
Abstract
Vascular plant endemism in the diverse vegetation zones and physiognomic habitat types of some islands or archipelagos is described and analysed in detail. The Juan Fernández Islands, Madeira Archipelago, Corsica and Madagascar, located in different regions and oceans of the world, present their own unique evolutionary histories and composition of habitats and endemics.
The concentration of endemics on Juan Fernández Islands, the Madeira archipelago, Corsica and additionally Tenerife is compared directly, as the absolute numbers of endemic taxa there are similar.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Juan Fernández Archipelago
1.1 Introduction
The Juan Fernández Archipelago is located around latitude 34°S offshore Chile, west of the Southern Cone of South America (Fig. 6.1). It comprises three small oceanic islands: Isla Robinson Crusoe (also known as Masatierra), situated at 670 km from the South American continent and encompassing 49 km2; the little Isla Santa Clara (2.2 km2), situated to the SSW of Robinson Crusoe; and Alejandro Selkirk (also known as Masafuera), situated 167 km to the west of the former two, and occupying an area of 53 km2. The biotic and especially botanical value of these islands has long being recognized (Johow 1896; Skottsberg 1953; Muñoz Pizarro 1969; Marticorena et al. 1998; Danton 2006). Due to the steady suggestions from early botanists, the archipelago has been declared a National Park in 1935, also getting the status of Biosphere Reserve in 1977.

The geographical position of the Juan Fernández Archipelago offshore Chile
Sadly, the history of intensive human occupation and introduction of pests has transformed this territory in one of the most threatened all over the world. Only in recent times nine species extinctions have been documented (Danton and Perrier 2005, 2006), and most of the endemic species are in critical danger according to national assessment following IUCN criteria.
1.2 Overall Plant Endemism of the Archipelago
The archipelago’s vascular flora comprises around 210 native species classified in 55 families and 110 native genera (Marticorena et al. 1998; Danton and Perrier2006). Among these taxa, the islands contain 2 endemic families (Lactoridaceae and Thyrsopteridaceae), 14 endemic plant genera, and 130 endemic taxa. This number of local endemic taxa is one of the highest levels of endemism per surface area reported globally for an island territory (Stuessy et al. 1992; Vargas et al. 2011). Most endemics pertain to the angiosperms, although many fern species also occur exclusively on these islands, including several species of Hymenophyllaceae, Blechnaceae and Dryopteridaceae among other endemic ferns such as the charismatic Dicksonia species. The endemic angiosperm species pertain mostly to the Asteraceae (28 species), including 22 species from the four endemic genera Dendroseris, Robinsonia, Yunquea and Centaurodendron. The species of these four genera have a remarkably growth form described as “rosette trees” (Skottsberg 1953; Hallé et al. 2004; Figs. 6.2, 6.3, and 6.4, Photo 6.1), which is rather uncommon among the Asteraceae.

Dendroseris litoralis (Drawing from Sierra Ráfols in Muñoz Pizarro 1971)

Robinsonia gracilis (Drawing from Sierra Ráfols in Muñoz Pizarro 1971)

Yunquea tenzii (Drawing from Sierra Ráfols in Muñoz Pizarro 1971)
The overall endemism for the archipelago is not homogenous across the three islands: Robinson Crusoe (RC) Island is the one that contains most endemic species, reaching a number of 90. From these, most pertain to the Asteraceae (19 species), followed by the Hymehoplyllaceae (5 species). Conspicuous endemic species from the archipelago are restricted to this island, like Lactoris fernandeziana (Lactoridaceae), Juania australis (Arecaceae), three Eryngium species (Apiaceae), the two Centaurodendron species, the only one species from the genus Yunquea (Asteraceae), and most of the Robinsonia and Dendroseris species (Asteraceae).
Alejandro Selkirk (AS) Island contains less endemic species than Masatierra (Robinson Crusoe), but with 65 endemic species the number is considerable. They pertain mostly to the Asteraceae (6 Erigeron species, 3 Dendroseris species and 1 Robinsonia). From these endemic Asteraceae only Erigeron fernandezianus is also found in Robinson Crusoe Island. Conspicuous species not found elsewhere are the only orchid of the archipelago: Gavilea insularis (Photo 6.3), and also Peperomia skottsbergii, Haloragis masafuerana (including two varieties) and Gunnera masafuerae. Indeed, many species endemic to this island have been named with the specific epithet “masafuerana” or “masafuerae”.
The little island of Santa Clara harbours four endemic species, shared with Masatierra: Wahlenbergia berteroi, Chenopodium sanctae-clarae, Dendroseris litoralis and D. pruinata.
The islands Masatierra (RC) and Masafuera (AS) have 25 endemic taxa in common, and several of these are conspicuous and dominant elements of the vegetation, like the trees Drimys confertifolia (Winteraceae) and Coprosma pyrifolia (Rubiaceae) and the ferns Blechnum cycadifolium and Rumohra berteroana. Interestingly, several endemics show a vicariant pattern between both islands, having a closely related but different species occupying comparable habitats on each island: this is the case for Fagara mayu/F. externa; Sophora fernandeziana/S. masafuerana; Berberis corymbosa/B. masafuerana; Nothomyrcia fernandeziana/Myrceugenia schulzei. This is the case also for some ferns at species level (Dicksonia berteroana/D. externa) and at infraspecific rank: Synammia intermedia subsp. intermedia/S. intermedia subsp. masafuerana; Megalastrum inaequalifolium var. inaequalifolium/M. inaequalifolium var. glabrior. Nevertheless, the phylogenetic relationship of these vicariant species is not always as close as one could intuitively suspect, like the case of Nothomyrcia fernandeziana that seems to be more closely related to neotropical species of the genus Blepharocalyx than to Myrceugenia schulzei from Alejandro Selkirk Island (Murillo-Aldana and Ruiz 2011). Interestingly, both Myrtaceae species are the main component of the Fernandezian vegetation, to the point that a specific name for this forest has been proposed, namely the Fernandezian “myrtisylva” (Danton 2006). This vegetation unit seems to be the habitat for the highest number of endemic species, at least in Robinson Crusoe Island. This contribution seeks to analyze if this is really the case and whether there is a similar situation in Selkirk Island, which shows a higher diversity of habitats and apparently a lower level of human-induced impacts.
1.3 Material and Methods
The assessment of plant endemism for each different habitat in both main islands was based on available vegetation schemes. Both islands have an early vegetation map drawn at 1:50,000 scale (Skottsberg 1953) that has an immense historical value but a limited practical use. For the case of Robinson Crusoe, this map has been updated by Greimler et al. (2002) on the base of standard phytosociological methods. This last map, at a scale of 1:30,000, classifies the vegetation in 17 classes, including many anthropogenic units. For the purpose of the present study these classes have been reclassified in nine classes, nesting the units with minor areal importance. For the case of Alejandro Selkirk Island, a vegetation map at scale 1:12,000 based on aerial photographs and field work has been recently published by Greimler et al. (2013). Our own vegetation classification presented here on the base of field observations, puts the emphasis on the fern formations. The vegetation classification has been completed there on the base of field observations. Santa Clara Island has been omitted from the analysis since it harbours only an herbaceous carpet composed mainly of weeds. Vegetation units were superimposed to available species distribution information, by means of the software ArcGIS 9.3. A topological revision was undertaken to assure the consistency of the spatial units, avoiding overlapping and blank spaces, and granting a correct area calculation.
The assessment of plant endemism for each different habitat in both main islands was based on available vegetation schemes. Both islands have an early vegetation map drawn at 1:50,000 scale (Skottsberg 1953) that has an immense historical value but a limited practical use. For the case of Robinson Crusoe, this map has been updated by Greimler et al. (2002) on the base of standard phytosociological methods. This last map, at a scale of 1:30,000, classifies the vegetation in 17 classes, including many anthropogenic units. For the purpose of the present study these classes have been reclassified in nine classes, nesting the units with minor areal importance. For the case of Alejandro Selkirk Island, a vegetation map at scale 1:12,000 based on aerial photographs and field work has been recently published by Greimler et al. (2013). Our own vegetation classification presented here on the base of field observations, puts the emphasis on the fern formations. The vegetation classification has been completed there on the base of field observations. Santa Clara Island has been omitted from the analysis since it harbours only an herbaceous carpet composed mainly of weeds. Vegetation units were superimposed to available species distribution information, by means of the software ArcGIS 9.3. A topological revision was undertaken to assure the consistency of the spatial units, avoiding overlapping and blank spaces, and granting a correct area calculation.
1.4 Results: Endemism Related to Habitats
The species-richest habitat, also containing the highest number of endemic species, is the montane forest of Robinson Crusoe Island. Physiognomically this forest resembles the Macaronesian laurisilva as well as the Metrosideros forest of the Hawaiian Islands (Skottsberg 1953). Danton (2006) proposed a proper name for the fernandezian forests, namely the Fernandezian Myrtisylva, reflecting both dominant species from the family Myrtaceae: Nothomyrcia fernandeziana in RC and Myrceugenia schulzei in AS. This myrtisylva has been divided since Skottsberg (1953) in “upper” and “lower” according to altitude, but the composition of both units is rather similar and the border between both units is very diffuse. Nevertheless, Greimler et al. (2002) recognized indeed a limit between both units, emphasizing the endemic character of their botanical components. One of the most remarkable aspects of this myrtisylva in its upper variant is the presence of numerous so called “rosette trees”, the name Skottsberg (1953) gave to this remarkable growth-form of umbrella like little trees (Fig. 6.3).
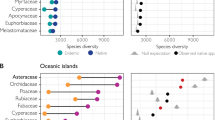
Robinson Crusoe Island contains nine main habitat types or vegetation units. Figure 6.5 shows the vegetation map, and Table 6.1 the number of endemics present in every unit. Photo 6.2 shows several aspects of the vegetation of the island.

Map of the different vegetation types of Robinson Crusoe Island
Rocks and eroded areas: these are highly eroded areas and rocky cliffs, barely vegetated, that occupy the largest area on the island, mostly at low altitude close to sea, also in Santa Clara Island. At several sites there are some plants, mostly weeds, that cover less than 10 % of the ground surface. The origin of these widespread eroded areas is a mixture of naturally exposed volcanic soil that is constantly being washed by the rain, and it also results from overexploitation of the environment by cattle and pests like rabbits. Nevertheless, some endemic species can be found in very low abundance, such as several Dendroseris and Wahlenbergia species, as well as the more abundant Ochagavia elegans, Eryngium inaccesum and Robinsonia gayana on cliffs exposed to the sea.
