
Summary
Plastids usually contain numerous copies of their genome. The reason of maintaining high copy numbers of the plastome and their changes resulting from developmental, cellular (e.g., nuclear ploidy levels), and environmental cues remains elusive. The DNA is contained in certain regions of the plastids, the nucleoids. Number and shape of nucleoids change during leaf and chloroplast development. Generally, a substantial increase of nucleoids and therefore plastome copy numbers per organelle and cell occurs very early in leaf development, which later on provides not only enough plastomes for the distribution during plastid division, but may also meet the increasing demand for plastid gene products during chloroplast biogenesis. Later in leaf development the fate of chloroplast DNA seems to be regulated in a species-specific manner. While some species further increase the amount of plastid DNA (at least per cell), others seem to decrease plastome copy numbers per cell and per organelle during chloroplast maturation. The amount of chloroplast DNA decreases during senescence.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
I. Introduction
Since the discovery of DNA in chloroplasts (Ris and Plaut 1962), its structure, abundance, and coding capacity have been extensively investigated (for reviews see Butterfass 1979; Herrmann and Possingham 1980; Sugiura 1995; Bendich 2004; Green 2011). The chloroplasts have evolved by endosymbiosis from once free-living cyanobacteria (Margulis 1970; Martin 2003). Compared to their free-living ancestors, the size of the genome (plastome) was greatly reduced during this process (to approximately 130–160 kbp; Sugiura 1995). For the most part this is due to the constant relocation of genes from the ancestral genome/plastome to the nuclear genome (Timmis et al. 2004; Kleine et al. 2009). However, basic genetic mechanisms of the nucleic acid metabolism and gene expression such as replication, transcription, and translation have been maintained (Mache and Lerbs-Mache 2001; Green 2011). The reported numbers of plastome copies per leaf cell vary from a few hundreds to more than 10,000 depending on the organism examined (Lamppa and Bendich 1979; Tymms et al. 1983; Leutwiler et al. 1984; Shaver et al. 2006; Zoschke et al. 2007). The number of plastome copies seems therefore to be regulated in a species-specific manner. The structure of the genome is not fully understood. A substantial part of the DNA can be isolated in form of circular DNA molecules having the expected size of the complete genome (‘plastid chromosome’). In addition, there are linear and circular di- and oligomers, but also smaller than genome size and more complex molecules (Bendich 2007). Thus, if we mention in the following ‘plastome copy numbers’ or ‘plastid/chloroplast genome copies’, we refer to multiples of the DNA amount representing one genome, but not to multiples of the ‘plastid chromosome’. Higher plants, with their many different tissues and cell types, have evolved mechanisms to adapt plastome copy numbers to the needs of the respective cell. This review addresses various aspects of the organization, structure, and amount of plastid DNA during leaf development.
II. Plastid DNA Is Contained in Nucleoids
A. The Number of Nucleoids and Their DNA Content per Organelle
Erwin Baur, Carl Correns (both in 1909) and Otto Renner (1934) demonstrated by genetic experiments that chloroplasts are individual genetic elements. In 1962, by using electron microscopic and cytochemical methods, Ris and Plaut (1962) indeed showed the presence of DNA within one or more irregularly shaped bodies in the chloroplast of Chlamydomonas moewusii. Subsequently, it was shown that the plastid DNA is replicated in a semiconservative way and contains genes for ribosomal RNAs, transfer RNAs as well as soluble and membrane-bound proteins (for references see Sugiura 1992). The appearance of plastid DNA in sub-plastomic bodies further sparked scientific interest. Various methods were used to obtain information on the structure and organization of the plastid DNA within the chloroplast, which ranged from early analyses of plastids isolated from thymidine labeled tissue by microscopic autoradiography to the later use of a DNA-specific, fluorescent stain, DAPI (4′,6-diamidino-2-phenylindole) (Herrmann 1970; Herrmann and Kowallik 1970; Dann et al. 1971; Kowallik and Herrmann 1972; Coleman 1978, 1979; James and Jope 1978; Kuroiwa et al. 1981).
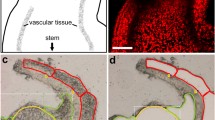
Similar to E. coli (Ogden et al. 1988; Landoulsi et al. 1990), plastid nucleoids are mostly situated at peripheral areas during active chloroplast division, since binding to the envelope membrane seems important for the replication of plastid DNA and equal distribution of the nucleoids (Kuroiwa et al. 1981; Miyamura et al. 1986). Later in development, the nucleoids were usually located within the organellar matrix associated with the thylakoids (Lindbeck et al. 1987). Membrane association is reported to enhance expression of several genes for photosynthetic components (Sato et al. 1993). The nucleoids of fully developed chloroplasts in leaves of angiosperms belong to the so-called SN-type (scattered nucleoids; Kuroiwa et al. 1981), which is distinguished by small nucleoids that are equally distributed throughout the plastid stroma (Fig. 11.1).

DAPI-stained mesophyll cell of a young rosette leaf of Arabidopsis thaliana. All chloroplasts contain numerous nucleoids. The nucleus exhibits bright fluorescence due to its high DNA content. (Courtesy of R.G. Herrmann and H. Golczyk.)
Interestingly, a correlation of the number of nucleoids, the amount of their DNA, and the size of the plastids has been reported (e.g., Herrmann 1970; Kowallik and Herrmann 1972; James and Jope 1978; Kuroiwa et al. 1981). Generally, polyploidization of the plastome within one organelle was shown by various isolation procedures, revealing multimeric copies and complex structures of the plastid DNA (Herrmann and Possingham 1980; Backert et al. 1995; Oldenburg and Bendich 2004; Scharff and Koop 2007). Kowallik and Herrmann (1972) furthermore suggested based on their in situ experiments, that nucleoids contain varying amounts of DNA even within one organelle. Similarly, individual polyploidization of plastid DNA of nucleoids, which can vary at least by a factor of ten, has been deduced from different fluorescent intensities after DAPI staining (Coleman 1978, 1979; James and Jope 1978; Miyamura et al. 1986).
Changes in DNA content, form, and number per organelle of nucleoids occur predominantly during chloroplast development. In proplastids and developing small chloroplasts only single or few mostly egg-shaped nucleoids have been observed. If kept in darkness, the nucleoids of the resulting etioplasts become cup-shaped and are often bound to the prolamellar body. Their size and DNA content increased about ten-times with an average number of ten nucleoids per etioplast. However, during chloroplast differentiation the nucleoids divided rapidly. Therefore, the average number of nucleoids per chloroplast increased up to 30 in fully developed leaves containing similar DNA amounts as in etioplasts (Herrmann 1970; Herrmann and Kowallik 1970; Kowallik and Herrmann 1972; James and Jope 1978; Scott and Possingham 1980; Kuroiwa et al. 1981; Miyamura et al. 1986; Sato et al. 1993; Fujie et al. 1994; Kato et al. 2007; Rauwolf et al. 2010).
B. Protein Components of Plastid Nucleoids
Plastid nucleoids are compact structures in which the plastid DNA is associated and organized with proteins (Kuroiwa 1991). During chloroplast development and conversions between different plastid types, nucleoids undergo changes in form, size, and location within the organelle (see above; Hashimoto 1985; Kuroiwa 1991), presumably mediated by modifications in the composition and abundance of certain nucleoid proteins (Kuroiwa and Suzuki 1981; Briat et al. 1982; Hansmann et al. 1985; Reiss and Link 1985; Nemoto et al. 1988, 1990; 1991; Nakano et al. 1993). Since nucleoids lack distinct membrane or envelope structures, it is often unclear if identified proteins are indeed inherent nucleoid proteins, e.g., commonly nuclear-localized histones found in nucleoid preparations may have been nuclear contaminants (Phinney and Thelen 2005). However, it is conceivable that nucleoid proteins fall into certain categories depending on their function. Early biochemical analyses indeed showed several nucleoid-associated proteins involved in DNA-, membrane-, and protein-binding (Hansmann et al. 1985; Bülow et al. 1987; Lindbeck et al. 1987), involved in regulating gene expression and replication (Lam et al. 1988; Khanna et al. 1992), and presumably in DNA packaging (Nemoto et al. 1990; Baumgartner and Mullet 1991).
