
Abstract
Nematode-trapping fungi of Orbiliaceae include those filamentous species forming trapping devices to prey on juveniles of nematodes. In this chapter, the taxonomic history of predatory orbiliaceous fungi is reviewed and the system of using trapping devices as the primary morphological criterion for generic delimitation is advocated. Following this taxonomic concept, keys for genera of Arthrobotrys, Drechslerella and Dactylellina, which include all reported species of predatory orbiliaceous fungi are presented. Totally, 54, 14 and 28 species from Arthrobotrys, Drechslerella and Dactylellina respectively, are morphologically described and illustrated. Known asexual-sexual connections (14) of predatory orbiliaceous fungi are summarized and their taxonomic descriptions and illustrations presented.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
In various natural environments, especially in soil, there are a wide and diverse range of microorganisms that function as antagonists of nematodes and include fungi, bacteria, viruses and rickettsias (Li et al. 2000). Nematophagous fungi or nematode-destroying fungi are antagonists of nematodes, and comprise a great variety of fungi belonging to widely divergent orders and families. Based on nematicidal mechanisms, these antagonists are assigned to the groups, endoparasitic fungi, opportunistic fungi, toxic fungi or nematode-trapping fungi (Li et al. 2000). Endoparasitic fungi are regarded as obligate parasites infecting nematodes via special spores and mycelia development takes place from spores within the nematode and only the reproductive-conidiophores are externalized (Li et al. 2000). Toxic fungi refer to those producing metabolites toxic to nematodes. Opportunistic fungi can colonize nematode reproductive structures and have the ability to seriously affect their reproductive capabilities (Siddiqui and Mahmood 1996).
Nematode-trapping fungi capture nematodes by trapping devices produced from the vegetative mycelia. The trap types reported include adhesive networks, adhesive knobs, constricting rings, non-constricting rings, adhesive branches, undifferentiated or unmodified adhesive hyphae, stephanocysts, spiny balls and acanthocytes (Zhang et al. 2001; Tzean and Liou 1993; Luo et al. 2004, 2007). The first five types of trap are known only to be produced by the predacious asexual morphs of Orbiliaceae of Ascomycota. Adhesive knobs are however, also formed by Zoophagus species of Zygomycetes (Li et al. 2000) and Nematoctonus species of basidiomycetes (Barron 1977).
Orbiliaceous members represent the majority of nematode-trapping fungi, which include 96 species, and are currently assigned in the asexual genera Arthrobotrys (53 species), Dactylellina (28 species) and Drechslerella (14 species). Traps of adhesive hyphae are restricted to the genera Stylopage and Cystopage of Zygomycetes (Drechsler 1941), while stephanocysts is restricted to the genus Hyphoderma of basidiomycetes (Tzean and Liou 1993). Spiny balls and acanthocytes are formed respectively by the macrobasidiomycetes Coprinus comatus (Luo et al. 2004, 2007) and Stropharia rugosoannulata (Luo et al. 2006). As mentioned above, the orbiliaceous nematode-trapping are the most complex and taxonomically unsettled.
In this chapter we focus on the largest group of nematode-trapping fungi in the Ascomycota. We discuss their historical classification and the taxonomic system currently accepted. All species of orbiliaceous nematode-trapping fungi are described and illustrated. Keys to the genera Arthrobotrys, Dactylellina and Drechslerella are provided in order to facilitate the readers to compare the species features. Furthermore, asexual-sexual state connections of the orbiliaceous nematode-trapping fungi are discussed in species descriptions and illustrations.
Systematics of Nematode-Trapping Fungi
Asexual nematode-trapping hyphomycetous fungi belong to a predatory fungal lineage. Distinctive trapping devices such as adhesive hyphae, adhesive knobs, adhesive networks, constricting rings, and non-constricting rings develop from extensions of the mycelia (Barron 1977). The trapping devices enable capture and cause subsequent mortality of nematodes, other small animals, and protozoans and the nematode-trapping fungi then obtain nutrients from these prey (Barron 1977). Since nematode-trapping fungi play an important role as antagonists of plant-parasitic and animal-parasitic nematodes, there is a great interest in using these fungi as biological control agents. Information concerning the systematic position of nematode-trapping fungi is therefore essential for understanding and exploiting them in biocontrol.
Asexual nematode-trapping hyphomycetous fungi are placed within the family Orbiliaceae (Orbiliales, Orbiliomycetes), based on morphological and molecular studies (Pfister 1997; Eriksson et al. 2003; Yu et al. 2011). The family Orbiliaceae was not known to be nematophagous until Pfister (1994) reported that a collection of Orbilia fimicola Jeng & Krug produced an Arthrobotrys asexual state named A. superba Corda. Subsequently, more and more connections between Orbiliaceae and predacious hyphomycetes have been established (Liu et al. 2005a; Mo et al. 2005a; Pfister 1995; Yu et al. 2006; Qiao et al. 2012; Li et al. 2009; Yu et al. 2009a). Connections between Orbiliaceae and other non-predacious hyphomycetous genera have also been reported (Liu et al. 2005b; Yu et al. 2009b, 2007a, 2007b, 2011; Mo et al. 2005b; Yang and Liu 2005). At least nine asexual genera have been linked to sexual Orbiliaceae species (Pfister 1997; Yu et al. 2011).
The family Orbiliaceae was shown to be monophyletic in the phylogenetic analyses of 18S and ITS rDNA sequences (Pfister 1997). Morphologically, the unique nature of the ascospores of this family, especially the inclusion of an elaborated spore body makes it distinguishable from other discomycetes. According to phylogenetic analysis of SSU rDNA sequences of 21 taxa, the Orbiliomycetes clustered near the base of the tree near the Pezizomycotina. A signature of six base pairs was consistently found in every studied species of Orbiliomycetes, which is a further evidence in the isolated position of the class (Eriksson et al. 2003). However, a range of conidial morphology is found in nematode-predacious and non-predacious asexual morphs in some phylogenetic clades. For example, in earlier classification systems, conidiophore characters were used to distinguish asexual genera. Within the series Auricolores in the sexual genus Orbilia, asexual morphs with a candelabrelloid or more simple type of conidiogenesis actually cluster, often separately, from those with an arthrobotryoid type. However, the arthrobotryoid type occurs scattered in several clades, therefore, this character does not permit a split of series Auricolores into two groups. Conidiophore types however, appear to characterize small groups within that series (Baral, pers. comm.).
Nematode-trapping fungi were formerly classified into a number of genera based on the morphology of conidia (shape, septa and size) and conidiophores (branching, modifications of the apex), such as Arthrobotrys, Dactylellina, Drechslerella, Monacrosporium, Dactylella, Gamsylella, Didymozoophaga, Trichothecium, Dactylaria, Candelabrella, Genicularia, Duddingtonia, Nematophagus, Monacrosporiella and Woroninula. This has led to a situation where species with diverse types of trapping devices have been assigned to one genus, while others with similar trapping devices can be found in different genera (Glockling and Dick 1994; Liu and Zhang 1994, 2003; Zhang et al. 1996a). The genus Arthrobotrys was first established for the type species, A. superba (Corda 1839), which is characterized by the formation of two-celled conidia on denticles in a whorled arrangement at the apex of the nodes of the simple, erect, septate conidiophore. Schenck et al. (1977) expanded the genus to include species with aseptate and multi-celled conidia. Scholler et al. (1999) subsequently limited Arthrobotrys to only those species with adhesive networks.
Subramanian (1963) erected Drechslerella for D. acrochaeta (Drechsler) Subram. based on the filiform apical appendage on its conidia. Although Liu and Zhang (1994) synonymized Drechslerella with Monacrosporium, detailed molecular analyses (Hagedorn and Scholler 1999) convinced Scholler et al. (1999) to propose new generic concepts for orbiliaceous nematode-trapping fungi based on mode of trapping device. We accept these concepts and recognize Drechslerella as characterized by forming three-celled constricting ring traps.
The genus Dactylellina was described by Morelet (1968) with Dactylellina leptospora (Drechsler) M. Morelet as type species. Species included in this genus were characterized by elongate, fusoid conidia, while microconidia and microconidiophores rarely formed. Dactylellina was recognized by Scholler et al. (1999) for species characterized by stalked adhesive knobs and included those species producing non-constricting rings. The knob-forming species of the nematode-trapping fungi however, have diverse configurations such as sessile knobs, stalked knobs, a combination of stalked knobs with non-constricting rings, or stalked knobs with proliferated knobs.
Earlier phylogenetic analyses based on ribosomal RNA (rRNA) gene sequences suggested that the trapping devices could rationalize the classification of nematode-trapping fungi (Rubner 1996). Indeed, lineages including species with stalked adhesive knobs, adhesive networks and constricting rings consistently separate from each other in molecular phylogenetic analysis. Recently, phylogenetic analyses based on ribosomal RNA (rRNA) and protein-coding gene sequences suggested that the trapping devices could serve as robust indicators of generic delimitation for these asexual fungi (Yang and Liu 2006; Ahrèn et al. 1998; Li et al. 2005; Scholler et al. 1999), different trapping devices-forming species were separated into different genetic lineages and new generic concepts of the taxonomy of predatory sexual Orbiliaceae were proposed.
Ahrén et al. (1998) found that nematode-tapping fungi clustered into three lineages: species with constricting rings, species with various adhesive structures (net, hyphae, knobs and non-constricting rings) and species have no trapping devices. Based on results obtained from morphological and molecular characters, Hagedorn and Scholler (1999) and Scholler et al. (1999) classified nematode-trapping fungi into four genera: Dactylellina characterized by stalked adhesive knobs including species characterised by non-constricting rings and stalked adhesive knobs; Gamsylella characterized by adhesive branches and unstalked knobs; Arthrobotrys characterized by adhesive networks and Drechslerella characterized by constricting rings. Furthermore, the systematic classification of nematode-trapping fungi was redefined based on phylogenies inferred from sequence analyses of 28S rDNA, 5.8S rDNA and ß-tubulin genes and an emended generic concept of nematode-trapping fungi is provided (Li et al. 2005). Arthrobotrys is characterized by adhesive networks, Dactylellina by adhesive knobs, and Drechslerella by constricting-rings. Phylogenetic placement of taxa characterized by stalked adhesive knobs and non-constricting rings is also confirmed in Dactylellina. Based on careful morphological observation of culture, Li et al. (2005) found that none of these taxa only formed unstalked adhesive knobs, which was just one temporary structure formed at the first stage in the fungus development. These unstalked knobs could then grow out to form branches, which may further change in different species, they proposed that unstalked adhesive knobs should not be treated as a unique type of trapping-device and this character should not be given taxonomic importance, and the genus Gamsylella is invalid. Species which produce unstalked adhesive knobs that grow out to form loops were transferred from Gamsylella to Dactylellina, and those which produce unstalked adhesive knobs that grow out to form networks were transferred from Gamsylella to Arthrobotrys.
Based on the combined phylogenetic analysis of rDNA and protein-coding genes and morphological studies, a hypothesis for the evolution of trapping-devices was also presented (Li et al. 2005). Predatory and non-predatory fungi appear to have been derived from non-predatory members of Orbilia. The adhesive knob is considered to be the ancestral type of trapping-device from which constricting rings and networks were derived via two pathways. In the first pathway adhesive knobs retained their adhesive material forming simple two-dimension networks, eventually forming complex three-dimension networks. In the second pathway adhesive knobs lost their adhesive materials, with their ends meeting to form non-constricting rings and they in turn formed constricting rings with three inflated-cells. Furthermore, using phylogenetic analysis of nucleotide sequences of three protein-coding genes (RNA polymerase II subunit gene, rpb2; elongation factor 1-α gene, ef1-α; and ß tubulin gene, bt) and ITS rDNA region, Yang et al. (2007) described the evolution of nematode-trapping cells of predatory fungi, they thought the initial trapping structure evolved along two lineages yielding two distinct trapping mechanisms: one developed into constricting rings and the other developed into adhesive traps. Among adhesive trapping devices, the adhesive network separated from the others early and evolved at a steady and gentle speed. The adhesive knob evolved through stalk elongation, with a final development of non constricting rings. The derived adhesive traps are indicated as a highly differentiated stage.
The non-predacious genus Dactylella was established by Grove (1884) on the basis of one species, D. minuta Grove. The genus is characterized by erect, simple, hyaline conidiophores with conidia produced singly at the apex. Conidia are ellipsoidal, fusoid or cylindrical, one-celled at first and later having 2 to many septa. This genus has been emended several times and both non-predacious and predacious fungi have been included (Subramanian 1963; Schenck et al. 1977; Zhang et al. 1994). Dactylella includes diverse taxa in morphology and behavior. There are considerable differences in conidiophore length and conidial size among species. Some species are saprotrophic, while others are oospore or nematode egg parasites (Zhang et al. 1994). Rubner (1996) revised the generic concept of Dactylella and excluded the nematode-trapping species. However, the classification has not commonly been accepted and several predacious species had been described under Dactylella (Liu and Zhang 2003). Chen et al. (2007a, b, c) emended the asexual state genus Dactylella complex based on ITS phylogeny, all the tested strains cluster into three monophylogenetic clades corresponding to three genera, e.g. Dactylella, Vermispora and a distinct group comprising species with very short conidiophores. The circumscription of Dactylella and Vermispora were emended and a new genus Brachyphoris characterized by very short conidiophores that are scarcely longer than conidia was established, which sexual states corresponding to the genus Hyalorbilia within Orbiliaceae, and other species separated into genera Dactylella and Vermispora, respectively.
Although trapping devices have proven as vital in generic delimitation of nematode-trapping fungi, species in the same genus are mainly delimited from each other by morphology of conidia and conidiophores. With more and more connections between nematode-trapping fungi and Orbiliaceae being established, phylogenetic analysis including existing asexual species indicated that conidial morphology showed only little correlation while the sexual state characters showed a high level of correlation. For instance, Arthrobotrys belong to Series Auricolores, which is defined by the ability to form adhesive networks and the asexual Trinacrium-type belong to species of Hyalorbilia, Orbilia subgenus Hemiorbilia and subgenus Orbilia (Baral, pers. comm.). Further studies are needed to set up a natural classification system including both asexual and sexual nematode-trapping fungi. In fact this is now a requirements of the International Code of Nomenclature for Algae, Fungi and Plants as two names can no longer be used for asexual and sexual morphs of the same fungus.
Key to Genera of Nematode-Trapping Fungi
1 Trapping-device a constricting ring, which consists of three inflated cells with a short, strong stalk Drechslerella
2 Trapping-device not a constricting ring, but various adhesive trapping devices 3
3 Trapping device unstalked adhesive knobs which develops into an adhesive network or adhesive networks only Arthrobotrys
4 Trapping device stalked adhesive knobs, some with non-constricting rings, or unstalked adhesive knobs which grow out to form adhesive branches and loops Dactylellina
Arthrobotrys Corda, Pracht-Fl. Eur. Schimmelbild.: 43 (1839) |
Type species: Arthrobotrys superba Corda, Pracht-Fl. Eur. Schimmelbild.: 43 (1839) |
Synonyms |
Monacrosporium Oudem., Ned. Kruidk. Arch., Ser.2, 4: 250 (1885) |
Lectotype species: Monacrosporium elegans Oudem., Ned. Kruidk. Arch., Ser.2, 4: 250 (1885), designated by Clements and Shear (1931) |
Didymozoophaga Soprunov & Galiulina, Mikrobiologiya 20: 493 (1951) (invalid; Art.36; illegit. Art. 52 ICBN) |
Type species: Didymozoophaga superba (Corda) Soprunov & Galiulina, Mikrobiologiya 20: 494 (1951) (invalid; Art. 36 ICBN). |
Arthrobotrys superba Corda, Pracht–Fl. Eur. Schimmelbildung.: 43 (1839) |
Candelabrella Rifai & R.C. Cooke, Trans. Br. mycol. Soc. 49 (1): 160 (1966) |
Type species: Candelabrella javanica Rifai & R.C. Cooke, Trans. Br. Mycol. Soc. 49: 160 (1966). |
Duddingtonia R.C. Cooke (1969), Trans. Br. mycol. Soc. 53 (2): 315 (1969) |
Type species: Duddingtonia flagrans (Dudd.) R.C. Cooke, Trans. Br. Mycol. Soc. 53: 315 (1969) |
Trichothecium flagrans Dudd., Trans. Br. Mycol. Soc. 32: 287 (1949) |
Genicularia Rifai & R.C. Cooke, Trans. Br. Mycol. Soc. 49: 153 (1966) [non Genicularia Rouss. ex Desv. 1808, non de Bary 1858] (nom. illeg.;. Art. 53 ICBN; replaced by Geniculifera) |
Type species: Genicularia cystosporia (Dudd.) Rifai & R.C. Cooke, Trans. Br. Mycol. Soc. 49: 154 (1966) |
Trichothecium cystosporium Dudd., Trans. Br. Mycol. Soc. 34: 600 (1951a) |
Nematophagus Mekht., Mikol. Fitopatol. 9 (3): 250 (1975) |
Type species: Nematophagus azerbaidzhanicus Mekht., Mikol. Fitopatol. 9: 250 (1975) |
Monacrosporiella Subram., Kavaka 5: 94 (1978) [1977] |
Type species: Monacrosporiella megalospora (Drechsler) Subram., Kavaka 7: 94 (1977). |
Dactylella megalospora Drechsler, Mycologia 46: 769 (1954) |
Woroninula Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 109 (1979) |
Type species: Woroninula polycephala (Drechsler) Mekht., Khishchnye nematofagovye Griby—Gifomitsety: 110 (1979) |
Dactylaria polycephala Drechsler, Mycologia 29: 530 (1937) |
Characteristics:
Mycelium fast growing. Hyphae septate, branching, hyaline. Conidiophores simple or branched, apex mainly with short denticles or with geniculate, candelabrelloid, or percurrent proliferations, rarely simple without modifications. Conidia either singly or in clusters at the apex of the conidiophore. Conidia holoblastic, hyaline, non-septate to multi-septate, mainly ellipsoidal, spindle-shaped or ovoid, rarely clavate, cylindrical, pyriform, or turbinate. Microconidia and microconidiophores frequently formed. Chlamydospores, when present, intercalary or rarely terminal, singly or in chains, thick-walled, sphaerical to ovoid, yellow pigmented. Sexual state when know, belonging to Orbilia. Trapping nematodes by means of adhesive networks or modified devices, with good saprotrophic capabilities. Formation of traps induced by nematodes in most species.
Key to Species of Arthrobotrys
1. Conidia produced singly at the apices of conidiophores or on subapical, lateral branches 2
1. Conidia not produced singly, often in groups 16
2. Conidia clavate, with 2–9 septa, mostly 3–7-septate, without equal-sized cells A. shizishana
2. Conidia other type 3
3. Conidia obovoid, obconical, broad turbinate or subglobose, ratio of length to width less than 2 4
3. Conidia fusiform, ratio of length to width more than 2 8
4. Conidia broadly turbinate to napiform, with 1–2 septa, distal cell largest A. janus
4. Central or distal cell of conidia largest 5
5. Conidia with 0–2 septa, or 0–3-septate 6
5. Conidia with 1–3 septa, or 2–3-septate 7
6. Conidia ellipsoid to obovoid to broadly turbinate, with 0–2 septa, 17.5–30 (23.2) × 12.5–20 (14.8) A. indica
6. Conidia globose to obovoid, with 0–3 septa, 20–44 (32) × 17–25 (20.4) μm A. sphaeroides
7. Conidia globose to turbinate, with 2–3 septa, 7–47.5 (32.2) × 17.5–27.5 (22) μm A. rutgeriense
7. Conidia subglobose, with 1–3 septa, 23.5–30 (27.6) × 17–25 (20) μm A. sinensis
8. Conidia with 4–12 septa, conidia longer A. multiformis
8. Conidia with 2–5 septa 9
9. Conidial less than 20 μm wide 10
9. Conidial wider than 20 μm 11
10. Conidia fusiform to ellipsoid, with 2–3 septa, 32.5–47.5 (41) × 12.5–17.5 (15.5) μm A. fusiformis
10. Conidia fusiform to ellipsoid, with 0–4 septa, mostly 3-septate A. salina
11. Conidia mostly 4-septate 12
11. Conidia mostly with 2 or 3–4 septa 13
12. Conidia broadly fusiform, with 2–4 septa, mostly 4-septate, with larger size A. mega lospora
12. Conidia fusiform, sometimes with 5 septa, 50–65 × 20–25 μm A. reticulata
13. Conidia turbinate, obovoid, fusiform, mostly 2–3-septate, with narrower and longer basal cell A. cookedickinson
13. Conidia fusiform to obpyriform 14
14. Conidia 2–5-septate, mostly 3–4-septate, 40–90 (54) × 15–27.5 (18) μm A. longiphora
14. Conidia mostly 2–3-septate 15
15. Conidia mostly 3-septate, 40–52–65 × 17–20–23 μm A. oudemansii
15. Conidia 1–4-septate, mostly-2 septate, obpyriform to fusiform A. huaxiensis
16. Conidiophores often merged into a bundle A. dendroides
16. Conidiophores not merged into bundles 17
17. Conidia 0–1-septate 18
17. Conidia 1- or more than 1-septate 23
18. Conidia lacking septa 19
18. Conidia 0–1-septate 20
19. Conidia obovoid to ellipsoid, 15–31 (23.6) × 10–20 (15.9) μm A. amerospora
19. Conidia elongate-ellipsoid, 11–16.8 × 5–6.6 μm A. nonseptata
20. Conidia produced from longer sterigmata, conidia occasionally 1-septate, 17.5–32.5 (22.57) × 2.75–7.5 (5.5) μm A. yunnanensis
20. Conidia producing from swollen denticles of apex 21
21. Conidia broadly ovoid to ellipsoid, 14.8–21.5 × 10.1–16.3 μm A. latispora
21. Conidia other type 22
22. Conidia ellipsoid, occasionally 1-septate, 12–28 × 10–15 µm A. botryospora
22. Conidia cylindrical to clavate 23
23.Conidiophores with apical as well as intercalary clusters of conidia, conidia cylindrical, occasionally clavate, minority 1-septate at centre A. anomala
23. Conidiophores with apical clusters of conidia only, conidia clavate, septum near the base A. pseudoclavata
24. Conidia always 1-septate 25
24. Conidia sometimes more than 1-septate 39
25. Conidiophores with longer sterigmata at the apex, of the candelabrelloid type 26
25.Conidiogenous loci denticles, or shorter sterigmata 27
26. Conidia obovoid to clavate, narrowed at septum A. javanica
26. Conidia elongate-obovoid or ovoid, not narrowed at septum, with narrower and slender basal cell, 33.5–57 × 11–15.5 μm A. shahriari
27. Conidiophores geniculation because of repeated elongation, each conidiogenous loci with 1–3 conidia 28
27. Conidiophores erect, not geniculation 30
28.Conidiogenous loci without obvious denticles, conidia narrow pyriform, 24–40 × 12.5–18.8 μm A. paucispora
28. Conidiogenous loci with obvious denticles 29
29. Conidia broadly pyriform, 25–35 × 18–24 μm, capturing nematodes by adhesive networks A. cystosporia
29. Conidia obpyriform, 24–32.5 × 12.5–20 μm, capturing nematodes by simple hyphae A. perpasta
30. Conidiogenous loci without obvious denticles, capturing nematodes by simple hyphae A. flagrans
30. Conidiogenous loci with typical denticles or sterigmata 31
31. Conidiophores with several groups denticles 32
31. Conidiophores with apical or occasionally subapical denticles or sterigmata 34
32. Conidia subellipsoid, not narrowed at septum, 7.5–27.5 (5.8) × 5–10.5 (6.6) μm A. superba
32. Conidia obovoid to pyriform 33
33. Conidia obovoid, not narrowed at septum, 28.5–32 (30) × 18–20.5 (20) μm A. obovata
33. Conidia pyriform to obovoid, narrowed at septum, 17–35 (23) × 8.5–16 (12.1) μm A. oligospora
34. Conidiophores unbranched 35
34. Conidiophores branched or occasionally branched 36
35. Conidia obconical, produced from denticles, 15–37.5 (8.4) × 7.5–14.5 (11.8) μm A. conoides
35. Conidia ellipsoid, produced from sterigmata, 20–47.5 (30.9) × 7–12.5 (10.3) A. musiformis
36. More than 10 conidia arranged in a close capitate head 37
36. 1–6 conidia arranged in a loose capitate head 38
37. Conidia pyriform, occasionally ovoid, 26–40 × 12–23 µm A. apscheronika
37. Conidia ellipsoid to ovoid, 10–20 (17.5) × 5–8 (6.2) µm A. cladodes
38. Conidia elongate-ovoid, produced from denticles, 22.5–32 × 11–22.5 µm A. chazarica
38. Conidia obovoid, produced from sterigmata, 20–27.5 (24.4) × 7.5–12.5 (10.8) µm A. robusta
39. Conidia with more septa, mostly 2–5-septate, capturing nematodes by simple two-dimensional networks A. dianchiensis
39. Conidia less than 4-septate, capturing nematodes by three dimensional networks
40. Conidia sometimes non-septate 41
40. Conidia at least 1-septate 42
41. Conidia multiform, broadly turbinate to narrow fusiform, ellipsoid, clavate, obovoid A. mangrovispora
41. Conidia fusiform, largest cell obvious A. micro scaphoides
42. Conidia fusiform or other type 43
42. Conidia other type, never fusiform 48
43. Conidia fusiform to other type 44
43. Conidia only fusiform 45
44. Conidia fusiform, obovoid or broadly conical, 3-septate, 37–55 (49) × 17.5–35 (28)
μm A. eudermata
44.Conidia pyriform or fusiform, 1–3-septate, mostly 1-septate A. guizhouensis
45. Conidia mostly 3-septate 46
45. Conidia mostly 4 or 3–4-septate 47
46. Conidia of two types, macroconidia arranged loosely, 3–4-septate, 30–50 (45.1) × 8–16.5 (12.2) μm A. polycephala
46. Conidia arranged closely, 1–4-septate, 30–60 (36.2) × 15–30 (20.2) μm A. thaumasia
47. Conidia sometimes curved, mostly 4-septate, 40–70 × 9–14 μm A. gampsospora
47. Conidia mostly 3–4-septate, 46–70 (62.3) × 21–29 (24.7) A. psychrophila
48. Sterigmata or denticles formed from flank of conidiophores 49
48. Conidiophores with apical and at least subapical clusters of conidia 50
49. Conidia pyriform to broadly clavate,1–3 septa, mostly 2-septate, 25–40 (17.5) × 7.5–19 μm A. clavispora
49. Conidia elongate-ovoid to ellipsoid, 1–3-septate, mostly 1–2 septa, 28.5–56 × 11.5–22.5 μm A. tabrizica
50. Conidia mostly 1-septate or with 1–2 septa 51
50. Conidia mostly 2-septate or with 2–3 septa 52
51. Conidia obovoid to ellipsoid, mostly 1–2-septate, 18–36 (28.1) × 12–20 (15.3) µm A. azerbaijanica
51. Conidia pyriform, 1–2-septate, mostly with 1 septum A. oviformis
52. Conidia broadly pyriform to subcylindrical, mostly 2–3-septate, occasionally 1-septate, 20–35 × 10–25 μm A. pyriformis
52. Conidia ellipsoid, obovoid or cymbiform 52
53. Conidia of two types, macroconidia ellipsoid or cymbiform,1–3-septate, mostly 2-septate A. scaphoides
53. Conidia ellipsoid to obovoid or cymbiform, 2–3-septate A. vermicola
Accepted Species of Arthrobotrys
1. Arthrobotrys amerospora S. Schenck, W.B. Kendr. & Pramer, Can. J. Bot. 55 (8): 979 (1977) [1976] |
Characteristics:
Mycelium hyaline, septate, branched. Conidiophores simple, erect, septate, unbranched, 75–250 µm long, producing 2–10 conidia singly from retrogressive conidiogenous loci on broad, conspicuous denticles at and near the apex. Conidia holoblastic, hyaline, obovoid, one-celled, 15–31 (23.6) × 10–20 (15.9) µm with a small truncate protuberance at the base. Germinating from the base, and often subsequently also from the apex. Chlamydospores yellowish, smooth-walled, sphaerical to elongate-ellipsoidal, 18–50 × 18–23 µm, usually intercalary, single or in chains. Capturing nematodes on adhesive hyphal loops.
Distribution:
Canada, China (Guansi), India, USA.
Notes:
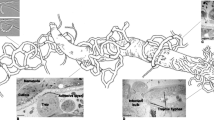
The description is based on the protologue. Among Arthrobotrys, there are five species with single celled conidia, i.e. A. amerospora (Schenck et al. 1977), A. anomala (Barron and Davidson 1972), A. botryospora (Barron 1979), A. yunnanensis (Mo et al. 2005a) and A. nonseptata (Yu et al. 2009b). However, only conidia of A. amerospora and A. nonseptata are consistently non-septate, and conidia of the other three species are occasionally 1-septate. The obovoid conidia of A. amerospora distinguishes it from the elongate-ellipsoidal conidia of A. nonseptata. (Fig. 3.1)

Arthrobotrys amerospora. a conidiophore; b conidia; c adhesive network. Bar = 10 µm
Characteristics:
Conidiophores short, erect or suberect, hyaline, indeterminate, 20–80 × 4–6 µm; conidia borne in an apical cluster or series of clusters, sometimes borne irregularly along the spore-bearing apex; conidia cylindric to long ellipsoid, occasionally clavate, hyaline, non-septate when attached, frequently forming a septum before germination, produced sympodially on pronounced denticles, 13–22 × 3–7 µm. Nematodes trapped by means of short, erect adhesive branches or more commonly by complex three dimensional adhesive networks.
Distribution:
Canada.
Notes:
The description is based on the protologue. Arthrobotrys anomala is characterized by cylindric to elongate ellipsoidal, 0 (–1)-septate conidia, which distinguish it from other species of Arthrobotrys. (Fig. 3.2)

Arthrobotrys anomala. a conidiophores; b conidia. Bar = 20 µm
Arthrobotrys anomala G.L. Barron & J.G.N. Davidson, Can. J. Bot. 50: 1773 (1972) |
Arthrobotrys apscheronica Mekht., Nov. Sist. Niz. Rast., 10: 174 (1973) |
Characteristics:
Conidiophores erect or curved, sometimes branched, occasionally repeated propagated, conidiophore apex expanded irregularly, bearing 12 (–16) conidia on denticles of apex; conidia pyriform, seldom ovoid, 1-septate at the centre of conidia, constricted at the septum. Nematodes trapped by three dimensional adhesive networks.
Distribution:
Russia.
Notes:
The description is based on the protologue. This species is similar to A. conoides in conidial shape, but it differs in having longer conidia (Fig. 3.3).

