
Abstract
The study of the relationships between agriculture and biodiversity is important to sustain biodiversity for the future. The landscape level has an influence, which has been until now mainly related to the importance of semi-natural elements. But in agricultural areas crop land is often dominant and acts on biodiversity by the resources it provides and the effects of disturbances induced by agricultural practices. The mosaic of crops is ephemeral and highly dynamic in space and time according to farming practices and crop rotations. The aim of this chapter is to assess the role of agricultural landscape heterogeneity on biodiversity. Landscape heterogeneity may be measured from different perspectives, considering non-cropped areas versus crop ones, or taking into consideration the dynamics of the mosaic of crops and agricultural practices. From studies on a long term ecological research site in Brittany, France, we present how these different approaches of landscape heterogeneity allow a better understanding of the diversity of processes driving biodiversity in agricultural landscapes. Most of all we underline the necessity to include knowledge of farming systems and farming practices in the analyses.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
Agriculture and biodiversity are two faces of the same coin. On the one hand, biodiversity is the basis of agriculture: plants that are grown, animals that are raised are species shaped over centuries for the benefit of people; they all depend on other living organisms. On the other hand, the fate of biodiversity in many landscapes of the world depends on its use and management by rural/farming communities. Some agricultural landscapes may be more diverse than “natural landscapes” and thus are a source of biodiversity, but many farming practices at field and landscape levels are a threat for biodiversity.
Between the 1960s and the 1990s the negative impacts of agriculture on biodiversity have become increasingly documented. Among the main drivers of biodiversity decline are agricultural practices and changes in landscape structure (Robinson and Sutherland 2002). For a long period, public policies neglected the problem but then, under pressure from environmentalists and the general public, regulations were established to protect rare or emblematic species. Nature reserves were created and were extended to Natura 2000 zones in the European Union as an ecological network of protected areas, designated to protect habitats and species present on red lists in negotiations between NGOs and policy makers. Even though farming techniques such as haying or grazing were used as integrative management techniques for those protected areas, the general concept remained to segregate the agricultural, productive areas from the nature protection areas (Fisher 2008). The growing insight regarding the benefits of ecosystem services led to an interest in species providing those services and to the idea that biodiversity should be managed and protected everywhere because it is of use everywhere. Nowadays the maintenance of beneficial insects and birds, pollination, water purification etc. are services that farmers must be aware of in their crop production and management of land.
The objective of this chapter is to tackle these different issues. We utilize results from the different projects carried out on a Long Term Ecological Research site, the “Zone Atelier Armorique”. Landscape ecology is the conceptual framework we use. That is to say we consider landscape patterns, their heterogeneity and connectivity, as major drivers of plant and animal population dynamics. Heterogeneity and connectivity are key concepts for biodiversity conservation and management (Burel and Baudry 2003) that need to be defined as specific metrics for the different questions and biodiversity groups we studied.
Landscape heterogeneity has many expressions. In the binary segmentation between semi-natural and cropping areas, heterogeneity increases if the share of the two components approaches 50 % of the area. The heterogeneity of the cultivated mosaic is also an important expression. This mosaic can be highly heterogeneous in space and time, as a result of the diversity of agricultural practices, and their spatial and temporal organizations by farmers (Vasseur et al. in press). The diversity of agricultural practices (cultivated species and varieties, rotations, technical operations) that can be observed at the landscape level, is due to farmers’ decisions (Joannon et al. 2008). Agricultural practices in a field follow crop management sequences and depend on cropping systems (pluriannual crop rotation and management). Cropping systems are furthermore spatially distributed on the farm territory according to environmental conditions in fields (e.g. soil type), spatial structure of field patterns on farms and logistic constraints (Thenail and Baudry 2004). As a result, the cropping systems mosaic is highly heterogeneous in space and time. This additional heterogeneity may be of great importance for species (insects, weeds, mammals) using crops for at least part of their life (Vasseur et al. in press).
Studies on the influence of landscape patterns on biodiversity have focused on spatial heterogeneity. On the contrary, temporal heterogeneity has been less studied in landscape ecology (Metzger 2008). Not considering this dimension is a limitation. Past landscape structure can affect present ecological processes, and there is often a time lag between landscape change and responses by organisms (Ernoult et al. 2006; Krauss et al. 2010; Auffret and Cousins 2011). Moreover, in dynamic landscapes the rate of habitat turnover and associated change in landscape structure can sometimes be more important for species survival than the spatial organization of resource patches (Fahrig 1992).
Along this gradient of heterogeneities, the respective role of landscape design (field size, shape, presence of hedgerows etc.) and of cropping practices in the management of biodiversity is a central question. This question is of importance to model population dynamics and to design management plans. A landscape mosaic is built from both design and practices; therefore it is more and more important to foster our capacity to disentangle their effects.
In this chapter, we present how different approaches of landscape heterogeneity, first oriented toward semi natural habitats, and then recognizing the role of crops and their dynamics, may give insights on the fate of biodiversity in agricultural landscapes. We will then discuss their relative efficiency according to landscape and species types. This will give clues for designing agri-environmental schemes for biodiversity.
2 The Role of Semi-Natural Elements in Agricultural Landscapes
When looking at the effect of landscape structure on biodiversity in agricultural landscapes the main emphasis has been put on considering the effect of semi natural elements (Billeter et al. 2008; Tscharntke et al. 2005a). They are considered as habitats, refuges, sources, corridors for many species that use crop fields for part of their life cycle and by species which are restricted to them (Deckers et al. 2005; Forman and Baudry 1984). This approach has been the first to be used by landscape ecologists who considered agricultural landscapes as sets of semi-natural elements embedded in a neutral agricultural matrix. We studied the effects of landscape structure, which is defined in this study by the proportion of semi-natural elements, on biodiversity measured with several taxa differing by their way of dispersal and their spatial scale of perception.
It soon appeared that biodiversity responses to this heterogeneity could be linked to farming systems, as it was proved that farming practices also played a major role in the decline of biodiversity. To assess this at the landscape level we compared landscape units, first with similar agricultural systems and contrasted landscape structures, and second, of similar landscape structure and contrasted farming systems.