To date, only few plastid nucleoid-associated proteins have been characterized in more detail. Although histone-like proteins with biochemical properties similar to bacterial HU proteins have been reported in higher plants (Briat et al. 1984; Nakano et al. 1993; Yurina et al. 1995; Phinney and Thelen 2005) and algae (Crevel et al. 1989; Grasser et al. 1997; Kobayashi et al. 2002), the function of these proteins in higher plant nucleoid formation is not yet clear. Interestingly, proteomic studies of nucleoids in pea chloroplasts revealed that one of the most abundant proteins was DCP68 (Phinney and Thelen 2005; cf. Majeran et al. 2012). This protein is a sulfite reductase with plastid DNA-binding and condensing activities (Cannon et al. 1999) suggesting it to have a bifunctional role in reductive sulfur assimilation and being a major component of organellar nucleoid organization (Chi-Ham et al. 2002). Furthermore, this study identified approximately 35 other proteins possibly associated with nucleoids including a family of DNA gyrases, as well as proteins involved in DNA repair, and plastid transcription and translation (Phinney and Thelen 2005). A plastid envelope DNA-binding protein (PEND) is thought to attach the nucleoids to the envelope membrane early in chloroplast development in order to provide an anchor for plastome replication and segregation during organelle division (Sato et al. 1993, 1998; Terasawa and Sato 2005; Wycliffe et al. 2005). However, the PEND protein, was not detected in maize nucleoids which did also not contain the bisulfite reductase (Majeran et al. 2012). Other candidates for anchoring the DNA/nucleoid to the membrane are MFP1, CND4 and TCP34 (Nakano et al. 1993; Jeong et al. 2003; Weber et al. 2006). CND41 was identified in chloroplast nucleoids of cultured tobacco cells (Nakano et al. 1993) and was more abundant in actively dividing plastids than in mature organelles (Nakano et al. 1997). However, more recent studies point to CND41 being a DNA-binding protease involved in Rubisco degradation and the translocation of nitrogen during senescence (Murakami et al. 2000; Kato et al. 2004). Plastid nucleoids are thought to further contain Whirly ssDNA-binding proteins (Krause et al. 2005), since they were co-purified with a chloroplast chromosome preparation (Pfalz et al. 2006) and shown to be associated with the thylakoid membrane (Prikryl et al. 2008). In Arabidopsis chloroplasts, Whirly proteins were shown to be important components of the plastid genome maintenance machinery, required for plastome stability (Maréchal et al. 2009; Cappadocia et al. 2010). Another nucleoid protein, YlmG, is proposed to be involved in chloroplast division and nucleoid partitioning and to have similar functions in cyanobacteria (Kabeya et al. 2010).
The eubacterial-type, plastid-encoded plastid RNA polymerase (PEP) can be isolated from plastids as both a soluble enzyme and an insoluble form, the ‘transcriptionally active chromosome’ (TAC; e.g., Suck et al. 1996; Krause and Krupinska 2000; Pfalz et al. 2006). The TAC is a membrane bound protein/DNA complex (Gruissem and Tonkyn 1993) and should form a major part of the nucleoid. Proteomic data of both TAC and plastid nucleoids demonstrated that the PEP and other proteins are parts of both complexes (e.g., Krause and Krupinska 2000; Phinney and Thelen 2005; Pfalz et al. 2006; Majeran et al. 2012).
Although nuclear-encoded plastid RNA-polymerases (NEP) have not yet been found in TAC/nucleoid preparations (for review see Liere and Börner 2007; Steiner et al. 2011; Majeran et al. 2012), there are some indications from studies with transcription inhibitors that NEP is as well part of plastid nucleoids (Sakai et al. 1998; Sato et al. 2009).
A recent investigation into the proteome of nucleoids of proplastids and mature chloroplasts from maize leaves by mass spectrometry revealed a core proteome of more than 150 proteins (Majeran et al. 2012). In agreement with and extending previous studies the identified proteins have functions related to DNA (replication, organization, and repair) to RNA (synthesis, processing, splicing, and editing), and translation (ribosomal proteins, ribosome assembly factors). Comparison of the protein compositions of nucleoids from proplastids vs. mature chloroplasts indicated a shift from RNA metabolism to translation and homeostasis as predominant functions to occur in nucleoids during chloroplast development (Majeran et al. 2012; also for more discussion and references on nucleoid proteins).
III. Amount of Plastid DNA
A. Genome Copy Numbers per Plastid and Cell Vary Between Species and Tissues
Precise information about genome sizes has been obtained by sequencing whole plastid chromosomes. This data, together with the determination of the amounts of DNA per cell and organelle, revealed that plastids generally contain many copies of their genome. However, large differences exist among the species with respect to the number of plastome copies in the organelles and cells (Table 11.1). Part of the documented variability is due to different nuclear DNA levels. There is evidence for a positive correlation between nuclear ploidy levels and the number of plastome copies (see Butterfass 1979; Herrmann and Possingham 1980; Rauwolf et al. 2010 for discussion of older data). Therefore, the differences in the number of plastome copies between Triticum monococcum and T. dicoccum species (Dean and Leech 1982) and Beta vulgaris genotypes (Rauwolf et al. 2010) shown in Table 11.1 are discussed as a response to different nuclear genome ploidies. In a recent study, Rauwolf et al. (2010) presented thorough analyses of the DNA amounts per plastid and their changes during development of sugar beet leaves. On an average, the amounts determined for plastids with diameters from 1 to 8 μm ranged from 0.15 to 4.9 × 102 pg DNA. This corresponds to approximately 12–330 plastomes per organelle with about 4–7 copies per nucleoid. Interestingly, although the nuclear genome/plastome ratio changed during development, the plastome copy numbers seemed to depend on nuclear ploidy levels within the different developmental stages (Butterfass 1979; Herrmann and Possingham 1980; Rauwolf et al. 2010). Nuclear ploidy levels were shown to often correlate with changes in cell size and plastid numbers (Barow 2006) suggesting a basic control level of plastome copy numbers by the same or similar pathways (Rauwolf et al. 2010). Interestingly, the change in nuclear ploidy from the tetraploid T. dicoccum to the hexaploid T. aestivum (Dean and Leech 1982) and the massive endopolyploidization occurring during leaf development in Arabidopsis do not result in an increase of plastome copy numbers (Zoschke et al. 2007; Rowan et al. 2009) indicating that higher nuclear ploidy levels are not necessarily coupled with an increase in copy numbers of the chloroplast genome.
A larger part of the variability in plastid genome copies as shown in Table 11.1 is certainly due to technical problems as apparent from variations in the results obtained for the same species by different laboratories and methods (e.g., number of plastid genomes per cell in Arabidopsis: Zoschke et al. 2007; Rowan et al. 2009; Beta vulgaris: Tymms et al. 1983; Rauwolf et al. 2010; Spinacia: Scott and Possingham 1983; Lawrence and Possingham 1986; Triticum aestivum: Boffey and Leech 1982; Miyamura et al. 1990). DAPI stained plastid DNA needs for quantification a DNA standard. Depending on the standard, the calculated DNA amounts may differ by 100 % (Oldenburg and Bendich 2004). Also binding of protein to DNA and/or different degrees of compaction of DNA affects the fluorescence of DAPI (Sekine et al. 2002). PCR and other methods based on total leaf DNA have the disadvantage of being not suited to differentiate between the various cell types. Isolation of plastids/chloroplasts and protoplasts may lead to DNA degradation (for references and discussion of technical problems see below). Not only technical problems, also variable growth conditions (e.g., light, see below) or differences in the age and developmental stage (even though Table 11.1 contains only data from mature leaves) may lead to apparently conflicting results. Nevertheless, large interspecies differences with respect to plastome copy numbers exist (Table 11.1). There are species with relatively low amounts of plastid DNA, e.g., Arabidopsis with less than 2,000 chromosome copies per leaf cell and less than 50 copies per mature chloroplast (Leutwiler et al. 1984; Draper and Hays 2000; Zoschke et al. 2007; Rowan et al. 2009). Species like rye (Avena sativa) represent the other extreme with about 10,000 plastome copies per cell and more than 100 copies per chloroplast (Hashimoto and Possingham 1989a).
