
Abstract
The Purkinje cell is the pivotal element of the cerebellar network, which integrates distinct extracerebellar inputs and generates the ultimate cortical output to be conveyed to the deep cerebellar nuclei. During development, the adult Purkinje cell phenotype is acquired through a complex sequence of ontogenetic processes, including migration from the ventricular neuroepithelium to the cortex, formation of the Purkinje cell plate and progressive arrangement into the final monolayer, axonal growth and expansion of the dendritic tree. Most of the distinctive features of Purkinje cells can be achieved even in dissociated cultures, suggesting that the acquisition of adult traits is regulated by the unfolding of a cell-autonomous program. On the other hand, the maturing Purkinje cells play a fundamental role in orchestrating the development of the entire cerebellum. Namely, Purkinje cells are required for the genesis or the survival of different populations of cerebellar and extracerebellar neurons. They contribute to regulate the morphogenic processes leading to construct the cortical layering and network. They provide positional information to extracerebellar afferent systems, so to direct the topographic arrangement of projection maps. These phenomena and the underlying mechanisms are described and discussed in the chapter, proposing that development of Purkinje cells is not just the acquisition of a mature neuronal phenotype, but represents an essential organizational event of the whole cerebellar ontogenesis.
Access provided by Autonomous University of Puebla. Download reference work entry PDF
Similar content being viewed by others



Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Introduction
Purkinje cells are one of the neuronal populations more easily identified by their distinctive and specific morphology. These neurons were discovered by the Czech anatomist Jan Evangelista Purkinje in 1837 (Purkinje 1837), who reported their monolayer disposition. Almost 50 years later, Camillo Golgi (1883) with his silver impregnation method revealed the extent and spatial orientation of the dendritic arbor. The description was completed by Santiago Ramón y Cajal (1888), who reported the occurrence of small thorns or dendritic spines and their peculiar distribution on the distal dendritic compartment.
The acquisition of the Purkinje cell phenotype is accomplished through a sequence of ontogenetic processes, comprising migratory phases, morphogenic phenomena and attainment of mature neuronal structure and connectivity. In spite of the obvious similarities with the development of other neuron populations, some unique features distinguish the maturation of Purkinje cells. These neurons are the pivotal element of the cortical circuitry, which integrates inputs from different extracerebellar sources and relays the resulting information to the cerebellar nuclei (Ramón y Cajal 1911; Ito 2006). To carry out this functional task, Purkinje cells must occupy a central position in the structure of the cerebellar network, and this has profound implications for the developmental mechanism of the entire cerebellum. Indeed, prior of assuming their adult computational role, Purkinje cells carry out a fundamental ontogenetic function in orchestrating the construction of the cortical architecture and the emergence of intra- and extracortical connectivity.
Newborn Purkinje cells migrate from the ventricular to the pial surface and aggregate in a primordial cortical layer, the Purkinje cell plate, which subsequently evolves into the final monocellular alignment. During their navigation toward the cortex, Purkinje cells participate to complex reciprocal displacements and interactions with other populations of cerebellar neurons, leading to set up the fundamental framework of the cerebellar structure. Accordingly, the cortico-nuclear projection, with a clear topographic arrangement, is established before birth. During late embryonic and early postnatal life, intense cross-talk between Purkinje cells and the precursors of granule cells in the external granular layer regulates the proliferation and maturation of this quantitatively important type of cortical neurons, their progressive integration into functional networks and the concomitant shaping of the cortical layers. At the same time, specific interactions between Purkinje cells and afferent axons dictate the topographic organization of the extracerebellar afferent systems. The next sections of this chapter will be devoted to discuss the present knowledge of these phenomena, and of the underlying mechanisms, trying to define the main phases of Purkinje cell maturation and their influence on the development of the surrounding cerebellar structure.
The Generation of Purkinje Cells in the Ventricular Neuroepithelium
Purkinje cells are generated by progenitors residing in the ventricular neuroepithelium of the embryonic cerebellar primordium (Ramón y Cajal 1911; Altman and Bayer 1997). The mechanisms that regulate the proliferation of these progenitors and their commitment to become Purkinje cells are treated in several chapters of this Handbook (Chap. 2, “Proneural Genes and Cerebellar Neurogenesis in the Ventricular Zone and Upper Rhombic Lip”, Chap. 3, “Zones and Stripes: Development of Cerebellar Topography”, Chap. 4, “Roof Plate in Cerebellar Neurogenesis”, Chap. 5, “Specification of Cerebellar and Precerebellar Neurons”). Therefore, only a few aspects, which are relevant for the successive phases of migration and differentiation, will be briefly summarized here.
All cerebellar neurons derive from two germinal neuroepithelia: the upper rhombic lip is the origin of glutamatergic neurons, whereas the ventricular neuroepithelium produces GABAergic phenotypes (Hoshino 2006; Carletti and Rossi 2008). Purkinje cells are born at the outset of cerebellar neurogenesis (between E10.5 and E12.5 in the mouse; Miale and Sidman 1961; Altman and Bayer 1997), whereas inhibitory interneurons are generated at later stages (Carletti and Rossi 2008). The acquisition of adult phenotypic traits follows the biological age of the neurons. Hence, Purkinje cells do not mature synchronously, and different developmental stages can coexist not only in the same lobule, but even among neighboring neurons.
The progenitors that proliferate in the cerebellar ventricular neuroepithelium are distinguished by the expression of the bHLH transcription factor Ptf1a (Obata et al. 2001; Hoshino et al. 2005; Hoshino 2006). Ptf1a appears not only to be required for the specification and the acquisition of the GABAergic phenotype (Hoshino et al. 2005), but also to prevent commitment toward glutamatergic lineages (Pascual et al. 2007). Downstream targets of Ptf1a include Neurogenin2 (Henke et al. 2009), Nephrin, and Neph3 (Nishida et al. 2010). Neurogenin2, together with neurogenin1, are transiently expressed in restricted domains of the ventricular neuroepithelium during the time-window of Purkinje cell genesis (Zordan et al. 2008). This spatio-temporal distribution pattern suggests that neurogenins are involved in the specification of phenotypically heterogeneous Purkinje cell subsets, which will be responsible to lay down the fundamental framework of the cerebellar topography (see below). On the other hand, Nephrin and Neph3 are cell surface molecules that may be important for the delamination of postmitotic neurons from the germinal neuroepithelium. Neph3 is equally expressed by all ventricular progenitors and their early-postmitotic derivatives in the subventricular zone. In the latter site, however, newborn Purkinje cells are distinguished from interneurons by differential expression of E-cadherin (Mizuhara et al. 2010) and of the transcriptional corepressor Corl2 (Minaki et al. 2008). The patterned regulation of these markers highlights distinct domains of the ventricular neuroepithelium and of the subventricular zone, corresponding to the sites of origin of the different GABAergic phenotypes. Under a functional point of view, however, these observations indicate that specific combinations of transcription factors and surface molecules regulate the sorting of different cell subtypes from the ventricular neuroepithelium. Although the precise role of these molecules is still unknown, it is likely that these heterogeneous neurochemical profiles modulate the sensitivity of the juvenile neurons to extrinsic cues that induce migratory behavior and trigger the unfolding of differentiation programs.
Migration of Purkinje Cells
Migration Toward the Cerebellar Cortex and Formation of the Purkinje Cell Plate
Newborn Purkinje cells leave the ventricular neuroepithelium to reach their final destination in the cerebellar cortex. At the stage when Purkinje cells initiate their migration, the superficial region of the cerebellar anlage is occupied by deep nuclear neurons (the so-called nuclear transitory zone), which have proliferated somewhat earlier (Miale and Sidman 1961; Altman and Bayer 1997) and are moving tangentially in the rostral direction, from the rhombic lip along a subpial stream (Machold and Fishell 2005; Fink et al. 2006). This phase, between E14 and E17, is characterized by a complex balance movement of Purkinje cells, that “ascend” toward the pia, and nuclear neurons, that “descend” toward the depth of the cerebellar primordium (Fig. 9.1; Altman and Bayer 1997; Bourrat and Sotelo 1986). The migratory route of Purkinje cells eventually terminates close to the pial surface, which is being occupied by the nascent external granular layer (Fig. 9.1). Here, Purkinje cells form a three–four- cell-thick aggregate, the Purkinje cell plate, which is first visible around E14.5 (Miyata et al. 2010) and will evolve into the final monocellular arrangement during the first postnatal week (Altman and Bayer 1997).


Migration of Purkinje cells to the cerebellar cortex. The classical view of Purkinje cell migration is displayed on the upper row. Newborn Purkinje cells (blue) migrate radially, along the processes of radial glia (red), from the ventricular neuroepithelium (VN) to the pial surface (PS), attracted by Reelin secreted by glutamatergic neurons of the cerebellar nuclei (green) that still occupy a subpial position. During this period, the nuclear neurons move toward their final location in the deep cerebellar nuclei (DCN), as indicated by the green dashed arrow. At the end of the process, Purkinje cells form the Purkinje cell plate (blue shading on the leftmost panel) underlining the nascent external granular layer (represented by the green cells on the leftmost panel). The new view, depicted in the lower row, proposes that Purkinje cells born in the posterior region of the cerebellar anlage first migrate tangentially, parallel to the pial surface. Once arrived at destination, they modify the posture of their cell bodies and initiate a second phase of radial migration, leading to form the Purkinje cell plate (Modified from Miyata et al. (2010))
Given the straight ventricular-to-pial direction of the route to the cortex, Purkinje cell migration has been classically considered to follow a strictly radial course along the processes of radial glia (Fig. 9.1; Altman and Bayer 1997). Evidence for this mechanism has been provided by numerous studies, which demonstrated direct apposition of young Purkinje cells to radial glia, elucidated functionally relevant cellular and molecular interactions, and highlighted differential time schedules of Purkinje cell migration in distinct cerebellar subdivisions (Miale and Sidman 1961; Rakic and Sidman 1970; Goffinet 1983; Yuasa et al. 1991, 1996; Hatten and Heintz 1995; Altman and Bayer 1997; Hatten 1999; Sotelo 2004).
In addition to the conducive scaffold provided by radial glia, the navigation of Purkinje cells is thought to be directed by the attractive action exerted by the glycoprotein Reelin (Miyata et al. 1997; Gilmore and Herrup 2000; Rice and Curran 2001), secreted by “overlying” subpial structures, including first the nuclear transitory zone and, later, the external granular layer (Miyata et al. 1996; Schiffmann et al. 1997). The Purkinje cell plate is located adjacent to latter structure, suggesting that Reelin is also important to arrest Purkinje cell migration and regulate the ensuing layering process (Miyata et al. 1997; Carletti et al. 2008). These notions are further supported by the cerebellar phenotype of reeler mice, as well as other neurological mutants that disrupt Reelin signaling, in which Purkinje cells fail to achieve their normal cortical allocation and remain clustered close to the region of the cerebellar nuclei (Mariani et al. 1977; see Rice and Curran 2001 for a review).
The notion that Purkinje cell migration follows a strict monophasic radial route has been recently challenged (Miyata et al. 2010). By adenoviral labeling in utero and time-lapse analysis on organotypic cultures of the mouse cerebellum, these authors propose that Purkinje cells born at E10.5 at the posterior edge of the ventricular neuroepithelium actually follow a tangential migratory route (Fig. 9.1). Between E12 and E14, these neurons navigate anteriorly, parallel to the pial surface, but deeper than the subpial migratory stream followed by rhombic lip derivatives (i.e. deep nuclei neurons; Altman and Bayer 1997; Sotelo 2004; Machold and Fishell 2005; Fink et al. 2006). Around E14.5, the Purkinje cells terminate their caudo-rostral navigation and undergo a profound morphological rearrangement (Fig. 9.1; Miyata et al. 2010): the leading process points toward the inner regions of the cerebellar primordium, the cell body modifies its posture to assume a radial orientation, and the trailing appendages are reshaped to leave a single process, directed perpendicular to the pial surface. The latter neurite, which directly abuts the Reelin-rich subpial region, takes the role of the leading process in a new radial migratory phase that will drive the neuron into the Purkinje cell plate.
