
Abstract
Livestock production is thought to be adversely affected by detrimental effects of extreme climatic conditions. Consequently, adaptation, mitigation and amelioration of detrimental effects of extreme climates have played a major role in combating the climatic impact in livestock production. While measures to reduce the growth of greenhouse gas emissions are an important response to the threat of climate change, adaptation to climate change will also form a necessary part of the response. The salient adaptation strategies are developing less sensitive breeds, improving water availability, improving animal health, promoting women empowerment, developing various policy issues, establishing early warning systems and developing suitable capacity building programmes for different stakeholders. Developing adaptation strategies is therefore an important part of ensuring that countries are well prepared to deal with any negative impacts that may occur as a result of climate change. The integration of new technologies into the research and technology transfer systems potentially offers many opportunities to further the development of climate change adaptation strategies. Adapting to climate change and reducing GHG emissions may require significant changes in production technology and farming systems that could affect productivity. Many viable opportunities exist for reducing CH4 emissions from enteric fermentation in ruminant animals and from livestock manure management facilities. To be considered viable, these emission reduction strategies must be consistent with the continued economic viability of the producer and must accommodate cultural factors that affect livestock ownership and management. This chapter also elaborates on ameliorative strategies that should be given consideration to prevent economic losses incurred due to environmental stresses on livestock productivity. Reducing the impact of climatic stresses on livestock production requires multidisciplinary approaches which emphasise animal nutrition, housing and animal health. Therefore, emphasis should be given to all three aspects of adaptation, mitigation and amelioration strategies to sustain livestock production under the changing climate scenario.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
While there is still much uncertainty surrounding the potential magnitude and likely impacts of climate change (CC), there is consensus in the global scientific community that some CC is already occurring and that further change is inevitable (IPCC 2013; Quere et al. 2014). CC is evident in both a change in average temperature and rainfall and changes in the frequency and severity of extreme weather events, such as frosts, heatwaves, droughts and flood (IPCC 2001). It is considered likely that continued greenhouse gas emissions at or above current rates will result in further global warming in this century (WMO 2014). Moreover, even if the atmospheric concentrations of all greenhouse gases and aerosols are stabilised at 2,000 levels, global temperatures are projected to continue rising (IPCC 2007).
While measures to reduce the growth of greenhouse gas emissions are an important response to the threat of CC, adaptation to CC will also form a necessary part of the response (Gerber et al. 2013). In this context, adaptation refers to strategies that act to reduce the adverse impacts of CC. Developing adaptation strategies is therefore an important part of ensuring that countries are well prepared to deal with any negative impacts that may occur as a result of CC. Given limited resources, adaptation strategies must target those populations most vulnerable to global change and equip those unable to adapt—generally the poorest—with the tools and incentives that will enable them to do so. Adaptation to climate variability has been an ongoing necessity for the agricultural sector. Existing strategies to manage climate variability present opportunities for meeting the challenges of future CC. Reducing the impact of CC on livestock requires a multidisciplinary approach with emphasis on animal nutrition, housing and animal health. It is important to understand the livestock responses to environment and analyse them in order to design modifications of nutritional and environmental management thereby improving animal comfort and performance. So a range of technologies are needed to match the different economic and other needs of livestock farmers. It is therefore important that we gear up all efforts to enhance the resilience of the farms and livestock through dissemination—adoption of various coping strategies and mechanisms so that the production and productivity levels of the farm and livestock are maintained even in challenging climatic conditions. While efforts are on to prevent further CC through livestock sectors by developing suitable mitigation strategies, equally important is to reduce the impact of already occurred CC on livestock population by developing suitable amelioration strategies. These consorted efforts need to be implemented simultaneously if we intend to sustain livestock production under the changing climate scenario. This chapter will address the salient adaptation, mitigation and amelioration strategies available to improve livestock production under CC perspectives.
2 Climate Change Affecting Livestock Economy
Several studies have shown significant and alarming negative impacts of CC and adaptation of livestock farmers in different parts of the world (Deressa et al. 2005; Kabubo-Mariara 2007). Various research findings indicate that the damaging effects of global temperature are increasing, and most damages are predicted to occur in the region already faced average high temperatures and low precipitation, frequent droughts and scarcity of both ground and surface water (IPCC 2001). Previous studies on CC and adaptation of livestock farmers have shown that CC affects livestock farming directly and indirectly (Kabubo-Mariara 2008). Direct effects have been observed to include retardation of animal growth, low-quality animal products including hides and skins and animal production in general. Indirect effects include general decline in quantity and quality of feedstuffs, for example, pasture, forage, grain severity and distribution of different species of livestock, and other effects such as increase in livestock diseases and pests. In particular, extreme temperatures resulting in drought have had devastating effects on livestock farming, and the vulnerable rural poor have been left with marginal pasture and grazing lands (Kabubo-Mariara 2005). The impacts of CC not only influence natural systems, habitats and species but also human economy and society. Therefore, governments must act in time to adapt to these changes in order to reduce damage in both natural and social systems and in this way avoid unnecessary costs associated with late action.
3 Significance of Understanding the Impact of Climate Change on Livestock Production
Potential direct and indirect impacts of CC on livestock production have not been thoroughly explored. Changes in crop availability and quality, which have been the primary focus of previous studies, affect animal production through changes in feed supplies. Analyses of direct impacts of CC on livestock production are few. Changes in climate would directly lead to reductions in summer-season milk production and conception rates in dairy cows. Because voluntary feed intake (VFI) is the primary factor influencing the production capacity of livestock, accurate prediction of the feed consumption of livestock under heat stress is a precursor to accurate assessment of changes in production resulting from changes to a warmer climate. Quantification of potential impacts of CC on livestock production allows producers to gain a better understanding of the magnitude of the changes in production levels faced under CC. Projected economic losses resulting from temperature-induced reductions in production may justify mitigation of these temperature increases through changes in management practices, such as installation of shades or sprinklers in feedlots or evaporative cooling of barns.
CC represents one of the greatest environmental, social and economic threats facing the planet today. In developing countries, CC will have a significant impact on the livelihood and living conditions of the poor. Increasing temperatures and shifting rain patterns reduce access to food and create effects that impact regions, farming systems, households and individuals in varying ways. Additional global changes, including changed trade patterns and energy policies, have the potential to exacerbate the negative effects of CC on some of these systems and groups. Thus, analyses of the biophysical and socioeconomic factors that determine exposure, adaptation and the capacity to adapt to CC are urgently needed so that policymakers can make more informed decisions.
4 Salient Adaptation Strategies for Livestock to Cope Up to Changing Climate
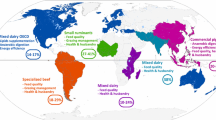
Table 22.1 describes the various adaptation strategies for livestock sector to counter the impact of climate change. Effective adaptation to climate variability and climate change is dependent on access to climate information for the coming seasons and years, to enable communities to make decisions for now and the future. Flexible planning in the face of a continuously changing climate—a key element of adaptive capacity—needs to be informed by climate forecasts and the effects of uncertainties and risks on different vulnerable groups and socio-economic sectors, so as to identify a range of response options. Scenario development of how livelihoods and sectors would be affected by probable climate futures contributes to making livelihoods more climate resilient and can be a first step towards mitigating the effects of climate-related disasters on communities. Figure 22.1 describes the different adaptation, mitigation and amelioration strategies to sustain livestock production under the changing climate scenario.

Strategies to sustain livestock production under changing climate scenario
4.1 Genetic Development of Less Sensitive Breeds
Genetic improvement is an evolutionary action; evolution should be defined as a continuous process of adaptation of the populations of organisms to the ever-changing geological, biological and climatic conditions. Because of the almost infinite number of combinations of environmental factors, organisms must have a great variety of genetic types that can deal with a range of climatic, nutritional or other conditions. In a word, any population must be genetically heterogeneous—i.e. with a great genetic diversity—in order to be able to survive under the challenge of the changing environment. This is the basis for the livestock genetic improvement. Genetic selection has been a traditional method to reduce effects of environment on livestock by development of animals that are genetically adapted to hot climates. There are additional possibilities for meeting this goal. Identification of specific genes that control traits related to thermotolerance makes it possible to select for thermal resistance without inadvertently selecting against milk yield. Possibly, there are also genes controlling cellular resistance to heat shock. Integration of marker-assisted selection into animal breeding systems should make selection for traits conferring thermotolerance more rapid. Functional genomics research is providing new knowledge about the impact of heat stress on livestock production and reproduction. Using functional genomics to identify genes that are up- or downregulated during a stressful event can lead to the identification of animals that are genetically superior for coping with stress and towards the creation of therapeutic drugs and treatments that target affected genes.
There is genetic variation among animals for cooling capability, which suggests that more heat-tolerant animals can be selected genetically. In spite of livestock being reared in tropical environment, there are local indigenous breeds of livestock which can effectively perform countering environmental extremes. These local breeds can perform well in adverse climatic condition like high temperature, drought and feed scarcity. Therefore, even under the changed climate scenario, the rich animal germplasm available may help to sustain the livestock productivity. Cross-breeding offers another opportunity. However, extensive cross-breeding studies have shown little heterosis for heat tolerance. Additional studies are needed to examine variability in heat tolerance of high yielding animals. Possibly improved herds could be developed when selected for milk yield and heat tolerance under local conditions.
In addition, there is a need to take up breeding programmes to develop CC-ready breed which performs better under stress due to climatic variability by using available rich germplasm. The recent advancement in global expression technologies (whole-genome arrays, RNA sequencing) is poised to be effectively utilised to identify those genes that are involved in key regulatory/metabolic pathway for thermal resistance and thermal sensitivity. Gene knockout technology will also allow better delineation of cellular metabolic mechanism required for acclimatisation to thermal stress in sheep and goat. By knowing the various genes responsible for thermotolerance, we can change the genetic structure of animal and drift towards superior thermotolerant ability.
4.2 Improving Water Availability
Water scarcity has become globally significant over the last 40 years or so and is an accelerating condition for one to two billion people worldwide (MEA 2005). Water and its availability and quality will be the main pressures on, and issues for, societies and the environment under CC (Wilk and Wittgren 2009). Nomadic people migrate seasonally to find water and grazing for their herds. Since ancient times, agricultural civilisations have invented techniques for water storage, transfer and irrigation (Wilk and Wittgren 2009). In general terms, the projected impacts of CC on water resources are a continuation of the impacts that have already been observed. At the same time, there are uncertainties of various magnitudes associated with the projections. The findings of several global models should be examined to reduce this uncertainty, especially in regard to regional or local effects (Wilk and Wittgren 2009; Thornton and Herrero 2010).