Arthrobotrys apscheronika. a conidiophore; b conidia; c adhesive network. Bar = 30 µm
Arthrobotrys azerbaijanica (Mekht.) Oorschot [as ‘azerbaidzhanica’], Stud. Mycol. 26: 70 (1985) |
Nematophagus azerbaijanicus Mekht. [as ‘azerbaidzhanicus’], Mikol. Fitopatol. 9: 250 (1975) |
Characteristics:
Conidiophores erect, branched or unbranched, repeatedly propagating at least once, apex expanded, bearing 10–12 conidia on short and slender denticles; conidia obovoid to ellipsoidal, 1–4-septate, constricted at septum, sometimes 1–2 cells expanded, 18–36 (28.1) × 12–20 (15.3) µm; Chlamydospores present. Nematodes trapped by three dimensional adhesive networks.
Distribution:
Russia.
Notes:
The description is based on the protologue. This species resembles A. pyriformis, but has wider conidia (conidia of A. pyriformis 17–38 × 6.5–11.5 μm, mostly 2–3-septate). (Fig. 3.4)

Arthrobotrys azerbaijanica. a conidiophore; b conidia; c adhesive network. Bar = 30 µm
Arthrobotrys botryospora G. L. Barron, Can. J. Bot. 57: 1371 (1979) |
Characteristics:
Colonies on CMA whitish. Mycelium spreading, vegetative hyphae hyaline, septate branched. Conidiophores hyaline, erect, septate, 250–350 µm long, 4.5–6.5 µm wide at the base, gradually tapering upwards to a width of 3–5 µm at the apex, simple or sparingly branched in strikingly loose capitate arrangement. Conidia 12–28 × 10–15 µm, hyaline, ellipsoid, non-septate or occasionally 1-septate (8 %) just below the centre of the spore and having a narrowly truncate protuberance at the base. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
Canada (Ontario), China (Anhui, Beijing, Guizhou, Hebei)
Material examined:
C9–1, C21–1, isolated from forest soil in Cangshan Mountain, Dali in 1999 by Lu Cao; 01–3, isolated from forest soil in Shigu, Lijiang in 1999 by Lu Cao.
Notes:
The only species of Arthrobotrys described with non-septate conidia are A. anomala and A. amerospora. Arthrobotrys anomala differs from A. botryospora in its cylindrical conidia. In A. amerospora, the conidia are larger than those of A. botryospora. The former also produce abundant chlamydospores, while chlamydospores are not produced in A. botryospora. In A. amersospora the conidia form in a solitary terminal cluster, produced from a retrogressive conidiogenous locus which remains relatively unswollen. In A. botryospora, successive production of conidia produces a swollen apex, with the main axis branching or elongating sympodially to produce a succession of spore clusters. (Fig. 3.5)

Arthrobotrys botryospora. a conidiophore; b conidia; c adhesive network. Bar = 20 µm
Arthrobotrys chazarica Mekht., Mycol. Res. 102: 683 (1998) |
Characteristics:
The fungus forms a delicate, fluffy, pale grey colony in pure culture on malt agar with pigments diffusing into the agar. Sporulation is abundant. Conidiophores simple, erect, branched. Conidia obovate to elongate-obovate, 1-septate, 22.5–32 × 11–22.5 μm, growing on very closely arranged, thin denticles. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
Azerbaijan (Baku).
Notes:
The description is based on the protologue. This species is similar to A. robusta in being 1-septate at the centre of the conidia, but conidia of A. robusta are mainly oblong-pyriform, and conidia are smaller. (Fig. 3.6)

Arthrobotrys chazarica. a conidiophore; b conidia; c adhesive network. Bar = 25 µm
Arthrobotrys cladodes Drechsler, Mycologia 29: 463 (1937) |
Trichothecium cladodes (Drechsler) Soprunov, Predacious fungi—Hyphomycetes and their use in the control of pathogenic nematodes: 113 (1958) |
Characteristics:
Colonies on CMA whitish, slow growing. Mycelium spreading, scanty; vegetative hyphae hyaline, septate, branched. Conidiophores hyaline, erect, septate, more or less branched, 68–236 μm long, 2.5–5 μm wide at the base, tapering gradually upwards to a width of 2–3 μm below the irregularly expanded, globose or somewhat coralloid apex, whereon are borne 10–20 conidia usually in a dense capitate arrangement. Conidia hyaline, ellipsoid or elongate obovoid, mostly 10–20 (17.5) × 5–8 (6.2) μm, 1-septate near the centre of the spore. Chlamydospores not formed. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Beijing, Guizhou, Hebei, Taiwan, Xizang, Yunnan), Cuba (Prov. Ciudad de La Habana), UK, USA (Wisconsin)
Material examined:
YMF1.00038, isolated from forest soil in Deqin, Yunnan, in 2002 by Jing Zhang; ①–33, isolated from field soil in Guiyang, Guizhou, in May 2002 by Ke-Qin Zhang; XZA–3, isolated from field soil in Zhouxian, Xizang in August 2002, by Minghe Mo. Permanent slide: H1–33.
Notes:
This species was first isolated from leaf mold of deciduous woods in Virginia and Maryland, which is similar to A. superba, A. oligospora and A. conoides in conidia shape, but branch mode of conidiophores differ. The apex of the A. cladodes conidiophore is irregularly expanded, on which 10–20 conidia form in a densely capitate arrangement. (Fig. 3.7)

Arthrobotrys cladodes. a, b conidia; d adhesive network; c, e conidiophore. Bar = 10 µm
Arthrobotrys clavispora (R.C. Cooke) S. Schenck, W.B. Kendr. & Pramer, Can. J. Bot. 55: 982 (1977) |
Dactylaria clavispora R.C. Cooke, Trans. Br. Mycol. Soc. 47: 307 (1964) |
Genicularia clavispora (R.C. Cooke) Rifai, Reinwardtia 7 (4): 367 (1968) |
Geniculifera clavispora (R.C. Cooke) Rifai, Mycotaxon 2 (2): 216 (1975) |
Nematophagus clavisporus (R.C. Cooke) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 107 (1979) |
Characteristics:
Conidiophores erect, unbranched, septate, 150–350 μm high, at the apex of each bearing 3–11 widely spaced conidia, pleurogenous on small sterigmatal branches. A single, terminal conidium is formed at the conidiophore apex, which continues to grow for a short distance so that the conidium is displaced laterally. A second conidium is then formed at the new apex followed by growth of the conidiophore displacing this conidium. This process is repeated giving rise to the arrangement of conidia typical of this species. Conidia pyriform to broadly clavate, 25–40 (17.5) × 7.5–19 μm, rounded distally and tapering proximally to a narrow, truncate base. They are 1–2-septate, about twice as many having one septum as two. In both forms of conidium the distal cell is usually the larger. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
France.
Notes:
The description based on the protologue. This species is characterized by widely spaced conidia borne pleurogenous on small sterigmatal branches. (Fig. 3.8)

Arthrobotrys clavispora. a conidiophores with conidia; b conidia; c adhesive network. Bar = 20 μm
Arthrobotrys conoides Drechsler, Mycologia 29 (4): 476 (1937) |
Arthrobotrys pravicovii (Soprunov) Sidorova, Gorlenko & Nalepina [as ‘pravicovi’], Bot. Zh. SSSR 49: 1598 (1964) |
Arthrobotrys pravicovii (Soprunov) Mekht. [as ‘pravicovi’], Dokl. Akad. Nauk Azerb. SSR 20 (6): 71 (1964) |
Arthrobotrys tortor Jarow., Acta Mycologica, Warszawa 4: 241 (1968) |
Trichothecium pravicovii Soprunov [as ‘Pravicovi’], Predacious fungi—Hyphomycetes and their use in the control of pathogenic nematodes: 117 (1958) |
Characteristics:
Colonies on CMA whitish then turned to yellowish; mycelium spreading, vegetative hyphae hyaline, septate, branched, except for occasional storage filaments that are densely filled with protoplasm and up to 12 μm wide, measuring mostly 2–8 μm in diameter. Conidiophores hyaline, erect, 5–10-septate, usually not branched, mostly 4–7.5 μm wide at the base, tapering gradually upwards to a width of 2.5–5 μm in attaining a height of 120–420 μm before bearing on a globose or more irregularly expanded apex as many as 30 conidia in dense capitate arrangement; subsequently often, following repeated elongation, giving rise successively to additional clusters of conidia. Conidia hyaline, obconical, 15–37.5 (28.4) × 7.5–14.5 (11.8) μm, somewhat flattened at the base, broadly rounded at the apex, 1-septate, usually perceptibly constricted at the at the septum. Chlamydospores yellowish, globose or prolate ellipsoidal, 17–24 μm in diameter, or sometimes narrower, oblong-cylindrical, 30–50 μm long and approximately 15 μm wide. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Guizhou, Hainan, Hubei, Tianjing, Xizang, Yunnan), France, Germany (Berlin), Netherlands, Russian, South Africa (North West Province), Spain (Chavio verde), USA.
Material examined:
8705, isolated from dunghill in Huaxi, Guizhou in 1988 by Ke-Qin Zhang; GZHS–3, isolated from forest soil in Huishui, Guizhou in 1996 by Ke-Qin Zhang; J9–1, J11–2, J5–3, isolated from soil in Jianshui, Yunnan in 1999 by Yanju Bi; TJ–W1Z03, isolated from soil in Wanglanzhuang, Tianjing in 2000 by Wenpeng Li; XZA–4, isolated from pasture soil in Xizang in August 2000 by Minghe Mo; DL1–1, isolated from forest soil in Dali, Yunnan in September 2002 by Jing Zhang; YMF1.00009, YMF1.00541, YMF1.00866, J49–1, isolated from field soil in Huaxi, Guizhou in 1996 by Ke-Qin Zhang; YMF1.00009, YMF1.00541, isolated from forest soil in Baoshan, Yunnan in June 2002 by Jing Zhang; YMF1.00551, YMF1.00866, isolated from forest soil in Lijiang, Yunnan in October 2002 by Jing Zhang. Permanent slide: J49–1.
Notes:
A. conoides has an extensive worldwide distribution, and was first isolated from delaying leaves from a greenhouse (Drechsler 1937). It resembles A. oligospora and A. superba in most morphological characters, with the exception of conidial shape and the position of septum. In A. conoides, conidia are elongate obconical, 1-septate in the lower third and constricted at the septum. In A. oligospora, conidia are obovoid, 1-septate near the base of the spore, and constricted at the septum. In A. superba, conidia are ellipsoidal, and 1-septate at the centre of the conidia.
The characters of examined strains match well with the original description (Drechsler 1937). Van Oorschot (1985) rechecked A. tortor deposited in CBS and ATCC, and found that A. tortor cannot be distinguished from A. conoides, and treated A. tortor as the synonym of A. conoides. (Fig. 3.9)

Arthrobotrys conoides. Drechsler a–e conidia; f conidiophore; g adhesive network. Bars = 10 µm; Strain number: YMF1.00009
Arthrobotrys cystosporia (Dudd.) Sidorova, Gorlenko & Nalepina, Bot. Zh. SSR 49: 1598 (1964) |
Trichothecium cystosporium Dudd. Trans. Br. Mycol. Soc. 34: 600 (1952) |
Arthrobotrys cystosporia (Dudd) Mekht.[as ‘cystosporium’], Dokl. Akad, Nauk Azerb. SSR 20 (6): 70 (1964) |
Genicularia cystosporia (Dudd.) Rifai, Mycotaxon 2 (2): 215 (1975) |
Characteristics:
The mycelium consist of straight, septate hyphae, 3–6 μm wide, sparingly branched. Conidiophores hyaline, erect or geniculate due to repeated propagated, usually not branched, attaining a height of 100–400 μm. The loose group of spores having the appearance of a panicle thus formed at each conidiogenous loci. The conidia hyaline, broadly pyriform, 25–35 × 18–24 μm, 1-septate near the base, proximal cell much larger. No chlamydospores were observed. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
Japan, UK.
Notes:
The description based on the protologue. Conidia of this species are arranged loosely, not grown from typical denticles, but from sterigmata. (Fig. 3.10)

Arthrobotrys cystosporia. a conidiophore; b conidia. Bar = 20 µm
Arthrobotrys cookedickinson (Cooke & Dickinson) Z.F. Yu, comb. nov. |
Monacrosporium cystosporum Cooke & Dickinson, Trans. Br. Mycol. Soc. 48: 623 (1965) |
MB 804791 |
Characteristics:
Vegetative hyphae hyaline, septate, branched. Conidiophores hyaline, erect, simple, 8–11-septate, 60–420 μm long, 5–7.5 μm wide at the base, gradually tapering upwards to a width of 3–5 μm at the apex, bearing a single conidium. Conidia hyaline, broadly clavate or broadly turbinate to obovoid, 30–52.5 (42) × 15–22.5 (17.6) μm, 1–3-septate, mostly 2–3-septate with an almost globose terminal cell, distally rounded and tapered to a long tail formed by the two basal cells, the proximal one being narrowly truncate. Chlamydospores present. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Anhui, Guizhou, Sichuan, Xizang, Yunnan), Germany (former FRG)
Material examined:
②–1–1, isolated from field soil in Huaxi, Guizhou in 1996 by Ke-Qin Zhang; XZM–2, isolated from soil in Xizang in August 2000 by Minghe Mo; DH4–2, isolated from soil in Ruili, Yunnan in October 2002 by Jing Zhang; YMF1.00024, isolated from forest soil in Jiuzhaigou, Sichuan in 2003 by Xuefeng Liu. Permanent slide: ②–1–1
Notes:
This species is transferred to Arthrobotrys from Monacrosporium and renamed as A. cookedickinson to avoid confusing with A. cystosporia (Dudd.) Mekht. (1964). Cooke & Dickinson indicated A. cookedickinson was closely related to A. eudermata, but conidia of the latter were described as being much larger. Based on our observation, A. cookedickinson resembles A. sphaeroides in conidial shape, but A. cookedickinson is characterised by tail-like basal cell, which is absent in A. sphaeroides. (Fig. 3.11)

Arthrobotrys cookedickinson. a conidiophore; b conidia; c adhesive network. Bar = 30 µm strain number: YMF1.00024
Arthrobotrys dendroides Kuthub. Muid & J. Webster Trans. Br. mycol. Soc. 84 (3): 564 (1985) |
Characteristics:
Colonies on CMA whitish, effuse, with scattered erect hyaline synnemata. Mycelium superficial, hyaline, septate, branched, 2.5–4 μm wide. Conidiophores simple, septate, erect, solitary or synnematous; mainly synnematous on natural substrata, synnemata 0.5–1.2 mm wide, 1 cm tall; conidiophores sometimes elongating sympodially to form additional heads of conidia at higher levels; when synnematous individual threads slender, septate, smooth, straight except near the apex, splaying out to become flexuous near apex, individual conidiophores up to 800 μm long and 2–3 μm wide. Conidiogenous cells polyblastic, integrated, terminal, raduliform. Conidia 10–20 (14.6) × 2.5–5 (4) μm, hyaline, ovate to oblong, 1-septate. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Yunnan), Malaysia (Kuala Lumpur), USA
Material examined:
YMF1.00010, isolated from forest soil in Shuangbai, Yunnan in February 2001 by Xuefeng Liu. Permanent slide: Sb02–4.
Notes:
A. dendroides is different from other species of Arthrobotrys because synnematous conidiophores even in cultures. (Fig. 3.12)

Arthrobotrys dendroides. a, b conidiophore cluster; c–h conidia; i–j conidiophore; k adhesive network. Bars = 10 µm; Strain number: YMF1.00010
Arthrobotrys dianchiensis (Y. Hao & K.Q. Zhang) Z. F. Yu, comb. nov. |
Dactylella dianchiensis Y. Hao & K.Q. Zhang, Mycotaxon 82: 236 (2004) |
MB 804792 |
Characteristics:
Colony on PDA initially whitish and turning orange white after 10 days of incubation, rapidly growing and extending 7 cm in diameter at 28 °C, 5 cm at 25 °C within 4 days. At the same time the reverse side of the media becomes faintly yellow to reddish orange. Colonies on CMA whitish, rapidly growing and extending to a diameter of 6 cm at 25 °C within 6 days. Mycelium hyaline, scanty, hyphae septate, branched, 2.5–5 μm wide. Conidiophores erect, simple or branched, 2–4-septate, 245–425 μm long, 5–7.5 μm wide at the base, gradually tapering upwards to a width of 2.5–5 μm at the apex, initially a width of 2.5–5 μm at the apex, later often producing a few short branches or spurs near the apex, and bearing 2–3 conidia in sympodial arrangement. Conidia hyaline, spindle-shaped or clavate, narrowly round at the distal end, truncate at the base, 37.5–100 (70) × 10–17.5 (14.3) μm, 1–7-septate, mainly 2–5-septate. The proportion of conidia with 1, 2, 3, 4, 5, 6 and 7 septa is 10, 17, 14, 24, 22, 9 and 4 %, respectively. Some conidia had small tubercles at the both ends and could germinate from these tubercles. Conidia could produce secondary conidiophores and secondary conidia. The secondary conidia are spindle-shaped, 23.9 × 5 μm and 1-septate. Capturing nematodes by means of adhesive two dimensional networks.
Distribution:
China (Yunnan)
Material examined:
YMF1.00471, isolated from fresh water soil in Dianchi, Yunnan in 2002 by Yu’e Hao.
Notes:
The description is based on the protologue. A. dianchiensis is similar to A. multiformis in primary conidial shape, but it can be distinguished from A. multiformis by the size and septation of primary conidia. (Fig. 3.13)

Arthrobotrys dianchiensis. a–e conidiophore; f–h, i–j conidia; k conidiophore growing from conidia; l–m germinating conidia; n adhesive network. Bars = 10 µm; Strain number: YMF1.00571
Arthrobotrys eudermata (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 102 (1999) |
Dactylella eudermata Drechsler, Mycologia 42 (1): 40 (1950) |
Dactylella eudermata (Drechsler) Seifert & W.B. Kendr., in Seifert, Kendrick & Murase, Univ. Waterloo Biol. Ser. 27: 30 (1983) |
Genicularia eudermata (Drechsler) Rifai, Reintiwarda 7 (4): 367 (1968) |
Geniculifera eudermata (Drechsler) Rifai, Mycotaxon 2 (2): 216 (1975) |
Golovinia eudermata (Drechsler) Mekht., Dokl. Akad. Nauk Azerb. SSR 27 (2): 73 (1971) |
Monacrosporium eudermatum (Drechsler) Subram., J. Indian bot. Soc. 42: 293 (1964) [1963] |
Characteristics:
Colonies on CMA whitish, rapidly growing. Mycelium spreading, vegetative hyphae hyaline, septate, branched, 1.8–7.5 μm wide. Conidiophores hyaline, 2–8-septate, erect, simple, 400–680 μm long, 5–8 μm wide at the base, gradually tapering upwards to a width of 2–3 μm at the apex, bearing a single conidium or 2–4 additional conidia. Conidia globose to broadly turbinate or broadly spindle-shaped, rounded at the distal end and truncate at the base, the central cell the largest, 37–55 (49) × 17.5–35 (28) μm, 1–3-septate; microconidia ellipsoid, aseptate, rounded at the distal end and truncate at the base, 10–15 × 5–6 μm. Chlamydospores present on aged cultures, globose, forming in chain or clusters, smooth-walled, yellow. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Anhui, Beijing, Fujian, Guangxi, Hebei, Huibei, Huinan, Shangdong, Xizang, Yunnan, Zhejiang), Burkina Faso, Germany (Berlin-Dahlem), Spain (La Palma, Tenerife)
Material examined:
J42–1, J38–2, isolated from soil in Jianshui, Yunnan in 1999 by Yanju Bi; XZM–4, isolated from forest soil in Xizang in August 2000 by Minghe M; YMF1.00120, YMF1.00542, YMF1.00545, isolated from soil in Lijiang, Yunnan in September 2002 by Jing Zhang. Permanent slide: LJ3–4.
Notes:
A. eudermata resembles A. psychrophila and A. thaumasia in conidial shape, but differs in conidiophores and width of conidia, respectively. Conidia of A. eudermata are considerably wider than in A. thaumasia. Moreover, in A. eudermata, conidiophores are simple, bearing a single conidium at the apex, or 2–4 additional conidia, while in A. thaumasia, conidiophores are usually branched near the apex, bearing 3–15, sometimes up to 25 conidia on blunt sterigmata in loose capitate arrangement. The difference between A. eudermata and A. psychrophila is that conidia of A. psychrophila are longer than in A. eudermata. (Fig. 3.14)

Arthrobotrys eudermata. a–e conidia; f conidiophore; g adhesive network. Bars = 10 µm; Strain number: YMF1.00545
Arthrobotrys flagrans (Dudd.) Sidorova, Gorlenko & Nalepina, Bot. Zh. SSSR: 1598 (1964) |
Trichothecium flagrans Dudd. Trans. Br. mycol. Soc. 32 (3–4): 287 (1950) |
Arthrobotrys flagrans (Dudd.) Mekht., Dokl. Akad. Nauk Azerb. SSR 20 (6): 70 (1964) |
Duddingtonia flagrans (Dudd.) R.C. Cooke, Trans. Br. Mycol. Soc. 53 (2): 316 (1969) |
Characteristics:
Colonies effused, scanty, with few aerial hyphae. Conidiophores erect, straight, unbranched, attaining 150 μm long, bearing 2–6 conidia at the apex. Conidia obconical to ellipsoidal, 1-septate, the proximal cell broadly truncate, 25–50 × 10–15 μm. Intercalary chlamydospores present abundantly. Capturing nematodes by means of adhesive three dimensional hyphae.
Distribution:
Germany, Soviet, UK.
Notes:
The description is based on the protologue. This species has not typical denticles or sterigmata which exist among other species of Arthrobotrys, but it has typical adhesive three dimensional hyphae. (Fig. 3.15)

Arthrobotrys flagrans. a conidiophore; b conidia; c chlamydospores. Bars = 20 µm
Arthrobotrys fusiformis (R.C. Cooke & C.H. Dickinson) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 102 (1999) |
Monacrosporium fusiforme R.C. Cooke & C.H. Dickinson [as ‘fusiformis’], Trans. Br. mycol. Soc. 46: 628 (1965) |
Golovinia fusiformis (R.C. Cooke & C.H. Dickinson) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 160 (1979) |
Characteristics:
Vegetative hyphae hyaline, septate, branched. Conidiophores hyaline, erect, single, 3–7-septate, 250–390 μm long, 5 μm wide at the base, gradually tapering upwards to a width of 2.5–3 μm at the apex, bearing a single conidium, sometimes a second conidium formed on an about 15 μm branch near the conidiophore apex. Conidia hyaline, fusiform-ellipsoidal, 32.5–47.5 (41) × 12.5–17.5 (15.5) μm, 2–3-septate, rounded at the distal end and truncate at the base. Chlamydospores not observed. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Anhui, Hebei, Guizhou, Tianjing, Yunnan, Zhejiang), USA
Material examined:
YMF1.00034, isolated from field soil in Balitai, Tianjingin 2001 by Wenpeng Li. Permanent slide: LT–11–03–3
Notes:
According to Cooke & Dickinson (1965), A. fusiformis is similar to A. psychrophila and A. salina, but the dimensions, the shape and the septation of conidia distinguish it from these species. (Fig. 3.16)

Arthrobotrys fusiformis. a–b conidiophore; c–j conidia; k adhesive network. Bars = 10 µm; Strain number: YMF1.00034
Arthrobotrys gampsospora (Drechsler) S. Schenck, W.B. Kendr. & Pramer, Can. J. Bot. 55 (8): 982 (1977) |
Dactylaria gampsospora (Drechsler) de Hoog & Oorschot, Stud. Mycol.26: 110 (1985) |
Dactylaria gampsospora Drechsler, Sydowia 15 (1–6): 9 (1962) |
Monacrosporium gampsosporum (Drechsler) Xing Z. Liu & K.Q. Zhang, Mycol. Res. 98 (8): 865 (1994) |
Woroninula gampsospora (Drechsler) Mekht., Khishchnye Nematofagovye Griby- Gifomitsety (Baku): 112 (1979) |
Characteristics:
Mycelium colourless, branched, septate. Conidiophores erect, mostly 150–626 μm high, 5–7 μm wide at the base, tapering to 2–3 μm at the apex, bearing several conidia in a loose head on tooth–like projecting often 3–5 μm long. Conidia hyaline, spindle-shaped, curved, 1–4-septate, mostly 4-septate, 25–76 × 7–16 μm. Microconidiophores often 1–3-septate, 75–125 μm high, 4–5 μm wide at the base, tapering to a width of 1.2–2 μm long at the apex, furnished distally with several tooth-like projections 1.5–2 μm long on which bearing microconidia. Microconidia colourless, elongate obovoid, tapering towards the base, broadly rounded at the distal end, 10–17 × 4–6 μm. Chlamydospores present, 8–21 × 6–17 μm Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
Canada, USA.
Notes:
The description is based on the protologue. This species is different from other species of Arthrobotrys in its curved and more septate conidia. (Fig. 3.17)

Arthrobotrys gampsospora. a conidiophore; b conidia; c adhesive network. Bar = 20 µm
Arthrobotrys guizhouensis K.Q. Zhang, Acta Mycol. Sin. 13: 101 (1994) |
Characteristics:
Colonies on CMA initially whitish and turned to reddish, rapidly growing. Mycelium spreading, dense, vegetative hyphae hyaline, septate, branched, mostly 3–9 μm wide. Conidiophores hyaline, erect, branched, 5–10-septate, 120–370 μm long, 5–7.5 μm wide at the base, tapering upwards to a width of 4–5 μm before bearing on irregularly expanded apex whereon form conidia; subsequently often, following repeated elongation, giving rise successively to additional clusters of conidia. Two type of conidia appear, macroconidia fusoid to ellipsoidal, 34–42.5 × 14–17.5 μm, 1–3-septate, proportion of conidia with 1, 2, and 3 is 95, 4 and 1 %, respectively. Microconidia pyriform, 1-septate, 19–30 × 8.4–14 μm. Chlamydospores yellowish, globose, 7.5–12.5 μm in diameter. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Guizhou, Yunnan).
Material examined:
8736, isolated from soil in Guiyang, Guizhou in September 1987 by Ke-Qin Zhang; GZFJ-48, isolated from forest soil in Fanjing Mountain, Guizhou in 1996 by Ke-Qin Zhang;YMF1.00014, isolated from field soil in Wenshan, Yunnan in April 2002 by Zhiwei Zhao; LJ1–9-2, isolated from soil in Lijiang, Yunnan in September by Jun Zhang. Permanent slide: P4. 50–1.
Notes:
A. guizhouensis morphologically resembles A. oviformis and A. vermicola. However, A. guizhouensis differs from A. oviformis in irregularly expanded conidiophores at the apex, and larger conidia. It also differs from A. vermicola in branched and irregularly expanded conidiophores at the apex, and mostly 1-septate conidia. In A. vermicola, conidiophores are unbranched and not expanded at the apex, conidia are mostly 2–3-septate. (Fig. 3.18)

Arthrobotrys guizhouensis. a conidiophore; b conidia; c adhesive network. Bars = 30 µm
Arthrobotrys huaxiensis (K.Q. Zhang, Xing Z. Liu & L. Cao) Z.F. Yu, comb. nov. |
Monacrosporium guizhouense K.Q. Zhang, Xing Z. Liu & L. Cao, Mycol. Res. 100: 275, 1996 |
MB 804790 |
Characteristics:
Colonies on CMA pink. Hyphae hyaline, thin-walled, septate, frequently branched, 1.5–5 μm wide. Conidiophores hyaline, septate, single, simple, erect, 172–350 μm long, 3.5–6 μm wide at the base, gradually tapering upwards to a width of 2–2.5 μm at the apex, bearing a single conidium. Conidia hyaline, obpyriform to fusiform, 1–4-septate, mostly 2-septate, 30.5–71.5 (52.7) × 18.5–28.5 (23.9) μm. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Guizhou)
Material examined:
GAU1001, isolated from soil in Huaxi, Guizhou in October 1993 by Ke-Qin Zhang.
Notes:
This species traps nematodes by means of three dimensional adhesive networks, which was transferred from Monacrosporium to Arthrobotrys based on the present classification and renamed as A. huaxiensis Z.F. Yu to avoid confusing with A. guizhouensis K.Q. Zhang (1994). A. huaxiensis resembles A. cookedickinson and A. fusiformis in conidial septation, but differs in its fusiform to obpyriform conidia, conidial size and also in its unbranched conidiophores. (Fig. 3.19)

Arthrobotrys huaxiensis. a conidiophore; b chlamydospore; d–h conidia; i adhesive network. Bar = 10 µm; Strain number: YMF1.00014
Arthrobotrys indica (Chowdhry & Bahl) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 102 (1999) |
Monacrosporialla indica Chowdhry & Bahl [as ‘indicum’], Curr. Science 51: 895 (1982) |
Characteristic:
Vegetative hyphae hyaline, septate, branched, 2–3 μm wide. Conidiophores hyaline, erect, simple or occasionally branched, 8–20-septate, 150–350 μm long, 3–6 μm wide at the base, gradually tapering upwards to a width of 1–2 μm at the apex. Conidia hyaline, elliptic or broadly top–shaped, 0–2-septate, rounded at the distal end and truncate at the base, a navel at the base, 17.5–30 (23.2) × 12.5–20 (14.8) μm. Chlamydospores present. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Guizhou, Xizang), India (Delhi)
Material examined:
Sjz3.11.1, isolated from field soil in Huaxi, Guizhou in 1996 by Ke-Qin Zhang; XZM–7, isolated from soil in Xizang in August 2000 by Minghe Mo.
Notes:
A. indica resembles A. eudermata and A. cookedickinson in conidial shape, but the dimension of conidia in A. eudermata is larger than in A. indica. A. indica produces navels at the base of conidia while this character is absent in A. cookedickinson. (Fig. 3.20)

Arthrobotrys indica. a conidiophore; b–m conidia; n adhesive network. Bars = 10 µm; Strain number: XZM-7
Arthrobotrys janus (S.D. Li & Xing Z. Liu) Z.F. Yu, comb. nov. |
Monacrosporium janus S.D. Li & X.Z. Liu, Mycol. Res. 107: 890 (2003) |
MB 804791 |
Characteristics:
Colonies growing rapidly on PDA, attaining 8 cm diameter within 7 days at 25 °C, producing orange reddish pigment resulting in medium colored. Aerial mycelium fluffy, whitish when young and becoming light yellowish when aged. Hyphae hyaline, septate branched. Conidiophores hyaline, simple, slender, erect, 185–250 μm long, 5–6.5 μm wide at the base, tapering to 1.5–2.5 μm wide at the apex, bearing a single conidium. Conidia broadly turbinate to napiform, 1–2-septate but mostly (> 65 %) 1-septate at basal part, 15–26 (22.5) × 17.5–37.5 (28.5) μm. The distal cell are sphaerical and much bigger than others, proximal cell obconic, tapering to a short truncate base, central cell disciform if any. Chlamydospores sphaerical to ellipsoid, intercalary, 7.5–25 (15.5) × 4–11.5 (7.5) μm. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Shandong)
Material examined:
YMF1.01312, isolated from field soil in Mixian, Shandong in September 2000 by Shidong Li.
Notes:
A. janus closely resembles, but differs from, A. indica, A. sphaeroides and A. sinensis in conidial morphology. According to the original description of these species, the conidia of A. indica are 22–30 × 14–20 μm, and mostly 2-septate with distinct basal hila (Chowdhry and Bahl, 1982); those of A. sphaeroides are 28–42 × 19–29 μm, usually 2-septate with one basal and one terminal septum (46 %) or 1-septate with only one basal septum (29 %) (Castaner 1968); and those of A. sinensis are 25–30.5 × 15–18 μm with 37 % 3-septate, 42 % 2-septate, and 21 % 1-septate (Liu and Zhang 1994). While conidia of A. janus are 20–25 × 24.5–32.5 μm with 1–2 transverse septa, mostly (> 65 %) are 1-septate in the basal part. When compared morphologically, A. janus can be easily distinguished from A. sinensis by its conidial shape and septation. This fungus also similar in conidial size, shape and septation to A. cystosporia (conidia 24–35.5 × 18–25 μm), A. obovata (28.5–32 × 18–20.5 μm) and A. perpasta (24–32.5 × 12.5–20 μm), but differ in that the conidiophores of all the three latter species are geniculately branched (Duddington 1951a, Rifai and Cooke 1966, Zhang et al. 1996a). (Fig. 3.21)

Arthrobotrys janus. a–c. conidiophore; d–e conidia; f adhesive network. Bars: a = 35 µm, b–f = 20 µm; Strain number: YMF1.01312
Arthrobotrys javanica (Rifai & R.C. Cooke) Jarow., Acta Mycologica, Warszawa 6 (2): 373 (1970) |
Candelabrella javanica Rifai & Cooke, Trans. Br. mycol. Soc. 49 (1): 162 (1966) |
Characteristics:
Colonies on CMA almost hyaline; mycelium spreading, vegetative hyphae hyaline, septate, branched, smooth–walled, 3–7 μm wide. Conidiophores erect, straight, subulate, hyaline, smooth, 5–10-septate, 220–400 μm long, about 5–7.5 μm wide at the base, gradually tapering to 2.5–5 μm at the apex, enlarged slightly as ramification took place and bearing a lax, short, candelabrum-like apical branching system. Conidia arose singly as blown-out ends of the conidiophore and of the subsequently developed growing points, 20–37.5 (27.9) × 7.5–10 (8.8) μm, smooth-walled and hyaline, narrowly obovoid or clavate, 1 septum, slightly constricted at the septum, rounded distally with the obconical basal cell tapering to a truncate base. Chlamydospores yellowish, subglobose, 7.5–15 μm in diameter. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Guizhou, Xizang, Yunnan), France (Limousin), Indonesia (Java)
Material examined:
Dian5 (4), isolated from fresh water soil in Dianchi Lake, Yunnan in 1996 by Ke-Qin Zhang et al; YMF1.00015, YMF1.00548, isolated from field soil in Lincang, Yunnan in April 1999 by Lu Cao; MW17–2, isolated from forest soil in Xishuangbanna, Yunnan in 1999 by Yanju Bi; XZA–5, isolated from field soil in Xizang in August 2000 by Minghe Mo; DH6–2, isolated from soil in Ruili, Yunnan in October 2002 by Jing Zhang. Permanent slide: LL4–1.
Notes:
Rifai and Cooke (1966) established a new genus Candelabrella Rifai and Cooke (1966) based on long, distinctive, subcylindrical conidial pegs and the candelabrum-like branching of the conidiophore. However, this genus was not accepted by most scholars, so this species was transferred into Arthrobotrys (Jarowaja, 1970). A. javanica resembles A. musiformis in the candelabrum-like branching of the conidiophores, but differs in conidia shape. In A. javanica, conidia are 20–37.5 (27.9) × 7.5–10 (8.8) μm, narrowly obovoid or clavate, while in A. musiformis, conidia ellipsoid, mostly slightly curved, 20–47.5 (30.9) × 7–12.5 (10.3) μm. (Fig. 3.22)

