) influences or local postmortem drift of the otherwise pelagic ammonoids (Tanabe 1979; Cecca 1992, Batt 1989, 1993; Lukeneder and Reháková 2004). Stratigraphic units with percentages of ‘Leiostraca’ ranging from 5 to 10% reflect environments from the shelf or upper slope, as indicated by the ammonoid genera analysis from the Vocontian Trough (SE-France; Bulot 1993; Reboulet 1996). Similar fluctuations in the proportions of ‘Leiostraca’ and ‘Trachyostraca’, presumably reflecting a comparable interplay of environmental factors, have been documented by Sarti (1986a, b) from Northern Italy, by Christ (1960), Wendt (1963) and D’Arpa and Meléndez (2004) from Western Sicily and Hungary, by Cecca et al. (1990), Cecca (1988, 1992), Fernandez-Lopez and Melendez (1996) from the Western Tethys, and by Stevens (1997) from New Zealand.
As stated in Buckman’s Law of Covariation, the ammonoid shell shape and ornamentation are typically correlated. Compressed, involute forms display a fine ornamentation, while more depressed, evolute morphotypes show stronger ornamentation (Yacobucci 2004). Such covariation implies a link between the morphogenesis of shell shape and ornamentation rather than an ecological control. In contrast, there is evidence that ornament growth is controlled by genetics and that shell shape is merely influenced by environmental factors. This is known as Buckman’s Paradox. Yacobucci (2004) suggested that shell shape and rib growth are controlled by different processes. Ribbing appears to be more constrained than shell shape, consistent with the view that ornamentation is more tightly controlled by the genetic growth programme in ammonoids (Yacobucci 2004).
More recently, Doghuzhaeva et al. (2010) suggested a middle pelagic habitat for lytoceratids (i.e., Eogaudryceras) from the Aptian of Russia, also suggesting that they were capable of diurnal vertical migration and hovering (see also Mutvei and Dunca 2007). These suggestions were based on narrow siphuncles with long septal necks and shell analyses (e.g., wrinkle layer, myostracum) combined with external features such as numerous thick collars stabilising the conch. Prominent collars feign a strong ribbing of an otherwise almost smooth to fine-ribbed form. Similarities in lifestyle were proposed in Eogaudryceras with the deep-water Spirula (Doghuzhaeva et al. 2010, Chap. 18.5.2).
8.2 Sexual Dimorphism and Sea-Level
Sexual dimorphism occurs with sex ratios up to 100:1 for either macroconchs or microconchs (Callomon 1980; Howarth 1992; Davis et al. 1996; Westermann 1996; Davis et al. 1996; Klug et al. 2015b). The sex ratios may diverge considerably even between closely related species in the same assemblage (e.g., Stephanoceras; Westermann 1996). In such cases, the dimorphs might have lived in segregated swarms. In normal occurrences of adult ammonoids (both dimorphs), dispersed or abundant, the animals died of old age (Westermann 1996). If sexual dimorphism can be observed in a single species from adjoining areas, one can speculate that adult females left hypoxic, cold basinal waters to spawn on the warm, solid, and oxygenated bottom of the adjoining platform, and died afterwards. A considerable amount of ammonoid taxa was, at least sporadically, separated according to sex before mating and/or during spawning. Callomon (1980) also suggested that female ammonoids return to their primary habitats. A sexual and age (juvenile and adult) separation seems to have existed in numerous ammonoid taxa (Kennedy and Cobban 1976). These authors suggested that earlier ontogenetic stages inhabited offshore waters, contrasted by shallower areas in adults, hence exhibiting an ontogenetic change of habitats. This is somewhat supported by stable isotope data (Lukeneder et al. 2010; Fig. 18.9).
Two Early Cretaceous ammonoid taxa from Austria (Northern Calcareous Alps) were examined with respect to the evolution of shape as well as morphology and environment (Lukeneder and Harzhauser 2003; Lukeneder 2004). Both ammonoids outnumber other ammonoid taxa and were deposited in mass-occurrences: the planispiral Olcostephanus (98%; Lukeneder 2004) and the criocone Karsteniceras (91% Lukeneder 2003; see Chap. 18.8.4).
The antidimorphs of the Valanginian Olcostephanus lived in two different environments and adapted morphologiclly to these somewhat different environments (shallow to deep gradient; Lukeneder 2004). Lithological differences around the Olcostephanus Level are due to an altered paleooceanography and reflect sea-level fluctuations (transgressive facies) during the Early Cretaceous (Hoedemaeker 1990; Lukeneder 2004); Lukeneder and Reháková 2004. Faunal turnovers, mass-occurrences, and migrations have often been considered to be controlled by transgressive/regressive cycles in various ammonoid groups (Rawson 1981, Hoedemaeker 1990; Lukeneder 2004). Studies conducted in SE France provide evidence of ammonoid distribution linked to facies and point to faunal assemblage variations between basin and outer shelf (Bulot 1993; Reboulet 1996; Reboulet and Atrops 1997). In Olcostephanus, several species are restricted to the outer shelf facies while others occur in basinal facies (Bulot 1993). Data from the western Tethys confirm this facies-link of Olcostephanus (N Caucasus in Kvantaliani and Sakharov 1986; Spain in Company 1987; Switzerland in Bulot 1989, 1992) and underline a general trend for the entire Tethyan Realm (Bulot and Company 1990).
Variations in the ratios between ‘Leiostraca’ and ‘Trachyostraca’ (Fig. 18.1) in Austria were investigated by Lukeneder and Reháková (2004). The Olcostephanus Level (upper bathyal or deep sublittoral) shows a value from 5–10% of ‘Leiostraca’ (lytoceratids, phylloceratids). The changes in ammonoid faunal spectra from the Steinmühl Formation (Lukeneder and Reháková 2004) to the Schrambach Formation reflect a complex of changes. This includes an altered pelagic influence, the sedimentological change related to the prograding development of a fan system, eustatic changes in sea level, and changes in bioproductivity (Lukeneder and Harzhauser 2003; Lukeneder and Reháková 2004).
8.3 Dysaerobic to Anaerobic Environments
As noted by Westermann (1996), a crucial component in correlating ammonoid occurrences is the oxygen content of the water column. Ammonoids are widely distributed in well-oxygenated water, rare in dysoxic layers and absent in anoxic environments. Such facies are characterized by the trace fossil content, sulphur values (e.g., pyrite formation) and total organic carbon contents. Hypoxic (oxygen-poor or dysaerobic, O2 = 0.3–1.0 ml/l; anaerobic O2 < 0.3 ml/l) sea water conditions are important in benthic habitats (mostly bivalves) and for pelagic organisms (e.g., ammonoids) throughout the Phanerozoic (Westermann 1996). The geochemical and biotic conditions are important in the seawater, at the seawater–sediment interface, and in the sediment of such areas (Boucot 1981; Morris 1979, 1980; Wilde and Berry 1984; Wignall 1987, 1990; Jenkyns 1988; Brett et al. 1991; Oschmann 1991; Savrda and Bottjer 1991; Tyson and Pearson 1991; Wignall and Hallam 1991, 1993; Kauffman et al. 1992; Monnet 2009).
Ammonoids, as indicators for environmental conditions, reacted on changes in water geochemistry. Anoxic, and in parts dysoxic, conditions, induced the deposition of black shales. These are commonly characterized by the absence of ammonoids, depending on where the oxygen-depleted layers were located in the water column (e.g., near-bottom or in the water column). If bottom waters were anoxic, demersal and benthic forms were absent; only nektopelagic or planktic ammonoids appear in the sediment or laminated facies. The vanishing of demersal forms during anoxic to dysoxic conditions was shown by Batt (1993; see also Monnet 2009). The high metabolism of ammonoids, especially of larger forms, excluded most of them from dysoxic–anoxic conditions. Exceptions were reported repeatedly (Rieber 1977; Donovan 1993; Vašíček and Wiedmann 1994; Cecca 1997, 1998; Lukeneder 2003, 2005, 2007; Lukeneder and Smrečková 2006) for small heteromorphs. They were probably adapted to such conditions, at least for short times for preying or during migration phases. Such small demersal ammonoids were perhaps able to survive briefly in dysoxic waters as reported for the Cenozoic nautilid Aturia (Schlögl et al. 2011) and the Recent Nautilus (Wells et al. 1992). A similar strategy was suggested for several species of Baculites (Westermann 1977, 1996): speculatively, they normally inhabited aerobic waters but could dive shortly into dysoxic zones to feed near the sea floor. Well-oxygenated waters probably enhanced the growth of eggs. Accordingly, full oxygenation of the primary habitat is required for early ontogenetic development (Westermann 1996; Laptikhovsky et al. 2013). Adult female ammonoids are thought to have spawned in well-oxygenated habitats.
During the Devonian nekton revolution (Klug et al. 2010), ammonoids started to explore various ecological strategies (demersal, planktic, nektic) and to occupy various habitats in the water column. This also depended on the oxygen content and is reflected in black shale deposition, affecting the presence of distinct taxa. Boston and Mapes (1991) postulated for Carboniferous anoxic–dysoxic occurrences that very young ammonoids were benthic inhabitants, unable to escape the periodic benthic anoxia, but Mapes and Nützel (2009) revised this view profoundly. Adult ammonoids lived in the water column or were capable of surviving brief hypoxic events (Boston and Mapes 1991), like extant Nautilus (Wells et al. 1992).
Late Cretaceous black shales from Hokkaido (offshore shelves and deep basins) were deposited in the deep waters of the continental margin (Hirano 1986, 1993; Westermann 1996). They yield abundant desmoceratids, with either planktic Desmoceras (inflated) or nektic Pachydesmoceras (compressed). Abundant Desmoceras were overcome by deepwater benthic anoxia, indicating a demersal habitat (Westermann, 1990, 1996). Heteromorphs like Turrilites were suggested as being planktic vertical migrants (Westermann 1996). Hirano (1993) reported that ammonoid populations were affected by oceanic anoxic conditions prevailing during the Cenomanian-Turonian boundary in Japan. Desmoceras japonicum died out, enabling the spread of Tragodesmoceroides subcostatus. The latter species arose from the former by migrating into shallow, oxygenated waters. Thus, the possible escape strategies of Cretaceous ammonoids (Hirano 1988, 1993) were a) to survive in other niches of the outer shelf to upper slope environments, b) to undergo phyletic evolution and c) to adapt to changing oxygen conditions. More recently, Kawabe (2003) showed, for Albian-Cenomanian assemblages from Japan, the correlation from depressed (low-energy, offshore) and compressed shell morphologies (high-energy regimes) in desmoceratids (Desmoceras) and gaudryceratids (Zelandites, nektic, nearshore) from the same localities (inner shelf, compressed vs. slope, depressed forms). Kawabe (2003) also noted that the shell ornamentation does not depend on lithofacies within these faunas, but that whorl section varies with lithofacies, reflecting environmental conditions (e.g., high or low energy, waves, currents; see Jacobs et al. 1994).
Unstable bottom-water conditions prevailed in some more or less hypoxic epeiric basins (ca. 50–100 m depth), marked by the deposition of black-shale facies (Little et al. 1991; Oschmann, 1991; Wignall and Hallam 1991; Westermann 1996). Westermann (1996) noted a major second component in correlating ammonoid occurrences and oxygenation of the water column, namely the salinity. Nektobenthic ammonoids could not survive when oxygen levels became low or salinity changed drastically. The range of tolerable salinity in modern cephalopods is 16–41‰ (Forsythe and Van Heukelem 1987; Cochran et al. 2003).
The Givetian in the USA (New York, Pennsylvania) shows that the total number of species increased when the oxygenation of bottom waters improved (Kammer et al. 1986; Brett et al. 1990, 1991). Within such facies, the presence of in situ aptychi can be important evidence for the autochthonous deposition of ammonoids (Frye and Feldman 1991). Cephalopod abundance peaks within a dysaerobic biofacies (House and Price 1985). The presence of all size classes and exceptional preservation suggested to House and Price (1985) a demersal swimming habit, with the animals preying on epibenthos (Westermann 1996). Additionally, bactritids were planktic (vertical migrants), living in midwaters (Westermann 1996).
Triassic dysoxic conditions prevailed in Carnian times (e.g., Reingrabner facies). A remarkable site for that is the Polzberg locality described by Glaessner (1931) and Krystyn (1991) in Austria. Krystyn (1991) suggested a relatively shallow intraplatform basin with dysoxic or even occasional anoxic conditions, and termed it a Konservat Lagerstätte based on the enormous amounts of well-preserved fishes, conodont clusters, fecal pellets, coleoids and mass occurrences of ammonoids (Austrotrachyceras) with in situ buccal masses (Doghuzaeva et al. 2007). Benthic life is represented by mass occurrences of the bivalve Halobia (comparable to Posidonia in the Toarcian Posidonia Shale) and rare crustaceans. Nektic life is dominated by the almost monospecific ammonoid fauna with Austrotrachyceras (n > 1000, all size classes), accompanied by bony fishes and sharks. The ammonoids and members of other nektic groups were able to live above the oxygen-deficient layers present throughout the sedimentary succession of this locality. Austrotrachyceras was preyed upon by fishes and ichthyosaurs, as is evident from coprolites with numerous incorporated specimens (own observations).
The Toarcian Posidonia Shales are characteristic for NW Europe and other parts of the world (Jenkyns 1988). These shales consist of laminated black shales, marly shales and bioturbated (hypoxic) carbonates (Urlichs et al. 1979). Abundant oxyconic harpoceratids (max. habitat depth c. 50 m, Hewitt 1996; Westermann 1996), platyconic hildoceratids (> 60 m; Hewitt 1996; Westermann 1996), typical serpenticonic dactylioceratids (c. 70 m; Hewitt 1996; Westermann 1996), dominate over Phylloceras and Lytoceras (i.e., ‘Leiostraca’). Squid predation (in the upper water column) is indicated by clusters of harpoceratinid fragments (Mehl 1978b). Abundance peaks of Dactylioceras (c. 70 m, Hewitt 1996; Westermann 1996) might be traces of mass mortalities by anoxic admixing during storms (Westermann 1996). Harpoceratids and hildoceratids frequently containing in situ aptychi hint at hypoxic habitats or rapid burial under such conditions (Westerman 1996). Normally coiled dactylioceratids such as Peronioceras were rare in this highly bituminous facies, suggesting that they might have been more bottom-dependent than Dactylioceras (Schmidt–Effing 1972; Loh et al. 1986; Westermann 1996). Posidonia Shales of Germany were deposited in shallow seas characterized by varying salinities (low salinity due to freshwater input) and periodic anoxic events (Brumsack 1991; Röhl et al. 2001; Schmid–Röhl and Röhl 2003). Few groups tolerated short anoxic or dysoxic pulses. Among the adapted organisms were fishes and rare ammonoid taxa: they survived such environmental changes (i.e., salinity and oxygen; Westermann 1996). Facies distributions and faunal ammonoid assemblages confirm a planktic lifestyle of Dactylioceras (Tintant et al. 1982). A euryhaline habit was suggested (Westermann 1996) for Dactylioceras (e.g., surface-water habitat and dispersal across shallow shelves; Elmi and Almeras 1984). More probably, these dactylioceratids were midwater drifters as indicated by their shell and siphuncle strength, which is average for Ammonitina (Westermann 1996). The global distribution of Dactylioceras species in all facies (Schmidt–Effing 1972) supports a planktic habit in epeiric and neritic seas as well as in the oceans. This is in contrast to Keupp’s (1997, 2000) interpretation based on the huge number of apertural injuries in Dactylioceras (e.g., caused by crustaceans on the sea floor), assuming a demersal life. Harpoceratinae and Hildoceratinae were suggested as being nektic (Westermann 1996).
A black shale sequence was reported from the Middle Jurassic (Bathonian–Callovian) Los Molles Formation in Argentina (e.g., Neuquén Basin; Riccardi and Westermann 1991; Riccardi et al. 1992). The invertebrate fauna consists mainly of abundant adult ammonoids with very low diversity but high intraspecific variation (Westermann 1996). Adult sphaeroceratids (Eurycephalites, Stenocephalites) and reineckeiids (Neuqueniceras) were interpreted as adults that died normally, whereas the juveniles were killed by single events (e.g., anoxia) in masses. Young stages probably lived in deeper waters that were exposed to periodic upwelling and, after death, were drifted and accumulated by bottom-water currents (Westermann 1996). In the basin center (max. depth), abundant small Ptychophylloceras (oceanic to mesopelagic; Westermann 1996) occur. O isotope data in similar forms of phylloceratins (Hypophylloceras, Phyllopachyceras) from the Campanian of Japan yielded depth ranges for that ammonoid group of 100–300 m (Chap. 18.5.6, Fig. 18.11). This is the upper mesopelagic to middle epipelagic zone. From estimates of habitat paleodepth in the Neuquén Basin, Ptychophylloceras inhabited shallower areas (Westermann 1996; Hewitt 1993, 1996).
The Callovian to Tithonian black shales (Lower Oxford Clay and the Kimmeridge Clay of England; Cope 1967, 1974; Callomon 1985; Duff 1975; Hudson and Martill 1991; Morris 1979; Oschmann 1991; Wignall 1990; Wignall and Hallam 1991) yield monospecific ammonoid assemblages with Kosmoceras. These are usually adult and strongly dimorphic with a high intraspecific variation. Depth estimates for the Kimmeridgean area are 30–50 m (Westermann 1996). Assuming normal salinity, the sluggish ammonoids lived at 17–20 °C (max. 16–23 °C), inhabiting deeper, slightly oxygen-deficient (upper dysaerobic) waters, away from predators (O'Dor and Wells 1990). The more serpenticonic and less sculptured kosmoceratids are found in the deepest facies (Marchand et al. 1985), which does not imply exlusively demersal habitats (Westermann 1996).
The Kimmeridgian to Tithonian of the Antarctic Peninsula, the anaerobic Nordenskjold Formation, was deposited in a deep basin (Doyle and Whitham 1991). The small (10–110 mm) ammonoids there belong to coarsely sculptured, serpenticonic perisphinctids. 70% of all ammonoids are infested by bivalves syn vivo. Mass mortality events occured, caused by storm-induced admixing of anoxia (Westermann 1996). Higher in the section, small, smooth, discoconic haploceratids lack epizoans, suggesting that the perisphinctids were planktic drifters in the upper dysaerobic zone. The ‘leiostracan’ Haploceras questionably swam in the aerobic zone, as assumed previously based on its cosmopolitan distribution (Westermann 1996). It was immune to drag-increasing epizoans (smooth surface; perhaps a special periostracum). In deep turbiditic sedimentation areas, the ammonoid fauna consists of dominantly finely costate Virgatosphinctes and Lithacoceras. Accessories were costate, platyconic berriasellids, planorbiconic olcostephanids, as well as haploceratids. Again, only perisphinctids carry epizoans. These perisphinctids are mesodomic, compressed planorbicones to discocones that were presumably moderately mobile and perhaps bottom-feeders.
The Albian Mowry Shale of USA (Western Interior) includes hoplitid assemblages (Neogastroplites; Reeside and Cobban 1960; Batt 1989). It appears that the immature, sluggish hoplitids lived in midwater of the shallow, epeiric sea above dysaerobic bottom waters (Landman and Waage 1993). More recently, Monnet (2009; inferred by Batt 1989) showed that, in the Late Cenomanian of the Western Interior Seaway (USA), nektobenthic forms such as Acanthoceras and Mammites (strongly ornamented) were the first to vanish during anoxic/hypoxic bottom conditions (Oceanic Anoxic Event 2). Subsequently, ancyloconic heteromorphs (Hamites, Allocrioceras) of the middle pelagic zone were eliminated by an upward migration of the oxygen minimum zone (OMZ). Nonetheless, Monnet (2009) showed that this is not a worldwide phenomenon and thus not the rule or main mechanism for ammonoid extinctions. This was illustrated in examples from Tunisia contradicting the ‘rising-anoxia’ model. In fact, Monnet (2009) reported an increase in ammonoid taxa during the formation of black shales in Tunisia during the late Cenomanian.
In contrast, an oxygen-dependent lifestyle was indicated by Landman et al. (2012) for scaphitids, e.g., from the Campanian Western Interior (Pierre Shale, USA). Hoploscaphites and benthic fauna did not occur during phases of dysoxic bottom waters: only Placenticeras and Baculites were present, both inhabiting higher zones in the water (see also Bearpaw Shale in Canada, Tsujita and Westermann 1998). During phases of good oxygenation, Hoploscaphites (juveniles and adults) recolonized the basinal areas, co-occurring with other ammonoids, except during spawning events, subsequently followed by mass mortality of females (Landman et al. 2003). A deeper habitat for Hoploscaphites was also supported by stable isotope data, in contrast to the shallower habitat of Baculites (Cochran et al. 2010a, b; Kruta et al. 2014; Fig. 18.9). Henderson and Price (2012) challenged that shallow habitat interpretation after investigating baculitids (Sciponoceras) from the Late Cretaceous of Australia. The authors argued for a nektobenthic mode of life in this baculitid, based on relatively positive δ18O data (mean calculated paleotemperature 21 °C; Henderson and Price 2012). These data contrasted with those for bivalves and gastropods. The authors proposed an adaptation among baculitids as specialists in demersal predation (see also Kruta et al. 2011).
Santonian to Campanian bituminous, laminated shales of the Bearpaw Formation and coeval Pierre Formation (both Western Interior, USA) contain abundant ammonoids. The main faunal components are Baculites and large Placenticeras. Lithofacies characterized by diverse benthos indicates better-oxygenated bottom water, yielding scaphitids and accumulations of neanic to juvenile baculitids (Landman 1987; Westermann 1996). Placenticeras meeki is strongly attacked in shallow, surface waters by predators (e.g., mosasaurs; 10–20% of all specimens; Kauffman 1990, Hewitt and Westermann 1990b; Westermann 1996). Oxygen and carbon isotope analyses (Chap. 18.5) revealed markedly reduced salinities of the surface waters inhabited by P. meeki (Westermann 1996). This clearly indicates that the large predator Placenticeras occasionally lived in brackish surface waters (e.g., < 30 m below surface; Westermann 1996). This contrasts with the deeper habitats assumed by Ziegler (1967, 1981) and Westermann (1990) above wave base. Shell characteristics of P. meeki are a complicated suture, low sutural amplitude index (Batt 1991), low resistance to hydrostatic pressure, and a high predation rate (Hewitt and Westermann 1990b). Baculites was also commonly attacked by mosasaurs, evident from several punctures (Kauffman 1990; Westermann 1996; Tsujita and Westermann 2001). This points to a pelagic habitat (Westermann 1977, 1996) in the aerobic waters. Baculites is therefore suggested as being capable of diving rapidly (5–100 m deep into dysoxic zones), feeding on the bottom. The maximal depth range for Baculites was corroborated by isotope data (Fatherree et al. 1998; Lukeneder et al. 2010; Fig. 18.9): relatively shallow from 50–100 m in epipelagic seas (Fig. 18.7, Table 18.1). Baculites apparently shifted into shallower waters after reaching maturity (Fatherree et al. 1998; Lukeneder et al. 2010). In contrast to the pure surface-water dweller Hypancanthoplites (Lukeneder et al. 2010), it thus changed its depth preference and seems to have started as a juvenile in the lower epipelagic zone. Similar depths for Baculites were given (40–120 m) based on SiSI values (Hewitt 1996; Westermann 1996). Abundant but smaller species and juveniles of the giant species were suggested to be vertical migrants (Klinger 1980; Batt 1989; Westermann 1990), probably mostly in midwater. The giant adults (> 0.8–1 m) of B. cuneatus, in turn, became mainly demersal swimmers.
8.4 R-Strategy in Oxygen-Depleted Habitats
The Barremian heteromorph ammonoid Karsteniceras (N Calcareous Alps, Austria) evolved under intermittent oxygen-depleted conditions associated with stable, salinity-stratified water masses; it exhibited mass occurrences (Lukeneder 2003, 2005, 2007; Lukeneder and Smrečková 2006). Lithological and geochemical analyses (e.g., dark laminated shales; CaCO3, TOC, S) were combined with investigations of trace fossils, microfossils and macrofossils. This led to the assumption of an invasion of an opportunistic Karsteniceras community (with sexual dimorphism, most probably applicable to the whole leptoceratoid group) during unfavorable conditions (Lukeneder 2003, 2007; Lukeneder and Tanabe 2002). The significant effect of autecological stress, caused either by changes in abiotic environmental factors (oxygen content, salinity, depth) or by synecological stress associated with biotic competitors occupying the same environment was shown by Lukeneder (2003, 2007). Jaw apparatuses are found in situ or isolated but are associated with a mass-occurrence of the genus Karsteniceras, reflecting a fast and autochthonous deposition. Low energy on the sea floor (absence of bottom currents) and dysaerobic conditions prevented predators from isolating the shells from the jaw apparatuses and led to the unusual preservation of the ammonoid conch-jaw association. The positions of the lower jaw in the body chamber of Karsteniceras indicate that the dead ammonoid bodies were not subjected to long postmortem drift; they rapidly became waterlogged and sank to the sea floor. These exceptional preservational features are typical for ‘Konservat–Lagerstätten’ (Seilacher et al. 1976), which always show exceptional preservation of articulated hard parts and/or soft body parts .