2.1 Comparing Landscapes with Contrasted Landscape Structures
In Brittany, France, as in most places of north western Europe, agricultural landscapes changed dramatically in the 1960s and up to 1980s due to the rapid intensification of agriculture (Robinson and Sutherland 2002). This led to an important decrease in semi natural habitats (Meeus 1993). To assess the effects of these changes on biodiversity we compared the gamma diversity of landscapes that differed by their amount of semi natural elements and where agricultural systems were similar.
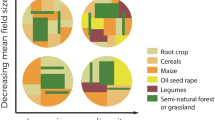
The study area, the Long Term Ecological Research (LTER) site “zone atelier Armorique”, is located in northern Brittany, south of the Mont Saint Michel Bay, France (48° 36′ N, 1° 32′ W). A hedgerow network (bocage) characterizes the landscape and agriculture is oriented toward milk production. Three units differing by field size, the density of the hedgerow network, and the relative abundance of grassland versus cropland have been delineated (Fig. 14.1). We used global indices such as percentage cover of woodlots, grasslands, crops, hedgerows, heterogeneity (Baudry and Burel 1982) to verify that units are different and have some kind of internal homogeneity (Table 14.1).

Parts of the three landscape units: the quantity of semi natural areas: woodlots, hedgerows and permanent grassland, decreases from (a) to (b) and (c) as stated in Table 14.1
We surveyed several groups of organisms which perceive the landscape at different spatial scales, and have different ways of dispersal and different life spans. These were two families of diptera: Chironomidae, and Empididae, carabidae, herbs, breeding passerines, woody plants and small mammals. All these groups have been sampled using specific sampling methods. Birds were counted according to “IPA” method (Blondel et al. 1970), small mammals were studied by using the pellets of the Barn owl (Tyto alba), carabids were caught with interception traps, diptera with yellow attractive traps and plants were identified in hedgerows, with 3014 relevés for woody plants and 455 relevés for herbs.
The results are presented in Table 14.2. They show that the different groups react differently to the changes in landscape structure. Diptera only decrease as semi natural areas decrease, there are few changes but not consistent with the landscape structure gradient for carabids breeding passerines and herbs, and no or almost no differences for woody plants and small mammals. When looking at community similarity between units (a) and (c) Burel et al. (1998) showed that three classes of taxonomic groups could be identified. First, communities of diptera Empididae and Chironomidae lose species from unit (a) to unit (c). For Empididae, species with small wings and a low power of dispersal are not present in landscapes where the distance between water courses and the closest hedgerows, two elements needed to accomplish their life cycle, is high as in unit (c) (Morvan, N. unpublished data). Second, communities of carabidae and herbs do not vary that much in species richness but there is a shift in species composition, some are only present in the site with a high proportion of semi natural areas, while others are only present where this proportion is low. Large apterous carabid forest species characterize unit (a) with a high quantity of semi natural areas, while smaller winged species, adapted to disturbances characterize unit (c) with a high proportion of crops (Aviron et al. 2005). Third, communities of breeding passerines and small mammals have almost the same species in all the units. For small mammals Millan-Pena et al. (2003) showed that if the species were the same their relative abundance varied. Forest species such as the bank vole (Clethrionomys glareolus) are relatively more abundant in dense hedgerow network landscapes, while crop species such as the Field vole (Microtus agrestis) are more abundant in sparse ones.
In these bocage landscapes, differing by the proportion of semi natural areas the response of biodiversity varies according to the different groups. For some of them, the high proportion of semi natural areas leads to an increase in species richness which has been shown in other studies for several taxa (Weibull et al. 2000; Schweiger et al. 2005). But according to dispersal ability and longevity of species not all of them react that way. This may be due to the fact that the gradient of landscape structure is not very long, the total proportion of semi natural areas varying only from 25 to 45 % of the total area, contrary to other studies where it may vary from 10 up to 80 % (Gabriel et al. 2005). Our results emphasize that species richness per se is not always a good indicator for measuring a community’s response to landscape changes. Similarity indices or relative abundance permit to identify changes in biodiversity even when species richness remains the same. Those indices underline that different groups react differently to the same changes in landscape structure. Nevertheless, for all of them the proportion of semi natural elements had an effect on the structure of the communities.
2.2 Comparing Landscapes with Similar Compositions and Contrasted Farming Systems
To assess the role of farming systems at the landscape level we compared biodiversity among landscape units of similar landscape structure but contrasted farming systems.
We studied landscapes located in the Côtes d’Armor, an administrative unit located in the northern part of the Brittany region. Its area is 700,000 ha, 440,000 of which are devoted to agriculture. It is a very dynamic agricultural area specialized in both milk production and hogs and poultry indoor production. Crops sustain mainly husbandry with maize and grassland for cows, and cereals for hogs and poultry (http://draaf.bretagne.agriculture.gouv.fr/Les-Cotes-d-Armor,203). We first selected 11 landscape units which represented the whole diversity of the landscape structures present in the area. They differ by their composition, total length of hedgerows, connectivity of the hedgerow network and heterogeneity of the mosaic. Carabid beetles have been surveyed in order to measure their gamma diversity in hedgerows and we tested for the influence of landscape structure and farming systems on it (Millan-Pena et al. 2003). We then compared sites dominated by cropland, which we split into two groups. They were both characterized by a low proportion of semi natural areas, were similar in landscape composition but differed in landscape configuration. The first one was characterized by large fields, with maize as the dominant crop and a low connectivity of the hedgerow network, while the second was characterized by smaller fields, with wheat and oat as dominant crops and a more connected hedgerow network. Ten hedgerows were sampled per site during the summer of 2001. We compared carabid species composition between the two “cropland” groups. They were characterized by two distinct clusters of species, maize-dominated landscapes hosted species occurring in rather moist and shaded habitats such as Brachinus scolopeta, while cereal-dominated landscapes hosted typical crop field species such as Pterostichus melanarius.
The abundance of carabids significantly differed between the two types of landscapes (t = 5,82, p = 0.01). The average abundance was 693 individuals per site in the cereal-dominated landscapes, and 333 for the maize-dominated ones. Species assemblages of the most different sites were compared using the ten most abundant species found at each site. These top ten species accounted for 81.6–87.8 % of the total catch, depending on the site. The top ten species present in the maize-dominated landscapes only accounted for 13.7 % in the cereal-dominated ones, reflecting the strong effect of this shift in farming system and change in landscape configuration (Table 14.3).