This complex sequence of morphogenic and migratory phenomena implies that Reelin signaling becomes particularly important during the last phase, when Purkinje cells modify their orientation to switch from tangential to radial migration. Indeed, in the reeler cerebellum tangential migration is unaffected, whereas the ensuing reorientation of Purkinje cells completely fails (Miyata et al. 2010). Interestingly, a similar mechanism has been demonstrated in the olfactory system, where Reelin induces neuroblasts of the rostral migratory stream to switch from tangential chain migration to radial individual migration (Hack et al. 2002).
These findings suggest that Purkinje cell migration toward the cerebellar cortex is more complex than previously envisaged. Nonetheless, several issues remain to be elucidated. For instance, it may be possible that the tangential phase is also trailed by radial glia, whose processes in the posterior part of the cerebellar anlage have an oblique orientation. In addition, it is still unknown whether the same phenomena also apply for Purkinje cells derived from anterior regions of the ventricular neuroepithelium. Finally, the proposed mechanism implies that Reelin exerts its function through short-range interactions rather than as a long-distance positional cue. Appropriate operation of such a mechanism requires a precise spatio-temporal distribution of Reelin signaling in the nascent cerebellar cortex. However, this cannot be immediately reconciled with the observation that large numbers of Purkinje cells achieve a correct cortical allocation after complete ablation of the external granular layer (Jensen et al. 2002), or when Reelin is ectopically expressed in reeler cerebella (Magdaleno et al. 2002). Notably, in these conditions the Purkinje cell plate forms normally and only the subsequent layering of Purkinje cells is impaired.
On the whole, Purkinje cell migration is influenced by complex interactions with different structures (Magdaleno et al. 2002; Sotelo 2004). The initial targeting to the cerebellar cortex is achieved at early ontogenetic stages and is likely controlled by signals issued from the nuclear transitory zone. Cues from the external granular layer, however, become relevant at later stages to direct the final arrangement of Purkinje cells and the ensuing layout of the cortical cytoarchitecture. Except for the fundamental role played by Reelin in these processes, the relevant molecular signaling is largely unknown.
Formation of the Purkinje Cell Monolayer
Successive waves of Purkinje cells progressively accumulate in the Purkinje cell plate throughout embryonic development. After birth, together with the expansion of the cortical lobules, the plate gradually evolves into the final monocellular arrangement, which is definitively achieved at about the end of the first postnatal week in rodents (Altman and Bayer 1997). The emergence of the Purkinje cell monolayer has been attributed to mechanical constraints exerted by the expanding internal granular layer and the simultaneous stacking of parallel fibers in the molecular layer (Altman and Winfree 1977; Altman and Bayer 1997). In spite of the attractive simplicity of this mechanism, which is partially corroborated by the characteristic misalignment of Purkinje cells in hypogranular cerebella (see Altman and Bayer 1997), this process likely involves other structures and interactions. For instance, in the cerebellum of math-1-null chimeras, characterized by a patchy distribution of the external granular layer, Purkinje cells fail to align in the cortical regions where this layer is absent (Jensen et al. 2004). In addition, experiments in which the geometry of Reelin signaling is artificially manipulated show that Purkinje cell alignment can be induced by creating a Reelin-rich zone overlying the Purkinje cell plate (Miyata et al. 1997), whereas it is defective if the protein is released by cells located deep in the cerebellar tissue (Magdaleno et al. 2002). On the whole, these observations indicate that during these phases of cortical histogenesis Purkinje cells continue to advance radially toward the pial surface to match the concomitant expansion of the outgrowing cortex (Fig. 9.2). This movement, however, must be synchronous in order to prevent the dispersal of Purkinje cells that would hamper the proper development of the cortical layers.


Formation of the Purkinje cell monolayer. The cartoon depicts the progressive movement of Purkinje cells (blue), required to match the outgrowth of the whole cerebellar mass, during late embryonic and early postnatal development. It is proposed that Purkinje cells undergo a progressive radial displacement, regulated by the overlying external granular layer (green). This process induces a gradual thinning of the Purkinje cell plate, leading to the formation of the monolayer at about the end of the first postnatal week
Heterochronic transplantation experiments indicate that synchronicity of Purkinje cell migration, and notably the timing of arrival in the cortex, is a relevant factor to determine the correct alignment (Carletti et al. 2008). Embryonic Purkinje cells grafted to P1 or P8 cerebella are still able to integrate in the recipient cortex by achieving the correct polarity and orientation. These neurons also have typical monoplanar dendrites extended up to the pial surface like their neighboring counterparts. Nevertheless, their cell bodies are ectopically positioned in the recipient molecular layer, and the extent of this displacement is proportional to the delay of their arrival to the cortex (i.e. Purkinje cells grafted to P8 cerebella occupy more superficial positions than Purkinje cells grafted to P1 recipients). The same phenomenon occurs when rat Purkinje cells are homochronically grafted to the embryonic mouse cerebellum, which develops at a faster pace (Carletti et al. 2008). In all these conditions, the late arriving Purkinje cells, that reach the cortex when the molecular layer is already expanding, bypass their natural position and terminate their course only when they come in contact with the external granular layer. Therefore, this layer provides short-range signals that direct the slow radial progression of Purkinje cells required to match the cortical outgrowth and, at the same time, restrain this movement in order to maintain the correct alignment. In this way, Purkinje cells are continuously attracted toward the pial surface of the expanding cortex and arranged into the final monolayer, thanks to the progressive thinning of the Purkinje cell plate (Fig. 9.2).
Development of the Purkinje Axon
The axon of the Purkinje neuron comprises two major domains corresponding to the main target fields: (1) the corticofugal axon projecting to the deep cerebellar nuclei and to a number of target nuclei in the brainstem (De Camilli et al. 1984); (2) the intracortical plexus formed by recurrent collateral branches that terminate within the cerebellar cortex (Ramón y Cajal 1911). The two compartments develop at distinct ontogenetic stages. The stem Purkinje axons elongates during embryonic life, concomitant with the migration toward the cortex. The intracortical branches are formed during postnatal development, after the establishment of the cortico-nuclear projection.
Development of the Corticofugal Purkinje Axon
The initial phases of Purkinje axonogenesis remained largely obscure up to recent times because of the lack of efficient tools to visualize and identify single Purkinje cells with their processes. The classical view postulates that the development of the neurite parallels the radial migration of newborn Purkinje cells from the ventricular neuroepithelium to the pial surface. In this process, the corticofugal axon originates as the trailing process of the migrating neuron, which is left behind to take contact with the deep nuclei (Fig. 9.1; Altman and Bayer 1997). Accordingly, the cortico-nuclear connections are established around birth (Eisenman et al. 1991), and synaptic responses can be recorded from deep nuclear neurons shortly after (Gardette et al. 1985).
This view has been recently questioned by the analysis of individual Purkinje cells in vivo and in vitro (Fig. 9.1; Miyata et al. 2010). According to this study, during the tangential phase of Purkinje cell migration, the leading process is actually the future axon, which is also distinguished by the expression of axon-specific markers, such as neurofilaments and tau. Later, when the Purkinje cell modifies the posture of the perikaryon to enter the Purkinje cell plate, the axon is directed deep toward the cerebellar nuclei, whereas the prospective dendrite takes the role of the leading, pially directed process. Therefore, axonogenesis starts very soon after the birth of the Purkinje cell, and the neurite actively navigates toward its target rather than lagging behind the parent neuron, as proposed by Miale and Sidman (1961). The latter authors, by describing an inexistent crossing of the nuclear transitory mass by Purkinje cells, hypothesized that the early segregation of the deep nuclear cells imposed the further segregation of Purkinje cells into clusters. The observations of Miyata et al. (2010) are consistent with additional evidence obtained by genetic tagging of developing mouse Purkinje cells in situ (Sillitoe et al. 2009). The latter report shows that axonogenesis starts a few hours or days after the birth of the Purkinje cell. In addition, a clear cortico-nuclear projection, with an evident topographic arrangement, is already detectable at E14.5 and is firmly established 3 days later.
The age when these connections become functional is still unknown. Similarly, very little is known about the molecular cues that guide the nascent Purkinje axons toward their nuclear targets. The initial phases of axonogenesis are normal in reeler mice (Miyata et al. 2010), ruling out the possibility that Reelin-mediated attraction is involved in Purkinje axon guidance. Recently, it has been observed that Unc5c is strongly expressed in cerebellar progenitors, particularly in deep nuclear and Purkinje cell progenitors. From E13.5 onwards, the protein is expressed also in their axons, in an adjacent area to the “nuclear transitory zone,” down to the hook bundle, suggesting that netrin1 could participate to the guidance of these axons (Kim and Ackerman 2011). The study of Unc5c−/− mice corroborates that this netrin receptor may repel the hook bundle from the floor plate, even though netrin1 alone does not seem to be enough and that other unknown ligands should be also at play (Kim and Ackerman 2011). In any instance, the precocious interactions between specific Purkinje cell subsets and defined nuclear regions imply that the cortico-nuclear topography is established before Purkinje cells reach their final cortical allocation (Sillitoe et al. 2009). As a consequence, it is likely that the guidance of outgrowing Purkinje neurites is regulated by short-range axon–target interactions rather than long-distance attractive cues.
It has to be emphasized that classical growth cone-driven path finding is not the only mechanism that sustains the developmental growth of Purkinje stem axons. In fact, at the ontogenetic stage when the cortico-nuclear connection is established the cerebellum is still far from reaching its final size. As a consequence, most of the length of the adult Purkinje axons is actually due to interstitial elongation, which occurs postnatally under the mechanical stretching exerted by the outgrowing tissue (Rossi et al. 2007; de Luca et al. 2009). Because of the fast postnatal expansion of the cerebellar mass, the rate of Purkinje axon elongation observed during the first week after birth (up to over 200 μm/day) is about twofold than that observed during embryonic development (de Luca et al. 2009).
Development of the Intracortical Plexus of the Purkinje Axon
The development of the intracortical Purkinje axon was first described by Ramón y Cajal (1911), who reported the initial redundancy of these branches and their subsequent pruning to achieve the final arrangement of the infra- and supra-ganglionic plexus. More recently, it has been shown that recurrent collaterals sprout as interstitial ramifications of the corticofugal axon during the first postnatal week (Gianola et al. 2003). Multiple processes bud from a moderately swollen segment of the stem neurite, situated along its course across the granular layer. The site of branch sprouting is characteristically located at 150–200 μm from the cell body (Gianola et al. 2003), corresponding to the future position of the first node of Ranvier (Clark et al. 2005). Interestingly, this mode of collateral branching is also maintained when isolated Purkinje cells are grown in culture (Fig. 9.3; Gianola et al. 2003), suggesting that collateral branches originate from defined neuritic segments, specifically primed for this task (see also Szebenyi et al. 1998).