As more and more stresses are placed on our natural resources through effects of CC, a renewed use of water harvesting would have positive outcomes (Masike 2007). Water harvesting is a technique of developing surface water resources that can be used in dry regions to provide water for livestock, for domestic use, and for agroforestry and small-scale subsistence farming (Thames and Cluff 1982). Water harvesting offers a method of effectively developing the scarce water resources of arid regions. It is also a relatively inexpensive method of water supply that can be adapted to the resources and needs of the rural poor. A successful harvesting system must be (a) technically sound, properly designed and maintained, (b) economically feasible for the resources of the user and (c) capable of being integrated into the social traditions and abilities of the users.
The simplest technique is to use water-ponding dikes which slow down surface runoff, allow infiltration and increase soil moisture, and promote significant vegetation growth for habitat cover and forage. The advantages of water-ponding dikes are that they are simple to install and cost-effective and make use of water that would be lost to evaporation. The use of stock tanks as water sources on rangeland for cattle grazing is a traditional method that has one of the least expensive construction costs among a variety of possible methods. In arid regions, it places reliance on trapping surface runoff that will otherwise be lost back into the atmosphere through evaporation. By confining this surface runoff in a pond with small surface area and water depth of up to 2 m, evaporation and infiltration losses are both reduced over what would normally occur if the water was spread out and infiltrated into a stream bed.
The principles of integrated water resources management (IWRM) should be promoted and guide climate adaptation strategies. IWRM provides a useful framework for planning well-coordinated and targeted adaptation measures to CC. It is a systematic process to the sustainable development and equitable allocation of water resources through a holistic approach to water management. Successful IWRM strategies include, among others, capturing societal views, reshaping planning processes, coordinating land and water resources management, recognising water quantity and quality linkages, combining the use of surface water and groundwater, protecting and restoring natural systems, addressing impediments to the flow of information, and considering CC (Wilk and Wittgren 2009).
National adaptation programmes for action need to be integrated with other national development plans and adopt a river basin perspective, including transboundary cooperation in cases of multinational rivers. The local level is crucial in climate adaptation, and institutional reforms must be crafted accordingly. Mechanisms should be put in place to make sure that adaptation efforts respect, protect and promote fundamental human rights. Fresh and flexible funding should be found to speed up investment in water management of vulnerable developing countries, in particular the least developed countries, to meet both the present Millennium Development Goals and the consequences of observed and projected CC.
4.3 Improving Animal Health
As a result of globalisation and CC, the world is currently facing an unprecedented increase of emerging and re-emerging animal diseases and zoonoses (animal diseases transmissible to humans). A changing climate will cause changes in the patterns of endemic diseases in livestock. Indeed, a better understanding of the effect of CC on animal health and production is crucial and good for recommendations on how to lessen its potential impact. Unfortunately, the determinants of resilience and adaptation that already reduce this impact are often poorly understood even though they are not unique but are needed regardless. For example, adaptive capacity could be increased in the broader context of developing appropriate policy measures and institutional support to help the livestock owners to cope with all animal health problems. In fact, the development of an effective and sustainable animal health service, with associated surveillance and emergency preparedness systems and sustainable animal disease control and prevention programmes, is perhaps the most important and most needed adaptive strategy. This will safeguard livestock populations from the threats of CC and climate variability.
Improving the governance of animal health systems in both the public and private sector is the most effective response to this alarming situation. The recently experienced animal disease crises have provided a clearer understanding of the benefits to the international community of applying the appropriate animal health policies and programme in order to safeguard public health and ensure food safety.
The animal health service of FAO addresses four animal health-related issues: transboundary diseases, vectorial diseases, veterinary public health (including food safety) and veterinary services. The latter must join forces and encourage the more active participation of the private sector defining complementary roles for each with specific responsibilities in order to improve and/or maintain the overall sanitary status of a country. The following points are very valuable if one considers tackling disease outbreaks that arise as a result of CC:
-
(a)
Improved monitoring and surveillance are required to detect changing patterns of diseases and to respond accordingly.
-
(b)
Improved diagnosis, forecasting and effective vaccines to protect our livestock from this increasing threat of disease.
-
(c)
Combine hazard maps with vulnerability maps for better prioritisation of areas/populations for interventions.
-
(d)
Development of ideal decision support frameworks involving development of risk maps for targeted surveillance and development of prediction models for outbreaks.
-
(e)
Institutional measures need to be sensitised for disease surveillance, and integration of experts of climate and health working groups needs to be prioritised for developing disease control technologies.
-
(f)
Potential interventions are needed in developing livestock value chain involving development of diversified livelihood options and safety nets comprising insurance schemes.
-
(g)
Understanding in depth the epidemiology of important disease-causing pathogens and diagnostics should be interfaced with very sophisticated geographic information system technology and both statistically and process-based modelling approaches to produce risk maps for disease at an appropriate temporo-spatial scale (e.g. Bergquist and Rinaldi 2010; Fox et al. 2012).
-
(h)
Validate models and quantify trends, through improved surveillance, ideally active rather than passive, of animal health disease issues, whether exotic or endemic. This places a requirement on improved diagnostic capabilities, with the potential to be rapid, high-throughput and cost-effective at an appropriate regional/national/international scale (Skuce et al. 2013).
-
(i)
In the context of the endoparasites and ectoparasites specifically, vaccines to protect animals from infection, although not a complete solution, would be highly desirable. However, development of such vaccines has proven to be extremely technically demanding for a number of reasons (e.g. Vercruysse et al. 2007).
4.4 Female Empowerment
Division of rights and responsibilities affects incentives and ability to adopt new technologies and practices to increase production and productivity. We need to understand this better to develop appropriate technologies and design more effective interventions. In recent years, there have been increasing concerns about CC and its impacts on food security and on human lives. While much of the earlier discussions on CC centred on scientific and technical aspects such as greenhouse gas emissions, ozone depletion and environmental impacts of global warming, attention is now increasingly encompassing the economic and social aspects of CC. Women and the poor are likely to be most affected by CC. The constraints they face in crop-livestock systems are not being adequately addressed in research programmes due to the lack of a systematic approach and low capacities to integrate gender in a meaningful way.
Participatory research involving women in livestock sector should be increased. Women play important roles as producers of food, managers of natural resources, income earners and caretakers of household food and nutrition security. The control of assets including livestock and income derived from these assets by women has positive consequences for their decision-making within the household and for household well-being. Women play a much stronger role than men in the ecosystem management services and food security. Women are powerful agents of change and their leadership is critical. They have a significant role in dealing with issues such as energy consumption, deforestation, burning of vegetation, population growth, economic growth, developing scientific research and technologies and policy making, among others.
Women play a critical but often overlooked role in livestock production. Livestock are the most important asset for women. But unfortunately, most of the livestock are owned by men. With respect to livestock, women are heavily involved in small ruminant production, and it is easy for them to get into production. They need information on improved feeding practices, management practices and marketing. Hence, consorted efforts are needed in defining the complexities and changes associated with livestock ownership. This will ensure closing of gender asset gap. Considerable efforts are also needed to define the exact role of women in livestock keeping and encouraging women’s participation in livestock markets. Further efforts are needed to integrate women, livestock, nutrition and health. Proper mechanisms should be put in place for securing women’s access and control of livestock and other assets. This can be easily achieved by creating index-based livestock insurance for women and women-owned livestock as well as by developing a separate fund for both women and livestock. Promoting women’s participation in both formal and informal livestock and livestock product markets can easily improve the confidence of women to take livestock as their primary livelihood activities.
4.5 Developing Policy Issues
The successful implementation of adaptation strategies to CC requires government support in terms of support by developing various policy issues. According to Antle (2010), there are two sets of policies that need to be developed. The first set of policies for designing adaptation strategies comprises (a) agriculture subsidy and trade policies, (b) production and income insurance policies and disaster assistance, (c) soil and water conservation policies and ecosystem services, (d) environmental policies and agricultural land use, (e) tax policies, (f) energy policies and (g) greenhouse gas mitigation policies. The second set of policies is concerned to facilitate adaptation. This comprises (a) estimating adaptation costs and reassessing impacts, (b) identifying adaptation strategies and related research needs, (c) identifying and estimating the vulnerability of ecosystem services to CC and adaptive responses, (d) providing public information about long-term climate trends and their economic implications and (e) implications of climate change and mitigation policies for agriculture and food sector.
4.6 Climate Change Communication and Early Warning System
Strengthening climate information and early warning systems (EWS) for climate-resilient development and adaptation to CC is to help farmers respond to both short-term/rapid-onset climatic hazards (e.g. cyclones, floods and storms) and long-term/slow-onset hazards (e.g. drought and long-term CC). To enhance the resilience of the population and the national economy, urgent action and measures needed to be taken to address the deficiencies and strengthen CC early warning system. The expected outcomes from the early warning systems are (a) enhanced capacity of hydrometeorological services and networks for predicting CC events and risk factors, (b) effective, efficient and targeted delivery of climate and CC information including early warnings and (c) improved and timely preparedness and responses of various stakeholders to forecast climate-linked risks and vulnerabilities. Further, the success of the early warning system depends on the following four elements: (1) risk knowledge, systematically collecting data and undertaking risk assessments; (2) monitoring and predicting, developing hazard monitoring and early warning services, including weather and hydrological monitoring equipment, improving forecast capabilities and the use of these technologies within agricultural advisories, flood-risk monitoring and supply chain management; (3) disseminating information, communicating risk information and reliable warnings to potentially affected locations through traditional and new media; and (4) responding to warnings, building national and community response capabilities to act effectively when warnings are received.
4.7 Capacity Building Programme
The success of adaptation strategies depends upon how effectively those strategies are being transferred to the ultimate target groups, the poor and marginal farmers. There is a need to improve the capacity of livestock producers and herders to understand and deal with climate change increasing their awareness of global changes. In addition, training in agroecological technologies and practices for the production and conservation of fodder improves the supply of animal feed and reduces malnutrition and mortality in herds. Developing suitable capacity building programme (CBP) is very crucial for successful implementation of adaptation strategies. CBP should address in detail the basic understanding of climate change science, climate change impacts on biodiversity and ecosystems and implication for conservation and sustaining eco-services and strategies for assessing vulnerability and adaptation. Figure 22.2 describes the various approaches and components of CBP. CBP (a) should be country driven and issue based, (b) should occur within a framework of integrated interdisciplinary problem solving, (c) is much more than training which requires institutional strengthening and human resource development and (d) should encourage potential for interaction and dialogues among diverse stakeholder groups. Building capacity of different stakeholders in parallel with the above adaptation strategies would help widen research and development communities to put climate, vulnerability and adaptation information into active use for pro-poor development.