These results show that farming systems have an effect on biodiversity at the landscape scale. The effects of farming systems on biodiversity have mainly been studied for comparisons between conventional and organic systems and looking at the effect of the surrounding landscape on alpha diversity (Weibull et al. 2000; Purtauf et al. 2005). The main results are that diversity is higher in complex landscapes with a high proportion of semi natural areas whatever the system, and that organic systems enhance diversity in simple landscapes. Our results deal with gamma diversity and systems that are both conventional but with different crop and husbandry productions. They are intensive agricultural systems, but the one dominated by maize and milk production is less favorable for carabid species than the one dominated by cereal crops. This may be due to the differences in farming practices, with higher inputs in maize fields, or to the configuration of the landscape with larger fields and a lower connectivity of the hedgerow network when maize is dominant.
3 The Role of the Cropping System Mosaic
We have shown that semi-natural elements contribute to produce landscape structures of ecological importance. But, because many species use crops (including grasslands) during their life cycle, the heterogeneity of the cropping systems mosaic is potentially important from an ecological point of view (Kennedy and Storer 2000; Benton 2003). At a given time, this mosaic can be viewed as a spatially heterogeneous mosaic of cropped habitats with varying resources for species (food resources, host plants, shelter), and of disturbances with direct effects on species survival (e.g. insecticide spraying). The cropping system mosaic also generates a spatiotemporal heterogeneity at different time scales (Burel and Baudry 2005). Crop phenology and farming practices lead to fast asynchronous variations of resources from field to field within a year; over several years, crop rotation and management succession result in spatio-temporal variations of resources availability, localization and accessibility, i.e. landscape connectivity for species. In this shifting mosaic, habitat patches are ephemeral regarding the life span of many species. Species survival will therefore depend on their ability to find and colonize new suitable resource patches to supplement or complement habitats and complete their life cycle (Dunning et al. 1992; Wissinger 1997) as well as availability of ephemeral, but suitable habitats over years.
In the following sections, we illustrate how the cropping systems mosaic can influence the movement of organisms, their population dynamics and species diversity at the community level, at infra- and/or- pluriannual time scales. We utilize results from empirical and modeling studies conducted on the LTER “Zone Atelier Armorique” for several insect taxa: two species with contrasted habitat requirements and dispersal abilities, i.e. a grassland butterfly and a carabid beetle of cropped habitats, and the community of wild bees. We will emphasize the temporal dimension by stating the effects of the changing crop mosaic within a year studying insect movements between crops, and looking at the spatial and temporal distribution of organisms during one rotation cycle.
3.1 Effects of Crop Phenology and Farming Practices at the Infra-Annual Time Scale
Species movements between habitat patches depend not only on functional landscape connectivity (Kindlmann and Burel 2008), but also on the dynamics of resource quality in patches (Schooley and Branch 2011). For insects using annual crops or grasslands, whether they are phytophageous, nectariferous or predatory, crop cover states control the availability of biotic and abiotic resources (e.g. Alston et al. 1991). These cover states vary throughout the season from crop sowing and growth to harvest. These changes induce quick and frequent changes in insect movements and distributions in the cultivated mosaic (Kennedy and Storer 2000).
We observed these processes for the Meadow brown (Maniola jurtina L.), a grassland butterfly species, in response to grassland mowing (Aviron et al. 2007). This common species has no strong host-plants requirements (Vane Wright and Ackery 1981) but has a limited mobility like many endangered butterfly species (Brakefield 1982). We conducted a mark-release-recapture experiment at various herbaceous patches (grasslands, lane banks and road verges) to study butterfly movements and distribution before and after the mowing of two studied grasslands. This survey showed that the mowing of grasslands can lead to changes in butterfly movements between herbaceous patches. This is illustrated by the decreased exchange rates of butterflies between one of the mown grasslands (G4) and surrounding patches (Fig. 14.2a). Mowing also resulted in localized drops of butterfly abundances in mown grasslands (G2 and G4), and a concentration of butterflies in certain unmown, accessible grasslands (G3 and G8; Fig. 14.2b). These localized changes in butterfly abundances in mown and unmown grasslands can probably be explained by a redistribution of butterflies from disturbed grasslands into remnant suitable herbaceous patches.

Exchange rates of butterflies between studied herbaceous patches (a) and total number of butterflies captured in patches (b) before and after grassland mowing (adapted from Aviron et al. 2007)
To compensate for these local and abrupt changes in resource availability on a given patch, the presence of alternative suitable and accessible resource patches will be crucial for species to realize their life cycle (Kennedy and Storer 2000; Men et al. 2004; Carrière et al. 2006; Bressan et al. 2010). Some asynchrony between farming practices and crop cover states might allow to compensate for the ephemeral suitability of crops, by ensuring a temporal continuity of resources for species. A study of a generalist predatory carabid species with limited mobility (Pterostichus melanarius Illiger) in annual crops illustrates these processes (Vasseur 2012). Carabid movements were surveyed at the edges between different types of annuals crops (winter cereals, spring maize and pea) with contrasted cover states during the activity-period of carabids. Bidirectional interception traps, adapted from Hawthorne et al. (1998), were used to sample carabid movements between six adjacent crops. The interception traps were open continuously and collected weekly from early May to early September. As an example, Fig. 14.3 displays the orientation of carabid movements at field edges between a pea crop and two adjacent maize fields. It shows that, in the early summer, carabid beetles move more frequently from maize fields (with bare soil at this period) to pea fields (with dense vegetation cover). In late July, an inversion of the orientation of carabid movements at edges is observed, i.e. more movements from pea to maize fields, in relationship with pea harvest and vegetation growth in maize. Thus, P. melanarius seems to move throughout its activity-period from crops with unsuitable, sparse vegetation to crops with dense cover. This suggests that adjacent annual crops with asynchronous vegetation growth and timing of harvest could provide a temporal continuity of resources for this generalist predatory species. The persistence of carabid populations in a cultivated mosaic within a year might not necessarily require permanent habitats, but complementary cropped habitats that are spatially and temporally connected.

Direction of carabid movements at edges between pea and adjacent maize fields from May to September
3.2 Effect of Crop Rotation and Management Succession Over Years
The pluri-annual effects of the spatio-temporal heterogeneity of the cropping systems mosaic on biodiversity are still mainly unknown. However, crop rotation and management succession over years will determine the temporal availability of suitable cultivated resources patches for species. Moreover, the ability of species to spatially and temporally complement or supplement their resources between cropped habitats during their life cycle will partly drive their survival from one year to another, and therefore over the long term (Rusch et al. 2011; Thorbek and Topping 2005).