Development of Purkinje axon in vitro. The confocal images show three examples of dissociated Purkinje cells maintained for 2, 4 or 21 days in vitro (DIV). Note that the growth of the axon progresses according to the same phases of axonogenesis in vivo, leading to develop a prominent terminal tree and recurrent collateral arbors (Modified from Gianola et al. (2003))
The newly formed branches ascend toward the Purkinje cell layer and develop large terminal networks, extended both in the granular and in the molecular layer (Gianola et al. 2003). The recurrent ramifications are preferentially directed toward the base of the lobule and innervate nearby Purkinje cells, thus forming a chain of functionally connected neurons running from the apex to the base of the lobule (Watt et al. 2009). These GABAergic synapses, which are initially excitatory and become inhibitory at a later time (Orduz and Llano 2007), drive activation waves that travel through the network of Purkinje cells along the main axis of the folium (Watt et al. 2009). The functional meaning of these waves is still unknown, but they are likely involved in activity-dependent mechanisms, required for the maturation of the cerebellar cortical circuitry.
During the second postnatal week, the majority of collateral branches and terminal ramifications are retracted to shape the adult neuritic arrangement (Gianola et al. 2003). At the end of this process, each corticofugal axon retains only one or two recurrent branches, whose terminal arbors are precisely confined within the upper granular or lower molecular layer. This remodeling also involves a profound reorganization of connectivity, including the withdrawal of most Purkinje-to-Purkinje synapses and the consequent disappearance of the traveling waves of activity (Watt et al. 2009). In the adult mice, recurrent Purkinje axons primarily contact interneurons in the upper granular or lower molecular layer (Palay and Chan-Palay 1974; King and Bishop 1982; Watt et al. 2009).
The remodeling of intracortical Purkinje axons is coincident with myelination, which progresses in a retrograde manner from the central white matter toward the cerebellar cortex and is completed at the end of the second postnatal week (Reynolds and Wilkins 1988; Bouslama-Oueghlani et al. 2003; Gianola et al. 2003). Procedures that block or delay myelinogenesis, such as ablation of oligodendrocytes or application of neutralizing antibodies against the myelin-associated growth-inhibitory protein Nogo-A, impair the pruning of Purkinje intracortical plexus, which fail to achieve the normal adult pattern (Gianola et al. 2003). Indeed, intracortical Purkinje axons express receptors for myelin-associated inhibitory proteins (Foscarin et al. 2009), indicating that environmental regulatory molecules contribute to shape and maintain the typical laminar distribution of the Purkinje intracortical plexus (Buffo et al. 2000; Rossi et al. 2006).
Intrinsic Mechanisms and Environmental Control of Purkinje Axon Development
The major phases of Purkinje cell axonogenesis described above are faithfully reproduced when these neurons are transplanted to extracerebellar regions or cultured as dissociated cells (Fig. 9.3; Gianola et al. 2003, 2009; de Luca et al. 2009). Accordingly, the main compartments of the Purkinje axon, i.e. the stem neurite with its terminal arbor and the collateral branches with the recurrent plexus, are developed even in such ectopic conditions. This suggests that the Purkinje axon phenotype is coded by an intrinsic program, which is unfolded by the developing neuron regardless of the surrounding environment.
The spontaneous growth properties of Purkinje cells change according to their age (de Luca et al. 2009). Neurons taken from E17 embryos and plated in dissociated cultures elongate their stem axon and appear less inclined to break into terminal ramifications. In contrast, neurons isolated at P1 grow at a faster rate and, after a period of stem axon elongation, develop extensive arbors. Most embryonic Purkinje cells are unable to switch spontaneously from the elongating to the arborizing mode of neuritic growth, even after 2 weeks in vitro. Nevertheless, they can be readily induced to form large terminal arbors by co-culturing with granule cells, suggesting that the execution of the intrinsic growth program is regulated by extrinsic signals.
Other environmental cues also influence the outgrowth of collateral and terminal branches. As mentioned above, the processes that induce the budding of collateral branches in vivo are essentially replicated when Purkinje cells are placed in dissociated cultures (Gianola et al. 2003). Nevertheless, the number of ramifications issued from each neurite can be influenced by addition of external factors. For instance, application of Anosmin-1 to the culture medium increases the number of collateral branches and favors the expansion of terminal arbors, whereas incubation with neutralizing antibodies reverses these effects (Gianola et al. 2009). In the developing cerebellum, Purkinje cells and deep nuclear neurons express Anosmin-1, suggesting that it may act as a target-derived branch-inducing factor, as shown in the olfactory system (Soussi-Yanicostas et al. 2002). Altogether, these observations indicate that the main distinctive traits of the Purkinje axon are determined by the endogenous properties of the parent neuron and are expressed in any milieu. The execution of the intrinsic neuritogenic program, however, is influenced by environmental signals that modulate the growth competence of the Purkinje neurons during different phases of axonogenesis.
Development of the Purkinje Dendritic Tree
In addition to the axon, Purkinje cells in dissociated cultures also develop dendritic trees with typical morphological features. This distinctiveness makes Purkinje cells excellent models for depicting intrinsic versus extrinsic mechanisms implied in dendritogenic processes. Here, the arguments will be discussed that lead to the conclusion that Purkinje cell dendritic differentiation happens in two main phases: an early one characterized by intensive and continuous remodeling, which is mainly controlled by intrinsic factors, and a late one required for the acquisition of the ultimate dendritic pattern, which depends both on intrinsic and extrinsic factors, represented by hormones, neurotrophins and, interactions with afferent axons (notably parallel fibers).
Sequential Phases of Dendritic Differentiation
During the last phase of radial migration, Purkinje cells adopt a bipolar elongated shape, and it is only when they settle in the Purkinje cell plate that the real development of the dendritic tree starts. This process, that involves a profound remodeling of the leading process of the radially migrating neuron, is typically characterized by a number of distinct phases (Fig. 9.4; Armengol and Sotelo 1991):
-
1.
“Simple-fusiform cells”: from the end of migration up till the day of birth (P0); the cells keep their bipolar shape even though apical dendrites may branch.
Fig. 9.4 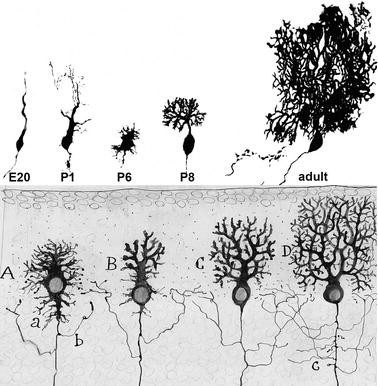
Development of the Purkinje cell dendritic tree. Upper half: Purkinje cells from rat cerebellum taken at different stages of dendritic development, from embryonic day 20 (E20) to adulthood (around P60). The cells have been filled with HRP either by retrograde labeling (E20–P6) or by intracellular iontophoretic injection (P8 to adult). These micrographs illustrate the early phases of dendritic remodeling passing progressively from “simple-fusiform” cells at the end of the migration into the Purkinje cell plate (E20), to “complex-fusiform” cells (P1) and regressively to “stellate cell with disoriented dendrites” (P6). The ultimate dendritic arborization, with the flattening of the dendrites (P8) and the acquisition of the espaliered tree (adult) occurs during the second and third postnatal weeks. Lower half: Composite drawing by Santiago Ramon y Cajal illustrating the development of Purkinje cell dendrites. Cajal has only described the second phase, from Purkinje cells with stellate shape and disoriented dendrons (A), through the phase of young cell with flattened trees (B, C), to the adult espaliered tree (D). The material for upper micrographs was taken from: Armengol and Sotelo 1991 (E20–P6), P1 corresponds to Fig. 9.4C, and P6 to Fig.6F; Crepel et al. 1980 (P8 to adult), P8 corresponds to Fig. 9.4, and adult to Fig. 10. The Cajal drawing is reproduced with permission from Legado Cajal. Instituto Cajal (CSIC). Madrid (Spain)
-
2.
“Complex-fusiform cells”: the bipolar elongated cells with branched apical dendrites start generating new and thinner processes that radiate from the cell bodies. Coincidentally, there is a regression in length of the primitive apical dendrite.
-
3.
“Regressive-atrophic dendrites”: this phase, between P2 and P3-P4, is characterized by a fast and intense pruning of all or most of the dendritic processes. This regression phase announces the beginning of the building up of the ultimate dendritic arbor.
-
4.
The Ramón y Cajal “stellate cells with disoriented dendrites.” This phase, the first recognized by Ramón y Cajal (1890) (Fig. 9.4), is characterized by the explosive outgrowth of perisomatic protrusions emerging in all directions. It is during this phase that Purkinje cell bodies reach their monolayer disposition. The processes arising from the lateral and basal aspects of the cell bodies are rapidly withdrawn to leave only one to three primary segments arising from the apical pole. The loss of lateral processes is partially compensated by the appearance of numerous thin spine-like pseudopodia, the postsynaptic elements for the climbing fibers in their perisomatic nest stage (Larramendi and Victor 1967; Chedotal and Sotelo 1992).
-
5.
The Ramón y Cajal “young Purkinje cells with flattened dendritic trees”: characterized by the vertical orientation of their apical dendritic tree (P6–P10), followed by its lateral growth (P10–P15), reaching the adult width by P13. From P15 to P30 there is an important change in the orientation of the growth plane, from transverse to vertical, which brings a significant extension in height of the dendritic tree (Berry and Bradley 1976).


In summary, Purkinje cells pass through consecutive constructive and recessive developmental phases to acquire the adult configuration of their dendritic arbors. The intrinsic versus extrinsic determinants involved in the building up of either the first postnatal steps (from simple-fusiform stage to the regressive atrophic dendrites), or the second phase (from regressive and stellate with disoriented dendrites to flattened dendritic trees) will be discussed hereafter.
Intrinsic Determinants Regulate the First Phase of Purkinje Cell Dendritogenesis
The reshaping of Purkinje cell dendrites occurs simultaneously with synaptogenesis, either by receiving olivocerebellar afferents (Mason et al. 1990; Chedotal and Sotelo 1992, 1993; Morara et al. 2001; Sotelo 2008), or contacting their postsynaptic targets in the deep nuclei (Gardette et al. 1985). In living animals, Purkinje cells are embedded in an intricate environment, and are constantly interacting with astrocytic processes, afferent or efferent inputs, which complicates the identification of the relative contribution of intrinsic and extrinsic factors in dendritogenesis. An optimal way to distinguish between such different influences is to analyze the development of isolated Purkinje cells in vitro. In spite of the poor survival of P0 Purkinje cells in purified cultures (Baptista et al. 1994), the neurons that survive for 21 days in vitro remain arrested in their dendritic differentiation and do not generate identifiable dendrites. Most importantly, however, at 4 days in vitro Purkinje cells have much longer, smoother and less branched arbors than at 21 days, suggesting that the pruning or regressive process, occurring between P2 and P5 in vivo, also takes place in purified Purkinje cell cultures (see Figures 9.4A, B in Baptista et al. 1994).
Elimination of all climbing fibers and most of the mossy fibers by transection of the inferior cerebellar peduncle in newborn rats in vivo provokes some changes, but does not alter substantially the mature architecture of Purkinje cell dendrites (Sotelo and Arsenio-Nunes 1976). Therefore, removal of extracerebellar afferents does not disrupt the overall course of Purkinje cell dendrite maturation. Similarly, the absence of extracerebellar afferents in organotypic cultures does not prevent Purkinje cells to go through the same dendritic developmental phases as those described in vivo (Boukhtouche et al. 2006; Poulain et al. 2008). Finally, quantitative analysis of segment length, bifurcation ratio and topology reveals that the growth of Purkinje dendrites proceeds normally until P7 in the staggerer and weaver cerebella, both devoid of granule cells (Bradley and Berry 1978). Thus, the important remodeling process occurring in early dendritogenesis seems to occur in the absence of parallel and even climbing fibers, being mainly regulated by intrinsic factors.