Arthrobotrys javanica. a–b conidiophore; c–f conidia; g adhesive network. Bars = 10 µm; Strain number: YMF1.00015
Arthrobotrys latispora Hong Y. Su & X.Y. Yang, in Su, Liu, Li, Cao, Chen & Yang, Mycotaxon 117: 32 (2011) |
Characteristics:
Colonies grew rapidly on CMA medium, attaining 6 cm diameter in 8 days at 25 °C. Mycelium spreading, vegetative hyphae hyaline, septate and branched, mostly 2–4 μm wide. Conidiophores colourless, erect, simple, septate frequently, 60–120 μm high, 2–5 μm wide at the base and 1.8–4 μm at the apex, producing 4–12 conidia singly from conidiogenous loci on conspiculus, partly superposed nodes at and near the apex. Conidia colourless, broadly ovoid-oval, broadly rounded at the apex, rounded truncate at the narrowed base, 1-septate or aseptate at the centre, 14.8–21.5 × 10.1–16.3 (average18.3 × 13.5) μm (living state). Approximately 41 % of the conidia were non-septate and 59 % 1-septate. Chlamydospores sphaerical to ellipsoidal. Nematodes are trapped by three dimensional adhesive networks.
Distribution:
China (Yunnan)
Material examined:
YMF1.03168, isolated from soil in Yongping, Yunnan in October 2009 by Hongyan Su.
Notes:
A. latispora is characterized by its broad, ovoid or slightly oval, partly aseptate conidia, which are borne on nodes. Based on conidial shape, A. latispora most closely resembles A. amerospora. The conidia of both species are ovoid, but differ in conidial size and septation. (Fig. 3.23)

A. latispora. a–e conidiophore; f–g conidia; h adhesive network. Bars = 10 µm; strain number: YMF1.00538
Arthrobotrys longiphora (Xing Z. Liu & B.S. Lu) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 102 (1999) |
Monacrosporium longiphorum Xing Z. Liu & B.S. Lu, Mycosystema 6: 65 (1993) |
Characteristics:
Colonies on CMA white, growing moderately. Mycelium effuse, vegetative hyphae smooth-walled, hyaline, septate. Conidiophores hyaline, erect, septate, simple or sparsely branched, up to 200–550 μm long, 5 μm wide at the base, gradually tapering upwards to a width of 2–3 μm at the apex, bearing a single conidium. Conidia hyaline, fusiform, 2–5-septate, mainly 3–4-septate, conidia holoblastic, 40–90 (54) × 15–27.5 (18) μm. Chlamydospores not observed. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Hebei, Sichuan, Xigjiang, Yunnan)
Material examined:
YMF1.00538, isolated from soil in Baoshan, Yunnan in October 2002 by Jing Zhang. Permanent slide: BS5–19
Notes:
This taxon most closely resembles A. psychrophila in having 3–4-septate conidia and in conidial shape, but the conidia size especially width of the original description of A. longiphora (38–65 × 13–22 μm) is less than that of A. psychrophila (46–71 × 21–29 μm). The taxon also resembles A. reticulata and A. megalospora in spore shape, but differs in that the taxon has 3–4-septate conidia, while A. reticulata and A. megalospora have mainly 4-septate conidia. (Fig. 3.24)

Arthrobotrys longiphora. a–b conidiophore; c–e conidia; f adhesive network. Bars = 10 µm. strain number: YMF1.00538
Arthrobotrys mangrovispora Swe, Jeewon, Pointing & K.D. Hyde, Bot. Mar. 51 (4): 332 (2008) |
Characteristics:
Mycelium spreading, scanty, irregularly projecting, hyaline, septate, branched. Conidiophores 70–330 μm high, 1.5–5 μm wide at the base, hyaline, erect, septate, bearing first a single conidium at the apex, later additional conidia from the lateral branches or longer geniculate branches below the apex, commonly producing 1–6 conidia; each long denticle or branch bearing a single conidium. Conidia 25– (38.9)–50 × 12– (17.3)–24 μm, colourless, variable in shape; broadly turbinate to elongate-fusoid, ellipsoidal or fusiform-ellipsoidal, clavate, broadly obovoid, elongate–obovoid, 1–3-septate; proportions of conidia with 0, 1, 2 or 3 septa are 6, 23.5, 50.0 and 20.5 %, respectively. Chlamydospores present. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Hong Kong)
Notes:
The description is based on the protologue. A. mangrovispora closely resembles, but differs from, A. eudermata, A. sphaeroides, A. microscaphoides and A. scaphoides in conidial morphology. According to the original descriptions of these species, the conidia of A. eudermata are 31–40 × 19–28.5 μm, and mostly 3-septate (Chowdhry and Bahl 1982). Those of A. sphaeroides are 28–42 × 19–29 μm, usually 2-septate with one basal and one terminal septum or 1-septate with only one basal septum (Castaner 1968), and those of A. scaphoides are 26–83 × 12–17 μm with 1–3 septa (Peach 1952). A. microscaphoides has conidia that are 25–50 × 12–24 μm with 1–3 transverse septa and most (53 %) are 2-septate. Of the above mentioned species, A. mangrovispora most closely resembles A. scaphoides, but can be distinguished by the following differences: A. scaphoides forms broadly fusiform conidia with 3 septa: non-septate microconidia and chlamydospores are absent, while A. mangrovispora possesses broadly turbinate to elongate-fusiform conidia and produces chlamydospores. (Fig. 3.25)

Arthrobotrys mangrovispora. a conidia; b conidiophore; c adhesive network. Bars = 5 µm
Arthrobotrys megalospora (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 103 (1999) |
Dactylella megalospora Drechsler, Mycologia 46: 769 (1954) |
Golovinia megalospora (Drechsler)Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 155 (1979) |
Monacrosporium megalospora (Drechsler) Subram., Kavaka 5: 94 (1978) [1977] |
Monacrosporium megalospora (Drechsler) Subram., J. Indian bot. Soc.42: 293 (1964) [1963] |
Characteristics:
Mycelium spreading; vegetative hyphae colourless, branched, septate at moderate intervals, mostly 2–6 μm wide. Conidiophores colourless, erect, often containing 1–7 cross walls, commonly 6–8 μm wide at the base, tapering gradually upwards to a distal width of 2–3 μm, 350–450 μm high, unbranched and bearing a single conidium at the apex. Sometimes 1–5 conidia formed on 10–40 μm long branch near the conidiophore apex. Conidia colourless, broadly fusoid or elongate-ellipsoidal or obovoid, mostly 40–75 long and 18–35 wide, commonly divided by cross-walls into 3–6 cells one of which greatly exceeds the others in length and width; the large cell somewhat variable in position, but occurring most often in median position in the especially distinctive large, broadly fusoid, quadriseptate conidia, frequently 55–75 μm long, and 23–35 μm wide, that are produced singly on nematode-infested substratum. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
USA (Florida)
Notes:
The description is based on the protologue. This species resembles A. psychrophyla in conidiophore and conidial shape, but conidia of the latter mainly 3–4-septate, conidia of the former are 2–5-septate. (Fig. 3.26)

Arthrobotrys megalospora. a conidiophore; b conidia; c adhesive network. Bars = 30 µm
Arthrobotrys microscaphoides (Xing Z. Liu & B.S. Lu) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 103 (1999) |
Monacrosporium microscaphoides Xing Z. Liu & B.S. Lu, Mycosystema 6: 68 (1993) |
Characteristics:
Colonies on CMA hyaline. Mycelium effuse, vegetative hyphae smooth-walled, hyaline, septate, and branched. Conidiophores hyaline, erect, septate, simple or with several branches normally at the apex, 230–460 μm long, 3–5 μm wide at the base, gradually tapering upwards to a width of 2–2.5 μm at the apex, bearing a single conidium or occasionally several conidia. Conidia hyaline, cymbiform, 0–3-septate, mostly 2-septate, 22.5–45 (27.2) × 10–20 (13.9) μm. The proportion of conidia with 0, 1, 2, and 3 septa is 4, 27, 55, 7 %, respectively. Chlamydospores present in aged cultures, in chain. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Beijing, Hainan, Henan, Sichuan, Yunnan)
Material examined:
YMF1.00028, isolated from soil in Lincang, Yunnan in 1999 by Lu Cao; JLCB, isolated from forest soil in Changbai Mountain, Jilin in 1999 by Ke-Qin Zhang; YMF1.00546, isolated from soil in Yinjiang, Yunnan in October 2002 by Jing Zhang. Permanent slide: L32–6
Notes:
This fungus most resembles A. scaphoides in its fusiform or boat-like shaped spores, but the conidia of A. microscaphoides is smaller than those of A. scaphoides. Although the conidia of A. scaphoides are 23–39 × 12–17 μm in Peach’s (1952) description covered the range of A. microscaphoides 23–39 × 8–15.5 μm in original description (Liu and Lu 1993), but the average length and width of conidia of A. scaphoides is much longer than that of A. microscaphoides according to our measurement of the conidia of the figure in Peach’s paper. (Fig. 3.27)

Arthrobotrys microscaphoides. a–e conidia; f conidiophore; g nematode trapped by adhesive network. Bars = 10 µm; Strain number: YMF1.00546
Characteristics:
Colonies on CMA whitish, rapidly growing and extending to a diameter of 7–8 cm at 25 °C within 7 days. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched. Conidiophores hyaline, simple, erect, septate, 80–220 μm long, bearing a single conidium at the apex. Conidia elongate-fusiform, 4–12-septate, 47–198 × 7–20 μm, solitary. Conidia may raise to secondary conidiophores up to 50–120 μm long, on apices of which bearing clavate to cylindric-clavate, 0–1 septum, secondary conidia, 15–35 × 2.5–5 μm. Chlamydospores present in older cultures. Capturing nematodes by three dimension adhesive networks.
Distribution:
Canada (Manitoba), China (Hebei, Guizhou, Yunnan), Japan
Material examined:
CBS773.84 (A. multiformis), IFO32554 (A. iridis), 89019–1 (A. iridis), AN–1101 (A. iridis), provided by Dr. Xingzhong Liu. YMF 1.01477, isolated from soil in Xundian, Yunnan in April 2004 by Ye’e Hao. Permanent slide: YMF1.01477.
Notes:
A. multiformis resembles Dactylellina leptospora in its rather delicate, spreading mycelium and terminally produced, single conidia which are mostly straight, elongate-fusiform to cylindrical, and with many septa; both taxa produce secondary conidia. However, Da. leptospora traps nematodes by non-constricting rings and adhesive knobs, while A. multiformis traps by means of adhesive three-dimensional networks. Conidia of Da. leptospora are 40–105 × 4–5.8 μm, with 5–15 septa; those of A. multiformis, 35–90 × 4–7.5 μm with 4–12 septa. Secondary conidia of Da. leptospora are somewhat clavate 20–50 × 4–5.8 μm, 3–8-septate; those of A. multiformis are cylindric to clavate, 20–25 × 5 μm, 0–1-septate, and borne singly or on short sympodially proliferated denticles. A. multiformis, Dactylella ramiformis and Dactylella iridis share most morphological characteristics based on our observed strain, so we treat them as conspecific under the name A. multiformis. (Fig. 3.28)

Arthrobotrys multiformis. a–e conidiophore; f–k conidia; l chlamydospore; m adhesive network. Bars = 10 µm; Strain number:YMF1.01477
Arthrobotrys nonseptata Z.F. Yu, S.F. Li & K.Q. Zhang, Mycotaxon 109: 249 (2009) |
Arthrobotrys musiformis Drechsler, Mycologia 29 (4): 481 (1937) |
Candelabrella musiformis (Drechsler), Rifai & R.C. Cooke, Trans. Br. mycol. Soc. 49 (1): 163 (1966) |
Dactylella musiformis (Drechsler) Matsush., Microfungi of the Solomon Islands and Papua-New–Guinea (Osaka): 22 (1971) |
Characteristics:
Colonies on CMA whitish, rapidly growing. Mycelium spreading, vegetative hyphae, septate, branched, mostly 1–10 μm wide. Conidiophores hyaline, septate, erect, not branched blow, 104–640 μm long, 5–7.5 μm wide at the base, gradually tapering upwards to a width of 2.5–5 μm near the apex, where are borne on divergent, slightly tapering, simple or branched sterigmata, mostly 1–2.5 μm wide, 3–35 μm long, usually 5–15 conidia in loose capitate arrangement. Conidia hyaline, ellipsoid, straight or slightly curved, broadly rounded at the wider distal end, tapering noticeably towards the slightly protruded base, 20–47.5 (30.9) × 7–12.5 (10.3) μm, 1-septate close to the base of the spore. Chlamydospores yellow, globose or less frequently ellipsoidal, mostly 14–22 μm in diameter. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
Brazil (Sao Paulo), China (Anhui, Beijing, Guangxi, Guizhou, Hubei, Shangdong, Sichuan, Taiwan, Xizang, Yunnan), Ecuador (Tungurahua), France (Limousin), Netherlands (Den Haag, Meerdink–bos near Winterswijk), Nigeria (Ibadan), UK (Scotland), USA (Maryland, Virginia)
Material examined:
Z327, Z377, isolated from soil in Guiyang, Guizhou in 1992 by Ke-Qin Zhang; H101, H093, isolated from field soil in Kunming, Yunnan in 1991 by Ke-Qin Zhang; GZFJ–15, AHHS–22, isolated from soil in Fanjing Mountain, Guizhou and Huangshan Mountain, Anhui in 1996 by Ke-Qin Zhang; DL2–2, isolated from forest soil in Dali, Yunnan in September 2002 by Jing Zhang; XM43–1, isolated from forest soil in Xiangmi, Yunnan in 1999 by Yanju Bi; XZA–6, isolated from humus in Xizang in August 2000 by Minghe Mo; YMF1.00581, YMF1.00122, YMF1.00575, isolated from forest soil in Ruili and Lijiang, Yunnan in October 2002 by Jing Zhang. Permanent slide: 187.
Notes:
A. musiformis resembles Drechslerella dactyloides in having ellipsoid, straight or slightly curved conidia, but differs in trapping-devices. The former captures nematodes by three dimensional adhesive networks, while the later captures nematodes by constricting rings. A. musiformis also resembles A. javanica in candelabrum-like branching of the conidiophore and trapping-devices, but differs in conidia. In A. musiformis, conidia are ellipsoid, mostly slightly curved, 20–47.5 (30.9) × 7–12.5 (10.3) μm, while in A. javanica, conidia are 20–37.5 (27.9) × 7.5–10 (8.8) μm, narrowly obovoid or clavate.
Strain H093 can produce microconidiophores and microconidia from mycelium or from conidia. Microconidiophores 40–50 μm long, microconidia oviform, single-cell, 12–14 × 4.8 µm. These characters were not described in previous references, but other characters match A. musiformis. (Fig. 3.29)

Arthrobotrys musiformis. a–c conidia; d conidiophore; e adhesive network. Bars = 10 µm; Strain number: YMF1.00575
Characteristics:
Colonies slow-growing on CMA medium, attaining less than 35 mm diameter in 10 days at 25 °C. Vegetative hyphae hyaline, septate, 3.5–4 μm wide, aerial mycelium sparse, hyaline, septate, branched, 2.5–4 μm wide. Conidiophores erect, septate, unbranched, 40–120 μm high, 2–4 μm wide in the lower part, 1.5–2 μm wide at the apex, producing 3–10 conidia from retrogressive conidiogenous loci on conspicuous denticles at and near the apex. Conidia hyaline, aseptate, 11–16.8 × 5–6.6 μm, elongate ellipsoid, constricted at the base by forming a small truncate protuberance. Nematodes are captured by means of three dimensional adhesive networks.
Distribution:
China (Yunnan)
Material examined:
YMF1.01852, Yimen, Yunnan, Yimen, in Aug. 2006 by Z.F. Yu.
Notes:
There are five known species of Arthrobotrys with aseptate or occasionally 1-septate conidia. Conidia of A. amerospora (Schenck et al. 1977) are consistently non-septate, those of A. anomala (Barron and Davidson 1972), A. botryospora (Barron 1979) and A. yunnanensis (Mo et al. 2005) are occasionally 1-septate. A. nonseptata differs from other four species in conidial shape and size. Among four species, A. nonseptata most resembles A. yunnanensis in regard to conidial shape. Both species were anamorph state of Orbilia sp., and conidia vary in shape, but part conidia of both species are elongate, ellipsoid. In addition, the denticles of A. yunnanensis are longer than those of A. nonseptata. (Fig. 3.30)

Arthrobotrys nonseptata. a–c conidiophore; d conidia; e adhesive network. Bars: a–b, d = 10 µm, c = 50 µm, e = 20 µm
Arthrobotrys oligospora Fresen., Beitr. Mykol. 1: 18 (1850) |
Arthrobotrys superba var. oligospora (Fresen.) Coem. |
Didymozoophaga oligospora (Fresen.) Soprunov & Galiulina, Mikrobiol., Reading 20: 493 (1951) |
Arthrobotrys obovata K.Q. Zhang & Xing Z. Liu, in Zhang, Liu, Cao & Gao, Mycol. Res. 100 (5): 529 (1996) |
Characteristics:
Colonies on CMA initially whitish and turned to tinged pink after about one month, rapidly growing, attaining a diameter of 9 cm within 5 days at 25 °C. Mycelium hyaline, vegetative hyphae septate, branched. Conidiophores single or branched, erect, septate, 200–400 μm long, 5–7.5 μm wide at the base, gradually tapering to 2.5–3 μm wide below the first node, branches up to 8–38 μm long, conidiogenous loci with denticle nodes; conidiophores usually continue development by repeated elongation and successive production of 2–9 additional denticle nodes, often containing 5–15 conidia in loose capitate arrangement. Conidia hyaline, obovoid, 1-septum in about one third from the basal end, broadly rounded at the apex, obconical at the base, not constricted at the septum, 28.5–32 (30) × 18–20.5 (20) μm. The basal cells are very small, usually 5–6.5 μm long, and the distal cells are much larger, usually 20.5–25.5 μm long. Chlamydospores absent. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Guizhou, Yunnan)
Material examined:
CBS618.95. YMF1.00011, isolated from forest soil in Fanjing Mountain, Guizhou in June 1994 by Ke-Qin Zhang. Permanent slide: FJ92.
Notes:
This fungus closely resembles A. cystosporia and A. perpasta in conidial shape and size but differs in conidiogenous loci and conidial arrangement. In A. cystosporia and A. perpasta, conidiogenous loci are simple denticles, and conidia are widely spaced arrangement, while in A. obovata, conidiogenous loci are denticle nodes and conidia are loose capitate arrangement. (Fig. 3.31)

Arthrobotrys obovata. a–b conidiophore; c–h conidia; i adhesive network. Bars = 10 µm; Strain number: YMF1.00011
Characteristics:
Colonies on CMA whitish or reddish, rapidly growing. Mycelium spreading, dense, vegetative hyphae hyaline, septate, branched. Conidiophores single, erect or curved slightly, septate, 110–440 μm long, 5–10 μm wide at the base, gradually tapering upwards to a width of 3.5–5 μm below the first node; conidiophores usually continue development by repeated elongation and successive production of 1–6 additional denticle nodes, where on forming several conidia. Conidia hyaline, pyriform or obovoid, 1-septate, constricting at the septum, broadly rounded at the apex, 17–35 (23) × 8.5–16 (12) μm. The basal cell is smaller than the distal cell. Chlamydospores yellow, globose, thick-walled, coarse, 15–24 μm in diameter. Capturing nematodes by means of adhesive three dimensional networks.
Distribution:
cosmopolitan
Material examined:
Z254, isolated from root rubble in Zunyi, Guizhou in 1992 by Ke-Qin Zhang et al.; H014, H058, isolated from forest soil in Guiyang, Guizhou in 1992 by Ke-Qin Zhang et al; ZE102, isolated from Oxford, UK in 1991 by Ke-Qin Zhang; GZDF–3, isolated from field soil in Dafang, Guizhou in 1996 by Ke-Qin Zhang; BJXS–5, isolated from forest soil in Fragrant Mountain, Beijing in 1996 by Ke-Qin Zhang; J8–1, isolated from soil in Jianshui, Yunnan in 1999 by Yanju Bi; XZA–7, isolated from pasture soil in Xizang in August 2000 by Minghe Mo; DL1–3, isolated from soil in Dali, Yunnan in September 2002 by Jing Zhang; YMF1.00012, YMF1.00110, YMF1.00544, YMF1.00051, YMF1.00056, isolated from soil in Dali and Deqin, Yunnan in September 2002 by Jing Zhang. Permanent slide: SJt3.10.8.
Notes:
A. oligospora resembles A. conoides and A. superba in most morphological characters except conidia shape and the position of septum. In A. oligospora, conidia are obovoid, 1-septate near the base of the spore, and constricted at the septa. In A. conoides, conidia elongate obconical, 1-septate about one third from the basal end, and constricted at the septum. In A. superba, conidia are elliptical, 1-septate at the centre of the spore, and not constricted at the septum. (Fig. 3.32)

Arthrobotrys oligospora. a, e conidiophore; b–d conidia; f adhesive network. Bars = 10 µm; Strain number: YMF1.00544
Arthrobotrys oudemansii M. Scholler Hagedorn & A. Rubner, Sydowia 52 (1): 60 (2000) |
Arthrobotrys elegans (Oudem.) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 102 (1999) |
Golovinia elegans (Oudem.) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 142 (1979) |
Monacrosporium elegans Oudem., Aanwinsten Fl. Mycol. Nederl. 4: 48 (1884) |
Characteristics:
Conidiophores 150–265–400 μm high, bearing a single conidium on the apex or some additional conidia on small perpendicular and sometimes longer, geniculate branches. Conidia top-shaped, 40–52–65 × 17–20–23 μm, commonly three-septate, the inequidistantly distributed septa demarcating a small apical cell, a strongly ventricose and large central cell, a parabasal and a basal cell. Microconidia obovoid, protracted at the base, 13–16.5 × 5–6 μm, aseptate. Chlamydospores not observed. Trapping nematodes by means of three dimensional networks.
Material examined:
CBS 299.94, from soil of a laurel forest, Las Mercedes, Tenerife, coll./isol, G.Lysek,1989. CBS300.94, from moss cushion, Berlin–Dahlem, coll./isol.S.Behnke, 1992, by Rubner (1996).
Distribution:
Germany, Netherlands, Sweden.
Notes:
This species is the lectotype species of Monacrosporium. The size of conidia of all other isolations are within that of type strain, only some strains have branches. (Fig. 3.33)

Arthrobotrys oudemansii. a conidia; b microconidia; c conidiophores. Bars: a = 30 µm, b = 10 µm, c = 50 µm
Arthrobotrys oviformis Soprunov Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 136 (1958) |
Nematophagus oviformis (Soprunov) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 106 (1979) |
Characteristics:
Colonies on CMA whitish or yellowish. Mycelium hyaline, vegetative hyphae branched, septate. Conidiophores hyaline, 6–9-septate, branched, 200–600 μm long, 2.5–5 μm wide, conidiogenous loci with denticle nodes; conidiophores usually continue development by repeated elongation and successive production of several additional denticle nodes, where on forming 20–39 conidia. Conidia pyriform, broadly rounded at the apex, 1–2-septate, mainly 1-septate at the centre of the spore, slightly constricted at the septum, 7.5–22.5 (15.8) × 5–10 (6.6) μm. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
Burkina Faso (Houet), China (Anhui, Beijing, Guangxi, Guizhou, Hebei, Heilongjiang, Hubei, Neimeng, Xizang, Yunnan), Russian.
Material examined:
Z12, isolated from soil in Yushan, Guangxi in 1987 by Ke-Qin Zhang; 8739, isolated from field soil in Huaxi, Guiyang in 1988 by Ke-Qin Zhang; BJHD–3, isolated from soil in Haidian, Beijing, in 1996 by Ke-Qin Zhang; XbL15, isolated from forest soil in Kunming, Yunnan in April 1999 by Lu Cao; XZA–8, isolated from pasture soil in Xizhang in August 2000 by Minghe Mo; YMF1.00121, DL1–3, isolated from forest soil in Dali, Yunnan in September 2002 by Jing Zhang. Permanent slide: XbL15–3.
Notes:
A. oviformis resembles A. oligospora in almost all morphological characters except branched conidiophores. Basically the conidia of A. oviformis are not easy to distinquish from those of A. oligospora. However, A. oviformis has 2-septate conidia, in A. oligospora conidia are consistently 1-septate. (Fig. 3.34)

Arthrobotrys oviformis. a–c conidia; d–e conidiophore; f adhesive network. Bars = 10 µm; Strain number: YMF1.00121
Arthrobotrys paucispora (R.C. Cooke) Jarow., Acta Mycologica, Warszawa 6 (2): 381 (1970) |
Genicularia paucispora R.C. Cooke, in Rifai & R.C. Cooke, Trans. Br. Mycol. Soc. 49: 157 (1966) |
Genicularia paucispora (R.C. Cooke) Rifai, Mycotaxon 2 (2): 215 (1975) |
Characteristics:
Conidiophores simple, erect, septate, straight at first but sometimes becoming geniculate through repeated subapical proliferation, 300 μm long and 4 μm wide, bearing widely spaced conidia. Conidia narrowly obpyriform, smooth, thin-walled, 1-septate, 24–40 × 12.5–18.8 μm, with a distinct constriction at the septum, distally rounded and tapered below to an obconical truncate base, In pure culture on cornmeal agar they were smaller, 25–33.8 × 12.5–16.3 μm and were sometimes curved or asymmetrical. capturing nematodes by means of three dimensional adhesive networks.
Distribution:
UK.
Notes:
The description is based on the protologue. This species has not typical denticles, conidia of which borne from small sterigmata of conidiogenous loci. (Fig. 3.35)

Arthrobotrys paucispora (R.C. Cooke) Jarow a geniculate conidiophore with conidia; b conidiophores and terminal conidium. Bar = 20 µm
Arthrobotrys perpasta (R.C. Cooke) Jarow., Acta Mycologica, Warszawa 6 (2): 381 (1970) |
Genicularia perpasta R.C. Cooke, in Rifai & R.C. Cooke, Trans. Br. Mycol. Soc. 49: 156 (1966) |
Genicularia perpasta (R.C. Cooke) Rifai, Mycotaxon 2 (2): 216 (1975) |
Characteristics:
Conidiophores erect, straight, septate, hyaline and typically unbranched, up to 400 μm long, later became geniculate or sinuous, bearing widely spaced conidia. Conidia plump, obpyriform, 24–32.5 × 12.5–20 μm, smooth and thin-walled, hyaline or subhyaline, 1-septate, sometimes constricted at the septum. In pure culture on cornmeal agar conidia were smaller, 21.5–30 × 12.5–19 μm. Capturing nematodes by simple, branched adhesive hyphae.
Distribution:
UK.
Notes:
The description is based on the protologue. This species resembles A. cystosporia in conidiogenous structure and conidial shape, but differs from the latter by trapping device and the size of conidia. (Fig. 3.36)

Arthrobotrys perpasta. a conidiophore; b conidium. Bar = 20 µm
Arthrobotrys polycephala (Drechsler) Rifai, Reinwardtia 7 (4): 371 (1968) |
Dactylaria polycephala Drechsler, Mycologia 29: 530 (1937) |
Woroninula polycephala (Drechsler) Mekht, Khishchnye Nematofagovye Griby— Gifomitsety (Baku): 110 (1979) |
Characteristics:
Colonies on CMA whitish, rapidly growing and extending to a diameter of 10 cm at 25 °C within one week. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched, mostly 1.8–4.5 μm wide. Conidiophores hyaline, septate, at first erect, later often more or less procumbent, 90–270 μm long, 4–5 μm wide at the base, gradually tapering upwards to a width of 2.5–3 μm below the first denticle node producing 1–5 conidia, then following repeated elongation often giving rise successively to additional conidial clusters. Initially forming ellipsoidal-shaped conidia, hyaline, rounded at the distal end, somewhat acute at the base,1–3-septate, 12.5–25 (20) × 3–5 (4.5) μm; then forming fusoid-shaped conidia, 2–4-septate, occasionally 2-septate, often 4-septate, most often 3-septate, sometimes slightly constricted at the septa, 30–50 (45.1) × 8–16.5 (12.2) μm; proportion of conidia with 2, 3 and 4 septa is 3, 79 and 18, respectively. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Hubei, Xizang, Yunnan), Cuba, Germany (Berlin), USA (Maryland)
Material examined:
W7230–3–06, isolated from field soil in Wuhan, Hubei in February 2001 by Xuefeng Liu et al; XZA–9, isolated from forest soil in Xizang, in August 2000 by Minghe Mo; YMF1.00035, YMF1.00600, isolated from forest soil in Jiuzhaigou, Xichuan in April 2004 by Xuefeng Liu. Permanent slide: W7230–3–06.
Notes:
The conidia of A. polycephala slightly resemble those of A. musiformis in general shape, but differ in having larger and more abundant septate. The fungus was initially assigned to Dactylaria based on the plural septation of its conidia (Drechsler 1937). However, it was then transferred to Arthrobotrys based on trapping devices, nodose and repeatedly capitate forms of conidiophores (Rifai 1968). (Fig. 3.37)

Arthrobotrys polycephala. a–c conidiophore; e–m conidia; d chlamydospores; n adhesive network. Bars = 10 µm; Strain number: YMF1.00035
Arthrobotrys pseudoclavata (Z.Q. Miao & Xing Z. Liu) J. Chen, L.L. Xu, B. Liu & Xing Z. Liu, Fungal Diversity 26 (1): 123 (2007) |
Dactylella pseudoclavata Z.Q. Miao & Xing Z. Liu, Can. J. Bot. 81: 453 (2003) |
Characteristics:
Colonies on CMA, slightly brown on top and light brown to yellowish below, effuse extending to a diameter of 7.5 cm at 25 °C within 7 days. Mycelium spreading, vegetative hyphae hyaline, septate, branched, prostrate, 2.5–7 μm wide. Conidiophores erect, simple or occasionally branched, hyaline, septate, 150–300 μm long, 6–7.5 μm wide at the base, gradually tapering upwards to a width of 4–5.5 μm at the apex. The conidiogenous cell produced 1–4 conidia holoblastically. Conidia 30–45 × 8–11 μm, obclavate, round to blunt at distal end, constricted into bottleneck shape at the base, with zero or one septum, which was generally at lower centre or at the septum. Some conidia broke into two parts at the septum. Chlamydospores were abundant in aged culture on PDA, intercalary or catenulate, yellowish to brown, globate or subglobate. Capturing nematodes by means of three dimensional adhesive networks.
Distribution
China (Beijing)
Material examined:
HMAS 84442, isolated from field soil in Haiding, Beijing in February 1998 by Zuoqing Miao.
Notes:
A. pseudoclavata is characterized by obclavate, 0–1-septate conidia with round distal end and bottleneck-shaped base, simple conidiophores bearing 1–4 conidia at apex, and adhesive networks as the nematode-trapping device. These characters can distinguish it from other Arthrobotrys species. (Fig. 3.38)

Arthrobotrys pseudoclavata. a–p conidia; q–s conidiophore; t–u conidiophore without protoplasm; v conidia with breakpoint at septum; w chlamydospore; x adhesive network; y nematode trapped by adhesive network. Bars = 20 µm
Arthrobotrys psychrophila (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 104 (1999) |
Dactylaria psychrophila Drechsler, Mycologia 36 (2): 161 (1944) |
Genicularia psychrophila (Drechsler) Rifai, Reinwardtia 7 (4): 367 (1968) |
Geniculifera psychrophila (Drechsler) Rifai, Mycotaxon 2: 216 (1975) |
Golovinia psychrophila (Drechsler) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 151 (1979) |
Monacrosporium psychrophilum (Drechsler) R.C. Cooke & C.H. Dickinson, Trans. Br. mycol. Soc. 48: 622 (1965) |
Characteristics:
Mycelium spreading; vegetative hyphae hyaline, septate, mostly 2–6 μm wide. Conidiophores hyaline, erect, septate, mostly 150–500 μm high, 5–9 μm wide at the base, 2.5–4.5 μm wide at the apex, sometimes simple and sometimes bearing near the apex a branch (occasionally 2 branches) up to 35 μm long, often on giving rise terminally to 1–2 conidia elongating once or twice to produce 1 or 2 additional conidia, and thus frequently coming to bear 3 or 4 conidia in a loose head. Conidia hyaline, ellipsoidal or fusoid-ellipsoidal, rounded at the distal end, somewhat truncate at the proximal end, when developed under favorable conditions measuring mostly 46 to 70 μm (average 62.3 μm) in length and 21–29 μm (average 24.7 μm) in greatest width, containing from 1–5 cross-walls but mostly divided by 3 or cross-walls into 4–5 cells whereof one, as a rule– the penultimate cell usually in triseptate specimens and the median cell usually in quadriseptate specimens- greatly exceeds the others in size. Capturing nematodes by means of three dimensional adhesive networks.
Sexual state:
Orbilia auricolor (Bloxam ex Berk) Sacc., Syll. Fung., 8: 625,1889
Distribution:
USA.
Notes:
The description is based on the protologue. This species resembles A. oudemansii, but differs from the latter by mainly 3–4-septate conidia, while conidia of A. oudemansii are mainly 3-septate. (Fig. 3.39)