Terms often used in ammonoid ecology such as allochthonous and autochthonous, must be used with caution when linked with fossil cephalopods because shells could have been subject to major postmortem drift. In most cases, the water depth or primary habitat is unknown. A certain likelihood concerning the autochthonous deposition of ammonoid specimens, hence the primary depositional area, is warranted by the preservation of in situ buccal masses located in the body chamber (Schindewolf 1958; Bandel 1988; Frye and Feldman 1991; Summesberger et al. 1996; Lukeneder and Tanabe 2002).
Karsteniceras inhabited areas of stagnant water with low dissolved oxygen, showing abundance peaks during times of oxygen depletion, which partially hindered other invertebrates from settling. An invasion of the r-strategist Karsteniceras assemblage during unfavorable conditions over the sea bed was proposed. The host layers do not reflect catastrophic mass mortality. The specimens are not concentrated in single horizons, but are abundant in layers of 3–10 cm thickness, reflecting 100,000 of years (calculations based on ammonoid zonation).
Taking the speculations on ammonoid life-habitats into account, demersal forms feeding from the sea floor should be rare or absent in the anoxic to dysoxic levels of the water column (Batt 1993). Vašíček and Wiedmann (1994) noted the possibility that the biotope of the Leptoceratoidinae was close to stagnant, poorly oxygenated environments. This has been interpreted to reflect the opportunistic bahavior of some heteromorphs (bochianitids, leptoceratoids) in unfavorable environments (Cecca 1997, 1998). Ecology, paleobiology and habitats of small heteromorphs such as leptoceratoids and spiroceratids were interpreted to have occupied various habitats (Thieuloy 1966) such as nektic above an anoxic bottom (Rieber 1977), on the distal shelf (Westermann 1990, 1996), in turbiditic environments (Vašíček and Wiedmann 1994), and demersal, capable of life in dysoxic waters (Lukeneder 2003, 2005) .
8.5 Oxygenated Shallow-Water Habitats
Carbonate platforms are a special environment for ammonoid occurrences. It is relatively shallow and well oxygenated, mostly well agitated by bottom or surface currents. Some early ammonoid taxa like Devonian clymeniids were suggested to appear within such environments. They exhibited a morphological depth gradient with coarse ribbing in shallow and fine ribbing in deeper habitats (Korn 1986, 1988, 1992; Westermann 1996) .
Middle Triassic ceratitids were the main inhabitants of shallow, epeiric platforms. They probably tolerated a wide range of salinities (euryhaline). Sutural simplification occurred in several immigrant clades from the deeper Tethys in the German Muschelkalk (Urlichs and Mundlos 1985; Klug et al. 2005). Coarse sculpture is also commonly associated with a basin-slope habitat, whereas a basinal pelagic habitat is usually indicated by smooth or weakly sculptured discocones. A few taxa (Monophyllites, Psilosturia) probably lived on the continental slope of the oceanic region (Westermann 1996), but based on their shell strength were still epipelagic. In the Late Triassic, radiation affected almost all morphs. The proportion of oceanic ‘Leiostraca’ greatly increased, and the first heteromorphic ammonoids appeared with torticone, orthocone and gyrocone morphotypes (Westermann 1996).
A more Recent study on Triassic ceratitid ammonoids was performed on a mass-occurrence (Kasimlarceltites; n> > 300 million; M and m, embryonic to adults) from Turkey (Lukeneder and Lukeneder 2014; Lukeneder et al. 2014; Lukeneder and Mayrhofer 2014). Kasimlarceltites is morphologically similar (i.e., serpenticone, highly evolute; Fig. 18.6, 18.12) and taxonomically closely related to other celtitid ammonoids including Aplococeras, Lecantites and Celtites. These genera were interpreted by Assereto (1969) as shallow ‘platform faunal associations’ from Latemar or Marmolada platforms (Dolomites, Italy). The celtitids are rare to absent in deeper basinal environments (Assereto 1969). The ammonoid-facies relation was interpreted to be controlled by ecological factors (Brack and Rieber 1993, 1996). Kasimlarceltites probably inhabited environments on the inner- to mid-ramp, adjacent to the Carnian platform (Lukeneder and Lukeneder 2014; Lukeneder and Mayrhofer 2014; Fig. 18.12). They were influenced by this environment and may have migrated near the sea floor, comparable to extant Nautilus. Assumptions on a facies or depth gradient dependency in celtitids (Kasimlarceltites) fit well into the interpretation of the Turkish material. For ammonoid assemblage-facies relations in the Mesozoic Era see Westermann (1996). Numerous layers are dominated by small ammonoids (< 30 mm) accompanied by shallow-water faunal components (transported ammonoids, bivalves, corals, gastropods, sponges) derived from the platform to inner ramp. They were therefore interpreted as being redeposited from shallower areas down to a shelf margin or mid-ramp position (Lukeneder et al. 2012). In contrast to Assereto (1969), additional findings of celtitids were presented by Manfrin et al. (2005) from adjacent basinal series surrounding the Latermar platform. The faunas from ammonoid layers were described as storm deposits. These were transported and comprise mixed faunal elements (pelagic ammonoids and benthic gastropods; Manfrin et al. 2005). This is almost identical to the assemblages found at Aşağiyaylabel. The use of such assemblages as characteristic elements for platform deposition or even primary habitat of celtitids is still arguable and needs more taxonomic and correlation work. Hence, a facies dependency of Kasimlarceltites in the carbonate member at Aşağiyaylabel must be considered with reservation. Taphonomic analyses rather point at allochthonous deposition after considerable transport (current, turbidites, mass flow) within ammonoid-mass occurrences (Lukeneder et al. 2014). The occurrence of single layers comprising mostly transported conchs of juvenile or exclusively adult shells of Kasimlarceltites, and beds bearing mostly adults, support the hypothesis that different ontogenetic stages inhabited different habitats (Lukeneder and Mayrhofer 2014; Lukeneder et al. 2014).

The gregarious ceratitid ammonoid Kasimlarceltites krystyni (max. diameter 35 mm) in assumed spawning grounds and the nektic actinopterygiid fish Saurichthys (max. length 1 m) in shallow waters of a Triassic carbonate platform. Reconstruction (by 7reasons Media Company) based on studies of Upper Triassic mass occurrences of the ammonoids in the Taurus Mountains (SW Turkey; Lukeneder and Lukeneder 2014)
Similar hypotheses on the relation of closely resembling morphotypes versus habitat and lifestyle of serpenticone morphotypes were given for Paraceltites (Permian; Spinosa et al. 1975), Psiloceras (Jurassic; Westermann 1996), and Celtites (Triassic; Rieber 1973, 1975). These were presented and commented upon in Westermann (1996). Spinosa et al. (1975) proposed a “tropical platform limestone” depositional area for such serpenticone ammonoid types and a presumable planktic lifestyle was assumed (Fig. 18.6, 18.12) for such costate, celtitid morphotypes (Rieber 1973, 1975; Westermann 1996). As postulated by Klug (2002) and Klug and Korn (2004), ontogenetic alterations in morphology reflect, at least in early ammonoid taxa, a change in mode of life. This is because these changes appear several times independently .
9 Autecology in Ammonoids
Neumayr (1883), Uhlig (1911), Scott (1940), Ziegler (1967, 1981), Kennedy and Cobban (1976), and, more recently, Westermann (1990, 1996) have studied the autecology of ammonoids and related ecological factors . This relation is denoted by the variation and ratio of shell morphologies and assemblages in general, related to tectonically caused or eustatic sea level fluctuations (Enay 1980; Enay and Mangold 1982; Marchand and Thierry 1986, 1997; Lominadze and Sakharov 1988; Wiedmann 1988a; Cecca 1992; Bulot 1993; Rawson 1993; Hantzpergue 1995; Reboulet and Atrops 1995; Reboulet 1996; Enay and Cariou 1997; Pucéat et al. 2003). Such environmental changes are accompanied by changes in many other ecological parameters. This has consequences for the organisms living in the affected environment. Neige et al. (1997) stressed the close relation between disparity and (eustatic) sea-level changes. These authors presented a new approach based on Callovian ammonoid faunas of France (Lominadze and Sakharov 1988, Callovian of Caucasus). Here, the morphospace calculations reflected the most important conch parameters (diameter, whorl height and width, etc). Mathematical estimations by Neige et al. (1997) resulted in a more precise use of ammonoid morphotypes as paleodepth indicators .
Oxygen is a biolimiting element for metazoans and one of the most important factors influencing species diversity and abundance in the marine realm. Batt (1993) suggested that the relationship between ammonoid shell morphotypes and position within the water column provides a useful tool in interpreting fluctuations in O levels in marine depositional systems of the Cretaceous Western Interior Seaway (N America). Suggested habitats in Batt (1993) were based on siphuncle strength index (SiSI), septal amplitude index (SAI) and septal complexity (Batt 1991, 1993; Ballentine 2007). As noted by Batt (1991) such indices are only valuable in combination with other shell morphological features (e.g., ribbing). The author concluded that the presence of nektobenthic morphotypes (e.g., Scaphites, Baculites) indicates oxygenation of the sea floor and lower parts of the water column, while that of pelagic morphotypes (e.g., Watinoceras, Borrisiakoceras) or shallow-water ammonoids (e.g., Placenticeras) may be used to interpret oxygenation higher in the water column. A nektobenthic mode in Scaphites is probably indicated by the highly injured specimens noted from Landman and Waage (1986; see also Landman 1986). Baculites was given with a depth range from 40–120 m based on SiSI values (Hewitt 1996; Westermann 1996; Fig. 18.9). The occurrence of shallow middle- and upper water column pelagic ammonoids in deeper-water stratigraphic intervals yielding no nektobenthic ones appears to be a useful indicator of oxygenated conditions within the upper part of the water column at a time when benthic environments were anoxic. The additional absence of ancyloconic heteromorphs in sediments deposited in water deeper than 100 m that contain upper-water pelagics indicates that anoxia extended to within 100 m of the surface (Batt 1993). Changes in ammonoid morphotype representation during upward migration of a poorly oxygenated water mass, hence fully oxygenated water column appears with all morphotypes (pelagic and nektobenthic). Bottom near anoxic conditions first eliminated nektobenthic ammonoids, and continued with anoxic conditions up to 100 m of the surface hampered life of pelagic forms (megaplanktic ancyloconic heteromorphs). Finally, only pelagic ammonoids from the upper water column are represented in sediments (Batt 1993; Reboulet et al. 2000, 2005; Lukeneder and Grunert 2013) .
10 Epizoans Versus Postmortem Epicoles in Ammonoids
Studying the spatial relationships among organisms that lived with each other provides insight into the synecology, autecology, and biostratinomic processes of these organisms. It also sheds light on their paleoenvironment and paleocommunity structure. Epibiosis is one of the few well-preserved biotic interactions in the fossil record dating back to the Early Paleozoic (Palmer 1982). A crucial aspect is to detect if infestation occurred before or after death. This determines whether an autecological or synecological relation is indicative for paleodepth, behavioral or habitat assumptions. Organisms that infested ammonoid shells within the water column are also referred to as pseudoplankton (Wignall and Simms 1990). If infestation occured postmortem, it can be speculated whether shell was infested while drifting or after it sunk to the sediment surface.
Davis et al. (1999) introduced terms to distinguish between the different aspects of epizoans. According to them, an organism that settles an lives on another one is an epizoan. By contrast, if the substrate object or organism is dead, Davis et al. (1999) suggested to call the animal living on it an epicole. These terms will be used subsequently. Countless examples of epizoa and epicoles on ammonoids have been described and some examples will be listed.
Ammonoid assemblages from the Early Cretaceous of Austria (Chap. 18.8.2; Lukeneder and Harzhauser 2003; Lukeneder 2004) yield abundant specimens of Olcostephanus guebhardi showing unique encrustations by placopsilinid foraminifera, bryozoans, serpulids, and crinoids (Phyllocrinus). The pattern of infestation clearly documents a 100% preference of these taxa for the inner shell surface, characterizing them as an epicole cavity-dwelling paleocommunity (Lukeneder and Harzhauser 2003). The encrustation in the body chamber furnishes evidence for postmortem settlement. O. guebhardi is suggested to have preferred shallow water and was most abundant in depths of 40–100 m (Bulot pers. comm. 2002). Major redeposition of the Austrian ammonoid shells is evident based on a mixture of olcostephanids with encrustations on their upper side and specimens with epibionts on their lower side within single layers. The shells sunk to the sea floor after death and became partly filled with sediment on the side of the shell facing the sediment. Thus, epicoles could not settle these areas of the body chamber. Only the ‘sediment free’ upper shell parts could be inhabited (Placopsilina, bryozoans, serpulids). This mode of infestation indicates that encrustation during postmortem drift can be excluded (Lukeneder and Harzhauser 2003). Phyllocrinus is one of the most frequent species associated with Olcostephanus. The calices of this sessile crinoid are often found in the sediment-infill of the ammonoids and the adherent holdfasts are exclusively found on the upper, outer side of the shell. In this soft bottom environment, the empty shells of Olcostephanus served as hardgrounds.
A more complex history is reported from the Valanginian to Aptian deposits of the Dolomites (N Italy). These deposits yield a rich ammonoid fauna showing unique epifaunal encrustations by an ahermatypic, solitary scleractinian coral (Lukeneder 2008). The coral encrusted only the outer shell surfaces; the inner surface remained unaffected. Shells of dead ammonoids sank to the bottom and became colonized by the coral (and serpulid) larvae, as documented by the location of the epibionts only on the upper side. Encrustation by the bivalve Placunopsis represents a different situation because both sides of the ammonoid shells were affected. This points to encrustation of floating or repeatedly turned (dead) or swimming (alive) ammonoids (Lukeneder 2008; Misaki et al. 2013). Such long-term infestation in the water column contrasts with coral settlement on the sea floor. Ammonoid specimens encrusted by Placunopsis never exhibit encrustation by corals (Merkt 1966; Meischner 1968).
The so-called coelobites are a somewhat neglected part of the vast spectrum of epibioses. They are defined by Ginsburg and Schroeder (1973) as cavity-dwelling organisms, often found in cryptic habitats. Studies on such habitats of modern environments have focused on coral reefs and reef-associated submarine caves (Jackson and Winston 1982; Rasmussen and Brett 1985; Meesters et al. 1991). Despite the smaller scale and limited spatial resource of the ammonoid shells, several parallels with modern cryptic habitats are obvious. As emphasized by Wunsch and Richter (1998) and Richter and Wunsch (1999), cavities and other cryptic habitats are “spatially confined habitats”, which provide living space for low-light adapted organisms. Among these, especially cryptic suspension feeders predominate. Cryptic habitats provide shelter for sessile species from predation and physical disturbance (Gischler and Ginsburg 1996). In modern cryptic habitats, the coelobite’s mode of life is mainly adopted by sponges, algae, serpulids, tunicates, bryozoans, and foraminifera. Several taxa such as placopsilinid foraminifera and the bryozoan Stomatopora have been shown to be characteristic constituents of all these paleocommunities. These taxa were also recorded adhering to ammonoid shells (Lukeneder and Harzhauser 2003).
Can such encrustation yield information on paleodepth and ammonoid habitats? The ecological depth ranges of the encrusting organism groups are enormous. The main encrusters on ammonoids are bivalves (e.g., oysters, Exogyra, Placunopsis, Gervillia, Anomia; Larson 2007; Misaki et al. 2013), gastropods (limpets), brachiopods (discinids; Seilacher 1982), serpulids, tunicates, solitary corals, bryozoans, and foraminifera. Larval stages often settle on secondary hard grounds (ammonoid shells). Unfortunately, synecological relationships (i.e., infestation during life) of ammonoid and encrusting taxa also can rarely be used to conclude on habitat or depth. Oysters (Merkt 1966), for example, inhabit depths from almost 0–850 m (Rooij et al. 2010), the bivalve Placunopsis hundreds of meters, serpulids can live in thousands of meters. Serpulids were able to grow with ammonoids in a synecological manner during life, on the ventral side of the shell. This makes them useful to estimate ammonoid growth rates and ages (Schindewolf 1934; Andrew et al. 2011). Andrew et al. (2011) suggest a distinct infestation by serpulids at different habitat depths for the Jurassic, mostly of living, juvenile ammonoids (Promicroceras) from the UK (Dorset). The dependency or at least preference of several serpulid species for shallower waters (6–12 m) led to the assumption that juvenile Promicroceras inhabited shallower areas: they were more highly infested than adults. Andrew et al. (2011) concluded that Promicroceras was negatively influenced by the parasitic serpulids, by the higher body weight and the increasing drag, dying at smaller diameters at an age of 2–3 years.
Synecological infestation by foraminifera potentially points to paleodepths of 15–70 m for Placopsilina attached to the ammonoid Olcostephanus (Lukeneder and Harzhauser 2003). Correspondingly, O. guebhardi was suggested to have preferred shallower shelf areas and was most abundant at depths of 40–100 m (pers. comm. Bulot 2002). Home scars of patellogastropods (limpets) on ammonoid shells were shown by Kase et al. (1994, 1998). Limpets, when settled postmortem on ammonoids, point to a drifting phase because limpets inhabit shallow areas and therefore attached near the sea surface (see Westermann and Hewitt 1995).
Maeda and Seilacher (1996) reported syn vivo and postmortem epibiont assemblages. The encrustation by inoceramids, oysters or Gervillia of both the flanks and venter in living ammonoids is remarkable (e.g., Jurassic Lytoceras and Harpoceras; Cretaceous Buchiceras; Seilacher 1960; Heptonstall 1970; Seilacher 1982; Maeda and Seilacher 1996; Larson 2007). One-side-only infestation by oysters was also reported as being caused by horizontal drifting of ammonoid shells on the water surface (e.g., Pectinatites; Donovan 1989). Sinking history can also be detected by analyzing the encrusting community and its position (Reyment 1973, Kauffmann 1978; Seilacher 1982). Kauffmann (1978) postulated the ‘benthic island’ infestation on sea floor for the Jurassic Posidonia Shale ammonoids, but this was refuted by Seilacher (1982) and Maeda and Seilacher (1996) based on the encrustation of both flanks (e.g., Lytoceras) in the water column. Further examples of syn vivo epizoans are discussed in Naglik et al. (2015).
11 Trophic Level
The trophic level of ammonoids was perhaps some kind of a mid-order omnivore level. They exhibit a range of feeding strategies from planktotrophic to microphagous (i.e. lower trophic levels). They were preyed upon by vertebrates (ichthyosaurs, mosasaurs, sharks, fishes; Fig. 18.13) and invertebrates (other cephalopods, crustaceans). Ammonoids ate possibly everything they got hold of from the lower trophic levels, leading to the interpretation as demersal herbivores, scavengers via microphagous carnivores to active, nektonic carnivores (Kennedy and Cobban 1976).
As noted by Westermann (1996), numerous feeding strategies can be envisioned for neanic and juvenile ammonoids. These speculatively fed on mobile prey with ejectile tentacles, visually on essentially planktic prey (ammonoid hatchlings, pseudoplanktic and planktic microorganisms such as ostracods or microgastropods), with non-ejectile tentacles or velar webs, and tactile or chemosensory feeding by ‘pseudoscavenging’ of organic particles floating and sinking at all depths (Reyment 1988; Westermann 1996). Reyment (1988) imagined a more scavenging and browsing mode, concluding that especially orthocones and heteromorph forms were incapable of active predation. The preying behavior is crucial for reconstructing the ammonoid habitat because ammonoids might have actively followed prey. This can cause ontogenetic or diurnal migrations, preying on different resources during their lifespan, or preying at night when some other predators sleep, comparable to Recent Nautilus.
Most recently, spectacularly preserved specimens of the Late Cretaceous heteromorph Baculites from South Dakota (USA) were reported by Kruta et al. (2011). They discovered buccal masses with radulae containing plankton found within. This represents an important indication for the trophic habits of these heteromorph ammonoids. One of the specimens revealed a planktic snail and three tiny planktic crustaceans (Isopoda) in its buccal mass (Kruta et al. 2011). As already stated by Westermann (1996), knowledge on stomach or crop contents is scarce (Lehmann and Weitschat 1973; Lehmann 1975, 1976, 1985, 1988; Summesberger 2000; Keupp 2007). See Klug and Lehmann (2015) for a review.
Numerous problems related to the dietary and feeding habits for the diversely shaped buccal organs are still unsolved, even for some extant cephalopods (Nesis 1986, 1987; Lehmann 1988). Westermann (1996) noted that besides the micro and mesophagy documented by crop/stomach contents, macrophagy and even duraphagous strategies might have also been present in ammonoids (Schindewolf 1958; Lehmann et al. 1980; Tanabe 1983; Nesis 1986; Tanabe and Fukuda 1987; Lehmann and Kulicki 1990; Hewitt 1993; Seilacher 1993; Keupp 2007). Westermann (1996) suggested that most ammonoids belonged to the pelagic food-web. Ammonoids fed directly on microplankton, perhaps at the base of the photic zone, themselves falling prey to larger ammonoids and vertebrates. An important feature for the ammonoid/prey interaction was therefore the beginning formation of a distinct stratification including a well-defined pycnocline in Cretaceous oceans (Hay 2008), as in Recent oceans. Other groups were vertical migrants in deep water, the mesopelagic zone. There, some fed on mesopelagic organisms including sluggish juvenile ammonoids, whereas others might have caught zooplankton with arms connected by velar skins (Westermann 1996). Only relatively few consumed soft-bodied epibenthos or preyed like Nautilus on shelly benthos.
The relation between changes in ammonoid assemblages (radiation of new taxa, appearance of additional morphogroups, size variations) and evolutionary events was interpreted by several authors (Elmi and Alméras 1984; Reboulet 1996, 1998, 2001; Reboulet and Atrops 1997; Cecca 1997, 1998; Lehmann 2000; Guex 2001; Reboulet et al. 2005). They considered the relation to be caused or at least influenced by changes of trophic levels (food, nutrients, energy, etc.). Reboulet et al (2005), however, noted that the assumptions on ammonoid trophic levels in the former papers were speculative. Reboulet et al. (2000, 2003) interpreted Valanginian assemblages by performing a quantitative and integrated high-resolution analysis of macrofauna, nannofossils, microfacies, trace fossils, and total organic carbon. The resulting absolute abundance variations reflected the trophic resources in the oceanic waters.
A trophic level specifies the position of an organism in the food web, often generated by the nutrient supply into that system. Accordingly, the water mass can be characterized as oligotrophic (i.e., less nutrients, less organic production, high oxygen content), mesotrophic (i.e., increasing nutrients, increasing productivity and enough oxygen), and eutrophic (i.e., high nutrient values, high organic production, decreasing oxygen). Nutrient supply creates energy, whose availability alters biodiversity (Tittensor et al. 2011). Less nutrients are available at great distances from land and in deeper waters in contrast to nearshore and shallow areas.
Abundance data on ammonoids have been collected (Reboulet et al. 2005) from the Albian of the Vocontian Basin (France) across an anoxic interval (OAE 1d, Breistroffer interval). Reboulet et al. (2005) suggested that these were influenced by trophic changes. The Breistroffer deposits do not record eutrophication of surface waters associated with expansion of the oxygen minimum zone. Rather, the deposits record changes from mesotrophic to more oligotrophic conditions in surface waters (Reboulet et al. 2000). Ammonoids are abundant and morphologically diverse (i.e., seven morpho-groups) in a section from proximal areas (platform environments) and the pelagic realm (open-marine water column). Ammonoid morphology was interpreted (Reboulet et al. 2005) to indicate aspects of their modes of life (Westermann 1996; Cecca 1997; Klug 2001), although all were attributed to the epipelagic zone. For example, Hypophylloceras (Phylloceratina) and Tetragonites (Lytoceratina) were placed by Reboulet et al. (2005) in the deepest layers of the epipelagic zone within the oceanic domain (150–200 m depth). Recent data on oxygen isotopes (Moriya et al. 2003; reinterpreted herein, Fig. 18.11) yielded wider depth ranges for both genera: 100–300 m (upper mesopelagic to middle epipelagic) in Hypophylloceras and 80–180 m in Tetragonites (lower to middle epipelagic). Tetragonites probably inhabited slightly shallower layers, as also figured by Reboulet et al. (2005).
Ammonoid taxa can be linked to changes in trophic conditions as inferred from a study on calcareous nannofossils (Reboulet et al. 2005). Heteromorphs, which were dominant during mesotrophic conditions, could have been more competitive than involute/evolute planispirals (normally coiled) when environmental conditions became more unstable. Lechites (orthocone) was assigned to an epipelagic habitat (i.e., distal paleoenvironments); it was interpreted to be a vertical migrant, able to move with increasing nutrients to meso- and eutrophic layer conditions in surface waters, and also down in order to avoid oligotrophic surface waters, preying on food-rich layers in deeper waters (Reboulet et al. 2005). More proximal paleoenvironments were suggested for Scaphites, which maybe was a vertical migrant. Quasiplanktic Turrilitoides and Mariella (torticones) mainly inhabited neritic paleoenvironments, occupying more distal paleoenvironments with mesotrophic conditions (increasing nannofossil content) in the surface water column. Anisoceras and Hamites (quasiplanktic U-shaped heteromorphs) lived in distal, epipelagic habitats. They were more competitive when oligotrophic conditions prevailed in surface waters. Mortoniceras (planispiral) was interpreted to inhabit the lower part of the epipelagic zone with a deep nektonic mode of life (Reboulet et al. 2005).