Herbaceous weed assemblages: this unit is widespread at low to mid elevations (50–400 m asl) on both northeastern and southwestern slopes of the island, reaching the westernmost part and Santa Clara Island. One of the dominant species is the weed Acaena argentea that forms dense carpets leaving little space for other taxa. It was inadvertently introduced from the mainland during the nineteenth century and represents today “a main act in the tragedy of the island flora” (Greimler et al. 2002: 275). Several endemics occur in this unit; always in very scarce populations are the rare Apium fernandezianum and Chenopodium crusoeanum. It is highly possible that this unit represents different levels of ecosystem degradation, e.g. in Quebrada El Lápiz in the northeast, where the last exemplar of Dendroseris neriifolia serves as an indicator for better conditions for forest development in the past.
High altitude myrtisylva (= endemic upper montane forest, sensu Greimler et al. 2002) is situated mainly above 400 m, along the main ridge that divides the island, on deep and moist soils and also on vertical escarpments with less soil development. It is dominated by the endemic trees Nothomyrcia fernadeziana, Drimys confertifolia, Fagara mayu, Coprosma pyrifolia and C. oliveri, Rhaphithamnus venustus, together with the endemic palm Juania australis and the iconic species Lactoris fernandeziana. The unit is the most difficult to access on the island, and concentrates in ca. 800 ha the highest level of endemism in the whole archipelago (69 species). Most endemic species pertaining to the rosette-tree Asteraceae are to be found in this unit, such as both Centaurodendron species, Dendroseris berteroana and D. pinnata, different Robinsonia species, and the iconic Yunquea tenzii, known only from the summit of El Yunque massive.
Low altitude myrtisylva (= endemic lower montane forest. sensu Greimler et al. 2002) is situated mainly below 400 m, on the dryer northwestern and southeastern parts of RC. The dominant tree is Nothomyrcia fernandeziana, often accompanied by Drimys confertifolia and Fagara mayu. Boehmeria excelsa is common on slopes adjacent to the bottoms of the “quebradas”. This unit is widely affected by the aggressive intrusion of alien plants; therefore, most of it has been currently classified by Greimler et al. (2002) as “endemic forest with aliens”.
Myrtisylva with aliens (endemic forest with aliens, sensu Greimler et al. 2002) is a unit that corresponds mainly to the lower altitude myrtisilva between 140 and 420 m, but is dominated by alien trees and shrubs from the species Aristotelia chilensis and Rubus ulmifolius. In spite of the high invasiveness of the mentioned species, a high number of endemics still survive under these conditions, the trees in a dramatic competition for light and space, and the herbaceous plants in very low abundances.
Ugni molinae shrubland is a unit that is dominated by the alien species Ugni molinae, a fleshy-fruit shrub that is native in the continent and has turned out to be the third worse pest in the island. It occupies mainly ridges and wind-exposed slopes between 100 and 700 m. Several endemic species that used to occupy this habitat still appear in the understory, but in low abundance, such as Gaultheria racemulosa at lower altitudes and Blechnum cycadifolium, Robinsonia gracilis and Ugni selkirkii above 400 m.
Rubus-Aristotelia shrubland is a unit that shows the extreme aspect of ecosystem degradation, located on low altitudes between 50 and 400 m. The native myrtisylva has been almost totally replaced by the alien species Aristotelia chilensis and Rubus ulmifolius. Aristotelia, which is native on the mainland, is supposed to have been taken to the archipelago in the middle of the nineteenth century, while the first report of Rubus ulmifolius as a weed was made by Looser (1927). During the second half of the twentieth century both species have turned out to be so invasive with the help of birds that disperse the fleshy fruits everywhere, that dramatic projections suggest that they could completely replace the native vegetation during the next 70 years (Dirnböck et al. 2003). Of course, the level of endemism in this unit is one of the lowest on the island.
Native grassland is a unit that comprises several patches in the lowest areas of the central part of the island between 20 and 460 m. It is dominated by tussock-forming native grasses, such as Nassella laevissima, Piptochaetium bicolor and occasionally Nassella neesiana, together with a high presence of alien herbs. This unit, would have been much more expanded towards the western side of the island under natural conditions. However, the area there is currently covered only by a weed assemblage. Several endemic grasses should have been found in this unit, such as Podophorus bromoides, but they got so rare that this last one has been classifies as “extinct”.
Town and forest plantations: in and around the town San Juan Bautista, there are cultivated areas of Eucalyptus globulus, Cupressus goveniana, and Pinus radiata that were planted around the village to help stopping the erosion once the original forest was removed. Several other introduced trees, like Acacia dealbata and A. melanoxylon, are also occur in this unit, and the risk of invasion of these newly arrived species is still unknown. Very few endemic species are found in this unit. The number of introduced species on the island increases constantly due to the artificial “enrichment” of the town’s gardens. Nevertheless, some species that are getting very rare in nature have been successfully cultivated in the gardens of the village, like Dendroseris litoralis, and many more of them in the greenhouses and gardens of the National Park’s installations.
Alejandro Selkirk Island contains 12 main habitat types or vegetation units, plus the tiny occasional settlement and Eucalyptus plantation on the entrance to Quebrada Las Casas. Figure 6.6 shows the vegetation map, and Table 6.2 the number of endemics present in every unit (cf. Photos 6.2 and 6.3).

Map of the different vegetation types of Alejandro Selkirk Island

Robinsonia gayana (Asteraceae) as representative of the ‘rosette tree’ growth form (Photographed by Sergio Elórtegui Francioli on Robinson Crusoe Island)

Representative photographs of the Juan Fernández Archipelago. (a) Santa Clara Island: grasslands and endemic species Dendroseris pruinata, (b) High altitude myrtisylva in Robinson Crusoe Island, (c) Myrtisylva in Alejandro Selkirk Island, (d) Grasslands on top of A. Selkirk Island, (e) Deep quebradas on the eastern side of A. Selkirk Island, (f) Fern scrub on top of the cliffs of the western side of A. Selkirk Island (Photographed by Sergio Elórtegui Francioli)

A photographic selection of plant species endemic to the Juan Fernández Archipelago. (a) Thyrsopteris elegans (Thyrsopteridaceae), (b) Drimys confertifolia (Winteraceae), (c) Rhaphithamnus venustus (Verbenaceae), (d) Wahlenbergia berteroi (Campanulaceae) endemic to RC and Santa Clara Islands, (e) Lactoris fernandeziana (Lactoridaceae) endemic to RC Island, (f) Plantago fernandezia (Plantaginaceae) endemic to RC Island, (g) Myrceugenia schulzei (Myrtaceae) endemic to AS Island, (h) Sophora masafuerana (Fabaceae) endemic to AS Island, (i) Gavilea insularis (Orchidaceae) endemic to AS Island (Photographed by Sergio Elórtegui Francioli)
Grassland occupies the vastest extent on the island, especially above the quebradas of the eastern side between 0 and 800 m asl. It is covered by a carpet of native and introduced grasses, such as Nassella laevissima, Aira praecox and Anthoxantum odoratum. Presence of endemic species is rather high (30), including Uncinia douglasii, Megalachne berteroana and Cardamine kruesselii.
Fern Scrub occupies one of the largest amounts of area (1,500 ha) in the upper central and northern part of the island above 800 m. Dominant characteristic species is Lophosoria quadripinnata, with the continuous presence of Gaultheria racemulosa and the introduced grass Anthoxantum odoratum. The ample presence of this grass is an indicator of the high level of fragmentation of this unit due to the huge presence of goats, estimated at around a hundred individuals. Nevertheless, the unit shows the highest amount of endemic species (38) of the island, including Thyrsopteris elegans, Gunnera masafuerae (in locally humid conditions), Berberis masafuerae, Erigeron ingae, patches of Peperomia fernandeziana and the sole island orchid Gavilea insularis in a very restricted spot called La Cuchara-Morro El Pasto (Photo 6.3).
Fern forest is located above the deep valleys of the southern portion of the island, acting as a transition towards fern scrub. It is characterized by the tree fern Dicksonia externa, and other ferns such as Blechnum cycadifolium, Blechnum chilensis, Rumohra berteroana, Pteris berteroana and several Hymenophyllum species. Other species here are Galium masafueranum, Gunnera masafuerae, few exemplars of Robinsonia masafuerae and patches of Drimys confertifolia. On the border of this unit with the cliffs of the southeast there is the only known exemplar of Dendroseris gigantea.
High altitude forest is situated mainly between 700 and 1,000 m, in the upper part of the valleys and plateau on the southern portion of the island. Dominant endemic tree species are Drimys confertifolia, Dicksonia externa, Blechnum cycadifolium and many different Hymenophyllum species, together with a rich array of mosses and Hepaticae, including Marchantia.
Pristine myrtisylva occurs mainly between 100 and 650 m, in the valleys of the western part of the island. The unit is widely dominated by the endemic tree Myrceugenia schulzei, joined by Coprosma pyrifolia, Rhaphithamnus venustus, and Fagara externa. Endemic herbaceous species are Peperomia skottsbergii and Haloragis masafuerana. It seems to be one of the least degraded units of the island due to difficult access, but the overall level of endemism is relative low as it covers only limited area. It is also possible that the richness of this unit actually is higher but it has been rarely surveyed due to the aforementioned difficulties. In spite of the fact that Santalum fernandezianum is considered to have gone extinct long ago, there is a small possibility that a living exemplar of this species might be found in this unit.
Degraded myrtisylva seem to be the degraded version of the former one, situated mainly between 100 and 650 m, in the more accessible valleys of the eastern part of the island. The level of anthropogenic impacts is high, related to overgrazing and fires, to the point that physiognomically it resembles an open forest rather than a myrtisylva. Nevertheless, some very rare endemic species such as Ranunculus caprarum have been reported here, only with few individuals.
Cliff vegetation is situated mainly on the western side of the island, between 400 and 1,000 m. It shows a high number of endemic species (23 taxa). Characteristic endemic species are Coprosma pyrifolia, Rhaphithamnus venustus, Acaena masafuerana and Galium masafueranum, together with Uncinia costata and Erigeron turricola. From the endemic Asteraceae, Dendroseris macrophylla and D. regia are to be found at very few sites. It is possible that Dendroseris gigantea, and the putatively extinct Chenopodium nesodendron could have been common in this unit once.
Coastal vegetation occupies a thin band in contact with the sea, along the coast surrounding most of the island, characterized by a scanty herbaceous vegetation of Nassella laevissima, Brassica oleracea, Juncus procerus, together with the fern Asplenium obtusatum. Herbaceous endemic species typical of the unit are Erigeron rupicola, Nicotiana cordifolia, Spergularia masafuerana and Spergularia confertiflora.
Rocks and eroded areas occupy highly degraded sites above the coastal vegetation with introduced and native grasses. In spite of this, several endemic herbaceous species can be found here, such as Haloragis masafuerana or Wahlenbergia masafuerae.
Creek bottom vegetation, occurring in the deepest part of the quebradas of the eastern side, is adapted to extreme shady conditions. It is composed by many different ferns, such as Blechnum longicauda, and several Peperomia species, together with Gunnera masafuerae, Urtica glumeruliflora, Erigeron fernandezianus, E. stuessyi, Wahlenbergia tuberosa and scanty exemplars of Sophora masafuerana. In lower parts of this unit the invasive Aristotelia is rapidly gaining space.