Green tissues of leaves, cotyledons and stems contain more plastome copies per organelle than non-green tissues like epidermis, root tissues, meristems or the vegetative and generative cells of pollen (Table 11.2). Since the number of plastids per cell is usually much lower in non-green tissues than in photosynthetically active tissues (e.g., Leutwiler et al. 1984; Pyke and Leech 1992, 1994; Pyke et al. 1994; Marrison et al. 1999; Pyke 1999; Rowan et al. 2004; Rauwolf et al. 2010), non-green cells contain much less plastid genomes than green cells (see references in Table 11.2; Lamppa and Bendich 1979; Cannon et al. 1986; Isono et al. 1997). Thus, for mesophyll cells of mature spinach leaves more than 16,000 plastome copies have been reported, while epidermis cells of the same leaves were found to possess only somewhat over 600 copies (Lawrence and Possingham 1986). Amyloplasts in potato tubers appear to be an exception. These non-green plastids were reported to contain with 195 plastome copies much more plastid DNA than leaf chloroplasts with only 22 copies (Scott et al. 1984). In a mixotrophic suspension culture of spinach cells, however, the copy number of plastomes in chloroplast-containing cells of the stationary phase was 5,940 contrasting with only 1,125 copies in amyloplast-containing cells of the log phase (Aguettaz et al. 1987). A similar ratio was calculated for plastome copies in leaf vs. root cells of Arabidopsis (Isono et al. 1997) and potato (Scott et al. 1984).
B. Variation in Amount and Structure of Plastid DNA During Leaf Development
1. The Fate of Plastid DNA Differs Between Species
There is convincing evidence for the increase in the number and size of plastids and in the amount of DNA per plastid, and consequently per cell, during the earliest stages of leaf development when green, photosynthetically active chloroplasts develop from the smaller, undifferentiated proplastids in the presence of light (Table 11.3). This holds true for both dicotyledenous and monocotyledenous plants. Among the latter, only leaves of graminaceous species have been investigated so far. They are particularly suitable for studies on leaf development since new cells are produced only from the basal meristem; i.e., the leaf cells are in a linear array with the youngest near the base and the oldest at the tip of the blade (Boffey et al. 1979). However, the position of a cell within leaf indicates its age also in dicots. Therefore, several studies on plastome copies in leaf cells included not only leaves of different ages but also material from different leaf sections (Table 11.3). An increase in plastome copies is also observed during the development of etioplasts from proplastids in young cells; i.e., when leaves develop in complete darkness (Bennett and Radcliffe 1975; Baumgartner et al. 1989; Hashimoto and Possingham 1989b; Miyamura et al. 1990; Oldenburg et al. 2006; Shaver et al. 2008; Zheng et al. 2011). A rise in the amount of DNA per developing chloroplast/etioplast is only possible if the rate of plastid DNA replication exceeds the rate of plastid division, a condition that is normally not fulfilled in non-green tissues (Table 11.2).
Also light seems to affect the DNA content in a species-specific manner (Table 11.3). The reduction of plastid DNA copies during chloroplast maturation per chloroplast and cell as detected by DAPI staining depends on light in maize and M. truncatula (Oldenburg et al. 2006; Shaver et al. 2008; Zheng et al. 2011), a phenomenon that was also observed in barley leaves by dot blot hybridization (Baumgartner et al. 1989). The light-induced decrease of DNA per cell was neither observed in leaves of rye (Avena sativa), soybean, and wheat (Cannon et al. 1986; Hashimoto and Possingham 1989a; Miyamura et al. 1990), nor in cultured leaf discs of spinach. In the latter material, light stimulated the synthesis of chloroplast DNA (Hashimoto and Possingham 1989b).
After having reached a high level per chloroplast and cell early in leaf development, the chloroplast DNA may adopt different, species-specific courses during further leaf development: (1) it may keep the high level until senescence; (2) copy numbers per plastid may decline to a lower level caused by ongoing plastid division without replication of the DNA while the copy number per cell remains relatively stable, subsequently the DNA may keep the lower level in all chloroplasts until senescence; (3) copy numbers per plastid and per cell may be reduced to a lower level by ongoing plastid division without replication of the DNA and in addition due to active degradation of part of the DNA, subsequently the DNA may keep the lower level per cell and in all chloroplasts until senescence; (4) the DNA content per chloroplast may rapidly decline due to active degradation and plastid division resulting in the complete loss of DNA in most chloroplasts long before senescence. Those four courses are proposed based on published data (Table 11.3), however, they remain speculation. It is obvious that there are differences between the species with respect to the fate of plastid DNA during leaf maturation. Yet, in most cases (Table 11.3) there are not enough data available to unequivocally group the species into one out of those four routes, since, unfortunately, comprehensive data sets are missing concerning the earliest and/or latest stages of leaf development; i.e., just those stages which are expected to show drastic alterations in plastid DNA amounts.
An example for course 1 (see above) may be the fate of plastid DNA in sugar beet (Beta vulgaris) leaves. Rauwolf et al. (2010) report a steady increase in the DNA content of plastids and cells during leaf development reaching highest levels in mature leaves confirming and extending older data (Herrmann 1970; Herrmann and Kowallik 1970; Kowallik and Herrmann 1972; cf. Tymms et al. 1983). Also rye plastid DNA may follow this way. Hashimoto and Possingham (1989a) measured the fluorescence of DAPI-stained plastid DNA and calculated the number of plastomes in individual chloroplasts and cells along the developmental gradient in the blades of primary leaves. They observed a rise of plastome copies both per chloroplast and per cell over the first basal 10 mm without further alteration in the level per cell during maturation of leaves and chloroplasts up to the tip. There was a slight decline in the DNA content per plastid between 10 and 30 mm, but no further change up to the leaf tip. The fate of chloroplast DNA in tobacco (Nicotiana tabacum) leaves may meet course 1 (Li et al. 2006) or course 2 (Rowan and Bendich 2009). Courses 2–4 are similar to each other: after a rise follow a descent in plastid DNA levels before senescence starts, but rate and extent of the decline in DNA content differ. Most species are reported to exhibit a decline in the DNA content per plastid during leaf maturation (Table 11.3). The fate of plastid DNA may follow course 2 in pea, wheat and spinach. The drop in plastome copy numbers per organelle during leaf maturation is suggested to result from the division of plastids without corresponding replication of DNA. There is no indication for significant reductions of plastid DNA per cell and therefore not for active degradation of DNA in these species (Lamppa et al. 1980; Boffey and Leech 1982; Lawrence and Possingham 1986; Miyamura et al. 1990). Barley may adopt course 3 (Baumgartner et al. 1989), also Medicago truncatula (although the observed decline in plastid DNA per leaf cell is only about 50–60 %; Shaver et al. 2008), and rice (Oryza sativa; Sodmergen et al. 1989, 1991). During the first 96 h after imbibition, number and size of chloroplasts and the amount of plastid DNA increased in the second rice leaf reaching a level of about 100 copies per plastid and 1,800 per cell. While chlorophyll content, chloroplast number and size remained stable, the number of plastome copies per chloroplast and cell dropped down to approximately 20 and 550, respectively, during the following 72 h. At this point of time, 168 h after imbibition, DAPI did not stain the chloroplast DNA anymore. Aging of the second leaves starts later at 240 h after imbibition. Therefore, degradation of plastid DNA was suggested to trigger senescence (Sodmergen et al. 1991).