Different approaches for capacity building programme
5 Mitigation Strategies to Reduce Livestock-Contributed Climate Change
5.1 Management Strategies
Any methane reduction strategies must be confined to the following general framework, viz. development priority, product demand, infrastructure, livestock resource and local resources (Sejian et al. 2012a). The most attractive emission mitigation projects must balance the needs in all of these areas, so that no one factor creates a constraint on continued improvement in production efficiency and the resulting CH4 emission reductions (Hristov et al. 2013). Within this framework, CH4 emission mitigation options for enteric fermentation can encompass a wide range of activities across these areas. However, underlying these activities must be specific options for improving the production efficiency of the livestock. Without these options, CH4 emissions cannot be reduced. Best management practices are defined as those that (a) minimise and mitigate impacts and risks to the environment by maintaining or improving quality of soil, water, air and biodiversity; (b) ensure the health and sustainability of natural resources used for agriculture production; and (c) support long-term economic and environmental viability of the agriculture industry. There are several management strategies that may be employed for livestock to reduce enteric methane emissions. Adoption of the basic livestock management principles offers the best opportunity of improving production efficiency while also reducing emissions. Within the context of these fundamental livestock management principles, specific techniques for improving production efficiency and reducing CH4 emissions include the following (USEPA 1993): (1) enhanced nutrition through mechanical and chemical feed processing, (2) balanced nutrition through strategic supplementation, (3) intake of production-enhancing agents, (4) improved production through enhanced genetic characteristics, (5) increased production efficiency through improved reproduction and (6) improved grassland and rangeland management.
5.1.1 Animal Manipulation by Reducing Livestock Numbers
The countries which are committed to reduce the enteric methane emission from the livestock, reducing animal number is the best possible way, but it is totally unacceptable for those countries which rely heavily on livestock production for their national economy (Sejian et al. 2011a). Shifting of old-age cattle from heifers in the cattle herds can efficiently increase the productivity and decrease the enteric methane production due to high intake and passage rate of ingested feed materials that can lead to lower enteric emission. Hegarty (2001) points out that if animal numbers do not decrease in response to the improved productivity, then emissions from the sector will increase rather than decrease. Sheep population has been reduced from 57.9 million in 1990 to 45.2 million in 2000, while dairy cattle and beef cattle population has increased slightly. The net outcome was a decline in ruminant CH4 emission from 1.45 to 1.31 Tg/year from 1990 to 2000 (Sejian et al. 2011a). The application of biotechnological aids can meet out the loss of reduced animal number, for example, the use of recombinant bST (bovine somatotropin) leads to an increase in milk production up to 10–20 %, and therefore, animal number can be reduced to lower the total enteric emission (Clemens and Ahlgrimm 2001). Kirchgessner et al. (1995) estimated that overall CH4 emissions could be decreased by reducing animal numbers while maintaining milk production.
5.1.2 Animal Breeding with Low CH4 Emissions
Genetic selection of animals that consume less feed or produce less CH4 per unit of feed is a management strategy that may be used to reduce enteric CH4 emissions. Pinares-Patino et al. (2007) established that there are differences between individual animals in the quantity of CH4 they emit per unit of dry matter intake. This finding has resulted in the establishment of research programmes aimed at exploiting these differences. Animal effects on fermentation could be via the saliva, feed processing (e.g. comminution) or flow rate through the rumen. It is possible that the animal’s impact on fermentation is genetically determined, and if this is the case, it may be possible to obtain markers that can be used to select low methane emitters.
5.1.3 Increasing the Efficiency of Livestock Production
Improving the efficiency of ruminant animal performance will generally lead to a reduction of CH4 emitted per unit of animal product. There are two aspects of this: genetic improvement of the animals themselves to achieve more product per unit of feed intake, as has been achieved with pigs and poultry, and nutritional manipulation via increased feed intake and appropriate feed composition. This can be achieved in three ways: (a) increasing feed intake: increasing feed intake decreases the methane emission per unit of feed intake. Kirchgessner et al. (1995) reported that as milk yield increases, methane emitted per unit of milk yield decreases. By feeding animals ad libitum, it is possible to both maximise efficiency and reduce methane emission per unit of product. This is because as intake increases, the methane emission associated with the essential, but non-productive, requirements for maintenance is diluted. By improving animal production efficiency, emissions per unit product can be reduced by 25–75 % depending on animal management practices (Bowman et al. 1992). In addition, improved productivity can allow managers to reduce the size of the herd necessary to produce a certain quantity of product (O’Mara 2004); (b) nutritional manipulation: as described above, decreasing dietary fibre and increasing starch and lipid will reduce methane emission. Generally, diets of higher digestibility have these characteristics. Generally, dairy cows were given feeds of increasing digestibility to achieve the same level of milk production. The animals would have eaten less of the higher digestibility diets and thus produced less total methane and reduced methane emitted per unit of milk produced. Improving the nutritive value of the feed given to grazing animals by balancing the diet with concentrates, or by breeding-improved pasture plants, should result in reduced methane emission; and (c) metabolic efficiency by production-enhancing agents: production-enhancing agents are available for use to increase production efficiency in cattle. Bovine somatotropin (bST) for dairy cows is a naturally occurring growth hormone produced by the pituitary gland. Recombinant bST, an identical molecule, is produced biotechnologically and has been shown to increase milk production in US dairy cows. In general, the use of bST leads to an increase in milk production of 10–20 %, and therefore, animal numbers can be reduced to lower total enteric emissions (Clemens and Ahlgrimm 2001). Johnson et al. (1996) estimated that the use of bST to improve US dairy cattle productivity could result in decreased CH4 emissions (% of GEI) by about 9 %.
5.1.4 Grazing Management
Implementing proper grazing management practices to improve the quality of pastures will increase animal productivity and lower CH4 per unit of product. McCaughey et al. (1997) observed that CH4 production was greatest for steers continuously grazing at low stocking rates (1.1 steer ha–1; 306.7 L d–1) and least for steers grazing continuously at high stocking rates (2.2 steers ha–1; 242.2 L d–1). At higher stocking rates, forage availability and intake are low. When pastures were rotationally grazed, stocking rates had no effect on CH4 production (L d–1) (McCaughey et al. 1997). At low stocking rates (1.1 steer ha–1), CH4 production (L ha–1 d–1) was 9 % lower on rotational grazing than continuous grazing (McCaughey et al. 1997). Measurements of CH4 production from grazing beef cows indicated a 25 % reduction in CH4 losses with alfalfa-grass pastures (7.1 % of GEI) compared to grass-only pastures (9.5 % of GEI) (McCaughey et al. 1999). Early grazing of alfalfa-grass pastures reduced CH4 production (% GEI) by 29–45 % in steers compared to grazing at mid and late seasons (Boadi et al. 2004). In most productive husbandry systems, the herbage digestibility tends to be maximised by agricultural practices such as frequent grazing and use of highly digestible forage cultivars. Consequently, in these systems, the primary factor which modifies the C flux returned to the soil by excreta is the grazing pressure, which varies with the annual stocking rate, i.e. mean number of livestock units per unit area (Soussana et al. 2004). Secondary effects of grazing on the C cycle of a pasture include: (1) the role of excretal returns which, at a moderate rate of grazing intensity, could favour nutrient cycling and increase primary production, especially in nutrient-poor grasslands (De Mazancourt et al. 1998) and (2) the role of defoliation intensity and frequency and of treading by animals, which both reduce the leaf area and then the atmospheric CO2 capture.
5.1.5 Improving Nutrition
Methane production was reduced (−7 % and −40 %) by increasing DMI and the proportion of concentrate in the diet (Rowlinson et al. 2008; Kingeston-Smith et al. 2010). The use of more digestible forage (less mature and processed forage) resulted in a reduction of methane production (−15 % and −21 %). Methane production was lower with legume than with grass forage (−28 %). Legumes generally have higher dry matter intakes and produce more milk solids. This reduces methane emissions per unit of milk or meat production. Methane emissions are also commonly lower with higher proportions of forage legumes in the diet, partly because of the lower fibre content, the faster rate of passage, and in some cases, the presence of condensed tannins (CTs) (Beauchemin et al. 2008). Forage quality has a significant impact on enteric CH4 emissions (Ulyatt et al. 2002; Sejian et al. 2011b). There is also evidence that using clovers and grasses with high water-soluble carbohydrates (WSC) in animal diets can directly reduce methane emissions (Lovett et al. 2004). It has been demonstrated that increasing the WSC content in perennial ryegrass by 33 g/kg reduces methane production in vitro by 9 % (Rowlinson et al. 2008). Also, increasing the proportion of concentrate in the diet will generally reduce rumen pH and as methanogens are pH sensitive; this will also tend to reduce methane emission. The proportion of concentrate within the diet has been reported to be negatively correlated with methane emissions (Lovett et al. 2005; McAllister and Newbold 2008). An increase in feeding level also was reported to induce lower CH4 losses. This is caused mainly by the rapid passage of feed out of the rumen (Mathison et al. 1998; Hegarty 2001). Grinding or pelleting of forages to improve the utilisation by ruminants has been shown to decrease CH4 losses per unit of feed intake by 20–40 % when fed at high intakes (Johnson et al. 1996). The lowered fibre digestibility, decreased ruminally available organic matter and faster rate of passage associated with ground or pelleted forages can explain the decline in CH4 production (Le-Liboux and Peyraud 1999). In summary, there are several promising strategies to reduce methane emissions through forage selection, but these need further investigation, particularly at a whole-farm level.
5.1.6 Improved Grassland and Rangeland Management
Increasing the digestibility of cell walls in forages has been suggested as a means to lower CH4 losses, but in fresh grass and grass silage, the scope of this approach appears to be rather limited. There is evidence that fresh grass results in lower CH4 losses than grass silage, but no direct comparisons exist between fresh grass and grass silage. Mainly forage diets are often supplemented with sugar-based concentrates to provide a rapidly available source of energy for the rumen microbes or to increase the palatability of the diet DM and hence stimulate the digestibility (Mills et al. 2001). CH4 production in ruminants tends to increase with maturity of forage fed, and CH4 yield from the ruminal fermentation of legume forages is generally lower than the yield from grass forages (McAllister et al. 1996; Moss et al. 2000). Shifting the animals from grass to legume plant species tends to decrease the enteric emission due to lower proportion structural carbohydrates and faster rate of passage which shifts the fermentation pattern towards higher propionate production (Johnson and Johnson 1995). Continuous grazing of improved pasture managed with a high stocking rate (2.2 steers ha-1) resulted in 21 % lower daily CH4 emissions as compared to the low stocking rate (1.1 steers ha-1) when measured as emissions per animal per day, but these differences were not evident when measured as emissions per unit gain or as % GEI (McCaughey et al. 1997).