Role of ROR-Alpha in the Formation of Purkinje Cell Dendrites
The study of the staggerer cerebellum has provided a first attempt to identify transcription factors that regulate the early phases of Purkinje dendrite formation. The molecular target of the staggerer mutation is the Retinoid-Related Orphan Receptor alpha (RORα) (Hamilton et al. 1996). As expected, transgenic mice with inactivation of the RORα gene share their phenotype with staggerer mice (Steinmayr et al. 1998). In this mutant, the cerebellum is highly atrophic and the number of remaining Purkinje cells at birth is only about 25% of the normal. The few surviving Purkinje cells exhibit dendrites smaller than normal, not confined to a single plane, and devoid of spiny branchlets, while the vast majority of granule cells have either missed their proliferation or have died (Sotelo and Changeux 1974). More importantly, Purkinje cells display a wide spectrum of shapes, ranging from unipolar to multipolar, which have been considered as remnants of the immature stage: the unipolar Purkinje cells resembling “simple-fusiform” cells and the multipolar ones represent “complex-fusiform” cells, in which the normal process of regression of early somatic protrusions fails to occur and instead they are transformed into long-lasting dendritic branches.
More direct evidence in favor of the implication of RORα in the regressive events during Purkinje cell dendritogenesis has been obtained in organotypic cultures, using a recombinant nonreplicative lentiviral-derived vector encoding either hRORα1 (Lenti-hRORα1) or Green Fluorescent Protein (for control), under the control of the phospho-glycerate kinase promoter (Boukhtouche et al. 2006). The lentiviral-mediated hRORα1 overexpression in fusiform Purkinje cells from P0 wild-type mice induces an accelerated progression of dendritic differentiation, leading to reach the mature pattern already at 7 days in vitro. In addition, while in control slices at 3 days in vitro, almost all Purkinje cells are still fusiform (94.3%), 58.8% of hRORα1-overexpressing neurons have already reached the regressive atrophic stage. Remarkably, RORα is necessary for the regressive events, because staggerer RORα-deficient Purkinje cells, which remain in the embryonic fusiform stages in nontransfected slices from RORα knock-out mice, complete normal dendritogenesis in transfected cultures (Boukhtouche et al. 2006). Therefore, RORα expression in fusiform Purkinje cells is crucial for dendritic regression.
It is likely that RORα controls a set of genes that interact with the cytoskeleton. SCLIP, a phosphoprotein member of the stathmin family interacting with microtubules (Curmi et al. 1999), might be one of the links between RORα and the cytoskeleton, since it also influences the growth of Purkinje cell dendrites. As revealed with a similar lentiviral-mediated approach on cerebellar organotypic cultures, depletion of SCLIP promotes primitive dendrite regression and inhibits further dendrite formation (Poulain et al. 2008). On the whole, the above reviewed evidence emphasize that the early phase of continuous outgrowth of the dendritic tree, with the stemming of multiple primary branches and the subsequent regressive phase of almost complete pruning, is controlled by intrinsic factors, among which RORα is the first to be known.
Extrinsic and Intrinsic Factors That Control the Late Phase of Purkinje Dendritic Growth
The late phase of Purkinje cell differentiation, with the progressive disappearance of the somatic processes and the rapid outgrowth and branching of the ultimate dendrites, is the one that provides these neurons with their characteristic espaliered dendritic trees. Although intrinsic factors contributing to the acquisition of this complex and original shape remain largely unknown, the number of identified extrinsic factors has largely increased over the last few years. Among them, afferent inputs (mainly parallel fibers, even though climbing fibers also contribute to the dendritic sculpting; Bradley and Berry 1976), trophic factors and hormones are the principal regulators of the orientation and growth of mature dendritic trees (Kapfhammer 2004; Sotelo and Dusart 2009). In addition, the neurotransmitter glutamate also exerts multiple actions. Activation of NMDA receptors promotes granule cell survival through a BDNF-mediated mechanism (Hirai and Launey 2000). This increases the interaction between granule and Purkinje cells, thus increasing the complexity of the dendritic tree. On the contrary, activation of metabotropic glutamate receptors (particularly those of class I) provokes a strong inhibition of the dendritic growth, generating Purkinje cells with small stubby dendrites (Sirzen-Zelenskaya et al. 2006). More recently, it has been shown that treatment with the GABAA agonists muscimol provided Purkinje dendrites with longer and less branched morphology, while GABAA antagonists resulted in shorter dendrites, in an endocannabinoid-dependent manner (Kawaguchi et al. 2010). Finally, the Calcitonin gene-related peptide stimulates the growth of Purkinje dendrites in culture through a mechanism that involves astrocytes (D’Antoni et al. 2010).
Several genes have been disclosed that influence the last phase of Purkinje dendrite growth. For instance, a deletion in exon 7 of Nna1 gene, which encodes a nuclear protein containing a zinc carboxypeptidase domain, is the genetic alteration responsible for the spontaneous “Purkinje cell degeneration” (pcd) mutant mouse (Fernandez-Gonzalez et al. 2002). In this mutant, Nna1 dramatically increases intranuclear localization of lysyloxidase propeptide, which provokes the underdevelopment of Purkinje cell dendrites by interfering with NF-kB/RelA signaling and microtubule-associated protein regulation of microtubule stability (Li et al. 2010). In Drosophila, Flamingo (a 7-pass transmembrane cadherin) is known for its prominent role in the growth and patterning of dendritic trees. To determine its function in mammalian Purkinje cells, Shima and co-workers (2004) have knocked down its homolog Celsr2 in organotypic cultures of P10 rat cerebella. The transduced Purkinje cells developed less complex trees with an ultimate phenotype of dendritic branch retraction, suggesting that Celsr2 may also regulate the growth and maintenance of Purkinje dendrites.
Role of Parallel Fibers in the Monoplanar Disposition and Branching Pattern of Purkinje Cell Dendrites: Comparison of Control and Agranular Cerebella
The key role of parallel fibers in the formation of Purkinje cell dendritic trees has been established many years ago through the study of different models of agranular cerebella. Indeed, the early disappearance of postmigratory granule cells, either by death of granule cell precursors or by inhibition of their proliferation, prevents all kinds of interactions between parallel fibers and Purkinje cells, allowing the examination of adult dendritic arbors grown in the absence of this input. The models used for these studies comprise: rat cerebellum exposed to postnatal X-irradiation (Altman and Anderson 1972), the weaver mouse (Hirano and Dembitzer 1973; Rakic and Sidman 1973; Sotelo 1973, 1975), and the central cerebellar mass of the releer mouse (Mariani et al. 1977; Sotelo 1978). In these conditions, the abnormalities of Purkinje dendrites are very similar and consist of: (1) severe reduction of the total length of the dendritic tree (atrophy); (2) nonplanar disposition of the dendrites, which become random with frequent inversion of the entire tree (loss of the espaliered shape); (3) lack of subdivision into a proximal and a distal dendritic compartment (spiny branchlets never form and proximal branches are studded with spines; Sotelo 1978). Since in these three situations the causes of the agranularity are very different and the only common thread in the developmental history of these neurons is the lack of granule cells, it can be concluded that the observed defects in the Purkinje cell dendrites are solely due to the lack of parallel fibers (Rakic and Sidman 1973; Sotelo 1975, 1978).
The dendritogenesis of embryonic Purkinje cells grafted to the adult pcd mutant cerebellum, devoid of this class of neurons, recapitulates most of the phases of normal development, although the duration of the last two phases is shorter (Sotelo et al. 1990). Since dendritic growth occurs ectopically, mainly within the molecular layer, grafted Purkinje cells do not adopt normal shapes, but remain atrophic, resembling those in the agranular cerebellum. In addition, the majority of donor neurons exhibit multipolar, short dendrites with few branches, resulting from the interaction between the growing dendrites and the adult, target devoid, parallel fiber varicosities.
Embryonic Purkinje cells grafted to adult hosts enter the cerebellum exclusively through the pial surface (Sotelo and Alvarado-Mallart 1991). However, when Pukinje cells are placed deep into the cerebellar parenchyma of perinatal hosts (before the expansion of the internal granular layer), they can still reach the cortex following the normal migratory route. These transplanted neurons become fully integrated in the host cerebellum (see section on “Formation of the Purkinje Cell Monolayer”), occupy correct positions and develop normal espaliered trees (Carletti et al. 2008). These results emphasize that, although the cerebellar milieu remains receptive for Purkinje cells even in adulthood, correct placement of donor neurons, allowing normal interactions with afferent and efferent fibers of the host and the acquisition of normal dendritic trees, depends on the timing of their migration, related to cerebellar developmental dynamics and granule cell layering.
Purkinje Cells as the Organizers of the Architecture and Projectional Arrangement of the Cerebellum
The central role played by Purkinje cells in cerebellar function is paralleled by their importance during development. Here, evidence will be presented indicating that Purkinje cells are the organizers of the whole ontogenetic process of the cerebellum, at least of the cortex, including neurogenesis, arrangement of the cortical layering, and topographic organization of its efferent–afferent projections. In other terms, without Purkinje cells there is no cerebellum.
Failure in the Differentiation of Purkinje Cells Leads to the Apparent Disappearance of the Cerebellum
Ptf1a, the transcription factor expressed by the cells of the ventricular neuroepithelium of the embryonic cerebellar primordium, is the disease-causing gene of neonatal diabetes mellitus associated with pancreatic and cerebellar agenesis (Sellick et al. 2003). In homozygous Ptf1a−/− mice, the cerebellar plate is extremely reduced in size at E16.5, leading to agenesis of the cerebellum at birth (Sellick et al. 2004). Another mouse strain carrying a mutation of Ptf1a, has been obtained during transgenesis experiments by Hoshino and collaborators (2005). In these mice, named “cerebelless,” the cerebellum is almost completely absent, except for a tiny rudimentary region, which exclusively contains glutamatergic neurons of the deep nuclei (reduced to about 40% of the normal number). Inferior olivary and pontine nuclei are also lacking, as expected for mice deprived of targets for climbing and mossy fibers (Hoshino et al. 2005).
In the ventricular neuroepithelium of cerebelless mice, GABAergic neurons, comprising both Purkinje cells and inhibitory interneurons, are generated, but are unable to progress in their differentiation and massively die soon after their birth (Hoshino et al. 2005). On the other hand, the severe reduction of glutamatergic deep nuclear neurons depends on the loss of the Purkinje cell input. Similarly, the complete absence of granule cells can be attributed to the lack of the proliferation-stimulating action exerted by Sonic Hedgehog (Shh) secreted by Purkinje cells (see the next section below). Therefore, even though glutamatergic neurons outnumber GABAergic neurons in the cerebellum, failure to develop the latter types completely disrupts cerebellar development. Notably, analysis of Ptf1a mutants shows that Purkinje cells are necessary for the proliferation of granule cell precursors and for the stabilization of glutamatergic nuclear neurons. These effects, together with the lack of GABAergic interneurons, prevent the formation of the cerebellar cortex, inducing further degeneration in the major extracerebellar afferent systems.
Purkinje Cells Stimulate Granule Cell Neurogenesis
Studies performed many years ago indicated that the presence of Purkinje cells was required for the normal proliferation of granule cells. Early interference with the Purkinje cells located under the external granular layer, either in spontaneous mutations such as staggerer (Mallet et al. 1976), early X-irradiation (Das 1977), or immunosurgery (cited in Sotelo 2004), greatly reduces the proliferation of granule cell precursors. Similarly, alterations in the location of Purkinje cells underlining the deepest strata of the external granular layer, like in the reeler mutant mouse, also massively reduce the number of generated granule cells (Mariani et al. 1977). With the advent of more selective approaches provided by molecular biology, it has been definitively demonstrated that the presence of Purkinje cells is indispensable for the proliferation of granule cell precursors. Thus, the use of diphtheria toxin, targeted to Purkinje cells with the specific promoter L7 (Smeyne et al. 1995), has provided transgenic cerebella, in which cortical bands containing Purkinje cells alternate with bands devoid of these neurons. The latter gaps in the Purkinje cell layer are coincidental with thinned zones in the overlying external granular layer, underscoring the mitogenic role of Purkinje cells.