Arthrobotrys psychrophila. a conidiophore; b conidium; c adhesive network. Bar = 50 µm
Arthrobotrys pyriformis (Juniper) Schenk, W.B. Kendr. & Pramer, Can. J. Bot. 55 (8): 984 (1977) |
Dactylaria pyriformis Juniper, Trans. Br. Mycol. Soc. 37: 437 (1954) |
Dactylariopsis pyriformis (Juniper) Mekht., Khishchnye Nematofagovye Griby— Gifomitsety (Baku): 119 (1979) |
Characteristics:
Colonies on CMA whitish. Mycelium spreading, vegetative hyphae hyaline, septate, branched, 2–5 μm wide. Conidiophores erect, hyaline, septate, 150–550 μm long, 4–8 μm wide at the base, tapering upwards to a width of 2.5–4 μm at the apex with denticle nodes; conidiophores usually continue development by repeated elongation and successive production of 2–3 additional denticle nodes, often containing 4–5 conidia. Conidia hyaline, elongate-pyriform, broadly rounded at the distal end, 17–38 × 6.5–11.5 μm, mostly 2–3-septate, occasionally 1-septate. Chlamydospores thick-walled, globose or ellipsoidal, 20–35 10–2 μm, yellowish with dense globuliferous content. Capturing nematodes by three dimensional adhesive networks.
Distribution:
China (Yunnan), Canada (Quebec), Ecuador (Baños), Germany (Berlin), Netherlands (Noordoostpolder), UK (England)
Material examined:
YMF1.00018, isolated from forest soil in Shennongngjia, Hubei in 2003 by Wei Zhou; Z125, isolated from forest soil in Xishuangbanna, Yunnan in 1994 by Ke-Qin Zhang. Permanent slide: LH31.
Notes:
The specific epithet of A. pyriformis is based on the pyriform-shaped conidia. It resembles Dr. brochopaga in having 2–3-septate conidia, but differs in trapping devices and conidial shape. A. pyriformis produces elongate-oviform conidia and three dimensional adhesive networks while Dr. brochopaga produces cylindrical or elongate ellipsoidal conidia and constricting rings. Oorshot (1985) restudied the holotype (CBS204.83), and observed conidiophore branching, less expanded at the conidiogenous loci, and smaller conidia than in the original description (15–25 × 5–8 μm). The characters of our strain YMF1.00018 match well with the original description. (Fig. 3.40)

Arthrobotrys pyriformis. a–c conidiophore; d–i conidia; j adhesive network. Bars = 10 µm; Strain number: YMF1.00018
Arthrobotrys reticulata (Peach) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 104 (1999) |
Dactylella reticulata Peach, Trans. Br. Mycol. Soc. 33 (1–2): 148 (1950) |
Golovinia reticulata (Peach) Mekht., Khishchnye Nematofagovye Griby-Gifomisety (Baku): 143 (1979) |
Monacrosporium reticulatum (Peach) R.C. Cooke & C.H. Dickinson, Trans. Br. Mycol. Soc. 48: 622 (1965) |
Characteristics:
Colonies on CMA whitish. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched, 4–5 μm. Conidiophores hyaline, erect, septate, simple, only infrequently branched at the base, 250 μm long, bearing a single conidium on the apex. Conidia hyaline, ellipsoid, with one of the central cells being largest, 50–65 × 20–25 μm, mostly 4-septate, sometimes 5-septate. Chlamydospores present in aged cultures, in chains or clusters, globose to subglobose. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Anhui, Guizhou, Hebei, Shanxi), UK (near Harpenden Common)
Material isolated:
FJ46, isolated from forest soil in Fanjing Mountain, Guizhou in 1994 by Ke-Qin Zhang;
Notes:
Our isolate matches the original description (Peach, 1950). This species can be distinguished from similar species by its fusiform and 5-septate conidia. (Fig. 3.41)

Arthrobotrys reticulata. a conidia. Bar = 30 µm
Arthrobotrys robusta Dudd., Trans. Br. mycol. Soc. 34 (4): 598 (1952) [1951] |
Characteristics:
Colonies on CMA whitish, rapidly growing and extending to a diameter of 7.5 cm at 25 °C within one week. Mycelium spreading, vegetative hyphae hyaline, septate, branched, mostly 2–7 μm wide. Conidiophores erect, septate, branched, 200–300 μm long, 2–2.5 μm wide at the base, tapering upwards to a width of 1–2 μm at the slightly expanded apex, where on forming a compact of conidia (up to 6). Conidia oblong-pyriform, hyaline, 20–27.5 (24.4) × 7.5–12.5 (10.8) μm, 1-septate at the centre of spore. Chlamydospores not observed. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Anhui, Beijing, Guizhou, Hebei, Heilongjiang, Jilin, Liaoning, Neimeng, Shanxi, Yunnan), Canada (Quebec), Ecuador (Baños), Germany (Berlin), Netherlands (Noordoostpolder), UK (England).
Material examined:
Z241, ZX41, isolated from forest soil in Guizhou and Shanxi in 1991 by Ke-Qin Zhang; ZS–3–2, isolated from forest soil in Xishan, Yunnan in June 1999 by Lu Cao; DQ4–8–2, isolated from soil in Xundian, Yunnan in September 2002 by Jing Zhang. Permanent slide: XS–3–2.
Notes:
A. robusta resembles A. superba and A. oligospora in conidia, but differs in the freely branched conidiophores and in the absence of nodal development of conidia in whorls down the conidiophores. A. robusta also resembles A. cladodes in its general habit, differing from the former in both size of conidia and the absence of enlarged storage hyphae. (Fig. 3.42)

Arthrobotrys robusta. a–b conidiophore; c adhesive network; d–g conidia. Bars = 10 µm; Strain number: DQ4-8-2
Arthrobotrys rutgeriense (R.C. Cooke & Pramer) Z.F. Yu, comb.nov |
Monacrosporium rutgeriense R.C. Cooke & Pramer, [as ‘rutgeriensis’] Phytopathology 58: 544, 1968 |
MB 804793 |
Characteristics:
Colonies on CMA white. Vegetative hyphae hyaline, septate, branched. Conidiophores hyaline, erect, simple, septate, 67.5–350 μm long, 3–8 μm wide at the base, gradually tapering upwards to a width of 2–3 μm at the apex, bearing a single conidium. Sometimes conidiophores branched at the apex and two conidia were each borne singly and terminally on these branches, which up to 15 μm long. Conidia hyaline, globose to broadly turbinate, 27–47.5 (32.2) × 17.5–27.5 (22) μm, rounded at the distal end and tapered proximally to a narrow, truncate at the base, 2–3-septate. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Guizhou, Xizang, Yunnan), USA
Material examined:
⑨–13, isolated from field soil in Huaxi, Guizhou in 1996 by Ke-Qin Zhang; XZM–9, isolated from soil in Xizang in August 2000 by Minghe Mo; YMF1.00040, isolated from soil in Baoshan, Yunnan in October 2002 by Jing Zhang. Permanent slide: ⑨–13
Notes:
A. rutgeriense resembles A. eudermata in conidial shape and septation, but differs markedly from it in conidial size, although there is a slight overlap between the two species. A. rutgeriense can be distinguished from A. cookedickinson in having predominantly 2–3-septate conidia in contrast to mainly 2-septate conidia in the latter species. (Fig. 3.43)

Arthrobotrys rutgeriense. a–j conidia; k germinating conidia; l–m conidiophore; n adhesive network. Bars = 10 µm; Strain number: YMF1.00040
Arthrobotrys salina (R.C. Cooke & C.H. Dickinson) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 104 (1999) |
Monacrosporium salinum R.C. Cooke & C.H. Dickinson, Trans. Br. Mycol. Soc. 48: 626 (1965) |
Golovinia salinum (R.C. Cooke & C.H. Dickinson)Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 145 (1979) |
Characteristics:
Single conidium produce from apex of conidiophores; conidia fusiform to ellipsoid, 32.5–52.5 × 12.5–17.5 μm, 0–4-septate, mainly 3-septate; chlamydospores not observed; capturing nematodes by means of three dimensional adhesive networks.
Distribution:
UK.
Notes:
The description is based on the protologue. This species varies in shape, septa and size of conidia, which result in difficulty of species identification. (Fig. 3.44)

Arthrobotrys salina. a conidia. Bar = 30 µm
Arthrobotrys scaphoides (Peach) S. Schenck, W.B. Kendr. & Pramer, Can. J. Bot. 55 (8): 984 (1977) |
Dactylaria scaphoides Peach, Trans. Br. Mycol. Soc.35 (1): 19 (1952) |
Monacrosporium scaphoides (Peach) Xing Z. Liu & K.Q. Zhang, Mycol. Res. 98 (8): 865 (1994) |
Woroninula scaphoides (Peach) Mekht, Khishchnye Nematofagovye Griby-Gifomitsety (Baku): 113 (1979) |
Characteristics:
Mycelium spreading, growing slowly, reaching 30 mm in diameter at 28 °C after 12 days. Vegetative hyphae hyaline, septate, 4–6 μm wide. Conidiophores erect, hyaline and septate, unbranched, 4.5–5.6 μm wide at base and tapering to a width of 4–4.5 μm, at a distance of 80–200 μm from the base producing 1–6 conidia, occasionally up to 10 conidia, in a loose capitate arrangement, then following repeated elongation often giving successively rise to up to 3 additional conidial clusters by branching at or just below slightly swollen warted nodules, producing more or less geniculate conidiophores up to 365–430 μm long. Conidia hyaline, 36.6–79.3 (57) × 11–17.5 (14) μm (Chinese strain), (50–)60–80 (–86) × (13–)14–15 (–16) μm (Dutch strain, both vital state in water), fusiform, not or slightly curved, 1–6-septate, mainly 2–3-septate, the proportion of conidia with 1, 2, 3, 4, 5 and 6 septa is 1.3, 48.8, 37.5, 10, 1.3 and 1.3 % respectively (Chinese strain), 2–3 (–4)-septate (Dutch strain), central cell mostly distinctly longer and wider than other cells. Three dimensional adhesive networks observed when nematodes added. Chlamydospores not observed in cultures.
Distribution:
China, Netherlands
Material examined:
YMF1.01895: Gansu Province, Jiuquan City, alt. 2450 m, from soil, under Malus asiatica Nakai (Rosaceae), from a private plantation. VIII.2006, YMF 1.01895, permanent slide culture (YMF 1.01895); H.B. 6972: The Netherlands: Zeeland, old harbor facing the Hertoginpolder, Verdronken Land van Saeftinghe, most eastern part, alt. 0 m, on previous year’s leaves of Scirpus maritimus L. (Cyperaceae), 23.III.2001,G. Van Ryckegem (H.B. 6972b, dry specimen, associated with A. oudemansii M. Scholler et al. and its sexual state).
Notes:
Within A. scaphoides, the Chinese strain shows a closer relationship to strain H.B. 6972: the ITS rDNA sequence analysis demonstrates that there is 99.6 % similarity between them (2 nucleotide variance in ITS1-5.8s-ITS2 region), but 98.7 % between the Chinese strain and CBS 226.52 (type strain, 7 nucleotide variances), and 99 % between the strain from Scirpus and the type strain from Typha (5 nucleotide variance). Between the Chinese strain of A. scaphoides and other morphologically close species, the similarity is 84.2 % to A. microscaphoides Xing Z. Liu & B.S. Lu, 84.9 % to A. thaumasia (Drechsler) S. Schenck et al., (CBS 322.94), and 85.7 % to A. gampsospora. (Fig. 3.45)

Arthrobotrys scaphoides. a conidiophores; b conidiophores with conidia; c adhesive network; d conidia. Bars: a, d = 10 µm, b, c = 50 µm. Strain number:YMF1.01895
Arthrobotrys shahriari (Mekht.) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 104 (1999) |
Candelabrella shahriari Mekht, Mycol. Res. 101: 334 (1997) |
Characteristics:
Colony growing rapidly, aerial mycelium initially white, becoming pale cream. Conidiophores erect, simple or branched and the apex candelabrum-like with 3–9 branches each bearing a conidium at the apex. The conidiophores rarely recommenced growth after the group of conidia had been produced and a second head was then formed above the first. Conidia elongate-obovoid or elongate-ellipsoidal, straight or sometimes curved, rounded distally and tapering proximally to a truncate base, 1-septate, 33.5–57 × 11–15.5 μm. Globose terminal chlamydospores also formed and adhesive traps formed rarely in pure culture. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
Azerbaijan (Tabriz)
Notes:
The description is based on the protologue. This species can be distinguished from other similar species by the candelabrum-like apex of the conidiophores and 1-septate, elongate-obovoid or elongate-ellipsoidal conidia. (Fig. 3.46)

Arthrobotrys shahriari. a conidiophore; b conidia; c adhesive network. Bar = 40 µm
Arthrobotrys shizishanna (X.F. Liu & K.Q. Zhang) J. Chen, L.L. Xu, B. Liu, Fungal Diversity 26 (1): 124 (2007) |
Dactylella shizishanna X.F. Liu & K.Q. Zhang, Fungal Diversity 14: 104 (2003) |
Characteristics:
Colonies on CMA whitish, slow growing and extending to a diameter of 3.5 cm within 15 days at 25℃. Aerial hyphae scant, hyaline, septate, branching, commonly 2.5–3.7 μm wide. Conidiophores growing from mycelium on the substratum, single, erect, rarely branched, 35–200 μm long, 1.5–2.5 μm wide at the base, tapering upwards gradually to a distal width of 0.5–1 μm, bearing a single conidium. Conidia hyaline, clavate, gradually narrowing at the basal end, obtuse at the distal end, straight or sometimes slightly curved, sometimes constricted at the septa, 2–9-septate, mostly 3–7-septate, 22.5–73.8 (50.6) × 5–10 (6.6) μm. The proportion of conidia with 3, 4, 5, 6 and 7 septa is 10, 30, 33, 18 and 6 %, respectively. Chlamydospores present in aged cultures. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Hubei)
Material examined:
YMF1.00022, isolated from field soil in Shizishan, Hubei in February 2001 by Xuefeng Liu.
Notes:
A. shizishanna is easily distinguished other Arthrobotrys species because having the most septate conidia. (Fig. 3.47)

Arthrobotrys shizishana. a–b conidiophore; c chlamydospore; d–k conidia; l adhesive network. Bars = 10 µm; Strain number: YMF1.00022
Arthrobotrys sinensis (Xing Z. Liu & K.Q. Zhang) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 104 (1999) |
Monacrosporium sinense Xing Z. Liu & K.Q. Zhang, Mycol. Res. 98: 863 (1994) |
Characteristics:
Colonies growing on CMA rapidly, attaining a diameter of 6 cm within 7 days at 25 °C, with sparse aerial hyphae. Vegetative hyphae hyaline, septate, branched, mostly 2.5–9.5 μm wide. Conidiophores hyaline, simple, erect, septate, occasionally branched, mostly 200–500 μm long, 4.5–5.5 μm wide at the base, tapering to 2.5–3 μm wide at the apex, bearing a single conidium. Conidia hyaline, subsphaerical to obovoid, 1–3-septate, 37 % of conidia 3-septate, 42 % 2-septate and 21 % 1-septate, 23.5–30 (27.6) × 17–25 (20) μm. Chlamydospores sphaerical to ellipsoidal, terminal to intercalary in chains, mostly 20–24 × 17–25 μm. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Anhui, Jiangxi, Xizang, Yunnan)
Material examined:
YMF1.00025, isolated from field soil in Huaxi, Guizhou in 1996 by Ke-Qin Zhang; XZM–10, isolated from soil in Xizang in August 2000 by Minghe Mo. Permanent slide: CK9–1
Notes:
This fungus closely resembles A. cookedickinson and A. indica in its relatively small conidia and conidial shape, but differs in size of conidia. In A. cookedickinson, conidia are 32.5–50 × 14–23 μm, in A. indica conidia are 22–30 × 14–20 μm, while in A. sinense, conidia are 25–30.5 × 15–18 μm. (Fig. 3.48)

Arthrobotrys sinensis. a–b conidiophore; c–l conidia; m adhesive network. Bars = 10 µm; Strain number: YMF1.00025
Arthrobotrys sphaeroides (Castaner) Z.F. Yu, comb. nov. |
Monacrosporium sphaeroides Castaner, Am. Midl. Nat. 80: 282 (1968) |
MB804795 |
Characteristics:
Colonies on CMA white, growing rapidly. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched, mostly 2–5 μm wide. Conidiophores hyaline, erect, simple, septate, 200–460 μm long, 5–8 μm wide at the base, gradually tapering upwards to a width of 2.5–3 μm at the apex, bearing a single conidium. Conidia hyaline, sphaeroid to obovoid, 20–44 (32) × 17–25 (20.4) μm, 0–3-septate. Chlamydospores in chains or clusters. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Guizhou, Xizang, Yunnan), USA
Material examined:
YMF1.00030, isolated from field soil in Huaxi, Guizhou in 1996 by Ke-Qin Zhang; YMF1.00539, isolated from soil in Baoshan, Yunnan in September by Jing Zhang. Permanent slide: ④–5–1.
Notes:
This fungus mostly resembles A. cookedickinson. In A. cookedickinson, conidiophores are simple or branched, and conidia are clavate to turbinate and 32.5–50 × 14–23 μm. In A. sphaeroides conidiophores are always simple, conidia are sphaeroid to obovoid and 28–42 × 19–29 μm. Comparisons between A. cookedickinson and A. sphaeroides indicate that the latter is sufficiently different to be considered a distinct species. (Fig. 3.49)

Arthrobotrys sphaeroides. a–g conidia; h conidiophores with conidia; i–j chlamydospore; k adhesive network. Bars = 10 µm; Strain number: YMF1.00030
Arthrobotrys superba Corda, Pracht-Fl. Eur. Schimmelbild.: 43 (1839) |
Didymozoophaga superba (Corda) Soprunov & Galiulina, Mikobiology, Reading 20: 493 (1951) |
Characteristics:
Colonies on CMA whitish to reddish, rapidly growing. Mycelium dense, spreading, vegetative hyphae hyaline branched, septate. Conidiophores hyaline, smooth, 6–9-septate, rarely branched, erect, 180–600 μm long, 2.5–7.5 μm wide, apex swelling, bearing 20–39 conidia. Conidia elliptical, 1-septate at the centre of spore, slightly constricted at the septum, broadly rounded at the distal end, 7.5–27.5 (15.8) × 5–10.5 (6.6) μm. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Anhui, Guizhou, Hainan, Hubei, Jilin, Tianjing, Xizang, Yunnan), cosmopolitan.
Material examined:
Z19, isolated from field soil in Huaxi, Guizhou in 1996 by Ke-Qin Zhang; JLCB–1, isolated from forest soil in Changbai Mountain, Jilin in 1996 by Ke-Qin Zhang; LT–10, isolated from soil in Huaguoshan, Lincang in 1999 by Yanju Bi; J–1qz07, isolated from soil in Liqizhuang, Tianjing in 2000 by Wenpeng Li; XZA–10, isolated from pasture soil in Xizang in August 2000 by Minghe Mo; YMF1.00016, DQ4–2, isolated from forest soil in Xundian, Yunnan in September 2002 by Jing Zhang. Permanent slide: ⑨–19.
Notes:
Arthrobotrys superba was established as the type species of Arthrobotrys (Corda 1839), which is the asexual state of Orbilia fimicola (Pfister 1994). Arthrobotrys superba resembles A. conoides and A. oligospora in most morphological characters except in conidia shape and the position of septum. In A. superba conidia are elliptical and 1-septate at the centre. In A. conoides conidia are elongate obconical, 1-septate about one third from the basal end, constricted at the septum. In A. oligospora conidia are obovoid, 1-septate near the base of the spore, and constricted at the septum. (Fig. 3.50)

Arthrobotrys superba. a conidiophore; b–f conidia; g adhesive network. Bars = 10 µm; Strain number:YMF1.00016
Arthrobotrys tabrizica (Mekht.) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 104 (1999) |
Nematophagus tabrizicus Mekht, Mycol.Res.102: 683 (1998) |
Characteristics:
Colony growing rapidly, velvety, initially white, changing through pale pink to intensive pink after 2 wk. Conidiophores simple, erect, branched, curved, with several swollen to distinctly denticulate conidiogenous regions forming close conidial heads; these proliferating to form additional conidiogenous regions. Conidia elongate-ovate to elongate-doliform or ellipsoidal, 1–2 (–3)-septate, 28.5–56 × 11.5–22.5 μm, with a pronounced protuberant base. Ellipsoidal to cylindrical intercalary chlamydospores present. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
Azeraijan (Tabriz)
Notes:
The description is based on the protologue. This species produces simple or branched conidiophores with several conidial clusters. Conidia are 1–3-septate; 2-septate conidia are more dominant in the original isolation plate than when the fungus was grown on malt agar. (Fig. 3.51)

Arthrobotrys tabrizica. a conidiophore; b conidia; c adhesive network. Bar = 25 µm
Arthrobotrys thaumasia (Drechsler) S. Schenck, W.B. Kendr. & Pramer, Can. J. Bot. 55 (8): 984 (1977) |
Dactylaria thaumasia Drechsler, Mycologia 29 (4): 522 (1937) |
Candelabrella thaumasia (Drechsler)Rifai, Reinwardtia 7 (4): 369 (1968) |
Golovinia thaumasia (Drechsler) Mekht., Mikol. Fitopatol. 1: 276 (1967) |
Monacrosporium thaumasium (Drechsler) de Hoog & van Oorschot [as ‘thaumasia’], Stud. Mycol. 26: 120 (1985) |
Characteristics:
Colonies on CMA white to reddish. Mycelium spreading, vegetative hyphae hyaline, septate, branched, 2–8 μm wide. Conidiophores hyaline, erect, septate, 195–460 μm long, 4–8 μm wide at the base, gradually tapering upwards to a width of 2–4 μm at the apex, simple or often somewhat branched near the apex, and there borne on blunt sterigmata, mostly 2–10 μm long and 2–3 μm wide, usually 3–15, rarely up to 25 conidia in loose capitate arrangement. Conidia hyaline, top-shaped, rounded at the apex, tapering towards the frequently somewhat protruding truncate base, 30–60 (36.2) × 15–30 (20.2) μm, 1–4-septate, mostly 3-septate. Chlamydospores yellow, globose or ellipsoidal. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Anhui, Beijing, Guangdong, Guizhou, Hebei, HuBei, Hunan, Jiangsu, Xizang, Yunnan, Zhejiang), Baukina Faso (Houet), Germany (Berlin–Dahlem), Netherlands (Schipborg), Thailand, UK (England), USA (Norfolk)
Material examined:
L3–5, isolated from soil in Licang, Yunnan in 1999 by Yanju Bi; XZM–11, isolated from soil in Xizang in August 2000 by Minghe Mo; YMF1.00026, YMF1.00547, YMF1.00586, isolated from soil in Baoshan, Yunnan in October 2002 by Jing Zhang; Sjz3.11.20, isolated from soil in Huaxi, Guiyang in 1996 by Ke-Qin Zhang. Permanent slide: Sjz3.11.20
Notes:
Arthrobotrys thaumasia resembles A. elegans in conidial shape and number of septa, but differs in conidiophores and conidial length. The conidia of A. thaumasia are longer than those of A. elegans. Moreover, in A. thaumasia, conidiophores branch near the apex, bearing blunt sterigmata with 3–15 conidia in a loose capitate arrangement, while in A. elegans conidiophores bear a single conidium at the apex, or occasionally form a branch near the apex with a conidium. (Fig. 3.52)

Arthrobotrys thaumasia. a conidiophore; b–e conidia; f chlamydospore; g adhesive network. Bars = 10 µm; Strain number: YMF1.00547
Arthrobotrys vermicola (R.C. Cooke & Satchuth.) Rifai, Reinwardtia 7 (4): 371 (1968) |
Dactylaria vermicola R.C. Cooke & Satchuth., Trans. Br. Mycol. Soc. 49: 27 (1966) |
Nematophagus vermicola (R.C. Cooke & Satchuth.) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 105 (1979) |
Characteristics:
Colonies on CMA whitish. Mycelium spreading, vegetative hyphae hyaline, septate, branched. Conidiophores erect, unbranched, septate, 140–470 μm long, forming 3–6 conidia in a loose capitate arrangement. Each conidium is formed on a small protuberance. The conidiophores often recommence growth after the first group of conidia had been produced, and a second head is then formed about 75 μm above the first. This process is repeated so that three to four groups of conidia are produced on each conidiophore, the latter then becoming somewhat geniculate. Conidia elongate-ellipsoidal to broadly fusiform, hyaline, 25–50 × 17.5–25 μm, bluntly rounded distally and tapering proximally to a narrow truncate base, 1–3-septate; proportion of conidia with 1, 2 and 3 septa is 4, 64 and 32 %, respectively. In 2-septate conidia the median cell, and in 3-septate conidia the penultimate cell, is usually larger than the others. Very rarely a 4-septate conidium was observed. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
Cosmopolitan
Material examined:
Z296, Z290, Z279–1, isolated from soil in Beijing, and Guiyang, Guizhou during 1991–1992 by Ke-Qin Zhang; YNBN–2, isolated from forest soil in Xishuangbanna, Yunnan in 1996 by Ke-Qin Zhang; J31–1, isolated from soil in Jianshui, Yunnan in 1999 by Yanju Bi; BS3–35, isolated from soil in Baoshan, Yunnan in October 2002 by Jing Zhang; YMF1.00533, YMF1.00534, YMF1.00554, from soil in Dehong, Yunnan in July 2003 by Hong Luo. Permanent slide: BS3–35.
Notes:
Arthrobotrys vermicola resembles A. guizhouensis (Zhang 1994) in the conidial shape and trapping devices, but differs in conidia septa and conidiophore form. In A. vermicola, conidia are mostly 2–3-septate, produced along the conidiophores, with 3–6 conidia being located at each denticle node of the unbranched conidiophores in a loose capitate arrangement. In A. guizhouensis, conidia are mostly 1-septate, produced at the irregular expanded apex of the branched conidiophores. (Fig. 3.53)

Arthrobotrys vermicola. a conidiophore; b conidia; c adhesive network. Bars = 10 µm; Strain number: YMF1.00534
Arthrobotrys yunnanensis M.H. Mo & K.Q. Zhang, Fungal Diversity 18: 109 (2005) |
Characteristics:
Colonies growing rapidly on CMA and attaining 6 cm diameter in 5 days at 28ºC and mycelia spreading at the rate of 0.5 cm per 24 h, conidiophores and conidia are produced after 4 days. Mycelium scanty, spreading, vegetative hyphae hyaline, septate and branched, mostly 2–4 µm wide. Conidiophores hyaline, erect, septate, simple, frequently 60–200 µm long, 2–5 µm wide at the base and 1.5–2.4 µm at the apex, producing 1–5 conidia singly from conidiogenous loci on conspicuous denticles at and near the apex. Conidia hyaline, elongate ellipsoid-cylindrical or slightly clavate, broadly rounded at the apex, rounded to truncate at the narrowed base, sometimes constricted gradually at the distal part of conidia, usually aseptate, occasional 1-septate (< 5 %) at the centre, 17.5–32.5 (22.6) × 2.75–7.5 (5.5) µm. Chlamydospores sphaerical to ellipsoidal, intercalary. Capturing nematodes by means of three dimensional adhesive networks.
Distribution:
China (Yunnan)
Material examined:
MF1.00593, isolated from soil in Yuliang, Yunnan in August 2003 by Minghe Mo.
Notes:
Arthrobotrys yunnanensis differs from the three species of Arthrobotrys which form aseptate, or occasionally 1-septate conidia, A. anomala, A. amerospora and A. botryospora. Arthrobotrys yunnanensis is characterized by predominantly elongate ellipsoid-cylindrical or slightly clavate, non-septate conidia which are borne on distinct and long denticles. Arthrobotrys yunnanensis and A. anomala have similar conidial shapes, but differ in the size of conidia. The conidia of A. anomala form one septum only when detached, while in the former species they are aseptate when detached. Arthrobotrys yunnanensis is easy to distinguish from A. botryospora and A. amerospora by their different conidia and conidiophores (modification at apex). Furthermore, unlike A. botryospora, A. yunnanensis does not capture nematodes by adhesive hyphae, and unlike A. amerospora, which only produces aseptate conidia, A. yunnanensis occasionally produces 1-septate conidia. (Fig. 3.54)

Arthrobotrys yunnanensis. a–c conidiophore; d–f conidia; g–h adhesive network; i–j chlamydospore. Bars = 10 µm; Strain number: YMF1.00593
Species of the genus Dactylellina
Dactylellina arcuata (Scheuer & J. Webster) Ying Yang & Xing Z. Liu, Mycotaxon 97: 158 (2006) |
Dactylella arcuata Scheuer & J. Webster, Mycol. Res. 94: 720 (1990) |
Arthrobotrys arcuata (Scheuer & J. Webster) Yan Li, in Li, Hyde, Jeewon, Cai, Vijaykrishna & Zhang, Mycologia 97 (5): 1042 (2005) |
Gamsylella arcuata (Scheuer & J. Webster) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 108 (1999) |
Monacrosporium arcuata (Scheuer & J. Webster) A. Rubner, Stud. Mycol.39: 49 (1996) |
Dactylellina M. Morelet, emend. M. Scholler, Hagedorn & A. Rubner |
Type species: Dactylellina leptospora (Drechsler) M. Morelet-Bull. Soc. Sci. Nat. Archéol. Toulon Var 178: 6 (1968) |
Dactylella leptospora Drechsler, Mycologia 29: 507 (1937) |
Dactylosporium Mekht. (1967) [non Harz 1871] (nom. illeg., Art. 53 ICBN) |
Type species: Dactylosporium leptosporum (Drechsler) Mekht., Mikol. Fitopatol. 1: 278 (1967) |
Dactylella leptospora Drechsler, Mycologia 29: 507 (1937). |
Kafiaddinia Mekht. (1978) |
Type species: Kafiaddinia fusarispora Mekht., Mikol. Fitopatol.12: 8 (1978). |
Laridospora Nawawi, (1976) |
Type species: Laridospora appendiculata (Anastasiou) Nawawi— Trans. Br. Mycol. Soc. 66: 344 (1976) |
Dactylella appendiculata Anastasiou— Pacif. Sci. 18: 202 (1964) |
Characteristics:
Mycelium slow-growing. Hyphae septate, branching, hyaline. Saprotrophic or predatory by trapping nematodes or other animals by means of stalked adhesive knobs, sometimes in combination with stalked three-celled non-constricting rings. Traps generally spontaneously formed. Conidiophores mostly simple; branches, when present, often near the apex, apex mostly simple without modifications or sometimes with short denticles. Conidia either formed singly or in clusters on the apex of the conidiophore, holoblastic, hyaline, 1–7 (15) septate, mostly cylindrical, ellipsoidal, fusiform or spindle-shaped, rarely clavate or obconical. Microconidia and microconidiophores very rarely formed. Chlamydospores absent. Sexual state belonging to the genus Orbilia Fr. (Helotiales).
Key to Species of Dactylellina
1. Trapping organs adhesive knobs or knobs and non-constricting rings 5
1. Trapping organs others 2
2. Trapping organs simple two-dimension networks D. gephyropaga
2. Trapping organs adhesive branches, verruca, adhesive knobs and networks 3
3. Trapping organs adhesive branches D. cionopaga
3. Trapping organs adhesive verruca, knobs and networks 4
4. Trapping organs adhesive verruca D. robustum
4. Trapping organs knobs and networks D. arcuata
5. Trapping organs adhesive knobs and non-constricting rings 6
5. Trapping organs adhesive knobs 10
6. Near apex of conidiophores producing short branches, with conidia in strikingly loose capitate arrangement D. candidum
6. Conidia singly grows at apices of conidiophores, or at apices of subapical lateral branches 7
7. Conidia elongate-fusiform to cylindrical, with 5-15 septa D. leptospora
7. Conidia fusiform, mostly 4 or 5-septate 8
8. Conidia 3–6-septate, mostly 4 septate, largest cell not obvious D. sichuanensis
8. Conidia with largest cell 9
9. Conidia 2–4-septate, mostly 4-septate D. lysipaga
9. Conidia 3–8-septate, mostly 5-septate D. illaqueata
10.Adhesive unstalked, growing out to form branches, conidia with mostly 3-4 septa D. phymatopaga
10. Adhesive stalked or with short stalk 11
11.Conidiophores with apical cluster of conidia or as well as intercalary cluster, conidia arranged into capitate heads 12
11. Conidiophores unbranched, or occasionally with very short subapical lateral branches, conidia singly arranged 18
12. Conidiophores with apical cluster of conidia as well as intercalary cluster
D. entomopaga
12. Conidiophores with apical cluster of conidia 15
13. Conidiophores branched, which apex with denticles or sterigmata 14
13. Conidiophores unbranched 18
14. Conidia obovoid to elongate-cylindrical, 0–4-septate, mostly 3-septate
D. hertziana
14. Conidia ovoid to clavate, 1-septate D. ferox
15.Conidiophore of the candelabrelloid type D. huisuniana
15. Apex of conidiophores with sterigmata, without intercalary cluster of conidia 16
16. Conidia cylindrical D. haptospora
16. Conidia not cylindrical 17
17. Conidia fusiform, mostly 3-septate, adhesive knobs joined together to form loops
D. lobata
17. Conidia elongate-fusiform or clavate, straight or curve, mostly 6–8-septa
D. daliensis
18. Conidia typically fusiform 21
18. Conidia fusiform to other type 19
19. Distally end of conidia bluntly rounded 20
19. Conidia fusiform to cylindrical, mostly 4–7-septate D. formosana
20. Conidia obconical to clavate, distally end bluntly rounded, 3-septate
D. asthenopaga
20. Conidia ellipsoid to fusiform, 2-4-septate D. mammillata
21. Conidia 1–3-septate, often with thorn-like appendage D. appendiculata
21.Conidia with more than 2 septa 22
22. Shape of conidia varies on CMA, with multiform conidia D. mutabilis
22. Conidia mostly fusiform 23
23. Conidia 4–9-septate, mainly 6–7-septate D. multiseptatum
23. Conidia mostly 4–septate or 2–3-septate 24
24. Conidia mostly 2–-septate 25
24. Conidia mostly 4–septate 26
25. Conidia 3–4-septate, mostly 2–3-septate, 29.5-48.7 × 10.7-15.4 μm D. drechsleri
25. Conidia 3–5-septa, mostly with 3 septa, with shorter stalk adhesive knobs
D. quercus
26. Conidia relatively long, conidia 1–6-septate, mostly 4-septate, 56-97 × 8.5-16
D. copepodii
26. Conidia about 50 μm long 27
27. Conidia 3–5-septate, mostly 4-septate, 33-50 (43.7) × 7.4-13.3 (10.7) D. parvicolla
27. Conidia 2–4-septate, mostly 4 septate, 37.5-62 × 8.7-19.3 D. ellipspspora
Accepted Species of Dactylellina
Dactylellina appendiculata (Anastasiou) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 110 (1999) |
Dactylella appendiculata Anastasiou, Pacif. Sci. 18: 202 (1964) |
Monacrosporium tentaculatum A. Rubner &W. Gams, Stud. Mycol.39: 97 (1996) |
Laridospora appendiculata (Anastasiou) Nawawi, Trans. Br. Mycol. Soc. 66 (2): 344 (1976) |
Characteristics:
Conidia fusiform, often with thorn on it, 20–59 μm long, 10–24 μm wide, 1–3-septate. Capturing nematodes by adhesive hyphae.
Distribution:
Russia
Notes:
This species has not been reported from other regions since it was first reported from Russia. The type strain cannot be located, but we still treat it as an accepted species. (Fig. 3.55)