We have categorized barley and rice as course 3 because the lowest reported plastome numbers per chloroplast are 50 and 20, respectively (Baumgartner et al. 1989; Sodmergen et al. 1991). All mature chloroplasts in cells from the middle section of primary barley leaves showed intensive fluorescence of the DNA/nucleoids after DAPI staining (Scott et al. 1982). It is not clear from the presented data how the number of 20 plastome copies per organelle fits to the observation that almost all mature chloroplasts could not be stained by DAPI in rice (Sodmergen et al. 1991). Like maize (Zea mays, Oldenburg and Bendich 2004; Oldenburg et al. 2006; Zheng et al. 2011), and perhaps Arabidopsis (Rowan et al. 2004, 2009), rice could also be an example for course 4, or vice versa, chloroplast DNA in all those species might actually meet fate 2 or 3. The proposed courses 3 and 4 differ from each other in the amount of DNA per chloroplast after its decline but well before the onset of senescence: a low level in most chloroplasts compared to young, dividing plastids in case of course 3 and none in most chloroplasts in case of course 4. If the lack of DAPI fluorescence would indeed indicate the absence of DNA from chloroplasts, then rice, maize and Arabidopsis would adopt course 4, since for all three species the decline in plastid DNA content is reported to result in chloroplasts without DAPI-stainable DNA (the vast majority of chloroplasts was found to contain no DNA as far as it is detectable by DAPI staining; Sodmergen et al. 1991; Oldenburg and Bendich 2004; Rowan et al. 2004; Evans et al. 2010). DAPI staining combined with a highly sensitive detection technique was demonstrated to detect DNA of the size of a single plastid genome (Miyamura et al. 1986). This does not necessarily mean that such sensitivity can be achieved under all conditions. In other species (which we would classify as species that adopt course 1 or 2), DNA/nucleoids can easily be observed after DAPI staining in virtually all chloroplasts of mature leaves (e.g., Scott and Possingham 1980; Scott et al. 1984; Cho et al. 2004; Rauwolf et al. 2010; Fulgosi et al. 2012). Thus, a dramatic reduction or even lack of DNA in all or most mature chloroplast (courses 3 and 4) is not a general feature of chloroplast and leaf development in contrast to a recent proposal (Rowan and Bendich 2009).
2. Do Mature Chloroplasts Lack DNA?
Data of several studies challenge the idea that there is no or nearly no DNA in mature chloroplasts of potential course four-species like maize and Arabidopsis. DNA blot hybridization and quantitative real-time PCR (qPCR) contradict the conclusions drawn from DAPI data by indicating no or only minor changes in the DNA content of chloroplasts during development (Draper and Hays 2000; Li et al. 2006; Zoschke et al. 2007; Rowan et al. 2009; Evans et al. 2010; Zheng et al. 2011). Rowan et al. (2009) proposed that the disagreement between data obtained by different methods might result from the fact that only green tissue was usually analyzed by DAPI staining whereas qPCR and hybridization were applied to DNA originating from all leaf tissues. It is easily imaginable that in one cell type (e.g., in green mesophyll cells) the DNA per chloroplast decreases during leaf maturation and the nuclear DNA remains stable, while in other cells (e.g., all non-green cells) the nuclear DNA increases due to endopolyploidization, but the number of plastome copies per plastid does not change. Rowan et al. (2009) suggested that under such conditions quantitative PCR with total DNA as template would not detect the decline of chloroplast DNA. However, a four-fold decline of chloroplast DNA in 50 % of the cells (about 50 % of cells in wheat leaves are mesophyll cells, Dean and Leech 1982; Arabidopsis leaves consist of about 50 % green and 50 % non-green cells, Rowan et al. 2009), and a stable plastid DNA content in the remaining 50 % of cells (non-green tissues contain much less DNA per plastid and per cell, Table 11.2) would be easily detectable by PCR.
More recently, a second explanation was proposed for the deviating results of DAPI staining on one hand and blot hybridization and PCR on the other hand – the potential existence of copies of chloroplast sequences within the nuclear (and mitochondrial) genome (Zheng et al. 2011). However, copies of chloroplast DNA sequences in nuclear and mitochondrial genomes were taken into account in the qPCR and hybridization studies by Li et al. (2006) and Zoschke et al. (2007); yet, no changes in chloroplast DNA content were observed during leaf development.
Thirdly, a possible explanation for the contradicting results may be that under certain conditions DAPI staining does not provide correct results. Rowan and Bendich (2009) could rule out that DNase acting during isolation on chloroplasts would lead to the observed loss of DNA, since the demise of chloroplast DNA was also observed by DAPI staining in situ. However, Evans et al. (2010) suggested that DAPI might have no access to chloroplasts at later stages of development and therefore does not stain their DNA. In addition, changes in structure and organization of chloroplast DNA during leaf development might lead to altered interactions with DAPI (Sekine et al. 2002; Oldenburg and Bendich 2004; Rowan et al. 2004; Shaver et al. 2006).
Data obtained in another series of experiments are also not in agreement with the lack of DNA in most chloroplasts of mature leaves suggested by the missing DAPI staining. The lack of DNA in chloroplasts would preclude transcription. Yet, active chloroplast transcription is commonly observed in mature chloroplasts. The transcription rates of several Arabidopsis chloroplast genes (including house-keeping and photosynthesis genes) were measured by run-on assays with chloroplasts from cotyledons and rosette leaves of different ages. The activities of all genes were highest in young and mature leaves exhibiting full photosynthetic activity (6–18-day-old) and still reached 25–50 % of their maximal values in old leaves at the beginning of senescence (Zoschke et al. 2007). Similarly, the transcriptional activity of chloroplast genes was determined in chloroplasts isolated from different sections of maize leaves representing the developmental gradient from very young to mature cells. Transcription of all investigated genes was highest in the cells at the tip of leaves, i.e. in the oldest cells with mature chloroplast (Cahoon et al. 2004). Furthermore, analyses of the transcriptional activity of chloroplast genes in barley during plant development and artificially induced senescence (prolonged dark treatment) showed active transcription of most tested genes in old or senescent leaves (Krupinska and Falk 1994; Krause et al. 1998). According to the data obtained by DAPI staining, most mature chloroplasts do not contain DNA, while a few chloroplast showed still a high fluorescence indicating the presence of many genome copies (Oldenburg and Bendich 2004; Zheng et al. 2011). These few chloroplasts might theoretically be responsible for the observed transcription activities in chloroplast isolated from mature or senescent leaves (Krupinska and Falk 1994; Krause et al. 1998; Cahoon et al. 2004; Zoschke et al. 2007). Why should transcription be needed in a few chloroplasts and not in the vast majority of chloroplasts? Chloroplasts without DNA seem imaginable since most mitochondria of higher plants do not possess the complete genome and might function (for a short time?) even without DNA (Preuten et al. 2010; Wang et al. 2010). Yet, the situation of mitochondria is not comparable to the situation of chloroplasts since mitochondria can theoretically exchange components like genes and gene products by frequent fission and fusion, i.e. the individual mitochondria seem to exist only for short periods of time (Arimura et al. 2004; Sheahan et al. 2005). Fusion of chloroplasts has not been observed, and stromules interconnecting chloroplasts (Gray, Chap. 9) might not be formed frequently enough to be a substitute for organellar fusion as a means of exchange (see Chap. 7; Hanson and Sattarzadeh 2011). At least in mature leaves of barley all chloroplasts were found to be transcriptionally active. By a combination of autoradiography, light microcopy and electron microscopy, Siemenroth et al. (1981) showed that all chloroplasts in 8–10-day-old primary leaves incorporated on a similar scale labeled uridine into RNA.
Taken together, the data suggest that all chloroplasts in mature leaves do contain substantial amounts of DNA, and the apparent loss of DNA in chloroplasts during maturation and long before senescence (‘course four’) may be an artefact due to technical problems with staining the DNA by DAPI. Obviously, the situation calls for more carefully designed experiments.