5.1.7 Longevity/Extended Lactation
Extended lactation can reduce the energy demand of cows and methane by approximately 10 %. Extended lactation has other benefits, such as reducing peak workload; cow health problems (due to less calving) and less heifer replacements are required. Milk in the extended lactation phase is higher in milk solids, making the milk more valuable per litre and there may be price incentives for milk produced outside peak supply months. Extended lactation is always considered as an option for herd, and selected breeds suited for extended lactation. The longer the cows stay in a herd, the lower the number of replacements required, and thus, the lower the total farm methane emissions. An example of a 100-cow farm is where the average number of lactations varies from 2.5 to 5. It is assumed that dairy cow emissions are 118 kg/year, while the rearing of a replacement heifer to calve at 2 years old results in methane emissions of 100 kg. This shows that total farm emissions of CH4 from enteric fermentation decline from 15,800 to 13,800 kg/year (0.127 less) as the average number of lactations increases from 2.5 to 5. This does not factor in the higher yield of the older cows which would further reduce emissions per kg of milk. Thus, any measures which reduce involuntary culling should be encouraged.
5.2 Nutritional Interventions
Methanogenesis is essential for an optimal performance of the rumen because it avoids hydrogen accumulation, which would lead to inhibition of dehydrogenase activity involved in the oxidation of reduced cofactors. Fermentation is an oxidative process, during which reduced cofactors (NADH, NADPH, FADH) are reoxidised (NAD1, NADP1, FAD1) through dehydrogenation reactions releasing hydrogen in the rumen. As soon as produced, hydrogen is used by methanogenic archaea, a microbial group distinct from Eubacteria, to reduce CO2 into CH4 according to the depicted equation. Acetate and butyrate production results in a net release of hydrogen and favours CH4 production, while the propionate formation is a competitive pathway for hydrogen use in the rumen. It was established that CH4 production can be calculated from stoichiometry of the main volatile fatty acid (VFA) formed during fermentation (Demeyer and Fievez 2000). The metabolic pathways involved in hydrogen production and utilisation as well as the methanogenic community are important factors that should be considered when developing strategies to control CH4 emissions by ruminants. Any given strategy has to address one or more of the following goals: (1) reduction of hydrogen production that should be achieved without impairing feed digestion and (2) stimulation of hydrogen utilisation towards pathways producing alternative end products beneficial for the animal and/or an inhibition of the methanogenic archaea (numbers and/or activity). Table 22.2 describes the potential mitigation strategies to reduce enteric methane emission.
5.2.1 Feeding Management
Feeding management is one of the most important strategies for CH4 mitigation in ruminants. An integrated approach that considers the rumen microbiota, the animal and the diet seems the best approach to find a long-term solution for reducing enteric CH4 production by ruminants. Both the amount of digestible nutrients ingested and the composition of the diet were found to be major factors governing methane production (Blaxter and Clapperton 1965). The developed equations (Yates et al. 2000), demonstrate that increasing the energy density of the diet (e.g. by increasing ratio of concentrates to forage) decreases methane production per unit of digestible energy ingested. Increasing energy density also increases productivity, thereby also contributing to decreased carbon per unit of product.
5.2.2 Inducing Acetogenesis and Feeding Probiotics
Reductive acetogenesis is a natural mechanism of hydrogen utilisation that coexists with methanogenesis in the gastrointestinal tract of many animals. This pathway is also the dominant one in several hindgut-fermenting mammals (human, rabbit, hamster, rat) but also in foregut fermenting such as kangaroos (Klieve and Joblin 2007). The final product of the reaction, acetate, has the additional advantage of being a source of energy for the animal. However, in the rumen environment, acetogens are less numerous and less efficient than methanogens in the competition for reducing equivalents. This is probably because acetogens need a higher concentration of hydrogen in the medium to reduce CO2 into acetate than that required for methanogens to reduce CO2 into CH4. In addition, the former reaction is thermodynamically less favourable (Weimer 1998). Attempts to increase the natural rumen population of acetogens have been assayed but without success (Van Nevel and Demeyer 1996). The use of acetogens as probiotics has also been tested by several authors with and without the addition of methanogen inhibitors to favour competition (Nollet et al. 1998; Lopez et al. 1999). Live yeast, the most commonly used probiotic in ruminant production, has not been extensively tested for their effect on CH4 production (Chaucheyras-Durand et al. 2008). The few reports available used strains selected for effects other than CH4 reduction, and the results are contradictory (Doreau and Jouany 1998; Chaucheyras-Durand et al. 2008).
5.2.3 Defaunation
Defaunation is the complete removal of protozoa from the rumen ecology and consequently reduces methane release by 20–30 % (Kreuzer et al. 1986). Association and cross-feeding between ruminal protozoa and archaea have been established and are the basis for suggesting defaunation as a CH4 mitigation strategy. Defaunation also markedly increases the total bacterial number, whereas it reduces the number of methanogens which may be due to the loss of preferable colonisation sites for them at which they associated ecto- and endosymbiotically with protozoa. Ruminal protozoa may also play an important role in methane production, particularly when cattle are fed high-concentrate diets. Ruminal methanogens have been observed attached to protozoal species suggesting possible interspecies hydrogen transfer. However, the response in CH4 production to partial or complete defaunation has been variable. Morgavi et al. (2010) calculated 10 % decrease in CH4 production due to defaunation, but the data from that study were extremely variable. Research with beef cattle reported no effect on rumen methanogen abundance despite a 65 % difference in protozoal numbers between a high-forage and a high-starch, lipid-supplemented diet (Popova et al. 2011). Similarly, a 96 % reduction in ruminal protozoa had no effect on methanogenic archaea in dairy cows treated with lauric acid (Hristov et al. 2011). Apart from lauric acid and coconut oil (Hristov et al. 2009, 2011; Hollmann and Beede 2012), and some vegetable oils with a high proportion of unsaturated fatty acids such as linseed (Doreau and Ferlay 1995), there have been no effective and practical defaunating agents tested comprehensively in vivo. Defaunation of the rumen of cattle fed a barley diet decreased methane production by approximately one half (Whitelaw et al. 1984). However, defaunation of animals receiving high-forage diets (Itabashi et al. 1984) did not reduce methane losses.
5.2.4 Mitigation Through Chemical Inhibitors
5.2.4.1 Nitrates
Leng (2008) provided a comprehensive review of the earlier literature on nitrates. Recent research with sheep (Nolan et al. 2010; Van Zijderveld et al. 2010) and cattle (Van Zijderveld et al. 2011; Hulshof et al. 2012) has shown promising results with nitrates decreasing CH4 production by up to 50 %. Nitrate persistently decreased CH4 production from lactating dairy cows during 4 successive 24-day periods (Van Zijderveld et al. 2011). In a short-term study, addition of 22 g nitrate/kg DM in the diet reduced methane emission by 32 % in Nellore×Guzera beef steers fed with sugarcane-based diets. However, in practice, the use of nitrate should be limited to diets naturally low in protein, in which addition of an NPN source is favourable (Hulshof et al. 2012). Nitrates may serve as a terminal electron acceptor and therefore may behave as alternate hydrogen sink and can be converted to ammonia and used in the rumen as a source of nitrogen. Nitrates (NO3) themselves are not very toxic, but nitrites (NO2), which they are converted to, are toxic. The nitrate and nitrite along with CO2 are the hydrogen acceptors in the rumen. Microorganisms are able to increase nitrate reductase activity in the rumen three to five times the normal values, with 3–5 day acclimatisation period, but toxicity at larger dose limits its application. The harmful effect is due to the oxidising properties of nitrite, whereby blood haemoglobin is converted into methaemoglobin, which is a poor oxygen carrier, resulting in anoxia in the animals. This methaemoglobin can be decreased by the β 1-4 galacto-oligosaccharides which are also proved to be methane inhibitors (Sar et al. 2004).
5.2.4.2 Sulphates
In the rumen, sulphate is reduced to sulphide. As hypothesised for marine sediments, sulphate-dependent methane oxidation with the use of methane and/or acetate is thermodynamically more favourable in anaerobic conditions like rumen. Sulphate-reducing bacteria (SRB) in the large intestine of human and pig outcompete those of methanogenic bacteria (MB) and thus reduce methane production. As free sulphate levels in digesta appear to affect the relationship between MB and SRB, sulphate levels in the rumen are likely to be insufficient for SRB to outcompete MB. Stoichiometric calculations show that reducing methane emissions in a sheep by 50 % would require ingestion of 0.75 moles of sulphate or nitrate per day. Adding sulphate to the diet of sheep also reduced CH4 production, and when both nitrate and sulphate were added, the effect on CH4 production was additive (Van Zijderveld et al. 2010).
5.2.4.3 Organic Acids
Fumaric and malic acids, the direct metabolic precursors of propionate, have also been studied as alternative hydrogen sinks in the rumen (Molano et al. 2008; Foley et al. 2009; Van Zijderveld et al. 2011). Inclusion of malic and fumaric acids or their sodium salts or the intermediates of carbohydrate degradation in diets results in shifting rumen fermentation towards propionate and hence less methane production (Castillo et al. 2004). Addition of sodium fumarate consistently decreased methane production in vitro by 2.3–41 % (Ungerfeld et al. 2007). Similarly, malate, which is converted to fumarate in rumen, stimulated propionate formation and also inhibited methanogenesis in some in vitro studies (Carro and Ranilla 2003; Tejido et al. 2005), although other studies have failed to find clear reductions of methanogenesis in vitro (Gómez et al. 2005; Ungerfeld and Forster 2011). In vivo effects of adding organic acids such as fumarate, aspartate and malate or oxaloacetate to the diet on methane mitigation are quite variable (Ungerfeld et al. 2003; McGinn et al. 2004). Wood et al. (2009) noted 60–76 % reductions in methane emissions by supplementing fumarate at 100 g/kg to growing lambs, while Foley et al. (2009) observed the reductions of only 6 % and 16 %, when the diet of beef heifers was supplemented with malic acid at 37.5 and 75 g/kg, respectively. In contrast, no effects of fumaric or malic acid on methane emissions were observed in other studies (Beauchemin and McGinn 2006; Foley et al. 2009). The effect of organic acid supplementation on methane reduction appears to be influenced by the roughage to concentrate ratio and the type of cereal grain being fed in diet (Carro and Ranilla 2003; Gómez et al. 2005; Tejido et al. 2005). Newbold and Rode (2005) tested 15 potential precursors of propionate, including pyruvate, lactate, fumarate, acrylate, malate and citrate in short-term batch cultures. Sodium acrylate and sodium fumarate produced the most consistent effect decreasing CH4 production by 8–17 %. Free acids rather than salts were more effective in reducing CH4.