Dactylellina appendiculata. a adhesive hyphae; b conidia. Bar = 30 µm
Characteristics:
Mycelium colourless, growing rapidly on 0.1 % malt extract agar. Conidiophores colourless, single, septate, erect, bearing only one conidium at the apex or a bundle of 2–8 conidia on denticles or short branches. Occasionally an additional ‘whorl’ of conidia formed around the central of conidiophores. Conidia colourless, usually 3-septate, fusiform, widest below the centre (30–)35–54 × 4–6 μm; end cells longer than central cells. Occasionally 2 septa conidia with cells of about equal length were found. Capturing nematodes by adhesive anastomotic branches.
Distribution:
UK.
Notes:
The description is based on the protologue. This species differs from other species of Dactylellina by its special trapping device. (Fig. 3.56)

Dactylellina arcuata. a conidia; b adhesive branches; c conidiophore. Bar = 20 µm
Dactylellina asthenopaga (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 110 (1999) |
Dactylella asthenopaga Drechsler, Mycologia 29: 498 (1937) |
Dactylariopsis asthenopaga (Drechsler) Mekht., Mikol Fitopatol, 1: 279 (1967) |
Monacrosporium asthenopaga (Drechsler) A. Rubner, Stud. Mycol. 39: 50 (1996) |
Characteristic:
Conidiophores hyaline, septate, erect, 100–200 μm, mostly 125–175 μm high, 2.5–4 μm wide at the base, tapering gradually upwards to a width of approximately 1.5 μm, mostly unbranched and terminating in a single conidium, but occasionally giving off one or two branches some distance below the apex, and then bearing two or three conidia. Conidia hyaline, obconical or clavate, truncate at the narrow proximal end, broadly rounded at the distal end, 20–46 μm, mostly 26–36 μm (average 31.5 μm) long, 6.5–9.5 μm (average 8.2 μm)wide, containing 1–5-septa, but most often and most typically containing 3 septa, the basal of the 4 cells then delimited averaging about 8 μm in length, the antepenultimate cell about 7.5 μm, the penultimate cell about 9.8 μm, and the apical cell about 6.2 μm. Capturing nematodes by adhesive knobs.
Distribution:
Australia, Netherlands, UK.
Notes:
The description is based on the protologue. Within Dactylellina species which have adhesive knobs as trapping device, Da. asthenopaga is characterized by rarely unbranched conidiophores, and obconical or clavate conidia. (Fig. 3.57)

Dactylellina asthenopaga. a conidia; b germinating conidia with adhesive knobs; c conidiophore. Bars = 50 µm
Dactylellina candidum (Nees) Yan Li, in Li, Hyde, Jeewon, Cai, Vijaykrishna & Zhang, Mycologia 97 (5): 1042 (2005) |
Dactylium candidum Nees, Syst Pilze Schw 58, 1816: Fries Syst. mycol. 3: 44, 829 |
Arthrobotrys candida (Nees) S. Schenck, W.B. Kendr. & Pramer. Can. J. Bot. 55 (8): 982 (1977) |
Candelabrella candida (Nees) Rifai, Reinwardtia 7 (4): 369 (1968) |
Dactylaria candida (Nees) de Hoog, Stud. Mycol. 26: 102 (1886) |
Dactylaria candida Nees, Syst. Pilze (Würzburg): 58 (1816) [1816–17] |
Monacrosporium candidum (Nees) Xing Z. Liu & K.Q. Zhang, Mycol. Res. 98 (8): 864 (1996) |
Dactylaria haptotyla Drechsler, Mycologia 42: 48, 1950 |
Monacrosporium yunnanense K.Q. Zhang, Xing Z. Liu & L. Cao, Mycol Res 100: 275 (1996) |
Dactylellina yunnanensis (K.Q. Zhang, Xing Z. Liu & L. Cao) M. Scholler, Sydowia 51: 108 (1999) |
Monacrosporium chiuanum Xing Z. Liu & K.Q. Zhang, Mycol. Res. 98: 863, 1994 |
Characteristics:
Colonies hyaline initially and turned whitish or faintly pink after 15 days of incubation on CMA and extending a diameter of 4 cm at 25℃ within 10 days. Mycelium hyaline, scanty, vegetative hyphae septate and branched. Conidiophores hyaline, erect, 5–7-septate, branched, 100–335 μm long, 2–3.7 μm wide at the base, gradually tapering upwards to a distal width of 1–2 μm, initially bearing a single terminal conidium, later often forming 2–12 (mostly 3–5) short branches near the apex with conidia in strikingly loose capitate arrangement. Conidia hyaline, narrow spindle-shaped or fusoid, gradually narrowing at the basal end, truncate at the base, 2–5-septate, mainly 4-septate, 27.5–57.5 (35) × 7.5–12.5 (9) μm. The proportion of conidia with 2, 3, 4 and 5 septa is 2, 10, 81 and 7 %, respectively. Chlamydospores not observed in cultures. The fungus produces two types of trapping organ: non-constricting ring and adhesive knobs.
Distribution:
worldwide.
Material examined:
D9-1, D12-1, isolated from soil in Wuding, Yunnan in 1999 by Yanju Bi; YMF1.00543, isolated from forest foil in Lijiang, Yunnan in October 2002 by Jing Zhang; YMF1.00036, isolated from soil in Zhongdian, Yunnan in 1999 by Lu Cao; YMF1.000579, isolated from forest soil in E’ mei Mountain, Sichuan in 2002 by Jinshi Deng. Permanent slide: DH5-5.
Notes:
Rubner (1996) treated Monacrosporium candidum, M. haptotylum and M. sclerohyphum as conspecific under the name M. haptotylum. M. yunnanense K.Q. Zhang, X.Z. Liu & L. Cao is similar to M. candidum in having spindle-shaped, mostly 4-septate conidia, and branched conidiophores, bearing 3–10 conidia in a loose capitate arrangement. The only difference being that M. candidum and M. yunnanense form both stalked adhesive knobs and non-constricting rings, while M. haptotylum and M. sclerohyphum form only stalked adhesive knobs. These four taxa are phylogenetically closely related receiving high support, based on the rDNA, β-tubulin and combined datasets and this grouping agrees well with the morphological characters of the taxa (Li et al. 2005). These four taxa can therefore be considered as conspecific and should be named Da. candidum based on our results.
No material of M. chiuanum could be obtained to study the relationship with other related species based on molecular analyses. Comparing the morphological characters of M. chiuanum from original description with M. candidum, M. yunnanense, M. haptotylum and M. sclerohyphum, the difference among them is only in the 4–5 septa and occasionally branched conidiophores, which were not the crucial distinctive features to definite a new species. M. chiuanum was also regarded as a synonym of M. haptotylum by Rubner (1996). Based on the discussion above, we treat it as a synonym of Da. candidum.
(Fig. 3.58)

Dactylellina candidum. a conidia; b adhesive knobs and non-constricting ring; c conidiophore. Bar = 20 µm
Dactylellina cionopaga (Drechsler) Ying Yang & Xing Z. Liu, Mycotaxon 97: 158 (2006) |
Dactylella cionopaga Drechsler, Mycologia 42: 30 (1950) |
Characteristics:
Mycelium sparse, hyphae, septate, 2–5 µm wide; Conidiophores hyaline, erect, 2–11 septate, 170–330 µm high, 5–7 µm wide at the base, gradually tapering upwards, 3 µm at the apex, sometimes branching and bearing a conidium at the apex of each branch. Conidia were broad fusoid, rounded at the apex, truncate at the base, 2–6-septate, mainly 3–4-septate, 35–60 × 13–21 µm. Chlamydospores not observed. Capturing nematode by adhesive branches.
Distribution:
China (Yunnan, Sichuan), USA.
Material examined:
TJ-1qz08, isolated from soil in Liqizhuang, Tianjing in 2000 by Wenpeng Li; YMF1.00569, isolated from soil in Deqin, Yunnan in September 2002 by Jing Zhang;YMF1.00580, isolated from forest soil in Jiuzhaigou, Sichuan in 2002 by Jingshi Deng;Hn2-1-3, isolated from field soil in Hainan in 2002 by Ke-Qin Zhang.
Notes:
Among the species of nematode-trapping fungi, only Da. cionopaga and Da. gephyrophaga form both unstalked adhesive knobs and simple two-dimension networks. Drechsler distinguished Da. cionopaga from Da. gephyrophaga by unstalked adhesive knobs, which seldom link to a scalariform network, while they commonly fuse in the latter. Conidiophores were found to be generally shorter, conidia noticeably longer in Da. cionopaga than in Da. gephyrophaga. Conidia of Da. cionopaga show a conspicuous vacuole and vary in their septation more than in Da. gephyrophaga. (Fig. 3.59)

Dactylellina cionopaga. a conidia; b adhesive branches; c conidiophore. Bar = 30 µm
Dactylellina copepodii (G.L. Barron) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 110 (1999) |
Dactylella copepodii G. L. Barron, Can. J. Bot. 68: 692 (1990) |
Monacrosporium copepodii (G. L. Barron) A. Rubner, Stud. Mycol. 39: 54 (1996) |
Characteristics:
Conidiophores erect, slender, hyaline, septate, unbranched, and 160–360 μm tall, 2–2.5 μm wide for most of their length and somewhat wider 3.5 (4) μm at the base; Each conidiophore bears a solitary, terminal conidium at its apex. Conidia fusiform, broadest in the central region, and tapering to a narrow connective tissue at the base and an acutely rounded apex at the distal end, 1–6-septate, mainly 4 septate, 56–97 × 8.5–16 μm. Capturing nematode by adhesive terminal cells growing on branches.
Distribution:
New Zealand
Notes:
The description is based on the protologue. Among species of Dactylellina, only D. copepodii can trap copepods and is characterized by long branches with adhesive knobs at the apex. (Fig. 3.60)

Dactylellina copepodii. a conidia; b adhesive knobs; c conidiophore. Bar = 25 µm
Dactylellina daliensis H.Y. Su, Mycotaxon 105: 314 (2008) |
Characteristics:
Colonies white on CMA, growing rapidly, obtaining a diameter of 6 cm at 25 °C within 7 days. Mycelium sparse, hyphae hyaline, septate, branched, 2–4-septate, 21–101 µm high, 1.8–2.5 µm wide at the base, gradually tapering upwards to a width of 1.5–1.7 µm at the apex where forming 1–7 apical conidia. Conidia hyaline, elongate-fusoid or clavate, straight or somewhat curved, round at the distal end, truncate at the base, 1–13 (mainly 6–8) septate, 28.7–51.5 (40.8) × 3.2–5.2 (4.2) µm. Chlamydospores not observed in culture. Trapping nematodes by stalked adhesive knobs and non-constricting rings.
Distribution:
China (Yunnan)
Notes:
The description is based on the protologue. This species resembles only two other species in the Dactylellina genus: Da. leptospora, Da. haptospora. Both Da. daliensis and Da. leptospora produce elongate-fusoid conidia and capture nematodes by the same kinds of trapping devices. However, unlike Da. daliensis, which forms terminal cluster of conidia, Da. leptospora mainly bears a single terminal conidium, occasionally producing an additional conidium on a short branch attached to the main axis near the conidiophores apex. Furthermore, the conidial of Da. daliensis are shorter than those of Da. leptospora. Both Da. daliensis and Da. haptospora can bear terminal conidial clusters and form stalked adhesive knobs as trapping devices. However, Da. daliensis differs from Da. haptospora by producing an additional type of trapping device, in non-constricting rings. (Fig. 3.61)

Dactylellina daliensis. a–i conidiophores; j–p conidia; q adhesive knobs; r non-constricting rings. Bars: a = 50 µm; b = 25 µm; c, e = 12.5 µm; d, f–r = 5 µm
Dactylellina drechsleri (Tarjan) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 110 (1999) |
Dactylella drechsleri Tarjan, Mycopathologia 14: 143 (1961) |
Monacrosporium drechsleri (Tarjan) R.C. Cooke & C.H. Dickinson, Trans. Br. Mycol. Soc. 48 (4): 623 (1965) |
Golovinia drechsleri (Tarjan) Mekht., Mikol. Fitopatol. 1: 276 (1967) |
Characteristics:
Mycelium usually profuse, hyaline, slowly spreading; vegetative hyphae septate, 4 μm (3.2–5.3 μm) wide. Conidiophores hyaline, erect, sparingly septate, 145 μm (93–184 μm) long, each terminating in single conidium but often forming one or two branches, well below apex, also bearing solitary conidia. Conidia hyaline, somewhat fusiform, usually tapering conical with finely truncate base, but broadly obtuse to bluntly rounded at distal end, 29.5–48.7 (39.6) × 10.7–15.4 (13.1), 2–4-septate, mainly 3-septate. Capturing nematodes by stalked adhesive knobs, knobs 8–10 (9) × 7.2–9.6 (8.3) μm.
Distribution:
USA.
Notes:
The description is based on the protologue. Among Dactylellina species with adhesive knobs as trapping devices, Da. drechsleri can be recognized by its 3 septate conidia, while others are usually 4 septate. (Fig. 3.62)

Dactylellina drechsleri. a–b, e–h conidia; c–d conidiophore; i adhesive knobs. Bars = 10 µm; Strain number:YMF1.00573
Dactylellina ellipsospora (Preuss) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 110 (1999) |
Dactylella ellipsospora (Preuss) Grove, J. Bot. Lond. 24: 200 (1886) |
Menispora ellipsospora Preuss in Sturm, Deutschl. Fl. Abt. 3:H.8:T.47 (1848) |
Monacrosporium ellipsosporum (Preuss) R.C. Cooke & C.H. Dickinson, Trans. Brit. Mycol. Soc., 48: 623 (1965) |
Characteristics:
Colonies on CMA white to reddish, slow growing. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched, mostly 2–3 μm wide. Conidiophores hyaline, erect, septate, unbranched, 100–330 μm long, 3–4.5 μm at the base, gradually tapering upwards to a width of 2–2.4 μm at the apex, bearing a single conidium. Conidia broadly spindle-shaped, hyaline, 40–57.5 (48.3) × 10–17.5 (13) μm, 2–4-septate, mostly 4-septate, the central cell being the largest, rounded on the apex. Chlamydospores not observed. Capturing nematodes by means of adhesive knobs.
Distribution:
Cosmopolitan.
Material examined:
Z29, isolated from forest soil in Tianzhu, Guizhou in 1988 by Zhang Ke-Qin ZJHZ-1, isolated from field soil in Hangzhou, Zhejiang in 1999 by Ke-Qin Zhang; XZM-3, isolated from forest soil in Xizang in August 2000 by Minghe Mo; YMF1.00032, isolated from soil in Ruili, Yunnan in October 2002 by Jing Zhang; 5102-1, isolated from field soil in Wuhan, Hubei in February 2001 by Xuefeng Liu.
Notes:
Da. ellipsospora resembles Da. lysipaga in conidial characters, but differs from the latter in trapping devices. Da. ellipsospora produces adhesive knobs, while Da. lysipaga produces both adhesive knobs and non-constricting rings. The material examined almost matches the original description. (Fig. 3.63)

Dactylellina ellipsospora. a conidia; b–c conidiophore; d adhesive knobs. Bars = 10 µm; Strain number: YMF1.00032
Dactylellina entomopaga (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 110 (1999) |
Arthrobotrys entomopaga Drechsler, Mycologia 36: 392 (1944) |
Arthrobotrys pauca J. S. McCulloch, Trans. Br. Mycol. Soc. 68: 176 (1977) |
Dactylariopsis entomopaga (Drechsler) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 122 (1979) |
Characteristics:
Mycelium spreading, colourless, septate, 2–3 μm wide. Conidiophores erect, colourless, meagerly septate, 75–175 μm tall, 3–4.5 μm wide at the base, about 2.5 μm wide farther upwards, often somewhat inflated at the top from which given off 3–8 simple or branched sterigmata of 2–7 μm long, whereon borne collectively 3–10 conidia in loose capitate arrangement; additional conidial clusters often being produced following renewed axial elongation. Conidia colourless, cylindrical or somewhat clavate, 15–28 × 4.5–5.5 μm, broadly rounded at the apex, often minutely pedicellate below, 1-septate, the 2 cells not pronouncedly unequal as a rule, even though the lower cell is often slightly longer than the upper one. Capturing nematodes by stalks ellipsoidal adhesive knobs measuring 8–13 × 4.5–8 μm.
Distribution:
Australia, USA.
Notes:
The description is based on the protologue. Among Dactylellina species with adhesive knobs as trapping devices, this species is characterized by loose capitate arranged conidia and repeated extended conidiophores. (Fig. 3.64)

Dactylellina entomopaga. a. conidia; b. adhesive knobs; c. conidiophores. Bar = 30 µm
Characteristics:
Colonies on Czapek yeast white to pale pink-orange, mycelium hyaline; repent and aerial hyphae, 4.5–6.5 μm wide, septate at intervals of 22–35 (–45) μm. Conidiophores macronematous, mononematous, erect, septate, often branched, (33.5)44–144 (–466) μm long, 5–7 μm wide at the base and 3–4 μm farther upwards, producing (2–)4 (–10)conidia on 2–4.5 μm long denticles. Conidia hyaline, obovoidal to clavate, 1-septate, slightly constricted at the septum which is usually in the centre, (13–)15–18 (24.5) × (5–)6–8 (–9) μm. Predatory on springtails (Gressittacantha terranova) by stalks adhesive knobs measuring (18–)20–25 (–31) × (16.5–)17–21 (–21.5) μm.
Distribution:
Italy
Notes:
The description is based on the protologue. The trapping device of this species is hard to induce in pure culture. Its adhesive knobs are 3–4 times larger than other species. Because living cultures are not available, the species may be doubtful. (Fig. 3.65)

Dactylellina ferox. a. conidia; b. adhesive knobs; c. conidiophores. Bars = 20 µm
Characteristics:
Colonies thin, white to orange white, 3.5–4.2 cm diameter in 1 wk. Mycelium hyaline, septate, branched, 2–4 μm wide. Conidiophores erect, unbranched, short, septate, hyaline, 17–90 μm long, 2.5–4.5 μm wide at the base, tapering to subterminal denticle. Conidia hyaline, fusoid, cylindrical, straight or slightly curved, (2)4–7 (8)-septate, 30–70 × 5–7.5 μm, tapering towards both ends. Chlamydospores absent. Capturing nematodes on adhesive knobs.
Distribution:
China (Taiwan)
Notes:
The description is based on the protologue. Morphologically Da. formosana resembles Da. copepodii and Da. leptospora. They share common features such as the ability to capture micro fauna, producing fusoid, cylindrical, multiseptate, solitary apical conidia on unbranched conidiophores. However, in contrast to Da. formosana, Da. leptospora has thinner, longer, more frequently septate conidia, which produce secondary conidia when germinating, and trap nematodes by adhesive knobs and also by non-constricting rings (Drechsler 1937). Although both Da. copepodii and Da. formosana prey micro fauna by adhesive knobs, the former traps copepods as compared to nematodes. Da. copepodii also has conidia with larger intercalary cells and secondary adhesive knobs which arise from germinated conidia. These characteristics readily separate it from Da. formosana (Barron 1990). (Fig. 3.66)

Dactylellina formosana. a. adhesive knobs on hypha trapping nematodes; b. conidia; c. conidiophore. Bars = 30 µm
Dactylellina gephyropaga (Drechsler) Ying Yang & Xing Z. Liu, Mycotaxon 97: 158 (2006) |
Dactylella gephyropaga Drechsler, Mycologia 29: 512 (1937) |
Arthrobotrys gephyropaga (Drechsler) Yan Li, in Li, Hyde, Jeewon, Cai, Vijaykrishna & Zhang, Mycologia 97 (5): 1042 (2005) |
Gamsylella gephyropaga (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 110 (1999) |
Golovinia gephyropaga (Drechsler) Mekht., Mikol Fitopatol.1: 276 (1967) |
Monacrosporium gephyropaga (Drechsler) Subram., [as ‘gephyropagum’] J. Indian bot. Soc.42: 293 (1964) [1963] |
Characteristics:
Colonies on CMA whitish, slow growing and extending to a diameter of 4 cm at 25 °C within ten days. Mycelium scanty, spreading, vegetative hyphae hyaline, septate, branched, mostly 2–5 μm wide. Conidiophores hyaline, erect, commonly unbranched, 2–11-septate, 75–360 μm long, 5–7 μm wide at the base, gradually tapering upwards to a width of 3 μm wide at the apex, bearing a single conidium, sometimes after repeated elongation forming one or two additional conidia. Conidia hyaline, mostly broadly spindle-shaped or somewhat clavate, broadly rounded at the distal end, but at the narrower proximal end tapering towards the bluntly truncate base, commonly 30–67.5 (45) × 10–23.5 (16) μm, 2–6-septate, mostly 3–4-septate. Chlamydospores not observed. Two types of trapping devices observed in culture: adhesive branches and simple two dimension networks.
Distribution:
China (Anhui, Beijing, Guizhou, Hainan, Hebei, Taiwan, Tianjing, Yunnan), Portugal (Quinta de Sao Pedro), Netherlands (Valthernond), Spain (Tenerife)
Material examined:
TJ-1qz08, isolated from soil in Liqizhuang, Tianjing in 2000 by Wenpeng Li; YMF1.00569, isolated from soil in Deqin, Yunnan in September 2002 by Jing Zhang; YMF1.00580, isolated from forest soil in Jiuzhaigou, Sichuan in 2002 by Jingshi Deng; Hn2-1-3, isolated from field soil in Hainan in 2002 by Ke-Qin Zhang. Permanent slide: Hn2-1-3.
Notes:
Among the species of Dactylellina, only Da. cionopaga and Da. gephyrophaga form adhesive branches and simple two-dimension networks, but Da. cionopaga seldom form simple two dimension networks. For other differences between both species refer to Da. cionopaga. (Fig. 3.67)

Dactylellina gephyropaga. a. conidia; b. adhesive branches; c. adhesive network; d. conidiophore. Bars = 40 µm
Characteristics:
Mycelium spreading; the vegetative hyphae hyaline, septate, 1.3–4.5 μm wide. Conidiophores hyaline, erect, septate, 50–150 μm high, 1.7–2.7 μm wide at the base, very gradually tapering upwards to a width of about 1.5 μm, then terminating often in a slightly branched more or less geniculate part formed by repeated elongation following successive apical production of conidia up to 15 in number in loosely capitate arrangement. Conidia hyaline, elongate-cylindrical, 35–60 × 2.2–3.7 μm, more rarely containing 5–8 cross-walls. Capturing nematodes by unicellular adhesive knobs and non-constricting rings. Adhesive subsphaerical or prolate ellipsoidal, 6–10 × 5–8.5 μm.
Distribution:
China (Guizhou, Yunnan), USA.
Notes:
The description is based on the protologue. This species differed from other Dactylellina species by elongate cylindrical and very narrow conidia. (Fig. 3.68)

Dactylellina haptospora. a–d conidiophore; e–h conidia; i adhesive knobs; j chlamydospores. Bars = 10 µm; Strain number: YMF1.00560
Dactylellina hertziana (M. Scholler & A. Rubner) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 111 (1999) |
Arthrobotrys hertziana, M. Scholler & A. Rubner, Mycol.Res.103: 764 (1999) |
Characteristics:
Colonies and reverse whitish to pink, slow-growing, attaining less than 8 cm diameter in 17 days at 25 °C, producing aerial mycelium in concentric rings only on nutrient-rich media. Odor lacking. Morphological characteristics on CMA/2; Hyphae 3–5 μm wide, hyaline, septate, branched, often anastomosing. Conidiophores (150)–170–400 (–440) μm high, 3–5 μm wide in the lower part, erect, septate, mostly branched near the apex or at the bottom, branches slightly bent upwards near the branch base, then clearly erect. Fertile tips with weakly swollen nodes forming up to seven conidia on denticles of 1.5–2.5 (–4) μm long. Conidia hyaline, obovoid to ellipsoid-cylindrical, equi- or mostly inequidistantly (0–)3 (–4) septa, (16–)16.5–22–27.5 (–28.5) × (7.5–)7.5–9.5–11.5 (–12) μm, l/w ratio = 2.5. Nematodes are trapped by stalked or unstalked adhesive knobs. Chlamydospores not seen.
Distribution:
Spain
Notes:
The description is based on the protologue. This species is characterized by branched conidiophores and obovoid to ellipsoid-cylindrical conidia of loose capitate arrangement. (Fig. 3.69)

Dactylellina hertziana. a. conidia; b. adhesive knobs; c. conidiophores. Bar = 15 µm
Dactylellina huisuniana (J.L. Chen, T.L. Huang & Tzean) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 111 (1999) |
Dactylella huisuniana J.L. Chen, T.L. Huang & Tzen, Mycol. Res. 102: 1269 (1998) |
Characteristics:
Colonies on oat meal agar (OMA) 35–40 mm in 17 d at 25 °C, effuse, velvety, very pale orange; reverse very pale orange white to pale orange. Mycelium mostly immersed, branched, septate, smooth, subhyaline to pale orange in mass, 1–4.5 μm wide;. Conidiogenous cells terminal or subterminal, lateral, with denticles up to 9 μm long, 1.5 μm wide, with sympodial proliferations, scattered and often branched, elongate, candelabrelloid at the apex in age, bearing 1–6 conidia. Conidia predominantly fusiform, 3-septate, occasionally clavate-fusoid and 2-septate or 4-septate, smooth, hyaline, 29.5–41 × 4.5–7.5 μm, the distal second cell the widest, base attenuated, truncated, 1–1.5 μm wide. Trapping nematodes by ovoid to subglobose adhesive knobs of 5–7 μm diameter.
Distribution:
China (Taiwan)
Notes:
The description is based on the protologue. This species is characterized by sympodial proliferations conidiophores with candelabrelloid apex. (Fig. 3.70)

Dactylellina huisuniana. a–g conidia; h–i conidiophore; j adhesive knobs. Bars = 10 µm; Strain number: YMF1.00574
Dactylellina illaqueata D.S. Yang & M.H. Mo, in Yang, Chen, Huang, Mo & Zhang, Mycotaxon 94: 215 (2005) |
Characteristics:
Mycelium scanty, spreading, vegetative mycelium colourless, septate, mostly 1.5–2.5 µm wide. Conidiophores colourless, erect, unbranched, often 95–250 µm high, 2.2–2.6 µm wide at base, and gradually tapering upwards to a width of 1.8–2.1 µm at the apex, bearing a single conidium on the apex, occasionally two conidia. Conidia colourless, elongate fusiform, narrowly obtuse at the distal end, truncate at the base, the central cell swelling obviously, 25.5–117.5 (66.5) × 5.5–15.2 (14.1) μm, 3–8-septate, mainly 5-septate. The proportion of conidia with 3,4,5,6,7, and 8 septa accounts for 9.1, 15.2, 63.6, 6.1, 3 and 3 %, respectively. When induced with nematodes, the fungus produced non-constricting rings and stalked adhesive knobs. Chlamydospores sphaerical to ellipsoidal, intercalary.
Distribution:
China (Yunnan)
Notes:
Da. illaqueata is mainly characterized by its 5-septate conidia singly borne on the unbranched conidiophores and this species resembles Da. yunnanensis and Da. lysipaga in conidial shape. However, Da. yunnanensis usually forms short denticles at the apex of conidiophores and bears 2–5 conidia, and Da. lysipaga produces the conidia mainly with 2–4-septate. In comparison with Da. leptospora, the conidia of Da. illaqueata usually have a wider central cell (average 14.1 μm) than that of Da. leptospora (4–5.8 μm). In addition, conidia of Da. leptospora have more septa (5–15) than those of Da. illaqueata (3–8, mainly 5 septa). (Fig. 3.71)

Dactylellina illaqueata. a–c conidiophore; d–l conidia; m–n adhesive knobs and non-constricting rings; o. chlamydospore. Bars = 10 µm
Dactylellina leptospora (Drechsler) M. Morelet, Bull. Soc. Sci. nat. Arch. Toulon & du Var 178: 6 (1968) |
Dactylella leptospora Drechsler, Mycologia 29: 507 (1937) |
Dactylosporium leptospora (Drechsler) Mekht., Mikol. Fitopatol. 1: 277 (1967) |
Monacrosporium leptosporum (Drechsler) A. Rubner, Stud. Mycol. 39: 79 (1996) |
Characteristics:
Colonies on CMA whitish, rapidly growing. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, mostly 1.2–3 μm, rarely up to 4 μm wide. Conidiophores hyaline, erect, septate, 75–250 μm long, 2.5–5 μm wide at the base, tapering upwards very slightly, 1–2 μm wide at the apex, mostly simple and bearing a single terminal conidium, but occasionally bearing an additional conidium on a short branch attached to the main axis near its apex. Conidia hyaline, mostly straight, elongate fusoid or cylindrical, 30–92 (64) × 4–6 (4.5) μm, 4–15-septa, after falling off, often giving rise individually to 1 or 2 globose knobs mostly on scarcely modified distal prolongations, more rarely on lateral stalks; often too, producing on slightly branched conidiophores, 50–125 μm long, 2–3 μm wide at the base and 1.3–1.8 μm wide at the apex, secondary conidia mostly 10–35 × 4–5.8 μm, 3–8-septate. Two types of trapping devices were observed in culture: stalked adhesive knobs and non-constricting rings.
Distribution:
China (Beijing, Guizhou, Hubei, Xizang, Yunnan), India, Russian, USA
Material examined:
SJt3.9.68, isolated from field soil in Huaxi, Guizhou in 1996 by Ke-Qin Zhang; XZD-2, XZD-3, isolated from soil in Xizang in August 2000 by Minghe Mo; DL1-1-1, isolated from forest soil in Dali, Yunnan in September 2002 by Jing Zhang; YMF1.00114, isolated from soil in Kunming, Yunnan in 2002 by Lu Cao; J34-1, J34-2, isolated from forest soil in Jianshui, Yunnan in 1999 by Yanju Bi; YMF1.00115, YMF1.00117, YMF1.00042, YMF1.00562, isolated form soil in Dali and Lijiang, Yunnan in September 2002 by Jing Zhang. Permanent slide: STt3.9.68.
Notes:
The fungus is easy to distinguish from other species because of its elongate-fusiform to cylindrical and greater number of conidial septa. Dactylaria dasguptaii and Kafiaddinia fusariispora were treated as synonym of Dactylaria leptospora because of similar trapping devices and conidial characteristics (Rubner, 1996). All the characteristics of examined material matched well with original distribution of Da. leptospora. (Fig. 3.72)

Dactylellina leptospora. a–c, f conidia; d–e conidiophore; g adhesive knobs; h non-constricting rings. Bars = 10 µm; Strain number: YMF1.00042
Dactylellina lobata (Dudd.) Yan Li, in Li, Hyde, Jeewon, Cai, Vijaykrishna & Zhang, Mycologia 97 (5): 1042 (2005) |
Dactylella lobata Dudd., Trans. Br. mycol. Soc. 34 (4): 489 (1951) |
Gamsylella lobata (Dudd.), M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 108 (1999) |
Monacrosporium lobata (Dudd.) A. Rubner, Stud. Mycol.39: 80 (1996) |
Characteristics:
Mycelium septate, occasionally branched, 2–5 µm wide. Conidiophores erect, single, 250 μm high. Conidia fusiform, mainly with 3 septa, 32–54 × 8–12 μm, each at the apex of an erect conidiophore. Trapping nematodes by adhesive unstalked knobs, which sometimes joined together to form loops.
Distribution:
UK.
Notes:
The description is based on the protologue. In this species, knobs often form a string, which resemble adhesive branches of Da. cionopaga, but the adhesive cell of the latter is rectangle. In addition, conidia of the two species are different. (Fig. 3.73)