3. Chloroplast DNA Levels Decline During Senescence
Regardless of the reported differences in the fate of chloroplast DNA during leaf development, the data show consistently that leaf cells maintain hundreds or thousands copies of the plastome (Table 11.3) and that chloroplast genes are active at least until the start of pigment degradation, i.e. the visible onset of senescence (e.g., Jiang et al. 1993; Zoschke et al. 2007; Evans et al. 2010). There are only a few studies, which followed the further fate of chloroplast DNA during senescence. In soybean (Glycine max), the chloroplast DNA content per organelle and per cell starts to decline when Rubisco large subunit (rbcL) mRNA levels, Rubisco protein levels and Rubisco activity decrease (Jiang et al. 1993). In Arabidopsis, the level of rbcL mRNA starts to drop down and rbcL transcription declines at about the stage when the degradation of chlorophyll is initiated, while there is still no significant decline of chloroplast DNA. At this stage, the accumulation of psbA transcripts remained high, though psbA transcription was damped (Zoschke et al. 2007). During later stages of senescence, in yellow Arabidopsis leaves having lost more than 80 % of their chlorophyll, the levels of chloroplast DNA and rbcL mRNA further decrease, whereas the number of chloroplasts and the psbA mRNA level remain stable (Evans et al. 2010). Similarly, a steady decrease of DAPI staining of plastid nucleoids and of the rbcL gene as detected by DNA-blot hybridization was observed during natural senescence of maple (Acer pseudoplatanus) leaves (Fulgosi et al. 2012). Taken together, the data suggest that senescing leaf cells keep plastid DNA as long as they keep their chloroplasts.
IV. Conclusions
Plastids normally contain many copies of their genome. To date the reason of maintaining high copy numbers of the plastome and their changes resulting from developmental, cellular (e.g., nuclear ploidy levels), and environmental cues remains elusive. It has been suggested that the adjustment of plastome copy numbers is a regulatory response to varying demands of plastid gene expression (Bennett and Radcliffe 1975; Scott and Possingham 1983), in particular of the genes for ribosomal RNA (Bendich 1987). However, the transcriptional activity of plastid genes seems not generally to be correlated with plastome copy numbers (Eberhard et al. 2002; Zoschke et al. 2007). A massive increase in plastome copy numbers per organelle and cell occurs very early in leaf development accompanied by a rise in the number of nucleoids per plastid. This increase is likely owing to the increased demand for plastid gene products during chloroplast biogenesis as mentioned above, but also to provide enough plastome copies for distribution to the daughter organelles during the phase of very active plastid division (Kuroiwa 1991). The available data suggests a species-specific fate of chloroplast DNA later during leaf development. Some species may further increase plastome numbers (at least per cell) during leaf maturation, while others may decrease the amount of plastid DNA per organelle and per cell before senescence. During senescence, the amount of chloroplast DNA per cell and organelle decreases. There is no proof for the assumption that degradation of DNA triggers senescence in leaves (Sodmergen et al. 1989), since rather large copy numbers of the plastome and chloroplast gene expression can still be observed at later stages of senescence. We have now a rather detailed understanding of the process of plastid division (Miyagishima 2011), but near to nothing is known about replication of plastid DNA (Parent et al. 2011) or the enzymes that degrade plastid DNA (Sodmergen et al. 1989). An attractive and challenging field of future research will be to find out how cells can control plastid numbers and plastid DNA levels in response to endogenous and environmental cues.
Abbreviations
- CND41 –:
-
41 kDa chloroplast nucleoid DNA binding protein;
- DAPI –:
-
4′,6-diamidino-2-phenylindole;
- NEP –:
-
Nuclear-encoded plastid RNA-polymerase;
- PEND –:
-
Plastid envelope DNA-binding protein;
- PEP –:
-
Plastid-encoded plastid RNA-polymerase;
- ppGpp –:
-
Guanosine 5′-diphosphate 3′-diphosphate;
- qPCR –:
-
Quantitative real-time PCR;
- RubBisCO –:
-
Ribulose-1,5-bisphosphate carboxylase oxygenase;
- SN-type –:
-
Scattered nucleoid type; TAC – Transcriptionally active chromosome
References
Aguettaz P, Seyer P, Pesey H, Lescure A-M (1987) Relations between the plastid gene dosage and the levels of 16S rRNA and rbcL gene transcripts during amyloplast to chloroplast change in mixotrophic spinach cell suspensions. Plant Mol Biol 8:169–177
Arimura S, Yamamoto J, Aida GP, Nakazono M, Tsutsumi N (2004) Frequent fusion and fission of plant mitochondria with unequal nucleoid distribution. Proc Natl Acad Sci USA 101:7805–7808
Backert S, Dorfel P, Börner T (1995) Investigation of plant organellar DNAs by pulsed-field gel electrophoresis. Curr Genet 28:390–399
Barow M (2006) Endopolyploidy in seed plants. Bioessays 28:271–281
Baumgartner BJ, Mullet JE (1991) Plastid DNA synthesis and nucleic acid-binding proteins in developing barley chloroplasts. J Photochem Photobiol B 11:203–218
Baumgartner BJ, Rapp JC, Mullet JE (1989) Plastid transcription activity and DNA copy number increase early in barley chloroplast development. Plant Physiol 89:1011–1018
Bendich AJ (1987) Why do chloroplasts and mitochondria contain so many copies of their genome? Bioessays 6:279–282
Bendich AJ (2004) Circular chloroplast chromosomes: the grand illusion. Plant Cell 16:1661–1666
Bendich AJ (2007) The size and form of chromosomes are constant in the nucleus, but highly variable in bacteria, mitochondria and chloroplasts. Bioessays 29:474–483
Bennett J, Radcliffe C (1975) Plastid DNA replication and plastid division in the garden pea. FEBS Lett 56:222–225
Boffey SA, Leech RM (1982) Chloroplast DNA levels and the control of chloroplast division in light-grown wheat leaves. Plant Physiol 69:1387–1391
Boffey SA, Ellis JR, Selldén G, Leech RM (1979) Chloroplast division and DNA synthesis in light-grown wheat leaves. Plant Physiol 64:502–505
Briat JF, Gigot C, Laulhere JP, Mache R (1982) Visualization of a spinach plastid transcriptionally active DNA-protein complex in a highly condensed structure. Plant Physiol 69:1205–1211
Briat JF, Letoffe S, Mache R, Rouviere-Yaniv J (1984) Similarity between the bacterial histone-like protein HU and a protein from spinach chloroplasts. FEBS Lett 172:75–79
Bülow S, Reiss T, Link G (1987) DNA-binding proteins of the transcriptionally active chromosome from mustard (Sinapis alba L.) chloroplasts. Curr Genet 12:157–159
Butterfass T (1979) Patterns of chloroplast reproduction. A developmental approach to protoplasmic plant anatomy. Springer, Wien
Cahoon AB, Harris FM, Stern DB (2004) Analysis of developing maize plastids reveals two mRNA stability classes correlating with RNA polymerase type. EMBO Rep 5:801–806
Cannon G, Heinhorst S, Weissbach A (1986) Plastid DNA content in a cultured soybean line capable of photoautotrophic growth. Plant Physiol 80:601–603