3.2.1 The Influence of Crop Rotations on Solitary Bees
To persist in a landscape, wild bees require nectar and pollen as food for brood and adults as well as suitable nesting sites (Westrich 1996). Intensive agriculture negatively affects the quality of bee habitat in several ways: (1) increasing crop field area results in the loss of suitable habitats including grasslands that are known to be highly beneficial habitats for bees (Klemm 1996; Steffan-Dewenter et al. 2002); (2) fertilizers, herbicides and intensive grazing reduce floral resources (De Snoo and Van der Poll 1999); (3) harvesting and tillage impede the nesting of most ground-nesting species (Shuler et al. 2005; Morandin et al. 2007); (4) some pesticides induce direct mortality or sublethal effects (Desneux et al. 2007). However, the crop mosaic can offer a great amount of easily available food resources when mass flowering crops such as oilseed rape or sunflower are cultivated (Westphal et al. 2003).
In order to better understand how landscape patterns influence solitary beeFootnote 1 communities, we took into account both spatial and temporal heterogeneities of the crop mosaic in addition to the commonly studied semi-natural elements (wooded elements and long-term grasslands) (Le Féon et al. 2011). Thus we considered the proportion of semi-natural elements, of oilseed rape and non-flowering crops at the moment of bee sampling and the proportion of two types of crop rotations. For the last variable, a crop rotation map summarising land-use history over a period of five years was realized. As cereals and grassland are the dominant land uses in the LTER site we distinguished two classes of crop fields: the fields that were sown only with cereals (wheat and maize) during the last five years and fields where the crop rotation included from one to four years of grassland (referred to as “mixed fields”). Solitary bees were trapped on 50 field margins, 15 of which were along oilseed rape fields and the 35 others were randomly located along other fields. Landscape composition was quantified in square windows centered on sampling points. Three window sizes were chosen, covering the range of relevant scales for flight and foraging distances of solitary bees (400, 800 and 1,200 m in width).
We found contrasted effects of non-flowering crops according to the type of margin and the spatial scale (Fig. 14.4). Solitary bee abundance in margins of oilseed rape fields deeply increased with the proportion of non-flowering crops at the moment of bee sampling at the finest spatial scale while it remained unchanged in margins of non-oilseed rape fields (Fig. 14.4a). This result shows that the attractiveness of mass-flowering crops depends on the quality of the surrounding landscape: the use of oilseed rape by solitary bees is higher when the surrounding area provides few floral resources.

Relationship between a solitary bee abundance and proportion of non-flowering crop fields during bee sampling (spring 2007) in 400 × 400 m windows; b solitary bee abundance and proportion of long-term grasslands in 1,200 × 1,200 m windows; c solitary bee species richness and proportion of “mixed fields” (“Mixed fields” are fields where crop rotation included from one to four years of temporary grassland during the period 2003–2007) in crop rotation in 800 × 800 m windows; d solitary bee species richness and proportion of “cereal fields” (“Cereal fields” are fields only sown with cereals (maize and wheat) during the period 2003–2007) in crop rotation in 1,200 × 1,200 m windows. Predictions returned by the Poisson-family models are shown by solid lines for significant relationships only (P ≤ 0.05, test F). × = oilseed rape field margins. • = non-oilseed rape field margins
Long-term grasslands and crop rotation influence local richness and abundance of bees at large spatial scales. Probably due to a masking effect of mass-flowering crops, these influences are only detected in margins of non-oilseed rape fields. Solitary bee abundance increased with the increasing proportion of long-term grasslands (Fig. 14.4b). Moreover solitary bee abundance and species richness increased with the increasing proportion of “mixed fields” (at least one year of grassland in the past five years) (Fig. 14.4c), whereas the proportion of fields only sown with cereals during the last five years had the opposite effect (Fig. 14.4d). The positive effect of long-term grassland is already known in landscape scale studies on wild bees (e.g. Steffan-Dewenter et al. 2002; Morandin et al. 2007). This type of fields is typically likely to provide wild flowers and suitable nesting sites. The originality in our results is to show that introducing temporary grasslands in cereal rotations is beneficial to bees. As they are generally sown with Poaceae species only and fertilized, the suitability of temporary grasslands for bees remains to be supported by further data. Nevertheless, the introduction of this cover type in cereal rotations could imply a less intensive farming system, potentially beneficial to solitary bees, thanks to (1) reduced pesticides and fertilizer inputs over the whole rotation cycle (2) greater floral resources in properly managed temporary grasslands (3) less disturbed soils better suited for ground-nesting bees. Our result is consistent with Steffan-Dewenter (2001) and Kuussaari et al. (2011) who showed the positive effect of the introduction of set-asides in cereal rotations on pollinator insects.
To sum up our findings, the composition of the landscape at the time of sampling had a direct impact on the spatial distribution of solitary bees only at the finest scale (400 m). On the contrary, when considering the landscape structure over several years (crop rotations and semi-natural elements like long-term grasslands), the effects occurred at the larger scales (800 and 1200 m). Therefore, our results are in agreement with the hierarchy theory that predicts that spatial and temporal scales are correlated. Phenomena occurring at coarse spatial scales are related to slower processes than phenomena occurring at smaller spatial scales (Allen et al. 1987). The maintenance of populations, a slow process, may be due to “large” spatio-temporal patterns, while feeding behavior, a fast process, is related to fine scale patterns (presence of a mass flowering crop in a given field). Our study showed that examining the heterogeneity of the agricultural mosaic over a whole crop rotation cycle was relevant to better understand the effects of agriculture on solitary bee communities. This approach allowed considering the cumulative effects of field cover and it demonstrated that introducing less intensive covers such as temporary grasslands in cereal rotations positively influences solitary bee communities.