Early Cretaceous size variations (on species level) along a depth gradient are known from S France (Bulot 1993, 1995; Reboulet 1996, 2001; Reboulet and Atrops 1997). These ammonoids exhibited larger conchs in Valanginian shallow-water facies (Provence platform) and smaller ones in deepwater facies (Vocontian basin). Reboulet (2001) noted a size dependency on higher hydrostatic pressure (i.e., emptying rate of cameral fluid; Ward 1987) and cooler waters. This led to dwarfed forms in the basinal areas. Reboulet (2001) assigned the Valanginian ammoinoid taxa (macro- and microconchs; Acanthodiscus, Busnardoites, Karakaschiceras, Leopoldia, Lytoceras, Neocomites, Neolissoceras) to a deep nektonic to nektobenthic mode of life. Acanthodiscus radiatus is much more common on the shallow platform than in the basin (Reboulet 1996). It was segregated by its dimorphs: macroconchs dominate on the platform whilst microconchs are abundant in both habitats. A. radiatus shows the same size in shallow and deep-water facies, probably reflecting a migrant lifestyle between different environments. Neocomites peregrinus appears in equal numbers in both environments and is thus thought to be more tolerant of environmental changes. Reboulet (2001) proposed that the main factor in ammonoid segregation was water temperature, as in Recent Nautilus (Ward 1987) and coleoids (Mangold 1989). Thus, temperature may have triggered sexual maturity and the corresponding growth; food resources were also suggested to have influenced size differentiation. The important factor of food availability was also noted by Bucher et al. (1996). Speculating on the assumptions above, nutrient supply is higher on shallow platforms (see Cecca 1998). Accordingly, planktic activity increased and more food was available, so that ammonoid size increased in such environments (Reboulet 2001).
Trends in ammonoid diversity, habitat, and trophic levels were compared by Lukeneder and Grunert (2013) from the Trento Plateau in Italy and from the Betic Cordillera in Spain (Company et al. 2005). This yielded new insights into the paleoenvironments, which were determined by trophic fluctuations. Both data sets agree in the strongly increasing diversity of epipelagic vertical migrants and planktic drifters during the Late Hauterivian (Pseudothurmannia mortilleti and P. picteti subzones; Faraoni Level). Company et al. (2005) and Lukeneder and Grunert (2013) related this distinct turnover to phytoplankton (radiolarian) bloom during the Faraoni event and during the coeval 2nd-order peak transgression in the lower P. mortilleti Zone. The increase in trophic resources favored the diversification of planktic ammonoids. In the Puez area, similar blooms in productivity are expressed as radiolarian wackestones. This fact, however, does not explain why this habitat group remains diverse in the aftermath of the Faraoni event in the upper P. mortilleti and P. picteti subzones when oligotrophic conditions were established (Lukeneder and Grunert in 2013). These contrasting trends show the difficulties in interpreting ammonoid habitat groups and feeding strategies. Company et al. (2005) explained the extinction of deep nektic groups around the Faraoni Level by an oxygen-minimum zone that developed due to phytoplankton blooms. Trends in radiolarian and nannoconid abundances suggest a turnover in the trophic structure of the surface water from eutrophication around the Faraoni event to oligotrophic conditions. These changes in primary productivity, however, are also reflected in the organic matter with values up to 7.0% TOC. Company et al. (2005) developed a classification of ammonoids into six life-habitat groups: planktic drifters, epipelagic vertical migrants, epipelagic nektic ammonoids, mesopelagic vertical migrants, mesopelagic nektic ammonoids, and nektobenthic ammonoids. Members of all these groups occur in the studied interval (Lukeneder and Grunert 2013). Planktic drifters are represented by Crioceratites, Pseudothurmannia, Paracostidiscus, Paraspiticeras, Karsteniceras, Hamulinites, Sabaudiella and Megacrioceras; Honnoratia, Hamulina, and Anahamulina were epipelagic vertical migrants. Lytoceras and Protetragonites were possibly mesopelagic vertical migrants, and the mesopelagic nektic ammonites include Phylloceras, Phyllopachyceras, Kotetishvilia, and Neolissoceras. Barremites, Plesiospitidiscus, and Discoidellia were epipelagic nektic ammonoids. Finally, Abrytusites and Astieridiscus are nektobenthic forms.
The analysis of ammonoid diversity and life-habitat groups (Westermann 1996; Company et al. 2005) suggests a strong influence of sea-level on the plateau. While epi- and mesopelagic ammonoids occur commonly in the Crioceratites krenkeli, P. mortilleti, and P. picteti subzones, a severe reduction in all groups occurred during the sea-level lowstand of the Spathicrioceras seitzi and Pseudothurmannia ohmi subzones. The Faraoni event is heralded by an ammonoid faunal turnover characterized by an increasing diversity of epipelagic ammonoids. Although a similar trend was observed in the Betic Cordillera in Spain, the trend differs in the common occurrence of deep nektic ammonoids, related to a better oxygenated environment on the Trento Plateau.
12 Implications for Ecology from Traces of Predation
Shell damage is an indicator for predator-prey interactions (Kowalewski 2002) involving predators within distinct and characteristic ammonoid habitats. Ammonoids are both predators and prey, foraging and being attacked in the same habitat. Mapes and Chaffin (2003) summarized the sublethal and lethal shell damage from recent and fossil cephalopods (Keupp and Hoffmann 2015). Predation in the water column can involve ichthyosaurs (Fig. 18.13), mosasaurs, nothosaurs, sharks or bony fishes, and invertebrates including other ammonoids, nautiloids, coleoids (Landman and Waage 1986; Kröger 2000, 2002a, b, c; Keupp 2000, 2006; Larson 2003, 2007; Tichy and Urbanek 2004; Andrew et al. 2010; Landman et al. 2012; Kauffman and Sawdo 2013) and arthropods (Keupp and Hoffmann 2015). As noted by Klompmaker et al. (2009) for Mesozoic and Andrew et al. (2010) for Jurassic ventral bite marks, they are most probably caused by predatory attacks on living animals by teuthoids (Yomogida and Wani 2013). These authors argued that the combination of an intact aperture and the absence of shell chips (i.e., broken parts of corresponding injury) excludes scavengers as producers of such injuries.

Reconstruction of a Lower Jurassic ichthyosaur hunting on ammonoids, biting into the shell of Dactylioceras in the epipelagic water zone of the Liassic sea (based on studies of Posidonia shales in SW Germany). Upper Cretaceous mosasaur bite marks (max. puncture diameter 3 cm; lower right corner) on a Campanian Placenticeras shell (cast replica, max. diameter 30 cm) from the Western Interior Seaway in South Dakota
Recently, remarkable bite marks on both flanks (i.e., by both jaws of a predatory fish) were reported on Jurassic Oxycerites (Richter 2009). Some ammonoids are thought to be demersal feeders, scavenging on the sea floor (Reyment 1988): there, they were attacked by crustaceans (Radwansky 1996; Kröger 2002b, c; Keupp 2006), causing sublethal to lethal injuries (Keupp and Hoffmann 2015). Keupp (2000) also attributed the apertural injuries in Dacytlioceras to crustaceans on the sea floor (Keupp 1997), assuming a demersal habitat. This significantly contrasts with Westermann (1996), who proposed that Dactylioceras was a planktic form. The same former result was presented for the entire Callovian assemblage (perisphinctids, kosmoceratids) from northern France (Keupp 1992): all ammonoids were assumed to be nektobenthic to benthic (habitat dependency in accordance to frequency of injuries). That author used sublethal shell injuries of ammonoid shells, caused by crustaceans to draw conclusions on paleopathology . Keupp and Schobert (2011) even suggested a special crab (Palaeopagurus) as the predator on Liassic ammonoids (Amaltheus, Amauroceras) from Germany, which were assumed to have been demersal forms inhabiting near-bottom areas. Palaeopagurus is as the hermit crab also known to inhabit the body chambers of Jurassic and Cretaceous ammonoid shells on the sea floor (Germany and UK; Fraaije 2003; Jagt et al 2006).
As noted by Keupp (2000), shell damage on the rocky sea floor caused by the animal itself can be ruled out in most injuries (Bayer 1970; Checa and Westermann 1989; Zatoń 2010). A predator-prey dependency (Vermeij 1977; Keupp 2000a, 2006) puts pressure on the prey and probably causes it to evolve new strategies such as coarser ribbing, a more complex suture or increasing coiling (i.e., from straight to involute). Nonetheless, ecological pressure can also cause strategic changes, as seen in Recent Nautilus: it migrates at night to prey on various resources, and may then prey when its main enemies such as many fishes sleep (Carlson et al. 1984; Saunders 1984; Ward et al. 1984; Dunstan et al. 2011).
Shell damage by marine reptiles was documented from Middle Jurassic Kosmoceras (Ward and Hollingworth 1990), Late Cretaceous Placenticeras and Sphenodiscus (Kauffman and Kiesling 1960; Kauffman 1990; Hewitt and Westermann 1990b; Stewart and Carpenter 1990, 1999), and the nautiloid Eutrephoceras (Kauffman and Sawdo 2013). Kauffman and Sawdo (2013) argued that reports of mosasaur predation on Early Maastrichtian nautiloids from the Western Interior are rare because of the prey’s predominantly deep, epibenthic habitat. They deduced this from modern Nautilus and applied it to the Cretaceous Eutrephoceras. Home scars of patellogastropods (i.e., round marks in a line) located on ammonoid shells have been used to falsify the mosasaur bite theory by Kase et al. (1994; 1998). In several cases, however, there appears to be no doubt about the mosasaur predation (Keupp 1991; Hewitt and Westermann 1990b; Fig. 18.13). Interestingly, patellogastropods, when attached to ammonoids (after their death) show a drifting history: limpets live in shallow areas and must have attached at the sea surface. Plesiosaurs were reported to prey on ammonoids by Sato and Tanabe (1998; see also Keupp 2000) from the Cretaceous of Japan (stomach content). Fish predators have been inferred (Ward 1981) and documented in Late Jurassic Haploceratidae by aptychi in holostean feces (Mehl 1978a). Jurassic ammonoids from the shallow sea in Solnhofen were preyed upon by teleosts such as Gyrodus or Dapedium (Keupp 2000). They broke parts off the aperture to expose the soft body, as observed in Recent Nautilus preyed upon by parrotfishes (Saunders et al. 1987), triggerfishes, and groupers (Mapes and Chaffin 2003). Injuries inflicted by fishes can be sublethal or lethal. Sublethal injuries can also reflect octopod attacks (e.g., borings in the shell; Mapes and Chaffin 2003). A more peculiar ‘attack’ is observed in extant Nautilus during mating (Arnold 1985). In this case, the partner bites and damages the apertural edge, which is also visible in fossil conchs. The mechanisms that damage extant Nautilus (sediment loading, within-sediment transport, collision during floating, predation) were analyzed by Wani (2004). He concluded that these mechanisms produce distinct and characteristic shell damage, hence being comparable to biostratinomic and taphonomic features in fossil materials.
Stomach contents of plesiosaurs from the Late Cretaceous of Japan represent important evidence for lethal injuries (Massare 1987; Sato and Tanabe 1998). They contain pieces of broken shell material from Scaphites and other ammonoids.
Keupp (2000) noted healed injuries caused by fish attacks on Kranaosphinctes and by crustaceans on Prososphinctes (Oxfordian, Madagascar). Teuthoid predators were suspected to have produced clusters of uniformly sized shell fragments of Early Jurassic harpoceratids and Late Jurassic Gravesia (Mehl 1978b). They probably also caused most of the peristomal mantle injuries in Jurassic to Cretaceous ammonoids. Unless these were lethal, they caused the well-known shell abnormalities. Evidence for predation (predator not specified) of Early Cretaceous ammonoids is also given by Reboulet and Rard (2008). Klug (2007) reported sublethal injuries on Devonian ammonoids that lived in the water column; that damage was likely caused by other cephalopods or phyllocarids. Shark attacks were reported as a possible cause for goniatid injuries in Carboniferous ammonoids (USA; Mapes and Hansen 1984; Mapes et al. 1995).
A remarkable ammonoid occurrence was documented by Mapes and Dalton (2002) from the Mississippian of Arkansas (USA). Ammonoid accumulations (e.g., Emstites, Cluthoceras) in carbonate concretions form halos around body chambers of Rayonnoceras (Actinocerida) and were interpreted as prey of nautiloid predators (based on stomach contents); scavenging was excluded by Mapes and Dalton (2002). Those assumptions were mainly based on missing body chambers, the random orientation of ammonoids, and the rarity of ammonitellae in such concretions. My own interpretation of that material is that ammonoids were redeposited into the broken phragmocone and body chambers of Rayonnoceras (fossil trap).
As noted by several authors, increased coiling and sculpture might have enhanced shell resistance towards breakage during predation attempts (Ward 1981; Klug 2007). Tight coiling implies an increased resistance against breakage of entire whorls when compared to loosely coiled shells (Nützel and Frýda 2003 and references therein). Consequently, a simple change in morphology had a major impact on ecological fitness, partially explaining the evolutionary success of ammonoids (Kröger 2005). Coiling probably enabled even early forms to increase their maximum swimming velocity compared to cephalopods with orthoconic shells (e.g., Jacobs 1992a; Jacobs and Chamberlain 1996; Westermann 1996; Korn and Klug 2002; Klug and Korn 2004). Predation and damage in relation to coiling in Carboniferous nautiloids and ammonoids was treated by Mapes and Chaffin (2003): predation levels were much higher in the latter. Swimming velocity was assumed to be higher in nearshore morphotypes (compressed) of Scaphites (Cretaceous, USA). Higher velocities are assumed to be essential for life in shallower areas (Jacobs et a. 1994), contrasting to the decreased velocity values of the depressed morphs of deeper areas (offshore). The more compressed morphotypes can better escape potential predators.
13 Conclusions
Ammonoid migrations within and between different habitats are ontogenetically induced and detectable by stable isotope data (δ13C, δ18O) and morphology. Ammonoids generally start with a perhaps planktic embryonic stage (floating egg masses), followed by the hatchlings, which differentiated in mode of life; they lived either planktic, nektic, or nektobenthic (demersal). Antidimorphic pairs (female/male, macroconch/microconch) were apparently separated into different habitats, at least during spawning periods. Females are thought to have laid 100–500000 eggs (r-strategy) on the sea floor or in the water column; ammonoid eggs and hatchlings were thus an important constituent of the plankton. Eggs were perhaps enclosed in egg sacks, anchored (in pouches) below the storm wave base or floated as egg masses in the upper water column. Hatchlings had a nektoplanktic lifestyle in the water column.
Several taxonomic groups were probably gregarious, some migrated over long distances and across depth levels. This conclusion has been strengthened by stable isotope analyses. Female ammonoids appear to have returned to their home-grounds for spawning and died afterwards (i.e., semelparous). This resembles the situation in extant neritic coleoids. Planktic egg masses and hatchlings/young juveniles floated for months within the water column, driven by ocean currents (surface or bottom-water currents). During the nektoplanktic juvenile phase and subsequent stages, they fed on various nutrients including nanno- and micro-plankton members as trophic resources. Taxonomic groups can therefore appear or vanish due to changes in trophic levels. Ammonoids inhabited most known parts of ancient seas (lack of knowledge from paleo-oceans), from euryhaline to brackish waters, and probably tolerated brief hypoxic events like extant cephalopods such as Nautilus and Spirula.
The best-estimate for habitat depths of ammonoids is between c. 20–500 m according to the literature and recent stable isotope data (δ18O). Maximal depth (implosion depth) estimated from siphuncle and shell wall strengths are now recognized to be reliable for application to ammonoid paleobathymetry. Nonetheless, some literature values are apparently too deep and the ranges inaccurate.
Bathymetric conclusions were drawn from δ18O values for several ammonoid morphogroups. The range in Cadoceras (Jurassic, Callovian) is 50–700 m. It therefore started in the lower epipelagic and migrated deeper during ontogeny into the upper mesopelagic. With a depth range of 10–60 m, the ontogeny of Hypacanthoplites (Cretaceous, Aptian) starts in deeper epipelagic layers, followed by shallower epipelagic stages. Nowakites (Cretaceous, Santonian) is interpreted to have inhabited depths from 50–300 m. This implies migrations from deeper mesopelagic layers to shallower epipelagic habitats. The depth range given in the literature for Baculites (Cretaceous, Campanian) was specified as being relatively shallow (50–100 m) in the epipelagic zone. Perisphinctes (Jurassic, Oxfordian) apparently inhabited almost the same habitat in the middle epipelagic zone (50–100 m), although they migrated to deeper areas as young juveniles and inhabit again cooler waters as adults.
Two main groups with different ontogenetic strategies based on the δ18O data were established by Lukeneder et al. (2010). The first is the wcw–type (warm–cool–warm type) of Cadoceras. Its lif strategy resembles those of Nautilus and Sepia , which migrate from shallow into deeper environments and back in ontogeny. The second is the cw–type (cool–warm type) of Hypacanthoplites. It resembling the first two migration phases of the cwc–type of Spirula (cool–warm-cool type), which migrates from deeper into shallower waters and back again.
δ18O values as proxies for paleotemperature and paleohabitat-depth estimates show that Campanian taxa from Japan of all morphogroups and ammonoid families (Tetragonites, Damesites, Hauericeras, Polyptychoceras) inhabited shallower waters than indicated earlier. This contrasts with the opinion of Moriya et al. (2003), who assumes a bottom-dwelling demersal lifestyle for all Late Cretaceous ammonoids they measured. They therefore suggested that the ammonoids from Japan undertook no long- or short-term vertical migrations. They measured only the body chambers of the specimens. This hinders any conclusions on life-history trends. Applying the scheme of Lukeneder et al. (2010) on ammonoid and extant cephalopod habitats, water depth and temperature yielded a somewhat divergent picture: dispersal during ontogeny is evident. The range for all included taxa from the Cretaceous ammonoids of Hokkaido would then be 70–350 m, i.e. exclusively lower to middle epipelagic (the middle epipelagic zone dominates). The exception is the Phylloceratina (Hypophylloceras). They were restricted from the upper mesopelagic to middle epipelagic, a range from 100–300 m; this contrasts with the much deeper values given in the literature (up to 600 m). This means that the difference in depth and habitat between Phylloceratina, Lytoceratina, Ammonitina and Ancyloceratina are not as large as previously thought. Nonetheless, a shallowing gradient is evident from smooth to fine-ribbed types of Phylloceratina (Hypophylloceras, Phyllopachyceras) and Lytoceratina (Tetragonites, Gaudryceras), over stronger ribbed types of the Ammonitina (Eupachydiscus, Yokoyamaoceras) to heteromorph Ancylocetratina (Polyptychoceras). At least Polyptychoceras (i.e., Pp1 to Pp2) appears to have migrated during ontogeny from the lower epipelagic at 200 m to shallower epipelagic zones at 60 m.
The conclusion in Moriya et al. (2003) that all species of the taxonomically diverse assemblage investigated were entirely demersal throughout their lives was challenged by δ18O data presented by Henderson and Price (2012) for Late Cretaceous ammonoids (Acanthoceras, Euomphaloceras) from Australia. The highly negative δ18O values were interpreted as indicating the growth of strongly ornamented morphogroups in near-surface waters. This conflicts with the interpretation of a deep marine nektobenthic life habit and with data from Late Cretaceous (Campanian) planispiral taxa recorded by Moriya et al. (2003).
Stable isotope data from Cretaceous (Albian) ammonoids of Madagascar (Eotetragonites, Cleoniceras, Desmoceras, Douvilleiceras) were given by Zakharov et al. (2011). Concerning the data as ontogenetic rows, the interpretation regarding the paleotemperature and habitat of the Albian Madagascan sea (tropical–subtropical zone) reflects habitats from upper mesopelagic to lower epipelagic for these ammonoids, with a general depth range from 70–400 m. The Lytoceratina with Eotetragonites appear to be the only inhabitants of the upper mesopelagic layers at 200–400 m. Contrastingly, Westermann (1996) suggested a shallower epipelagic range from 100–180 m based on siphuncle index estimations. The Albian members of the Ammonitina lived within the lower epipelagic zone. There are two finely ribbed forms: Cleoniceras shows a depth range from 100–200 m (max. depth 250 m in Westermann 1996) with a constant ontogenetic trend, whereas Desmoceras ranges from 70–250 m (min. depth 180 m in Westermann 1996). The ontogenetic trend in Desmoceras shows a major habitat change, i.e., a deepening in adult stages. Strongly ribbed Ancyloceratina like Douvilleiceras are relatively shallow-water inhabitants of the middle epipelagic at depths of 70–90 m, slightly increasing in ontogeny. The Albian nautiloid Camytoceras inhabited a depth of c. 180–200 m (not ontogenetically measured) in the range of extant Nautilus.
Based on δ18O values for ammonoids, the rule of thumb that ‘Trachyostraca’ dominated above 200 m depth in the neritic epipelagic zone, whereas ‘Leiostraca’ dominated in the lowermost epipelagic and deeper mesopelagic zones, appears to be valid. There were, however, exceptions. In general, ammonoids were mostly adapted to neritic areas associated with the continental shelf. Crucial for future work on stable isotope trends in mobile and migrating ammonoids is an understanding of ocean stratification during the Jurassic and Cretaceous (Hay 2008). Knowledge about the presence or absence of a pycnocline and hence the thermal structure of the ocean bodies will improve our understanding of δ18O data from cephalopod shells in the Mesozoic.
A major difference in the modes of life of many ammonoids compared to the extant cephalopods Nautilus, Sepia, and Spirula occured in the latest adult stage. All extant examples tend to retreat into the deepest environments as mature adult animals (Spirula, Sepia) or remain there throughout their post-juvenile phase (Nautilus). In contrast, all measured ammonoids except for Perisphinctes display a clear tendency to migrate into shallower environments in their latest ontogenetic stage (mature, fully grown; Lukeneder et al. 2010). Sexually dimorphic ammonoids might have reached maturity at different ages, earlier in males and later in females. Additional stable isotope analyses of antidimorphic pairs will help to test this hypothesis.
Caution should be exercised when estimating or assuming an ammonoid–facies dependency. Ammonoids were probably dependent on a specific environment, reflected today in the lithology/facies of the host rock. Ammonoids were probably capable of migrating between different habitats, or at least of escaping from hostile environmental changes (e.g., oxygen depletion, anoxia), giving rise to new populations. Such new faunal constituents are marked by distinct morphological adaptations to shallow or deeper environments, predominantly triggered by water energy and hydrostatic changes, and less bound to facies. Highly morphologically variable species challenge the paradigms that explicitly require a dependency of shell morphology and a special mode of life. Single species exhibiting numerous morphotypes, for example from oxycone to serpenticone detected in a single rock-layer were also able to inhabit the same habitat (pelagic areas) without any separation into morphogroups reflecting special facies dependence. An expansion of shallow-water areas can, however, influence morphological evolution of ammonoid shells, mainly during sea level transgressions onto shelves.
The main prey for ammonoids was most likely zooplankton from different water layers, from mid-water to benthic areas. Ostracods, foraminifera, tiny ammonoids (e.g., mandibles of juveniles hint at cannibalism ; Summesberger 2000) and planktic crinoids have been detected in stomach/crop content (Jäger and Fraaye 1997; Keupp 2007; see also Milson 1994), indicating that they were important food sources. The feeding strategies of ammonoids suggest that they probably ate everything they could reach, foraging as demersal herbivores, scavengers or even as active nektic carnivores.
The implications from ammonoid shell damage caused by predators are twofold. Predation can take place directly in the water column by mosasaurs, sharks, bony fishes and invertebrates (other cephalopods) or near the sea floor by, e.g., crustaceans, causing sublethal or lethal injuries. Abundant injuries caused by crustaceans hint at a (at least temporal) demersal life or benthic resting times. This recognition significantly changed the opinions presented in the past. A more demersal, nektobenthic to benthic life (indicated by the frequency of injuries) was suggested by some authors. Some ammonoids speculatively undertook vertical diurnal migrations, crossing different oceanic strata of the epipelagic to upper mesopelagic zones. This is characteristic for many planktic ocean dwellers and for nektobenthic genera such as Nautilus and Spirula.
Different ammonoid groups show major differences (size, morphology) in embryonic stages, not assessing where the eggs were stored (i.e., water column or benthic areas). The spawning strategy in ammonoids (r-strategy, 100–500 000 eggs per mother animal) contrasts with that of extant nautiloids (K-strategy, < 10 eggs or capsulae). The dispersal mechanisms of the planktic hatchlings and subsequent neanic ammonoids were entirely passive and depended on currents. Dispersal distance ranged from tens of kilometers in enclosed epeiric seas, over a few hundred kilometers on shelf seas, to up to 1000–2000 km in the open ocean. Planktic stages drifted passively, making up a large part of the Mesozoic plankton in shelf seas and adjoining oceans.