Dactylellina lobata. a. conidia; b. adhesive knobs; c. hypha infect nematodes and produce adhesive knobs. Bar = 30 µm
Dactylellina lysipaga (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 111 (1999) |
Dactylella lysipaga Drechsler, Mycologia 29: 503 (1937) |
Golovinia lysipaga (Drechsler) Mekht., Mikol. Fitopatol.1: 277 (1967) |
Monacrosporium lysipagum (Drechsler) Subram., J. Indian Bot. Soc. 42: 293 (1963) |
Characteristics:
Colonies on CMA whitish. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched, mostly 1.5–3.5 μm wide. Conidiophores hyaline, septate, erect, simple, 125–760 μm long, 3–5 μm wide at the base, gradually tapering upwards to a width of 1–2 μm, bearing a single conidium, but occasionally producing a second conidium on a short branch attached to the main axis some distance below the apex. Conidia hyaline, sometimes obovoid-fusoid, but much more frequently and more typically rather symmetrically fusoid, somewhat acutely rounded at the apex, truncate at the narrow base, 2–4-septate, mostly with 3–4 septa, 27.5–85 (40.7) × 9–17.5 (11) μm. Capturing nematodes by means of adhesive knobs and non-constricting rings.
Distribution:
China (Beijing, Guizhou, Hebei, Yunnan), Portugal (Azores)
Material examined:
Z23, isolated from field soil in Tianzhu, Guizhou in 1988 by Ke-Qin Zhang; YMF1.00535, isolated from soil in Baoshan, Yunnan in October 2002 by Jing Zhang; P7.1.1, isolated from forest soil in Wenshan, Yunnan in July 2002 by Zhiwei Zhao. Permanent slide: P7.1.1
Notes:
Da. lysipaga resembles Da. ellipsospora in conidial shape, septate, but the conidia of the former are smaller than those of the latter. Moreover, Da. lysipaga forms both adhesive knobs and non-constricting rings, while Da. ellipsospora forms only adhesive knobs. Da. lysipaga also resembles Da. candidum in conidial shape and trapping devices, but differs in conidiophores. In Da. lysipaga, conidiophores simple, bear a single conidium at the apex, occasionally forming additional one next the apex. While in Da. candidum, conidiophores form short branches near the apex with conidia in strikingly loose capitate arrangement. The conidia of present isolate are longer than in the description of Subramanian [28–55 (40.7) × 9–14 (11.6) μm] (1963). (Fig. 3.74)

Dactylellina lysipaga. a–b conidiophore; c–e conidia; f adhesive knobs and non-constricting rings. Bars = 10 µm; Strain number: YMF1.00535
Dactylellina mammillata (S.M. Dixon) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 111 (1999) |
Dactylella mammillata S.X. Dixon, Trans. Br. Mycol. Soc. 35: 144 (1952) |
Golovinia mammillata (S.X. Dixon) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 159 (1979) |
Monacrosporium mammillata (S.X. Dixon) R.C. Cooke & C.H. Dickinson[as ‘mammillata’] |
Trans. Br. Mycol. Soc. 48 (4): 622 (1965) |
Characteristics:
Mycelium scanty; hyphae hyaline, septate. Conidiophores 160–280 μm high, conidia borne singly on the apex of conidiophores. Conidia ellipsoidal to fusiform, obtuse rounded at the apex, 3–4-septate, 30–60 × 9–17 μm. Capturing nematodes by stalks adhesive knobs 6.5–9 × 6.5–7 μm.
Distribution:
Canada, Germany, UK.
Notes:
This species is most similar to Da. ellipsospora, but differs in having a rounded conidia apex and 3–4-septate conidia, while Da. ellipsospora most often has 4-septate, occasionally 3 or 2 septate conidia. (Fig. 3.75)

Dactylellina mammillata. a. adhesive knobs; b. conidia. Bar = 30 µm
Dactylellina mutabilis (R.C. Cooke) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 111 (1999) |
Monacrosporium mutabile R. C. Cooke [as ‘mutabilis’], Trans. Br. Mycol. Soc. 53: 318 (1969) |
Golovinia mutabilis (Cooke) Mekht., Khishnchnye Nematofagovye Griby–Gifomitesty (Baku): 161 (1979) |
Characteristics:
Conidiophores simple, septate, erect, bearing a single conidium at the apex. Conidia ellipsoidal to fusiform, 50–60 × 16–17.5 μm, with 4 septa, with the median cell markedly larger than the rest. Conidia taper to a narrowly rounded cell distally and proximally to a narrowly truncate base. In pure culture on cornmeal agar conidia are markedly different in size and shape from those on the nematode-infested substrate, being 37.5–50 × 12.5–15 μm. In addition, about 14 % of conidia were 3 septate. Curved, malformed spores were frequent. Capturing nematodes by stalks adhesive knob about 5 μm in diameter.
Distribution:
Samoan Islands
Notes:
The description is based on the protologue. Among species of Dactylellina with 4 septate conidia, Da. mutabilis can be distinguished from other species by densely crowded, stalked, adhesive knobs. (Fig. 3.76)

Dactylellina mutabilis. a. adhesive knobs; b. conidia. Bar = 20 µm
Dactylellina multiseptata (H.Y Su & K.Q. Zhang) Z.F. Yu, comb. nov. |
Monacrosporium multiseptatum H.Y. Su & K.Q. Zhang. Mycotaxon 92: 193 (2005) |
MB 804794 |
Characteristics:
Colonies on CMA plates growing quickly, attaining 6 cm diameter within 7 days at 25 °C. Mycelium spreading, vegetative hyphae hyaline, septate and branched, mostly 1.8–2.5 µm wide. Conidiophores erect, simple, septate, 170–260 µm long, 2.5–3 µm wide at the base, gradually tapering upwards to a width of 1.7–2 µm at the apex, bearing a single conidium, occasionally two conidia. Conidia hyaline, elongate fusiform to straight clavate, sometimes apparently curved, narrowly round at the distal end, tapering towards the narrow truncate at the base, somewhat constricted at septa, 67.5–132.5 (91.6) × 3.8-17.5 (15.5) µm, 4–9-septate, mainly 6–7- septate. The proportion of conidia with 4, 5, 6, 7, 8 and 9 septa is 1.8, 3.6, 23, 45, 25.1 and 2 % respectively. Chlamydospores sphaerical to ellipsoidal, intercalary, about 5–8 × 4–6 µm. In the presence of nematodes the fungus forms stalked adhesive knobs which are unicellular, subsphaerical. The knobs usually 11 µm long, 10 µm wide, with a stalk of 20–67.5 µm.
Distribution:
China (Yunnan)
Material examined:
YMF1.00127, isolated from soil in Yunnan in 2003 by HongYan Su.
Notes:
Da. multiseptata resembles but distinctly differs from two nematode-trapping fungi, Da. candidum and Da. ellipsospora in conidial shape and type of predacious device. Da. multiseptata produces larger conidia [67.5–132.5 (91.6) × 3.8–17.5 (15.5) µm] and more septa (usually 6–8) than Da. candidum [33–50 (43.7) × 7.4–13.3 (10.7) µm, mainly 4-sepate] and Da. ellipsospora [37.5–62 (48.3) × 8.7–19.3 (13) µm, often with 4 septa]. Additionally, Da. multiseptata does not form denticles on the apex of conidiophores, while Da. candidum and Da. ellipsospora often produce 2–5 short branches near the apex. (Fig. 3.77)

Dactylellina multiseptata. a conidiophores; b–k conidia; l–m adhesive knobs; n chlamydospores. Bars = 10 µm
Dactylellina parvicolla (Drechsler) Yan Li, in Li, Hyde, Jeewon, Cai, Vijaykrishna & Zhang, Mycologia 97 (5): 1042 (2005) |
Dactylella parvicollis Drechsle, Sydowia 15: 13 (1962) |
Gamsylella parvicollis (Drechsler) M. Scholler, Sydowia 51: 109 (1999) |
Golovinia parvicollis (Drechsler) Mekht., Khishchnye nematofagovye Griby-Gifomitsety: (Baku)162 (1979) |
Monacrosporium parvicolla (Drechsler) R.C. Cooke & C.H. Dickinson [as ‘parvicollis’], Trans Br Mycol Soc 48 (4): 622 (1965) |
Characteristics:
Mycelium colourless, sterile hyphae branched, septate, 1.5–3.8 μm wide. Conidiophores colourless, erect, mostly 130–290 μm long, 3–4.5 μm wide at the base, tapering gradually upwards, 1.8–2.2 μm wide at the apex bearing a single conidium. Conidia colourless, commonly spindle-shaped, rounded at the distal end, narrowly truncate at the base, 35–45 × 8–14 μm, 2–4- septate, mainly 4-septate. Capturing nematodes by globose or obovoid or prolate ellipsoid stalks adhesive knobs measuring 6.5–11 × 5.5–9 μm.
Distribution:
China (Beijing, Fujian, Hubei, Shandong, Xizang, Yunnan), Ecuador (National Parc Cotopaxi), Germany (Berlin-Dahlem, Berlin), USA (California, Colorado).
Material examined:
YMF1.00028, YMF1.00029, YMF1.00546, isolated from field soil in Wuhan, Hubei in 2001 by Xuefeng Liu; XZM-8, isolated from forest soil in Xizhang in August 2000 by Minghe Mo.
Discussion:
Da. parvicolla can be distinguished from related species of Dactylellina by its 3–5 septate (mostly 4-septate), spindle-shaped conidia. The conidial dimensions of the present isolate is wider than those of the type strain (29–47 × 6–10.5 μm). (Fig. 3.78)

Dactylellina parvicolla. a conidiophore; b–k conidia; l–m adhesive knobs. Bars = 10 µm; Strain number: YMF1.00029
Dactylellina phymatopaga (Drechsler) Yan Li, in Li, Hyde, Jeewon, Cai, Vijaykrishna & Zhang, Mycologia 97 (5): 1042 (2005) |
Dactylella phymatopaga Drechsler, Mycologia 46: 775 (1954) |
Gamsylella phymatopaga (Drechsler) M. Scholler, Sydowia 51: 109 (1999) |
Golovinia phymatopaga (Drechsler) Mekht., Khishchnye nematofagovye Griby-Gifomitsety (Baku): 165 (1979) |
Monacrosporium phymatopagum (Drechsler) Subram.[as ‘phymatophagum’], J. Indian bot. Soc 42: 293 (1963) |
Characteristics:
Mycelium scanty; vegetative hyphae hyaline, branched, septate at moderate intervals, 1.8–3.5 μm wide. Conidiophores hyaline, erect, septate, 150–500 μm long, 5–9 μm wide at the base, gradually tapering upwards to a width of 2.5–4.5 μm at the apex, bearing a single conidium, occasionally forming 1–2 short branches near the apex with conidia. Conidia hyaline, spindle-shaped, truncate and rather narrow at the base, rounded at the distal end, 46–71 (62.3) × 21–29 (24.7) μm, 1–5-septate, mostly 3–4-septate. Capturing nematodes by means of unstalked adhesive knobs, which can grow out to form branches.
Distribution:
China (Beijing, Hebei, Jilin, Sichuan, Yunnan), Germany (Berlin Dahlem, Berlin), India, Netherlands (Naaldwijk), Sweden.
Material examined:
N4-1, isolated from forest soil in Jiuzhaigou, Sichuan in April 2003 by Xuefeng Liu; YMF1.1413, isolated from forest soil in Dehong, Yunnan in 2003 by Hong Luo. Permanent slide: N4-1
Notes:
This species closely resembles Da. ellipsospora with its solitary conidia, which are spindle-shaped and 4 septa; however, conidia are much longer in Da. ellipsospora. The trapping devices of Da. phymatopaga are not the typical adhesive knobs like those of Da. ellipsospora, which are obovoid rather than globose and can grow out to form adhesive branches. (Fig. 3.79)

Dactylellina phymatopaga. a. conidia; b. adhesive knobs; c. conidiophores. Bar = 40 µm
Dactylellina quercus Bin Liu, Xing Z. Liu & W.Y. Zhuang [as ‘querci’], FEMS Microbiol. Lett. 245 (1): [99] (2005) |
Characteristics:
Colonies colourless on PDA or CMA, reached to 60–65 mm in diameter on PDA and 45 mm CMA after 20 days culture at 23–25 °C. Aerial mycelium sparse, hyphae hyaline, septate, branched, 2.5–3.5 µm wide. Conidiophores mostly 130–180 µm high, 5–6.5 µm wide at the base, 1.5–2.5 µm at the apex, sometime branched near the apex, bearing a single conidium. Conidia were commonly spindle-shaped, slightly rounded at the distal end, narrowly truncate at the base, 25–32.5–40 (–50) × 8–9.5–12 μm, with 3–5 and mainly 3-septate, but the central cell is not larger than the others. Chlamydospores not observed. Trapping nematodes by means of stalked knobs (0–5.5 μm), knobs produced frequently on the nutritional agar plates even without challenging with nematodes.
Sexual state:
Orbilia querci
Distribution:
China (Beijing, Huairou County)
Notes:
This species resembles Da. drechsleri, Da. ellipsospora and Da. haptotyla in the shape of conidia and adhesive knobs, but differs in that it lacks the largest cell in conidia and in having shorter stalks of adhesive knobs. The sequence divergence of the ITS1 region between Da. quercus and the other knobs forming species tested is 23.8–33.4, which separates Da. quercus from its related species. (Fig. 3.80)

Dactylellina quercus. a colony on PDA; b–f conidiophores and conidia, note the single conidium produced on each apex of the conidiophores; g–h stalked knobs. Bars: a = 2 cm; b = 20 μm; c–h = 10 μm
Characteristics:
Colonies on CMA white, slow growing. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched. Conidiophores hyaline, erect, septate, unbranched, 150–450 μm long, 5–8 μm wide at the base and tapering to 3–4 μm wide at the apex, bearing a single conidium. Conidia hyaline, spindle-shaped, 68–85 × 20–30 μm, 3–5-septate. Chlamydospores not observed. Capturing nematodes by means of stout unicellular unstalked adhesive knobs, 16–24 × 9–10 μm.
Distribution:
China (Yunnan), West Australia.
Material examined:
YMF1.1413, isolated from forest soil in Ruili, Yunnan in September 2003 by Hong Luo.
Notes:
Da. robusta resembles Da. phymatopaga (Drechsler) Y. Li (2005), in trapping devices and spore shape, but differs in the larger size of the hyphae, traps and conidia. Da. robusta is easily recognized by its large conidia and its characteristic trapping devices, which just are unicellular adhesive knobs. In Da. phymatopaga, trapping devices can grow out to form adhesive branches. (Fig. 3.81)

Dactylellina robusta. a. conidia; b. adhesive branches; Bar = 20 µm
Dactylellina sichuanensis Yan Li, K.D. Hyde & K.Q. Zhang, in Li, Jeewon, Hyde, Mo & Zhang, Mycol. Res. 110 (7): 792 (2006) |
Characteristics:
Colonies on PDA villiform, whitish, extending to 5 cm diameter at 25 °C, 7 cm diameter at 28℃ and not growing at −4 °C or 35℃ within 7 d. The reverse side of the media is of the same colour. Colonies on CMA whitish, slow-growing, and extending to a diameter of 4 cm at 25 °C, 6 cm 28 °C and not growing at −4 °C or 35 °C within 10 d. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched, mostly 2.5–5 µm wide. Conidiophores hyaline, erect, simple, septate, 139–198.5 µm (168.5 µm) high, 2–2.5 µm wide at the base, gradually tapering upwards to a width of 0.5–1 µm at the apex, bearing one single conidium. Conidia 35–82.5 µm (53.5 µm) × 7.5–17.5 µm (13 µm), hyaline, spindle-shaped, straight or sometimes slightly curved, acutely narrowing towards and truncate at the base, the central cell the largest, (3–)4 (–6) septa; proportion of conidia with 3,4,5 and 6 septa are 14, 76, 9 and 1 %, respectively. Two types of trapping devices were observed in culture: stalked adhesive knobs, 20–50 µm long, unicellular, subsphaerical or prolate ellipsoidal; non-constricting rings approximately circular, composed of three subequal arcuate cells. Chlamydospores were not observed in culture.
Distribution:
China (Sichuan)
Notes:
This species is distinct in having two types of trapping devices and larger conidia from similar species within the same genus. Da. sichuanensis is morphologically similar to Da. lysipaga, which also possesses two types of trapping devices, simple conidiophores and spindle-shaped conidia. However, the main differences between these two are the size of the conidia and the number of conidial septa. Da. sichuanensis produces larger conidia than Da. lysipaga. Only 2–4 septa are present in Da. lysipaga whereas Da. sichuanensis is usually characterized by more than four septa. (Fig. 3.82)

Dactylellina sichuanensis. a–b conidiophore; c–p, r conidia; q adhesive knobs and non-constricting rings. Bars = 20 µm; Strain number: YMF1.00234
Species of the Genus Drechslerella
Drechslerella anchonia (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 99 (1999) |
Arthrobotrys dactyloides Drechsler, Mycologia 29 (4): 486 (1937) |
Arthrobotrys anchonia Drechsler, Mycologia 46: 762 (1954) |
Dactylariopsis dactyloides (Drechsler) Soprunov [as ‘dactyloidea’], Predacious fungi-Hyphomycetes and their use in the control of pathogenic nematodes: 142 (1958) |
Nematophagus anchonius (Drechsler)Mekht., Khishchnye Nematofagovye Griby-Gifomitsety, (Baku): 108 (1979) |
Drechslerella Subram. emend. M. Scholler, Hagedorn & A. Rubner |
Type species: Drechslerella acrochaeta (Drechsler) Subram.–J. Ind. Bot. Soc. 42: 299 (1963) |
Dactylella acrochaeta Drechsler—Mycologia 44: 541 (1952) |
Dactylariopsis Mekht. 1967 |
Type species: D. brochopaga (Drechsler) Mekht.—Mikol. Fitopatol. 1: 278 (1967) |
Dactylaria brochopaga Drechsler—Mycologia 29: 517, 1937 |
Golovinia Mekht. 1967 |
Type species: Golovinia bembicodes (Drechsler) Mekht.—Mikol. Fitopatol.1: 257 (1967) |
Dactylella bembicodes Drechsler—Mycologia 29: 491 (1937) |
Characteristics:
Mycelium slow-growing. Hyphae septate, branching, hyaline. Conidiophores simple or rarely branched, apex mainly simple without modifications, rarely with short denticles or with geniculate proliferations. Conidia mainly formed singly at the apex of the conidiophores, rarely in clusters. Conidia holoblastic, hyaline, one to four septate, mainly ellipsoidal, obovoidal, spindle-shaped, or top-shaped, rarely clavate, or cylindrical. Microconidia and microconidiophores rarely observed. Chlamydospores, when present, intercalary and in chains, thick-walled, sphaerical to ovoid, yellow pigmented. Sexual state, when known, belonging to the genus Orbilia Fr. (Helotiales). Trapping nematodes by means of spontaneously formed three-celled, stalked, constricting rings; saprotrophic capacities weak.
Key to Species of Drechslerella
1. Conidiophores usually with swollen tips and denticles, bearing apical clusters of conidia 2
1. Tips of conidiophores bearing a single conidium, occasionally with very short subapical lateral branches 5
2. Conidia elongate-obovoid, mostly 1-septate, occasionally 2-septate D. anchonia
2. Conidia other types 3
3. Conidia digitiform 4
3. Conidia elongated-ellipsoid, 0–1-septate, mostly 1-septate D. yunnanensis
4. Conidia cylindrical to clavate, mostly 3-septate D. brochopaga
4. With two types of conidia, macroconidia clavate, 1-septate D. dactyloides
5 Conidiophores of the genicularioid type, conidia ellipsoid D. effusa
5. Conidiophores unbranched or occasionally with subapical lateral branches 6
6. Conidia ellipsoid or broadly obovoid 7
6. Conidia spindle-shaped 9
7. Conidia broadly obovoid, with 1-septate at the base D. polybrochum
7. Conidia ellipsoid or wide digitiform 8
8. Having two types of conidia, macroconidia ellipsoid, 2-septate D. heterospora
8. Conidia ellipsoid or wide digitiform, straight or curved, 1–3-septate
D. stenobrocha
9. Conidia 2-septate 10
9. Septa of conidia more than 2 septa 11
10. Conidia 26.5–45 × 14–18.5 μm, bearing an appendage at conidia distal apex D. acrochaetum
10. Conidia 25–52.5 (33.2) × 12.5–29 (17.3) μm, conidiophores with globose apex
D. doedycoides
11. Conidia mostly 3-septate 12
11. Conidia mostly 4-septate 13
12. Conidia narrower, 42.5–62.5 (47) × 15–22.5 (16.9) D. inquisitor
12. Conidia 2–4-septate, mostly 3-septate, 40–57.5 (51) × 15.5–35 (24.6)
D. aphrobrocha
13. Conidia 36–43.2 (40) × 16.8–21.6 (20.5) D. bembicodes
13. Conidia 2–5-septate, 45.6–55.2 (49.5) × 16.8–21.6 (19.8) D. coelobrochum
Accepted Species of Drechslerella
Drechslerella acrochaeta (Drechsler) Subram., J. Indian Bot. Soc. 42: 299 (1964) [1963] |
Dactylella acrochaeta Drechsler, Mycologia 44 (4): 541 (1952) |
Golovinia acrochaeta (Drechsler) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 138 (1979) |
Monacrosporium acrochaetum (Drechsler) R.C. Cooke, Trans. Br. Mycol. Soc. 50 (2): 317 (1967) |
Characteristics:
Colonies on CMA rather slow growing and attaining a diameter of 1 cm within one week at 25 °C. Hyphae hyaline, septate, branched. Conidiophores hyaline, erect, septate, simple, 110–320 μm long, 2–4 μm wide, bearing a single conidium at the apex. Conidia hyaline, broadly spindle-shaped, rounded at the distal end and truncate at the base, 1–2-septate, 26.5–45 × 14–18.5 μm, bearing distally an appendage approximate alignment with the spore axis. Trapping nematodes by means of constricting rings.
Distribution:
China (Guizhou, Hebei, Jilin, Yunnan), USA
Material examined:
95X9–1, isolated from forest soil in Xishuangbanna Yunnan in June 1994 by Xingzhong Liu.
Notes:
The conidial appendage was observed in culture in our isolate. Conidia shape, septa and size of examined material matched well with original description (Drechsler 1952). (Fig. 3.83)

Drechslerella acrochaeta. a. conidia; b. chlamydospore; c. constricting rings; d. conidiophore. Bar = 30 µm
Characteristics:
Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched, 2–5 μm. Conidiophores hyaline, erect, 3–8-septate, simple, 350–500 μm long, 4–6 μm wide at the base, tapering gradually upwards to a width of 2.5–3.5 μm at the apex, there bearing 3–8 conidia in radiating capitate arrangement; conidia hyaline, elongate obovoid, rarely bi-septate, usually 1-septate with the proximal cell commonly 8 to 18 μm (average 11.5) long and the distal cell commonly 17–27 (average 23.5) μm long, 29–43 (35) × 15–19 (16.8) μm, capturing nematodes by means of constricting rings.
Distribution:
USA
Material examined:
No live culture was obtained, so the description is based on the protologue.
Notes:
Among species of Drechslerella, there are four species with conidia in a radiating capitate arrangement at the apex of conidiophores, including Dr. anchonia, Dr. brochopaga, Dr. dactyloides, Dr. yunnanensis. Within these species, Dr. anchonia can be distinguished from the other three species by elongate obovoid and 1-septate conidia. (Fig. 3.84)

Drechslerella anchonia. a. conidia; b. constricting rings; c. conidiophore. Bar = 30 µm
Drechslerella aphrobrocha (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 99 (1999) |
Dactylella aphrobrocha Drechsler, Mycologia 42: 20 (1950) |
Characteristics:
Colonies on CMA slow growing and extending a diameter of 2.6 cm within 7 days at 25 °C. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched, mostly 1.7–3.7 μm. Conidiophores hyaline, erect, 2–5-septate, simple, 162–350 μm long, 3–7.5 μm wide at the base, tapering gradually upwards to a width of 2.5–5 μm at the apex, bearing a single conidium and sometimes a second one on a short perpendicular branch below the apex. Conidia hyaline, broadly spindle-shaped, the central cell the largest, rounded at the distal end and somewhat truncate at the base, 2–4-septate, mostly 3-septate, 40–57.5 (51) × 15.5–35 (24.6) μm, bearing appendages occasionally. Chlamydospores not observed. Capturing nematodes by means of constricting rings, which consist of three cells, outer diameter 20–27.5 μm, inner diameter15–17.5 μm.
Distribution:
Carolina, China (Beijing, Guizhou, Jiling, Yunnan), India, North USA
Material examined:
YMF1.00119, isolated from forest soil in Lijiang, Yunnan in September 2002 by Jing Zhang. Permanent slide: LJ2–11–②
Notes:
The conidia of our isolate matched the original description (41–55 × 17–26 μm) (Drechsler 1950) and is slightly larger than Subramanian (1963) description (46.7 × 21.9 μm). Conidial length and width of Dr. aphrobrocha is very similar to Dr. coelobrocha, but Drechsler (1950) emphasized the greater conidial length and the protracted apical and basal cells of the latter (Rubner 1996). (Fig. 3.85)

Drechslerella aphrobrocha. a–c conidiophore; d–k conidia; l–m constricting rings; n nematode trapped by constricting. Bars = 10 µm; Strain number: YMF1.00119
Drechslerella bembicodes (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 99 (1999) |
Dactylella bembicodes Drechsler, Mycologia 29 (4): 491 (1937) |
Monacrosporium bembicodes (Drechsler) Subram. J. Indian Bot. Soc.42: 293 (1964)[1963] |
Golovinia bembicodes (Drechsler) Mekht., Mikol. Fitopatol. 1: 275 (1967) |
Characteristics:
Mycelium spreading, vegetative hyphae hyaline, septate, mostly 2–5 μm wide. Conidiophores hyaline, erect, septate, 250–450 μm long, 5–7.5 μm wide at the base, tapering gradually upwards to a width of 2–3 μm at the apex, where bear a single conidium. Conidia hyaline, broadly spindle-shaped, broadly rounded at the apex, tapering towards the slightly protruding truncate base, 36–43.2 (40) × 16.8–21.6 (20.5) μm, 3–4-septate, mostly 4-septate. Microconidia developing on conidiophores with several (3–7) subterminal denticles, obovoid, 10–19 × 4–5 μm, usually 1-septate. Chlamydospores not observed. Trapping nematodes by means of constricting rings.
Distribution:
China (Guizhou), India, Netherlands (Schovenhorst near Putten), New Zealand, USA
Material examined:
YMF1.01429, isolated from soil in Kuankuoshui, Guizhou in October 2003 by Yu’e Hao. Permanent slide: YMF1.1429D.
Notes:
The conidia size of our isolate is slightly smaller than in the original description [34–48 (42) × 16–23 (20) μm] (Drechsler 1937). (Fig. 3.86)

Drechslerella bembicodes. a–c conidia and conidiophore; d–f conidia; g constricting rings; h nematode trapped by constricting. Bars = 10 µm; Strain number: YMF1.01429
Drechslerella brochopaga (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 99 (1999) |
Arthrobotrys gracilis (Dudd.) S. Schenck, W.B. Kendr & Pramer, Can. J. Bot. 55 (8): 983 (1977) |
Candelabrella brochopaga (Drechsler) Subram., Kavaka 5: 95 (1977)[1978] |
Dactylaria gracilis Dudd., Trans. Br. Mycol. Soc.34 (2): 194 (1951) |
Dactylariopsis brochopaga (Drechsler) Mekht, Mikol. Fitopatol. 1: 278 (1967) |
Dactylariopsis gracilis (Dudd.) Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 117 (1979) |
Dactylella brochopaga Drechsler, Mycologia 29: 517 (1937) |
Characteristics:
Colonies on CMA initially whitish then turning reddish. Mycelium spreading; vegetative hyphae hyaline, septate, branched. Conidiophores erect, single, septate, 77.5–582.5 μm high, mostly 2.5–5 μm wide at the base, gradually tapering upwards to a width of 2–3 μm at the apex, there bearing on short blunt sterigmata 2–12 μm, mostly 3–8 conidia in radiating capitate arrangement or less often and less typically producing up to 15 conidia in more scattered, irregularly racemose arrangement. Conidia 20–45 (30) × 5–12.5 (6) μm, hyaline, straight of slightly curved, cylindrical or elongate ellipsoidal, broadly rounded at the distal end, usually tapering noticeably towards the somewhat truncate base, 1–3-septate, mainly 3-septate. Capturing nematodes by means of constricting rings.
Distribution:
China (Beijing, Guizhou, Hainan, Hebei, Jiangsu, Jilin, Yunnan), USA (Florida)
Material examined:
Hn38–2–4, isolated from field soil in Haikou, Hainan, China in August 2002 by Ke-Qin Zhang; MW13-5, isolated from forest soil in Mengla, Yunnan, China in 1999 by Yanju Bi; YMF1.00008, isolated from field soil in Simao, Yunnan in September 2000 by Yanju Bi; BS3-3, YMF1.00557, isolated from soil in Baoshan, Yunnan in October 2002 by Jing Zhang. Permanent slide: Hn38-3-4
Notes:
This species resembles Dr. dactyloides in conidia shape, but it mainly has 3 septa while the later often has 1 septum. It occasionally forms conidial traps to trap nematodes including Acrobeles, Acrobeloides, Cephalobus, Diplogaster, Diploscapter, Picctus, Rhabditis and Mononchus. The conidial trap is smaller in diameter than the constricting ring. Dackman & Nordbring (1992) indicated that conidial traps were formed as an adaption to a poor environment. (Fig. 3.87)

Drechslerella brochopaga. a–b, g conidiophore; c–f conidia; h constricting rings. Bars = 10 µm; Strain number: YMF1.00008
Drechslerella coelobrocha (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 99 (1999) |
Dactylella coelobrocha Drechsler, Mycologia 39: 17 (1947) |
Monacrosporium coelobrochum (Drechsler) Subram., J. Indian Bot. Soc. 42: 293 (1963) |
Golovinia coelobrocha (Drechsler) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 141 (1979) |
Characteristics:
Mycelium spreading, vegetative hyphae hyaline, septa, mostly 2–3 μm. Conidiophores hyaline, erect, containing 1–3 septa, mostly in the basal portion, usually unbranched, commonly 180–470 μm long, 3.5–6 μm wide at the base, tapering gradually upwards to a width of 1.5–3 μm at the apex, bearing a single conidium, and sometimes on renewed elongation forming a second conidium. Conidia hyaline, mostly broadly fusiform and strongly ventricose in the centre, 2–5-septate, mostly 4-septate, 45.6–55.2 (49.5) × 16.8–21.6 (19.8) μm, appendages sometimes wholly absent, but at other times present in numbers from 1 to 5, usually arising from the short parabasal cell at right angles to the spore axis, hyaline, filiform, straight, rather rigid. Capturing nematodes by means of constricting rings.
Distribution:
China (Guizhou), USA
Material examined:
YMF1.01480, isolated from soil in Kuankuoshui, Guizhou in October 2003 by Yu’e Hao.
Notes:
The conidial size of Dr. coelobrocha is very similar to Dr. aphrobrocha, but Drechsler (1950) emphasized the greater conidial length, the protracted apical and basal cells in Dr. coelobrocha. This species resembles Da. ellipsospora in conidial shape and septa, but the latter traps nematodes by means of adhesive knobs. Compared with original description, the present isolate has slightly smaller conidia. (Fig. 3.88)