3.2.2 Using Models to Predict the Influence of Management Successions and Crop Rotations on Biodiversity
In landscapes where too many habitat patches are simultaneously disturbed each year, the limited habitat complementation/supplementation processes might result in a population decrease or even extinction in the long term (Vasseur et al. in press). We used an existing spatially-explicit model to simulate the yearly and pluri-annual dynamics of populations of the Meadow brown (Maniola jurtina) under different scenarios of habitat disturbance extent (i.e. percent cover of grasslands mown in a 1 km² landscape) (Aviron et al. 2007). Simulations were run on the landscape unit where empirical data on the effect of mowing on butterfly movements were available, in order to validate the model’s predictions. Our results show that when habitat suppression through mowing occurs during the activity period of butterflies (in summer), butterfly populations get rapidly extinct if a large amount of grassland habitats (80 %) is simultaneously disturbed each year (Fig. 14.5). On the contrary, the synchronous disturbance of a lower amount of grasslands (20 %) each year allows population persistence and increase over the years, probably due to higher possibilities of habitat complementation/supplementation for butterflies (Fig. 14.5). Thus, the long-term persistence of butterfly populations could be strongly affected by the extent of habitat destruction and of direct disturbances caused by mowing.

Scenarios of butterfly habitat disturbance (20 and 80 % of grasslands mown each year) (a) and predicted evolution of total butterfly abundances over 10 years for the two scenarios of habitat disturbance and in absence of mowing (b) (derived from Aviron et al. 2007)
Complementation of resources by species is not only dependent on the availability of alternative suitable resource patches each year but also on their accessibility (Dunning et al. 1992). Thus, the spatial organization of asynchronous, complementary cropped habitats each year might be crucial for long-term population persistence as well. We used a spatio-temporally explicit model to simulate the dynamics of carabid populations (P. melanarius) in a cropping system mosaic characterized by a rotation of three annual crops (maize-wheat-barley) differing mainly by their period of sowing and harvest (“ANGORA” model, Vasseur 2012). Simulations were run on virtual landscapes (grids of 45 × 45 fields) with similar compositions each year (33 % of each crop type) but contrasted spatial organizations of the crop mosaic (i.e. 15 vs. 84 % of total interfaces between complementary crops) (Fig. 14.6a). The results show that, in crops mosaics with similar compositions, the population growth rate over 30 years is higher in mosaics where adjacency between complementary crops, i.e. with asynchronous cover states (maize-wheat, maize-barley, barley-wheat), is promoted (Fig. 14.6b). Adjacency between complementary crops each year increases survival of carabid beetles until the end of the reproductive period, ensuring a more important renewal of carabid populations. It also allows, in the latter part of carabid activity period, a rapid colonization of new, suitable patches (maize crops). Thus, the spatio-temporal heterogeneity of the cropping systems mosaic is likely to increase landscape spatio-temporal connectivity for this carabid species. Over the long term, this process could result in a homogenization of populations’ spatial distribution, and enhance their resilience to frequent local extinctions (Fig. 14.7).

Scenarios of spatial organization of the crop mosaic (15 and 84 % of interfaces between complementary crops: maize-wheat, wheat-barley and maize-barley) (a) and predicted growth rate of carabid populations (mean ± 95 % CI) after 30 years of simulation (10 cycles of rotation) for the two scenarios (b). (Details of the ANGORA model given in Vasseur 2012)

Spatial distribution of carabid populations in the crop mosaic after 30 years of simulation (10 cycles of rotation) for the two scenarios of crop spatial organization (15 and 84 % of interfaces between complementary crops: maize-wheat, wheat-barley and maize-barley) (Vasseur 2012)
4 Discussion
4.1 Semi Natural Elements
Our results confirm that semi-natural or more natural elements (Fahrig et al. 2011) have an impact on biodiversity for all considered taxa. This has been shown by previous results on the effect of the amount of these elements, considered as a measure of landscape complexity, on several taxonomic groups. Most of the studies show a positive effect of semi natural elements, and conclude that complex agricultural landscapes favor biodiversity as measured by the number of species (Tscharntke et al. 2005a, b). The use of several taxa permits to discuss this assumption; if the number of some small, short-lived species increases with changes in landscape composition and configuration, in the same landscapes there is no response by longer-lived ones. The first category may perceive the landscape at a finer spatio-temporal scale and thus react to changes that are not perceived by the other categories. Nevertheless, even if the total number of species is not related to the quantity of semi-natural areas, community structure changes. There is a shift in species or in the relative abundance of species. This points out the necessity to adapt the measure of biodiversity to the target species and to the intensity of landscape change. Species richness is a good indicator for strong gradients of landscape changes and for fine spatio-temporal grain species. Otherwise, some more sensitive measures such as the composition of communities or the relative abundance of species are needed to highlight the response of biodiversity to landscape structure changes.
In rural landscapes, all parts of the mosaic are influenced by agricultural activities. Even semi natural areas such as woodlots, hedgerows and even more permanent grassland depend on the farming system. They may be sprayed by pesticides from the crops, enriched by fertilizers from the upper fields; woodlot boundaries and hedgerows are pruned not to shadow crops (Lotfi et al. 2010), etc. It is thus an illusion to draw a strong boundary between semi-natural and productive areas. This is shown by our work on landscapes with similar compositions and different farming systems, with a strong response of species abundance and composition to changes in production type and farming practices. It is of overall importance when comparing landscapes with different structures to explicitly characterize farming activities. This has been done within a European project, green veins, with 24 landscape units, distributed along a double gradient of farming intensity and amount of semi-natural areas. The more important factor to explain biodiversity was then the intensity of the farming system (Billeter et al. 2008).
4.2 Cropping System Mosaic
Beyond the effects of farming system intensity, our results show that the spatio-temporal organization of crop covers, farming practices and crop rotations affect biodiversity. Until now, this issue has mainly been addressed for crop pest species (Carrière et al. 2006; Bresson et al. 2010; Kennedy and Storer 2010), but our results on pollinators, predatory arthropods and butterflies show that the heterogeneity of the cropping systems mosaic permits the persistence of beneficial organisms, and of species of conservation interest. For them, the diversity of farming practices and rotations, together with semi-natural elements, ensures habitat complementation and/or supplementation in space and time, and determines landscape connectivity. Over the years, crop rotations will not only control the degree of stability of resources for species, as shown for bee communities, but also their temporal accessibility, as illustrated by our modeling studies. Benefits of the cropping systems mosaic are expected for species in semi-natural elements, especially in landscapes where uncultivated elements are sparse. This has been underlined in previous studies, which showed that landscape connectivity for forest species increases when crops are grown high and dense (Fitzgibbon 1997; Ouin et al. 2000). The relative contribution of semi-natural and cultivated elements for biodiversity is however, likely to vary in time due to crop turnover and associated changing suitability of the cropping systems mosaic (Holzschuh et al. 2011).