The postmortem history of ammonoids can change our picture of an assemblage in diverse ways. Shells that reach the sea surface after death can drift over long distances or long times. Postmortem drifting is enhanced when predators or scavengers remove the soft body from the shell, increasing buoyancy. This obscures the primary habitat and depth of death. In contrast, those shells that became negatively buoyant at a certain depth and sank to sea floor reveal a more reliable nearly autochthonous picture of the primarily deeper habitat. The path of a shell after the animal’s death is controlled by several environmental factors such as depth at death, the initial buoyancy of the empty shell, the morphological and physiological parameters of the shell, and the rate of sea water influx into the phragmocone. Sinking history can also be detected by analyzing the encrusting community and its position. Buccal masses found within body chambers provide a useful criterion for evaluating postmortem transport. In situ buccal masses (upper and lower jaws) are suggested to reflect an in situ deposition of the ammonoid.
New stable isotope data (δ13C, δ18O) extracted from original shell materials (aragonite) revealed different ammonoid lifestyles (e.g., wcw–type, cw–type, wc-type), ontogenies, and varying habits in different habitats. Additional work on these isotopes and on the composition of ammonoid shells will enhance our knowledge on lifestyles and habitats. Exact ontogenetic measurements of δ13C and δ18O from neanoconchs to adult stages, from ammonitellae to terminal apertures, are crucial in future investigations on ammonoids.
References
Abel O (1916) Paläobiologie der Cephalopoden. Aus der Gruppe der Dibranchiaten. Verlag Gustav Fischer, Jena
Alberti M, Fürsich FT, Pandey DK, Ramkumar M (2011) Stable isotope analyses of belemnites from the Kachchh Basin, western India: paleoclimatic implications for the Middle to Late Jurassic transition. Facies 58:261–278
Anderson TF, Popp BN, Williams AC, Ho LZ, Hudson JD (1994) The stable isotopic record of fossils from the Peterborough Member, Oxford Clay Formation (Jurassic), UK: Palaeoenvironmental implications. J Geol Soc Lond 151:125–138
Andrew C, Hoew P, Paul CRC, Donovam SK (2010) Fatally bitten ammonoids from the lower Lias Group (Lower Jurassic) of Lyme Regis, Dorset. Proc York Geol Soc 58:81–94
Andrew C, Hoew P, Paul CRC, Donovam SK (2011) Epifaunal worm tubes on Lower Jurassic (Lower Lias) ammonites from Dorset. Proc Geol Assoc 34–46
Arkell W (1957) Introduction to Mesozoic Ammonoidea. In: Moore RC (ed) Treatise on Invertebrate Paleontology Part L Mollusca 4 Cephalopoda Ammonoidea. The University of Kansas Press, New York, pp. L81–L129
Arnold JM (1985) Shell growth, trauma and repair as an indicator of life history for Nautilus. Veliger 27:386–396
Assereto R (1969) Sul significato stratigrafico della ‘Zona ad Avisianus’ del Trias medio delle Alpi. Boll Soc Geol Italia 88:123–145
Auclair A-C, Lecuyer C, Bucher H, Sheppard SMF (2004) Carbon and oxygen isotope composition of Nautilus macromphalus: a record of thermocline waters off New Caledonia. Chem Geol 207:91–100
Ballentine CM (2007) A mathematical analyses of some indices used to classify ammonite shells. Lethaia 40:197–198
Bandel K (1988) Operculum and buccal mass of ammonites. In: Wiedmann J, Kullmann J (eds) Cephalopods–present and past. Schweizerbart, Stuttgart
Bandel K, Hoefs J (1975) Die Isotopenzusammensetzung von Molluskenschalen am Beispiel der Gastropoden. Neues Jahrb Geol Paläontol Monatshefte 1975:1–11
Bandel K, Landman NH, Waage KM (1982) Micro-ornament on early whorls of Mesozoic ammonites: implications for early ontogeny. J Paleontol 56:386–391
Batt RJ (1987) Pelagic biofacies of the Western Interior Greenhorn Sea (Cretaceous). Evidence from ammonites and planktonic foraminiferas [Ph.D. Thesis]: University of Colorado, p 778
Batt RJ (1989) Ammonite shell morphotype distribution in the Western Interior Greenhorn sea and some paleoecological implication. Palaios 4:32–42
Batt RJ (1991) Sutural amplitude of ammonite shells as a paleoenvironmental indicator. Lethaia 24:219–225
Batt RJ (1993) Ammonite shell morphotypes as indicators of oxygenation in ancient epicontinental seas: example from Late Cretaceous Greenhorn Cyclothem (U.S.A.). Lethaia 26:49–64
Bayer U (1970) Anomalies in Aalenian and Bajocian ammonites as clues to their mode of life. Neues Jahrb Geol Paläontol Abh 135:19–41
Bayer U (1977) Cephalopod septa I. Constructional morphology of the ammonite septum. Neues Jahrb Geol Paläontol Abh 154:290–366
Bayer U (1982) Ammonite maneuverability-a new look at the functions of shell geometry. Neues Jahrb Geol Paläontol Abh 164:154–156
Bayer U, McGhee GR Jr (1984) Iterative evolution of Middle Jurassic ammonite faunas. Lethaia 17:1–16
Becker RT (1993) Anoxia, eustatic changes, and Upper Devonian to lowermost Carboniferous global ammonoid diversity. In: House MR (ed) The Ammonoidea: Environment, Ecology, and Evolutionary Change. The Systematics Association, special vol 47. Clarendon Press, Oxford
Bettencourt V, Guerra A (1999) Carbon- and oxygen-isotope composition of the cuttlebone of Sepia officinalis: a tool for predicting ecological information? Marine Biol 133:651–657
Bettencourt V, Guerra A (2001) Age studies based on daily growth increments in statoliths and growth lamellae in cuttlebone of cultured Sepia officinalis. Marine Biol 139:327–334
Birkelund T (1981) Ammonoid shell structure. In: House MR, Senior J (eds) The Ammonoidea. Systematics Association, special vol 18. Academic Press, London
Boletzky S v (1974) The “larvae” of Cephalopoda: a review. Thalassia Jugosl 10:45–76
Boletzky SV (1987) Juvenile behaviour. In: Boyle PR (ed) Cephalopod life cycles, vol II. Academic Press, London
Boletzky SV (1992) Evolutionary aspects of development, life style, and reproductive mode in incirrate octopods (Molluscs, Cephalopoda). Rev Suisse Zool 99:755–770
Bond PN, Saunders WB (1989) Sublethal shell repair in Upper Mississippian ammonoids. Paleobiol 15:414–428
Böse E (1928) Cretaceous ammonites from Texas and northern Mexico. Bull Univ Tex 2748:143–312
Boston WB, Mapes RH (1991) Ectocochleate cephalopod taphonomy. In: Donovan SK (ed) Processes of fossilization. Bellhaven Press, London
Boucot AJ (1981) Principles of benthic marine paleoecology. Academic Press, New York
Boyajian G, Lutz T (1992) Evolution of biological complexity and its relation to taxonomic longevity in the ammonoidea. Geology 20:983–986
Brack P, Rieber H (1993) Towards a better definition of the Anisian/Ladinian boundary: new biostratigraphic data and correlations of boundary sections from the Southern Alps. Eclogae Geol Helvetiae 86:415–527
Brack P, Rieber H (1996) The ‘High resolution Middle Triassic ammonoid standard scale’ proposed by Triassic researchers from Padova–a discussion of the Anisian/Ladinian boundary interval. Albertiana 17:42–50
Brand U (1986) Paleoenvironmental analysis of Middle Jurassic (Callovian) ammonoids from Poland: trace elements and stable isotopes. J Paleontol 60:293–301
Brett CE, Miller KB, Baird GC (1990) A temporal hierarchy of paleoecologic processes within a Middle Devonian epeiric sea. In: Miller W (ed) Paleocommunity Temporal Dynamics: The Longterm Development of Multispecles Assemblages. Paleontol Soc Spec Publ 5:178–203
Brett CE, Dick VB, Baird GC (1991) Comparative taphonomy and paleoecology of Middle Devonian dark gray and black shale facies from western New York. State Mus Bull 469:5–36
Brumsack HJ (1991) Inorganic geochemistry of the German ‘Posidonia Shale’: Palaeoenvironmental consequences. In: Tyson RV, Pearson TH (eds) Modern and ancient continental shelf anoxia. Geol Soc Spec Publ 58:353–362
Bruna GD, Martire L (1985) La successione giurassica (Pliensbachiano-Kimmeridgiano) delle Alpi Feltrine (Belluno). Riv Ital Paleontol Stratigr 91:15–62
Bucher H, Landman NH, Klofak SM, Guex J (1996) Mode and rate of growth in ammonoids. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid Paleobiology. Topics in Geobiology 13, Plenum, New York
Bulot L (1989) Les Olcostephaninae (Ammonitina, Cephalopoda) dans le Crétacé inférieur du Jura Suisse et Francais. In: Museé d’Histoire Naturelle de Genève Résumés (ed) Réunion Commune APF-SPS, 21–22 Octobre 1989, Geneva, p 5
Bulot L (1992) Les Olcostephaninae Valanginiens et Hauteriviens (Ammonitina, Cephalopoda) du Jura Franco-Suisse: systematique et interet biostratigraphique. Rev Paléobiol 11:149–166
Bulot L (1993) Stratigraphic implications of the relationship between ammonites and facies: examples taken from the Lower Cretaceous (Valanginian-Hauterivian) of the western Tethys. In: House MR (ed) The Ammonoidea: Environment, Ecology, and Evolutionary Change, Systematics Association, special vol 47. Clarendon Press, Oxford
Bulot L (1995) Les formations à ammonites du Crétacé inférieur dans le SE de la France (Berriasien-Hauterivien): Biostratigraphie, paléontologie et cycles sédimentaires [PhD thesis]: Paris, University Paris, p. 374
Bulot L, Company M (1990) Les Olcostephanus du groupe atherstoni (Ammonitina, Cephalopoda): potential d’utilisation pour les correlations biostratigraphiques à longue distance. In: VI J Soc Esp Paleontol rés, p. 34
Burnaby TP (1966) Allometric growth of ammonoid shells: a generalization of the logarithmic spiral. Lett Nat 209:904–906
Callomon JH (1980) Dimorphism in ammonoids. In: House MR, Senior JR (eds) The Ammonoidea, Systematics Association, special vol 18. Academic Press, London
Callomon JH (1985) The evolution of the Jurassic ammonite family Cardioceratidae. Spec Pap Palaeontol 33:49–90
Carlson BA, McKibben, JN, deGruy, MV (1984) Telemetric investigation of vertical migration of Nautilus belauensis. Pac Sci 38:183–188
Casadío S, Concheyro A (1992) Facies y ambientes de sedimentación en el límite Cretácico-Terciario de La Pampa, Argentina. Act VIII Cong Latin Geol Salamanca 4:30–34
Cecca F (1988) Ammonites méditerranéennes du Tithonique inférieur de l'Ardèche (Sud-Est de la France): analyse des afflux téthysiens. Geobios 21:169–186
Cecca F (1992) Ammonite habitats in the Early Tithonian of Western Tethys. Lethaia 25:257–267
Cecca F (1997) Late Jurassic and Early Cretaceous uncoiled ammonites: trophism-related evolutionary processes. Comptes Rendus Acad Sci Paris, Ser II 325:629–634
Cecca F (1998) Hypothesis about the role of the trophism in the evolution of the uncoiled ammonites: the adaptive radiations of the Ancyloceratina (Ammonoidea) at the end of the Jurassic and in the Lower Cretaceous. Rend Fis Acc Lincei 9:213–226
Cecca F, Cresta S, Pallini G, Santantonio M (1990) Il Giurassico di Monte Nerone (Appennino marchigiano, Italia Centrale): biostratigrafia, litostratigrafia ed evoluzione paleogeografica. In: Pallini G, Cecca F, Cresta S, Santantonio M (eds) Fossili, Evoluzione e Ambiente. Atti de Secondo Convegno Internazionale, Pergola ottobre 1987, Ed Comitato Centenario R Piccinini, Ancona
Cecca F, Fozy I, Wierzbowski A (1993) Ammonites et paléoécologie: étude quantitative d’associations du Tithonien inférieur de la Tethys occidentale: In: Elmi S, Mangold C, Alméras Y (eds) 3ème Symposium International, Céphalopodes Actuels et Fossiles (Lyon, 1990), Geobios Mém Spéc 15:39–48
Ceriola L, Milone N (2007) Cephalopods age determination by statolith reading: a technical manual. Adria Med Technical Documents 22, Rome, 78 p
Challier L, Dunn M R, Robin J-P (2005) Trends in age-at-recruitment and juvenile growth of cuttlefish, Sepia officinalis, from the English Channel. ICES J Mar Sci 62:1671–1682
Chamberlain JA (1976) Flow patterns and drag coefficients of cephalopod shells. Paleobiology 19:539–563
Chamberlain JA Jr (1980) Hydrodynamical design of fossil cephalopods. In: House MR, Senior JR (eds) The Ammonoidea: the evolution, classification, mode of life and geological usefulness of a major fossil group. The Systematics Association, special, vol 18. Academic Press, London
Chamberlain JA Jr (1991) Cephalopod locomotor design and evolution: the constraints of jet propulsion. In: Rayner JMV, Wooton RJ (eds) Biomechanics and evolution. Cambridge University Press, Cambridge
Chamberlain JA Jr, Moore WA Jr (1982) Rupture strength and flow rate of Nautilus siphuncular tube. Paleobiology 8:408–5452
Chamberlain JA Jr, Weaver JS (1978) Equations of motion for post-mortem sinking of cephalopod shells. Math Geol 10:673–689
Chamberlain JA Jr, Ward PD, Weaver JS (1981) Postmortem ascent of Nautilus shells: implications for cephalopod paleogeography. Paleobiology 7:494–509
Checa AG (2003) Fabrication and function of ammonite septa—comment on Lewy. J Paleontol 77:790–791
Checa AG, Garcia-Ruiz JM (1996) Morphogenesis of the septum in ammonoids. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid Paleobiology. vol 13 of Topics in Geobiology, Plenum, New York
Checa A, Westermann GEG (1989) Segmental growth in planulate ammonites: inferences on costae function. Lethaia 22:95–100
Cherel Y, Hobson KA (2005) Stable isotopes, beaks and predators: a new tool to study the trophic ecology of cephalopods, including giant and colossal squids. Proc Biol Soc 272:1601–1607
Cherel Y, Ridoux V, Spitz J, Richard P (2009) Stable isotopes document the trophic structure of a deep-sea cephalopod assemblage including giant octopod and giant squid. Biol Lett 5:364–367
Chikaraishi Y, Ogawa NO, Kashiyama Y, Takano Y, Suga H, Tomitani A, Miyashita H, Kitazato H, Ohkouchi N (2009) Determination of aquatic food-web structure based on compound-specific nitrogen isotopic composition of amino acids: Liminology and Oceanography. Methods 7:740–750
Chirat R (2000) The so-called “cosmopolitan palaeobiogeographic distribution” of Tertiary Nautilida of the genus Aturia Bronn 1838: the result of post-mortem transport by oceanic palaeocurrents. Palaeogeogr Palaeoclimatol Palaeoecol 157:59–77
Chirat R (2001) Anomalies of embryonic shell growth in post-Triassic Nautilida. Paleobiology 27:485–499
Christ HA (1960) Beiträge zur Stratigraphie und Paläontologie des Malm von Westsizilien. Mém Soc Paléont Suisse 77:1–138
Cichowolski M, Pazos PJ, Tunik MA, Aguirre-Urreta MB (2011) An exceptional storm accumulation of nautilids, Lower Cretaceous, Neuquén Basin, Argentina. Lethaia 45:121–138
Clari PA, Marini P, Pastorini M, Pavia G (1984) Il Rosso Ammonitico Inferiore (Baiociano-Calloviano) nei Monti Lessini Settentrionali (Verona). Riv Ital Paleontol Stratigr 90:15–86
Clarke MR (1969) Cephalopoda collected on the SOND cruise. J Mar Biol Assoc UK 49:961–976
Cochran JK, Rye DM, Landman NH (1981) Growth rate and habitat of Nautilus pompilius inferred from radioactive and stable isotope studies. Paleobiology 7:469–480
Cochran JK, Landman NH, Turekian KK, Michard A, Schrag DP (2003) Paleoceanography of the Late Cretaceous (Maastrichtian) Western Interior Seaway of North America: evidence from Sr and O isotopes. Palaeogeogr Palaeoclimatol Palaeoecol 191:45–64
Cochran JK, Kallenberg K, Landman NH, Harries PJ, Weinreb D, Turekan KK, Beck AJ, Cobban WA (2010a) Effect of diagensis on the Sr, O and C isotopic composition of Late Cretaceous mollusks from the Western Interior of North America. Am J Sci 310:69–88
Cochran JK, Landman NH, Harries PJ, Larson NL, Garb MP, Klofak SM, Myers C, Brezina J (2010b) Stable isotopes in well-preserved shells of a methane seep fauna from the Upper Cretaceous (Campanian) Pierre Shale, U.S. Western Interior. 8th International Symposium, Cephalopods—Present and Past, 30 August–3 September, 2010 Dijon, Abstract Volume, 119
Collignon M (1963) Atlas des fossils caractéristiques de Madagascar (Ammonites), Albien. Serv Géol Tananarive 10:184
Collignon M (1964) Atlas des fossils caractéristiques de Madagascar (Ammonites), Cenomanien. Serv Géol Tananarive 11:152
Collins D, Ward PD (2010) Adolescent growth and maturity in Nautilus. In: Saunders WB, Landman NH (eds) Nautilus. the biology and paleobiology of a living fossil. Topics in Geobiology, 6, Springer Press, New York (reprinted from Saunders WB, Landman NH (eds) 1987 with additions) pp. 421–432
Company M (1987) Los Ammonites del Valanginiense del sector oriental de las Cordilleras Béticas (SE de Espana) [PhD thesis]: Universidad de Granada, Granada, p 294
Company M, Aguado R, Sandoval J, Tavera JM, Jimenez de Cisneros C, Vera JA (2005) Biotic changes linked to a minor anoxic event (Faraoni Level, Latest Hauterivian, Early Cretaceous). Palaeogeogr Palaeoclimat Palaeoecol 224:186–199
Cope JCW (1967) The palaeontology and stratigraphy of the lower part of the Upper Kimmeridge Clay of Dorset. Bull Br Mus Nat Hist Geol 15:1–79
Cope JCW (1974) Upper Kimmeridge ammonite faunas of wash area and a subzonal scheme from the lower part of the Upper Kimmeridgian. Bull Geol Surv GB 1974:29–37
Crocker KC, DeNio, MJ, Ward, PD (1985) Stable isotopic investigations of early development in extant and fossil chambered cephalopods I. Oxygen isotopic composition of eggwater and carbon isotopic composition of siphuncle organic matter in Nautilus. Geochim Cosmochim Acta 49:2527–2532.