The observation that the gene encoding Shh is expressed by Purkinje cells, while its receptor Patched (Ptch) is expressed by granule cell precursors, suggests that the pathway Shh–Gli–Ptch regulates the proliferation of these precursors. In line with this possibility, treatment of granule cell precursors with Shh prevents their differentiation and induces a potent, long-lasting proliferative response, whereas application of neutralizing anti-Shh antibodies in vivo reduces BrdU incorporation and gives rise to an agranular cerebellum, together with deficient Bergmann glia differentiation and abnormal Purkinje cell dendritogenesis. These results clearly indicate that Shh, secreted by Purkinje cells, is responsible for the proliferation of granule cell precursors (Dahmane and Ruiz i Altaba 1999; Wallace 1999; Wechsler-Reya and Scott 1999).
The use of DNA microarrays to identify more eventual targets of Shh in granule cell precursors points to cyclin D1 and N-myc. Both genes are robustly induced by Shh, N-myc being induced even in the presence of the protein synthesis inhibitor cycloheximide (Oliver et al. 2003). Moreover, Atoh1 plays the role of regulator of the signal transduction pathway of Shh, and its deletion prevents the formation of medulloblastoma (Flora et al. 2009).
Although extracellular matrix proteins, such as vitronectin, expressed in the lower external granular layer are able to block Shh activity (Pons et al. 2001), the mechanisms that inform granule cell precursors to exit the cell cycle and switch from a proliferative state in the upper layer to differentiation in the lower layer is still poorly understood. Indeed, increasing the levels of cAMP and the activity of PKA can antagonize the response to Shh. The G-protein-coupled receptor GPR3 (GPCR21), a member of a family of G-protein-coupled receptors, can stimulate adenylate cyclases (Tanaka et al. 2007). It has been reported that increase of GPR3 antagonizes Shh in vitro, and its inhibition by siRNA in vivo stimulates Shh-induced proliferation of granule cell precursors. Therefore, this orphan receptor could constitute one of the signals that mediate the transition between proliferation and differentiation of granule cell precursors (Tanaka et al. 2009).
Modular Organization of the Input/Output Projections in the Cerebellum: The Olivocerebellar System
In addition to the classical organization of the cerebellar cortex into lobes and lobules forming the folia that emerge from its folding, the adult cerebellum is topographically organized into parasagittal bands and transverse zones that, although lacking visible anatomical frontiers, have fixed positions outlined by molecular markers (Sillitoe and Joyner 2007). This three-dimensional organization should match a hidden projectional heterogeneity emerging from the complex input-output organization (Voogd 1969). Indeed, extracerebellar afferent fibers (without considering the modulatory monoaminergic systems) terminate into narrow parasagittally oriented cortical modules, whose Purkinje cells innervate specific regions of the cerebellar nuclei, matching with the functional heterogeneity and microzonal organization of the cerebellum (Oscarsson 1979). The entire olivo-cortico-nuclear network is organized in defined modules, where clusters of neurons in the inferior olive project to specific Purkinje cell bands in the cortex and to discrete groups of nuclear neurons innervated by the same Purkinje cells. Subsets of the latter nuclear neurons, which are GABAergic, send their axons back to the original neuronal cluster in the inferior olive. Their terminal bulbs synapse straddling the dendrites of olivary neurons coupled by gap junctions, to regulate the level of electrotonic coupling (Angaut and Sotelo 1989), which seems to be essential for learning-dependent timing in cerebellar motor control (van der Giessen et al. 2008).
Adult Cerebellum: Purkinje Cell Biochemical Heterogeneity, Parasagittal Stripes of Protein Expression
The advent of histochemical and immunohistochemical techniques has disclosed that, parallel to the topographic arrangement of extracerebellar projections in parasagittal modules, Purkinje cells are also neurochemically heterogeneous (Hawkes et al. 1985) and distributed into alternating subsets that either possess or lack the corresponding antigens (Gravel et al. 1987). Over the last decades, numerous markers expressed by alternating bands of Purkinje cells have been reported, leading to describe a complex three-dimensional pattern, in which each cortical compartment is distinguished by a specific combination of markers (Apps and Hawkes 2009). By integrating these observations with the findings obtained with other anatomical, physiological or molecular methods in developing and adult animals, Apps and Hawkes (2009) have formulated the “one-map hypothesis,” concluding that each approach “reveals different facets of a common topography.”
Over the last decades, a molecular coding of the cerebellum has been progressively disclosed. The three-dimensional complexity of this nervous center is arranged around a basic coordinate system that, as clearly described by Sillitoe and Joyner (2007), can be broken down into morphology and molecular coding. The precise anatomo-molecular description of this complexity is out of the scope of this chapter, and can be appraised in numerous reviews (Hawkes and Gravel 1991; Hawkes et al. 1992; Armstrong and Hawkes 2000; Sillitoe and Joyner 2007; Apps and Hawkes 2009).
One Congruent Map or Several Independent Maps?
One of the main questions, about the functional significance of the parasagittal-striped disposition of heterogeneous Purkinje cells, is to reveal whether the cortical compartmentation correlates with the arrangement of the projection maps of extracerebellar afferents. The first investigations that addressed this question were focused on the olivocerebellar map, by combining anterograde tracing of the olivocerebellar pathway and anti-zebrin immunostaining to highlight Purkinje cell bands (Fig. 9.5; Gravel et al. 1987; Wassef et al. 1992a). Since the olivocerebellar projection is discontinuous (Buisseret-Delmas and Angaut 1993), a small random injection of an axonal tracer in the inferior olive results in the labeling of several sagittal bands. These bands have boundaries (local discontinuities) that are characteristic of the topography of this projection and pre-date the actual injection. Under these conditions, it has been shown that the boundaries of anterogradely traced axons and those of immunoreactive Purkinje cell subsets are strongly correlated (Fig. 9.5; Gravel et al. 1987; Wassef et al. 1992a). Statistical analysis confirms that the coincidence between olivocerebellar and Purkinje cell bands is much too high to result from a random distribution and, therefore, both maps are congruent (Wassef et al. 1992a).


Congruence between Purkinje cell bands and the olivocerebellar projection to the cerebellar cortex. (a) Coronal section through the caudal vermis of an adult rat injected with tritiated amino acids in the inferior olive, and the sections treated for zebrin I immunoreactivity and autoradiography. The 3H-labeled climbing fibers are visualized in dark field (white points), whereas the zebrin immunostaining is identified in bright field, to allow the conjoined visualization of both markers (brown color cells). The labeled clusters of climbing fibers and the zebrin positive or negative compartments of Purkinje cells have numerous common boundaries, too many to be due to hazard, indicating that both maps are congruent. (b) Another coronal section of the rat cerebellar vermis, in which climbing fibers are visualized by biotynylated dextran amine tracing (black), while Purkinje cell bands are highlighted by zebrin immunostaining (brown). The correspondence between Purkinje cell compartments and the parasagittally oriented strips of climbing fibers is evident. Scale bars, 100 μm. The material for the micrograph in the upper half was published in a book chapter (Wassef et al. 1992a) Figs. 1–10. The lower micrograph is unpublished material by Ferdinando Rossi
Recently, the matching between Purkinje cell and olivocerebellar maps has been tested for more precise anatomical regions, and for its modular organization. It has been shown that the projections emerging from specific parts of the medial or dorsal accessory olive are congruent with narrow, longitudinal stripes of Purkinje cells in the zebrin-negative C1 and C3 zones of the anterior and posterior cerebellar cortex. These Purkinje cells project to specific regions of the cerebellar nuclei, which also receive direct projections from collaterals of the same climbing fibers (Pijpers et al. 2005). This way, the functional modular organization of the cerebellum (its olivo-cortico-nuclear connections) appears tightly related to the disposition of neurochemically distinct Purkinje cell subsets.
To assess whether the adult pattern of Purkinje cell heterogeneity is independently expressed or is induced by the olivocerebellar afferents, the cerebellar anlagen of E12–E15 rat embryos, which have not yet been contacted by climbing fibers (Wassef et al. 1992b), have been ectopically grafted into cavities placed in adult neocortex (Wassef et al. 1990). The grafts, which evolve into minicerebellar structures in complete absence of climbing fibers, once immunostained with anti-zebrin antibodies display a clear arrangement of alternating bands of immunopositive and immunonegative Purkinje cells. These results show that the neurochemical profile and the parasagittal segregation of Purkinje cell subsets can be achieved in the absence of olivocerebellar fibers, indicating that the postsynaptic Purkinje neurons direct the building up of the afferent projection map.
Organization of the Cerebellar Cortical Layering; Transient Biochemical Heterogeneity of Purkinje Cells and Inferior Olivary Neurons During Embryonic Development
One problem to conclude that the congruence between Purkinje cell arrangement and the olivocerebellar projection map reflects an ontogenetic organizational process is that the expression of adult Purkinje cell markers begins too late to discern any transition between immature and adult patterns. Nevertheless, a number of proteins and glycoproteins that in the adult cerebellum are selectively expressed in the entire population of Purkinje cells (calbindin, cyclic GMP-dependent protein kinase, Purkinje cell–specific glycoprotein; PEP-19) show a peculiar developmental expression pattern that may help to address this issue. Analysis of Purkinje cell differentiation in the embryonic rat cerebellum with antibodies against these “Purkinje cell markers” has disclosed that, during transient developmental periods, only subsets of Purkinje cells are labeled (Wassef and Sotelo 1984; Wassef et al. 1985). The different subsets are arranged in alternating, bilaterally symmetric, positive and negative parasagittal clusters, which follow dynamic developmental changes. Each antibody provides a distinct but reproducible pattern, even though some boundaries can be common to two or more markers. This peculiar organization of the cerebellum persists until P5, when all Purkinje cells express these markers and the clusters disappear. This transient segregation of immature Purkinje cells indicates the establishment of an intrinsic compartmentation of the early developing cerebellar cortex. The basic compartment is not the immunopositive or negative cluster, but the intersection of such clusters where, by definition, all these neurons are identical.
Within the context of the chemoaffinity hypothesis (Sperry 1963), the early compartmentation of the cerebellar cortex needs, as a counterpart, a similar acquisition of positional identities in inferior olivary neurons. Indeed, this counterpart does exist. Starting at E16 in the rat, before the entry of olivocerebellar axons in the cerebellar parenchyma, clusters of inferior olivary neurons also display a transient biochemical heterogeneity (Wassef et al. 1992b), defining an intrinsic compartmentation of this nucleus (Wassef et al. 1990).
These observations indicate that the neurochemical heterogeneity of Purkinje cells and inferior olivary neurons arise independently, and that the transient biochemical individualization of subgroups in these two immature neuronal populations could contribute to recognition mechanisms. More importantly, an anatomical relationship between this developmental compartmentation and the organization of the olivocerebellar projection has been found (Wassef et al. 1992c), allowing the formulation of the “matching hypothesis” (Sotelo 2004): the acquisition of subpopulation-type identities in inferior olivary neurons and in the Purkinje cells would direct the construction of the projection map by matching of appropriated positional cues (Sotelo and Wassef 1991a; Sotelo and Chédotal 1997). A clear example of this mechanism is provided by the climbing fiber projection arising from the dorsal cap of the inferior olive. These axons, which terminate in vermal lobules IX and X and in the flocculus, show transient parvalbumin immunoreactivity around birth. In newborn rats, the projection has already attained the adult arrangement. Immunostaining with anti-PEP-19 antibodies, to visualize Purkinje cell clusters, shows that the medial parvalbumin immunoreactive climbing fiber band of lobule X is coextensive with a PEP-19 negative Purkinje cell cluster, revealing a clear projectional relationship between the biochemical parcellations of the cerebellum and of the inferior olive (Wassef et al. 1992c).