4.3 The Role of Farming Systems
We have shown that farming systems interact with biodiversity in several ways and at different scales from fields to farm to groups of farms. This is summarized in Table 14.4. At the field scale, only species spending part of their life in crops are concerned. Their populations are driven by the food and microclimatic resources within the field. Processes in the mosaic of a few adjacent fields drive the same type of species, the mosaics control movements from field to field, therefore the possibilities to find food and shelter.
At a wider scale (about 100–1,000 ha), the controlling structure is the crop mosaic and associated semi-natural elements. The different landscape patterns offer different habitats and resources and, therefore, select the species that can thrive. Both species living in semi-natural elements and cropland species are concerned. When a landscape pattern changes by addition or removal of elements or by a new spatial distribution, the species that are not adapted vanish while new ones can come.
At the regional scale, the differentiation of farming systems implies a diversity of production and management practices in terms of inputs and disturbances such as soil tillage and harvesting. This is another major cause of species distribution.
4.4 Guidelines for policies
In terms of policies, it has been demonstrated that agri-environmental policies implemented at the field scale only are inefficient (Kleijn et al. 2006). The ignorance of the landscape context explains a large part of this failure (Concepción et al. 2008). An important point that is not integrated in the design of those policies is that the overall (gamma) diversity of a region depends on the diversity of landscapes and farming systems at all scales.
By deciphering the drivers of biodiversity in terms of landscape patterns and farming systems, our research shows that both are important and that field scale processes are controlled by external factors. Therefore, biodiversity objectives must be set at those different scales, taking into account the regional diversity.
In the European Union, policies related to agricultural practices already exist, as in the nitrate directive that makes compulsory the presence of a catch crop in winter in areas where nitrate leaching is a problem. Within the cross-compliance of the Common Agricultural Policy, farmers must record their use of fertilizers and pesticides that must be kept below a certain level. Crop diversification is an objective of the Common Agricultural Policy reform. In France, the implementation of grassy strips along streams is a first step toward a landscape scale management of water and biodiversity. To further enhance biodiversity, policies should include a limitation of field size and soil disturbance, such as long-term grassland and minimum tillage. The ban of herbicides in field margin management should be part of the package.
Heterogeneity is an important variable to enhance biodiversity (Benton et al. 2003). In the course of this chapter we show how “visible” heterogeneity linked to land cover, especially semi-natural elements and “hidden” heterogeneity (Vasseur et al. in press) resulting from farming practices play a major role to maintain high levels of biodiversity. In Table 14.5, we present the strengths and weaknesses of these two approaches of biodiversity
5 Conclusion
To conclude we may state that until now most regulations to enhance or conserve biodiversity have been aiming at increasing (Aviron et al. 2009) or managing extensively (Kleijn et al. 2011) semi-natural elements. But at an era when food production must increase to feed a growing world population it is important to identify practices at field and landscape levels that will favor biodiversity without retrieving land from production. One may expect a threshold of amount of semi natural areas, 0–5 %, below which biodiversity remains low whatever the practices, as the regional species pool will be low. Above this threshold, crop spatial heterogeneity and environmentally friendly practices will increase biodiversity. For high amounts of semi natural elements, 20 % onwards (Tscharntke et al. 2005b), biodiversity will be high, even simply by keeping the current farming activities (Leroux et al. 2008). In many parts of the world, agricultural landscapes fall within the second category. It is time to define policies that will encourage agricultural practices and systems that maximize biodiversity for its own sake and for the services it provides to our societies. It must also be acknowledged that all these policies will not increase all species, but may be targeted toward certain groups.
Notes
- 1.
Wild bees comprise of social species (Bombus sp.) and solitary bees (even if different forms of primitive or advanced social behavior exist in some species). Our study only focuses on solitary bees, which represent more than 80 % of wild bee species in Europe.
References
Allen TFH, O’Neill RV, Hoekstra TW. Inter level relations in ecological research and management: some working principles from hierarchy theory. J Appl Syst Anal. 1987;14:63–79.
Alston DG, Bradley JR, Schmitt DP, Coble HD. Relationship of Heliothis zea predators, parasitoids and entomopathogens to canopy development in soybean as affected by Heterodera glycines and weeds. Entomol Exp Appl. 1991;58:279–88.
Auffret AG, Cousins SAO. Past and present management influences the seed bank and seed rain in a rural landscape mosaic. J Appl Ecol. 2011;48:1278–85.
Aviron S, Burel F, Baudry J, Schermann N. Carabid assemblages in agricultural landscapes: impacts of habitat features, landscape context at different spatial scales and farming intensity. Agric Ecosyst Environ. 2005;108:205–17.
Aviron S, Kindlmann P, Burel F. Conservation of butterfly populations in dynamic landscapes: the role of farming practices and landscape mosaic. Ecol Model. 2007;205:135–45.
Aviron S, Nitsch H, Jeanneret P, Buholzer S, Luka H, Pfiffner L, Pozzi S, Schüpbach B, Walter T, Herzog F. Ecological cross compliance promotes farmland biodiversity in Switzerland. Front Ecol Environ. 2009;7:247–52.
Baudry J, Burel F. La mesure de la biodiversité spatiale: utilisation dans les évaluations d’impact. Acta Oecologica Oecologica Applicata. 1982;177–90.
Benton TG, Vickery JA, Wilson JD. Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol. 2003;18:182–8.
Billeter R, Liira J, Bailey D, Bugter R, Arens P, Augenstein I, Aviron S, Baudry J, Bukacek R, Burel F, Cerny M, De Blust G, De Cock R, Diekotter T, Dietz H, Dirksen J, Dormann C, Durka W, Frenzel M, Hamersky R, Hendrickx F, Herzog F, Klotz S, Koolstra B, Lausch A, Le Coeur D, Maelfait JP, Opdam P, Roubalova M, Schermann A, Schermann N, Schmidt T, Schweiger O, Smulders MJM, Speelmans M, Simova P, Verboom J, Van Wingerden WKRE, Zobel M. Indicators for biodiversity in agricultural landscapes: a pan-European study. J Appl Ecol. 2008;45:141–50.