Currie ED (1942) Growth stages in the ammonite Promicroceras marstonense Spath. Proc R Soc Edinb B 61:344–367
Currie, ED. (1944) Growth stages in some Jurassic ammonites. Trans R Soc Edinb 61:171–198
Dagys AS, Weitschat W (1993) Extensive intraspecific variation in a Triassic ammonoid from Siberia. Lethaia 26:113–122
Daniel, TL, Helmuth BS, Sunders WB, Ward PD (1997) Septal complexity in ammonoid cephalopods increased mechanical risk and limited depth. Paleobiology 23: 470–481
D’Arpa C, Meléndez G. (2004) Oxfordian biostratigraphy and ammonite associations from west Sicily: biostratigraphic succession of genus Gregoryceras ad correlation with Tethyan Perisphinctid scale. Riv Ital Paleontol Strati 110:225–267
Davis RA (1972) Mature modification and dimorphism in selected Late Paleozoic ammonoids. Bull Am Paleontol 62:26–130
Davis RA, Furnish WM, Glenister BF (1969) Mature modification and dimorphism in Late Paleozoic ammonoids. In: Westermann GEG (ed) Sexual dimorphism in fossil metazoa and taxonomic implications. Schweizerbart, Stuttgart
Davis RA, Landman NH, Dommergues J-L, Marchand D, Bucher H (1996) Mature modifications and dimorphism in ammonoid cephalopods. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid paleobiology. Plenum, New York
Davis RA, Mapes RH, Klofak SM (1999) Epizoa on externally shelled cephalopods. In: Rozanov AY, Shevyrev AA (eds) Fossil Cephalopods: recent advances in their study. Russian Academy of Sciences, Palaeontological Institute, Moskva
De Baets K Klug C Korn D Landman NH. (2012) Early evolutionary trends in ammonoid embryonic development. Evolution 66:1788–1806
De Baets K Klug C Monnet C (2013) Intraspecific variability through ontogeny in early ammonoids. Paleobiology 39: 75–94
De Baets K, Landman NH, Tanabe K (2015) Ammonoid embryonic development. This volume
Dennis KJ, Cochran JK, Landman NH, Schrag DP (2013) The climate of the Late Cretaceous: new insights from the application of the carbonate clumped isotope thermometer to Western Interior Seaway macrofossil. Earth Planet Sci Lett 362:51–65
Denton EJ (1974) On the buoyancy and the lives of modern and fossil cephalopods. Proc R Soc Lond B 185:273–299
Denton EJ, Gilpin-Brown JB (1973) Floatation mechanisms in modern and fossil cephalopods. Adv Mar Biol 11:197–268
Ditchfield PW (1997) High Northern palaeolatitude Jurassic-Cretaceous palaeotemperature variation: new data from Kong Karls Land, Svalbard. Palaeogeogr Palaeoclimatol Palaeoecol 61:237–254
Doguzhaeva LA (1988) Siphuncular tubes and septal necks in ammonite evolution. In: Wiedmann J, Kullmann J (eds) Cephalopods–present and past. Schweizerbart, Stuttgart
Doguzhaeva LA (2002) Adolescent bactritoid, orthoceratoid, ammonoid and coleoid shells from the Upper Carboniferous and Lower Permian of the South Urals. In: Summesberger H, Histon K, Daurer A (eds) Cephalopods–present and past. Abh Geol B-A 57: 9–55
Doguzhaeva LA, Mutvei H (1989) Ptychoceras—a heteromorphic lytoceratid with truncated shell and modified ultrastructure (Mollusca: Ammonoidea). Palaeontogr Abt A 208:91–121
Doguzhaeva LA, Mutvei H (1991) Organization of the soft body in Aconeceras (Ammonitina), interpreted on the basis of shell morphology and muscle scars. Palaeontogr Abt A Palaeoz Stratigr 218:17–33
Doguzhaeva LA, Mutvei H. (1992) Radula of the Early Cretaceous ammonite Aconeceras (Mollusca: Cephalopoda). Palaeontogr A 223:167–177
Doguzhaeva LA, Mutvei H (1993) Structural features in Cretaceous ammonoids indicative of semiinternal or internal shells. In: House MR (ed) The Ammonoidea: Environment, Ecology, and Evolutionary Change. Systematics Association, special vol 47. Clarendon Press, Oxford
Doguzhaeva LA, Mapes RH, Summesberger H, Mutvei H (2007) The preservation of body tissues, shell, and mandibles in the ceratitid Ammonoid Austrotrachyceras (Late Triassic), Austria. In: Landman NH, Davis RA, Mapes RH (eds) Cephalopods Present and Past: new insights and fresh perspectives. Springer, Dordrecht
Doguzhaeva LA, Bengtson S, Mutvei H (2010) Structural and morphological indicators of mode of life in the Aptian lytoceratid ammonoid Eogaudryceras. In: Tanabe K, Shigeta Y, Sasaki T, Hirano H (eds) Cephalopods Present and Past. Tokai University Press, Tokyo
Donovan DT (1985) Ammonite shell form and transgression in the British Lower Jurassic. In: Bayer U, Seilacher A (eds) Sedimentary and evolutionary cycles, vol. 1. Springer–Verlag, Berlin
Donovan SK (1989) Taphonomic significance of the encrustation of the dead shell of recent Spirula spirula (Linneé) (Cephalopoda: Coleoidea) by Lepas anatifera Linné (Cirripedia: Thoracia). J Paleontol 63:698–702
Donovan DT (1993) Ammonites in 1991. In: House MR (ed) The Ammonoidea. environment, ecology and evolutionary change. Systematics Association, special vol. 47. Clarendon Press, Oxford
Doyle P, Whitham AG (1991) Palaeoenvironments of the Nordenskjold Formation: an Antarctic Late Jurassic–Early Cretaceous black shale-tuff sequence. In: Tyson RV, Pearson TH (eds) Modern and ancient continental shelf anoxia. Geol Soc Spec Publ 58:397–414
Duff KL (1975) Palaeoecology of a bituminous shale—the Lower Oxford Clay of central England. Palaeontology 18:443–482
Dunstan A, Bradshaw CJA, Marshall J (2011) Nautilus at Risk—Estimating population size and demography of Nautilus pompilius. PLoS ONE 6(2):e16716. doi:10.1371/journal.pone.0016716
Dutton A, Huber BT, Lohmann KC, Zinsmeister WJ (2007) High-resolution stable isotope profiles of a dimitobelid belemnite: implications for a paleodepth habitat and late Maastrichtian climate seasonality. Palaios 22:642–650
Ebel K (1983) Calculations on the buoyancy of ammonites. Neues Jahrb Geol Paläontol Monatshefte 1983:614–640
Ebel K (1985) Gehäusespirale und Septenformen bei Ammoniten unter Annahme vagil benthischer Lebensweise. Paläontol Z 59:109–123
Ebel K (1990) Swimming abilities of ammonites and limitations. Paläontol Z 64:25–37
Ebel K (1999) Hydrostatics of fossil ectochochleate cephalopods and its significance for the reconstruction of their lifestyle. Paläontol Z 73:277–288
Ebel K (1992) Mode of life and soft body shape of heteromorph ammonites. Lethaia 25:179–193
Eichler R, Ristedt H (1966a) Isotopic evidence on the Earth Life History of Nautilus pompilius (Linné). Science 153:734–736
Eichler R, Ristedt H (1966b) Untersuchungen zur Frühontogenie von Nautilus pompilius (Linné). Paläontol Z 40:173–191
Elmi S (1993) Area-rule, boundary layer and functional morphology of cephalopod shells (Ammonoids). Geobios Mem Spéc 15:121–138
Elmi S, Alméras Y (1984) Physiography, palaeotectonics and palaeoenvironment as control of changes in ammonite and brachiopod communities (an example from the Early and Middle Jurassic of western Algeria). Pelaeogeogr Palaeoclimatol Palaeoecol 47:347–360
Elmi S, Benshili K (1987) Relations entre la structuration tectonique, la composition des peuplements et l’évolution; exemple du Toarcian du Moyen–Atlas meridional (Maroc). Boll Soc Paleontol Ital 26:47–62
Enay R (1980) Paléobiogéographie et ammonites jurassiques: “rythmes fauniques” et variations du niveau marin; voies d’échanges, migrations et domaines biogéographiques. Mém Hors sér Soc Géol France 10:261–81
Enay R, Cariou E (1997) Ammonite faunas andpalaeobiogeography of the Himalayan belt during the Jurassic: initiation of a Late Jurassic austral ammonite fauna. Palaeogeogr Palaeoclimat Palaeoecol 134:1–38
Enay R, Mangold C (1982) Dynamique biogéographique et évolution des faunes d’ammonites au Jurassique. Bull Soc Géol France 24:1025–1046
Epstein S, Buchsbaum R, Lowenstam HA, Urey HC (1953) Revised carbonate-water isotopic temperature scale. Bull GSA 64:1315–1326
Etches S, Clarke J, Callomon J (2009) Ammonite eggs and ammonitellae from the Kimmeridge Clay Formation (Upper Jurassic) of Dorset, England. Lethaia 42:204–217
Fabricius F, Friedrichsen H, Jacobshagen V (1970) Paläotemperaturen und Paläoklima in Obetrias und Lias der Alpen. Int J Earth Sci 59:805–826
Fatherree JW, Harries PJ, Quinn TM (1998) Oxygen and carbon isotopic “dissection” of Baculites compressus (Mollusca: Cephalopoda) from Pierre Shale (Upper Campanian) of South Dakota: implications for paleoenvironmental reconstructions. Palaios 13:376–385
Fernandez-López S (1991) Taphonomic concepts for a theoretical biochronology. Rev Esp Paleontol 6:37–49
Fernández-López S (1997) Ammonites, taphonomic cycles and stratigraphic cycles in carbonate epicontinental platforms. Cuad Geol Iber 23:95–136
Fernández-López S, Meléndez G (1994) Abrasion surfaces on internal moulds of ammonites as palaeobathymetric indicators. Palaeogeogr Palaeoclimat Palaeoecol 110:29–42
Fernández-López S, Meléndez G (1995) Taphonomic gradients in Middle Jurassic ammonites of the Iberian Range (Spain). Geobios Mém Spéc 18:155–165
Fernandez-López S, Melendez G (1996) Phylloceratina ammonoids in the Iberian Basin during the Middle Jurassic: a model of biogeographical and taphonomic dispersal related to relative sea-level changes. Palaeogeogr Palaeoclimat Palaeoecol 120:291–302
Fernandez-López S, Melendez G (2004) Fossilization of ammonites and sedimentary events in deep environments of carbonate platform (highest Middle to lowest Upper Oxfordian, Iberian Range, Spain). Riv It Paleontol Strati 110:219–230
Fernandez-López SR, Henriques MH, Linares A, Sandoval J, Ureta MS (1999) Aalenian Tmetoceras (Ammonoidea) from Iberia. taxonomy, habitats and evolution In: Olóriz F, Rodríguez-Tovar FJ (eds) Advancing Research on Living and Fossil Cephalopods. Plenum, New York
Forester RW, Caldwell WGE, Oro FH (1977) Oxygen and carbon isotopic study of ammonites from Late Cretaceous Bearpaw Formation in southwestern Saskatchewan. Can J Earth Sci 14:2086–2110
Forsythe JW, Van Heukelem WF (1987) Growth. In: Boyle PR (ed) Cephalopod life cycles, Vol. 2. Academic Press, New York
Fourcade E, Azema J, Cecca F, Bonneau M, Peybernes B, Dercourt J (1991) Essai de reconstitution cartographique de la Paléogéographie et des Paléoenvironnements de la Tethys au Tithonique supérieur (138 Ã 135 Ma). Bull Soc Géol Fr 162:1197–1208
Fraaije RHB (2003) The oldest in situ hermit crab from the Lower Cretaceous of Speeton, UK. Palaeontology 46:53–58
Frye CJ, Feldman RM (1991) North American Late Devonian cephalopod aptychi. Kirtlandia 49:49–71
Galacz A, Horwath E (1985) Sedimentary and structural evolution of the Bakony Mountains (Transdanubian Central Range, Hungary): Paleogeographic implications. Acta Geol Hung 28:85–100
Geary D, Brieske TA, Bemis B (1992) The influence and interaction of temperature, salinity, and upwelling on the stable isotopic profiles of strombid gastropod shells. Palaios 7(1):77–85
Geczy B (1982) The Davoi Zone in the Bakony Mountains, Hungary. Ann Univ Sci Budap Sect Geol 21:1–11
Geczy B (1984) Provincialism of Jurassic ammonites; examples from Hungarian faunas. Acta Geol Hung 27:379–389
Geosecs Atlantic, Pacific and Indian Ocean Expeditions (1987) Shore-based data and graphics GEOSECS Executive Committee IDOE National Science Foundation 7
Geraghty M, Westermann GEG (1994) Composition and origin of Jurassic ammonite concretions from Alfeld, Germany: a biogenic alternative. Paläontol Z 68:473–490
Ginsburg RN, Schroeder JH (1973) Growth and submarine fossilization of algal cup reefs, Bermuda. Sedimentology 20:575–614
Gischler E, Ginsburg RN (1996) Cavity dwellers (coelobites) under coral rubble in southern Belize barrier and atoll reefs. Bull Mar Sci 58:570–589
Glaessner M (1931) Eine Crustaceenfauna aus den Lunzer Schichten Niederösterreichs. Jb Geol BA 81:467–486
Goodwin DH, Schöne BR, Dettman DL (2003) Resolution and fidelity of oxygen isotopes as palaeotemperature proxies in bivalve mollusk shells: models and observations. Palaios 18:110–125
Greenwald L, Ward PD, Greenwald OE (1980) Cameral liquid transport and buoyancy control in chambered Nautilus (Nautilus macromphalus). Nature 286:55–56.
Greenwald KP, Cook CB, Ward P (1982) The structure of the chambered Nautilus siphuncle: the siphuncular epithelium. J Morphol 172:5–22
Grossman EL, Ku T (1986) Oxygen and carbon isotope fractionation in biogenic aragonite: temperature effects. Chem Geol 59:59–74
Grunert P, Harzhauser M, Rögl F, Sachsenhofer R, Gratzer R, Soliman A, Piller WE (2010) Oceanographic conditions triggering the formation of an Early Miocene (Aquitanian) Konservat-Lagerstätte in the Central Paratethys Sea. Palaeogeogr Palaeoclimatol Palaeoecol 292:425–442
Guex J (2001) Environmental stress and atavism in ammonoid evolution. Eclogae Geol Helv 94:321–328
Gygi RA (1986) Eustatic sea level changes of the Oxfordian (Late Jurassic) and their effect documented in sediments and fossil assemblages of an epicontinental sea. Eclogae Geol Helv 79:455–491
Haimovici M, Brunetti NE, Rodhouse PG, Csirke J, Leta RH (1998) Illex argentinus. In: Rodhouse PG, Dawe EG, O’Dor RK (eds) Squid recruitment dynamics. The genus Illex as a model. The commercial Illex species. Infiuences on variability. FAO Fish Tech Pap 376:27–58
Hamada T (1964) Notes on the drifted Nautilus in Thailand. Contr Geol Palaeont Southeast Asia, 21, Sci Pap Coll Geb Educ 14:255
Hamada T (1984) Further notes on Nautilus drifts. Contr Geol Palaeont SE Asia 25:263
Hantzpergue P (1995) Faunal trends and sea-level changes: biogeographic patterns of Kimmeridgian ammonites on the Western European Shelf. Geol Rundsch 84:245–54
Hassan MA, Westermann GEG, Hewitt RA, Dokainish MA (2002) Finite-element analysis of simulated ammonoid septa (extinct Cephalopoda): septal and sutural complexities do not reduce strength. Paleobiology 28:113–126
Hay WW (2008) Evolving ideas about the cretaceous climate and ocean circulation. Cretac Res 29:725–753
Hayasaka S, Oki K, Tanabe K, Saisho T, Shinomiya A (1987) On the habitat of Nautilus pompilius in taon strait (Philippines) and the Fiji Islands. In: Saunders W B, Landman N H (eds) Nautilus: The Biology and Paleobiology of a Living Fossil. Plenum Press, New York
He S, Kyser TK, Caldwell GE (2005) Paleoenvironment of the Western Interior Seaway inferred from δ18O and δ13C values of molluscs from Cretaceous Bearpaw marine cyclothem. Palaeogeogr Palaeoclimatol Palaeoecol 217:67–85
Henderson RA, Price GD (2012) Paleoenvironment and paleoecology inferred from oxygen and carbon isotopes of subtropical mollusks from the late Cretaceous (Cenomanian) of Bathurst Island, Australia. Palaios 27(9):617–626
Hengsbach R (1978) Bemerkungen über das Schwimmvermögen der Ammoniten und die Funktion der Septen. Sitzungsberichte Ges Naturforschungs Freunde Berl Neue Folge 18:105–117
Heptonstall WB (1970) Buoyancy control in ammonoids. Lethaia 3:317–328
Hewitt RA (1988) Significance of early septal ontogeny in ammonoids and other ectocochliates. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Hewitt RA (1993) Relation of shell strength to evolution. In: House MR (ed) The Ammonoidea: Environment, Ecology, and Evolutionary Change. Systematics Association, special vol. 47. Clarendon Press, Oxford
Hewitt RA (1996) Architecture and strength of the ammonoid shell. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid paleobiology. Plenum, New York
Hewitt RA, Stait B (1988) Seasonal variations in septal spacing of Sepia officinalis and some Ordovician actinocerid nautiloids. Lethaia 21:283–394
Hewitt RA, Westermann GEG (1987) Functions of complexly fluted septa in ammonoid shells. II. Septal evolution and conclusions. Neues Jahrb Geol Paläontol Abh 174:135–169
Hewitt RA, Westermann GEG (1988) Stress and strain in Nautilus shells: some limitations on the buoyancy control and vertical migration of ectocochliates. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Hewitt RA, Westermann GEG (1990a) Nautilus shell strength variance as an indicator of habitat depth limits. Neues Jahrb Geol Paläontol Abh 179:73–97
Hewitt RA, Westermann GEG (1990b) Mosasaur tooth marks on the ammonite Placenticeras from the Upper Cretaceous Bearpaw Formation of Alberta. Can J Earth Sci 27:469–472
Hewitt RA, Westermann GEG (1997) Mechanical signifcance of ammonoid septa with complex sutures. Lethaia 30:205–212
Hewitt RA, Westermann GEG (2003) Recurrences of hypotheses about ammonites and Argonauta. J Paleontol 77:792–795
Hewitt RA, Westermann GEG, Checa A, Zaborski PM (1994) Growth rates of ammonoids estimated from aptychi. Geobios Mém Spéc 15:203–208
Hirano H (1986) Cenomanian and Turonian biostratigraphy of the off-shore facies of the Northern Pacific-an example of the Oyubari area, central Hokkaido, Japan. Bull Sci Eng Res Lab Waseda Univ 113:6–20
Hirano H (1988) Evolutionary mode of some late cretaceous ammonites in offshore waters. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past, Schweizerbart, Stuttgart
Hirano H (1993) Phyletic evolution of desmoceratine ammonoids through the Cenomanian–Turonian oceanic anoxic event. In: House MR (ed) The Ammonoidea: Environment, Ecology, and Evolutionary Change, Systematics association, special vol 47. Clarendon Press, Oxford
Hirano H, Okamoto T, Hattori K (1990) Evolution of some Late Cretaceous desmoceratine ammonoids. Trans Proc Palaeontol Soc Jpn NS 157:382–411
Hobson KA, Cherel Y (2006) Isotopic reconstruction of marine food webs using cephalopod beaks: new insight from captively raised Sepia officinalis. Can J Zool 84:766–770
Hoedemaeker PJ (1990) The Neocomian boundaries of the Tethyan Realm based on the distribution of ammonites. Cretac Res 11:331–342
Hoefs J (2004) Stable isotope geochemistry. (Fifth revised and updated edition). Springer Verlag, New York, p. 244
Hoffmann R, Lemanis R, Naglik C, Klug C (2015) Ammonoid buoyancy. This volume
House MR (1985) A new approach to an absolute time scale from measurements of orbital cycles and sedimentary microrhythms. Nature 316:721–725
House MR (ed) (1993a) The Ammonoidea: Environment, Ecology, and Evolutionary Change. Systematics Association, special vol 47. Clarendon Press, Oxford, p 354
House MR (1993b) Fluctuations in ammonoid evolution and possible environmental causes. In: House MR (ed) The Ammonoidea: Environment, Ecology, and Evolutionary Change. Systematics Association, special vol 47. Clarendon Press, Oxford
House MR, Price JD (1985) New Late Devonian genera and species of tornoceratid goniatites. Palaeontology 28:159–188
House MR, Senior JR (eds) (1981) The Ammonoidea: the evolution, classification, mode of life and geological usefulness of a major fossil group. Systematics Association, special vol 18. Academic Press, London
Howarth MK (1992) The ammonite family Hildoceratidae in the Lower Jurassic of Britain. Palaeontogr Soc Monogr 590(146):1–200
Hudson JD, Martill DM (1991) The lower Oxford Clay: Production and preservation of organic matter in the Callovian (Jurassic) of central England. In: Tyson RV, Pearson TH (eds) Modern and ancient continental shelf anoxia. Geol Soc Spec Pap 58:363–379
Hyatt A (1894) Phylogeny of an acquired characteristic. Am Philos Soc 32 143:349–647
Ifrim C, Stinnesbeck W (2010) Migration pathways of the late Campanian and Maastrichtian shallow facies ammonite Sphenodiscus in North America. Palaeogeogr Palaeoclimatol Palaeoecol 292(1):96–102
Ishii T (1981) Shells of Nautilus drifted ashore after an interval of 11 years. J Malac Soc Jpn 12:37–39
Jackson JBC, Winston JE (1982) Ecology of cryptic coral reef communities. I. Distribution and abundance of major groups of encrusting organisms. J Exper Mar Biol Ecol 57:135–147
Jacobs DK (1992a) Shape, drag, and power in ammonoid swimming. Paleobiology 18:203–220
Jacobs DK (1992b) The support of hydrostatic load in cephalopod shells. Adaptive and ontogenetic explanations of shell form and evolution from Hooke 1695 to the present. In: Hecht MK, Wallace B, MacIntyre RJ (eds) Evolutionary biology, vol 26. Plenum, New York
Jacobs DK, Chamberlain JA (1996) Buoyancy and hydrodynamics in ammonoids. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid Paleobiology. Plenum, New York
Jacobs DK, Landman NH (1993) Nautilus—a poor model for the function and behavior of ammonoids? Lethaia 26:101–112
Jacobs DK, Landman NH, Chamberlain JA (1994) Ammonite shell shape covaries with facies and hydrodynamics: iterative evolution as a response to changes in basinal environment. Geology 22:905–908
Jäger M, Fraaye R (1997) The diet of the Early Toarcian ammonite Harpoceras falciferum. Palaeontology 40:557–574
Jagt JWM, Bakel BWM van, Fraaije RHB, Neumann C (2006) In situ fossil hermit crabs (Paguroidea) from northwest Europe and Russia. Preliminary data on new records. Rev Mex Cienc Geol 23:364–369
Jansa LF, Emos P, Tcholke BE, Gradstein E, Sheridan RE (1979) Mesozoic-Cenozoic sedimentary formations of the North American Basin, western North Atlantic. In: Talman M, Hay W, Ryan WBE (eds) Deep drilling results in the Atlantic Ocean Continental Margins and Paleoenvironment, Maurice Ewing Series 3, American Geophysical Union, Washington, DC
Jeletzky JA, Zapfe H (1976) Coleoid and Orthocerid Cephalopods of the Rhaetian Zlambach Marl from the Fischerwiese near Aussee, Styria (Austria). Ann Naturhist Mus Wien 71:69–106
Jenkyns H (1988) The early Toarcian (Jurassic) anoxic event. Stratigraphic, sedimentary, and geochemical evidence. Am J Sci 288:101–151
Jones DS, Williams DF, Romanek CS (1986) Life history of symbiont-bearing giant clams from stable isotope profiles. Science 231:46–48
Jordan R, Stahl W (1970) Isotopische Paläotemperatur-Bestimmungen an jurassischen Ammoniten und grundsätzliche Voraussetzungen für diese Methode. Geol Jahrb 89:33–62
Kahn P, Kant R (1975) Biometrische Untersuchungen zur Lobenentwicklung. Paläontol Z 49:287–297
Kaltenegger W (1967) Paläotemperaturbestimmungen an aragonitischen Dibranchiatenrostren der Trias. Naturwissenschaften 54:515
Kaltenegger W, Preisinger A, Rögl F (1971) Palaeotemperature determinations of aragonitic mollusks from the Alpine Mesozoic. Palaeogeogr Palaeoclimatol Palaeoecol 10:273–285
Kammer TW, Brett, CE, Boardman DR IS, Mapes RH (1986) Ecologic stability of the dysaerobic biofacies during the Late Paleozoic. Lethaia 19:109–121
Kanie Y, Fukuda Y, Nakayama H, Seki K, Hattori M (1980) Implosion of living Nautilus under increased pressure: Paleobiology 6(1):44–47
Kant R (1975) Biometric analysis of ammonoid shells. Paläontol Z 49:203–220
Kant R, Kullmann J (1988) Changes in conch form in the Paleozoic ammonoids. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Kase T, Shigeta F, Futakami M (1994) Limpet home depressions in Cretaceous ammonites. Lethaia 25:49–58
Kase T, Johnson PA, Seilacher A, Boyce JP (1998) Alleged mosasaur bite marks on late Cretaceous ammonites are limpet (patellogastropod) home scars. Geology 26:947–950
Kashiyama Y, Ogawa NO, Chikaraishi Y, Kashiyama N, Sakai S, Tanabe K, Ohkouchi N (2010) Reconstructing the life history of modern and fossil nautiloids based on the nitrogen isotopic composition of shell organic matter and amino acids. In Tanabe K, Shigeta Y, Sasaki T, Hirano H, (eds.) Cephalopods Present and Past. Tokai Unive Press, Tokyo
Kauffman EG (1977) Geological and Biological Overview: Western Interior Cretaceous Basin, U.S.A. In: Kauffman EG (ed) Field Guide: North American Paleontological Convention II; Cretaceous facies, faunas, and paleoenvironments across the Western Interior Basin: The Mountain geologist 14:75–79
Kauffman EG (1978) Benthic environments and paleoecology of the Posidonienschiefer (Toarcian). Neues Jahrb Geol Paläontol Abh 157:18–36
Kauffman EG (1990) Mosasaur predation on ammonites during the Cretaceous–an evolutionary history. In: Boucot AJ (ed) Evolutionary paleobiology of behaviour and coevolution. Elsevier, New York