5.2.4.4 Ionophores
Among ionophore antibiotics, monensin is the most studied in ruminants, besides lasalocid, salinomycin, nigericin and gramicidin. Monensin specifically targets bacteria producing H2 and formate. It reduces the amount of H2 available for methanogenic bacteria and attaches to the cell membrane of ruminal bacteria and protozoa, resulting in a decrease in the proportion of acetate relative to propionate in the rumen and thereby effectively lowering CH4 production by up to 76 % in vitro and to an average of 18 % in vivo (Van Nevel and Demeyer 1996; Hristov et al. 2003). However, the inhibitory effect on methane production appears to be dose dependent. Odongo et al. (2007) reported that feeding monensin to dairy cows (24 mg/kg diet) caused 7 % reduction in methane compared to control animals and the reductions sustained for 6 months with no adaptation detected. In a meta-analysis of 22 controlled studies, monensin (given at 32 mg/kg DM) reduced CH4 emissions by 19 ± 4 g/animal/day (P < 0.001) in beef steers (Appuhamy et al. 2013). The corresponding reductions in dairy cows were 6 ± 3 g/animal/day (P = 0.065) for monensin given at 21 mg/kg DM. Overall, the conclusion of that analysis was that monensin had stronger anti-methanogenic effect in beef steers than in dairy cows (mostly fed forage-based diets), but the effects in dairy cows can be improved by dietary modifications and increasing monensin dose. Another meta-analysis has shown a consistent decrease in acetate: propionate ratio with monensin addition in high grain diets fed to beef cattle (Ellis et al. 2012), which may lead to a reduction in CH4 emission per unit of feed. Hook et al. (2009) observed that long-term (up to 6 months) monensin supplementation did not affect either the number or diversity of methanogens in the rumen of dairy cows, confirming that monensin is able to suppress methanogenesis through an indirect effect on methanogens which may be due to inhibition of protozoa, which produce hydrogen and are colonised by methanogens. Ionophores inhibit Gram-positive microorganisms responsible for supplying methanogens with substrate for methanogenesis and reduce protozoal numbers (Hook et al. 2009). There can be a risk that ionophores get absorbed from rumen and reach animal products (i.e. meat or milk). So, it has been banned in the European Union since January 2006 and it is being currently strongly questioned in many countries.
5.2.4.5 Halogenated Compounds
Halogenated methane analogues such as BCM, chloral hydrate, amichloral, 2-bromoethane sulphonate, chloroform and cyclodextrin are highly effective at reducing methane production, though methanogen species differ in their responsiveness (McAllister and Newbold 2008; Mitsumori et al. 2011; Knight et al. 2011). Though these compounds can be highly effective, the effect of these chemicals is transitory with no significant long-term reduction in methane production (McAllister and Newbold 2008). The BCM can inhibit methanogenesis by reacting with coenzyme B, which functions at the last step of the methanogenic pathway. Recently, Abecia et al. (2012) confirmed the methane-reducing effects of BCM in lactating dairy goats and reported a 33 % reduction with no effect on rumen bacteria, protozoa and methanogens. Some compounds such as bromine analogue of coenzyme M were potent methane inhibitors in vitro, but the inhibition was not persistent in vivo, suggesting adaptation of methanogenic populations (van Nevel and Demeyer 1995). An adaptation of methanogens to quaternary ammonium compounds has also been demonstrated (Tezel et al. 2006). Data by Knight et al. (2011) showed an immediate and dramatic drop in CH4 production in dry cows administered with chloroform; however, CH4 production gradually increased to about 62 % of the pretreatment levels by day 42, suggesting adaptation to chloroform by the rumen ecosystem, but in contrast, the effect of BCM appeared to persist in the studies by Tomkins et al. (2009) and Abecia et al. (2012). However, a banned compound, such as BCM (an ozone-depleting agent), cannot be recommended as a CH4 mitigating agent, but compounds with similar mode of action could be developed.
5.2.4.6 Prebiotics
In ruminants, the role of prebiotics has not yet been quantified, but they are used in rumen manipulation along with nitrate and probiotics and had potential to reduce methane production. They are speculated to enhance the propionate production by stimulating Selenomonas, Succinomonas and Megasphaera with simultaneous inhibition of acetate producers such as Ruminococcus and Butyrivibrio (Mwenya et al. 2004). Administration of galacto-oligosaccharide (GOS) supplementation decreased nitrite accumulation in rumen and plasma and nitrate-induced methaemoglobin while retaining low methane production. Eleven percent reduction in methane emission (litres/day) in GOS-supplemented diet compared to control diet has been reported (Zhou et al. 2004). Inclusion of GOS increased propionate production and decreased CH4 (Santoso et al. 2004).
5.2.4.7 Nitro compounds
Growth of nitro-metabolising bacterium Denitrobacterium detoxificans was supported by the electron acceptors such as 3-nitropropanol, 3-nitropropionate, nitrate, 2-nitropropanol, nitroethane, nitroethanol or 3-nitro-1-propyl-β-D-glucopyranoside (miserotoxin). These less toxic nitro compounds may have the potential to be used as an alternative to decrease CH4 production in ruminants (Gutierrez-Banuelos et al. 2007; Brown et al. 2011). In the presence of the appropriate nitro compound, formate, lactate and H2 served as electron donors of D. detoxificans. Nitroethane is a potent anti-methanogenic compound on three gastrointestinal microbial sources (i.e. chicken ceca, bovine rumen and ovine rumen). Nitroethane has been safely administered to cattle and has the potential to serve as an alternative electron acceptor within the rumen (Anderson et al. 2003). More recently, the effect of 3-nitrooxypropanol (3NP) and ethyl-3-nitrooxypropanol on rumen fermentation and methane emission has been studied using Rusitec fermenters and in in vivo trials (Haisan et al. 2013; Martinez-Fernandez et al. 2013). The 3NP compound decreased CH4 production per unit of DMI in sheep in respiration chambers (24 % reduction; Martinez-Fernandez et al. 2013) and dairy cows using the SF6 technique (a dramatic 60 % decrease; Haisan et al. 2013).
5.2.5 Mitigation Through Forage Feeding
Lucerne hay decreases CH4 emissions by 21 % (Benchaar et al. 2001) when expressed as % of digestible energy. McCaughey et al. (1999) observed that beef cattle reared on grazing decrease 10 % CH4 production by unit of product when grasses were replaced by a mixture of lucerne and grasses (70: 30). The authors concluded that this was due to the higher intake observed for lucerne-fed animals, which was related with a higher digestibility rate and an increased passage of feed particles out of the rumen. This effect on methanogenesis is not a characteristic of all legumes; for instance, clover (white and/or red) did not differ from ryegrass on CH4 emissions of growing cattle (Beever et al. 1989) or dairy cows (Van Dorland et al. 2007). Several authors have shown that including tannin-rich legumes (sainfoin, lotus, sulla) and shrubs in the diet contributes to a decrease in methanogenesis due to the presence of condensed tannins (Waghorn et al. 2002). Forage preservation and processing also affect enteric CH4 production, but limited information with regard to these effects is available in the literature. Methanogenesis tends to be lower when forages are ensiled than when they are dried and when they are finely ground or pelleted than when coarsely chopped (Beauchemin et al. 2008).
5.2.6 Dietary Lipids
Vegetable and animal lipids are also considered useful in terms of reduced rumen methanogenesis (Beauchemin et al. 2007; Brask et al. 2013). Ten to 25 % reduction of methane may be achievable through the addition of dietary oils to the diets of ruminants (Beauchemin et al. 2008). Possible mechanisms by which added lipid can reduce methane production include: (a) by reducing fibre digestion (mainly in long-chain fatty acids), (b) by lowering dry matter intake (if total dietary fat exceeds 6–7 %), (c) through direct inhibition of activities of different microbes including methanogens, (d) through suppression of rumen protozoa and (e) to a limited extent, through biohydrogenation of unsaturated fatty acids (Beauchemin et al. 2008; Eckard et al. 2010). The addition of different oils (soya, coconut, canola, linseed, rapeseed, sunflower, etc.) to ruminant diets has been shown to reduce methane production between 18 % and 62 % in Rusitec fermenters (Dohme et al. 2000), sheep (Ding et al. 2012), beef cattle (O’Mara 2004; McGinn et al. 2004; Beauchemin and McGinn 2006) and dairy cows (Hristov et al. 2009; Sejian et al. 2011b; Brask et al. 2013). Copra meal gives comparable decreases in CH4 to refined coconut oil (Jordan et al. 2006).
Comparison of the effects of different fatty acids revealed that lauric, myristic and linoleic acids were the most potent reducers of methanogenesis (Jordan et al. 2006; Ding et al. 2012), and the ability of lauric acid to decrease cell viability of Methanobrevibacter ruminantium has been recently reported by Zhou et al. (2013). A wide range of essential oils (derived from garlic, thyme, oregano, cinnamon, rhubarb, frangula, etc.) has been shown to decrease methane production in vitro in a dose-dependent manner, but at high doses, the decrease in methanogenesis was accompanied by adverse effects on fermentation such as reduction in VFA production and feed digestibility (Busquet et al. 2005; Patra and Yu 2012). The lack of response in vivo is partly attributed to the adaptation of microbes (Bodas et al. 2012) but also to the use of lower doses compared to those in the in vitro experiments. Ohene-Adjei et al. (2008) also do not exclude the possibility of influence of essential oils in the activity of methane-producing genes.
5.2.7 Concentrate Supplementation
Increasing the level of concentrate in the diet leads to a reduction in CH4 emissions as a proportion of energy intake or expressed by unit of animal product (milk and meat). Methane losses appear relatively constant for diets containing up to 30–40 % concentrate (6–7 % of GE intake) and then decrease rapidly to low values (2–3 % of GE intake) for diets containing 80–90 % concentrate (Lovett et al. 2005; Martin et al. 2007). Replacing structural carbohydrates from forages (cellulose, hemicellulose) in the diet with nonstructural carbohydrates (starch and sugars) contained in most energy-rich concentrates is associated with increases in feed intake, higher rates of ruminal fermentation and accelerated feed turnover, which results in large modifications of rumen physio-chemical conditions and microbial populations. This results in a lower CH4 production because the relative proportion of ruminal hydrogen sources declines whereas that of hydrogen sinks increases. Concerning the effect of the nature of concentrate on methanogenesis, few direct comparisons have been carried out. Concentrates rich in starch (wheat, barley, maize) have a more negative effect on CH4 production than fibrous concentrates (beet pulp).
5.2.8 Propionate Enhancers
Within the rumen, hydrogen produced by the fermentation process may react to produce either methane or propionate. By increasing the presence of propionate precursors (e.g. pyruvate, oxaloacetate, malate, fumarate, citrate, succinate, etc.), more of the hydrogen is used to produce propionate and methane production is reduced (O’Mara 2004). Moreover, the stoichiometric balance of VFA, CO2 and CH4 indicates that acetate and butyrate promote CH4 production whereas propionate formation conserves H2. Therefore, when CH4 production is decreased, propionate production is increased. Propionate precursors can be introduced as a feed additive for livestock receiving concentrates. The propionate precursor malate also occurs naturally in grasses, and research is being conducted to identify affordable natural sources, e.g. alfalfa and engineered feedstocks with high concentrations of propionate precursors. As propionate precursors naturally occur in the rumen, they are likely to be more readily acceptable than antibiotic or chemical additives.