Organization of the Three-Dimensional Architecture of the Projectional Maps; Validation of the “Matching Hypothesis” with the Mossy Fiber System
Mossy fiber neurons are distributed from the spinal cord to the pons in several precerebellar nuclei. Even though these axons mainly establish synaptic contacts with granule cells, and less often with Golgi and unipolar brush cells, the mossy fiber projection is also organized according to a parasagittal banding pattern, similar to that followed by climbing fibers (Pijpers et al. 2006). The relationships between mossy fiber terminal fields and Purkinje cell antigenic compartments have been also explored by anterograde axonal tracing and zebrin II immunoreactivity in the rat (Ji and Hawkes 1994) and in the mouse cerebellum (Akintunde and Eisenman 1994). These studies revealed species-specific differences, since in the rat the matching between mossy fibers and Purkinje cell bands was stricter than in the mouse. In both species, however, the congruence was less precise than that of the olivocerebellar projection.
During development of the rat, the spinocerebellar mossy fibers enter the cerebellum at the end of fetal life, and the adult topographic pattern only emerges during the first postnatal week, after a transitional period from P3 to P7 (Arsénio-Nunes and Sotelo 1985). During the latter period, spinocerebellar axons invade the gray matter forming a kind of loose and dispersed bands (the “protocolumnar” stage), which undergo a progressive reshaping to reach their ultimate columnar pattern by P7.
Quite remarkably, in the developing mouse cerebellum, immature mossy fibers profusely branch in the cerebellar cortex and terminate with growth cone-like structures with long filopodia in contact with Purkinje cell bodies, particularly at the end of the first postnatal week or at the middle of the second (Mason and Gregory 1984). These transient synaptic contacts are functional (Takeda and Maekawa 1989) and might be a key element in the settling of the topography of this projection. Indeed, analysis of mutant mice with agranular cerebella has allowed discriminating which among granule cells and Purkinje cells is the organizer of the topography. Thus, in the weaver cerebellum, a recessive mutation of an inward rectifying potassium channel (Patil et al. 1995) primarily affects the granule cells and, hence, most of the premigratory granule cells degenerate. On the contrary in the staggerer mouse 75% of Purkinje cells die and, consequently, granule cells are also reduced (see above). In both instances the cerebella are agranular and most of the natural targets of mossy fibers are missing, but it is only in the staggerer that Purkinje cell population is severely reduced and abnormal. Accordingly, important differences in the organization of the mossy fiber projection are evident in these mice. While in the weaver the projection is segregated in parasagittal columns, in the staggerer it is more or less homogeneously distributed, without any obvious columnar pattern (Arsenio-Nunes et al. 1988). Most importantly, while in the weaver the characteristic parasaggittal compartmentation of Purkinje cells is well preserved, in the staggerer zebrin-positive Purkinje cells are missing (Sotelo and Wassef 1991b). Therefore, as for climbing fibers, Purkinje cells are the organizer elements of the spinocerebellar projectional map.
In conclusion, the congruence between climbing and mossy fiber projectional maps and the Purkinje cell parasagittal organization is maintained even in the weaver cerebellum, deprived of granule cells, but containing more than 50% of Purkinje cells. This provides a strong argument to validate the “matching hypothesis” and allows the conclusion that Purkinje cell arrays constitute the scaffolding through which extracerebellar afferent fibers and Purkinje cell axonal projections attain the normal organization during development.
Conclusions
The evidence described in this chapter shows that Purkinje cell development has a dual significance, in which the acquisition of mature traits is combined with a complex organizational influence on the whole cerebellar ontogenesis. Although many of the cellular interactions underlying these phenomena are now becoming clear, the molecular cues and signaling pathways are only partially elucidated. Indeed, in spite the important role played by a restricted number of main actors, such as Reelin in migratory events or Shh in progenitor proliferation, the mechanisms by which Purkinje cells exert their organizational function have still to be elucidated. For instance, while the existence of heterogeneous subsets of Purkinje cells and extracerebellar afferents is well established, the mechanisms that mediate their reciprocal recognition and specific interaction in the formation of map topography are still uncertain. Similarly, it is unclear how chemoaffine recognition interacts with activity/experience-dependent events that contribute to shape cerebellar connectivity at late ontogenetic stages. These, as well as many others, are unanswered questions of general interest in developmental neurobiology. Once more, the simple, stereotyped structure of the cerebellar network will provide the best ground to address such fundamental issues.
References
Akintunde A, Eisenman LM (1994) External cuneocerebellar projection and Purkinje cellzebrin II bands: a direct comparison of parasagittal banding in the mouse cerebellum. J Chem Neuroanat 7:75–86
Altman J, Anderson WJ (1972) Experimental reorganization of the cerebellar cortex. I. Morphological effects of elimination of all microneurons with prolonged x-irradiation started at birth. J Comp Neurol 146:355–406
Altman J, Bayer SA (1997) Development of the cerebellar system in relation to its evolution, structures and functions. CRC, Boca Raton/New York
Altman J, Winfree AT (1977) Postnatal development of the cerebellar cortex in the rat. V. Spatial organization of Purkinje cell perikarya. J Comp Neurol 171:1–16
Angaut P, Sotelo C (1989) Synaptology of the cerebello-olivary pathway. Double labelling with anterograde axonal tracing and GABA immunocytochemistry in the rat. Brain Res 479:361–365
Apps R, Hawkes R (2009) Cerebellar cortical organization: a one map hypothesis. Nat Rev Neurosci 10:670–681
Armengol JA, Sotelo C (1991) Early dendritic development of Purkinje cells in the rat cerebellum. A light and electron microscopic study using axonal tracing in “in vitro” slices. Brain Res Dev Brain Res 64:95–114
Armstrong CL, Hawkes R (2000) Pattern formation in the cerebellar cortex. Biochem Cell Biol 78:551–562
Arsénio Nunes ML, Sotelo C (1985) Development of the spinocerebellar system in the postnatal rat. J Comp Neurol 237:291–306
Arsenio-Nunes ML, Sotelo C, Wehrle R (1988) Organization of spinocerebellar projection map in three types of agranular cerebellum: Purkinje cells vs. granule cells as organizer element. J Comp Neurol 273:120–136
Baptista CA, Hatten ME, Blazeski R, Mason CA (1994) Cell-cell interactions influence survival and differentiation of purified Purkinje cells in vitro. Neuron 12:243–260
Berry M, Bradley P (1976) The growth of the dendritic trees of Purkinje cells in the cerebellum of the rat. Brain Res 112:1–35
Boukhtouche F, Janmaat S, Vodjdani G, Gautheron V, Mallet J, Dusart I, Mariani J (2006) Retinoid-related orphan receptor alpha controls the early steps of Purkinje cell dendritic differentiation. J Neurosci 26:1531–1538
Bourrat F, Sotelo C (1986) Neuronal migration and dendritic maturation of the medial cerebellar nucleus in rat embryos: an HRP in vitro study using cerebellar slabs. Brain Res 378:69–85
Bouslama-Oueghlani L, Wehrlé R, Sotelo C, Dusart I (2003) The developmental loss of the ability of Purkinje cells to regenerate their axons occurs in the absence of myelin: an in vitro model to prevent myelination. J Neurosci 23:8318–8329
Bradley P, Berry M (1976) Quantitative effects of climbing fibre deafferentation on the adult Purkinje cell dendritic tree. Brain Res 112:133–140
Bradley P, Berry M (1978) The Purkinje cell dendritic tree in mutant mouse cerebellum. A quantitative Golgi study of Weaver and Staggerer mice. Brain Res 142:135–141
Buffo A, Zagrebelsky M, Huber A, Skerra A, Schwab ME, Strata P, Rossi F (2000) Application of neutralising antibodies against NI-35/250 myelin-associated neurite growth inhibitory proteins to the adult rat cerebellum induces sprouting of uninjured Purkinje cell axons. J Neurosci 20:2275–2286
Buisseret-Delmas C, Angaut P (1993) The cerebellar olivo-corticonuclear connections in the rat. Prog Neurobiol 40:63–87
Carletti B, Rossi F (2008) Neurogenesis in the cerebellum. Neuroscientist 14:91–100
Carletti B, Williams IM, Leto K, Magrassi L, Rossi F (2008) Time constraints and positional cues in the developing cerebellum regulate Purkinje cell placement in the cortical architecture. Dev Biol 317:147–160
Chedotal A, Sotelo C (1992) Early development of olivocerebellar projections in the fetal rat using CGRP immunocytochemistry. Eur J Neurosci 4:1159–1179
Chedotal A, Sotelo C (1993) The “creeper stage” in cerebellar climbing fiber synaptogenesis precedes the “pericellular nest”, ultrastructural evidence with parvalbumin immunocytochemistry. Brain Res Dev Brain Res 76:207–220
Clark BA, Monsivais P, Branco T, London M, Hausser M (2005) The site of action potential initiation in cerebellar Purkinje neurons. Nat Neurosci 8:137–139
Crepel F, Delhaye-Bouchaud N, Dupont JL, Sotelo C (1980) Dendritic and axonic fields of Purkinje cells in developing and x-irradiated rat cerebellum. A comparative study using intracellular staining with horseradish peroxidase. Neuroscience 15:333–347
Curmi PA, Gavet O, Charbaut E, Ozon S, Lachkar-Colmerauer S, Manceau V, Siavoshian S, Maucuer A, Sobel A (1999) Stathmin and its phosphoprotein family: general properties, biochemical and functional interaction with tubulin. Cell Struct Funct 24:345–357
D’Antoni S, Zambusi L, Codazzi F, Zacchetti D, Grohovaz F, Provini L, Catania MV, Morara S (2010) Calcitonin gene-related peptide (CGRP) stimulates Purkinje celldendrite growth in culture. Neurochem Res 35:2135–2143
Dahmane N, Ruiz i Altaba A (1999) Sonic hedgehog regulates the growth and patterning of the cerebellum. Development 126:3089–3100
Das GD (1977) Experimental analysis of embryogenesis of cerebellum in rat. I. Subnormal growth following x-ray irradiation on day 15 of gestation. J Comp Neurol 176:419–434
De Camilli P, Miller PE, Levitt P, Walter U, Greengard P (1984) Anatomy of cerebellar Purkinje cells in the rat determined by a specific immunohistochemical marker. Neuroscience 11:761–817