High altitude steppe occurs above the “treeline” (fern-line), on the highest portion of the island between 1,160 and 1,380 m, with the characteristic presence of “magellanic” species of the shrubby cushions habit, such as Abrotanella linearifolia, Lagenophora hariotii, Myrteola nummularia, and Rubus geoides. Remarkably is the precipitation of snow at this altitude some times during the winter, which does not at all occur on Robinson Crusoe Island. The level of endemics is low, but endemic species characteristic to this unit are Urtica masafuerae (just few individuals) and Agrostis masafuerana.
Weed assemblages: Like in Robinson Crusoe Island, but somehow different, the tree aliens also occur on this island, but not as widespread as in the former island. Anyhow, the arboreal weed Aristotelia chilensis forms dense stands at middle altitudes on the eastern side of the island. Rubus ulmifolius is also entering in the quebradas, but still far not as dramatic as in Robinson Crusoe Island. The most widespread weed on the coast to the north and south of the settlement at the base of Quebrada Las Casas is Bahia ambrosioides (Asteraceae). The number of endemic species is drastically reduced in this unit to Spergularia confertiflora and very sparsely Spergularia masafuerana at the entrance to the quebradas, as well as Uncinia douglassi and Megalachne berteroana at middle altitudes.
Human settlement and plantation: this unit occupies a tiny portion at the base of Quebrada Las Casas, including a Eucalyptus plantation; the village is only sporadically occupied by fisherman. The only endemic species that has been reported there is the rare Cardamine kruesselii that occurs at very low presence at the entrance of several eastern quebradas.
1.5 Discussion and Notes on Conservation
Most of the units on Robinson Crusoe Island are characterized or dominated by alien species, with the high and low altitude myrtisylva being historically and systematically reduced due to invasive species. Both units plus the native grasslands occupy currently only a 23 % of Robinson Crusoe Island. Endemic species showing viable populations are today almost exclusively confined to the most inaccessible parts of the island: the slopes and summit around the El Yunque massive and the southeastern sector known as La Piña. This is why several conservation assessments suggest that at least 75 % of the endemic flora might be highly threatened (Stuessy et al. 1998; Cuevas and van Leersum 2001; Vargas et al. 2011). And physiognomically, it is highly possible that the myrtisilva as a whole might be replaced or severe invaded by alien species by the end of this twenty-first century (Dirnböck et al. 2003). Research leading to concrete actions towards ecological restoration is only recently being developed (Vargas et al. 2010).
In the case of Alejandro Selkirk Island the distribution of endemic species is more heterogeneous than in Robinson Crusoe, and many endemic species are not to be found in the myrtisylva but in the fern assemblages and in the vegetated cliffs and deep canyons. In fact, a relative pristine myrtisylva is restricted now to the most inaccessible parts on the western side. Even though the eastern myrtisylva are much more degraded, it seems that the intrusion of alien plant species is not as intensive as on Robinson Crusoe. But no assessment has been conducted to evaluate the potential spread velocity of these aliens, so the real threat for the endemic species remains uncertain.
The early vision of Skottsberg is still totally valid: …“I wish to draw attention to the fact that so many of the endemic types are very scarce. […]”. Of species from Masatierra, Podophorus bromoides and Phrygilanthus berteroi have not been found by later collections and no exact locality is known. Greigia berteroi, Chenopodium crusoeanum, Eryngium inaccesum, E. fernandezianum, and Dendroseris macrantha were reported from one locality each: of the latter only two specimens could be found […] other very rare plants are Peperomia margaritifera, Selkirkia berteroi, Plantago fernandezia, Robinsonia thurifera and Centaurodenron. One or more of these will probably share the fate of Santalum fernandezianum, which seems to be extinct.
Skottsberg continues regarding Masafuera Island: …“also some species seem to be rare: Peperomia skottsbergii, Ranunculus caprarum and Cardamine kruesselii were found in one locality each; Chenopodium nesodendron, Robinsonia masafuerae, and Dendroseris regia are very scarce, and their recovery is checked by the too numerous wild goats. Many of the non-endemic species are also rare” (Skottsberg 1921: 212–213).
The conservation of the fernandezian flora is not just a romantic environmentalists’ ideal, but an urgent need for better scientific understanding of the biogeography and evolution processes of island floras. The geographical relationships of the archipelago’s flora show an important relationship to the American continent, as one could expect for these oceanic islands. Nevertheless, other important relationships with far territories like Australasia have intrigued biogeographers for a long time (Bernardello et al. 2006). Furthermore, the presence of many endemic genera and two endemic families suggests an old continental character of the flora, and traditional palaeogeographical reconstructions still do not fit well with this high degree of endemism. They suggest a much wider hypothetical territory that once could had better connections to Australasia or to ancient Gondwanaland (Skottsberg 1956; Moreira-Muñoz 2011). Under the current paradigm of molecular biology, oceanic islands are crucial for the calibration of phylogenies, despite the problems these calibrations might have (Heads 2011). Some species are so uncommon, that their systematic position is still problematic. In the words of Carlquist, referring to Yunquea tenzii restricted to the summit of El Yunque: “it may become extinct, as has so much of the autochthonous Juan Fernández vegetation, without having been collected in the flowering state” (Carlquist 1958: 20). Aside from botanical information, detailed ecological studies that consider variables such as soils, solar incidence, relief and micro-site characteristics are still lacking, in spite of being crucial for developing effective conservation actions (Vargas et al. 2010).
1.6 Conclusions
The main question of this section was the distribution of endemic species in relation to the different habitats present in the islands of the Juan Fernández archipelago. The answer seems to be very obvious in the case of Robinson Crusoe Island: the endemic species are today concentrated, or better said, are restricted due to human intervention, to the high altitude myrtisylva located at the most inaccessible parts of the island. Nevertheless, some species continue to exist in the low altitude myrtysilva and other human-disturbed units, but in critically low abundances. For many endemic species the population number does not exceed ten living exemplars or even just one remaining exemplar. At least nine species have gone extinct in historical times and two of them during the last decade (Danton and Perrier 2006). These species are Podophorus bromoides, Santalum fernandezianum, Chenopodium nesodendron, Empetrum rubrum, Eryngium sarcophyllum, Notanthera heterophylla, Robinsonia berteroi, R. macrocephala, and Margyracaena skottsbergii. This last species is still conserved ex-situ. The threats for the endemics in Alejandro Selkirk Island seem to be a little less dramatic; endemics still occupy the different habitats of the island, concentrated in the most inaccessible parts of the cliffs and deep valleys.
National Park rangers fight daily not only against pests and invaders, but also against the prevalent national carelessness and the local ignorance, dreaming of the day at which the big impulse for ecosystem restoration will generate again the conditions for the flourishing of this widely recognized botanical treasure. And who knows, maybe the future conditions are so favourable as to ensure that a hidden sandal-wood seed decides that it is time to show that life indeed can re-emerge when and where one does not believe it anymore.
Acknowledgements Compilation of the species data base and cartography was undertaken thanks to Fondecyt project Nº 1120448. Juan Troncoso helped with the classification units, GIS calculations and map design. Rodrigo Vargas provided vegetation information for Masatierra. The park rangers and especially the park administrator don Iván Leiva have been for years of immense help in the field. Philippe Danton and Christophe Perrier provided always valuable information. Marcia Ricci also provided valuable information about the distribution of the species, while Mélica Muñoz-Schick and Alicia Marticorena from the SGO and CONC herbaria, provided information for checking the distribution of rare species. Patricio Novoa did a critical and much helpful revision on a first draft of the manuscript.
2 Madeira Islands
2.1 Physical Geography of the Madeira Archipelago
The Madeira archipelago (30–33°N) is a group of relatively old volcanic islands. The archipelago comprises Madeira as the largest island (737 km2) and with the highest summit (Pico Ruivo, 1,861 m asl), Porto Santo (42 km2), the Desertas (15 km2), many islets, and, in political terms, the small Salvage Islands (4 km2) which lie 300 km to the SE of Madeira. The islands and islets together cover an area of approximately 800 km2.

Aeonium glandulosum is common on sea cliffs and rocks below 300 (700) m asl. in Madeira (Photographed by Miguel Sequeira)

Isoplexis sceptrum is a rare shrub (50–400 cm tall) of the Madeiran laurisiva (Photographed by Miguel Miguel Sequeira). On Madeira many individuals are planted along levadas, roadsides and in gardens

Clethra arborea is one of the major components of laurel forest on Madeira and the Azores at altitudes between 300 and 1,000 m (Photographed by Carsten Hobohm in Ribera de Janela, Madeira)
The archipelago is located in the Atlantic Ocean SW of the Iberian Peninsula and W of North Africa. The distance from Madeira to the Iberian Peninsula is approximately 1,000 km and to Africa 600 km. The distance from the Selvage Islands to Africa is 300 km and to the Canary Islands 180 km.
The geological age differs from island to island and from one literature source to another. According to Press and Short (1994) the islands are 60–70 million years old, according to Jardim and Francisco (2000) the Madeira Islands are 18 million years old, whereas the Selvage Islands were formed about 27 million years ago (for details of Madeira’s geology cf. Burton and MacDonald 2008).
However, in contrast to the Canaries or Azores the islands have known no recent or historic volcanism. The terrain has suffered erosion for a very long time, is generally rugged and dissected by deep ravines. According to Mucina and Wardell-Johnson (2011) the landscapes of the Madeira Islands belong to the old stable landscapes of the world (OSLs) because of their old geological surface, relatively stable climate and the high predictability of the (non-) fire regime.
The climate is typical Mediterranean, with annual average temperatures between 6–10 °C at the summits and 15–20 °C on the southern coasts. Differences in precipitation and summer drought are the result of variation in altitude and geomorphology. High altitudes on Madeira Island are relatively wet (>1,600 mm/a), whereas Porto Santo and Desertas are relatively dry (<400 mm/a). The Salvage Islands can be prone to long periods of drought.
Press and Short (1994) counted 123 endemic vascular plant taxa (out of 1,226 indigenous and naturalized taxa), Borges et al. (2008) listed 154 species and subspecies (out of a total of 1,204 taxa). According to our own list (EvaplantE, latest version) the number is 134 (excluding endemic hybrids and varieties). Including the infraspecific categories varieties and natural hybrids, Jardim and Francisco (2000) counted 165 endemics; in their book they list 143 endemics – species, several subspecies and a few hybrids. However, some 120 species or 170 taxa including species, subspecies and hybrids, or 10–12 % of the flora are endemic to the Madeira archipelago including the Salvage Islands.
Six endemic genera occur on Madeira Islands (de Sequeira et al. 2007).