Cannon GC, Ward LN, Case CI, Heinhorst S (1999) The 68 kDa DNA compacting nucleoid protein from soybean chloroplasts inhibits DNA synthesis in vitro. Plant Mol Biol 39:835–845
Cappadocia L, Maréchal A, Parent J-S, Lepage E, Sygusch J, Brisson N (2010) Crystal structures of DNA-Whirly complexes and their role in Arabidopsis organelle genome repair. Plant Cell 22:1849–1867
Chi-Ham CL, Keaton MA, Cannon GC, Heinhorst S (2002) The DNA-compacting protein DCP68 from soybean chloroplasts is ferredoxin:sulfite reductase and co-localizes with the organellar nucleoid. Plant Mol Biol 49:621–631
Cho HS, Lee SS, Kim KD, Hwang I, Lim J-S, Park Y-I, Pai H-S (2004) DNA gyrase is involved in chloroplast nucleoid partitioning. Plant Cell 16:2665–2682
Coleman AW (1978) Visualization of chloroplast DNA with two fluorochromes. Exp Cell Res 114:95–100
Coleman AW (1979) Use of the fluorochrome 4′6-diamidino-2-phenylindole in genetic and developmental studies of chloroplast DNA. J Cell Biol 82:299–305
Crevel G, Laine B, Sautière P, Galleron C (1989) Isolation and characterization of DNA-binding proteins from the cyanobacterium Synechococcus sp. PCC 7002 (Agmenellum quadruplicatum) and from spinach chloroplasts. Biochim Biophys Acta 1007:36–43
Dann O, Bergen G, Demant E, Volz G (1971) Trypanocide Diamidine des 2-Phenyl-benzofurans, 2-Phenyl-indens und 2-Phenyl-indols. Liebigs Ann Chem 749:68–89
Dean C, Leech RM (1982) Genome expression during normal leaf development: I. cellular and chloroplast numbers and DNA, RNA, and protein levels in tissues of different ages within a seven-day-old wheat leaf. Plant Physiol 69:904–910
Draper CK, Hays JB (2000) Replication of chloroplast, mitochondrial and nuclear DNA during growth of unirradiated and UVB-irradiated Arabidopsis leaves. Plant J 23:255–265
Eberhard S, Drapier D, Wollman F-A (2002) Searching limiting steps in the expression of chloroplast-encoded proteins: relations between gene copy number, transcription, transcript abundance and translation rate in the chloroplast of Chlamydomonas reinhardtii. Plant J 31:149–160
Evans IM, Rus AM, Belanger EM, Kimoto M, Brusslan JA (2010) Dismantling of Arabidopsis thaliana mesophyll cell chloroplasts during natural leaf senescence. Plant Biol 12:1–12
Fujie M, Kuroiwa H, Kawano S, Mutoh S, Kuroiwa T (1994) Behavior of organelles and their nucleoids in the shoot apical meristem during leaf development in Arabidopsis thaliana L. Planta 194:395–405
Fulgosi H, Jezic M, Lepedus H, Peharec Stefanic P, Curkovic-Perica M, Cesar V (2012) Degradation of chloroplast DNA during natural senescence of maple leaves. Tree Physiol 32:346–354
Grasser KD, Ritt C, Krieg M, Fernández S, Alonso JC, Grimm R (1997) The recombinant product of the Chryptomonas phi plastid gene hlpA is an architectural HU-like protein that promotes the assembly of complex nucleoprotein structures. Eur J Biochem 249:70–76
Green BR (2011) Chloroplast genomes of photosynthetic eukaryotes. Plant J 66:34–44
Gruissem W, Tonkyn JC (1993) Control mechanisms of plastid gene expression. Crit Rev Plant Sci 12:19–55
Hansmann P, Falk H, Ronai K, Sitte P (1985) Structure, composition, and distribution of plastid nucleoids in Narcissus pseudonarcissus. Planta 164:459–472
Hanson MR, Sattarzadeh A (2011) Stromules: recent insights into a long neglected feature of plastid morphology and function. Plant Physiol 155:1486–1492
Hashimoto H (1985) Changes in distribution of nucleoids in developing and dividing chloroplasts and etioplasts of Arena sativa. Protoplasma 127:119–127
Hashimoto H, Possingham J (1989a) DNA levels in dividing and developing plastids in expanding primary leaves of Avena sativa. J Exp Bot 40:257–262
Hashimoto H, Possingham JV (1989b) Effect of light on the chloroplast division cycle and DNA synthesis in cultured leaf discs of spinach. Plant Physiol 89:1178–1183
Herrmann RG (1970) Multiple amounts of DNA related to the size of chloroplasts. I. An autoradiographic study. Planta 90:80–96
Herrmann RG, Kowallik KV (1970) Multiple amounts of DNA related to the size of chloroplasts. II. Comparison of electron-microscopic and autoradiographic data. Protoplasma 69:365–372
Herrmann RG, Possingham JV (1980) Plastid DNA – the plastome. In: Reinert J (ed) Results and Problems in Cell Differentiation, vol 10 Chloroplasts, pp. 45–96. Springer Verlag, Berlin/Heidelberg/New York
Isono K, Niwa Y, Satoh K, Kobayashi H (1997) Evidence for transcriptional regulation of plastid photosynthesis genes in Arabidopsis thaliana roots. Plant Physiol 114:623–630
James TW, Jope C (1978) Visualization by fluorescence of chloroplast DNA in higher plants by means of the DNA-specific probe 4′6-diamidino-2-phenylindole. J Cell Biol 79:623–630
Jeong SY, Rose A, Meier I (2003) MFP1 is a thylakoid-associated, nucleoid-binding protein with a coiled-coil structure. Nucl Acids Res 31:5175–5185
Jiang CZ, Rodermel SR, Shibles RM (1993) Photosynthesis, RuBisCO activity and amount, and their regulation by transcription in senescing soybean leaves. Plant Physiol 101:105–112
Jope CA, Hirai A, Wildman SG (1978) Evidence that the amount of chloroplast DNA exceeds that of nuclear DNA in mature leaves. J Cell Biol 79:631–636
Kabeya Y, Nakanishi H, Ishikawa T, Kondou Y, Matsui M, Miyagishima S-Y (2010) The YlmG protein has a conserved function related to the distribution of nucleoids in chloroplasts and cyanobacteria. BMC Plant Biol 10:57
Kato Y, Murakami S, Yamamoto Y, Chatani H, Kondo Y, Nakano T, Yokota A, Sato F (2004) The DNA-binding protease, CND41, and the degradation of ribulose-1,5-bisphosphate carboxylase/oxygenase in senescent leaves of tobacco. Planta 220:97–104
Kato Y, Miura E, Matsushima R, Sakamoto W (2007) White leaf sectors in yellow variegated2 are formed by viable cells with undifferentiated plastids. Plant Physiol 144:952–960
Khanna NC, Lakhani S, Tewari KK (1992) Identification of the template binding polypeptide in the pea chloroplast transcriptional complex. Nucleic Acids Res 20:69–74
Kinoshita I, Tsuji H (1984) Benzyladenine-induced increase in DNA content per chloroplast in intact bean leaves. Plant Physiol 76:575–578
Kleine T, Maier UG, Leister D (2009) DNA transfer from organelles to the nucleus: the idiosyncratic genetics of endosymbiosis. Annu Rev Plant Biol 60:115–138
Kobayashi T, Takahara M, Miyagishima S-Y, Kuroiwa H, Sasaki N, Ohta N, Matsuzaki M, Kuroiwa T (2002) Detection and localization of a chloroplast-encoded HU-like protein that organizes chloroplast nucleoids. Plant Cell 14:1579–1589
Kowallik KV, Herrmann RG (1972) Variable amounts of DNA related to the size of chloroplasts. IV. Three-dimensional arrangement of DNA in fully differentiated chloroplasts of Beta vulgaris L. J Cell Sci 11:357–377
Krause K, Krupinska K (2000) Molecular and functional properties of highly purified transcriptionally active chromosomes from spinach chloroplasts. Physiol Plant 109:188–195
Krause K, Falk J, Humbeck K, Krupinska K (1998) Responses of the transcriptional apparatus of barley chloroplasts to a prolonged dark period and to subsequent reillumination. Physiol Plant 104:143–152
Krause K, Kilbienski I, Mulisch M, Rodiger A, Schafer A, Krupinska K (2005) DNA-binding proteins of the Whirly family in Arabidopsis thaliana are targeted to the organelles. FEBS Lett 579:3707–3712