5.3 Manipulation of Rumen Microbial Ecosystem
5.3.1 Direct-Fed Microbials
Direct-fed microbials (DFM) used in ruminant nutrition are yeast-based products (YP). The notion of using YP to mitigate CH4 production has been discussed (Newbold and Rode 2006), but with the exception of some exciting and unconfirmed in vitro results (Chaucheyras et al. 1995), convincing animal data to support this concept are lacking. There have also been other attempts to inoculate the rumen with fungi (Candida kefyr) and lactic acid bacteria (Lactococcus lactis) along with nitrate supplementation to both control methanogenesis and possibly prevent nitrite formation, but no consistent animal data have been reported (Takahashi 2011).
5.3.1.1 Yeast Culture
Yeast cultures reduce methane production in three ways: (1) by reducing protozoa numbers, (2) by increasing butyrate or propionate production and (3) by stimulating acetogens to compete with methanogens or to co-metabolise hydrogen, thereby decreasing methane formation (Mwenya et al. 2004; Chaucheyras et al. 2008). However, only limited information is available on the effects of yeasts (i.e. Saccharomyces cerevisiae and Aspergillus oryzae) on methane production, and most of the studies were conducted in vitro. Carro et al. (1992) reported that supplementing Rusitec fermenters with S. cerevisiae reduced methane production and protozoa numbers with a 50:50 forage-concentrate diet, but no effects were observed with a 70:30 forage-concentrate diet, which would indicate that effects of probiotics may be diet dependent.
5.3.1.2 Methane Oxidisers
Methane-oxidising bacteria (methanotrophs) could also be introduced as direct-fed microbial preparations. The oxidation reaction would compete with the production of methane, which is a strictly anaerobic process. Methanotrophs are a unique group of methylotrophic bacteria, which utilise methane as their sole carbon and energy source (Hanson and Hanson 1996). Methane oxidisers from gut and non-gut sources could be screened for their activity in rumen.
5.3.2 Use of Bacteriocins
Bacteriocins are antimicrobial proteinaceous polymeric substances that are ubiquitous in nature and produced by a variety of Gram-negative and Gram-positive bacteria. They are typically narrow-spectrum antibacterial substances under the control of plasmid and play a role in competition among microbial species for niches within the rumen system. McAllister and Newbold (2008) reported that bacteriocins could prove effective in directly inhibiting methanogens and redirecting H2 to other reductive bacteria, such as propionate producers or acetogens. The most well-known bacteriocin is nisin. Nisin obtained from Lactobacillus lactis ssp. lactis has also been shown to decrease methane production in vitro. It is nearly as potent a methane inhibitor as monensin, and it was just as effective in decreasing the acetate to propionate ratio and has been reported that 36 % methanogenesis was reduced by the use of nisin (Callaway et al. 1997). A combination of nisin and nitrate, an alternative electron receptor, has been reported to reduce methane emissions in sheep (Sar et al. 2005). Bovicin HC5, the semi-purified bacteriocin produced by Streptococcus bovis HC5 from the rumen, has been reported to suppress methane production by 50 % in vitro (Lee et al. 2002; Sang et al. 2002), and even low concentration of bovicin HC5 (128 activity units ml−1) may be equally as useful as monensin in limiting methane production in the rumen (Lee et al. 2002). Mantovani and Russell (2002) suggested that bovicin HC5 inhibited a variety of Gram-positive bacteria and the spectrum of activity was similar to monensin. Recently, highly specific antibacterial activity of PRA-1 produced by Lactobacillus plantarum TUA1490L against methanogens was reported by Asa et al. (2010). The methane content was observed to decline with pediocin, enterocin and combinations of both after 24 h incubation. Some bacteriocins produced by lactic acid bacteria have been identified as an alternative group of antimicrobials for manipulation of the rumen microbial ecosystem and characterised biochemically and genetically (Chen and Hoocver 2003).
5.3.3 Fungal Metabolites
Secondary fungal metabolites from Monascus spp. reduced enteric methane emissions in sheep by 30 % in a short-term trial. Reduction of methane was accompanied, both in vitro and in vivo, by a shift in VFA pathways, decreasing the acetate to propionate ratio. The main microbial modifications observed were reduction in methanogen numbers, suggesting a specific and toxic effect on this microbial group. Methane emissions and the acetate to propionate ratio remained numerically less in the 2 weeks posttreatment as compared with measures before treatment (Morgavi et al. 2013).
5.3.4 Methane-Reducing Species
Mitsuokella jalaludinii has been demonstrated as an efficient methane-reducing agent in the rumen by competing with methanogens for hydrogen, necessary for growth by both (Mamuad et al. 2012). Moreover, Mitsuokella jalaludinii may not only decrease gas production in livestock but also improve ruminal fermentation and, in turn, improve feed efficiency. This may suggest that the favourable relationship between better feed efficiency and lower methane production may be due to an increased ability to reduce methane in the rumen as opposed to lower abundance of actual methanogenic species.
5.4 Phage Therapy
The lytic potential of phages and their genes make them an important tool for methane mitigation strategies. In contrast to nearly 300 phage genomes, only six archaeal phages are sequenced and described, and just four of them are from methanogens: Methanobacterium phage psi M1, M2 and M100 (Pfister et al. 1998) and also Methanothermobacter phage psi M100 (Luo et al. 2001). Little information is currently available on the genetic blueprint and gene functionality of archaeal, particularly methanogenic, phages, but more are being discovered using electron microscopy and in vitro techniques. McAllister and Newbold (2008) reported siphophages that can infect methanogens (Methanobacter, Methanobrevibacter and Methanococcus spp.), although these phages have not been isolated from rumen. Metagenomic surveys are expected to reveal the presence of embedded prophages and phage-like elements that would have otherwise remain unnoticed. An unanticipated outcome from sequencing the M. ruminantium genome was the discovery of prophage φ-mru having 69 phage-related proteins (Leahy et al. 2010). A gene encoding a putative lytic enzyme was identified, expressed and shown to lyse M. ruminantium. Such lytic enzyme is a potentially useful bio-controlling agent for manipulating rumen methanogenic populations (Leahy et al. 2010). Phages are host- and even strain-specific, so phage-based methane mitigation strategies could be developed without affecting other phylogenetically distinct microbes in the rumen. However, hosts and phages are also known to be involved in a rapid evolutionary race as the host changes to avoid infection and the phage changes to maintain infectivity. In combination with the application of other phage enzymes and structural components, a rotation system can be envisioned that may overcome the rapid adaptation mechanisms of microbes to phage challenges. More methanogenic phages need to be identified, sequenced and characterised to identify and employ such phage-based methane mitigation strategies.
5.5 Secondary Plant Metabolites
This category includes a variety of plant secondary metabolites (PSM), specifically tannins and saponins, which have been extensively studied for their CH4 mitigating potential (Staerfl et al. 2012; Bhatta et al. 2012, 2013a). As a separate chapter addresses the role of PSM in this book, only the salient features of PSM are being discussed in this chapter.
5.5.1 Tannins
Tannins are plant polyphenols of varying molecular size and exist in two forms in plants: hydrolysable and condensed tannins (CT). Tannins as feed supplements or as tanniferous plants have often, but not always (Beauchemin et al. 2007), shown potential for reducing CH4 emission by up to 20 % (Zhou et al. 2011; Staerfl et al. 2012). A meta-analysis of in vivo experiments with tannins by Jayanegara et al. (2012) reported a relatively close relationship between dietary tannin concentration and CH4 production per unit of digestible organic matter. According to Goel and Makkar (2012), the anti-methanogenic effect of tannins depends on the dietary concentration and is positively related to the number of hydroxyl groups in their structure. These authors concluded that hydrolysable tannins tend to act by directly inhibiting rumen methanogens, whereas the effect of condensed tannins on CH4 production is more through inhibition of fibre digestion. The mechanism to decrease methanogenesis seems to vary with the nature of CT, as Bhatta et al. (2013b) observed that Ficus benghalensis and Artocarpus integrifolia reduced methane production due to defaunation, but Azadirachta indica reduced methanogenesis by a direct effect on methanogens, whereas feeding up to 2 % of the dietary DM as quebracho tannin extract failed to reduce enteric methane emissions from growing cattle (Beauchemin et al. 2007).
5.5.2 Saponins
Saponins are naturally occurring surface-active glycosides with foaming characteristics, occurring in many plant species. They usually consist of a sugar moiety linked to a hydrophobic compound, either triterpenoid or steroid in nature (Francis et al. 2002). Saponins reduce methane production via inhibition of either protozoa or methanogens or both. These inhibited protozoa at relatively low concentrations, whereas higher concentrations were required to kill or suppress methanogenic archaea (Wina et al. 2005; Goel et al. 2008). Saponins are considered to have detrimental effects on protozoa through their binding with cholesterol present on the protozoal cell membrane leading to its disruption, breakdown, lysis and finally cell death (Francis et al. 2002). Because of their anti-protozoal activity, saponins might have the potential to reduce CH4 as protozoa have both an ecto- and endosymbiotic relationship with methanogens, and methanogens associated with protozoa are estimated to be responsible for 9–37 % of the total CH4 production in the rumen (McAllister and Newbold 2008). Anti-methanogenic activity of saponins is believed to occur by limiting hydrogen availability to methanogens and rechannelling of metabolic hydrogen from methane to propionate production in the rumen (Wina et al. 2005). In addition, saponins, due to their chemical structure, may display antibacterial properties by reducing the number of bacteria producing H2 thus resulting in the inhibition of H2 production—a substrate for methane formation (Wang et al. 2009).