2.2 Analysis of Floras and Databases
Important information about the endemics of the Madeira archipelago in EvaplantE (latest updated version) were extracted from Borges et al. (2008), de Sequeira et al. (2007), Jardim and Francisco (2000), Franquinho and Da Costa (1996), Press and Short (1994), Hansen and Sunding (1993), and Vieira (1992).
EvaplantE (latest updated version) and the literature cited above form the basis for the following analysis. During several excursions to Madeira and Porto Santo the second author had the opportunity to study the vegetation and to identify many of the endemic vascular plants in different landscapes and localities of the archipelago. Furthermore, we explored one of the most beautiful and impressive valleys in the world, the Janela valley, from the sea up to the highest parts. Many observations of species compositions in relation to their habitat types were noted and entered into the data base.
The importance of a specific habitat for endemics can be measured as the number of taxa which occur in this habitat – independent of the fact that most taxa occur in more than one habitat type.
2.3 Endemism Related to Landscapes and Habitats
The largest proportion of endemics on the Madeira archipelago occurs in rocky habitats and screes (82 taxa). The second largest group comprises forest endemics (41); these are followed by inhabitants of coastal and saline habitats (34), scrub and heath (21), habitats connected with running waters or waterfalls (12), and arable land, ruderal and settlement habitats (10). 4 Taxa (also) occur in grassland or grassy pastures, and 1 (also) in moorland (Vaccinium padifolium).
Examples of endemic inhabitants of rock and scree habitats are Aeonium glutinosum, Agrostis obtusissima, Aichryson divaricatum and dumosum, Armeria maderensis, Asplenium trichomanes ssp. maderense, Berberis maderensis, Bunium brevifolium, Bystropogon punctatus, Ceterach lolegnamense, Chamaemeles coriacea which inhabits both coastal and inland cliffs, Cheirolophus massonianus, Crepis vesicaria ssp. andryaloides, Deschampsia maderensis, Erysimum arbuscula and maderense, Euphorbia desfoliata, Geranium maderense and palmatum, Helichrysum melaleucum and monizii, Hymenophyllum maderense, Jasminium azoricum, Lotus argyrodes, Luzula elegans, Melanoselinum decipiens, Monanthes lowei, Monizia edulis, Musschia aurea and wollastonii, Parafestuca albida, Rumex bucephalophorus ssp. frutescens, Saxifraga maderensis and portosanctana, Sideritis candicans, Sinapidendron frutescens, Sonchus pinnatus, Tolpis macrorhiza and Viola paradoxa.
The following endemic taxa (also) occur in laurisilva, other forest types or woodland: Arachniodes webbianum, Bystropogon maderensis, Carduus squarrosus (also occurs on coastal cliffs), Carex lowei and malato-belizii, Cirsium latifolium, Convolvulus massonii which also occurs on low-altitude coastal rocks, Dactylorhiza foliosa, Deschampsia argentea on rocky slopes in damp woodland, Dryopteris aitoniana and maderensis, Echium candicans, Festuca donax, Goodyera macrophylla, Hedera maderensis ssp. maderensis, Ilex perado ssp. perado, Isoplexis sceptrum, Luzula seubertii, Normania triphylla, Polystichum drepanum, Rosa mandonii, Rubus grandifolius and vahlii, Ruscus streptophyllus, Sambucus lanceolata also occurs outside the forests and woodlands in thickets, just as Sideroxylon mirmulans, Sonchus fruticosus, Teline maderensis, Teucrium abutiloides and betonicum, Vaccinium padifolium, and Vicia capreolata. Vicia capreolata is found on Porto Santo and Desertas, also at low altitudes and independent of forest.
Coastal habitats such as coastal cliffs, maritime sands or organic layers on stony beaches are inhabited by e.g. Andryala crithmifolia, Argyranthemum haematomma and thalassofilum, Betula patula, Calendula maderensis, Chamaemeles coriacea, Crambe fruticosa (also some kilometers inland), Echium nervosum (also near roads), Helichrysum devium and obconicum, Limonium ovalifolium ssp. pyramidatum, Lobularia canariensis ssp. pyramidatum and ssp. rosula-venti, Lotus loweanus and macranthus which also occur in dry stony pastures or cornfields and other open and rocky places, respectively, Matthiola maderensis, Maytenus umbellata, Olea maderensis, Senecio incrassatus, Sinapidendron angustifolium which also grows a short distance inland, Sonchus ustulatus, and Urtica portosanctana.
Examples of endemics of heaths, thickets and scrub habitats are Bystropogon maderensis (also in laurisilva), Erica maderensis and scoparia ssp. maderinicola, Genista tenera, Lavandula stoechas ssp. maderensis, Marcetella maderensis, Sorbus maderensis, and again Vaccinium padifolium.
Some endemics are inhabitants of arable land, ruderal or urban habitats. Examples are Crepis noronhaea on roadsides, cultivated and waste ground, and Delphinium maderense in cornfields and on roadside banks.
Only few water plants occur on Madeira, and none of them are endemic to the archipelago. Several endemic taxa such as Carex lowei, Ceterach lolegnamense, Dryopteris aitoniana, Dryopteris maderensis, Geranium palmatum, Geranium rubescens, Melanoselinum decipiens, Oenanthe divaricata, Scrophularia hirta, Scrophularia racemosa and Solanum patens occur under wet and shady conditions in habitats which are connected to water ecosystems, e.g. river banks, dry river beds, or along levadas and beside streams. Some also occur in shady and wet forest, independent of the proximity to water bodies.
The average of the elevation minima is c. 300 m asl. and the maxima average at 1,000 m asl. Therefore, we assume that most endemics on Madeira occur between 300 and 1,000 m.
The high endemism of the archipelago as a whole can be explained by the age of the geological surface and the absence of historical or recent volcanism, by the rich environmental heterogeneity, isolation and – compared with the mainland of Europe or western North Africa – a relatively stable climate history. According to Mucina and Wardell-Johnson (2011) Madeira has old stable landscapes. Fossil records show that in the past laurisilva was part of the Tertiary flora, as it was in mainland Europe where this forest type was destroyed by climate change during the Pleistocene (see e.g. Kunkel 1993).
In general, differences in the endemism of different habitat types can be explained by the same arguments but smaller in time and space, e.g. the age of the current ecological conditions of a locality or habitat type. Ecological conditions, including meso-climate of coastal cliffs, of laurisilva, scrub and rocky habitats of the Madeira Islands, are much older than urban habitats, cornfields or roadsides. The area covered by a certain habitat type might also be relevant for the number of evolutionary processes and endemics. However, natural grassland and swamps are rare in Madeira’s landscapes. The grassland area was extended by human activities such as cutting, burning and grazing by domestic animals. This grassland mostly is species-poor, and endemics of the island are seldom to be found here (Photos 6.7 and 6.8).

Helichrysum obconicum, endemic to the island of Madeira, inhabits coastal rocks and sea cliffs (Photographed by Carsten Hobohm on basaltic rock near Porto Moniz, Madeira)

Plantago arborescens ssp. maderensis, endemic to the Madeira archipelago, is common throughout Porto Santo and grows among rocks, on banks, on roadsides and in other open areas below 1,000 m (Photographed by Carsten Hobohm near the northwestern coast of Porto Santo)
3 Corsica, Mediterranean
3.1 Physical Geography of Corsica
The Mediterranean island of Corsica lies south of the mainland of France between latitudes 41 and 43°N. The highest mountains on Corsica consist predominantly of granites, gabbros and volcanic rocks which date back 280–250 Ma. At that time, the mountain ranges of Corsica, Sardinia and the Balearic Islands were part of the Iberian subplate west of the localities where they are now. Some 35 Ma ago, these mountain ranges began to move to the east. When sea water from the Atlantic Ocean reclaimed the Mediterranean Basin some 5 Ma ago the area around Corsica was flooded and Corsica became an island. Before that time, Corsica was part of the mainland and already covered by vegetation. However, since its birth the island has experienced different warm and cold climate periods. During the Pleistocene glaciation periods the highest mountain tops and shady slopes were most likely permanently covered with snow and ice. Climate change scenarios suggest that, as a consequence of increasing and decreasing temperatures during Pleistocene cycles, plant populations had to move up- and downwards, and a loss of species diversity might have been the result, not only in the northern parts of Europe but also in Mediterranean regions (Gillespie and Clague 2009; Gottfried et al. 2002).
The Mediterranean Island of Corsica (8,750 km2) is situated between the coasts of mainland France to the North (160 km), Italy to the East (82 km) and the island of Sardinia to the South (12 km). More than 20 mountains reach a height of at least 2,000 m. The highest points are Monte Cintu (2,706 m), Monte Rotondo (2,622 m), and Monte Padro (2,393 m). Corsica is known as the most mountainous island in the Mediterranean Basin (Gillespie and Clague 2009; Gamisans and Marzocchi 1996).
Coastal regions are relatively dry (450–600 mm/a) in comparison with high mountain zones (>1,800 mm/a). Gamisans and Marzocchi (1996) distinguish three climate zones: a coastal zone of warm Mediterranean climate between the coastline and 600 m, with winter rain, summer drought and annual temperatures between 14 and 17 °C (1); a zone of cool Mediterranean climate between 600 and 1,200 m, with annual temperatures between 13 and 10 °C and precipitation rates of 800–1,500 mm/a (2); and a zone of relatively cold climate at elevations over 1,200 m, with high precipitation rates (>1,500 mm/a) and snowfall during winter (3).
A major part of the island is composed of crystalline rocks (e.g. granites, rhyolites), schists, ophiolites, serpentines and sedimentary limestone. Typical habitats comprise marine sands, dunes, saltmarshes and rocky shores at the coast. Large parts of the lowlands are cleared for agriculture. However, typical Mediterranean evergreen sclerophylleous forest, mixed forests and maquis remain in many parts of the island. The mountains are home to diverse mixed and broadleaf deciduous forests which are dominated by pine and oak species. The submontane zone is dominated by shrubs, the alpine zone by grassland, heaths with cushion plants, rocks and screes.
3.2 Analysis of Floras and Databases
The analysis of the composition of endemic vascular plants on the island of Corsica is based on Gamisans and Marzocchi (1996), Tutin et al. (1996a–e, 1964–1990), Bouchard (1968), EvaplantE (latest updated version which is based on diverse floras; cf. Bruchmann 2011; Hobohm and Bruchmann 2009) and Hobohm (2000).
According to the references cited it was possible to characterise most endemics in relation to habitat types and altitudes. In EvaplantE we use a relatively wide and conservative species concept in accordance with Tutin et al. (1996a–e). It is, therefore, possible to compare numbers of endemics from different regions on the basis of a consistent species concept.