Krupinska K, Falk J (1994) Changes in RNA-polymerase activity during biogenesis, maturation and senescence of barley chloroplasts. Comparative analysis of transcripts synthesized either in run-on assays or by transcriptionally active chromosomes. J Plant Physiol 143:298–305
Kuroiwa T (1991) The replication, differentiation, and inheritance of plastids with emphasis on the concept of organelle nuclei. Int Rev Cytol 128:1–62
Kuroiwa T, Suzuki T (1981) Circular nucleoids isolated from chloroplasts in a brown alga Ectocarpus siliculosus. Exp Cell Res 134:457–461
Kuroiwa T, Suzuki T, Ogawa K, Kawano S (1981) The chloroplast nucleus: distribution, number, size, and shape, and a model for the multiplication of the chloroplast genome during chloroplast development. Plant Cell Physiol 22:381–396
Lam E, Hanley-Bowdoin L, Chua NH (1988) Characterization of a chloroplast sequence-specific DNA binding factor. J Biol Chem 263:8288–8293
Lamppa GK, Bendich AJ (1979) Changes in chloroplast DNA levels during development of pea (Pisum sativum). Plant Physiol 64:126–130
Lamppa G, Elliot L, Bendich A (1980) Changes in chloroplast number during pea leaf development. Planta 148:437–443
Landoulsi A, Malki A, Kern R, Kohiyama M, Hughes P (1990) The E. coli cell surface specifically prevents the initiation of DNA replication at oriC on hemimethylated DNA templates. Cell 63:1053–1060
Lawrence ME, Possingham JV (1986) Microspectrofluorometric measurement of chloroplast DNA in dividing and expanding leaf cells of Spinacia oleracea. Plant Physiol 81:708–710
Leutwiler LS, Hough-Evans BR, Meyerowitz EM (1984) The DNA of Arabidopsis thaliana. Mol Gen Genet 194:15–23
Li W, Ruf S, Bock R (2006) Constancy of organellar genome copy numbers during leaf development and senescence in higher plants. Mol Genet Genomics 275:185–192
Liere K, Börner T (2007) Transcription of plastid genes. In: Grasser KD (ed) Regulation of transcription in plants, vol. Blackwell Publishing, Oxford, pp 184–224
Lindbeck A, Rose R, Lawrence ME, Possingham JV (1987) The role of chloroplast membranes in the location of chloroplast DNA during the greening of Phaseolus vulgaris etioplasts. Protoplasma 139:92–99
Mache R, Lerbs-Mache S (2001) Chloroplast genetic system of higher plants: chromosome replication, chloroplast division and elements of the transcriptional apparatus. Curr Sci 80:217–224
Majeran W, Friso G, Asakura Y, Qu X, Huang M, Ponnala L, Watkins KP, Barkan A, van Wijk KJ (2012) Nucleoid-enriched proteomes in developing plastids and chloroplasts from maize leaves: a new conceptual framework for nucleoid functions. Plant Physiol 158:156–189
Maréchal A, Parent JS, Véronneau-Lafortune F, Joyeux A, Lang BF, Brisson N (2009) Whirly proteins maintain plastid genome stability in Arabidopsis. Proc Natl Acad Sci USA 106:14693–14698
Margulis L (1970) Origin of eukaryotic cells: evidence and research implications for a theory. Yale University Press, New Haven, Conn
Marrison JL, Rutherford SM, Robertson EJ, Lister C, Dean C, Leech RM (1999) The distinctive roles of five different ARC genes in the chloroplast division process in Arabidopsis. Plant J 18:651–662
Martin W (2003) Gene transfer from organelles to the nucleus: frequent and in big chunks. Proc Natl Acad Sci USA 100:8612–8614
Miyagishima S-Y (2011) Mechanism of plastid division: from a bacterium to an organelle. Plant Physiol 155:1533–1544
Miyamura S, Nagata T, Kuroiwa T (1986) Quantitative fluorescence microscopy on dynamic changes of plastid nucleoids during wheat development. Protoplasma 133:66–72
Miyamura S, Kuroiwa T, Nagata T (1990) Multiplication and differentiation of plastid nucleoids during development of chloroplasts and etioplasts from proplastids in Triticum aestivum. Plant Cell Physiol 31:597–602
Murakami S, Kondo Y, Nakano T, SATO F (2000) Protease activity of CND41, a chloroplast nucleoid DNA-binding protein, isolated from cultured tobacco cells. FEBS Lett 468:15–18
Nagata N, Saito C, Sakai A, Kuroiwa H, Kuroiwa T (1999) The selective increase or decrease of organellar DNA in generative cells just after pollen mitosis one controls cytoplasmic inheritance. Planta 209:53–65
Nakano T, Sato F, Yamada Y (1993) Analysis of nucleoid-proteins in tobacco chloroplasts. Plant Cell Physiol 34:873–880
Nakano T, Murakami S, Shoji T, Yoshida S, Yamada Y, SATO F (1997) A novel protein with DNA binding activity from tobacco chloroplast nucleoids. Plant Cell 9:1673–1682
Nemoto Y, Kawano S, Nakamura S, Mita T, Nagata T, Kuroiwa T (1988) Studies on plastid-nuclei (nucleoids) in Nicotiana tabacum L. I. Isolation of proplastid-nuclei from cultured cells and identification of proplastid-nuclear proteins. Plant Cell Physiol 29:167–177
Nemoto Y, Kawano S, Kondoh K, Nagata T, Kuroiwa T (1990) Studies on plastid-nuclei (nucleoids) in Nicotiana tabacum L. III. Isolation of chloroplast-nuclei from mesophyll protoplasts and identification of chloroplast DNA-binding proteins. Plant Cell Physiol 31:767–776
Nemoto Y, Kawano S, Nagata T, Kuroiwa T (1991) Studies on plastid-nuclei (nucleoids) in Nicotiana tabacum L. IV. Association of chloroplast-DNA with proteins at several specific sites in isolated chloroplast-nuclei. Plant Cell Physiol 32:131–141
Ogden GB, Pratt MJ, Schaechter M (1988) The replicative origin of the E. coli chromosome binds to cell membranes only when hemimethylated. Cell 54:127–135
Oldenburg DJ, Bendich AJ (2004) Changes in the structure of DNA molecules and the amount of DNA per plastid during chloroplast development in maize. J Mol Biol 344:1311–1330
Oldenburg D, Rowan B, Zhao L, Walcher C, Schleh M, Bendich A (2006) Loss or retention of chloroplast DNA in maize seedlings is affected by both light and genotype. Planta 225:41–55
Parent J-S, Lepage E, Brisson N (2011) Divergent roles for the two PolI-like organelle DNA polymerases of Arabidopsis thaliana. Plant Physiol 156:254–262
Pascoe MJ, Ingle J (1978) Distinction between nuclear satellite DNAs and chloroplast DNA in higher plants. Plant Physiol 62:975–977
Pfalz J, Liere K, Kandlbinder A, Dietz K-J, Oelmüller R (2006) pTAC2, -6 and −12 are components of the transcriptionally active plastid chromosome that are required for plastid gene expression. Plant Cell 18:176–197
Phinney BS, Thelen JJ (2005) Proteomic characterization of a triton-insoluble fraction from chloroplasts defines a novel group of proteins associated with macromolecular structures. J Proteome Res 4:497–506
Preuten T, Cincu E, Fuchs J, Zoschke R, Liere K, Börner T (2010) Fewer genes than organelles: extremely low and variable gene copy numbers in mitochondria of somatic plant cells. Plant J 64:948–959
Prikryl J, Watkins KP, Friso G, van Wijk KJ, Barkan A (2008) A member of the Whirly family is a multifunctional RNA- and DNA-binding protein that is essential for chloroplast biogenesis. Nucleic Acids Res 36:5152–5165
Pyke KA (1999) Plastid division and development. Plant Cell 11:549–556
Pyke KA, Leech RM (1992) Chloroplast division and expansion is radically altered by nuclear mutations in Arabidopsis thaliana. Plant Physiol 99:1005–1008