de Luca A, Vassallo A, Benitez-Temino B, Menichetti G, Rossi F, Buffo A (2009) Distinct modes of neuritic growth in Purkinje neurons at different developmental stages: axonal morphogenesis and cellular regulatory mechanisms. PLoS ONE 4:e6848
Eisenman LM, Schalekamp MPA, Voogd J (1991) Development of the cerebellar cortical efferent projection: an in vitro study in rat brain slices. Brain Res Dev Brain Res 60:261–266
Fernandez-Gonzalez A, La Spada AR, Treadaway J, Higdon JC, Harris BS, Sidman RL, Morgan JI, Zuo J (2002) Purkinje cell degeneration (pcd) phenotypes caused by mutations in the axotomy-induced gene, Nna1. Science 295:1904–1906
Fink AJ, Englund C, Daza RAM, Pham D, Lau C, Nivison M, Kovalczyk T, Hevner R (2006) Development of the deep cerebellar nuclei: transcription factors and cell migration from the rhombic lip. J Neurosci 26:3066–3076
Flora A, Klisch TJ, Schuster G, Zoghbi HY (2009) Deletion of Atoh1 disrupts Sonic Hedgehog signaling in the developing cerebellum and prevents medulloblastoma. Science 326:1424–1427
Foscarin S, Gianola S, Carulli D, Fazzari P, Mi S, Tamagnone L, Rossi F (2009) Overexpression of GAP-43 modifies the distribution of the receptors for myelin-associated growth-inhibitory proteins in injured Purkinje axons. Eur J Neurosci 30:1837–1848
Gardette R, Debono M, Dupont JL, Crepel F (1985) Electrophysiological studies on the postnatal development of intracerebellar nuclei neurons in rat cerebellar slices maintained in vitro. I. Postsynaptic potentials. Brain Res 351:47–55
Gianola S, Savio T, Schwab ME, Rossi F (2003) Cell-autonomous mechanisms and myelin-associated factors contribute to the development of Purkinje axon intracortical plexus in rat cerebellum. J Neurosci 23:4613–4624
Gianola S, de Castro SF, Rossi F (2009) Anosmin-1 stimulates outgrowth and branching of developing Purkinje axons. Neuroscience 158:570–584
Gilmore EC, Herrup K (2000) Cortical development: receiving reelin. Curr Biol 10:R162–R166
Goffinet AM (1983) The embryonic development of the cerebellum in normal and reeler mutant mice. Anat Embryol 168:73–86
Golgi C (1883) Sulla fina anatomia degli organi centrali del sistema nervoso IV. Sulla fina anatomia delle circonvoluzioni cerebellari. Riv Sper Freniatr Med Leg 9:1–17
Gravel C, Leclerc N, Rafrafi J, Sasseville R, Thivierge L, Hawkes R (1987) Monoclonal antibodies reveals the global organization of the cerebellar cortex. J Neurosci Meth 21:145–157
Hack I, Bancila M, Loulier K, Carroll P, Cremer H (2002) Reelin is a detachment signal in tangential chain-migration during postnatal neurogenesis. Nat Neurosci 5:939–945
Hamilton BA, Frankel WN, Kerrebrock AW, Hawkins TL, FitzHugh W, Kusumi K, Russell LB, Mueller K, van Berkel V, Birren BW, Kruglyak L, Lander ES (1996) Disruption of the nuclear hormone receptor RORalpha in staggerer mice. Nature 379:736–739
Hatten ME (1999) Central nervous system neuronal migration. Annu Rev Neurosci 22:511–539
Hatten ME, Heintz N (1995) Mechanism of neural patterning and specialization in the developing cerebellum. Annu Rev Neurosci 18:385–408
Hawkes R, Gravel C (1991) The modular cerebellum. Prog Neurobiol 36:309–327
Hawkes R, Colonnier M, Leclerc N (1985) Monoclonal antibodies reveal sagittal banding in the rodent cerebellar cortex. Brain Res 333(2):359–365
Hawkes R, Brochu G, Doré L, Gravel C, Leclerc N (1992) Zebrins: molecular markers of compartmentation in the cerebellum. In: Llinas R, Sotelo C (eds) The cerebellum revisited. Springer, New York, pp 22–55
Henke RM, Savage TK, Meredith DM, Glasgow SM, Hori K, Dumas J, MacDonald RJ, Johnson JE (2009) Neurog2 is a direct downstream target of the Ptf1a-Rbpj transcription complex in dorsal spinal cord. Development 136:2945–2954
Hirai H, Launey T (2000) The regulatory connection between activity of granule cells NMDA receptors and dendritic differentiation of cerebellar Purkinje cells. J Neurosci 20:5217–5224
Hirano A, Dembitzer HM (1973) Cerebellar alterations in the weaver mouse. J Cell Biol 56:478–486
Hoshino M (2006) Molecular machinery governing GABAergic neuron specification in the cerebellum. Cerebellum 5:193–198
Hoshino M, Nakamura S, Mori K, Kawauchi T, Terao M, Nishimura YV, Fukuda A, Fuse T, Matsuo N, Sone M, Watanabe M, Bito H, Terashima T, Wright CVE, Kawaguchi Y, Nakao K, Nabeshima Y (2005) Ptf1a, a bHLH transcriptional gene, defines GABAergic neuronal fates in cerebellum. Neuron 47:201–213
Ito M (2006) Cerebellar circuitry as a neuronal machine. Prog Neurobiol 78:272–303
Jensen P, Zoghbi HY, Goldowitz D (2002) Dissection of the cellular and molecular events that position cerebellar Purkinje cells: a study of the math1 null-mutant mouse. J Neurosci 22:8110–8116
Jensen P, Smeyne R, Goldowitz D (2004) Analysis of cerebellar development in math1 null embryos and chimeras. J Neurosci 24:2202–2211
Ji Z, Hawkes R (1994) Topography of Purkinje cell compartments and mossy fiber terminal fields in lobules II and III of the rat cerebellar cortex: spinocerebellar and cuneocerebellar projections. Neuroscience 61:935–954
Kapfhammer JP (2004) Cellular and molecular control of dendritic growth and development of cerebellar Purkinje cells. Prog Histochem Cytochem 39:131–182
Kawaguchi K, Habara T, Terashima T, Kikkawa S (2010) GABA modulates development of cerebellar Purkinje cell dendrites under the control of endocannbinoid signaling. J Neurochem 114:627–638
Kim D, Ackerman S (2011) The UNC5C Netrin receptor regulates dorsal guidance of mouse hindbrain axons. J Neurosci 31:2167–2179
King JS, Bishop GA (1982) The synaptic features of horseradish peroxidase-labelled recurrent collaterals in the ganglionic plexus of the cat cerebellar cortex. J Neurocytol 11:867–880
Larramendi LN, Victor T (1967) Synapses on the Purkinje cell spines in the mouse. An electron microscopic study. Brain Res 5:15–30
Li J, Gu X, Ma Y, Calicchio ML, Kong D, Teng YD, Yu L, Crain AM, Vartanian TK, Pasqualini R, Arap W, Libermann TA, Snyder EY, Sidman RL (2010) Nna1 mediates Purkinje cell dendritic development via lysyl oxidase propeptide and NF-kB signaling. Neuron 68:45–60
Machold R, Fishell G (2005) Math1 is expressed in temporally discrete pools of cerebellar rhombic-lip neural progenitors. Neuron 48:17–24
Magdaleno S, Keshvara L, Curran T (2002) Rescue of ataxia and preplate splitting by ectopic expression of reelin in reeler mice. Neuron 33:573–586
Mallet J, Huchet M, Pougeois R, Changeux JP (1976) Anatomical, physiological and biochemical studies on the cerebellum from mutant mice. III. Protein differences associated with the weaver, staggerer and nervous mutations. Brain Res 103:291–312
Mariani J, Crepel F, Mikoshiba K, Changeux JP, Sotelo C (1977) Anatomical, physiological and biochemical studies of the cerebellum from Reeler mutant mouse. Philos Trans R Soc Lond B Biol Sci 281:1–28
Mason CA, Gregory E (1984) Postnatal maturation of cerebellar mossy and climbing fibers: transient expression of dual features on single axons. J Neurosci 4:1715–1735
Mason CA, Christakos S, Catalano SM (1990) Early climbing fiber interactions with Purkinje cells in the postnatal mouse cerebellum. J Comp Neurol 297:77–90
Miale IL, Sidman RL (1961) An autoradiographic analysis of histogenesis in the mouse cerebellum. Exp Neurol 4:277–296
Minaki Y, Nakatani T, Mizuhara E, Inoue T, Ono Y (2008) Identification of a novel transcriptional corepressor, Corl2, as a cerebellar Purkinje cell-selective marker. Gene Expr Patterns 8:418–423
Miyata T, Nakajima K, Aruga J, Takahashi S, Ikenaka K, Mikoshiba K, Ogawa M (1996) Distribution of a reeler gene-related antigen in the developing cerebellum: an immunohistochemical study with an allogeneic antibody CR-50 on normal and reeler mice. J Comp Neurol 372:215–228
Miyata T, Nakajima K, Mikoshiba K, Ogawa M (1997) Regulation of Purkinje cell alignment by reelin as revealed with CR-50 antibody. J Neurosci 17:3599–3609
Miyata T, Ono Y, Okamoto M, Masaoka M, Sakakibara A, Kawaguchi A, Hashimoto M, Ogawa M (2010) Migration, early axonogenesis, and Reelin-dependent layer-forming behavior of early/posterior-born Purkinje cells in the developing mouse lateral cerebellum. Neural Dev 5:23
Mizuhara E, Minaki Y, Nakatani T, Kumai M, Inoue T, Muguruma K, Sasai Y, Ono Y (2010) Purkinje cells originate from cerebellar ventricular zone progenitors positive for Neph3 and E-cadherin. Dev Biol 338:202–214
Morara S, van der Want JJ, de Weerd H, Provini L, Rosina A (2001) Ultrastructural analysis of climbing fiber-Purkinje cell synaptogenesis in the rat cerebellum. Neuroscience 108:655–671
Nishida K, Hoshino M, Kawaguchi Y, Murakami F (2010) Ptf1a directly controls expression of immunoglobulin superfamily molecules Nephrin and Neph3 in the developing central nervous system. J Biol Chem 285:373–380
Obata J, Yano M, Mimura H, Goto T, Nakayama R, Mibu Y, Oka C, Kawaichi M (2001) p48 subunit of mouse PTF1 binds to RBP-Jkappa/CBF-1, the intracellular mediator of Notch signalling, and is expressed in the neural tube of early stage embryos. Genes Cells 6:345–360
Oliver TG, Grasfeder LL, Carroll AL, Kaiser C, Gillingham CL, Lin SM, Wickramasinghe R, Scott MP, Wechsler-Reya RJ (2003) Transcriptional profiling of Sonic hedgehog response: a critical role for N-myc in proliferation of neuronal precursors. Proc Natl Acad Sci USA 100:7331–7336
Orduz D, Llano I (2007) Recurrent axon collaterals underlie facilitating synapses between cerebellar Purkinje cells. Proc Natl Acad Sci USA 104:17831–17836
Oscarsson O (1979) Functional units of the cerebellum: sagittal zones and microzones. Trends Neurosci 2:143–145
Palay SL, Chan-Palay V (1974) Cerebellar cortex. Cytology and organization. Springer, Berlin/Heidelberg/New York
Pascual M, Abasolo I, Mingorance-Le Meur A, Martinez A, Del Rio JA, Wright CV, Real FX, Soriano E (2007) Cerebellar GABAergic progenitors adopt an external granule cell-like phenotype in the absence of Ptf1a transcription factor expression. Proc Natl Acad Sci USA 104:5193–5198
Patil N, Cox DR, Bhat D, Faham M, Myers RM, Peterson AS (1995) A potassium channel mutation in weaver mice implicates membrane excitability in granule cell differentiation. Nat Genet 11:126–129
Pijpers A, Voogd J, Ruigrok TJ (2005) Topography of olivo-cortico-nuclear modules in the intermediate cerebellum of the rat. J Comp Neurol 497:670–682
Pijpers A, Apps R, Pardoe J, Voogd J, Ruigrok TJH (2006) Precise spatial relationships between mossy fibers and climbing fibers in the rat cerebellar cortical zones. J Neurosci 26:12067–12080