3.3 Endemism Related to Landscapes and Habitats
According to Gamisans and Marzocchi (1996) c. 2,978 vascular plant taxa – 2,092 species, 264 subspecies, 89 varieties, 82 hybrids – are indigenous or naturalised on the island of Corsica. 131 taxa are endemic to the island (4.4 %). 165 taxa are subendemics. These are restricted to Corsica and Sardinia (75), Corsica together with Sardinia and the Balearic Islands (11), Corsica together with small neighbouring mainland regions, or together with mainland regions and neighbouring islands (86). Only 48 island-endemic taxa are listed for Corsica in EvaplantE. The discrepancy between these figures and those of Gamisans and Marzocchi (1996) has to do with the breadth of the species concept. In Gamisans and Marzocchi (1996: 28 ff.), for example, eight species of the genus Limonium which are endemic to the island are named and shown in photographs. They belong to the species groups of Limonium articulatum, acutifolium and densiflorum. None of these groups is endemic to Corsica. Bouchard (1968) distinguished six species of the genus Limonium in total. According to Tutin et al. (~1996c) one is endemic to Corsica (and therefore listed in EvaplantE).
However, certain tendencies can be described independent of the interpretation of the species concept. Most endemics occur at altitudes between 1,000 and 2,200 m (average minimum altitude to average maximum altitude).
The highest proportion of endemics (c. 67 %) is associated with rocks and screes. The second largest group is found in grasslands (46 %); this is a relatively high proportion in comparison to other Mediterranean islands, and compared to the Canaries or Madeira (cf. Bruchmann 2011). A third (35 %) of the endemics are found in scrub and heaths, 19 % in freshwater habitats and wetlands, 10 % in forests, and only a few taxa occur in coastal habitats, mires or arable land. The sum of the percentages exceeds 100 % because the majority of the endemics are found in several habitat types. A few endemic taxa are rather habitat-specific, such as Armeria leucocephala, Bupleurum falcatum ssp. corsicum and Phyteuma serratum in rocky habitats or Alnus viridis ssp. suaveolens which dominates in subalpine scrub communities.
A comparison between the oceanic archipelago of Madeira and the continental island Corsica shows remarkable differences in the numbers of endemics, distribution patterns and habitat characteristics. Both regions are very mountainous and mainly influenced by a Mediterranean climate with winter rain and summer drought, at least at lower altitudes.
Madeira is much smaller than Corsica and richer in endemic vascular plants. This is most likely the result of a longer island history and a more continuous climate during the Pleistocene – stabilised by marine currents (cf. Mucina and Wardell-Johnson 2011). This interpretation is underlined by the fact that the ancient forest type laurisilva, which in the Tertiary also occurred in southern Europe, survived on Madeira, and a high percentage of endemics are found in these forest habitats.
A larger distance to the next landmass or mainland region clearly favours isolation. The Madeira archipelago is more isolated than Corsica. Genetic isolation favours speciation and speciation favours endemism. On the other hand, geographical separation successfully reduces dispersal events, and Madeira is poorer in vascular plant species overall; Corsica has more than twice the total number of vascular plant taxa in comparison with Madeira (cf. Borges et al. 2008; Gamisans and Marzocchi 1996), and a large species pool also favours speciation processes. However, the higher concentration of endemics on Madeira cannot be fully explained by the separation/isolation factor.
As in most regions of Europe, the highest proportion of endemics is found on rocks and screes. Grassland endemics constitute the second largest group on Corsica, whereas on Madeira the forest taxa are the second largest group. The high proportion of endemics associated with grasslands is astonishing because natural grassland on Corsica is rare, and most grassland habitats are the result of fertilisation, cutting, moving and/or grazing. No island endemic which occurs in grassland is found exclusively in this habitat type. Many of these taxa also occur in heathlands, scrub or rocky habitats. Scrub and heaths harbour a large percentage of the endemics on both islands, despite the fact that fire is a normal phenomenon on Corsica where scrub vegetation is a succession stage between fire and forest, whereas on the Madeira archipelago fire is fairly uncommon (favoured by plantations of pine and Eucalyptus globulus).
4 Madagascar
4.1 Physical Geography of Madagascar
Madagascar is the fourth largest island in the world (585,000–587,000 km2; Table 5.4), situated >400 km off the SE coast of Africa between latitudes 12 and 25.5°S. With rocks dating back 3,200 million years and an origin as continental island some 160 million years ago it is most likely the oldest island in the world. Most landscapes are flattened, leached, and old in the sense of Mucina and Wardell-Johnson (2011). The highest mountain is Tsaratanana (2,876 m asl).
In most parts of the island the climate is tropical warm or hot with summer rain. Madagascar is influenced by mostly dry trade-wind conditions in winter and wet monsoon-driven tropical winds in summer. Only a few regions are influenced by a more subtropical (S Madagascar) or orogenic climate (central mountains). Eastern regions with humid forests receive precipitation throughout the year, without a distinct dry period, but also with higher precipitation in summer than in winter. Precipitation rates can be more than 6,500 mm/a on the northeastern coasts, whereas the driest parts in the SW receive less than 400 mm/a. Mean air temperatures range from 26–29 °C in the North to 20–27 °C in the South. Temperatures in the high mountain regions average between 19 and 12 °C, and can drop below zero in winter (minimum: −12 °C; Wells in Goodman and Benstead 2003).
Madagascar is part of the “Madagascar and the Indian Ocean Islands Biodiversity Hotspot” (Mittermeier et al. 1999, 2005). The outstanding plant and animal richness of Madagascar can be explained by the age of the geological surface (i), by the fact that Madagascar is of continental origin (ii) and by long and uniterrupted evolutionary histories (iii). The number of vascular plant species is estimated at 10,000–14,000 including c. 80–90 % endemics (Cribb and Hermans 2009; Gillespie and Clague 2009; Burga and Zanola 2007). A project called Catalogue of the Vascular Plants of Madagascar aims to enumerate and evaluate the native and naturalised flora of the island. Based on this list, Callmander et al. (2011) recently registered 11,220 species and 1,626 infraspecific taxa in total, including 8,884 endemic and 2,336 non-endemic species. The level of endemism of the indigenous vascular flora is calculated as 82 %, or 79 % of all vascular plants including 331 naturalised species.
Today, Madagascar is burning almost always and everywhere because of charcoal production, because of the tradition of burning pastures and arable land to produce nutrients from the ash and – in the case of pastures – give space for the young grass shoots which are eaten by zebus, and because charcoal and wood are used for cooking and heating.
The conversion of primary vegetation and forest to arable and pastural landscapes under the pressure of an overwhelming population still continues.
4.2 Analysis
Basic facts and numbers come from Callmander et al. (2011), Gillespie and Clague (2009), Gribb and Hermans (2009), Burga and Zanola (2007), Moat and Smith (2007), Goodman and Benstead (2003), Schatz (2001), Koechlin et al. (1974). However, the massive explosion of knowledge about the natural history of Madagascar and the description of plants and animals which are new to science at the beginning of the third millenium indicates a high potential for further investigations.
The author undertook several excursions to Madagascar with a couple of hypotheses in his baggage and with the plan to see and understand the natural units – landscapes, habitats, ecological conditions, biodiversity – just a little better.
4.3 Endemism Related to Landscapes and Habitats
The flora of Madagascar is predominantly a woody flora. Despite the recent controversy surrounding the nature of the vegetation when humans first arrived, and the origin of the vast treeless areas on the Central High Plateu, the fact remains that even the driest and edaphically harshest areas in the southwest support woody vegetation. Moreover, the treeless expanses are especially depauperate in herbs, often consisting of only one or two species of grass widespread throughout Africa. With the exception of orchids - the majority of which are epiphytes on the woody vegetation - the Malagasy flora is relatively poor in herbaceous groups. Thus, trees provide the basic structure for most of the island’s flora and fauna, as well as fuel wood and housing for the overwhelming majority of Malagasy people (Schatz 2001: 1).
Tree species, large and small shrubs and other woody species together form the majority of the flora on Madagascar. Species endemism for trees and shrubs together reaches a rate of 92 %, whereas the level for herbs is 72 % (Callmander et al. 2011). Before people inhabited the island, most of the regions of Madagascar were covered by forest, woodland and dry thickets.
Nowadays, the forest of Madagascar forms a near-coastal ring and includes a few inland forests which are concentrated in montane areas. Different processes and environmental circumstances have been discussed to explain this phenomenon.
Madagascar has a peripheral ring of lowland to montane forest around an upland that is covered by fire-simplified grassland and savanna. These largely degraded and depauperate open landscapes are dominated by a few cosmopolitan fire-adapted grasses.
The remaining primary vegetation, species richness and endemism are concentrated in near-coastal rather than inland regions.
Such a concentric order of richness can be found in various regions of the world (cf. e.g. Foster et al. 2012; Mittermeier et al. 2005). Furthermore, a peripheral forest ring can also be found in other parts of the world, e.g. Australia and Borneo.
In Australia, all species-rich regions are situated near the coast, whereas the central inland regions are dominated by species-poorer woodlands, thickets, grasslands and semi-desert. In the case of Australia, the low species diversity of the central parts can be explained by alternating dry and wet climate periods during the Pleistocene and Holocene. These cycles affected inland regions more than coastal regions. Unfortunately, and in contrast to Australia, the Cenozoic climate history and the development of the Malagasy biomes during this period remain largely unknown (cf. Wells in Goodman and Benstead 2003; Beard et al. 2000).
Raes et al. (2009) discussed and analysed distribution patterns of endemic vascular plants and centres of endemism on Borneo and compared these patterns with findings in other regions in the world and other groups of organisms. For Borneo they concluded that regional endemism hotspots represent near-coastal areas which are characterised by a small range in annual temperature and high environmental heterogeneity and which are least affected by El Niño drought events. Relatively stable climate conditions maintaining endemism have been found for various groups of organisms in different regions of the world (Jetz et al. 2004; Gathorne-Hardy et al. 2002, cf. also Araujo et al. 2008; ter Steege et al. 2003)
However, there are remarkable differences between the distribution patterns of Madagascar and e.g. Australia or Borneo. Whereas the species-poorer inner areas of Australia receive less than 250 mm precipitation per year, almost all parts of Madagascar have precipitation rates which doubtless would enable forest growth (>>800 mm/a, e.g. Antananarivo c. 1,300 mm, Fianarantsoa >1,200 mm). Furthermore, the driest parts of the island in the South, with precipitation rates <500 mm/a, are very rich in plants and endemics (dry spiny bush and forest). In contrast to Borneo, the near-coastal regions of Madagascar represent very different climates, from a wet tropical climate in the East (with precipitation rates >2,000 mm per year) to subarid in the South (<400 mm).
It is therefore curious that in Madagascar, under totally different climate conditions, there should be a peripheral ring of forest, rich biodiversity and endemism around a largely species- and endemic-poor inland area with grassland and savanna. If this peripheral ring reflected high environmental heterogeneity and steep mountain slopes, then the biodiversity would easily be explained by habitat diversity. However, maps show that this is only the case in a few parts of the island. In many parts forest occurs in lowland areas with obviously low environmental heterogeneity. It is possible that the higher endemism in coastal areas could be explained by unfavourable inland conditions rather than by a uniform marine-influenced coastal climate.