Kauffman EG, Kiesling RV (1960) An Upper Cretaceous ammonite bitten by a mosasur. Contr Michigan Univ Mus Paleontol 15:193–248
Kauffman EG, Sawdo JK (2013) Mosasaur predation on a nautiloid from the Maastrichtian Pierre Shale, Central Colorado, Western Interior Basin, United States. Lethaia 46:180–187
Kauffman EG, Villamil T, Harries PJ, Meyer C (1992) The flat clam controversy: where did they come from? Where did they go? Paleontol Soc Spec Publ 6:159
Kawabe F (2003) Relationship between mid-Cretaceous (upper Albian-Cenomanian) ammonoid facies and lithofacies in the Yezo forearc basin, Hokkaido, Japan. Cretac Res 24:751–763
Kennedy WJ (1971) Cenomanian ammonites from southern England. Spec Pap Palaeont 8:1–133
Kennedy WJ, Cobban WA (1976) Aspects of ammonite biology, biogeography, and biostratigraphy. Spec Pap Palaeont 17:1–94
Kennedy WJ, Landman NH, Christensen WK, Cobban WA, Hancock JM (1998) Marine connections in North America during the Late Maastrichtian: palaeogeographic and palaeobiogeographic significance of Jeletzkytes nebrascensis Zone cephalopod fauna from the Elk Butte Member of the Pierre Shale, SE South Dakota and NE Nebraska. Cretac Res 19:745–775
Kennedy WJ, Landman NH, Cobban WA, Larson NL (2002) Jaws and radulae in Rhaeboceras, a late Cretaceous ammonite. In: Summesberger H, Histon K, Daurer A (eds) Cephalopods Present and Past B-A 57:113–132
Keupp H (1991) Bissmarken oder postmortale Implosionsstrukturen? Fossilien 1992:141–146
Keupp H (1992) Paläopathologie der Ammonitenfauna aus dem Obercallovium der Normandie und ihre palökologische Interpretation. Berl Geowiss Abh 3:171–189
Keupp H (1997) Paläopahologische Analyse einer „Population“ von Dactylioceras athleticum (Simpson) aus dem Unter-Toarcium von Schlaifhausen/Oberfranken. Berl Geowiss Abh E 25:243–267
Keupp H (2000) Ammoniten. Paläobiologische Erfolgsspiralen. Thorbecke, Stuttgart
Keupp H (2006) Sublethal punctures in body chambers of Mesozoic ammonites (forma aegrafenestra n. f.), a tool to interpret synecological relationships, particularly predator–prey interactions. Paläontol Z 80:112–123
Keupp H (2007) Complete ammonoid jaw apparatuses from the Solnhofen plattenkalks: implications for aptychi function and microphagous feeding of ammonoids. N Jahrb Geol Paläontol Abh 245:93–101
Keupp H, Hoffmann R (2015) Ammonoid paleopathology. This volume
Keupp H, Schobert J (2011) Gehäuseanomalien bei Klein-Ammoniten von Buttheim/Oberfranken. Fossilien 3:164–174
Kim S-T, O’Neil JR, Hillaire-Marcel C, Mucci A (2007) Oxygen isotope fractionation between synthetic aragonite and water: Influence of temperature and Mg2+ concentration. Geochim Cosmochim Acta 71:4704–4715
Klinger HC (1980) Speculations on buoyancy control and ecology in some heteromorph ammonites. In: House MR, Senior JR (eds) The Ammonoidea: The evolution, classification, mode of life and geological usefulness of a major fossil group. Systematics Association, special vol 18. Academic Press, London
Klofak SM, Landman NH, Mapes RH (1999) Embryonic development of primitive ammonoids and the morphology of the Ammonoidea. In: Olóriz F, Rodríguez-Tovar FJ (eds) Advancing Research on Living and Fossil Cephalopods. Plenum, New York
Klofak SM, Landman NH, Mapes RH (2007) Patterns of embryonic development in early to middle Devonian ammonoids. In: Landman NH, Davis RA, Mapes RH (eds) Cephalopods Present and Past: new insights and fresh perspectives. Springer, Dortrecht
Klompmaker AA, Waljaard, Fraaije RHB (2009) Ventral bite marks in Mesozoic ammonoids. Palaeogeogr Palaeoclimatol Palaeoecol 280:245–257
Klug C (2001) Life-cycle of some Devonian ammonoids. Lethaia 34:215–233
Klug C (2002) Conch parameters and habitats of Emsian and Eifelian ammonoids from the Tafilalt (Morocco) and their relation to global events. Abh Geol B-A 57:523–538
Klug C (2007) Sublethal injuries in Early Devonian cephalopod shells from Morocco. Acta Palaeont Polonica 52:749–759
Klug C, Hoffmann R (2015) Ammonoid septa and sutures. This volume
Klug C, Korn D (2004) The origin of ammonoid locomotion. Acta Palaeont Pol 49:235–242
Klug C, Lehmann J (2015) Soft-part anatomy of ammonoids: reconstructing the animal based on exceptionally preserved specimens and actualistic comparisons. This volume
Klug C, Schatz W, Korn D, Reisdorf A (2005) Morphological fluctuations of ammonoid assemblages from the Muschelkalk (Middle Triassic) of the Germanic Basin—indicators of their ecology, extinctions, and immigrations. Palaeogeogr, Palaeoclim Palaeoeco 221:7–34
Klug C, Kröger B, Kiessling W, Mullins GL, Servais T, Frýda J, Korn D, Turner S (2010) The Devonian nekton revolution. Lethaia 43:465–477
Klug C, Korn D, Landman NH, Tanabe K, De Baets K, Naglik C (2015a) Describing ammonoid conchs. This volume
Klug C, Zatoń M, Parent H, Hostettler, Tajika A (2015b) Mature modifications and sexual dimorphism. This volume
Kobashi T, Grossman EL (2003) The oxygen isotopic record of seasonality in Conus shells and its application to understanding late middle Eocene (38 Ma) climate. Paleontol Res 7:343–355
Kobayashi T (1954) A contribution toward Palaeo-Flumenology, Science of the Oceanic Current in the Past, with a description of a new Miocene Aturia from Central Japan. Jap J Geol Geogr Transact 25:35–59
Korn D (1986) Ammonoid evolution in late Famennian and early Tournaisian. Ann Soc Géol Belg 109:49–54
Korn D (1988) Oberdevonishe Goniatiten mit dreieckigen Innenwindungen. N Jahrb Geol Palaont Mh 1988:605–610
Korn D (1992) Relationship between shell form, septal construction and suture line in clymeniid cephalopods (Ammonoidea: Upper Devonian). N Jahrb Geol Paläontol Abh 185:115–130
Korn D (2010) A key for the description of Palaeozoic ammonoids. Fossil Rec 13:5–12
Korn D, Klug C (2001) Biometric analyses of some Palaeozoic ammonoid conchs. Berl Geowiss Abh E 36:173–187
Korn D, Klug C (2002) Ammoneae Devonicae. In Riegraf (ed). Fossilium Catalogus I: Animalia. Backhuys Publishers, Leiden, 138:375
Korn D, Klug C (2007) Conch form analysis, variability, morphological disparity, and mode of life of the Frasnian (Late Devonian) ammonite Manticoceras from Coumiac (Montagne nOire, France). In: Landman NH, Davis RA, Mapes RH (eds) Cephalopods Present and Past: new insights and fresh perspectives. Springer, Dortrecht
Korn D, Klug C (2012) Ammoneae Devonicae. In: Riegraf W (ed) Fossilium Catalugus I: Animalia. Blackhuys Publishers, Leiden
Korn, D, Klug, C, Mapes RH (2004) Cuboid Carboniferous ammonoids. Mitt Geol Palaöont Inst Univ Hambg 88:79–98
Kowalewski M (2002) The fossil record of predation: an overview of analytical methods. In: Kowalewski M, Kelley PH (eds) The fossil record of predation. Paleontol Soc Pap 8:1–42
Kröger B (2000) Schalenverletzungen an jurassischen Ammoniten—ihre paläobiologische und paläoökologische Aussagefähigkeit. Berl Geowiss Abh E 33:1–97
Kröger B (2001) Comments on Ebel’s benthic–crawler hypothesis for ammonoids and exticnt nautiloids. Paläontol Z 75:123–125
Kröger B (2002a) On the effciency of the buoyancy apparatus in ammonoids: evidences from sublethal shell injuries. Lethaia 35:61–70
Kröger B (2002b) On the ability of withdrawing of some Jurassic ammonoids. In: Summesberger H, Histon K, Daurer K (eds) Cephalopods Present and Past. Abh Geol B-A 67:199–244
Kröger B (2002c) Antipredatory traits of the ammonoid shell—Indications from Jurassic ammonoids with sublethal injuries. Paläontol Z 76:223–234
Kröger B (2005) Adaptive evolution in Paleozoic coiled cephalopods. Paleobiology 31:253–268
Kroh A, Harzhauser M (1999) An echinoderm fauna from the Lower Miocene of Austria: paleoecology and implications for Central Paratethys Paleobiogeography. Ann Nat Mus Wien 101A:145–191
Kruta I, Landman N, Rouget I, Cecca F, Tafforeau P (2011) The role of ammonites in the Mesozoic marine food web revealed by jaw preservation. Science 2011:70–72
Kruta I, Landman N, Cochran JK (2014) A new approach for the determination of ammonite and nautilid habitats. PLos ONE 9(1):e87479. doi:10.1371/journal.pone.0087479
Krystyn L (1991) Die Fossillagerstätten der alpinen Trias. In: Nagel D, Rabeder G (eds) Exkursionen im Jungpaläozoikum und Mesozoikum Österreichs. Österr Paläontol Ges, Wien
Kulicki C (1974) Remarks on the embryogeny and postembryonal development of ammonites. Acta Palaeontol Pol 19:201–224
Kulicki C (1979) The ammonite shell: Its structure, development and biological significance. Palaeontol Pol 39:97–142
Kulicki C (1996) Ammonoid shell microstructures. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid Paleobiology. Plenum, New York
Kulicki C, Doguzhaeva L (1994) Development and calcification of the ammonitella shell. Acta Palaeont Pol 39:17–44
Kulicki C, Landman NH, Heaney MJ, Mapes RH, Tanabe K (2002) Morphology of the early whorls of goniatites from the Carboniferous Buckhorn Asphalt (Oklahoma) with aragonitic preservation. In: Summesberger H, Histon K, Daurer A (eds) Cephalopods—present and past. Abh Geol B-A 57:225–255
Kullmann J (1981) Carboniferous goniatites. In: House MR, Senior JR (eds) The Ammonoidea. Systematics Association, special vol 18. Academic Press, London
Kullmann J, Scheuch J (1970) Wachstums-Änderungen in der Ontogenese paläozoischer Ammonoideen. Lethaia 3:397–412
Kullmann J, Scheuch J (1972) Absolutes und relatives Wachstum bei Ammonoideen. Lethaia 5:129–146
Kvantaliani IV, Sakharov AS (1986) Valanginian ammonites of the Northern Caucasus (Russ.). Geol Balc 163:55–68
Landman NH (1982) Embryonic shells of Baculites. J Paleontol 56:1235–1241
Landman NH (1985) Preserved ammonitellas of Scaphites (Ammonoidea, Ancyloceratina). Am Mus Novit 2815:1–10
Landman NH (1986) Shell abnormalities in scaphitid ammonites. Lethaia 19:211–224
Landman NH (1987) Ontogeny of Upper Cretaceous (Turonian-Santonian) scaphitid ammonites from the Western Interior of North America: systematics, developmental patterns, and life history. Bull Am Mus Nat Hist 185:117–241
Landman NH (1988) Early ontogeny of Mesozoic ammonites and nautilids. In: Wiedmann, J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Landman NH, Cobban WA (2007) Ammonite touch marks in Upper Cretaceous (Cenomanian-Santonian) deposits of the Western Interior Seaway. In: Landman NH, Davis RA, Mapes RH (eds) Cephalopods Present and Past: new insights and fresh perspectives. Springer, Dortrecht
Landman NH, Cochran JK (1987) Growth and longevity of Nautilus. In: Saunders WB, Landman NH (eds) Nautilus: The Biology and Paleobiology of a Living Fossil. Topics in Geobiology, vol 6. Springer, New York
Landman NH, Cochran, JK (2010) Growth and longevity of Nautilus. In: Saunders WB, Landman NH (eds) Nautilus: The Biology and Paleobiology of a Living Fossil. Topics in Geobiology, vol 6. Springer, New York
Landman NH., Klofak, SM (2012). Anatomy of a concretion: life, death, and burial in the Western Interior Seaway. Palaios 27:672–693
Landman NH, Waage KM (1986) Shell abnormalities in scaphitid ammonites. Lethaia 19:211–224
Landman NH, Waage KM (1993) Scaphitid ammonites of the Upper Cretaceous (Maastrichtian) Fox Hills Formation in South Dakota and Wyoming. Bull Am Mus Nat Hist 215:l–257
Landman NH, Rye DM, Shelton KL (1983) Early ontogeny of Eutrephoceras compared to Recent Nautilus and Mesozoic ammonites: evidence from shell morphology and light stable isotopes. Paleobiol 9:269–279
Landman NH, Saunders WB, Winston JE, Harries PJ (1987) Incidence and kinds of epizoans on the shells of living Nautilus. In Saunders WB, Landman NH (eds) Nautilus: The Biology and Paleobiology of a Living Fossil. Plenum, New York
Landman NH, Cochran, JK, Rye DM, Tanabe K, Arnold JM (1994) Early life history of Nautilus: evidence from isotopic analysis of aquarium reared specimens. Paleobiology 20:40–51
Landman NH, Tanabe K, Davis RA (eds) (1996a) Ammonoid Paleobiology. Plenum Press, New York, pp 857
Landman NH, Tanabe K, Shigeta Y (1996b) Ammonoid embryonic development. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid Paleobiology. Plenum, New York
Landman NH, Klofak SM, Sarg KB (2003) Variation in adult size of scaphitid ammonites from the Upper Cretaceous Pierre Shale and Fox Hill Formation. In: Harries PJ (ed) Approaches in high-resolution stratigraphic paleontology. Kluwer Academic, Dordrecht
Landman NH, Davis RA, Mapes RH (eds) (2007a) Cephalopods Present and Past: new insights and fresh perspectives. Springer, Dortrecht
Landman NH, Larson NL, Cobban WA (2007b) Jaws and radula of Baculites from the Upper Cretaceous (Campanian) of North America. In: Landman NH, Davis RA, Mapes RH (eds) Cephalopods Present and Past: new insights and fresh perspectives. Springer, Dortrecht
Landman NH, Mapes RH, Cruz C (2010) Jaws and soft tissues in ammonoids from Lower Carboniferous (Upper Mississippian) Cear Gulch Beds Montan, USA. In: Tanabe K, Shigeta Y, Sasaki T, Hirano H (eds) Cephalopods Present and Past. Tokai Univ Press, Tokyo
Landman NH, Cobban WA, Larson NL (2012) Mode of life and habitat of scaphitid ammonites. Geobios 45:87–98
Laptikhovsky VL, Rogov, MA, Nikolaeva SE, Arkhipkin AI (2013) Environmental impact on ectocochleate cephalopod reproductive strategies and the evolutionary significance of cephalopod egg size. Bull Geosci 88:83–93
Larson NL (2003) Predation and pathologies in the Late Cretaceous ammonite family Scaphitidae. Mid-Am Paleontol Soc 26:1–30
Larson NL (2007) Deformities in the Late Callovian (Late Middle Jurassic) ammonite fauna from Saratov, Russia. In: Landman NH, Davis RA, Mapes RH (eds) Cephalopods Present and Past: new insights and fresh perspectives. Springer, Dortrecht
Lécuyer C, Bucher H (2006) Stable isotope composition of late Jurassic ammonite shell: a record of seasonal surface water temperatures in the southern hemisphere? eEarth Discuss 1:1–7
Lécuyer C, Reynard B, Martineau F (2004) Stable isotope fractionation between mollusc shells and marine waters from Martinique Island. Chem Geol 213:293–305
Lehmann U (1966) Dimorphismus bei Ammoniten der Ahrensburger Lias-Geschiebe. Paläontol Z 40:26–55
Lehmann U (1975) Über Nahrung und Ernährungsweise der Ammoniten. Paläontol Z 49:187–195
Lehmann U (1976) Ammoniten-Ihr Leben und Ihre Umwelt. Enke, Stuttgart
Lehmann U (1980) Ammonite jaws and soft parts. In: House MR, Senior JR (eds) The Ammonoidea: the evolution, classification, mode of life and geological usefulness of a major fossil group. Systematics Association, special vol 18, Academic Press, London
Lehmann U (1985) Zur Anatomie der Ammoniten: Tintenbeutel, Kiemen, Augen. Paläontol Z 59:99–108
Lehmann U (1988) On the dietary habits and locomotion of fossil cephalopods. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Lehmann J (2000) Upper Albian ammonites from ODP leg 171B off northern Florida. Palaeontology 43:41–61
Lehmann U, Kulicki C (1990) Double function of aptychi (Ammonoidea) as jaw elements and opercula. Lethaia 23:325–331
Lehmann U, Weitschat W (1973) Zur Anatomie und Okologie der Ammoniten. Funde von Kropf und Kiemen. Paläontol Z 47:69–76
Lehmann U, Tanabe K, Kanie Y, Fukuda Y (1980) Über den Kieferapparat der Lytoceraten (Ammonoidea). Paläontol Z 54:319–329
Levitus (1994) World Ocean Atlas, an atlas of objectively analyzed fields of major ocean parameters at the annual, seasonal, and monthly time scales. http://iridl.ldeo.columbia.edu/SOURCES/.LEVITUS94/. Accessed 18 June 2012
Lewy Z (2002) The function of ammonite fluted septal margins. J Paleontol 76:63–69
Lewy Z (2003) Reply to Checa and to Hewitt and Westermann. J Paleontol 77:796–798
Little R, Baker DR, Leythaeuser D, Rullkottner J (1991) Keys to the depositional history of the Posidonia Shale (Toarcian) in the Hills Syncline, northern Germany. In: Tyson RV, Pearson TH (eds) Modern and ancient continental margin anoxia. Geol Soc Spec Publ 58:311–333
Loh H, Maul B, Prauss M, Riegel W (1986) Primary production, marl formation and carbonate species in the Posidonia Shale of NW Germany. Mitt Geol Palaont Inst Univ Hambg 60:397–421
Lominadze T, Sakharov AS (1988) Ecology of caucasian callovian ammonitida. In: Wiedmann J, Kullmann J (eds) Cephalopods—present and past. Schweizerbart, Stuttgart
Longinelli A (1966) Ratios of oxygen-18: oxygen-16 in phosphate and carbonate from living and fossil marine organisms. Nature 211:923–927
Longinelli A., Nuti S (1973) Revised phosphate-water isotopic temperature scale. Earth Planet Sci Lett 19:373–376
Lowenstein HA, Epstein S (1954) Paleotemperatures of the post-Albian Cretaceous as determined by the oxygen isotope method. J Geol 62:207–248
Lukeneder A (2003) The Karsteniceras level: dysoxic ammonoid beds within the Early Cretaceous (Barremian, Northern Calcareous Alps, Austria). Facies 49:87–100
Lukeneder A (2004) The Olcostephanus level: an Upper Valanginian ammonoid mass-occurrence (Lower Cretaceous, Northern Calcareous Alps, Austria). Acta Geol Pol 54:23–33
Lukeneder A (2005) The Early Cretaceous Karsteniceras level in the Vienna Woods (Northern Calcareous Alps, Lower Austria). Geol Carpathica 56:307–315
Lukeneder A (2007) Cephalopod evolution: a new perspective—implications from two Early Cretaceous ammonoid suborders (Northern Calcareous Alps, Upper Austria). Denisia 20:395–404
Lukeneder A (2008) The ecological significance of solitary coral and bivalve epibionts on lower Cretaceous (Valangianian–Aptian) ammonoids from the Italian dolomites. Acta Geol Pol 58:425–436
Lukeneder A (2012) Computed 3D visualisation of an extinct cephalopod using computer tomographs. Comput Geosci 45:68–74
Lukeneder A, Grunert P (2013) Palaeoenvironmental evolution of the Southern Alps across the Faraoni Level equivalent: new data from the Trento Plateau (Upper Hauterivian, Dolomites, N. Italy). Acta Geol Pol 63:89–104
Lukeneder A, Harzhauser M (2002) Shell accumulations of the Nautilidae Aturia (Aturia) aturi (Bast.) in the Lower Micocene Paratethys (Lower Austria). In: Summesberger H, Histon K, Daurer A (eds) Cephalopods Present and Past. Abh Geol B-A 57:459–466
Lukeneder A, Harzhauser M (2003) Olcostephanus guebhardi as cryptic habitat for an Early Cretaceous coelobite community (Valanginian, Northern Calcareous Alps, Austria). Cretac Res 24:477–485
Lukeneder S, Lukeneder A (2014) A new ammonoid fauna from the Carnian (Upper Triassic) kasimlar formation of the Taurus Mountains (Anatolia, Turkey). Palaeontology 57:357–396
Lukeneder A, Mayrhofer S (2014) Taphonomic implications from Upper Triassic mass flow deposits: 2-dimensional reconstructions of an ammonoid mass occurrence (Carnian, Taurus Mountains, Turkey). Geol Carpath 65/5:339–364
Lukeneder A, Reháková D (2004) Lower Cretaceous section of the Ternberg Nappe (Northern Calcareous Alps, Upper Austria): facies-changes, biostratigraphy and paleoecology. Geol Carpathica 55:227–237
Lukeneder A, Smrečková M (2006) An Early Cretaceous radiolarian assemblage: palaeoenvironmental and palaeoecological implications for the Northern Calcareous Alps (Barremian, Lunz Nappe, Lower Austria). Ann Naturhist Mus Wien 107A:23–57
Lukeneder A, Tanabe K (2002) In situ finds of aptychi in the Barremian of the Alpine Lower Cretaceous (Barremian, Northern Calcareous Alps, Upper Austria). Cretac Res 23:15–24
Lukeneder A, Harzhauser M, Müllegger S, Piller W (2008) Stable isotopes (δ18O and δ13C) in Spirula spirula shells from three major oceans indicate developmental changes paralleling depth distributions. Mar Biol 154:175–182
Lukeneder A, Harzhauser M, Müllegger S, Piller WE (2010) Ontogeny and habitat change in Mesozoic cephalopods revealed by stable isotopes (δ18O, δ13C). Earth Planet Sci Lett 296:103–114
Lukeneder S, Lukeneder A, Harzhauser M., Islamoglu Y, Krystyn L, Lein R (2012) A delayed carbonate factory breakdown during the Tethyan-wide Carnian Pluvial Episode along the Cimmerian terranes (Taurus, Turkey). Facies 58:279–296
Lukeneder A, Suttner TJ, Bertle RJ (2013) New ammonoid taxa from the Lower Cretaceous Giumal Formation of the Tethyan Himalaya (Northern India). Palaeontology 56:991–1028
Lukeneder S, Lukeneder A, Weber GW (2014) Computed reconstruction of spatial ammonoid-shell orientation captured from digitized grinding and landmark data. Comput Geosci 64:104–114
Maeda H (1993) Dimorphism of Late Cretaceous false-Puzosiinae ammonites, Yokoyamaoceras Wright and Matsumoto, 1954 and Neopuzosia Matsumoto, 1954. Trans Proc Palaeont Soc Japan N S 169:97–128
Maeda H, Seilacher A (1996) Ammonoid taphonomy. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid paleobiology. Plenum, New York
Maeda H, Mapes RH, Mapes G (2003) Taphonomic features of a Lower Permian beached cephalopod assemblage from Central Texas. Palaios 18:421–434
Manfrin S, Mietto P, Preto N (2005) Ammonoid biostratigraphy of the Middle Triassic Latemar platform (Dolomites, Italy) and its correlation with Nevada and Canada. Geobios 38:477–504
Mangold K (1989) Reproduction, croissance et durée de vie. In: Grassé PP (ed) Traité de Zoolgie, 5 (4). Anatomie, Systématique, Biologie. Céphalopodes. Masson, Paris
Mapes RH (1979) Carboniferous and Permian Bactritoidea (Cephalopoda) in North America. Univ Kans Paleontol Contrib Artic 64:l–75
Mapes RH (1987) Upper Paleozoic cephalopod mandibles: frequency of occurrence, modes of preservation, and paleoecological implications. J Paleontol 61:521–538
Mapes RH, Chaffin DT (2003) Predation on cephalopods. a genaral overview with case study from the Upper Carboniferous of Texas. In: Kelley PH, Kowalewski M, Hansen TA (eds). Predator-Prey Interactions in the Fossil record. Kluwer Academic, New York
Mapes RH, Dalton RB (2002) Scavening or Predation?—Mississippian ammonoid accumulations in carbonate concretion halos around Rayonnoceras (Actinoceratoidea—Nautiloidea) body chambers. In: Summesberger H, Histon K, Daurer A (eds) Cephalopods Present and Past. Abh der Geol B-A 57:407–422
Mapes RH, Hansen MC (1984) Pennsylvanian shark–cephalopod predation: a case study. Lethaia 17:175–183
Mapes RH, Nützel A (2009) Late Palaeozoic mollusc reproduction: cephalopod egg-laying behavior and gastropod larval palaeobiology. Lethaia 42:341–356
Mapes RH, Tanabe K, Landman NH, Faulkner CJ (1992) Upper Carboniferous ammonoid shell clusters: transported accumulations or in situ nests? Paleontol Soc Spec Pub 6:196
Mapes RH, Sims MS, Boardman DR II (1995) Predation on the Pennsylvanian ammonoid Gonioloboceras and ist implications for allochthonous vs. autochthonous accumulations of goniatites and other ammonoids. J Palaeontol 69:441–446
Mapes RH, Hembree DI, Rasor BA, Stigall A, Goirand C, de Forges BR (2010a) Modern Nautilus (Cephalopoda) taphonomy in a subtidal to backshore environment, Lifou (Loyalty Islands). Palaios 25:656–670
Mapes RH, Landman NH, Cochran K, Goiran C, Richer De Forges B, Renfro A (2010b) Early taphonomy and significance of naturally submerged Nautilus shells from the New Caledonia Region. Palaios 25:597–610
Marchand T (1984) Ammonites et paleoenvironnements: une nouvelle approche. Geobios Mém Spéc 8:101–107
Marchand D (1992) Ammonites et paléoprofondeur: les faits, les interprétations. Paleovox 1:49–68
Marchand D, Thierry J (1986) Relations entre les évènements calloviens et l’évolution des peuplements d’ammonites en Europe occidentale. Bull Centr Rech Elf Aquitaine 10:383–92
Marchand T, Thierry J (1997) Enregistrement des variations morphologiques et de la composition des peuplements d’ammonites durant le cycle régressif/transgressif de 2éme ordre Bathonien inférieur–Oxfordien inférieur en Europe occidentale. Bull Soc Géol Fr 168:121–132