Drechslerella coelobrocha. a–c conidia and conidiophore; d–k conidia; l–m constricting rings; n nematode trapped by constricting. Bars = 10 µm; Strain number: YMF1.01480
Drechslerella dactyloides (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 99 (1999) |
Arthrobotrys dactyloides Drechsler, Mycologia 29 (4): 486 (1937) |
Arthrobotrys anchonia (Drechsler), Mycologia 46 (6): 762 (1954) |
Dactylaria dactyloides (Drechsler) Soprunov [as ‘dactyloidea’], Hyphomycetes and their use in the control of pathogenic nematodes: 142 (1958) |
Dactylariopsis dactyloides (Drechsler) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 120 (1979) |
Nematophagus anchonius (Drechsler) Mekht., Khishchnye Nematofagovye Griby—Gifomitsety (Baku): 108 (1979) |
Characteristics:
Colonies on CMA whitish. Mycelium spreading, vegetative hyphae, branched, septa, mostly 2–5 μm wide. Conidiophores hyaline, septate, erect, mostly 230–650 μm long, 5–6 μm wide at the base, tapering gradually upwards to a width of 1.5–2 μm at the apex, bearing on sterigmata 1–5 μm long and 2–3 μm wide, forming 4–13 conidia usually in a single loose head, more rarely in two somewhat distinct clusters. Conidia 35–51.5 (42.1) × 6.5–8 (7.5) μm, hyaline, usually elongate ellipsoidal or somewhat digitiform, straight or slightly curved, tapering noticeably from the broadly rounded wider distal end, towards the narrower truncate basal end, single septum near the centre of the spore, but occasionally becoming older and shorter, measuring as much as 25–29 × 10–13 μm, then often 1–2-septate, spindly with an inflated central cell greatly exceeding the end cells in size. Capturing nematodes by means of constricting rings.
Distribution:
Canada, China (Beijing, Guizhou, Hebei, Taiwan, Tianjing, Yunnan, Xizhang), Germany (Berlin, Bonn, Jena), South Africa (North West Province), USA (Maryland).
Material examined:
Z258, Z285, Z279, H055, H056, H057, isolated from forest soil in Guizhou and Beijing, in 1992 by Ke-Qin Zhang; YMF1.00131, isolated from field soil in Xizang in 2000 by Minghe Mo; YMF1.00031, YMF1.00555, isolated from field soil in Xundian, Yunnan in March 1999 by Lu Cao; TJ–gzz05, isolated from soil in Gaozhuangzi, Tianjin in 2000 by Wenpeng Li; LJ2–9, isolated from soil in Lijiang, Yunnan in September by Jing Zhang. Permanent slide: D104.
Notes:
Drechslerella dactyloides resembles Arthrobotrys musiformis and A. javanica in conidia shape, but differs in modification of conidiophores, conidia size and trapping–devices. Drechslerella dactyloides forms constricting-rings, while Arthrobotrys musiformis and A. javanica form adhesive three-dimensional networks. In Drechslerella dactyloides, the apex of conidiophores bears a simple and short sterigmata. In Arthrobotrys musiformis and A. javanica, the apex of conidiophores often bears branched and long sterigmata. Drechslerella dactyloides can form conidia traps, which are smaller than constricting rings formed from hyphae, especially in the presence of nematodes. Higgins and Pramer (1967) described the process on conidia trap in detail. In addition, this species is a good material to control Meloidogyne javanica (Stirling et al. 1998). (Fig. 3.89)

Drechslerella dactyloides. a–b, h conidiophore; c–g, i–k conidia; l nematode trapped by constricting; m constricting rings. Bars = 10 µm; Strain number: YMF1.00555
Drechslerella doedycoides (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 99 (1999) |
Dactylella doedycoides Drechsler, Mycologia 32 (4): 454 (1940) |
Golovinia doedycoides (Drechsler) Mekht., Khishchnye Nematofagovye Griby—Gifomisety (Baku): 136 (1979) |
Monacrosporium doedycoides (Drechsler) R.C. Cooke & C.H. Dickinson, Trans. Br. Mycol. Soc. 48: 622 (1965) |
Characteristics:
Colonies on CMA whitish. Mycelium spreading, vegetative hyphae hyaline, septa, branched, 2–4 μm wide. Conidiophores hyaline, erect, 2–6-septate, 200–550 μm long, 5–8 μm wide at the base, tapering gradually upwards to a width of 2–4 μm wide, then expanding abruptly into a knob-like apex 3–5 μm wide, whereon single conidium are borne. Conidia hyaline, somewhat top-shaped, tapering noticeably towards the abruptly or somewhat concavely truncate base, 1–3-septate, mostly 2-septate, 25–52.5 (33.2) × 12.5–29 (17.3) μm. Capturing nematodes by means of constricting rings composed of three cells, out diameter 22–40 μm, inner diameter 10–20 μm.
Distribution:
China (Beijing, Guizhou, Yunnan), Portugal (Quinta de Sao Pedro), UK
Material examined:
YMF1.00553, isolated from soil in Lijiang, Yunnan in September 2002 by Jing Zhang. Permanent slide: LJ4-1
Notes:
The knob-like apex of fertile branches is the crucial feature to distinguish it from other species. Our isolate has the knob-like tips to the conidiophores as in the original description (Cooke and Dickinson, 1965). (Fig. 3.90)

Drechslerella doedycoides. a–d conidia; e–f conidiophore; g constricting rings. Bars = 10 µm; Strain number: YMF1.00553
Drechslerella effusa (Jarow.) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 99 (1999) |
Arthrobotrys effusa (Jarow.) S. Schenck, W.B. Kendr. & Pramer, Can. J. Bot. 55: 982, 1977 |
Dactylaria effusa Jarow., Bull. Acad. Polon. Sci., Cl. II. Sér. sci. biol. 16: 773 (1968) |
Geniculifera effusa (Jarow.) Oorschot, Stud. Mycol. 26: 93 (1985) |
Monacrosporium effusum (Jarow.) Xing Z. Liu & K.Q. Zhang, Mycol. Res. 98: 864 (1994) |
Characteristics:
Colonies on CMA forming a cobweb-like, gray-like coating. Mycelium spreading, scanty, delicate, vegetative hyphae hyaline, branched, thread-like, septate, 2–4 μm wide. Conidiophores straight, erect, simple, septate, 220–400 μm long, 5–7.5 μm wide at the base, gradually tapering upwards to a width of 2.5 μm at the apex, initially with a single apical spore, later bearing a loose head consisting of 2–12 (or more) conidia, supported by short wide, set apart sterigmata. Conidia ellipsoid, widely rounded at the distal end, and widely obtuse, somewhat truncate at the base, 32.5–45 (38.9) × 17.5–25 (21.4) μm, 1–2-septa. The proportion of conidia with 1 and 2 is 25 and 75 % respectively. Microconidia formed from aging hyphae in old culture, clavate, 18–26 × 9–10.5 μm, 0–1-septum.Capturing nematodes by means of constricting rings consisted of three cells, 25–30 μm in outer diameter, 15–20 μm in inner diameter, stalks 7.5–10 μm long, and 5 μm wide.
Distribution:
China (Sichuan), Canada (Manitoba)
Material examined:
YMF1.00583, isolated from forest soil in Jiuzhaigou, Sichuan in April 2003 by Xuefeng Liu. Permanent slide: N58–10.
Notes:
Drechslerella effusa resembles Dr. heterospora and Dr. doedycoides in conidial shape, but differs in conidial size (13–20 × 35–47 μm, and 19.7 × 30–41.4 μm, respectively). Moreover, microconidia present in Dr. effusa, but absent in Dr. doedycoides, while microconidia show somewhat cylindrical shape in Dr. heterospora. (Fig. 3.91)

Drechslerella effusa. a–b conidiophore; c–i conidia; j nematode trapped by constricting; k constricting ring. Bar = 20 µm; Strain number: YMF1.00583
Characteristics:
Colonies on CMA whitish, slow growing and extending to a diameter of 2.9 cm within 7 days at 25°C. Mycelium spreading, scanty, vegetative hyphae hyaline, septate, branched, mostly 2–4 μm. Conidiophores hyaline, erect, septate, simple, occasionally branched, 150–450 μm long, 5–6 μm wide at the base, gradually tapering upwards to a width of 2.5–5 μm at the apex, bearing a single conidium. Conidia hyaline, prolate ellipsoidal, slightly protuberant at the abruptly truncate base, rounded at the distal end, 1–2-septate, 17.5–45 (34) × 17.5–25 (20.4) μm. Microconidiophores often 15–25 μm long, with a cylindrical, curved and somewhat allantoid conidium, 23–40 (31.3) × 5.3–8 (6.8) μm, one septate. Chlamydospores regularly formed, yellowish, in chains. Capturing nematodes by means of constricting rings composed of three cells, 25–35 μm in outer diameter and 17.5–22.5 μm in inner diameter.
Distribution:
Chian (Jilin, Yunnan), USA
Material examined:
YMF1.00550, isolated from forest soil in Deqin, Yunnan in September 2002 by Jing Zhang. Permanent slide: DQ6–1
Notes:
This fungus can be distinguished from other Drechslerella species by its prolate ellipsoidal conidia and slightly protuberant at the abruptly truncate base. (Fig. 3.92)

Drechslerella heterospora. a–b conidia; c–d conidiophore; e constricting rings. Bars = 10 µm; Strain number: YMF1.00550
Drechslerella inquisitor (Jarow.) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 100 (1999) |
Dactylella inquisitor Jarow. [as ‘inquistor’], Acta. Mycologica, Warszawa 7: 4 (1971) |
Characteristics:
Mycelium spreading, vegetative hyphae hyaline, septate, branched, 1.8–2.7 μm wide. Conidiophores hyaline, erect, septate, 193–390 μm long, 5–7.5 μm wide at the base, gradually tapering upwards to a width of 2–4.5 μm at the apex, bearing one single conidium. Conidia hyaline, spindle-shaped, 42.5–62.5 (47) × 15–22.5 (16.9) μm, 1–4-septate, mostly 3-septate. Chlamydospores not observed. Capturing nematodes by means of constricting rings.
Distribution:
China (Yunnan), Poland
Material examined:
YMF1.00585, isolated from soil in Lushui, Yunnan in September 2002 by Jing Zhang. Permanent slide: NJ5–43–3
Notes:
Drechslerella inquisitor resembles Dr. bembicodes in having spindle-shaped and mostly 3 septate conidia. However, the conidia of Dr. inquisitor are much longer and narrower than those of Dr. bembicodes. Our isolate is smaller than the original description (Jarowaja 1971) [50–65 (55) × 18–24 (19) μm], and occasionally formed 1–2 short branches near the apex of the conidiophores. (Fig. 3.93)

Drechslerella inquisitor. a–f conidia; g conidiophore; h constricting rings. Bars = 10 µm; Strain number: YMF1.00585
Drechslerella polybrocha (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 100 (1999) |
Dactylella polybrocha (Drechsler) Mekht., [as‘ polybrochum ’]Dokl. Akad. Nauk Aerb. SSR, 20: 70 (1964) |
Golovinia polybrocha (Drechsler) Mekht., Khishchnye, Nematofagovye Griby Gifomitsety, 137 (1979) |
Monacrosporium polybrochum (Drechsler) Subram., Kavaka 5: 96 (1977)[1978] |
Orbilia tenebricosa (Svrček) Baral, in Yu, Zhang, Qiao, Baral, Weber & Zhang, Mycotaxon 96: 167 (2006) |
Patinella tenebricosa Svrček, česká Mykol. 31 (3): 135 (1977) |
Trichothecium polybrochum Drechsler, Mycologia 29 (4): 536 (1937) |
Characteristics:
Mycelium spreading, vegetative hyphae hyaline, septate, 2 to 4.5 μm wide; conidiophores hyaline, septate, erect, 275–400 μm long, 7 μm wide at the base, gradually tapering upwards to a width of 3 μm at the apex, bearing one single conidium. Conidia hyaline, broadly obovoid, 35 × 24 μm, divided by a single septum into a small obconical basal cell about 6.5 μm long, and a much larger distal cell, about 28.5 μm long, often enveloped in a hyaline layer of mucus 2 to 3 μm thick. Capturing nematodes by means of constricting rings.
Distribution:
USA
Notes:
The description is based on the protologue. This species differs from other species in having 1 septate, broadly obovoid conidia, which is often enveloped in a hyaline layer of mucus when attached on conidiophores. (Fig. 3.94)

Drechslerella polybrocha. a. conidia; b. constricting ring; c. conidiophore. Bars = 25 µm
Drechslerella stenobrocha (Drechsler) M. Scholler, Hagedorn & A. Rubner, Sydowia 51 (1): 100 (1999) |
Dactylella stenobrocha Drechsler, Mycologia 42: 10 (1950) |
Characteristics:
Mycelium scanty; vegetative hyphae hyaline, septate, branched 1.7–3.7 μm wide. Conidiophores hyaline, erect, 425–550 μm long, 4.5–6.5 μm wide at the base, gradually tapering upwards to a width of 2.5–3 μm at the apex, bearing a single conidium. Conidia hyaline, elongate ellipsoidal or broadly finger-shaped or sometimes slightly clavate, straight or slightly curved, somewhat rounded to truncate at the narrowed base, broadly rounded at the apex, 34–56.5 × 12.5–16.5 μm, 1–3-septate. Chlamydospores not present. Capturing nematodes by means of constricting rings.
Distribution:
China (Hebei), USA (Wyoming)
Notes:
The description is based on the protologue. This species can be distinguished from other species of Drechslerella by its ellipsoid or wide digitiform conidia. (Fig. 3.95)

Drechslerella stenobrocha. a. conidia; b. constricting rings; c. conidiophore. Bars = 30 µm
Drechslerella yunnanensis Z.F. Yu & K.Q. Zhang, Mycotaxon 110: 254 (2009) |
Characteristics:
Colonies white, growing slowly on CMA medium, attaining less than 25 mm diam. in 10 days at 25 °C. Vegetative hyphae hyaline, septate, 3.5–4 μm wide, aerial mycelium sparse, hyaline, septate, branched, 2.5–4 μm wide. Conidiophores hyaline, septate, erect, unbranched or occasionally branched below, 60–100 (–220) μm, 3.8–4.2 μm wide at the base, tapering gradually upwards to 1.5–2 μm near the apex, with 2–7 (–10) denticles of 2.3–4.2 μm long, each bearing one conidium in a capitate or racemose arrangement. Conidia hyaline, straight, elongate ellipsoidal, rounded at the apex, with a small truncate protuberance at the base, 7.8–12.9 (–17.8) × 3.3–4.2 μm, (0–)1-septate, proportion of aseptate conidia 17 %. Nematodes are captured by three-celled constricting rings. In the non-constricted state, the outer diameter is 18–21.9 µm, the inner diameter is 12–13.6 µm.
Sexual state:
Orbilia cf. orientalis.
Distribution:
China (Yunnan)
Material examined:
YMF1.01863, isolated from Orbilia cf. orientalis, collected in Yimen, Yunnan in August 2006 by Z. F. Yu.
Notes:
Within species of Drechslerella where conidia are arranged in a cluster at the apex of conidiophores, Dr. yunnanensis can be distinguished from the other three species by the size and shape of conidia. Details are as follows: Dr. brochopaga has much larger conidia (26–46 × 5–9 µm) with more (mainly 3) septa (Drechsler 1937), Dr. anchonia has obovoid, much larger conidia 29–43 (–35) × 15–19 (–16.8) µm with a single septum in the lower third (Drechsler 1954). Conidia of Dr. dactyloides are narrowly ellipsoidal or somewhat digitiform, often very slightly curved, and there are more conidia in the apex of conidiophores and conidia are much larger 32–48 × 7–9.5 μm (Drechsler 1937). (Fig. 3.96)

Drechslerella yunnanensis. a–d conidiophore; e conidia; f constricting rings. Bars: a–c, e–f = 10 µm, d = 50 µm; Strain number: YMF 1.01863
Asexual State-Sexual State Connections of Nematode-Trapping Fungi
Fungi are classified primarily based on the structures associated with sexual reproduction, which tend to be evolutionarily conserved. However, many taxa reproduce only asexually, which cannot easily be placed in a classification based on sexual characters, while some produce both asexual and sexual states. The International Code of Botanical Nomenclature (Vienna Code) (McNeill et al., 2006) previously permitted mycologists to give asexually reproducing fungi (asexual states) separate names from their sexual states (sexual states). When names are available for both the asexual and sexual states of the same taxon, the holomorph usually took the sexual state name. However, where several asexual states were associated with the same sexual morph, this nomenclature could not be followed. Although dual nomenclature has persisted for decades, the concept of permitting separate names for asexual states of fungi with a pleomorphic life-cycle has been an issue of debate since the phenomenon was first recognized in the mid-19th century. More recently, it was deemed that one fungus can only have one name and this is now being put into practice (Taylor, 2011). After 1 January 2013, one taxon can only have one name, the system of permitting separate names to be used for sexual and asexual states is no longer permissible (Hawksworth, 2011). The topic presented here focuses only on the asexual—sexual state connections, but not with name validity, as the latter is still being debated.
Drechsler (1937) was possibly the first to report an asexual state connected to a sexual state in nematode-trapping fungi. He obtained apothecia of an Orbilia-like species in a contaminated culture of Arthrobotrys superba when challenged with nematodes. The apothecia were flesh-coloured, 0.5-0.8 μm diameter, somewhat stalked and with a prominent marginal border and the cylindrical asci were 29-32 × 3 μm. Ascospores were 5 × 1.3 μm, hyaline and tear-shaped. This sexual state was similar to Orbilia fimicola Jeng & Krug in morphology. Zachariah (1983) obtained Orbilia-like apothecia when crossing an auxotrophic strain (D30A) of Drechslerella dactyloides with a prototrophic strain (D31P). However, the apothecia were sterile; no asci were formed. Since that time, at least 14 nematode-trapping asexual species have had credible connections established with Orbilia species and one with Patinella tenebricosa (Table 3.1). In the following, detailed descriptions of 12 asexual nematode-trapping fungi states connected to a sexual state are presented and most illustrated. Two of the connections identified by Pfister (1997), i.e. Drechslerella doedycoides—Orbilaria sp., and Drechslerella polybrochum—Patinella tenebricosa, lack taxonomic descriptions.
Arthrobotrys oligospora Fresen. var. oligospora |
Sexual morph: Orbilia auricolor (A. Bloxam ex Berk.) Sacc. |
Reference: Pfister and Liftik (1995) |
Arthrobotrys cladodes Drechsler var. macroides Drechsler |
Sexual morph: Orbilia auricolor (A. Bloxam ex Berk.) Sacc. |
Reference: Pfister and Liftik (1995) |
Asexual morph (Culture No. 90): Cultures of germinated ascospores on MEYE (Lilly and Barnett, 1951), white, becoming pale tan, forming hyphal strands on the surface and cottony patches, remaining white. On CMA growth sparse, remaining white. Conidiophores erect, branching, not proliferating; each branch terminating in a loose whorl of conidia. Conidia blastic, hologenous, 18–20 × 6–8 μm, elongate-ellipsoid, with a single septum, the cells nearly equal, without constrictions at the septa. Conidia basipetally produced at several closely spaced loci on a sometimes inflated, often distorted, terminal area of the conidiogenous cell; secession schizolytic, leaving a slight protuberant, unthickened, open denticle with a slight frill. Conidiogenous cell determinant, persistent. Cultures induced by nematodes (Cephalobus sp.) normally producing adhesive networks within 12 h.
Sexual morph (Specimen No. 90): Ascomatal specimens were collected by Pfister and Liftik in July, 1994, on decorticated wood from a swampy area, Purgatory, Tract, Westwood, Massachusetts. Ascomata up to 1 mm diameter, yellow to orange, pulvinate to turbinate; asci 8-spored, 30–40 × 3–4 μm, cylindric, J-, tapered towards the base and often forked, at the apex truncate without an obvious pore; ascospores curved, narrowly clavate, broad and rounded at one end, narrowing to an acute point at the other end, non-septate containing a single inclusion which stains intensely in cresyl blue, 8–12.5 × 0.9–1.5 μm; paraphyses branching below, abruptly swelling to become capitate above reaching a diameter of 3-3.5 μm; excipulum of angular cells.
Arthrobotrys cystosporia |
Sexual morph: Orbilia auricolor (A. Bloxam ex Berk.) Sacc. |
Asexual morph (Culture No. SBP88, Fig. 3.97): Colonies colourless, spreading on CMA, reaching 5 cm at 25 °C after 10 days. Vegetative hyphae hyaline, septate, 3.2–5.4 μm wide. Aerial mycelium sparse. Conidiophores hyaline, erect, simple, septate, 60–200 μm high, 2.5–5.5 μm wide at the base, gradually tapering upwards to a width of 1.8–2.5 μm at the apex. Conidia produced at the apex, sometimes in a cluster of 2–5 conidia. Primary conidia hyaline, broadly clavate or broadly turbinate to obovoid, 26–38.5 (31.4) × 13–22 (20) μm, with 2–3 septa. The second conidiophores from the primary conidia colourless, erect, simple, septate, 35–140 μm high, 2–4.5 μm wide at the base and 1.8–2.5 μm at the apex, producing 3–7 secondary conidia on conspicuous denticles loci at near the apex of conidiophores. The secondary conidia hyaline, elongate ellipsoid-cylindrical or slightly clavate, broadly rounded at the apex, rounded truncate at the narrowed base, 26–38.5 (31.4) × 13–22 (20) μm, 0–1-septate. Chlamydospores ellipsoidal. Trapping nematodes by three-dimensional adhesive networks. (Fig. 3.97)

Arthrobotrys cystosporia a–b, d conidiophores; c, h secondary conidia; e adhesive networks; g–i conidia; j chlamydospores. Bars: a, d, f = 20 μm; b, c, e, g–i = 10 μm. Strain number: SBP88
Sexual morph (Specimen No. SBP88, Fig. 3.98): Soils samples were collected from Binchuan County, Yunnan, China on March 21, 2008 by HongYan Su. Subsamples of 2–5 g were spread on CMA plates and stored at 28ºC. After incubation for 10 days, apothecia of an Orbilia sp. were observed on the soil granules, and later also on other areas of the plates. Apothecia superficial, sessile, pale cream. Disc 0.4–0.9 mm diameter, smooth, plane, margin even. Ectal excipulum composed from base to margin of globose or subglobose cells, 5–12 μm diameter, with thin or slightly thickened walls. Asci cylindric-clavate, tapered and often forked at the base, apex rounded or truncate-rounded, 8-spored, 35.5–39 (37) × 3.5–4.7 (4.1) μm in living state. Ascospores banana-shaped or narrowly clavate, medium curved, one end narrower, broadest above and slightly trapped to the rounded at proximal end, non-septate, containing an elongate tear-shaped spore body at the broader end, 7.5–11 (8.6) × 0.9–1.2 μm. Paraphyses hyaline, cylindric-clavate, often branched below, septate in the lower part, 1.4–1.8 (1.6) μm diameter, slightly expanded to 2.3–3 μm at the apex, which is not encrusted. (Fig. 3.98)

Orbilia auricolor a fresh apothecia; b Paraphyses; c–d cell morphology of ectal excipulum; e–g asci; h alive ascospores. Bars = 10 μm. Specimen number: SBP88
Asexual morph (Culture Nos. 45 and 55): Cultures of germinated ascospores on MEYE at first white, becoming rose within 1 wk, growth partially cottony with hyphal strands forming across the surface of the medium. On CMA growth is uniform but sparse, remaining white. Conidiophores erect, with whorls of conidia. Conidia blastic, hologenous, 14–28 × 10–14 μm, obovoid, 1-septate, often constricted at the septum, the proximal cell smaller than the distal cell. Conidia basipetally produced, at several closely spaced loci, on somewhat inflated areas of the conidiogenous cell; secession is schizolytic, leaving protuberant, open, unthickened denticles with frills. Conidiogenous cell are indeterminate, persistent, with hologenous sympodial, presumptive proliferation. Cultures with nematodes (Cephalobus sp.) produced the trap device of adhesive networks within 12 h.
Sexual morph (Specimen Nos. 45 and 55): Ascomatal specimens were obtained from a crustose lichen on a rock which had been incubated in a moist chamber. Taxonomic characteristics of specimen Nos. 45 and 55 were similar as that of specimen No. 90 according to description of Pfister and Liftik (1995).
Arthrobotrys vermicola (R.C. Cooke & Satchuth.) Rifai |
Sexual morph: Orbilia blumenaviensis (Henn.) Baral & E. Weber |
Reference: Qiao et al. (2012) |
Arthrobotrys psychrophilum |
Sexual morph: Orbilia auricolor (A. Bloxam ex Berk.) Sacc. |
Reference: Rubner (1996) |
Asexual morph (Culture Nos. CBS 319.94): The strain was isolated form soil collected from Ecuador (Province Cotopaxi, north of Latacunga, hacienda “La Cienega”). Cultures grew well on CMA medium. Macroconidia were formed after one week on mostly geniculate conidiophores, microconidia appeared later, especially from germinating macroconidia (and also on the fruit bodies). Conidiophores of macroconidia simple, 250–310 μm high, bearing up to 6 conidia. Macroconidia spindle-shaped and mostly 3-septate, with a ratio of length to width of 2.48, 47–52–59.5 × 17.5–21–22.5 μm. Microconidia ellipsoidal to obovoidal and mostly 1-septate, 15-18-22 × 5-6.5-7.5 μm. Chlamydospores in chains, thick-walled, smooth and yellow. When adding the nematode species Turbatrix aceti, the fungus produced adhesive networks.
Sexual morph (Specimen No. A.R.9312): Culture of CBS 319.94 on CMA medium amended with oat husks produced apothecia after two months. Mature apothecia deviating from normal Orbilia apothecia by strongly convex warted hymenia. Asci 45–58 × 3.5–4.2 μm, ascospores 7.5–13 × 1–1.3 μm. An ellipsoid refractive spore body is attached to the spore apex by a thin thread and 3–5 of the lower spores are inversely oriented within asci. Paraphyses slightly inflated at apex. Cortical cells of ectal excipulum terminated by 2–6 μm high glassy caps.
Arthrobotrys superba Corda |
Sexual morph: Orbilia fimicola Jeng & Krug. |
Reference: Pfister (1994) |
Deer dung was collected from Berkshire County in western Massachusetts and was placed in a moist chamber. After 21/2 mouths at room temperature and in natural light, the ascomata discovered on dung was used to project ascospores and obtain the asexual state culture. Pfister (1994) confirmed that taxonomic morphology of the sexual state agreed with the original description of Orbilia fimicola (Jeng and Krug, 1977) and he identified the asexual state as Arthrobotrys superba (Drechsler, 1937), but morphological details on both sexual state and asexual state had not been described.
Arthrobotrys yunnanensis M.H. Mo & K.Q. Zhang |
Sexual morph: Orbilia auricolor (A. Bloxam ex Berk.) Sacc. |
Reference: Mo et al. (2005) |
Asexual morph (Culture No. YMF1.00593, HT1.00593, Fig. 3.99): Colony derived from single ascospore, whitish and slow growing on CMA. Mycelium spreading, scanty; vegetative hyphae hyaline, septate, branched, except for occasional storage filaments that densely filled with protoplasm and up to 11 μm wide measuring mostly 2–7 μm in diameter. Conidiophores hyaline, erect, septate, more or less branched, 68–236 μm long, 2.5–5 μm wide at the base, tapering gradually upwards to a width of 2–3 μm below the irregularly expanded, globose or somewhat coralloid apex whereon are borne 10–20 conidia in usually dense capitate arrangement. Conidia hyaline, ellipsoid or elongate obovoid, mostly 10–20 (17.5) × 5–8 (6.2) μm, 1–septate approximately near in the centre of the spore. Chlamydospores not present. Cultures on CMA produce adhesive networks when nematodes were added (Panagrellus redivivus). (Fig. 3.99)

Arthrobotrys yunnanensis a–c conidiophores with short denticles; c an immature conidium attached to a conidiophore; d–f elongate-ellipsoid-cylindrical or slightly clavate conidia; g adhesive networks; h trapped nematode in adhesive networks; i–j sphaerical to ellipsoidal chlamydospores. Bars: a, d–g, i–j = 10 μm; b, c, h = 20 μm. Strain number: YMF1.00593
Sexual morph (Specimen No. Mo MH O 002, Fig. 3.100): Wet soil samples were from Mt Xiaobailong, Yiliang, Yunnan, China collected by MingHe Mo on 15 August 2003. Subsamples of 2–5 g were spread on CMA plates and stored at room temperature (about 20–28ºC). After incubation for 20 days, apothecia were observed on the soil granules, and later also on other areas of the plates. Apothecia superficial, sessile, pale cream. Disc 0.3–0.8 mm diameter, smooth, plane, margin even. Ectal excipulum composed from base to margin of globose or subglobose cells, 6–12 μm diameter, with thin or slightly thickened walls. Asci cylindric-clavate, tapered and often forked at the base, apex rounded or truncate-rounded, 8-spored, 30–45 × 3.5–5 μm in living state. Ascospores banana-shaped or narrowly clavate, medium curved, one end narrower, broadest above and slightly trapped to the rounded proximal end, non-septate, containing an elongate tear-shaped spore body at the broader end, 9 × 1.4 μm when shooting from asci and still in living state. Paraphyses hyaline, cylindric-clavate, often branched below, septate in the lower part, 1.5–2 μm diameter, slightly expanded to 2.5–3.5 μm at the apex, not encrusted at apex. (Fig. 3.100)

Orbilia auricolor a cluster of asci and paraphyses; b eight living ascospores projected from an ascus; c ascospores within the dead ascus; d a turgescent living ascus (left) and an empty ascus (right) after projection of spores; e paraphyses; f globose to subglobose ectal excipulum cells. Bars: a = 40 μm; b–d = 10 μm; e–f = 7 μm. Specimen number: Mo MH O 002
Arthrobotrys nonseptata Z.F. Yu, S.F. Li & K.Q. Zhang |
Sexual morph: Orbilia sp. |
Reference: Li et al. (2009) |
Asexual morph (Culture No. YMFT 1.01852, Fig. 3.101): Cultures were derived ascospores. Colonies slow-growing on CMA medium, attaining less than 35 mm diameter in 10 days at 25 °C. Vegetative hyphae hyaline, septate, 3.5–4 μm wide, aerial mycelium sparse, hyaline, septate, branched, 2.5–4 μm wide. Conidiophores erect, septate, unbranched, 40–120 μm high, 2–4 μm wide in the lower part, 1.5–2 μm wide at the apex, producing 3–10 conidia from retrogressive conidiogenous loci on conspicuous denticles at and near the apex. Conidia hyaline, non-septate, 11–16.8 × 5–6.6 μm, elongate ellipsoid, constricted at the base by forming a small truncate protuberance. Cultures on CMA produce adhesive networks when nematodes added (Panagrellus redivivus). (Fig. 3.101)

Arthrobotrys nonseptata a conidiophores with short denticles; b conidium attached to a conidiophore; c conidiophores bearing conidia in clusters; d conidia; e adhesive networks. Strain number: YMF1.01852
Sexual morph (Specimen No. YMFT 1.01852, Fig. 3.102): Fresh fruit bodies were collected by Y. Zhang on 18 August 2006 from decaying bark of a broad-leaved tree in forest dominated by Cyclobalanopsis glaucoides Schottky and Pinus armandii Franch., in Dalongkou Park of Yimen County, Yunnan Province, China. The authors confirmed that their specimen belongs to an unidentified species of the genus Orbilia and presented a morphological plate, but did not provide a taxonomic description. (Fig. 3.102)

Orbilia sp a apothecia; b cells of ectal excipulum; c vertical section of part apothecia; d–e cluster of dead asci and paraphyses with living spores; f paraphyses; g asci; h ascospores. Specimen number: YMFT 1.01852
Asexual morph (Culture No. YMF 1.03002, Fig. 3.103): Colonies growing rapidly on CMA medium, attaining 40 mm diameter in 6 days at 28 °C. Mycelium spreading, vegetative hyphae hyaline, septate and branched, mostly 3–6 μm wide. Conidiophores colourless, produced on the mycelium growing at the fringe of the plate, appressed or erect, branched, septate; 150–300 μm high when erect, often 700–1,500 μm when appressed, 4–8 μm wide at the base and 3.5–5 μm at the apex, often recommencing growth after the first group of conidia had been produced and a second head is then formed about 100 μm above the first. This process was repeated until 3–4 (–7) whorls of conidia were produced on a single conidiophore, each group with 2–8 conidia. Two types of colourless conidia were simultaneously formed which mainly differ in their length-width ratio but also in the number of septa: Type a with 1-septate, pear-shaped to obovoid, sometimes elongate ellipsoidal conidia being broadly rounded at the apex, rounded-truncate at the narrowed base, sometimes slightly gradually attenuated at the proximal end, (16.4–)20.4–28.2 × (9.8–)13.5–15.3 (–19) μm; Type b with 1–2 septa, elongate cylindric-clavate, obconical conidia being broadly rounded at the apex, (21.7–)30–34.5 (–39.2) × 12.7–14 μm. The proportion of conidial types a and b was 88 and 12 % respectively. Nematodes are trapped by three-dimensional adhesive networks. (Fig. 3.103)

Arthrobotrys vermicola a–b conidia; c conidiophore and conidia; d adhesive network. Bars = 10 μm. Strain number: YMF 1.03002
Sexual morph (Specimen No. YMFT 1.03002, Fig. 3.104): Specimen was collected on decaying angiosperm wood fallen on the ground of a broad-leaved subtropical evergreen forest located in Xiushan Forest Park of Tonghai County, Yunnan Province, China, in July 2007, by S.F. Li and J.W. Guo. Apothecia 0.7–1.5 mm in diameter, superficial, with a distinct stalk up to 0.2–0.3 mm high, scattered to gregarious on decayed wood, smooth, margin even, yellow and translucent throughout when moist, pale brown when dry. Ectal excipulum composed of globose or subglobose cells, near base 20–43 × 15–38 μm, on flanks 15–30 × 12–25 μm. Asci cylindric-clavate, often forked at the base, apex medium truncate (rounded in side view), 8-spored, 28–39 (–45) × 2.6–3.7 (–4.4) μm in dead state. Ascospores falcate, medium to strongly curved, lower end distinctly narrowed, upper end only slightly so, non-septate, containing a spore body at the broader end, with 1–2 minute lipid bodies in the centre, (6.2–)8–11 (–12) × 1–1.2 μm when shot from asci and still in living state. Spore body rounded or elongate ellipsoid, 0.9–1.6 × 0.7–0.8 μm (1.5–2.5 μm including invisible connecting part). Paraphyses hyaline, cylindrical, with distinctly widened, lanceolate apex, septate below, 1.7–2.8 μm wide at widest point, not encrusted. (Fig. 3.104)