We assume that the risk of lightning fires corresponds with the length of the dry season. Therefore, the probability of natural fire increases from rainforest areas in the N and E to dry spiny bush in the SW. This view is underlined by the distribution patterns of fire-resistant plants which are concentrated in the western and southern parts of the island. Some plants have notably thick barks that allows them to survive fires (e.g. Uapaca bojeri, Bismarckia nobilis, Hyphaene coriacea, Poupartia birrea ssp. caffra, and several species of Sarcolaenaceae), others are able to resprout from their woody base or tuberlike roots after fire (Labat and Moat 2003; Schatz 2001; Rauh 1973). Furthermore, many of the succulent plants withstand fire simply because they do not burn well.
Because natural fire is a more or less normal event in the dry spiny bush in the S of Madagascar many of the related vegetation units are well (pre)adapted to fire. On the other hand, anthropogenic fires are less effective in terms of fire spread in areas without, or with only short, dry periods, as is the case in the northern and eastern parts with humid forest and high precipitation rates. In the central parts of Madagascar, the vegetation is not well fire-adapted on the one hand but on the other hand it burns easily in dry periods in wintertime. This might be a reason for the poorness of the inland grasslands and savannas, which in comparison with high mountainous and coastal or near-coastal regions are strongly influenced by man and fire.
Binggeli (in Goodman and Benstead 2003) discussed the impact of several neophytes on native ecosystems in Madagascar. Many exotic and invasive plants have been introduced in recent centuries. Alien species are omnipresent in almost all habitats and environments. Nonetheless, it is not easy to calculate the direct influence and to estimate the interrelationship between increasing neophyte populations and decreasing populations of native or endemic taxa. In many cases, human alteration of environmental conditions can promote both effects. Thus, it remains uncertain which neophytes directly threaten native or endemic vascular plants.
The impact of neophytes in several parts of the primary forest is astonishingly low. We saw untouched rainforest with a few individuals of e.g. Clidemia hirta, Psidium guajava or other introduced plant species – only along the footpaths. And we saw dry spiny bush and dry forest without any associated neophytes.
Three species that either have or have had an important impact on natural vegetation and humans alike (Binggeli 2003: 260) are Eichhornia crassipes, Lantana camara and Opuntia monacantha. However, these species do not occur everywhere either. Eichornia inhabits eutrophic, slow-running or standing waters, and many waterbodies in Madagascar are not covered by this plant. Lantana camara is named as widespread and an important pest in agricultural lands, secondary shrublands and succession stages after clearings. This can be seen in many cultivated areas and along the roads. Nonetheless, many other shrublands do not harbour a single individual of Lantana camara. The appearance of Opuntia, especially in dry regions, is extremely complex and its occurrence is scattered; as it is not infected by cochineal everywhere, it is widely used as resource and fodder, planted as an impenetrable hedge and so on (Binggeli 2003). Furthermore, this plant does not occur in every dry habitat.
However, the extent to which neophytes affect endemic vascular plant taxa remains unclear, whereas there is no doubt as to the constant threat created by the elimination and transformation of habitats by man.
The dominant vegetation types at the beginning of our millenium (Moat and Smith 2007: 19 ff.) are plateau grassland – wooded grassland mosaic (41.7 %) and wooded grassland-bushland (22.9 %), which together cover approximately two thirds of the island (65 %), and degraded humid forest (9.8 %) and primary and secondary humid forest (8.1 %) which together amount to c. 18 %. Western dry forest covers 5.4 %, south western dry spiny forest thicket 3.1 %, western subhumid forest 0.7 %, mangroves 0.4 %, tapia forest 0.2 %, littoral forest and western humid forest are the smallest forest types which together cover less than 1 % of the island’s area. All forest types together make up c. 26 %. Approximately 4 % of the whole island is cultivated, 1 % is bare rock or soil, 1 % is wetlands.
Assuming that most parts of the island used to be covered by forest, the reduction in forest area can be estimated at c. 60–70 % of the whole area or more than two thirds of the island’s forest. The majority of the remaining forest is highly impacted by various human activities.
One third of Madagascar’s primary forest is estimated to have been lost since the 1970s. Less than 10 % of Madagascar’s area is covered by natural vegetation and most of the open landscapes are grazed by zebu cattle and other livestock.
The dry spiny forest-thickets in SW Madagascar appear to be the vegetation type with the highest level of endemism (95 %; Wells in Goodman and Benstead 2003, Gautier & Goodman in Goodman and Benstead 2003). Endemism is also extremely high in primary forest (89 %), very high in rocky habitats (82 %), intermediate in swamps (56 %), lower in aquatic, semi-aquatic (38 %) and coastal habitats (21 %), and very low in savanna, grassland (6 %) and urban habitats (c. 0 %). However, information about absolute numbers of endemic taxa related to different vegetation types or information about species ranges is still scarce. As mentioned above, the majority of the endemics clearly inhabit forest. Because of the distribution patterns and area of the humid forest and because this is indicated by the floras (e.g. Schatz 2001; Dransfield et al. 2006; Cribb and Hermans 2009, and others) we assume that this vegetation unit harbours most of the endemic vascular plant taxa found in Madagascar.
According to Ranarijaona (in Goodman and Benstead 2003: 251) c. 128 aquatic and semiaquatic (of a total of 338 species and varieties, or 38 %) vascular plant taxa are endemic to Madagascar. This number is relatively low compared to that for terrestrial plants (81–82 % or 8,000–10,000; see Callmander et al. 2011; Gautier and Goodman 2003). Ferry et al. (1999, cited in Gautier and Goodman 2003) suggest that the low number of endemic aquatic plants on Madagascar may be a result of Quaternary climate fluctuations: during dry periods freshwater and semi-aquatic habitats were essentially dry, eliminating local floras which depend on wet conditions. However, the small total area of water bodies and the fact that lakes and running waters are normally relatively young and discrete habitats are not the best conditions for promoting endemism.
One of eight mangrove tree species (Ceriops boiviniana) is considered to be endemic or possibly a synonym of Ceriops tagal (Roger & Andrianasolo in Goodman and Benstead 2003: 209).
However, the absolute numbers and percentages of endemics in coastal habitats, freshwater habitats, wetlands, grasslands and savannas are relatively low in comparison with rocky habitats, dry thickets and different forest types.
Plant families on Madagascar rich in genera and/or species, are e.g. Orchidaceae, Rubiaceae, Fabaceae, Euphorbiaceae, Sapindaceae, Malvaceae, Arecaceae.
Ferns and orchids represent two very species-rich plant groups on the island and comprise c. 1,400–1,500 species (at least 862 species of orchids, 563 species of pteridophytes). Endemism in orchids is slightly lower than 90 % and in ferns c. 45–47 %.
Both groups are extremely species- and endemic-rich compared to every other region in continental Africa of a similar size. These groups are rich in epiphytes, and show a diversity gradient with increasing numbers from dry to wet conditions (cf. Cribb and Hermans 2009, Rakotondrainibe 2003 in Goodman and Benstead 2003). This fact confirms the significance of humid forest for the survival of very many endemic vascular plants on Madagascar.
The same diversity gradient from dry to wet and SW to NE can be found in several other systematic groups, e.g. palms. On a global scale and in comparison with the whole of continental Africa, Madagascar is extremely rich in palms (Arecaceae). Of the 170 or more species (continental Africa: approximately 60) all but 5 are endemic and most of them occur in the more humid eastern and Sambirano regions (Dransfield et al. 2006, Dransfield & Beentje in Goodman and Benstead 2003).
Other plant families and genera, such as Didieraceae (4 endemic genera, 11 endemic species) and Aloe (c. 120 endemic species, more than 50 of which were described after 1990), radiated in relatively dry environments (Castillon and Castillon 2010; Petignat and Cooke 2009; Schatz 2001).
Madagascar is a little larger than France, for example, and has more than twice the number of plant species. France is a mainland region with temperate to subtropical (Mediterranean) and orogenic (Alps, Pyrenees) climates, and was strongly affected by Pleistocene glaciation cycles. Madagascar is an island with winter-dry or wet tropical climates which was less impacted by Pleistocene glaciation cycles. We assume that the differences in the number of vascular plant taxa can be better explained by differences in the evolutionary histories than by differences in warmth or other aspects of contemporary climates (cf. Sandel et al. 2011; Normand et al. 2011). Nor can the differences be explained by environmental heterogeneity or habitat diversity.
5 Comparison of Endemism and Habitat Affinities on Juan Fernández Islands, Madeira Archipelago, Tenerife and Corsica
The Juan Fernández archipelago (Chile), the Madeira archipelago (Portugal), and Tenerife (Canary Islands, Spain) are volcanic islands that originated in the ocean. Corsica (Mediterranean, France) is a continental island that became an island when the Mediterranean Basin was flooded. All islands are older than 1 million years, in most cases they originated a few or many million years ago (cf. Gillespie and Clague 2009, for the following comparison see also the literature cited above); the Juan Fernández Islands originated 1–4 Ma, Madeira Archipelago 18–70 Ma, Tenerife 7–50 Ma, and Corsica 5 Ma ago. According to the geological data, the Juan Fernández islands are younger than Corsica and Tenerife whereas the Madeira islands are the oldest. However, the exact numbers differ depending on the references (Table 6.3).
Juan Fernández (33°S), the Madeira islands (30–33°N) and Tenerife (28–29°N) lie c. 670, 600 or 275 km west of continents, respectively. Corsica (41–43°N) lies very close to the European mainland of France to the North (160 km), Italy to the East (58 km), and to the large continental island Sardinia to the South (12 km). The latter was also a mountain range before it became an island. Tenerife occupies a central position in the Canary Islands, and is c. 30–100 km from La Gomera, La Palma, El Hierro and Gran Canaria and 200–250 km from Fuerteventura and Lanzarote. The distance to the African mainland is approximately 275 km.
All islands are influenced by a Mediterranean climate with winter rain and summer drought. Mean temperatures from summits to warm coastal regions are c. 10–16 °C on Juan Fernández Islands, 6–20 °C on the Madeira archipelago, 9–21 °C on Tenerife, and 5–17 °C on Corsica. All islands can experience snow and frost on mountain tops in winter, irregularly on Juan Fernández Islands and Madeira, regularly on Tenerife and Corsica.
Precipitation rates differ from year to year but also from one to another locality on Juan Fernández Islands (c. 300–1,700 mm) with almost daily rain at high altitudes. Desertas, Porto Santo and the South of Madeira (Madeira archipelago) are relatively dry (<400 mm), whereas high mountain regions of Madeira Island receive more than 1,600 mm rain per year. Tenerife is very dry on southern coasts (100–200 mm), dry at high elevations (300–400 mm) and relatively wet at mid-altitudinal zones on northern slopes (c. 800 mm; Pott et al. 2003; Kämmer 1982). On Corsica the values range from 450 mm in the driest parts to >1,800 mm in alpine regions (Gamisans and Marzocchi 1996). Thus, compared to the other islands Tenerife is relatively dry (cf. minima, maxima, averages).