Marcinowski R, Wiedmann J (1985) The Albian ammonite fauna of Poland and its paleogeographical significance. Acta Geol Pol 35:199–218
Marcinowski R, Wiedmann J (1988) Paleogeographic implications of the Albian ammonite faunas of Poland. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Marchand D, Thierry J, Tintant H (1985) Influence des seuls et des hauts–fonds sur la morphology et l’évolution des ammonites. Inst Sci Terre Univ Dijon Bull Sect Sci 9:191–202
Marshall JD (1981) Zoned calcites in Jurassic ammonite chambers: trace elements, isotopes and neomorphic origin. Sedimentology 28:867–887
Martin AW, Catala-Stucki I, Ward DP (1978) The growth rate and reproductive behaviour of Nautilus macromphalus. N Jahrb Geol Paläontol Abh 156:207–225
Massare JA (1987) Tooth morphology and prey preference of Mesozoic marine reptiles. J Vert Paleontol 7:121–137
Matsumoto T, Futakami M, Tanabe K, Obata I (1981) Upper Turonian ammonite assemblages in the Pombets area, central Hokkaido. Bull Kitakyushu Mus Nat Hist 3:1–10
Matsuoka A, Anso J, Nakada K, Terabe K, Sato T (2010) Biometrical analysis on primary rib number of the Middle Jurassic ammonoid Pseudoneuqueniceras yokoyamai (Kobayashi & Fukada) and its allied forms. In: Tanabe K, Shigeta Y, Sasaki T, Hirano H (eds) Cephalopods Present and Past. Tokai University Press, Tokyo
McArthur JM, Mutterlose J, Price GD, Rawson PF, Ruffell A., Thirlwall MF (2004) Belemnites of Valanginian, Hauterivian and Barremian age: Sr-isotope stratigraphy, composition (87Sr/86Sr, δ13C, δ18O, Na, Sr, Mg), and palaeo-oceanography. Palaeogeogr Palaeoclimatol Palaeoecol 202:253–272
McConnaughey TA, Gillikin DP (2008) Carbon isotopes in mollusk shell carbonates. Geo-Mar Lett 28:287–299
Meesters E, Knijn R, Pennarts R, Roebers G, van Soest RWM (1991) Sub-rubble communities of Curacao and Bonaire coral reefs. Coral Reefs 10:189–197
Mehl J (1978a) Ein Koprolith mit Ammoniten-Aptychen aus dem Solnhofer Plattenkalk. Jahresb Wetterau Ges Naturkunde 129–130:85–89
Mehl J (1978b) Anhäufungen scherbenartiger Fragmente von Arnmonitenschalen im süddeutschen Lias und Malm und ihre Deutung als Frassreste. Ber Naturforsch Ges Freib Breisgau 68:75–93
Mehl J (1984) Radula and arms of Michelinoceras sp. from the Silurian of Bohemia. Paläontol Z 58:211–229
Meischner D (1968) Perniciöse, Epökie von Placunopsis auf Ceratites. Lethaia 1:156–174
Merkt J (1966) Über Austern und Serpeln als Epöken auf Ammonitengehäusen. N Jahrb Geol Palöont Abh 125:467–479
Milson CV (1994) Saccocoma, a benthic crinoid from the Jurassic Solnhofen Limestone, Germany. Palaeontology 37:121–130
Minagawa M, Wada E (1984) Stepwise enrichment of 15N along food chains: further evidence and the relation between delta 15N and animal age. Geochim Cosmochim Acta 48:1135–1140
Misaki A, Maeda H, Kumagae T, Ichida M (2013) Commensal anomiid bivalves on Late Cretaceous heteromorph ammonites from south-west Japan. Palaeontology 57:77–95
Monnet C (2009) The Cenomanian–Turonian boundary mass extinction (Late Cretaceous): new insights from ammonoid biodiversity patterns of Europe, Tunesia and the Western Interior (North America). Palaeogeogr Palaeoclimatol Palaeoecol 282:88–104
Moore EJ (1984) Molluscan paleontology and biostratigraphy of the lower Miocene upper part of the lincoln creek formation in southwestern Washington. Contr Sci Nat Hist Mus Los Angeles City 351:1–39
Moriya K (2015) Isotope signature of ammonoid shells. This volume
Moriya K, Nishi H, Kawahata H, Tanabe K, Takayanagi Y (2003) Demersal habitat of Late Cretaceous ammonoids: evidence from oxygen isotopes for the Campanian (Late Cretaceous) northwestern Pacific thermal structure. Geology 31:167–170
Morris KA (1979) A classification of Jurassic marine shale sequences: an example from the Toarcian (Lower Jurassic) of Great Britain. Palaeogeogr Palaeoclimatol Palaeoecol 26:117–126
Morris KA (1980) Comparison of major regions of organic–rich mud deposition in the British Jurassic. J Geol Soc Lond 137:157–170
Morton N, Nixon M (1987) Size and function of ammonoid aptychi in comparison with buccal masses in modern cephalopods. Lethaia 20:231–238
Mouterde R, Elmi S (1991) Caractères différentiels des faunes d’ammonites du Toarcien des bordures de la Téthys. Signification paléogéographique. Bull Soc Géol Fr 162:1185–1195
Müller AH (1969) Ammoniten mit ‘Eierbeutel’ und die Frage nach dem Sexual-Dimorphismus der Ceratiten (Cephalopoda). Mber Dt Akad Wiss Berlin 11:411–420
Mutvei H (1975) The mode of life in ammonoids. Paläontol Z 49:196–202
Mutvei H, Dunca E (2007) Connecting ring ultrastructure in the Jurassic ammonoid Quenstedtoceras with discussion on mode of life of ammonoids. In: Landman NH, Davis RA, Mapes RH (eds) Cephalopods Present and Past: new insights and fresh perspectives. Springer, Dortrecht
Naglik C, Tajika A, Chamberlain JA, Klug C (2015) Ammonoid locomotion. This volume
Navarro N, Neige P, Marchand D (2005) Faunal invasion as a source of morphological constraints and innovations? The diversification of the early Cardioceratidae (Ammonoidea; Middle Jurassic). Palaeobiology 31:98–116
Neige P, Marchand D, Bonnot A (1997) Ammonoid morphological signal versus sea-level changes. Geol Mag 134:261–264
Nesis KN (1986) On the feeding and causes of extinction of certain heteromorph ammonites. Paleontol Zh 1986:8–15 (Engl transl, Paleontol J 20:5–11)
Nesis KN (1987) Cephalopods of the world, squids, cuttlefishes, octopuses and allies. (transl from Russian). TFH Publications, Neptune City, p. 351
Neumayr M (1883) Über Klimatische Zonen während der Jura und Kreidezeit: Denk K Akad Wiss Math-Nat Kl 48:57–142
Niebuhr S, Jochiamski MM (2002) Stable isotope and trace element geochemistry of Upper Cretaceous carbonates and belemnite rostra (Middle Campanian, North Germany). Géobios 35:51–64
Nikolaeva SV (1999) Morphological diversity of ammonoids from the Lower Namutian of Central Asia. In: Olóriz F, Rodríguez-Tovar FJ (eds) Advancing research on living and fossil cephalopods. Plenum, New York
Nixon M (1988) The feeding mechanism and diets of cephalopods-living and fossil. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Nixon M (1996) Morphology of the jaws and radula in ammonoids. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid Paleobiology. Plenum, New York
NOAA National Oceanic and Atmospheric Administration (2014) National oceanographic data center (NODC). United States department of commerce. http://www.nodc.noaa.gov. Accessed 12 April 2014
Nützel A, Frýda, J (2003) Paleozoic plankton revolution: Evidence from early gastropod ontogeny. Geology 31:829–831
Oba T, Kai M, Tanabe K (1992) Early life history and habitat of Nautilus pompilius inferred from oxygen isotope examinations. Mar Bio 113:211–217
Obata I, Futakami M (1977) The Cretaceous sequence of the Manji Dome. Palaeontol Soc Jpn Spec Pap 21:23–30
O'Dor RK, Wells MJ (1990) Performance limits of ‘antique’ and ‘state-of-the-art’ cephalopods, Nautilus and squid. Am Mahcologicnl Union Program Abstr Annu Meet 56:52
Okamoto T (1984) Theoretical morphology of Nipponites (a heteromorph ammonite). Fossils (Kaseki) Palaeont Soc Jpn 36:37–51
Okamoto T (1988a) Analysis of heteromorph ammonoids by differential geometry. Palaeontology 31:35–52
Okamoto T (1988b) Changes in life orientation during the ontogeny of some heteromorph ammonoids. Palaeontology 31:281–294
Okamoto T (1988c) Developmental regulation and morphological saltation in the heteromorph ammonite Nipponites. Paleobiology 14:273–286
Okamoto T, Shibata M (1997) A cyclic mode of shell growth and its implications in a Late Cretaceous heteromorph ammonite Polyptychoceras pseudogaultinum (Yokoyama). Palaeontol Res 1:29–46
Olivero EB, Zinsmeister WJ (1989) Large heteromorph ammonites from the Upper Cretaceous of Seymour Island, Antarctica. J Paleontol 63:626–635
Olóriz E (1976) Kimmeridgiano-Tlithonico inferior en el sector central de las Cordilleras Béticas (Zona Subbética). Paleontologia, Bioestratigrafía [PhD thesis]. Universidad de Granada, Granada
Olóriz F, Palmqvist P (1995) Sutural complexity and bathymetry in ammonites: fact or artefact? Lethaia 28:167–170
Olóriz F, Rodríguez-Tovar FJ (eds) (1999) Advancing Research on Living and Fossil Cephalopods. Kluwer Academic, New York
Olóriz E, Marques B, Rodriguez-Tovar FJ (1991) Eustatism and faunal associations. examples from the south Iberian margin during the Late Jurassic (Oxfordian-Kimmeridgian). Eclogae Geol Helv 84:83–106
Olóriz, F, Caracuel, JE, Ruiz-Heras JJ, Rodríguez-Tovar FJ, Marques B (1996) Ecostratigraphic approaches, sequence stratigraphy proposals and block tectonics: examples from epioceanic swell areas in south and east Iberia. Palaeogeogr Palaeoclimat Palaeoecol 121:273–295
Olóriz F, Palmqvist P, Pérez-Claros A (1999) Recent advances in morphometric approaches to covariation of shell features and the complexity of suture lines in Late Jurassic ammonites, with reference to the major environments colonized. In: Olóriz F, Rodríguez-Tovar FJ (eds) Advancing Research on Living and Fossil Cephalopods. Kluwer Academic, New York,
Oschmann W (1991) Distribution, dynamics and palaeontology of Kimmeridgian (Upper Jurassic) shelf anoxia in western Europe. In: Tyson RV, Pearson TH (eds) Modern and ancient continental shelf anoxia. Geol Soc Spec Pap 58:381–395
Packard, A (1972) Cephalopods and fish: the limits of convergence. Biol Rev 47:241–307
Palmer TJ (1982) Cambrian to Cretaceous changes in hardground communities. Lethaia 15:309–323
Pamenter CB (1956) Imitoceras from the Exshaw Formation of Alberta. J Paleont 30:965–966
Payne AIL, Lipiński MR, Clarke MR, Roeleveld MAC (1998) Cephalopod biodiversity, ecology and evolution. South American Journal of Marine Science 20. Sea Fisheries Department of Environmental Affairs and Tourism, Cape Town, p 469
Podlaha OG, Mutterlose J, Veizer J (1998) Preservation of δ18O and δ13C in belemnite rostra from the Jurassic/Early Cretaceous successions. Am J Sci 298:324–347
Price GD, Mutterlose J (2004) Isotopic signals from late Jurassic-early Cretaceous (Volgian-Valanginian) sub-Arctic belemnites, Yatra River, Western Siberia. J Geol Soc Lond 161:959–968.
Price GD, Ruffel AH, Jones CE, Kalin RM, Mutterlose J (2000) Isotopic evidence for temperature variation during the Early Cretaceous (late Ryazanian-mid Hauterivian). J Geol Soc Lond 157:335–343
Price GD, Twitchett RJ, Smale Ch, Marks V (2009) Isotopic analysis of the life history of the enigmatic squid Spirula spirula, with implications for studies of fossil cephalopods. Palaios 24:273–279
Price GD, Fözy I, Janssen NMM, Pálfy (2011) Late Valanginian-Barremian (Early Cretaceous) palaeotemperatures inferred from belemnite stable isotope and Mg/Ca ratios from Bersek Quarry (Gerecse Mountains, Transdanubian Range, Hungary). Palaeogeogr Palaeoclimatol Palaeoecol 305:1–9
Pucéat E, Lecuyer C, Sheppard SMF, Dromart G, Reboulet S, Grandjean P (2003) Thermal evolution of Cretaceous Tethyan marine waters inferred from oxygen isotope composition of fish tooth enamels. Paleoceanography 18:7–12
Radwansky A (1996) The predation upon, and the extinction of, the latest Maastrichtian populations of the ammonite species Hoploscaophites constrictus (J. Sowerby, 1817) from the Middle Vistula Valley, Central Poland. Acta Geol Pol 46:117–135
Railsback BL, Anderson TF, Ackerly SC, Cisne JL (1989) Paleoceanographic modeling of temperature-salinity profiles from stable isotopic data. Paleoceanography 4(5):585–591
Rasmussen K, Brett C (1985) Taphonomy of Holocene cryptic biotas from St. Croix, Virgin Islands: information loss and preservational biases. Geology 13:551–553
Rawson PF (1981) Early Cretaceous ammonite biostratigraphy and biogeography. In: House MR, Senior JR (eds) The Ammonoidea. Systematics Association, special vol 18, Academic Press, London
Rawson PF (1993) The influence of sea level changes on the migration and evolution of early Cretaceous (pre-Aptian) ammonites. In: House MR (ed) The Ammonoidea: Environment, Ecology, and Evolutionary Change. Systematics Association, special vol 47. Oxford Science Publications, Oxford
Reboulet S (1996) L’évolution des ammonites du Valanginien–Hauterivien inférieur du bassin vocontien et de la plate-forme provençale (Sud-Est de la France): relations avec la stratigraphie séquentielle et implications biostratigraphique. Doc Labor Géol Lyon 137:371
Reboulet S (1998) Diversification des ammonites hétéromorphes. In: Gayet M, Otero O (eds) Paleodiversifications, Land and Sea Compared: International Symposium, Lyon, France (July 6–8, 1998), Abstr Vol, p. 54
Reboulet S (2001) Limiting factors on shell growth, mode of life and segregation of Valanginian ammonoid populations: evidence from adult-size variations. Geobios 34:423–435
Reboulet S, Atrops F (1995) Rôle du climat sur les migrations et la composition des peuplements d’ammonites du Valanginien supérieur du bassin vocontien (S-E de la France). Geobios Mém Spéc 18:357–365
Reboulet S, Atrops F (1997) Quantitative variations of the Valanginian ammonite fauna of the Vocontian Basin (southeastern France) between limestone-marls and within parasequence sets. Palaeogeogr Palaeoclimat Palaeoecol 135:145–155
Reboulet S, Rard A (2008) Double alignments of ammonoid aptychi from the Lower Cretaceous of Southeast France: Result of a post–mortem transport or bromalites? Acta Palaeontol Pol 53:261–274
Reboulet, S, Atrops F, Ferry S, Schaaf A (1992) Renouvellement des ammonites en fosse vocontienne à la limite Valanginien-Hauterivien. Geobios 25:469–476
Reboulet S, Proux O, Giraud F, Baudin F, Olivero D, Aucour AM (2000) Characterization and significance of ammonoid and nannoplankton assemblages during an ‘‘anoxic’’ event: the Breistroffer level (Upper Albian, SE France). In: Summesberger H, Kollmann H (eds) 6th International Cretaceous Symposium, Vienna, Austria (August 27–September 4), Abstr vol, p 111
Reboulet S, Mattioli, E, Pittet B, Baudin F, Olivero D, Proux O (2003) Ammonoid and nannoplankton abundance in Valanginian (early Cretaceous) limestone-marl successions from the southeast France Basin: carbonate dilution or productivity? Palaeogeogr Palaeoclimat Palaeoecol 201:113–139
Reboulet S, Giraud F, Proux O (2005) Ammonoid abundance variations related to changes in trophic conditions across the Oceanic Anoxic Event 1d (Latest Albian, SE France). Palaios 20:121–141
Reeside JB, Cobban WA (1960) Studies of the Mowry Shale (Cretaceous) and contemporary formations in the United States and Canada. US Geol Surv Prof Pap 355:1–126
Renz O (1972) Aptychi (Ammonoidea) from the Upper Jurassic and Lower Cretaceous of the western North Atlantic (site 105, leg 11, DSDP). In: Holister CD, Ewing JI et al (eds) Initial reports DSDP, No. 11, US Government Printing Office, Washington DC
Renz O (1973) Two lamellaptychi (Ammonoidea) from the Magellan Rise in the central Pacific. In: Winterer EL, Hewing JL (eds) Initial reports DSDP, No 17, US Government Printing Office, Washington DC
Renz O (1978) Aptychi (Ammonoidea) from the early cretaceous of the Blake–Bahama Basin, leg 44, hole 391c, DSDP. In: Benson WE, Sheridan RE (eds) Initial reports DSDP, No. 44, US Government Printing Office, Washington DC
Renz O (1979) Aptychi (Ammonoidea) and ammonites from the Lower Cretaceous of the western Bermuda Rise, leg 43, site 387, DSDP. In: Tucholke BE, Vogt PR (eds) Initial reports DSDP, No. 43, US Government Printing Office, Washington DC
Rexfort A, Mutterlose J (2006) Stable isotope records from Sepia officinalis—a key to understand the ecology of belemnites? Earth Planet Sci Lett 247:212–221
Reyment RA (1958) Some factors in the distribution of fossil cephalopods. Stock Contr Geol 1:97–184
Reyment RA (1973) Factors in the distribution of fossil cephalopods. Part 3: experiments wth exact models of certain shell types. Bull Geol Inst Univ Uppsala 4(2):7–41
Reyment RA (1980) Floating orientations of cephalopod shell models. Palaeontology 23:931–936.
Reyment RA (1988) A foraging model for shelled cephalopods. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Reyment RA (2008) Reyment, Richard A. 2008. A review of the post-mortem dispersal of cephalopod shells. Palaeontol Electr 11:12A:13p. http://palaeo-electronica.org/2008_3/148/index.html
Riccardi AC, Westermann GEG (1991) Middle Jurassic ammonite fauna of the Argentine-Chilean Andes, III: Bajocian-Callovian Eurycephalitinae, Stephanocerataceae. Palaeontogr A 216:1–110
Riccardi AC, Gulisano CA, Mojica J, Palacios O, Schubert C, Thomson MRA (1992) Western South America and Antarctica. In: Westermann GEG (ed) The Jurassic of the Circum-Pacific. Cambridge University Press, New York
Richter AE (2009) Ammoniten-Gehäuse mit Bissspuren. Berl Paläontol Abh 10:297–305
Richter C, Wunsch M (1999) Cavity-dwelling suspension feeders in coral reefs-a new link in reef trophodynamics. Mar Ecol Prog Ser 188:105–116
Rieber H (1973) Ergebnisse paläontologisch-stratigraphischer Untersuchungen in der Grenzbitumenzone (Mittlere Trias) des Monte San Giorgio. Eclogae Geol Helv 66:667–685
Rieber H (1975) Der Posidonienschiefer (Oberer Lias) von Holzmaden und die Grenzbitumenzone (Mittlere Trias) des Monte San Giorgio (Kanton Tessin, Schweiz). Jahrb Ges Naturkd Württemb 130:163–190
Rieber H (1977) Eine Ammonitenfauna aus der oberen Maiolica der Breggia-Schlucht (Tessin/Schweiz). Eclogae Geol Helv 70:777–787
Ritterbush KA, Bottjer DJ (2012) Westermann Morphospace displays ammonoid shell shape and hypothetical paleoecology. Paleobiology 38:424–446
Ritterbush KA, Lukeneder A, Hoffmann R, De Baets K (2014) Pelagic palaeoecology: the importance of recent constraints on ammonoid palaeobiology and life history. J Zool 292:229–241
Roetzel R, Mandic O, Steininger FF (1999) Lithostratigraphie und Chronostratigraphie der tertiären Sedimente im westlichen Weinviertel und angrenzenden Waldviertel. Arb Geol BA 1999:38–54
Röhl H, Schmid-Röhl A, Oschmann W, Frimmel A, Schwark L (2001) The Posidonia Shale (Lower Toarcian) of SW-Germany: an oxygen depleted ecosystem controlled by sea level and palaeoclimate. Palaeogeogr Palaeoclimat Palaeoeco 165:27–52
Romanek CS, Jones DS, Williams DF, Krantz DE, Radtke R (1987) Stable isotopic investigation of physiological and enviromnental changes recorded in shell carbonate from the giant clam Tridacna maxima. Mar Biol 94:385–393
Rooij D van, Mol L de, Guilloux E le, Wisshak M, Huvenne VAI, Moeremans R, Henriet JP (2010) Environmental setting of deep-water oysters in the Bay of Biscay. Deep Sea Res Part I: Oceanogr Res Pap 57:1561–1572
Roux M (1990) Underwater observations of Nautilus macromphalus off New Caledonia. Chamb Nautil Newsl 60:1
Roux M, Bouchet P, Bourseau J-P, Gaillard C, Grandperrin R, Guille A, Laurin B, Monniot C, Richer de Forges B, Rio M, Segonzac M, Vacelet J, Zibrowius H (1991) L’environnement bathyal au large de la Nouvelle-Calédonie: résultats préliminaires de la campagne CALSUB et conséquences paléoécologiques. Bull Soc Géol Fr 162:675–685
Sandoval J (1983) Bioestratigrafia y paleontologia (Stephanocerataceae y Perisphinctaceae) del Bajocense y Bathoniense en las Cordilleras Beticas [PhD thesis]. Universidad de Granada, Granada, p. 613
Sarti C (1986a) Faune e biostratigrafia del Rosso Ammonitico del Trentino centrale (Kimmeridgiano- Titoniano). Boll Soc Paleontol Ital 23:473–514
Sarti C (1986b) Considerazioni sul Rosso Ammonitico Veronese del Col Santino (M. Pasubio) e raffronti con altre successioni del Trentino. In: Pallini G (ed) Atti I Conv Int Fossili Evoluzione Ambiente. Pergola, ottobre 1984
Sarti C (1999) Whorl width in the body chamber of ammonites as a sign of dimorphism. In: Olóriz F, Rodríguez–Tovar FJ (eds) Advancing Research on Living and Fossil Cephalopods. Kluwer Academic, New York
Sato T, Tanabe K (1998) Cretaceous plesiosaurs ate ammonites. Nature 394:629–630
Saunders WB (1983) Natural rates of growth and longevity of Nautilus belauensis. Paleobiology 9:280–288
Saunders WB (1984) Nautilus belauensis growth and longevity: evidence from marked and recaptured animals. Science 224:990–992
Saunders WB (1995) The ammonoid suture problem: relationships between shell septum thickness and suture complexity in Paleozoic ammonoids. Paleobiology 21:343–355
Saunders WB, Landman NH (eds) (1987) Nautilus. The Biology and Paleobiology of a Living Fossil. Topics in Geobiology 6, Springer Press, New York
Saunders WB, Landman NH (eds) (2010) ditto (reprinted from 1987 with additions)
Saunders WB, Shapiro EA (1986) Calculation and simulation of ammonoid hydrostatics. Paleobiology 12:64–79
Saunders WB, Spinosa, C (1979) Nautilus movement and distribution in Palau, Western Caroline Islands. Science 204:1199–1201
Saunders WB, Swan RH (1984) Morphology and morphologic diversity of mid-Carboniferous (Namurian) ammonoids in time and space. Paleobiol 10:195–228
Saunders WB, Spinosa C, Davies LE (1987) Predation on Nautilus. In: Saunders WB, Landman NH (eds) (2010) Nautilus. The Biology and Paleobiology of a Living Fossil. Topics in Geobiology 6, Springer Press, New York (reprinted from Saunders WB, Landman NH (eds) 1987 with additions), pp. 201–212
Savrda CE, Bottjer DJ (1991) Oxygen-related biofacies in marine strata: an overview and update. In: Tyson RV, Pearson TH (eds) Modern and ancient continental shelf anoxia. Geol Soc Spec Pap 58:201–219
Schindewolf OH (1934) Über Epöken auf Cephalopoden-Gehäusen. Paläontol Z 16:15–31
Schindewolf OH (1958) Über Aptychen (Ammonoidea). Palaeontogr A 111:1–46
Schindewolf OH (1959) Adolescent cephalopods from the Exshaw Formation of Alberta. J Paleontol 33:971–976
Schlögl J, Chirat R, Balter V, Joachimski M, Hudáčková N, Quillévéré F (2011) Aturia from the Miocene Paratethys: an exceptional window on nautilid habitat and lifestyle. Palaegeogr Palaeoclimat Palaeoecol 308:330–338
Schmid-Röhl A, Röhl H-J (2003) Overgrowth on ammonite conchs: environmental implications for the Lower Toarcian Posidonia Shale. Palaeontology 46:339–352
Schmidt-Effing R (1972) Die Dactylioceratidae, eine Ammoniten-Familie des unteren Jura. Münst Forsch Geol Paläontol 25/26:l–254
Scott G (1940) Paleoecological factors controlling the distribution and mode of life of Cretaceous ammonoids in the Texas area. J Paleontol 14:299–323
Seilacher A (1960) Epizoans as a key to ammonoid ecology. J Paleontol 34:189–193
Seilacher A (1963) Umlagerung und Rolltransport von Cephalopoden-Gehäusen. Neues Jahrb Geol Paläontol Monatshefte 1963:593–615
Seilacher A (1982) Ammonites as habitats in the Posidonia Shale—floats or benthic islands? Neues Jahrb Geol Paläontol Monatshefte 1982:98–114
Seilacher A (1993) Ammonite aptychi: how to transform a jaw into an operculum. Am J Sci 293A:20–32
Seilacher A, Analib F, Dietl G, Gocht H (1976) Preservational history of compressed ammonites from southern Germany. Neues Jahrb Geol Paläontol Abh 152:307–356
Seuss B, Titschack J, Seifert S, Neubauer J, Nützel A (2012) Oxygen and stable carbon isotopes from a nautiloid from the middle Pennsylvanian (Late Carboniferous) impregnation Lagerstätte ‘Buckhorn Asphalt Quarry’—Primary paleo-environmental signals versus diagenesis. Palaeogeogr Palaeoclimat Palaeoecol 319/320:1–15
Shapiro EA, Saunders WB (1987) Nautilus shell hydrostatics. In: Saunders WB, Landman NH (eds) Nautilus. Plenum, New York
Shigeta Y (1993) Post–hatching early life history of Cretaceous Ammonoidea. Lethaia 26:23–46
Shimansky VN (1975) Cretaceous nautiloids. Acad Sci USSR Trans Palaeontol Inst 150:1–288 [In Russian]
Sornay J (1955) Ammonites nouvelles du Crétacé de la région des Monts du Mellègue (Constantine). Bull Serv Carte Geol Algér 1 ser Paléontol 18:1–40