5.6 Immunisation for Reducing Enteric Methane Mitigation
Host immunisation offers a diverse and eco-friendly solution to the problems associated with animal health. Therefore, developing vaccines against methanogens appears to be an alternative and attractive approach for extensive production systems. Vaccines against rumen archaea are based on the concept of a continuous supply of antibodies to the rumen through saliva. The vaccine also works by triggering an animal’s immune system to produce antibodies against methanogenic bacteria that live in the rumen of the animal. Wright et al. (2004) developed two vaccines, VF3 (based on three methanogenic strains 1Y, AK-87 and ZA-10) and VF7 (based on seven methanogens), that produced a 7.7 % reduction in methane emissions from sheep, despite targeting only a minority (20 %) of methanogens present within these host animals. Moreover, in Europe up to 23 % ruminal methane emission reduction was reported (Wedlock et al. 2010). They also created a vaccine based on five methanogens (Methanobrevibacter spp. strains 1Y, AK-87, M. millerae ZA-10, Methanomicrobium mobile BP and M. stadtmanae MCB-3) that was administered in three vaccinations to sheep (Williams et al. 2009). Surprisingly, immunisation with this second vaccine caused methane output to increase by 18 %, despite the fact that a larger proportion of the methanogenic population (52 %) was targeted. Vaccines against archaea have been successful in vitro (Wedlock et al. 2010) but not in vivo (Wright et al. 2004; Williams et al. 2009). Vaccines prepared from New Zealand and Australian methanogen strains proved unsuccessful in reducing CH4 production in ewe lambs (Clark et al. 2004). Thus, further work is needed to optimise the individual components of these vaccines such that the most potent methanogens are specifically targeted. Researchers believe that anti-methanogenic vaccines will only yield the short-term reductions in methanogens and/or methanogenesis, due to the possible proteolytic degradation and low persistence of host antibodies in rumen (Cook et al. 2008; Lascano and Cárdenas 2010). Vaccination of sheep with entodinial or mixed protozoa antigens reduced protozoa, and the released IgG antibodies against rumen protozoa remained active and continued to bind the target cells up to 8 h (Williams et al. 2008). Vaccines targeting single surface antigens may not be effective, as methanogenic archaea differ largely based on their host, diet as well as geographical regions. A new vaccine has been developed using subcellular fractions (cytoplasmic and cell wall-derived protein) of M. ruminantium M1 (Wedlock et al. 2010). Twenty sheep were vaccinated, then booster doses were given after 3 weeks and the antisera were found to agglutinate and decrease the growth of archaeal methanogens and methane production in vitro. The in vivo efficacy of the vaccine on methanogens is yet to be evaluated. New approaches have involved identification of genes encoding specific membrane-located proteins from M. ruminantium and using purified proteins (produced in Escherichia coli) as antigens to vaccinate sheep (Buddle et al. 2011).
In another approach, antisera were generated in sheep against subcellular fractions from M. ruminantium, which reduced microbial growth and CH4 production in vitro (Wedlock et al. 2010). Sequencing the genome of M. ruminantium has opened new opportunities for inhibition of rumen methanogens and the potential to mitigate ruminant CH4 emissions (Leahy et al. 2010). Based on liquid chromatography mass spectrometer, it was reported that most of the proteins were intracellular enzymes, particularly methyl-coenzyme M reductase, and these intracellular proteins would not be suitable as vaccine antigens owing to their inaccessibility for antibody binding. Since there is growing database for the genome sequences of rumen methanogens, the possibility of finding new target antigens/proteins using comparative and pangenomics analysis has increased. Furthermore, extensive research is needed to identify adjuvants that stimulate high titre of antibody and are suitable for formulating with protein antigens to produce a low-cost and effective vaccine. However, the vaccine-based inhibition method will have to pass the regulatory systems to guarantee animal health.
A team of researchers at Commonwealth Scientific and Industrial Research Organisation (CSIRO) in Australia have made an application for a worldwide patent for a method of improving the productivity of a ruminant animal by administering to the animal an immunogenic preparation effective to invoke an immune response to at least one rumen protozoan. The removal of one species of protozoan from the rumen will invoke the improvements in productivity associated with defaunation. It is also believed that by modifying the activity of the rumen protozoan, there will be an indirect effect on the activity of methanogens, due to their commensal relationship with rumen protozoa. Data from this work are not yet published, but it is anticipated that methane production could be reduced by as much as 70 %. The anti-methanogen vaccine is under development by CSIRO in Australia. A number of experimental vaccine preparations were given to sheep. The studies have shown that up to a 20 % reduction in methane production is possible, but the observed reductions are dependent on diet, climate and time of year. To date, sheep have been used as the experimental animal because of the ease of use and availability. Methane production was reduced by 11–23 % in vaccinated animals. CSIRO’s next task is to develop a vaccine and test its effectiveness in cattle.
5.7 Modelling of GHG in Livestock Farm
Livestock undoubtedly need to be a priority focus of attention as the global community seeks to address the challenge of CC. The magnitude of the discrepancy between estimates illustrates the need to provide the CC community and policymakers with accurate emission estimates and information about the link between agriculture and climate. Improving the global estimates of GHG attributed to livestock systems is of paramount importance. This is not only because we need to define the magnitude of the impact of livestock on CC but also because we need to understand their contribution relative to other sources. Estimates of GHG emission through experiments under different production system is practically impossible and with growing awareness of global warming and its continuous negative impact on agricultural production system demands immediate mitigation strategies to curtail such emissions. In this context, simulation models offer a great scope to predict accurately the GHG emission in farm as a whole (Sejian et al. 2011c). Such information will enable effective mitigation options to be designed to reduce emissions and improve the sustainability of the livestock sector while continuing to provide livelihoods and food for a wide range of people. A synthesis of the available literature suggests that the mechanistic models are superior to empirical models in accurately predicting the CH4 emission from dairy farms (Sejian and Naqvi 2012). The latest development in prediction model is the integrated farm system model which is a process-based whole-farm simulation technique (Chianese et al. 2009; Rotz et al. 2009).
6 Amelioration Strategies to Improve Livestock Production Under Changing Climate Scenario
Livestock producers have traditionally adapted to various environmental and climatic changes by building on their in-depth knowledge of the environment in which they live. However, the expanding human population, urbanisation, environmental degradation and increased consumption of animal source foods have rendered some of those coping mechanisms ineffective (Sidahmed 2008). Since CC could result in an increase of heat stress, all methods to help animals cope with or, at least, alleviate the impacts of heat stress could be useful to mitigate the impacts of global change on animal responses and performance (Sejian et al. 2012b).
6.1 Physical Protection
Physical protection with artificial or natural shade presently offers the most immediate and cost-effective approach for enhancing the productive and reproductive efficiency of animals (Sejian 2013). Evaporative cooling also can be effective. Various shade management systems have been evaluated extensively and generally result in improved feed intake and productivity. Sprinkling animal in the morning is more effective than sprinkling in the afternoon. Certainly, it is recommended to start cooling strategies prior to animal showing signs of heat stress (panting). Sprinkling of pen surfaces may be as much or more beneficial than sprinkling the animal. Cooling the surface would appear to provide a heat sink for animal to dissipate body heat, thus allowing animal to better adapt to environmental conditions than adapting to being wetted. In handling studies, moving animal through working facilities requires an expenditure of energy causing an elevation of average body temperature between 0.5 and 1.0 °C (0.9 and 1.8 °F), depending on the ambient conditions. So during hot days, minimal handling of animal is recommended for promoting animal comfort. Bedded barn facilities appear to be useful for buffering animal against the adverse effects of the environment under hot and cold conditions even though 2–4 °C higher temperature as well as THI level is maintained within the barn when compared to outside conditions, possibly by the decreased airflow through the building. The use of bedded barns does not reduce heat stress, as measured by the THI, but act as a shade to decrease the solar heat load on the animal.
6.2 Ideal Shed Requirements
Careful management which can alleviate heat stress is the best way to maintain high production levels in lactating cows in a hot environment. Cattle will produce milk and reproduce more efficiently if they are protected from extreme heat and particularly from direct sunshine. Good management includes the modification of the surrounding environment to reduce the impact of the environment and/or to promote heat loss from the animals (Shibata 1996). Thus, in tropical and subtropical climates, providing shade becomes an important factor. In many cases, the provision of shade may be the most economical solution of reducing high heat load. It is suggested that a well-designed shade structure should reduce the total heat load by 30–50 %. Although the benefits of providing shade to cattle will vary depending on factors such as breed, coat colour, weight, health and lactation status, producers may be able to increase production and improve pasture use and water quality by providing it. The amount of shade needed depends on the type and age of the cattle. Shade is a must for pasture-based grazing systems. It curtails heat stress, which is detrimental to cattle. Besides, if cattle are kept in a confined area, it should be free from mud and manure in order to reduce hoof infection to a minimum. Concrete floors or pavements are ideal where the area per cow is limited. However, where ample space is available, an earth yard, properly sloped for good drainage, is adequate. The size of this paved area depends on the orientation of the shade structure. If the longitudinal axis is east to west, then part of the floor under the roof should be in shade all day. Extending the floor by approximately one-third of its length on the east and on the west results in a paved surface being provided for the shaded area at all times. If the longitudinal axis is north to south, the paved area must be three times the roof area, i.e. one-third to the east, one-third to the west and one-third underneath. In regions where temperatures average 30 °C or more for up to 5 h per day during some period of the year, the east-west orientation is the most beneficial.
Shade cloth is typically used as the roof covering to allow air movement. Shade cloth patterns come in various weaves providing 30–90 % shade. One of the more common types is a woven polypropylene fabric which provides 80 % shade. While longevity is considerably less than that expected of permanent structures, shade cloth if properly maintained (kept tight) can last 5 years or longer. Shade cloth is commonly available in black, though lighter colours reflect more heat. The gable roof is more wind resistant than a single pitch roof and allows for a centre vent. A woven mat of local materials can be installed between the rafters and the corrugated iron roof to reduce radiation from the steel and to reduce temperatures just below the roof by 10 °C or more.
Cattle generally prefer shade from trees rather than constructed structures. Trees are effective at blocking incoming solar radiation, and moisture evaporating from their leaves helps cool surrounding air (Hahn 1981). Planting shade trees on the west side of pastures will provide protection from the afternoon sun. Feed and water can be located close to the existing or planned natural shade. Permanent shade can be provided by constructing barns or sheds. It is most often provided for dry lots and bull lots. Often in a grazing system, permanent shade is not located where it is needed, and it can be costly. The size of shade structures varies according to climatic conditions. Researchers worldwide generally recommend 19–27 ft2 of floor space per cow. However, for environments that are particularly hot and humid, floor space equivalent to 60–65 ft2 per cow is recommended. Space requirements are essentially doubled in hot and humid climates to provide additional open area for improved air movement.
Air movement increases the rate of heat loss from a cow’s body surface, as long as the air temperature is lower than the animal’s skin temperature. Fans should be tilted downward at a 20–30° angle (from vertical) to direct the flow of air onto the cows, and preferably they should be kept above the sprinklers to remove the moisture. Another cooling device that might be useful is using mist and fan system. In this cooling system, mist particles are sprayed onto the animal’s body to wet the hair. A fan is then used to evaporate the moisture, as a way of cooling the cows. The single use of a sprinkling and fan system, for 30 min before milkings, has proved to be useful to relieve dairy cows’ heat stress, in terms of efficiency to reduce the impact of heatwaves under a grazing system (Valtorta et al. 2002).