Orbilia blumenaviensis a asci; b paraphyses; c ascospores. Bar = 10 μm. Specimen number: YMFT 1.03002
Dactylellina parvicolla |
Sexual morph: Orbilia cunninghamii |
Reference: Liu et al. (2002) |
Asexual morph: Colonies reached to 25 mm in diameter on PDA after 10 days. Conidia sparse on the nutritionally rich media, spindle-shaped at maturity, 27.5–37.5 × 8–10 μm, mainly with 3–4 septa. Trapping nematodes by means of sessile knobs, occasionally the knob with a short stalk.
Sexual morph: Specimen was collected from Huai-rou County, Beijing, China on rotten wood of Quercus sp.. Apothecia 0.2–0.5 in diameter, asci 20–25 × 3–4 μm, subcylindrical to clavate, tapered and bifurcate at the base, ascospores 5–6 × 1–1.5 μm, rod-shaped to clavate.
Drechslerella brochopaga (Drechsler) M. Scholler, Hagedorn & A. Rubner |
Sexual morph: Orbilia orientalis (Raitv.) Baral |
Reference: Yu et al. (2006) |
Dactylellina quercus Bin Liu, Xing Z. Liu & W.Y. Zhuang |
Sexual morph: Orbilia quercus Bin Liu, Xing Z. Liu & W.Y. Zhuang |
Reference: Liu et al. (2005) |
Asexual morph (Culture No. AS 3.6762, Fig. 3.105): Colonies colourless on PDA or CMA, reached to 60–65 mm in diameter on PDA and 45 mm on CMA after 20 days culture at 23–25°C. Aerial mycelium sparse, hyphae hyaline, septate, branched, 2.5–3.5 μm wide. Conidiophores mostly 130–180 μm high, 5–6.5 μm wide at the base, 1.5–2.5 μm at the apex, sometime branched near the apex, bearing a single conidium. Conidia were commonly spindle-shaped, slightly rounded at the distal end narrowly truncate at the base, 25–50 × 8–12 μm, with 3–5 and mainly 3-septa, but the central cell is not much larger than the others. Chlamydospores not observed. Trapping nematodes by means of stalked knobs (0–5.5 μm), knobs sphaerical to subsphaerical, 8–12 × 7.5–10 μm. Knobs produced frequently on the nutritional agar plates even without challenging with nematodes. (Fig. 3.105)

Dactylellina quercus a colony on PDA; b–f conidiophores and conidia, note the single conidium produced on each apex of the conidiophores; g–h stalked knobs. Bars: a = 2 cm; b = 20 μm; c–h = 10 μm. Strain number: AS 3.6762
Sexual morph (Specimen No. HMAS 88781, Fig. 3.106): Specimen was collected from Labagoumen Forest Park of Huai-rou County, Beijing, China in July, 2002. Apothecia superficial on rotten wood, gregarious, sessile, 0.2–0.5 mm in diameter. Disc concave, smooth, translucent, whitish to pale yellow when fresh, brownish-yellow when dried. Ectal excipulum of textura angularis, 78–96 μm thick, cells isodiametric, 6–10 (–14) μm in diameter. Medullary excipulum of textura intricata, 27–35 μm thick. Subhymenium poorly developed. Asci cylindrical-clavate, narrower and tapered towards the base, sometime forked at the base, apex truncate to rounded, 18–30 × 2.5–3 μm. Ascospores hyaline, subcylindrical to cylindrical-clavate, straight or sometimes slightly curved, non-septate, usually overlapping tightly and biseriate within ascus, 5–6 × 0.8–1.2 μm in water, spore body (SB) tear-shaped to short rod-shaped, 1.1–1.8 × 0.7–1 μm. Paraphyses filiform with a clavate to capitate apex, hyaline, 18–22 μm long, 2–2.5 μm wide at apex and 1.5–2 μm wide below. (Fig. 3.106)

Orbilia quercus a dried apothecia; b vertical section of an apothecium; c–d Asci and ascospores, arrow indicates the SB in living ascospore; e asci; f paraphyses. Bars: a = 100 μm; b = 10 μm; c–f = 5 μm. Specimen number: HMAS 88781
Asexual morph (Culture No. YMF1.01829, Fig. 3.107): Cultures were derived from ascospores. Colonies colourless, spreading on CMA, reaching 48 mm after 14 days at 25 °C. Vegetative hyphae hyaline, septate, 3.5–4 μm wide. Aerial mycelium sparse, hyaline, septate, branched, 2.5–4 μm wide. Conidiophores hyaline, erect, septate, mostly 380–430 μm high, 3.5–4 μm at the base, tapering upwards to a width of 2–2.5 μm at the apex, there bearing short blunt denticles 2–10 μm in length, whereon bearing 3–10 conidia in radiating capitate arrangement, occasionally producing up to 13 conidia in more scattered, irregularly racemose arrangement. Conidia commonly cylindric-oblong, rounded at the distal end, shortly tapered at the somewhat truncate base, 20–36 × 5–7.5 μm, slightly to medium curved especially near distal end, with 1–3 septa, predominantly 3-septate, then terminal cells often exceeding the length of the central cells by a third or even a half. Cultures on CMA produce constricting rings adhesive networks when challenged with nematodes (Panagrellus redivivus). (Fig. 3.107)

Drechslerella brochopaga a–f conidiophores; g–l conidia; m constricting rings. Bars: a–d; g–l = 10 μm; e–f; m = 50 μm. Strain number: YMF 1. 01829
Sexual morph (Specimen No. YMFT1. 01829, Fig. 3.108): Specimens were collected on decaying bark of broadleaved tree in a forest at 848 m altitude, located in Jinggangshan Park of Jian City, Jiangxi Province, China, in October 2005 by Y. Zhang. Apothecia 0.8–1.2 mm in diameter, superficial, with a distinct stalk up to 0.2–0.3 mm high, scattered to gregarious on decayed bark, white and translucent throughout when moist, pale brown when dry. Disc slightly concave to flat, margin even and smooth. Ectal excipulum of angular or globose cells, 8–13.8 μm in diameter, with thin or only slightly thickened walls, towards margin of textura prismatica-angularis oriented at a high angle, marginal cells tipped by glassy processes up to 30–40 × 3.5–4.5 μm, curved outwards, agglutinated to form rounded teeth. Medullary excipulum rather thick, of texture globulosa (-prismatica) with intermingled hyphae, subhymenium poorly developed. Asci 29.5–38 × 3.2–4.5 μm, 8-spored, lower (2)3–5 (6) spores inversely oriented (with spore body towards ascus base), cylindric, rounded or truncate at the apex (depending on side of view), tapered and often forked at the base (L, hor H-shaped). Ascospores hyaline, non-septate, cylindric- ellipsoid, sometimes slightly tapered at lower end, 3–4 × 1.6–1.8 μm, with a refractive rod-shaped spore body (SB) at upper end in living mature ascospores, 1.2–1.4 × 0.3–0.5 μm in diameter. Paraphyses 1.3–2 μm wide, enlarged to 2.8–3.7 μm in diameter at the medium to rather strongly clavate or capitate apex, terminal cell much longer than lower cells, apex covered by a rough thin layer of exudates. (Fig. 3.108)

Orbilia orientalis a hymenium; b cells of ectal excipulum; c–e dead asci; f living ascospores with globose spore bodies; g–i Dead paraphyses; j–k cluster of dead asci and paraphyses with living spores. Bar: a–e, g–k = 10 μm; f = 5 μm. Specimen number: YMFT 1. 01829
Drechslerella yunnanensis Z.F. Yu & K.Q. Zhang |
Orbilia cf. orientalis (Raitv.) Baral. |
Reference: Yu et al. (2009) |
Asexual morph (Culture No. YMF1.01863, Fig. 3.109): Cultures were derived from ascospores. Colonies white, slow-growing on CMA medium, attaining less than 25 mm diameter in 10 days at 25 °C. Vegetative hyphae hyaline, septate, 3.5–4 μm wide, aerial mycelium sparse, hyaline, septate, branched, 2.5–4 μm wide. Conidiophores hyaline, septate, erect, unbranched or occasionally branched below, 60–100 (–220) μm, 3.8–4.2 μm wide at the base, tapering gradually upwards to 1.5–2 μm near the apex, with 2–7 (–10) denticles 2.3–4.2 μm long, each bearing one conidium in a capitate or racemose arrangement. Conidia hyaline, straight, elongate ellipsoidal, rounded at the apex, with a small truncate protuberance at the base, 7.8–12.9 (–17.8) × 3.3–4.2 μm, (0-)1 septate, proportion of aseptate conidia 17 %. Cultures on CMA produce the trap device of constricting rings when induced by nematodes (Panagrellus redivivus). (Fig. 3.109)

Drechslerella yunnanensis a–c conidiophores with short denticles; d conidiophores bearing conidia in clusters; e conidia; f constricting rings. Bar: a–c, e–f = 10 μm; d = 50 μm. Strain number: YMF 1.01863
The sexual morph (specimen No. YMFT 1.01863, Fig. 3.110) was collected as fresh fruit bodies by ZeFen Yu in August 2006 from decaying bark of a broad-leaved tree, in Dalongkou Park of Yimen County, Yunnan Province, China (N24°34, E101º00´, alt. 1580 m, coniferous-broadleaf forest dominated by Cyclobalanopsis glaucoides Schottky. and Pinus armandii Franch.). The sexual state is very similar to Orbilia orientalis in having cylindric-ellipsoid ascospores with a refractive, rod-shaped, laterally arranged spore body (SB) at the upper end. Also spore size is very close (protologue of O. orientalis: 3–4 × 1.5–2 µm, the specimen YMFT1.01863: 4.0–4.2 × 1.6–1.9 μm). The apothecial margin is only minutely crenulate and hardly bears any glassy processes, while in the O. orientalis type, the glassy processes are long and agglutinated, forming distinct teeth at the margin. (Fig. 3.110)

Orbilia cf. orientalis a apothecium; b cells of excipulum; c asci; d cluster of asci and paraphyses; e paraphyses; f ascospores. Bar: b–f = 10 μm. Specimen number: YMFT 1.01863
Conclusion
In the classification of nematode-trapping fungi, trapping devices are generally more informative than the morphologies of the conidia (shape, number and size of cells) and conidiophores (branching, modification of the apex). The latter characters were previously used in delimiting genera. We conclude that all orbiliaceous nematode-trapping fungi should be placed in the genera of Arthrobotrys, Dactylellina and Drechslerella based on their trap types. Arthrobotrys includes species producing adhesive networks, Dactylellina contain species forming stalked adhesive knobs, unstalked adhesive knobs, or non-constricting rings, while Drechslerella includes species producing constricting rings. Species lacking trap formation, especially members of the genus Dactylella, are excluded from the nematode-trapping fungi, although some share similar morphologies to species of Dactylellina.
A total of 96 species of orbiliaceous nematode-trapping fungi are described and illustrated in detailed, with 54 species from Arthrobotrys, 28 from Dactylellina and 14 from Drechslerella. Nematode-trapping fungi have been found in all parts of the world and in all climatic zones. Most of the nematode-trapping fungi annotated here were isolated from agricultural soils and forest soils, while some of them also inhabit freshwater. The majority of nematode-trapping fungi are fairly ubiquitous, with few species restricted geographically. Species forming adhesive networks are the dominant group.
Fourteen connections have been established between asexual nematode-trapping fungi and their orbiliaceous sexual state. Five asexual species are associated with Orbilia auricolor suggesting that O. auricolor is a species complex. It is difficult to distinguish between members of this complex based only morphological characters of the sexual state. Beside the morphological features, more molecular information, such as gene sequences of ITS, 28S rDNA, 5.8S rDNA and ß-tubulin, should be taken into account in the taxonomy of Orbilia.
References
Ahrèn, D., Ursing, B. M., & Tunlid, A. (1998). Phylogeny of nematode-trapping fungi based on 18S rDNA sequences. FEMS Microbiology Letters, 158, 179–184.
Anastasiou, C. J. (1964). Some aquatic fungi imperfecti from Hawaii. Pacific Science, 18, 202–206.
Barron, G. L. (Ed.). (1977). The Nematode-Destroying Fungi. Topics in Mycology No. 1. Canadian Biological Publications, Guelph.
Barron, G. L. (1979). Nematophagous fungi: A new Arthrobotrys with aseptate conidia. Canadian Journal of Botany, 55, 1371–1373.
Barron, G. L. (1990). A new predatory hyphomycete capturing copepods. Canadian Journal of Botany, 68, 691–696.
Barron, G. L., & Davidson, J. G. (1972). Nematophagous hyphomycetes: Arthrobotrys anomala sp. nov. Canadian Journal of Botany, 50, 1773–1774.
Castaner, D. (1968). Monacrosporiumsphaeroides, a new nematode-destroying hyphomycete from Iowa. The American Midland. Naturalist, 80, 280–283.
Chen, J. L., Huang, T. L., & Tzean, S. S. (1998). Dactylella huisuniana, a new nematode-trapping fungus from Taiwan. Mycological Research, 102(10), 1269–1273.
Chen, J., Xu, L. L., Liu, B., & Liu, X. Z. (2007a). Taxonomy of Dactylella complex and Vermispora. I. Generic concepts based on morphology and ITS sequences data. Fungal Diversity, 26, 73–83.
Chen, J., Xu, L. L., Liu, B., & Liu, X. Z. (2007b). Taxonomy of Dactylella complex and Vermispora. II. Generic concepts based on morphology and ITS sequences data. Fungal Diversity, 26, 85–126.
Chen, J., Xu, L. L., Liu, B., & Liu, X. Z. (2007c). Taxonomy of Dactylella complex and Vermispora. III. Generic concepts based on morphology and ITS sequences data. Fungal Diversity, 26, 127–142.
Chowdhry, P. N., & Bahl, N. (1982). Notes on Indian hyphomycetes—VII Monacrosporiealla. Current Science, 20, 894–896.
Cooke, R. C. (1969). Two nematode-trapping hyphomycetes, Duddingtonia flagrans gen. et. comb. nov. and Monacrosporium mutabilis sp. nov. Transactions of the British Mycological Society, 53, 315–318.
Cooke, R. C., & Dickison, C. H. (1965). Nematode-trapping species of Dactylella and Monacrosporium. Transactions of the British Mycological Society, 48, 621–629.
Cooke, R. C., & Pramer, D. (1968). Monacrosporium rutgeriensis sp. nov., a nematode-trapping hyphomycetes. Phytopathology, 58, 544–545.
Cooke, R. C., & Satchuthananthavale, V. (1965). Some nematode-trapping species of Dactylaria. Transactions of the British Mycological Society, 49, 27–32.
Corda, A. G. J. (1839). Pracht-Flora europaischer Schimmelbildungen, vol 43. Leipzig.
Dackman, C., & Norbring, B. (1992). Nematophagous fungi and their activities in soil. Soil Biochemistry, 7, 95-130
Dixon, S. M. (1952). Predacious fungi from rotton-wood. Transactions of the British Mycological Society, 35, 144–148.
Dowsett, J. A., Reid, J., & Kalkat, R. S. (1984). A new species of Dactylella. Mycologia, 76, 563–566.
Drechsler, C. (1937). Some hyphomycetes that prey on free-living terricolous nematodes. Mycologia, 29, 447–552.
Drechsler, C. (1940). Three new hyphomycetes preying on free-living terricolous nematodes. Mycologia, 32, 448–470.
Drechsler, C. (1941). Four hyphomycetes destructive to nematodes and rhizopods. Mycologia, 33, 248–269.
Drechsler, C. (1943). A new nematode-capturing Dactylella and several related hyphomycetes. Mycologia, 35, 339–362.
Drechsler, C. (1944a). Three hyphomycetes that capture nematodes in adhesive networks. Mycologia, 36, 138–171.
Drechsler, C. (1944b). A species of Arthrobotrys that captures springtails. Mycologia, 36, 383–397.
Drechsler, C. (1947). A nematode-strangling Dactylella with broad quadriseptate conidia. Mycologia, 39, 5–18.
Drechsler, C. (1950). Several species of Dactylella and Dactylaria that capture free-living nematodes. Mycologia, 42, 1–79.
Drechsler, C. (1952). Another nematode-strangulating Dactylella and some related hyphomycetes. Mycologia, 44, 533–556.
Drechsler, C. (1954). Some hyphomycetes that capture eelworms in southern states. Mycologia, 46, 762–782.
Drechsler, C. (1962). Some clampless hyphomycetes predaceous on nematodes and rhizopods. Sydowia, 15, 9–25.
Duddington, C. L. (1951a). Two predacious hyphomycetes. Transactions of the British Mycological Society, 34, 598–603.
Duddington, C. L. (1951b). Dactylella lobata, predacious on nematodes. Transactions of the British Mycological Society, 34, 489–491.
Eriksson, O., Baral, H., Currah, R., Hansen, K., Kurtzman, C., Lassoe, T., & Rambold, G. (2003). Notes on ascomycete systematics Nos 3580-3623. Myconet, 9, 91–103.
Glockling, S., & Dick, M. (1994). Dactylella megalobrocha, a new species of nematophagous fungus with constricting ring. Mycological Research, 98, 845–853.
Grove, W. B. (1884). New or noteworthy fungi. American Journal of Botany, 22, 195–201.
Hagedorn, G., & Scholler, M. (1999). A re-evaluation of predatory orbiliaceous fungi. I. Phylogenetic analysis using rDNA sequence data. Sydowia, 51, 27–48.
Hao, Y. E., Luo, J., & Zhang, K. Q. (2005). A new aquatic nematode-trapping hyphomycete. Mycotaxon, 89, 235–239.
Hawksworth, D. L. (2011). A new dawn for the naming of fungi: Impacts of decisions made in Melbourne in July 2011 on the future publication and regulation of fungal names. International Mycological Association Fungus, 2, 155–162.
Higgins, M. L., & Pramer, D. (1967). Fungal morphogenesis: ring formation and closure by Arthrobotrys dactyloides. Science, 155, 345–346.
Jarowaja, N. (1968). Dactylaria effusa sp. nov. -na new nematode-trapping hyphomycetes. Bulletin de l’Academie Polonaise des Sciences. Serie des Sciences Biologiques, 16, 773–778.
Jarowaja, N. (1970). The genus Arthrobotrys Corda. Acta Mycology, 6, 337–406.
Jarowaja, N. (1971). Dactylella inquisitor sp. nov. a new nematode-strangulating fungus. Acta Mycology, 7, 3–6.
Jeng, R. S., & Krug, J. C. (1977). A new species of Orbilia from Venezuela. Mycologia, 69, 832–835
Juniper, A. J. (1954). Dactylaria pyriformis sp. nov. Transactions of the British Mycological Society, 37, 437–440.
Kuthubutheen, A. J., & Webster, J. (1985). Arthrobotrys dendroides, a new nematode-trapping synnematous Arthrobotrys from Malaysia. Transactions of the British Mycological Society, 84, 563.
Li, S. D., Miao, Z. Q., Zhang, Y. H., & Liu, X. Z. (2003). Monacrosporium janus sp. nov., a new nematode-trapping hyphomycetes parasitizing sclerotia and hyphae of Sclerotinia sclerotiorum. Mycological Research, 107, 888–894.
Li, Y., Hyde, K. D., Jeewon, R., Cai, L., Vijaykrishna, D., & Zhang, K. Q. (2005). Phylogenetics and evolution of nematode-trapping fungi estimated from nuclear and protein coding genes. Mycologia, 97, 1034–1046.
Li, Y., Jeewon, R., Hyde, K. D., Mo, M. H., & Zhang, K. Q. (2006). Two new species of nematode-trapping fungi:relationships inferred from morphology, rDNA and protein gene sequence analyses. Mycological Research, 110, 790–800.
Li, S. F., Yu, Z. F., Zhang, Y., Guo, J. W., & Zhang, K. Q. (2009). Arthrobotrys nonseptata, a new asexual state from Orbilia sp. Mycotaxon, 109, 247–254.
Lilly, V. G., & Barnett, H. L. (1951). Physiology of the Fungi (pp. 464). New York: McGraw-Hill.
Liou, J. Y., Liou, G. Y., & Tzean, S. S. (1995). Dactylella formosana, a new nematode-trapping fungus from Taiwan. Mycological Research, 99, 751–755.
Liu, X. Z., & Lu, B. S. (1993). Two new species of Monacrosporium from China. Mycosystema, 6, 65–69.
Liu, X. Z., & Zhang, K. Q. (1994). Nematode-trapping species of Monacrosporium with special reference to two new species. Mycological Research, 98, 862–868.
Liu, X. Z., & Zhang, K. Q. (2003). Dactylella shizishanna sp. nov., from Shizi Mountain, China. Fungal Diversity, 14, 103–107.
Liu, B., Yang, Y., Liu, X. Z., & Zhuang, W. Y. (2002). Monacrosporium parvicolla, a knob-forming nematophagous hyphomycetes from Orbilia cunninghamii. In: The 3rd Asia-Pacific Mycological Congress on Biodiversity and Biotechnology, P. 75 (abstract).
Liu, B., Liu, X., & Zhuang, W. Y. (2005a). Orbilia querci sp. nov. and its knob-forming nematophagous asexual state. FEMS Microbiology Letters, 245, 99–105.
Liu, B., Liu, X., & Zhuang, W. Y. (2005b). A new species of Hyalorbilia and its asexual state from China. Novartis Foundation Symposium, 81, 145–155.
Luo, H., Mo, M. H., Huang, X. W., Li, X., & Zhang, K. Q. (2004). Coprinus comatus: A basidiomycete fungus forms novel spiny structures and infects nematodes. Mycologia, 96, 1218–1225.
Luo, H., Li, X., Pan, Y. B., Li, G. H., & Zhang, K. Q. (2006). Acanthocytes of Stropharia rugoso-annulata function as a nematode-attacking device. Applied and Environmental Microbiology, 72, 2982–2987.
Luo, H., Liu, Y. J., Fang, L., Li, X., Tang, N. H., & Zhang, K. Q. (2007). Coprinus comatus: Damages nematode cuticles mechanically with spiny balls and produces potent toxins to immobilize nematodes. Applied and Environmental Microbiology, 72, 3916–3923.
McCulloch, J. S. (1977). New species of nematophagous fungi from Queensland. Transactions of the British Mycological Society, 68, 173–179.
McNeill, J., Barrie, F. R., Burdet, H. M., Demoulin, V., Hawksworth, D. L., Marhold, K., Nicolson, D. H., Prado, J., Silva, P. C., Skog, J. E., & Wiersema, J. H. (Eds.). (2006). International Code of Botanical Nomenclature (Vienna Code) adopted by the Seventeenth Interational Botanical Congress, Vienna, Austria, July 2005. (Regnum Vegetabile No. 146.) Ruggell: A.R.G. GantnerVerlag.
Mekhtieva, N. A. (1973). Species novae fungorum carnivororum generis Arthrobotrys Corda. Nov Sist Niz Rast, 10, 174–176.
Mekhtieva, N. A. (1979). Khishchnye Nematofagovye Griby—Gifomitsety (Predacious Nematophagous Hyphomycetes). A Kademiya Nauk Azerbaidzhanskoi SSR. Baku.
Mekhtieva, N. A. (1997). A new nematophagous fungus, Candelabrella shahriari sp. nov., from south Azerbaijan. Mycological Research, 101, 334.
Mekhtieva, N. A. (1998). New nematophagous fungi from Azerbaijan. Mycological Research, 102, 683–684.
Miao, Z. Q., Liu, X. Z., Li, S. D., & He, M. X. (2003). Dactylella pseudoclavata sp. nov., a new nematode-trapping fungus. Canadian Journal of Botany, 81, 1–5.
Mo, M. H., Huang, X. W., Zhou, W., Huang, Y., Hao, Y. E., & Zhang, K. Q. (2005a). Arthrobotrys yunnanensis sp. nov., the fourth asexual state of Orbilia auricolor. Fungal Diversity, 18, 107–115.
Mo, M. H., Zhou, W., Huang, Y., Yu, Z. F., & Zhang, K. Q. (2005b). A new species of Dactylella and its sexual state. Mycotaxon, 91, 185–192.
Morelet, M. (1968). De aliquibus in mycologia novitatibus. Bulletin de la Société des Sciences Naturelles and d’Archéologie de Toulon and du Var, 178, 6.
Nees, V., & Esenbeck, C. G. D. (1817). Das system der Pilze und Schwämme. Würzburg.
Onofri, S., & Tosi, S. (1992). Arthrobotrys ferox sp. nov., a springtail-capturing hyphomycete from continental antarctica. Mycotaxon, 445–451.
Oorshot, C. A. N. (1985). Taxonomy of the Dactylaria complex V. A review of Arthrobotrys and allied genera. In Hoog, G.S. DE (Ed.).Taxonomy in the Dactylaria Complex, IV–VI, Studies in mycology 26: 61–96.
Oudemans, C. A. (1885). Aanwinstenvoor de flora mycological van Nederland. Ned Kruidk Archf Ser, 24, 236–257.
Peach, M. (1950). Aquatic predacious fungi. Transactions of the British Mycological Society, 33, 148–153.
Peach, M. (1952). Aquatic predacious fungi. Transactions of the British Mycological Society, 35, 19–23.
Pfister, D. H. (1994). Orbilia fimicola, a nematophagous discomycetes and its Arthrobotrys anamorph. Mycologia, 86, 451–453.
Pfister, D. H. & Liftik, M. E. (1995). Two Arthrobotrys anamorphs from Orbilia auricolor. Mycologia, 87, 684–688.
Pfister, D. H. (1997). Castor, Pollux and life histories of fungi. Mycologia, 89, 1–23.
Qiao, M., Zhang, Y., Li, S. F., Baral, H. O., Su, H. Y., Xu, J. P., Zhang, K. Q., & Yu, Z. F. (2012). Orbilia blumenaviensis and its Arthrobotrys asexual state. Mycological Progress, 11, 255–262.
Rifai, M. A. (1968). The hyphomycetes genus Dactylaria Sacc. Reninwardtia, 7, 357–374.
Rifai, M. A., & Cooke, R. C. (1966). Studies on some didymosporous genera of nematode-trapping hyphomycetes. Transactions of the British Mycological Society, 49, 147–168.
Rubner, A. (1996). Revision of predacious hyphomycetes in the Dactylella-Monacrosporium complex. Studies in Mycology, 39, 1–134.
Scheuer, C. H., & Webster, J. (1990). Dactylella arcuata sp. nov., a nematode-trapping hyphomycete. Mycological Research, 94, 718.
Schenck, S., Kendrick, W. B., & Pramer, D. (1977). A new nematode-trapping hyphomycetes and a revaluation of Dactylaria and Arthrobotrys. Canadian Journal of Botany, 55, 977–985.
Scholler, M., & Rubner, A. (1999). Arthrobotrys hertziana sp. nov. from the Canary Islands. Mycological Research, 103, 764–768.
Scholler, M., Hagedorn, G., & Rubner, A. (1999). A reevaluation of predatory orbiliaceous fungi. II. A new generic concept. Sydowia, 51, 89–113.
Scholler, M., Hagedorn, G., & Rubner, A. (2000). Arthrobotrys oudemansii comb. nov., a new name for a nematode-trapping fungus to avoid homonymy. Sydowia, 52, 59–60.
Siddiqui, Z. A., & Mahmood, I. (1996). Biological control of plant parasitic nematodes by fungi: A review. Bioresource Technology, 58, 229–239.
Soprunov, F. F. (1958). Kishchnyegriby-gifomitsetiiikhprimenenie v bor’be s patogennyminematodami (Predcious hyphomycetes and their application in the control of pathogenic nematodes). Akademiya Nauk Turkmenskoi SSR, Ashkhabad. (Translation published by S. Monson, Jerusalem).
Stirling, G. R., Nicol, J. M., & Reay, F. (1998). Advisory services for nematodes pests - operational guide. Rural Industries Research and Development Corporation Publication No. 99/41. Canberra. pp 120.
Su, H. Y., Li, Y., Zhang, K. Q., & Mo, M. H. (2005). Monacrosporium multiseptatum, a new species from China. Mycotaxon, 92, 193–196.
Su, H. Y., Hao, Y. E., Yang, X. Y., Yu, Z. F., Deng, J. S., & Mo, M. H. (2008). A new species of Dactylellina producing adhesive knobs and non-constricting rings to capture nematodes. Mycotaxon, 105, 313–316.
Su, H. Y., Liu, S. R., Li, Y. X., Cao, Y. H., Chen, M. H., & Yang, X. Y. (2011). Arthrobotrys latispora, a new nematode-trapping fungus from southwest China. Mycotaxon, 117, 29–36.
Subramanian, C. V. (1963). Dactylella, Monacrosporium and Dactylina. Journal of Indian Botanical Society, 42, 291–300.
Swe, A., Jeewon, R., Pointing, S. B., & Hyde, K. D. (2008). Taxonomy and molecular phylogeny of Arthrobotrys mangrovispora, a new marine nematode-trapping fungal species. Botanica Marina, 51, 331–338.
Tarjan, A. C. (1961). Growth characteristics of Dactylella drechsleri nov. sp., an adhesive-knobbed nematode-trapping hyphomycete from Florida. Mycopath. Mycopathology Mycology Applied, 14, 136–144.
Taylor, J. W. (2011). One Fungus = One Name: DNA and fungal nomenclature twenty years after PCR. International Mycological Association Fungus, 2(2), 113–120.
Tzean, S. S., & Liou, J. Y. (1993). Nematophagous resupinate basidiomycetous fungi. Phytopathology, 83, 1015–1020.
Yang, Y., & Liu, X. Z. (2005). Dactylella coccinella sp. nov., an asexual stateic species. Mycotaxon, 91, 127–132.
Yang, Y., & Liu, X. Z. (2006). A new generic approach to the taxonomy of predatory asexual stateic Orbiliaceae (Ascomycotina). Mycotaxon, 97, 153–161.
Yang, D. S., Chen, W. M., Huang, Y., Mo, M. H., & Zhang, K. Q. (2005). A new predatory fungus from China. Mycotaxon, 94, 215–217.
Yang, Y., Yang, E. C., An, Z. Q., & Liu, X. Z. (2007). Evolution of nematode-trapping cells of predatory fungi of the Orbiliaceae based on evidence from rRNA-encoding DNA and multiprotein sequences. Proceedings of National Academy of Science of USA of America, 104, 8379–8384.
Yu, Z. F., Zhang, Y., Qiao, M., Baral, H. O., Weber, E., & Zhang, K. Q. (2006). Drechslerella brochopaga, the asexual state of Orbilia (Hyalinia) orientalis. Mycotaxon, 96, 163–168.
Yu, Z. F., Qiao, M., Zhang, Y., Baral, H. O., & Zhang, K. Q. (2007a). Orbilia vermiformis sp. nov. and its asexual state. Mycotaxon, 99, 271–278.
Yu, Z. F., Zhang, Y., Qiao, M., & Zhang, K. Q. (2007b). Orbilia dorsalia sp. nov., the sexual state of Dactylella dorsalia sp. nov. Cryptogamie Mycologie, 28(1), 55–63.
Yu, Z. F., Qin, L., Zhang, Y., Qiao, M., Kong, Y. J., & Zhang, K. Q. (2009a). A new Drechslerella species isolated from Orbilia cf. orientalis. Mycotaxon, 110, 253–259.
Yu, Z. F., Kong, Y. J., Zhang, Y., Qiao, M., Guo, J. W., & Zhang, K. Q. (2009b). A new Dactylella species from Orbilia alba. Journal of Microbiology, 47(3), 265–269.
Yu, Z. F., Qiao, M., Zhang, Y., Qin, L., & Zhang, K. Q. (2011). Pseudotripoconidium, a new asexual state genus connected to Orbilia. Mycologia, 103, 164–173.
Zachariah, K. (1983). Ascocarp induction in a natural autotroph of a predatory fungus. Canadian Journal of Botany, 62, 3262–3266
Zhang, K. Q., Liu, X. Z., & Cao, L. (1996a). Nematophagous species of Monacrosporium from China. Mycological Research, 100, 274–276.
Zhang, K. Q., Liu, X. Z., Cao, L., & Gao, R. H. (1996b). A new species Arthrobotrys from China. Mycological Research, 100, 527–530.
Li, T. F., Zhang, K. Q., & Liu, X. Z. (2000). Taxonomy of nematophagous fungi (Chinese). Beijing: Science Press.
Zhang KQ, Liu XZ, Li TF. (Ed.). (2001). Biology of nematophagous fungi. Chinese Scientific & Technological Publication, Beijing, China.
Pfister DH. (1997). Castor, Pollux and life histories of fungi. Mycologia 89: 1–23
Zhang KQ, Liu XZ, Li TF. (Ed.). (2001). Biology of nematophagous fungi. Chinese Scientific & Technological Publication, Beijing, China.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Mushroom Research Foundation
About this chapter
Cite this chapter
Yu, Z., Mo, M., Zhang, Y., Zhang, KQ. (2014). Taxonomy of Nematode-Trapping Fungi from Orbiliaceae, Ascomycota. In: Zhang, KQ., Hyde, K. (eds) Nematode-Trapping Fungi. Fungal Diversity Research Series, vol 23. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-8730-7_3
Download citation
DOI: https://doi.org/10.1007/978-94-017-8730-7_3
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-8729-1
Online ISBN: 978-94-017-8730-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)