Pons S, Trejo JL, Martinez-Morales JR, Marti E (2001) Vitronectin regulates Sonic hedgehog activity during cerebellum development through CREB phosphorylation. Development 128:1481–1492
Poulain FE, Chauvin S, Wehrlé R, Desclaux M, Mallet J, Vodjdani G, Dusart I, Sobel A (2008) SCLIP is crucial for the formation and development of the Purkinje cell dendritic arbor. J Neurosci 28:7387–7398
Purkinje JE (1837) Neueste Untersuchungen aus der Nervenund Hirnanatomie. In: Sternberg K, von Krombhloz JV (eds) Bericht €uber die Versammlung deutscher Naturforscher und Ärzte im September 1837. Hasse, Prag, pp 177–180
Rakic P, Sidman RL (1970) Histogenesis of cortical layers in human cerebellum, particularly the lamina dissecans. J Comp Neurol 139:473–500
Rakic P, Sidman RL (1973) Organization of cerebellar cortex secondary to deficit of granule cells in weaver mutant mice. J Comp Neurol 152:133–161
Ramón y Cajal S (1888) Sobre las fibras nerviosas de la capa molecular del cerebelo. Rev Trim Histol Norm Patol 2:33–41
Ramón y Cajal S (1890) À propos de certain éléments bipolaires du cervelet avec quelques détails nouveaux sur l’évolution des fibres cérébelleuses. Int Msch Anat Physiol 7:12–30
Ramón y Cajal S (1911) Histologie du Système Nerveux de l’Homme et des Vertébrés, vol II. A. Maloine, Paris
Reynolds R, Wilkins GP (1988) Development of macroglial cell in rat cerebellum. II. An in situ immunohistochemical study of oligodendroglial lineage from precursor to mature myelinating cell. Development 102:409–425
Rice DS, Curran T (2001) Role of the Reelin signaling pathway in central nervous system development. Annu Rev Neurosci 24:1005–1039
Rossi F, Gianola S, Corvetti L (2006) The strange case of Purkinje axon regeneration and plasticity. Cerebellum 5:174–182
Rossi F, Gianola S, Corvetti L (2007) Regulation of intrinsic neuronal properties for axon growth and regeneration. Prog Neurobiol 81:1–28
Schiffmann SN, Bernier B, Goffinet AM (1997) Reelin mRNA expression during mouse brain development. Eur J Neurosci 9:1055–1071
Sellick GS, Garrett C, Houlston RS (2003) A novel gene for neonatal diabetes maps to chromosome 10p12.1-p13. Diabetes 52:2636–2638
Sellick GS, Barker KT, Stolte-Dijkstra I, Fleischmann C, Coleman RJ, Garrett C, Gloyn AL, Edghill EL, Hattersley AT, Wellauer PK, Goodwin G, Houlston RS (2004) Mutations in PTF1A causes pancreatic and cerebellar agenesis. Nat Genet 36:1301–1305
Shima Y, Kengaku M, Hirano T, Takeichi M, Uemura T (2004) Regulation of dendritic maintenance and growth by a mammalian 7-pass transmembrane cadherin. Dev Cell 7:205–216
Sillitoe RV, Joyner AL (2007) The three-dimensional complexity of the cerebellum. Annu Rev Cell Dev Biol 23:549–577
Sillitoe RV, Gopal N, Joyner AL (2009) Embryonic origins of ZebrinII parasagittal stripes and establishment of topographic Purkinje cell projections. Neuroscience 162:574–588
Sirzen-Zelenskaya A, Zeyse J, Kapfhammer JP (2006) Activation of class I metabotropic glutamate receptors limits dendritic growth of Purkinje cells in organotypic slice cultures. Eur J Neurosci 24:2978–2986
Smeyne RJ, Chu T, Lewin A, Bian F, S-Crisman S, Kunsch C, Lira SA, Oberdick J (1995) Local control of granule cell generation by cerebellar Purkinje cells. Mol Cell Neurosci 6:230–251
Sotelo C (1973) Permanence and fate of paramembranous synaptic specializations in “mutants” experimental animals. Brain Res 62:345–351
Sotelo C (1975) Anatomical, physiological and biochemical studies of the cerebellum from mutant mice. II. Morphological study of cerebellar cortical neurons and circuits in the weaver mouse. Brain Res 94:19–44
Sotelo C (1978) Purkinje cell ontogeny: formation and maintenance of spines. Prog Brain Res 48:149–170
Sotelo C (2004) Cellular and genetic regulation of the development of the cerebellar system. Prog Neurobiol 72:295–339
Sotelo C (2008) Development of “Pinceaux” formations and dendritic translocation of climbing fibers during the acquisition of the balance between glutamatergic and gamma-aminobutyric acidergic inputs in developing Purkinje cells. J Comp Neurol 506:240–262
Sotelo C, Alvarado-Mallart RM (1991) The reconstruction of cerebellar circuits. Trends Neurosci 14:350–355
Sotelo C, Arsenio-Nunes ML (1976) Development of Purkinje cells in absence of climbing fibers. Brain Res 111:289–295
Sotelo C, Changeux JP (1974) Transsynaptic degeneration “en cascade” in the cerebellar cortex of staggerer mutant mice. Brain Res 67:519–526
Sotelo C, Chédotal A (1997) Development of the olivocerebellar projection. Perspect Dev Neurobiol 5:57–67
Sotelo C, Dusart I (2009) Intrinsic versus extrinsic determinants of the development of Purkinje celldendrites. Neuroscience 162:589–600
Sotelo C, Wassef M (1991a) Cerebellar development: afferent organization and Purkinje cell heterogeneity. Philos Trans R Soc Lond B Biol Sci 331:307–313
Sotelo C, Wassef M (1991b) Purkinje cell heterogeneity in four cerebellar mutations revealed by Zebrin-1 expression. Soc Neurosci Abstr 17:918
Sotelo C, Alvarado-Mallart RM, Gardette R, Crepel F (1990) Fate of grafted embryonic Purkinje cells in the cerebellum of the adult “Purkinje cell degeneration” mutant mouse. I. Development of reciprocal graft-host interactions. J Comp Neurol 295:165–187
Soussi-Yanicostas N, de Castro F, Julliard AK, Perfettini I, Chédotal A, Petit C (2002) Anosmin-1, defective in the X-linked form of Kallmann syndrome, promotes axonal branch formation from olfactory bulb output neurons. Cell 109:217–228
Sperry RW (1963) Chemoaffinity in the orderly growth of nerve fiber patterns and connections. Proc Natl Acad Sci USA 50:703–710
Steinmayr M, Andre E, Conquet F, Rondi-Reig L, Delhaye-Bouchaud N, Auclair N, Daniel H, Crepel F, Mariani J, Sotelo C, Becker-Andre M (1998) Staggerer phenotype in retinoid-related orphan receptor alpha-deficient mice. Proc Natl Acad Sci USA 95:3960–3965
Szebenyi G, Callaway JL, Dent EW, Kalil K (1998) Interstitial branches develop from active regions of the axon demarcated by the primary growth cone during pausing behaviors. J Neurosci 18:7930–7940
Takeda T, Maekawa K (1989) Transient direct connection of vestibular mossy fibers to the vestibulocerebellar Purkinje cells in early postnatal development of kittens. Neuroscience 32:99–111
Tanaka S, Ishii K, Kasai K, Yoon SO, Saeki Y (2007) Neural expression of G protein-coupled receptors GPR3, GPR6, and GPR12 up-regulates cyclic AMP levels and promotes neurite outgrowth. J Biol Chem 282:10506–10515
Tanaka S, Shaikh IM, Chiocca EA, Saeki Y (2009) The GS-linked receptor GPR3 inhibits the proliferation of cerebellar granule cells during postnatal development. PLoS ONE 4:e5922
Van Der Giessen RS, Koekkoek SK, van Dorp S, De Gruijl JR, Cupido A, Khosrovani S, Dortland B, Wellershaus K, Degen J, Deuchars J, Fuchs EC, Monyer H, Willecke K, De Jeu MT, De Zeeuw CI (2008) Role of olivary electrical coupling in cerebellar motor learning. Neuron 58:599–612
Voogd J (1969) The importance of fiber connections in the comparative anatomy of the mammalian cerebellum. In: Llinas R (ed) Neurobiology of cerebellar evolution and development. AMA-ERF Institute for Biomedical Research, Chicago, pp 493–514
Wallace VA (1999) Purkinje-cell-derived Sonic hedgehog regulates granule neuron precursor cell proliferation in the developing mouse cerebellum. Curr Biol 9:445–448
Wassef M, Sotelo C (1984) Asynchrony in the expression of guanosine 3′:5′-phosphate-dependent protein kinase by clusters of Purkinje cells during the perinatal development of rat cerebellum. Neuroscience 13:1217–1241
Wassef M, Zanetta JP, Brehier A, Sotelo C (1985) Transient biochemical compartmentationof Purkinje cells during early cerebellar development. Dev Biol 111:129–137
Wassef M, Sotelo C, Thomasset M, Granholm AC, Leclerc N, Rafrafi J, Hawkes R (1990) Expression of compartmentation antigen zebrin I in cerebellar transplants. J Comp Neurol 294:223–234
Wassef M, Angaut P, Arsenio-Nunes ML, Bourrat F, Sotelo C (1992a) Purkinje cell heterogeneity: its role in organizing the topography of the cerebellar cortex connections. In: Llinas R, Sotelo C (eds) The cerebellum revisited. Springer, New York, pp 5–21
Wassef M, Chedotal A, Cholley B, Thomasset M, Heizmann CW, Sotelo C (1992b) Development of the olivocerebellar projection in the rat: I. Transient biochemical compartmentation of the inferior olive. J Comp Neurol 323:519–536
Wassef M, Cholley B, Heizmann CW, Sotelo C (1992c) Development of the olivocerebellar projection in the rat: II. Matching of the developmental compartmentations of the cerebellum and inferior olive through the projection map. J Comp Neurol 323:537–550
Watt AJ, Cuntz H, Mori M, Nusser Z, Sjöström PJ, Häusser M (2009) Traveling waves in developing cerebellar cortex mediated by asymmetrical Purkinje cell connectivity. Nat Neurosci 12:463–473
Wechsler-Reya RJ, Scott MP (1999) Control of neuronal precursor proliferation in the cerebellum by Sonic Hedgehog. Neuron 22:103–114
Yuasa S, Kawamura K, Ono K, Yamakuni T, Takahashi Y (1991) Development and migration of Purkinje cells in the mouse cerebellar primordium. Anat Embryol 184:195–212
Yuasa S, Kawamura K, Kuwano R, Ono K (1996) Neuron-glia interactions during migration of Purkinje cells in the mouse embryonic cerebellum. Int J Dev Neurosci 14:429–438
Zordan P, Croci L, Hawkes R, Consalez GG (2008) Comparative analysis of proneural gene expression in the embryonic cerebellum. Dev Dyn 237:726–735
Acknowledgments
The scientific work of Ferdinando Rossi is supported by grants from Ministero dell’Università e della Ricerca Scientifica e Tecnologica (MIUR-PRIN 2007 prog. nr. 2007F7AJYJ), Compagnia di San Paolo (Neurotransplant Project 2008; GABAGEN Neuroscience project 2009), Regione Piemonte (Project A14/05; Ricerca Sanitaria Finalizzata, 2008, 2009), Ataxia UK, Fondazione Cavaliere del Lavoro Mario Magnetto of Turin. The work of Constantino Sotelo is supported by MINISTERIO CIENCIA E INNOVACION (grants: MICINN BFU-2008-00588 and CSD2007-00023).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media Dordrecht
About this entry
Cite this entry
Sotelo, C., Rossi, F. (2013). Purkinje Cell Migration and Differentiation. In: Manto, M., Schmahmann, J.D., Rossi, F., Gruol, D.L., Koibuchi, N. (eds) Handbook of the Cerebellum and Cerebellar Disorders. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-1333-8_9
Download citation
DOI: https://doi.org/10.1007/978-94-007-1333-8_9
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-1332-1
Online ISBN: 978-94-007-1333-8
eBook Packages: Biomedical and Life SciencesReference Module Biomedical and Life Sciences