Blondel J, Ferry C, Frochot B. la méthode des indices d’abondance (IPA) ou des relevés d’avifaune par “station d’écoute”. Alauda. 1970;38:55–71.
Brakefield PM. Ecological studies on the butterfly Maniola jurtina in Britain. I—Adult behaviour, micro distribution and dispersal. J Anim Ecol. 1982;51:713–26.
Bressan A, Moral García FJ, Sémétey O, Boudon-Padieu E. Spatio-temporal pattern of Pentastiridius leporinus migration in an ephemeral cropping system. Agric For Entomol. 2010;12:59–68.
Burel F, Baudry J, Butet A, Clergeau P, Delettre Y, Le Coeur D, Dubs F, Morvan N, Paillat G, Petit S, Thenail C, Brunel E, Lefeuvre JC. Comparative biodiversity along a gradient of agricultural landscapes. Acta Oecologica. 1998;19:47–60.
Burel F, Baudry J. Landscape ecology: concepts, methods, and applications. Enfield: N.H. Science Publishers; 2003.
Burel F, Baudry J. Habitat quality and connectivity in agricultural landscapes: the role of land use systems at various scales in time. Ecol Ind. 2005;5:305–13.
Carrière Y, Ellsworth PC, Dutilleul P, Ellers-Kirk C, Barkley V, Antilla L. A GIS-based approach for area wide pest management: the scales of Lygus hesperus movements to cotton from alfalfa, weeds, and cotton. Entomol Exp Appl. 2006;118:203–10.
Concepción E, Díaz M, Baquero R. Effects of landscape complexity on the ecological effectiveness of agri-environment schemes. Landscape Ecol. 2008;23:135–48.
Deckers B, De Becker P, Honnay O, Hermy M, Muys B. Sunken roads as habitats for forest plant species in a dynamic agricultural landscape: effects of age and isolation. J Biogeogr. 2005;32:99–109.
De Snoo GR, Van der Poll RJ. Effect of herbicide drift on adjacent boundary vegetation. Agric Ecosyst Environ. 1999;73:1–6.
Desneux N, Decourtye A, Delpuech JM. The sublethal effects of pesticides on beneficial arthropods. Annu Rev Entomol. 2007;52:81–106.
Dunning JB, Danielson BJ, Pulliam HR. Ecological processes that affect populations in complex landscapes. Oikos. 1992;65:169–75.
Ernoult E, Tremauville Y, Cellier D, Margerie P, Langlois E, Alard D. Potential landscape drivers of biodiversity components in a flood plain: Past or present patterns? Biol Conserv. 2006;127:1–17.
Fahrig L. Relative importance of spatial and temporal scales in a patchy environment. Theor Popul Biol. 1992;41:300–14.
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ, Sirami C, Siriwardena GM, Martin J-L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett. 2011;14:101–12.
Fischer J, Brosi B, Daily G, Ehrlich P, Goldman R, Goldstein J, Lindenmayer D, Manning A, Mooney H, Pejchar L, Ranganathan J, Tallis H. Should agricultural policies encourage land sparing or wildlife-friendly farming? Front Ecol Environ. 2008;6(7):380–5.
Forman RTT, Baudry J. Hedgerows and hedgerow networks in landscape ecology. Environ Manage. 1984;8:499–510.
Fitzgibbon CD. Small mammals in farm wood lands: the effects of habitat, isolation and surrounding land-use patterns. J Appl Ecol. 1997;34:530–9.
Gabriel D, Thies C, Tscharntke T. Local diversity of arable weeds increases with landscape complexity. Perspect Plant Ecol, Evol Syst. 2005;7:85–93.
Hawthorne AJ, Hassall M, Sotherton NW. Effects of cereal headland treatments on the abundance and movements of three species of carabid beetles. Appl Soil Ecol. 1998;9:417–22.
Holzschuh A, Dormann CF, Tscharntke T, Steffan-Dewenter I. Expansion of mass-flowering crops leads to transient pollinator dilution and reduced wild plant pollination. Proceedings of the Royal Society B-Biological Sciences 2011;278:3444–3451.
Joannon A, Bro E, Thenail C, Baudry J. Crop patterns and habitat preferences of the grey partridge farmland bird. Agron Sustainable Dev. 2008;28:379–87.
Kennedy GG, Storer NP. Life systems of polyphagous arthropod pests in temporally unstable cropping systems. Ann Rev Entomol. 2000;45:467–493.
Kennedy GG, Storer NP. Life systems of polyphagous arthropod pests in temporally unstable cropping systems. Annu Rev Entomol. 2000;45:467–93.
Kindlmann P, Burel F. Connectivity measures: a review. Landscape Ecol. 2008;23:879–90.
Kleijn D, Baquero RA, Clough Y, Díaz M, Esteban J, Fernández F, Gabriel D, Herzog F, Holzschuh A, Jöhl R, Knop E, Kruess A, Marshall EJP, Steffan-Dewenter I, Tscharntke T, Verhulst J, West TM, Yela JL. Mixed biodiversity benefits of agri-environment schemes in five European countries. Ecol Lett. 2006;9:243–54.
Kleijn D, Rundlã FM, Scheper J, Smith HG, Tscharntke T. Does conservation on farmland contribute to halting the biodiversity decline? Trends Ecol Evol. 2011;26:474–81.
Klemm M. Man-made bee habitats in the anthropogenous landscape of central Europe-substitutes for threatened or destroyed riverine habitats? The conservation of bees. In: Matheson A, Buchmann SL, O’Toole C, Westrich P, Williams IH, editors. 1996. p. 17–34. Academic Press, London.
Krauss J, Bommarco R, Guardiola M, Heikkinen RK, Helm A, Kuussaari M, Lindborg R, Öckinger E, Pärtel M, Pino J, Pöyry J, Raatikainen KM, Sang A, Stefanescu C, Teder T, Zobel M, Steffan-Dewenter I. Habitat fragmentation causes immediate and time delayed biodiversity loss at different trophic levels. Ecol Lett. 2010;13:597–605.
Kuussaari M, Hyvönen T, Härmä O. Pollinator insects benefit from rotational fallows. Agric Ecosyst Environ. 2011;143:28–36.
Féon Le V, Burel F, Chifflet R, Henry M, Ricroch A, Vaissière BE, Baudry J. Solitary bee abundance and species richness in dynamic agricultural landscapes. Agric Ecosyst Environ. 2011. doi:10.1016/j.agee.2011.06.020.