Spaeth C, Hoefs J, Vetter U (1971) Some aspects of isotope composition of belemnites and related paleotemperature. Geol Soc Amer Bull 82:3139–3150
Spinosa PL, Furnish WM, Glenister GE (1975) The Xenodiscidae, Permian ceratitoid ammonoids. J Paleontol 49:239–283
Sprey AM (2002) Early ontogeny of three Callovian ammonite genera (Binatisphinctes, Kosmoceras (Spinikosmoceras) and Hecticoceras) from Ryazan (Russis). In: Summesberger H, Histon K, Daurer A (2002) Cephalopods Present and Past. Abh Geol B-A 57:225–255
Stahl W, Jordan R (1969) General considerations on isotopic paleotemperature determinations and analysis on Jurassic ammonites. Earth Planet Sci Lett 6:173–178
Stampfli GM, Borel GD (2002) A plate tectonic model for the Paleozoic and Mesozoic constrained by dynamic plate boundaries and restored synthetic oceanic isochrons. Earth Planet Sci Lett 196:17–33
Stephen DA, Stanton RJ (2002) Impact of reproductive strategy on cephalopod evolution. In: Summesberger H, Histon K, Daurer A (eds) Cephalopods Present and Past. Abh Geol B-A 57:151–155
Stephen DA, Bylund KG, Garcia P, McShinsky RD, Carter HJ (2012) Taphonomy of dense concentrations of juvenile ammonoids in the Upper Cretaceous Mancos Shale, east-central Utah, USA. Geobios 45:121–128
Stevens GR (1988) Giant ammonites: a review. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Stevens GR (1997) The Late Jurassic ammonite fauna of New Zealand. Institute of Geological and Nuclear Sciences monograph 18. N Z Geol Surv Paleontol Bull 74:1–217
Stevens K, Mutterlose J, Wiedenroth (2015) Stable isotope data (δ18O, δ13C) of the ammonite genus Simbirskites—implications for habitat reconstructions of extinct cephalopods. Palaeogeogr Palaeoclimatol Palaeoecol 417:164–175
Stevens K, Mutterlose J, Wiedenroth (2015) Stable isotope data (δ18O, δ13C) of the ammonite genus Simbirskites—implications for habitat reconstructions of extinct cephalopods. Palaeogeogr Palaeoclimatol Palaeoecol 417:164–175
Stewart JD, Carpenter K (1990) Examples of vertebrate predation on cephalopods in the Late Cretaceous of the Western Interior. In: Boucot AJ (ed) Evolutionary paleobiology of behavior and coevolution. Elsevier, Amsterdam
Stewart JD, Carpenter K (1999) Examples of vertebrate predation on cephalopods in the Late Cretaceous of the Western Interior. Bull S Cal Paleontol Soc 31:66–73
Stilwell JD, Henderson RA (2002) Description and paleobiogeographic significance of a rare Cenomanian molluscan faunule from Bathurst Island, northern Australia. J Paleontol 76:447–471
Summesberger H (2000) Ammoniten als Kannibalen. Das Naturhistorische 1, p. 10
Summesberger H, Jurkovsek B, Kolar-Jurkovsek T (1996) Aptychi associated with ammonites from the lipica-formation (Upper Cretaceous, Slovenia). Ann Naturhist Mus Wien 97A:1–19
Summesberger H, Histon K, Daurer A (2002) Cephalopods Cephalopods Present and Past. Abh Geol B-A 57:1–569
Swan RH, Saunders WB (1987) Function and shape in Late Paleozoic (mid-Carboniferous) ammonoids. Paleobiology 13:297–311
Tajika A, Wani R (2011) Intraspecific variation of hatchling size in Late Cretaceous ammonoids from Hokkaido, Japan: implication for planktic duration at early ontogenetic stage. Lethaia 44: 287–298
Tajika A, Naglik C, Morimoto N, Pascual-Cebrian E, Hennhöfer DK, Klug C (2015): Empirical 3D-model of the conch of the Middle Jurassic ammonite microconch Normannites, its buoyancy, the physical effects of its mature modifications and speculations on their function. Hist Biol, 11 pp. doi:10.1080/08912963.2013.872097
Tanabe K (1977) Functional evolution of Otoscaphites puerculus (Jimbo) and Scaphites planus (Yabe), Upper Cretaceous ammonites. Mem Fac Sci Kyushu Univ Ser D Geol 23:367–407
Tanabe K (1979) Palaeoecological analysis of ammonoid assemblages in the Turonian Scaphites facies of Hokkaido, Japan. Palaeontology 22:609–630
Tanabe K (1983) The jaw apparatus of Cretaceous desmoceratid ammonites. Palaeontology 26:677–689
Tanabe K, Fukuda Y (1987) The jaw apparatus of the Cretaceous ammonite Reesidites. Lethaia 20:41–48
Tanabe K, Landman NH (1996) Septal neck-siphuncular complex of ammonoids. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid Paleobiology. Plenum, New York
Tanabe K, Ohtsuka Y (1985) Ammonoid early internal shell structure: Its bearing on early life history. Paleobiology 11:310–322
Tanabe K, Shigeta Y (1987) Ontogenetic shell variation and streamlining of some Cretaceous ammonites, Trans Proc Palaeonr Soc Jpn N S 147:165–179
Tanabe K, Obata I, Futakami M (1978) Analysis of ammonoid assemblages in the Upper Turonian of the Manji area, central Hokkaido. Bull Nat Sci Mus Tokyo C 4:37–62
Tanabe K, Obata I, Futakami M (1981) Early shell morphology in some Upper Cretaceous heteromorph ammonites. Trans Proc Palaeontol Soc Jpn NS 124:215–234
Tanabe K, Landman NH, Weitschat W (1993a) Septal necks in mesozoic ammonoidea: structure, ontogenetic development and evolution. In: House MR (ed) The Ammonoidea. Environment, Ecology, and Evolutionary Change. Systematic Association special vol 47, Clarendon Press, Oxford
Tanabe K, Landman NH, Mapes, RH, Faulkner CJ (1993b) Analysis of a Carboniferous embryonic ammonoid assemblage-implications for ammonoid embryology. Lethaia 20:215–224
Tanabe K, Landman NH, Mapes RH (1994a) Early shell features of some Late Paleozoic ammonoids and their systematic implications. Trans Proc Palaeontol Soc Jpn NS 173:383–400
Tanabe K, Landman NH, Mapes RH, Faulkner CJ (1994b) Analysis of a Carboniferous embryonic ammonoid assemblage from Kansas. U.S.A.—implications for ammonoid embryology. Lethaia 26:215–224
Tanabe K, Shigeta Y, Mapes, RH (1995) Early life history of Carboniferous ammonoids inferred from analysis of fossil assemblages and shell hydrostatics. Palaios 10:80–86
Tanabe K, Landman NH, Yoshioka Y (2003) Intra- and interspecific variations in the early internal shell features of some Cretaceous ammonoids. J Paleontol 77:876–887
Tanabe K, Shigeta Y, Sasaki T, Hirano H (eds) (2010a) Cephalopods Present and Past. Tokai University Press, Tokyo
Tanabe K, Kulicki C, Landman NH, Kaim A (2010b) Tuberculate micro-ornamentation on embryonic shells of Mesozoic ammonoids: microstructure, taxonomic variation, and morphogenesis In: Tanabe K, Shigeta Y, Sasaki T, Hirano H (eds) Cephalopods Present and Past. Tokai University Press, Tokyo
Tanaka N, Monaghan MC, Rye DM (1986) Contribution of metabolic carbon to mollusc and barnacle shell carbonate. Nature 320:520–523
Tarutani T, Clayton RN, Mayeda TK (1969) The effects of polymorphism and magnesium substitution on oxygen isotope fractionation between calcium carbonate and water. Geochim Cosmochim Acta 33:987–996
Taylor BE, Ward PD (1983) Isotopic studies of Nautilus macromphalus Sowerby (New Caledonia) and Nautilus pompilius L. (Fiji). Palaeogeogr Palaeoclimatol Palaeoecol 41:1–16
Teichert C, Matsumoto T (1987) The ancestry of the genus Nautilus. In: Saunders WB, Landmann NH (eds) Nautilus: The Biology and Paleobiology of a Living Fossil. Topics in Geobiology 6, Springer, New York
Thieuloy JP (1966) Leptocères berriasiens du massif de la Grande-Chartreuse. Trav Lab Géol Fac Sci Univ Grenoble 42:281–295
Tichy G, Urbanek E (2004) Biss-Spuren eines Sauriers an Pinacoceras parma Mojsisovics, ein Ammonit der Halleiner Obetrias. GeoAlp 1:87–90
Tintant H, Marchand D, Mouterde R (1982) Relations entre les milieux marins et l’evolution des Ammonoides: Les radiations adaptives du Lias. Bull Soc Géol Fr 24:951–961
Tittensor DP, Rex MA, Stuart CT, McClain CR, Smith CR (2011) Species-energy relationships in deep-sea molluscs. Biol Letters 23:718–722
Toriyama R, Sato T, Hamada T, Komalarjun P (1964) Nautilus pompilius drifts on the west coast of Thailand. Jap J Geol Geogr 36: 149–161
Tourtelot HA, Rye RO (1969) Distribution of oxygen and carbone isotopes in the fossil of Late Cretecaous age, Western Interior region of North America. Geol Soc Am Bull 80:1903–1922
Trueman AE (1941) The ammonoid body chamber with special reference to the buoyancy and mode of life of the living ammonite. Quart J Geol Soc Lond 96: 339–383
Tsujita CJ, Westermann GEG (1998) Ammonoid habitats and habits in the Western Interior Seaway: a case study from the Upper Cretaceous Bearpaw Formation of southern Alberta, Canada. Palaeogeogr Palaeoclimat Palaeoecol 144:135–160
Tsujita CJ, Westermann GEG (2001) Were limpets or mosasaurs responsible for the perforations in the ammonite Placenticeras? Palaeogeogr Palaeoclimatol Palaeoecol 169:245–270
Tyson RV, Pearson TH (1991) Modern and ancient continental shelf anoxia: an overview. In: Tyson RV, Pearson TH (eds) Modern and ancient continental shelf anoxia. Geol Soc Spec Pub 58:l–26
Uchiyama K, Tanabe K (1999) Hatching of Nautilus macromphalus in the Toba aquarium, Japan. In: Olóriz F, Rodríguez-Tovar FJ (eds) Advancing Research on Living and Fossil Cephalopods. Kluwer Academic, New York
Uhlig V (1911) Über die sogen. borealen Typen des südandinen Reiches. Cent Mineral Geol Paläontol 15, 16, 17:483–490, 517–522, 536–548
Urey HC (1947) The thermodynamic properties of isotopic substances. J Chem Soc 1947:562–581
Urey HC, Lowenstam HA, Epstein S, McKinney CR (1951) Measurements of paleotemperatures and temperatures of the Upper Cretaceous, Denmark, and southeastern United States. GSA Bull 62:399–416
Urlichs M, Mundlos R (1985) Immigration of cephalopods into the German Muschelkalk basin and its influence on the suture lines. In: Bayer U, Seilacher A (eds) Sedimentary and Evolutionary Cycles. Lecture Notes in Earth Sciences. Springer, Berlin
Urlichs M, Wild R, Ziegler B (1979) Fossilien aus Holzmaden. Stuttg Beitr Naturkunde Ser C 11:1–34
Vašíček Z, Wiedmann J (1994) The Leptoceratoidinae: small heteromorph ammonites from the Barremian. Palaeontology 37:203–239
Vermeij GJ (1977) The Mesozoic marine revolution: evidence from snails, predators and grazers. Paleobiology 3:245–258
Wang Y, Westermann GEG (1993) Paleoecology of Triassic ammonoids. Geobios Mem Spec 15:373–392
Wani R (2004) Experimental fragmentation pattersn of modern Nautilus shells and the implications for fossil cephalopod taphonomy. Lethaia 37:113–123
Wani R (2007) How to recognize in situ fossil cephalopods: evidence from experiments with modern Nautilus. Lethaia 40:305–311
Wani R, Gupta NS (2015). Ammonoid taphonomy. In: Klug C, Korn D, De Baets K, Kruta I, Mapes RH (eds) Topics in Geobiology, vol 43. Ammonoid Paleobiology: From macroevolution to paleogeography. Vol 2, Part III - Ammonoids through time. Springer, Dordrecht
Wani R, Kase T, Shigeta Y, De Ocampo R (2005) New look at ammonoid taphonomy, based on field experiments with modern chambered Nautilus. Geology 33:849–852
Wani R, Kurihara K, Ayyasami K (2011) Large hatchling size in Cretaceous nautiloids persists across the end-Cretaceous mass extinction: new data of Hercoglossidae hatchlings. Cretac Res 32:618–622
Ward PD (1976a) Stratigraphy, paleoecology and functional morphology of heteromorph ammonites in the Upper Cretaceous Nanaimo Group, British Columbia and Washington. Dissertation, McMaster University of Hamilton, Ontario.
Ward PD (1980) Comparative shell shape distributions in Jurassic-Cretaceous ammonites and nautilids. Paleobiology 6:32–43
Ward PD (1981) Shell sculpture as a defensive adaptation in ammonoids. Paleobiology 7:96–100.
Ward PD (1986a) Rates and processes of compensatory buoyancy change in Nautilus macromphalus. Veliger 1986:356–368
Ward PD (1986b) Cretaceous ammonite shell shapes. Malacologia 27:3–28
Ward PD (1987) The natural history of Nautilus. Allen and Unwin, Boston
Ward PD (1990a) A review of Maastrichtian ammonite ranges. GSA Spec Pap 247:519–530
Ward PD (1990b) The Cretaceous/Tertiary extinctions in the marine realm: A 1990 perspective. GSA Spec Pap 247:425–432
Ward PD, Bandel K. (1987) Life history strategies in fossil cephalopods. In: life cycles. Academic Press, London
Ward DJ, Hollingworth NTJ (1990) The first record of a bitten ammonite from the Middle Oxford Clay (Callovian, Middle Jurassic) of Bletchley, Buckingshamshire. Mesozoic Res 2:153–161
Ward PD, Signor PW III (1983) Evolutionary tempo in Jurassic and Cretaceous ammonites. Paleobiology 9:183–198
Ward PD, Westermann GEG (1977) First occurrence, systematics and functional morphology of Nipponites from the Americas. J Paleontol 51:367–372
Ward PD, Westermann GEG (1985) Cephalopod paleoecology. In: Bottjer DJ, Hickman CS, Ward PD (org), Broadhead TW (ed) Mollusks, Notes for a short course. Univ Tenn Geol Sci Stud 13:1–18
Ward PD, Carlson B, Weekly M, Brumbaugh B (1984) Remote telemetry of daily vertical and horizontal movement of Nautilus in Palau. Nature 309:248–250
Warnke K, Keupp H (2005) Spirula—A window to the embryonic developments of ammonoids? Morphological and molecular indications for a palaeontological hypothesis. Facies 51:60–65
Warnke K, Plötner J, Santana JI, Rueda MJ, Llinas O (2003) Reflections on the phylogenetic position of Spirula (Cephalopoda): preliminary evidence from the 18S ribosomal RNA gene. Berl Paläobiol Abh 3:253–260
Warnke K, Oppelt A, Hoffmann R (2010) Stable isotopes during ontogeny of Spirula and derived hatching temperatures. Ferrantia 59:191–201
Watanabe T, Gagan MK, Correge T, Scott-Gagan H, Cowley J, Hantoro WS (2003) Oxygen isotope systematics in Diploastrea heliopora: new coral archive of tropical paleoclimate. Geochim Cosmochim Acta 67:1349–1358
Wefer G (1985) Die Verteilung stabiler Isotope in Kalkschalen mariner Organismen. Geol Jahrb 82:3–111.
Wefer G, Berger, WH (1991) Isotope paleontology: growth and composition of extant calcareous species. Mar Geol 100:207–248
Wells MJ (1983) Cephalopods do it different. New Sci 100:332–338
Wells MJ, Wells J, O’Dor RK (1992) Life at low oxygen tensions: the behaviour and physiology of Nautilus pompilius and the biology of extinct forms. J Mar Biol Assoc UK 72:313–328
Wendt J (1963) Stratigraphisch-paläontologische Untersuchungen im Dogger Westsiziliens. Boll Soc Paleontol Ital 2:57–145
Westermann GEG (1954) Monographie der Otoitidae (Ammonoidea). Geol Jahrb Beih 15:1–364.
Westermann GEG (1958) The significance of septa and sutures in Jurassic ammonite systematics. Geol Mag 45:441–455
Westermann GEG (1971) Form, structure and function of shell and siphuncle in coiled Mesozoic ammonoids. Life Sci Contrib R Ont Mus 78:1–39
Westermann GEG (1973) Strength of concave septa and depth limits of fossil cephalopods. Lethaia 6:373–403
Westermann GEG (1975a) Architecture and buoyancy of simple cephalopod phragmocones and remarks on ammonites. Paläontol Z 49:221–234
Westermann GEG (1975b) A model for origin, function and fabrication of fluted cephalopod septa. Paläontol Z 49:235–253
Westermann GEG (1977) Form and function of orthoconic cephalopod shells with concave septa. Paleobiology 3:300–321
Westermann GEG (1982) The connecting rings of Nautilus and Mesozoic ammonoids: Implications for ammonoid bathymetry. Lethaia 15:374–384
Westermann GEG (1989) New developments in ecology of Jurassic-Cretaceous ammonoids, in Pallini, G., Cecca, F., and Cresta, S., (eds), Fossili, evoluzione, ambiente, Att II Conventione Internationale Pergola 1987: Otra Vetere, Italy, Tecnostampa
Westermann GEG (1990) New developments in ecology of Jurassic-Cretaceous ammonoids. In: Pallini G, Cecca E, Cresta S, Santantonio M (eds) Fossili, Evolutione, Ambiente. Atti II Conv Int F E A Pergola 1987. Com Cent Raff Piccinini, Ostra Vetere
Westermann GEG (1993a) On alleged negative buoyancy in ammonoids. Lethaia 26:246
Westermann GEG (1993b) Global bio-events in mid-Jurassic ammonites controlled by seaways. In: House MR (ed) The Ammonoidea. Environment, Ecology and Evolutionary Change. Systematics Association, special volume 47, Clarendon Press, Oxford
Westermann GEG (1996) Ammonoid life and habitat. In: Landman NH, Tanabe K, Davis RA (eds) Ammonoid Paleobiology. Plenum, New York
Westermann GEG, Callomon JH (1988) The Macrocephalitinae and associated Bathonian and early Callovian (Jurassic) ammonoids of the Sula lslands and New Guinea. Palaeontogr A 203:l–90
Westermann GEG, Hewitt RA (1995) Do limpet pits indicate that desmoceratacean ammonites lived mainly in surface waters? Lethaia 28:24
Westermann GEG, Ward P (1980) Septum morphology and bathymetry in cephalopods. Paleobiol 6:48–50
Whittaker SG, Kyser TK, Caldwell GE (1987) Paleoenvironmental geochemistry of the Claggett marine Cyclothem in south-central Saskatchewan. Can J Earth Sci 24:967–984
Wiedmann J (1973) Evolution or revolution of ammonoids at Mesozoic system boundaries. Biol Rev 48:159–194
Wiedmann J (1988a) Plate tectonics, sea level changes, climate and the relationship to ammonite evolution, provincialism, and mode of life. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Wiedmann J (1988b) Ammonite extinction and the Cretaceous Tertiary boundary event. In: Wiedmann J, Kullmann J (eds) Cephalopods Present and Past. Schweizerbart, Stuttgart
Wiedmann J, Kullmann J (eds) (1988) Cephalopods Present and Past. Schweizerbart, Stuttgart
Wignall PB (1987) A biofacies analysis of the Gastrioceras cumbriense Marine Band (Namurian) of the Central Pennines. Proc York Geol Soc 46:111–121
Wignall PB (1990) Observations on the evolution and classification of dysaerobic communities. In: Miller W (ed) Paleocommunity Temporal Dynamics: The Long-term Development of Multispecies Assemblies. Paleontol Soc Spec Publ 5:99–111
Wignall PB, Hallam A (1991) Biofacies, stratigraphic distribution and depositional models of British onshore Jurassic black shales. In: Tyson RV, Pearson TH (eds) Modern and ancient continental shelf anoxia. Geol Soc Spec Pap 58:291–309
Wignall PB, Hallam A (1993) Griesbachian (earliest Triassic) palaeoenvironmental changes in the Salt Range, Pakistan and southeast China and their bearing on the Permo-Triassic mass extinction. Palaeogeogr Palaeoclimatol Palaeoecol 102:215–237
Wignall PB, Simms MJ (1990) Pseudoplankton. Palaeontology 33:359–378
Wilde P, Berry WBN (1984) Destabilization of the oceanic density structure and its significance to marine "extinction" events. Palaeogeogr Palaeoclimatol Palaeoecol 48:143–162
Wilmsen M, Mosavinia A (2011) Phenotypic plasticity and taxonomy of Schloenbachia varians (J. Sowerby, 1817). Paläontol Z 85:169–184
Wippich MGE, Lehmann J (2004) Allocrioceras from the Cenomanian (Mid-Cretaceous) of the Lebanon and its bearing on the palaeobiological interpretation of heteromorphic ammonites. Palaeontology 47:1093–1107
Wright CW, Calloman JH, Howarth MK (1996) Treatise on Invertebrate Paleontology, Part L, Mollusca 4 revised (Cretaceous Ammonoidea), GSA, Boulder. University of Kansas Press, Lawrence
Wright EK (1987) Stratification and paleocirculation of the Late Cretaceous Western Interior Seaway of North America. GSA Bull 99:480–490
Wunsch M, Richter C (1998) The CaveCam–an endoscopic underwater videosystem for the exploration of cryptic habitats. Mar Ecol Prog Ser 169:277–282
Yacobucci MM (1999) Plasticity of developmental timing as underlying cause of high speciation rates in ammonoids. In: Olóriz F, Rodríguez-Tovar FJ (eds) Advancing Research on Living and Fossil Cephalopods. Kluwer Academic, Plenum, New York
Yacobucci M (2004) Buckman’s Paradox: variability and constraints on ammonoid ornament and shell shape. Lethaia 37:57–69
Yahada H., Wani R (2013) Limited migration of scaphitid ammonoids: evidence from the analyses of shell whorls. J Paleontol 87(3):406–412
Yomogida S, Wani R (2013) Higher risk of fatality by predatory attacks in earlier ontogenetic stages of modern Nautilus pompilius in the Philippines: evidence from the ontogenetic analyses of shell repairs. Lethaia 46:317–330
Young RE, Vecchione M, Donovan DT (1998) The evolution of coloid cephalopods and their present biodiversity and ecology. In: Payne AIL, Lipinski MR, Clarke MR, Roeleveld MAC (eds) Cephalopod diversity, Ecology and Evolution. Afr J Mar Sci 20:393–420
Zaborski PMP (1982) Campanian and Maastrichtian sphenodiscid ammonites from southern Nigeria. Bull Br Mus Nat Hist (Geol) 36:303–332
Zakharov YD, Boriskina, N.G., Ignatyev AV, Tanabe K, Shigeta Y, Popov AM, Afanasyeva TB, Maeda H (1999) Palaeotemperature curve for the Late Cretaceous of the northwestern circum-Pacific. Cretac Res 20:685–997
Zakharov YD, Smyshlyaeva OP., Tanabe K, Shigeta Y, Maeda H, Ignatiev AV, Velivetskaya TA, Afanasyeva TB, Popov AM, Golozubov VV, Kolyadae AA, Cherbadzhi AK, Moriya K (2005) Seasonal temperature fluctuations in the high northern latitudes during the Cretaceous Period: isotopic evidence from Albian and Coniacian shallow-water invertebrates of the Talovka River Basin, Koryak Upland, Russian Far East. Cretac Res 26:113–132
Zakharov YD, Shigeta Y, Smyshlyaeva OP, Popov AM, Ignatiev AV (2006) Relationship between δ13C and δ18O values of the Recent Nautilus and brachiopod shells in the wild and the problem of reconstruction of fossil cephalopod habitat. Geosci J 10:331–345
Zakharov YD, Shigeta Y, Nagendra R, Safronov PP, Popov AM, Velivetskaya TA, Afanasyeva TB (2011) Cretaceous climate oscillations in the southern palaeolatitudes: new stable isotope evidence from India and Madagascar. Cretac Res 32:623–645
Zakharov YD, Melnikov ME, Popov AM, Pletnev SP, Khudik VD, Punina TA (2012) Cephalopod and brachiopod fossils from the Pacific: Evidence from the Upper Cretaceous of the Magellan Seamounts. Geobios 45:145–156
Zatoń M (2010) Sublethal injuries in Middle Jurassic ammonite shells from Poland. Geobios 43:365–375
Ziegler B (1963) Ammoniten als Faziesfossilien. Paläontol Z 37:96–102
Ziegler B (1967) Ammoniten-Ökologie am Beispiel des Oberjura. Geol Rundsch 56:439–464
Ziegler B (1981) Ammonoid biostratigraphy and provincialism: Jurassic-Old World. In: House MR, Senior JR (eds) The Ammonoidea: Systematics Assocoation, speial volume 18. Academic Press, London
Zinsmeister WJ (1987) Unusual nautilid occurrence in the upper Eocene La Meseta Formation, Seymour Island, Antarctica. J Paleontol 61:724–726
Acknowledgements
Studies were enhanced by the data generated from projects (P22109–B17, P20018–N10) financially supported by the Austrian Science Fund (FWF). The author wishes to thank Kazuyoshi Moriya (University of Kanazawa) for his extensive review of the manuscript and Christian Klug (University of Zurich) for his constructive comments and review. The author is grateful to Susanne Mayrhofer and Mathias Harzhauser (both Natural History Museum, Vienna) for discussions on specific problems on cephalopods and the life and habitat of ammonoids. The author thanks Kathleen A. Ritterbush (University of Southern California) for the calculation of a morphospace diagram. For important information and comments, I am grateful to Neil H. Landman (American Museum of Natural History, New York), Benjamin J. Linzmeier (University of Wisconsin-Madison) and two anonymous reviewers. Special thanks for editorial work and fruitful discussions on ammonoid life and habitat go to Christian Klug (University of Zurich) and Dieter Korn (Natural History Museum, Berlin).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Lukeneder, A. (2015). Ammonoid Habitats and Life History. In: Klug, C., Korn, D., De Baets, K., Kruta, I., Mapes, R. (eds) Ammonoid Paleobiology: From anatomy to ecology. Topics in Geobiology, vol 43. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-9630-9_18
Download citation
DOI: https://doi.org/10.1007/978-94-017-9630-9_18
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-9629-3
Online ISBN: 978-94-017-9630-9
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)