Juan Fernández Islands have extremely high levels of endemism, c. 62 % on average. Furthermore, the islands harbour 14 endemic vascular plant genera and even two endemic plant families. The Madeira archipelago harbours 10–12 % endemics (including 6 endemic genera). According to our own counts based on Izquierdo et al. (2004) Tenerife has 8.4 % endemics, including two genera. The rate for Corsica is 4.4 % (no endemic genus). The rate of endemism itself is quite a good indicator for isolation. The lower rates of endemism on Tenerife and Corsica can easily be explained by the nearby mainland or island landmasses. Both islands are much less isolated compared to the other two archipelagos. As the much higher level of endemism of the Juan Fernández Islands shows, the degree of isolation of this archipelago is high compared to Madeira Archipelago. This is not easily explicable, as both archipelagos are located west of and at a similar distance to the continent and are not surrounded by other islands or archipelagos.
However, the absolute number of endemic vascular plant taxa on Juan Fernández Islands (104 km2), the Madeira archipelago (800 km2), Tenerife (2,034 km2) and Corsica (8,750 km2) is similar (c. 123–154 species and subspecies). Thus, the density of endemics is higher on Juan Fernández Islands than on Madeira Islands or Tenerife and much higher in comparison to Corsica.
We assume that these differences have to do with differences in ecological stability over long evolutionary periods. Climate and geological surface stability are important factors in explaining the differences; however, we can only speculate on the details.
Climate, in general, is buffered and stabilised by marine water bodies, especially by strong marine currents. As the oceanic archipelagos lie on the periphery of a circular current system the situation of the Juan Fernández, Madeira, and Canary Islands seems to be comparable. One clear difference, however, is the existence of a circumpolar current (Antarctic Circumpolar Current or West Wind Drift) in the southern hemisphere, which has no counterpart in the northern hemisphere. This strong current system which moves the cold southern oceans from West to East is today the largest ocean current and the major driver of exchange of water between the oceans. Estimates of the onset of this current range from 6 to 41 Ma (Barker et al. 2007). The discussion about the influence of such currents on climate or, the other way round, about the influence of climate change on the existence and stability of currents is still in progress (Hofmann and Rahmstorf 2009; Levermann et al. 2007; Zickfeld et al. 2007). However, it is possible that in the past the climate of Juan Fernández Islands was more stable than that of the Madeira archipelago and Canary Islands (Santelices 1992). Corsica, on the other hand, was more impacted by glacial cycles than the other archipelagos because of its position close to the European landmass.
There are also differences connected with the stability of the geological surface: for example, recent volcanism – i.e. during the past few centuries – is known from Tenerife but not from the other islands.
The habitat preferences of the endemics on these islands also differ remarkably. On Juan Fernández Islands, endemics are concentrated in montane forests (myrtisylva) and fern-dominated units.
In the Madeira archipelago, on Tenerife and Corsica most endemics occur in rocky habitats and screes. In most regions of Europe the highest proportion of endemics occur in this group of habitats.
Similar to the Azores, the second large group on the islands of Madeira comprises forest endemics. This fact is unusual compared to most regions of continental Europe or compared to Mediterranean islands and to the neighbouring Canaries (cf. Bruchmann 2011). The laurel forest of Madeira, the Canaries and Azores survived during glaciation cycles, whereas this forest type disappeared on the European mainland. Furthermore, in contrast to the Mediterranean, fire is very uncommon and mostly a man-made event on Madeira; also, the forest still covers large parts of the island. In contrast to the Canary Islands there has been no recent volcanism, and the main and maximum precipitation rate is much higher.
The second large group on the Canary Islands comprises endemics of shrub- and heathlands (Bruchmann 2011). This is more or less typical for Mediterranean regions. We assume that this relationship is also typical for the island Tenerife alone, as this is the largest island of the archipelago. Furthermore, Tenerife is richest in terms of different habitat types and species. Thus, we assume that the values for the whole archipelago are transferable to Tenerife, even if we do not have an analysis of the habitat affinities of Tenerife itself.
The second largest group of endemics on Corsica is found in grasslands. This is also unusual compared to other Mediterranean islands. For example, the second largest group on Sardinia, Sicily and the Balearic Islands consists of endemics of coastal and saline habitats. The second largest group following the endemics of rocks and screes on Crete and Cyprus comprises endemics with preference for scrub and heath habitats. The relatively large group of grassland endemics on Corsica might be explained by the large proportion of acidic granites in the central mountain regions which, additionally, receive relatively high rainfall. Also, summer rainfall in the high mountain regions of Corsica is a normal event. These climatic and edaphic conditions together promote the existence of grassland in alpine regions and might be advantageous over e.g. dwarf shrub or thorn cushion plant communities.
References
Araujo MB, Nogues-Bravo D, Diniz-Filho JAF, Haywood AM, Valdes PJ, Rahbek C (2008) Quaternary climate changes explain diversity among reptiles and amphibians. Ecography 31:8–15
Barker PF, Filppelli GM, Florindo F, Martin EE, Scher HD (2007) Onset and role of the Antarctic circumpolar current. Deep-Sea Res II 54:2388–2398
Beard JS, Chapman AR, Gioia P (2000) Species richness and endemism in the Western Australian flora. J Biogeogr 27:1257–1268
Bernardello G, Anderson GJ, Stuessy TF, Crawford DJ (2006) The angiosperm flora of the Archipelago Juan Fernández (Chile): origin and dispersal. Can J Bot 84:1266–1281
Binggeli I (2003) Introduced and invasive plants. In: Goodman SM, Benstedad JP (eds) The natural history of Madagascar. The University of Chicago Press, Chicago/London, pp 257–268
Borges PAV, Abreu C, Aguiar AMF, Carvalho P, Jardim R, Melo I, Oliveira P, Sergio C, Serrano ARM, Vieira P (eds) (2008) A list of the terrestrial fungi, flora and fauna of Madeira and Selvagens archipelagos. Direccao Regional do Ambiente da Madeira and Universidade dos Acores, Funchal und Angro do Heroismo, 440pp
Bouchard J (1968) Flore pratique de La Corse, 2nd edn. Société des Sciences Historiques et Naturelles de la Corse, Bastia
Bruchmann I (2011) Plant endemism in Europe: spatial distribution and habitat affinities of endemic veascular plants. Dissertation, University of Flensburg. Flensburg. URL: www.zhb-flensburg.de/dissert/bruchmann
Burga C, Zanola S (2007) Madagaskar – Hot Spot der Biodiversität. Exkursionsbericht und Landeskunde, Schriftenreihe Physische Geographie Bodenkunde und Biogeographie 55. Geographisches Institut der Universität, Zürich, pp 1–201
Burton CJ, MacDonald JG (2008) A field guide to the geology of Madeira. Geological Society of Glasgow, Galsgow
Callmander MW, Phillipson PB, Schatz GE, Andriambololonera S, Rabarimanarivo M, Rakotonirina N, Raharimampionona J, Chatelain C, Gautier L, Lowry PP II (2011) The endemic and non-endemic vascular flora of Madagascar updated. Plant Ecol Evol 144(2):121–125
Carlquist S (1958) Anatomy and systematic position of Centaurodendron and Yunquea (Compositae). Brittonia 10(2):78–93
Castillon B, Castillon J-P (2010) Les Aloe de Madagascar. Published by the Authors, Reunion
Cribb P, Hermans J (2009) Field guide to the orchids of Madagascar. Kew Publishing, Kew
Cuevas JG, van Leersum G (2001) Proyecto conservación, restauración y desarrollo de las islas Juan Fernández, Chile. Rev Chil Hist Nat 74:899–910
Danton P (2006) La “myrtisylve” de l’archipel Juan Fernández (Chili), une forêt en voie de disparition rapide. Acta Bot Gallica 153(2):179–199
Danton P, Perrier C (2005) Notes sur la disparition d’une espèce emblématique: Robinsonia berteroi (DC.) Sanders, Stuessy & Martic. (Asteraceae), dans l’île Robinson Crusoe, archipel Juan Fernández (Chili). Le Journal de Botanique de la Société Botanique de France 31:3–8
Danton P, Perrier C (2006) Nouveau catalogue de la flore vasculaire de l’archipel Juan Fernández (Chili). Acta Bot Gallica 153:399–587
de Sequeira M, Jardim R, Silva M, Carvalho L (2007) Musschia isambertoi M. Seq., R. Jardim, M. Silva & L. Carvalho (Campanulaceae), a new species from the Madeira archipelago (Portugal). Anales del Jardim Botanico de Madeira 64(2):135–146
Dirnböck T, Greimler J, Lopez P, Stuessy TF (2003) Predicting future threats to the native vegetation of Robinson Crusoe Island, Juan Fernandez Archipelago, Chile. Cons Biol 17:1650–1659
Dransfield J, Beentje H, Britt A, Ranarivelo T, Razafitsalama J (2006) Field guide to the palms of Madagascar. Kew Publishing, Kew
Foster MN, Brooks TM, Cuttelod A, De Silva N, Fishpool LDC, Radford EA, Woodley S (eds) (2012) Key biodiversity area special series. J Threat Taxa 4(8):2733–2844
Franquinho LO, Da Costa A (1996) Madeira: Plantas e flores. Francisco Ribeiro & Filhos, Funchal
Gamisans J, Marzocchi J-F (1996) La flore endémique de la Corse. Edisud, Aix-en-Provence
Gathorne-Hardy FJ, Syaukani, Davies RG, Eggleton P, Jones DT (2002) Quaternary rainforest refugia in south-east Asia: using termites (Isoptera) as indicators. Biol J Linn Soc 75(4):453–466