Leroux X, Barbault R, Baudry J, Burel F, Doussan I, Garnier E, Herzog F, Lavorel S, Lifran R, Roger-Estrade J, Sarthou JP, Trommetter M. Agriculture et biodiversité: valoriser les synergies, Paris, éds. Quae. 2008.
Lotfi A, Javelle A, Baudry J, BUREL F. Interdisciplinary analysis of hedgerow network landscapes sustainability. Landscape Res. 2010;35:391–402.
Meeus JHA. The transformation of agricultural landscapes in Western Europe. Sci Total Environ. 1993;129:171–90.
Men XY, Ge F, Yardim EN, Parajulee MN. Evaluation of winter wheat as a potential relay crop for enhancing biological control of cotton aphids in seedling cotton. Biocontrol. 2004;49:701–14.
Metzger JP. Landscape ecology: perspectives based on the 2007 IALE world congress. Landscape Ecol. 2008;23:501–4.
Millan-Pena N, Butet A, Delettre Y, Paillat G, Morand P, Le Du L, Burel F. Response of small mammal community to changes in western French agricultural landscapes. Landscape Ecol. 2003;18:265–78.
Millan De La Pena N, Butet A, Delettre Y, Morant P, Burel F. Landscape context and carabid beetles (Coleoptera: Carabidae) communities of hedgerows in western France. Agric Ecosyst Environ. 2003;94:59–72.
Morandin LA, Winston ML, Abbott VA, Franklin MT. Can pastureland increase wild bee abundance in agriculturally intense areas? Basic Appl Ecol. 2007;8:117–24.
Ouin A, Paillat G, Butet A, Burel F. Spatial dynamics of wood mouse (Apodemus sylvaticus) in an agricultural landscape under intensive use in the Mont Saint Michel Bay (France) agriculture. Ecosyst Environ. 2000;78:159–65.
Purtauf T, Roschewitz I, Dauber J, Thies C, Tscharntke T, Wolters V. Landscape context of organic and conventional farms: Influences on carabid beetle diversity. Agric Ecosyst Environ. 2005;108:165–74.
Robinson RA, Sutherland WJ. Post war changes in arable farming and biodiversity in Great Britain. J Appl Ecol. 2002;39:157–76.
Rusch A, Valantin-Morison M, Sarthou J-P, Roger-Estrade J. Multi-scale effects of landscape complexity and crop management on pollen beetle parasitism rate. Landscape Ecol. 2011;26:473–86.
Schooley RL, Branch LC. Habitat quality of source patches and connectivity in fragmented landscapes. Biodivers Conserv. 2011;20:1611–23.
Shuler RE, Roulston TH, Farris GE. Farming practices influence wild pollinator populations on squash and pumpkin. J Econ Entomol. 2005;98:790–5.
Schweiger O, Maelfait JP, Van Wingerden W, Hendricks F, Billeter R, Speelmans M, Augenstein I, Aukema B, Aviron S, Bailey DW, Bukacek R, Burel F, Diekotter T, Dirksen J, Frenzel M, Herzog F, Liira J, Roubalova M, Bugter R. Quantifying the impact of environmental factors on arthropod communities in agricultural landscapes across organizational levels and spatial scales. J Appl Ecol. 2005;42:1129–39.
Steffan-Dewenter I, Tscharntke T. Succession of bee communities on fallows. Ecography. 2001;24:83–93.
Steffan-Dewenter I, Münzenberg U, Bürger C, Thies C, Tscharntke T. Scale-dependent effects of landscape context on three pollinator guilds. Ecology. 2002;83:1421–32.
Thenail C, Baudry J. Variation of farm spatial land use pattern according to the structure of the hedgerow network (bocage) landscape: a case study in northeast Brittany. Agric Ecosyst Environ. 2004;101:53–72.
Thorbek P, Topping CJ. The influence of landscape diversity and heterogeneity on spatial dynamics of agrobiont linyphiid spiders: an individual-based model. Biocontrol. 2005;50:1–33.
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C. Landscape perspectives on agricultural intensification and biodiversity—ecosystem service management. Ecol Lett. 2005a;8:857–74.
Tscharntke T, Rand TA, Bianchi F. The landscape context of trophic interactions: insect spillover across the crop-non crop interface. Ann Zool Fenn. 2005b;42:421–32.
Vane Wright RI, Ackery PR. The biology of butterflies. British Museum: Academic Press; 1981.
Vasseur C, Joannon A, Aviron S, Burel F, Meynard JM, Baudry J. The cropping systems mosaic: how does the hidden heterogeneity of agricultural landscapes drive populations? Agri Ecosyst Environ (in press).
Vasseur C. Contrôles exercés par les mosaïques de systèmes de culture sur les dynamiques du carabe Pterostichus melanarius Illiger (Coleoptera, Carabidae). Thèse de l’Université de Rennes 1. 2012. p. 205.
Weibull A-C, Bengtsson J, Nohlgren E. Diversity of butterflies in the agricultural landscape: the role of farming system and landscape heterogeneity. Ecography. 2000;23:743–50.
Westphal C, Steffan-Dewenter I, Tscharntke T. Mass flowering crops enhance pollinator densities at a landscape scale. Ecol Lett. 2003;6:961–5.
Westrich P. Habitat requirements of central European bees and the problems of partial habitats. Matheson A, Buchmann SL, O’Toole C, Westrich P, Williams IH, editors. The conservation of bees. London: Academic; 1996. p. 1–16.
Wissinger SA. Cyclic colonization in predictably ephemeral habitats: a template for biological control in annual crop systems. Biol Control. 1997;10:4–15.
Acknowledgments
LTER research is supported by the zone atelier program of INEE-CNRS, additional support is provided by INRA, the ministry of environment, the European Union and the National Research Agency. We thank three anonymous reviewers for their constructive comments.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Burel, F., Aviron, S., Baudry, J., Le Féon, V., Vasseur, C. (2013). The Structure and Dynamics of Agricultural Landscapes as Drivers of Biodiversity . In: Fu, B., Jones, K. (eds) Landscape Ecology for Sustainable Environment and Culture. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-6530-6_14
Download citation
DOI: https://doi.org/10.1007/978-94-007-6530-6_14
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-6529-0
Online ISBN: 978-94-007-6530-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)