Pyke KA, Leech RM (1994) A genetic analysis of chloroplast division and expansion in Arabidopsis thaliana. Plant Physiol 104:201–207
Pyke KA, Rutherford SM, Robertson EJ, Leech RM (1994) arc6, a fertile Arabidopsis mutant with only two mesophyll cell chloroplasts. Plant Physiol 106:1169–1177
Rauwolf U, Golczyk H, Greiner S, Herrmann RG (2010) Variable amounts of DNA related to the size of chloroplasts III. Biochemical determinations of DNA amounts per organelle. Mol Genet Genomics 283:35–47
Reiss T, Link G (1985) Characterization of transcriptionally active DNA-protein complexes from chloroplasts and etioplasts of mustard (Sinapis alba L.). Eur J Biochem 148:207–212
Renner O (1934) Die pflanzlichen Plastiden als selbständige Elemente der genetischen Konstitution. Ber. sächs. Akad. Wiss. Leipzig, Math.-phys. K1 86:241–266
Ris H, Plaut W (1962) Ultrastructure of DNA-containing areas in the chloroplast of Chlamydomonas. J Cell Biol 13:383–391
Rowan BA, Bendich AJ (2009) The loss of DNA from chloroplasts as leaves mature: fact or artefact? J Exp Bot 60:3005–3010
Rowan BA, Oldenburg DJ, Bendich AJ (2004) The demise of chloroplast DNA in Arabidopsis. Curr Genet 46:176–181
Rowan B, Oldenburg D, Bendich A (2009) A multiple-method approach reveals a declining amount of chloroplast DNA during development in Arabidopsis. BMC Plant Biol 9:3
Sakai A, Saito C, Inada N, Kuroiwa T (1998) Transcriptional activities of the chloroplast-nuclei and proplastid-nuclei isolated from tobacco exhibit different sensitivities to tagetitoxin: implication of the presence of distinct RNA polymerases. Plant Cell Physiol 39:928–934
Sato N, Albrieux C, Joyard J, Douce R, Kuroiwa T (1993) Detection and characterization of a plastid envelope DNA-binding protein which may anchor plastid nucleoids. EMBO J 12:555–561
Sato N, Ohshima K, Watanabe A, Ohta N, Nishiyama Y, Joyard J, Douce R (1998) Molecular characterization of the PEND protein, a novel bZIP protein present in the envelope membrane that Is the site of nucleoid replication in developing plastids. Plant Cell 10:859–872
Sato M, Takahashi K, Ochiai Y, Hosaka T, Ochi K, Nabeta K (2009) Bacterial alarmone, guanosine 5′-diphosphate 3′-diphosphate (ppGpp), predominantly binds the β′ subunit of plastid-encoded plastid RNA polymerase in chloroplasts. Chembiochem 10:1227–1233
Scharff LB, Koop H-U (2007) Targeted inactivation of the tobacco plastome origins of replication A and B. Plant J 50:782–794
Scott NS, Possingham JV (1980) Chloroplast DNA in expanding spinach leaves. J Exp Bot 31:1081–1092
Scott NS, Possingham JV (1983) Changes in chloroplast DNA levels during growth of spinach leaves. J Exp Bot 34:1756–1767
Scott NS, Cain P, Possingham JV (1982) Plastid DNA levels in albino and green leaves of the ‘albostrians’ mutant of Hordeum vulgare. Z Pflanzenphysiol 108:187–192
Scott NS, Tymms MJ, Possingham JV (1984) Plastid-DNA levels in the different tissues of potato. Planta 161:12–19
Sekine K, Hase T, Sato N (2002) Reversible DNA compaction by sulfite reductase regulates transcriptional activity of chloroplast nucleoids. J Biol Chem 277:24399–24404
Shaver J, Oldenburg D, Bendich A (2006) Changes in chloroplast DNA during development in tobacco, Medicago truncatula, pea, and maize. Planta 224:72–82
Shaver JM, Oldenburg DJ, Bendich AJ (2008) The structure of chloroplast DNA molecules and the effects of light on the amount of chloroplast DNA during development in Medicago truncatula. Plant Physiol 146:1064–1074
Sheahan MB, McCurdy DW, Rose RJ (2005) Mitochondria as a connected population: ensuring continuity of the mitochondrial genome during plant cell dedifferentiation through massive mitochondrial fusion. Plant J 44:744–755
Siemenroth A, Wollgiehn R, Neumann D, Börner T (1981) Synthesis of ribosomal RNA in ribosome-deficient plastids of the mutant “albostrians” of Hordeum vulgare L. Planta 153:547–555
Sodmergen KS, Tano S, Kuroiwa T (1989) Preferential digestion of chloroplast nuclei (nucleoids) during senescence of the coleoptile of Oryza sativa. Protoplasma 152:65–68
Sodmergen KS, Tano S, Kuroiwa T (1991) Degradation of chloroplast DNA in 2nd leaves of rice (Oryza sativa) before leaf yellowing. Protoplasma 160:89–98
Steiner S, Schröder Y, Pfalz J, Pfannschmidt T (2011) Identification of essential subunits in the plastid-encoded RNA polymerase complex reveals building blocks for proper plastid development. Plant Physiol 157:1043–1055
Suck R, Zeltz P, Falk J, Acker A, Kössel H, Krupinska K (1996) Transcriptionally active chromosomes (TACs) of barley chloroplasts contain the α-subunit of plastome encoded RNA polymerase. Curr Genet 30:515–521
Sugiura M (1992) The chloroplast genome. Plant Mol Biol 19:149–168
Sugiura M (1995) The chloroplast genome. Essays Biochem 30:49–57
Terasawa K, Sato N (2005) Occurrence and characterization of PEND proteins in angiosperms. J Plant Res 118:111–119
Timmis JN, Ayliffe MA, Huang CY, Martin W (2004) Endosymbiotic gene transfer: organelle genomes forge eukaryotic chromosomes. Nat Rev Genet 5:123–135
Tymms MJ, Scott NS, Possingham JV (1983) DNA content of Beta vulgaris chloroplasts during leaf cell expansion. Plant Physiol 71:785–788
Wang D-Y, Zhang Q, Liu Y, Lin Z-F, Zhang S-X, Sun M-X, Sodmergen (2010) The levels of male gametic mitochondrial DNA are highly regulated in angiosperms with regard to mitochondrial inheritance. Plant Cell 22:2402–2416
Weber P, Fulgosi H, Piven I, Müller L, Krupindka K, Turini P (2006) TCP34, a nuclear-encoded response regulator-like TPR protein of higher plant chloroplasts. J Mol Biol 357:535–549
Wycliffe P, Sitbon F, Wernersson J, Ezcurra I, Ellerstrom M, Rask L (2005) Continuous expression in tobacco leaves of a Brassica napus PEND homologue blocks differentiation of plastids and development of palisade cells. Plant J 44:1–15
Yurina NP, Belkina GG, Karapetyan NV, Odintsova MS (1995) Nucleoids of pea chloroplasts: microscopic and chemical characterization. Occurrence of histone-like proteins. Biochem Mol Biol Int 36:145–154
Zheng Q, Oldenburg DJ, Bendich AJ (2011) Independent effects of leaf growth and light on the development of the plastid and its DNA content in Zea species. J Exp Bot 62:2715–2730
Zoschke R, Liere K, Börner T (2007) From seedling to mature plant: Arabidopsis plastidial genome copy number, RNA accumulation and transcription are differentially regulated during leaf development. Plant J 50:710–722
Acknowledgments
The work of the authors was supported by Deutsche Forschungsgemeinschaft (SFB 429). We are thankful to Reinhold G. Herrmann (Munich) and Hieronim Golczyk (Lublin) for providing Fig. 11.1.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Liere, K., Börner, T. (2013). Development-Dependent Changes in the Amount and Structural Organization of Plastid DNA. In: Biswal, B., Krupinska, K., Biswal, U. (eds) Plastid Development in Leaves during Growth and Senescence. Advances in Photosynthesis and Respiration, vol 36. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-5724-0_11
Download citation
DOI: https://doi.org/10.1007/978-94-007-5724-0_11
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-5723-3
Online ISBN: 978-94-007-5724-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)