Gautier L, Goodman SM (2003) Introduction to the flora of Madagascar. In: Goodman SM, Benstedad JP (eds) The natural history of Madagascar. The University of Chicago Press, Chicago/London, pp 229–250
Gillespie RG, Clague DA (eds) (2009) Encyclopedia of islands. University Press of California, Berkeley
Goodman SM, Benstead JP (eds) (2003) The natural history of Madagascar. The University of Chicago Press, Chicago/London
Gottfried M, Pauli H, Reiter K, Grabherr G (2002) Potential effects of climate change on alpine and nival plants in the Alps. In: Körner C, Spehn EM (eds) Mountain biodiversity: a global assessment. Parthenion Publishing, Boca Raton et al, pp 213–223
Greimler J, López P, Stuessy TF, Dirnböck T (2002) The vegetation of Robinson Crusoe Island (Isla Masatierra), Juan Fernández archipelago, Chile. Pac Sci 56:263–284
Greimler J, López-Sepúlveda P, Reiter K, Baeza C, Peñailillo P, Ruiz E, Novoa P, Gatica A, Stuessy T (2013) Vegetation of Alejandro Selkirk Island (Isla Masafuera), Juan Fernández Archipelago. Chile Pac Sci 67(2):267–282
Gribb P, Hermans J (2009) Field guide to the orchids of Madagascar. Kew Publishing, Kew
Hallé F, Danton P, Perrier C (2004) Architectures de plantes de l’Ile Robinson Crusoe, Archipel Juan Fernández, Chili. In: Vester H, Romeijn P, van der Wal H (eds) 25 Jaar een “Boom der vrijheid” – Liber Amicorum, Prof Dr Ir R.A.A Oldeman, Treebooks 8. Treemail Publishers, Heelsum, pp 43–55
Hansen A, Sunding P (1993) Flora of Macaronesia. Checklist of vascular plants, Sommerfeltia 17. Botanical Garden and Museum, University of Oslo, Oslo, pp 1–295
Heads M (2011) Using island age to estimate the age of island-endemic clades and calibrate molecular clocks. Syst Biol 60:204–218
Hobohm C (2000) Biodiversität. Quelle & Meyer, Wiebelsheim
Hobohm C, Bruchmann I (2009) Endemische Gefäßpflanzen und ihre Habitate in Europa – Plädoyer für den Schutz der Grasland-Ökosysteme. RTG-Berichte 21:142–161
Hofmann M, Rahmstorf S (2009) On the stability of the Atlantic meridional overturning circulation. PNAS. www.pnas.org_cgi_doi_10.1073_pnas.0909146106. Downloaded 21 Sept 2012
Izquierdo I, Martín JL, Zurita N, Arechavaleta M (eds) (2004) Lista de especies silvestres de Canarias (hongos, plantas y animales terrestres) 2004. Consejeria de Medio Ambiente y Ordención Territorial. Gobierno de Canarias, Tenerife
Jardim R, Francisco D (2000) Flora Endémica da Madeira. Corlito/Setubal, Funchal
Jetz W, Rahbek C, Colwell RK (2004) The coincidence of rarity and richness as the potential signature of history in centres of endemism. Ecol Lett 7:1180–1191
Johow F (1896) Estudios sobre la flora de las Islas de Juan Fernández. Imprenta Cervantes, Santiago
Kämmer F (1982) Beiträge zu einer kritischen Interpretation der rezenten and fossilen Gefässpflanzenflora und Wirbeltierfauna der Azoren, des Madeira-Archipels, der Ilhas Selvagens, der Kanarischen Inseln und der Kapverdischen Inseln, mit einem Ausblick auf Probleme des Artenschwundes in Makaronesien. Polycopy, Freiburg i. Br, 179pp
Koechlin J, Guillaumet J-L, Morat P (1974 [1997]) Flore et végétation de Madagascar. Reprint. A.R.G. Gantner Verlag, Vaduz
Kunkel G (1993) Die Kanarischen Inseln und ihre Pflanzenwelt, 3rd edn. Gustav Fischer, Stuttgart
Labat JN, Moat J (2003) Leguminosae (Fabaceae). In: Goodman SM, Benstead JP (eds) The natural history of Madagascar. The University of Chicago Press, Chicago/London, pp 346–373
Levermann A, Mignot J, Nawrath S, Rahmstorf S (2007) The role of Northern sea ice cover for the weakening of the thermohaline circulation under global warming. J Clim 20:4160–4171
Looser G (1927) La zarzamora (Rubus ulmifolius) en Juan Fernández. Rev Chil Hist Nat 31:84–85, Santiago
Marticorena C, Stuessy TF, Baeza CM (1998) Catalogue of the vascular flora of the Robinson Crusoe or Juan Fernández Islands, Chile. Gayana Bot 55:187–211
Mittermeier RA, Myers N, Mittermeier CG, Gil PR (1999) Hotspots: earth’s biologically richest and most endangered terrestrial ecoregions. Cemex, Mexico City
Mittermeier RA, Gil PR, Hoffman M, Pilgrim J, Brooks T, Mittermeier CG, Lamoreux J, da Fonseconda GAB (2005) Hotspots revisited: earth’s biologically richest and most endangered terrestrial ecoregions. Cemex, Mexico City
Moat J, Smith P (eds) (2007) Atlas of the vegetation of Madagascar. Kew Publishing, Kew
Moreira-Muñoz A (2011) Plant geography of Chile, vol 5, Plant and vegetation. Springer, Dordrecht
Mucina L, Wardell-Johnson GW (2011) Landscape age and soil fertility, climate stability, and fire regime predictability: beyond the OCBIL framework. Plant Soil 341:1–23. doi:10.1007/s11104-011-0734-x
Muñoz Pizarro C (1969) El Archipiélago de Juan Fernández y la conservación de sus recursos naturales renovables. Museo Nacional de Historia Natural (Santiago) Serie Educativa 9:17–47
Muñoz Pizarro C (1971) Chile: Plantas en Extinción. Editorial Universitaria, Santiago
Murillo-Aldana J, Ruiz E (2011) Revalidación de Nothomyrcia (Myrtaceae), un género endémico del Archipiélago de Juan Fernández. Gayana Bot 68(2):129–134
Normand S, Ricklefs RE, Skov F, Bladt J, Tackenberg O, Svenning J-C (2011) Postglacial migration supplements climate in determining plant species ranges in Europe. Proc R Soc B 278:3644–3653
Petignat A, Cooke B (2009) Guide des plantes succulentes du Sud-Ouest de Madagascar. Arboretum d`Antsokay, Toliara
Pott R, Hüppe J, Wildpret de la Torre W (2003) Die Kanarischen Inseln: Natur- und Kulturlandschaften. Ulmer, Stuttgart
Press JR, Short MJ (1994) Flora of Madeira. The Natural History Museum, London
Raes N, Roos MC, Slik JWF, van Loon E, ter Steege H (2009) Botanical richness and endemicity patterns of Borneo derived from species distribution models. Ecography 32(1):180–192
Rauh W (1973) Über die Zonierung und Differenzierung der Vegetation Madagaskars, Tropische und Subtropische Pflanzenwelt 1. Steiner, Wiesbaden, pp 1–146
Sandel B, Arge L, Dalsgaard B, Davies RG, Gaston KJ, Sutherland WJ, Svenning J-C (2011) The influence of late quaternary climate-change velocity on species endemism. Science 334:660–664
Santelices B (1992) Marine phytogeography of the Juan Fernández archipelago: a new assessment. Pac Sci 46(4):438–452
Schatz GE (2001) Generic tree flora. Royal Botanic Gardens/Missouri, Botanical Garden, Kew Louis
Skottsberg C (1921) The phanerogams of the Juan Fernández Islands. In: Skottsberg C (ed) The natural history of the Juan Fernandez and Easter Islands, vol 2 (Botany). Almqvist & Wiksell, Uppsala, pp 95–241
Skottsberg C (1953) The vegetation of the Juan Fernández Islands. In: Skottsberg C (ed) The natural history of the Juan Fernández and Easter Islands, vol 2 (Botany). Almqvist & Wiksell, Uppsala, pp 793–960
Skottsberg C (1956) Derivation of the flora and fauna of Juan Fernández and Easter Island. In: Skottsberg C (ed) The natural history of the Juan Fernández and Easter Islands, vol 1. Almqvist & Wiksell, Uppsala, pp 193–405
Stuessy TF, Grau J, Zizka G (1992) Diversidad de plantas en las Islas Robinson Crusoe. In: Grau J, Ziska G (eds) Flora silvestre de Chile, Palmengarten Sonderheft 19, Frankfurt am Main, pp 54–66
Stuessy TF, Swenson U, Marticorena C, Matthei O, Crawford DJ (1998) Loss of plant diversity and extinction on Robinson Crusoe Islands, Chile. In: Peng CI, Lowry PP (eds) Rare, threatened and endangered floras of Asia and the Pacific Rim, Academia Sinica monographs 16. Institute of Botany, Academia Sinica, Taipei, pp 243–257
ter Steege H, Pitman N, Sabatier D, Castellanos H, van der Hout P, Daly DC, Silveira M, Phillips O, Vsquez R, van Andel T et al (2003) A spatial model of tree alpha-diversity and tree density for the Amazon. Biodivers Conserv 12:2255–2277
Tutin TG, Burges NA, Chater AO, Edmondson JR, Heywood VH, Moore DM, Valentine DH, Walters SM, Webb DA (1996a) Flora Europaea, vol 1: Psilotaceae-Platanaceae, 2nd edn (Reprint, first published 1993). Cambridge University Press, Cambridge
Tutin TG, Heywood VH, Burges NA, Moore DM, Valentine DH, Walters SM, Webb DA (1996b) Flora Europaea, vol 2: Rosaceae-Umbelliferae (Reprint, first published 1968). Cambridge University Press, Cambridge
Tutin TG, Heywood VH, Burges NA, Valentine DH, Walters SM, Webb DA (1996c) Flora Europaea, vol 3: Diapensiaceae-Myoporaceae (Reprint, first published 1968). Cambridge University Press, Cambridge
Tutin TG, Heywood VH, Burges NA, Valentine DH, Walters SM, Webb DA (1996d) Flora Europaea, vol 4: Plantaginaceae-Compositae (and Rubiaceae) (Reprint, first published 1976). Cambridge University Press, Cambridge
Tutin TG, Heywood VH, Burges NA, Valentine DH, Walters SM, Webb DA (1996e) Flora Europaea, vol 5: Alismataceae-Orchidaceae (Reprint, first published 1980). Cambridge University Press, Cambridge
Vargas R, Cuevas JG, Le-Quesne C, Reif A, Bannister J (2010) Spatial distribution and regeneration strategies of the main forest species on Robinson Crusoe Island. Rev Chil Hist Nat 83:349–363
Vargas R, Reif A, Faúndez MJ (2011) The forest of the Robinson Crusoe Island, Chile: an endemism hotspot in danger. Bosque 32(2):61–70
Vieira R (1992) Flora da Madeira: o interesse das plantas endémicas Macaronesicas, Coleccao Natureza e Paisagem 11. Serviço Nacional de Parques, Lisboa, 155pp
Zickfeld K, Lvermann A, Morgan MG, Kuhlbrodt T, Rahmstorf S, Keith DW (2007) Expert judgement on the response of the Atlantic meridional overturning circulation to climate change. Clim Chang 82:235–265
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Moreira-Muñoz, A., Francioli, S.E., Hobohm, C., da Silva Menezes de Sequeira, M.P. (2014). Endemism on Islands – Case Studies. In: Hobohm, C. (eds) Endemism in Vascular Plants. Plant and Vegetation, vol 9. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-6913-7_6
Download citation
DOI: https://doi.org/10.1007/978-94-007-6913-7_6
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-6912-0
Online ISBN: 978-94-007-6913-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)