6.3 Nutritional Interventions
There is a growing interest in strategically altering the diet in an attempt to improve the production of livestock in changing climatic stress (Sejian 2013; Shinde and Sejian 2013). A common strategy is to increase the energy and nutrient density (reduced fibre, increased concentrates and supplemental fat) of the diet as feed intake is markedly decreased during heat stress. In addition to the energy balance concern, reducing the fibre content of the diet is thought to improve the cow’s thermal balance and may reduce body temperature (FAO 2012). Because there is greater heat production associated with metabolism of acetate compared with propionate, there is a logical rationale for the practice of feeding low fibre rations during hot weather. Feeding more concentrate at the expense of fibrous ingredients increases ration energy density and reduces heat increment (Sejian et al. 2012b). Due to reduced feed intake, dietary protein levels may need to be increased during heat stress (West 1999). However, recent recommendations suggest that addition of dietary crude protein, more specifically rumen undegradable protein, is not helpful (Arieli et al. 2006). Increasing the amount of dietary fat has been a widely accepted strategy within the industry in order to reduce basal metabolic heat production. The heat increment of fat is over 50 % less than typical forages, so it is seemingly a rational decision to supplement additional lipid and reduce fibre content of the diet.
Water intake is vital for milk production (milk is ~87 % water) and also essential for thermal homeostasis. This stresses the importance of water availability. Keeping water tanks clear of feed debris and algae is a simple and cheap strategy to help cows remain cool and improve production (Baumgard and Rhoads 2007). Further, dietary supplementation with either lipoic or dihydrolipoic acid may improve heat tolerance and animal performance during heat stress by enhancing insulin action. However, the effectiveness of lipoic acid supplementation to alter glucose availability may be dependent on the magnitude and duration of heat stress events (Hamano 2012).
6.3.1 Mineral Supplementation
Unlike humans, bovines utilise potassium (K+) as their primary osmotic regulator of water secretion from sweat glands. As a consequence, K+ requirements are increased (1.4–1.6 % of DM) during the heat stress, and this should be adjusted for in the diet. In addition, dietary levels of sodium (Na+) and magnesium (Mg+) should be increased as they compete with K+ for intestinal absorption (West 2002). Among micronutrients, zinc is one of the most important in the body (Stefanidou et al. 2006). At the cellular level, zinc is essential for cell proliferation and survival and contributes to genomic stability and antioxidant defence. Zinc is a potent inducer of Hsp70 in cell culture (Hatayama et al. 1993), which is indispensable for adaptation (Voellmy 2004). Besides, chromium has proved its importance during thermal stress. Chromium facilitates insulin action on glucose, lipid and protein metabolism. Because glucose use predominates during heat stress, chromium supplementation may improve thermal tolerance or production in heat-stressed animals (Spears et al. 2012).
6.3.2 Antioxidant Supplementation
Nutritional tools such as antioxidant feeding (vit. A, selenium, zinc, etc.) and ruminant-specific live yeast can help. Studies have shown that addition of antioxidant in diets of sheep is able to reduce heat stress and is a good strategy to prevent mastitis, optimise feed intake and reduce the negative impact of heat stress on milk production. Moreover, the use of antioxidant such as vit. E, vit. A, selenium and selenium-enriched yeast helps in reducing the impact of heat stress on the oxidant balance, resulting in improved reproductive efficiency (Sejian et al. 2014). Both vitamin C and vitamin E protect the biological membranes against the damage of reactive oxygen species (ROS). The role of vitamin E as an inhibitor—“chain blocker”—of lipid peroxidation has been well established (Seyrek et al. 2004). Vitamin C along with electrolyte supplementation has also been reported to ameliorate the heat stress in buffaloes (Sunil Kumar et al. 2010).
6.4 Disease Surveillance Measures
Adaptation assessments are designed to identify options to reduce the current and projected health risks attributable to CC by preventing exposures to weather and climate hazards, reducing the consequences of exposure and/or reducing vulnerabilities. Assessing the potential infectious disease risks of CC also requires considering the non-climatic factors that drive their incidence and distribution, including demographics, socio-economic development, land use, urbanisation, technology and the political and health-care context (Weiss and McMichael 2004; Suk and Semenza 2011). There is an urgent need for public health and health care to develop adaptation strategies for the impacts of CC on infectious diseases (Zhang et al. 2008; Paaijmans et al. 2010).
One of the most critical obstacles to improving our understanding of climate-disease linkages is the lack of high-quality epidemiological data on disease incidence for many locations. Greater attention should be focused on areas where climate-sensitive diseases are often found or where outbreaks that appear to be weather related are more severe and more difficult to predict. Increased understanding of disease-climate linkages should permit public health workers to begin to shift focus from “surveillance and response” to a more proactive approach of “prediction and prevention”. Adaptive capacity can be improved through better surveillance if the resulting data are made available in a timely fashion and appropriate manner.
Age- and cause-specific mortality surveillance offers a baseline for recognising what is considered “normal” and allows recognition of unusual mortality periods or locations. Such death data have served as the basis for analysis of extreme weather events that occur over days to weeks. Floods, heatwaves and “cold snaps” are examples. Historical surveillance data permit evaluation of how these extreme weather periods may have resulted in deaths above what would have been expected for that time and place. Modern computing power is also being used to improve surveillance through geographical information systems (GIS) and spatial statistical analyses. More and more countries are using simple, low-cost GIS for management and mapping of their data. The health map programme of the World Health Organization (WHO) helps countries use GIS systems with standardised indicators to enhance comparability. These computer programs allow data entry and analysis at the local level and can be transmitted electronically to higher levels without loss of detail. When available, such data may be relevant to understanding the pattern of various diseases, depending on their transmission pattern and other relationships. Other kinds of surveillance data, derived from satellite images (remote sensing data), are often included in GIS and spatial statistical analyses that contribute to disease surveillance and pattern analysis. Another important contemporary surveillance method involves use of acute-care networks that employ innovative software for detecting known risks and unexpected events. These measures will ensure timely intervention of sudden disease outbreaks of livestock that might occur as a result of CC.
6.5 Advanced Reproductive Technologies
Infertility in the male caused by heat stress can be eliminated through the utilisation of artificial insemination (AI) with semen collected and frozen from males in cool environments. In females, the situation is more complicated; embryo transfer (ET) represents a method analogous to AI in that embryos can be collected from nonstressed cows and transferred to heat-stressed recipients (Rutledge 2001). ET can be an effective fertility-enhancing strategy for the heat-stressed cow because most of the effects of heat stress to reduce fertility occur before the blastocyst stage of development when embryos are typically transferred. It may also be possible to remove damaged follicles from the ovary by transvaginal aspiration to hasten the recovery period after heat stress. Estrus induction techniques offer the opportunity to induce fertile estrus in noncyclic animals to increase fertility. Different treatments have been utilised to induce estrus, such as prostaglandin, gonadorelin and progestagen. Better results have been obtained using progesterone-impregnated intravaginal device (PRID) plus pregnant mare serum gonadotropin (PMSG) and prostaglandin. Stem cell research in livestock is still in its infancy, and a number of technical hurdles need to be overcome before the full potential of research and therapeutic use can be realised. Induced differentiation of stem cells towards male and female gametes can be exploited for combating infertility problems. The embryo production and transfer technique is immensely useful in the production of embryonic stem cells and transgenic animals. It shall be highly useful to improve efficiency of this technology and study basic mechanisms involved in it using modern molecular and proteomics approaches.
Administrations of hormonal preparations have found to be more beneficial for improving reproductive efficiency in livestock. Pregnancy rate can also be improved in the summer and autumn in primiparous cows by the use of gonadotropin-releasing hormone and prostaglandin F2α to generate three consecutive 9-day follicular waves beginning at 50–60 days in milk (Friedman et al. 2011). The gonadotropin-releasing hormone (GnRH) administered at estrus onset is found more effective in primiparous cows and during the summer in increasing conception rate in cows with low body condition scores at AI (Kaim et al. 2003). In particular, administration of either follicle-stimulating hormone (FSH) or somatotropin during one estrous cycle in the autumn improves oocyte quality in the subsequent cycle (Roth et al. 2002). Among its many actions, insulin-like growth factor-1 (IGF-1) can protect cells from various forms of stress including elevated temperature. In particular, administration of IGF-1 to bovine embryos produced in vitro reduced the magnitude of effects of elevated temperature on inhibition of development and apoptosis (Jousan and Hansen 2004, 2007). Probably the easiest way to increase IGF-1 concentrations in lactating cows is to administer bovine somatotropin (bST). Sakatani et al. (2004) indicated that treatment of cows with bST can increase pregnancy rates of cows bred via timed AI, increase the survivability of embryos recovered from donors and increase the pregnancy rate of recipients treated with bST. ET offers the opportunity to expose embryos directly to IGF-1. Transfer of in vitro-produced embryos that were cultured with IGF-1 into heat-stressed, lactating recipients resulted in increased pregnancy and calving rates as compared to transfer of in vitro-produced embryos cultured without IGF-1 (Block and Hansen 2007).
6.6 Thermal Tolerance Genes in Livestock
Given the complexity of the traits related to adaptation to tropical environments, the discovery of genes controlling these traits is a very difficult task. One obvious approach of identifying genes associated with acclimation to thermal stress is to utilise gene expression microarrays in models of thermal acclimation to identify changes in gene expression during acute and chronic thermal stress. Another approach will be with single gene deletions exposed to a defined thermal environment. This permits the identification of those genes that are involved in key regulatory pathways for thermal resistance and thermal sensitivity. Finally, gene knockout models in single cells will also allow better delineation of the cellular metabolic machinery required to acclimate to thermal stress. Those genes identified as key to the process of thermal acclimation will then need to be mapped to their chromosomal location, and the sequences of these genes will need to be determined in order to see if there are single-nucleotide polymorphisms (SNPs) that are associated with changes in the coding for gene expression or protein function. Identification of SNPs that are associated with variation in animal resistance or sensitivity to thermal stress will permit screening of animal’s presence or absence of desirable or undesirable alleles. However, further research is needed to quantify the genetic antagonism between adaptation and production traits to evaluate the potential selection response.
Studies evaluating genes identified as participating in the cellular acclimation response from microarray analyses or genome-wide association studies have indicated that heat-shock proteins are playing a major role in adaptation to thermal stress. In mammalian cells, nonlethal heat shock produces increased thermotolerance through enhanced expression of heat-shock genes. Additional genes of interest which two or more studies have identified are the genes for fibroblast growth factor, solute carrier proteins, interleukins and tick-resistance genes. Genes which have only been identified by microarray analysis but not by genome-wide association studies include genes associated with cellular metabolism (phosphofructokinase, isocitrate dehydrogenase, NADH dehydrogenase, glycosyltransferase, transcription factor and mitochondrial inositol protein). Other genes of importance were thyroid hormone receptor, insulin-like growth factor II and annexin. Genes repressed in response to the environmental stress are mostly concerned with translation of genes for cytoplasmic ribosomal protein, DNA polymerase I, II and III, transcription, t-RNA synthetases, proteins required for processing ribosomal RNA and a subset of translation initiation factors. The identification of the variety of CER genes involved in stress responses suggests that these responses are aimed at production of additional energy (ATP), maintenance of environment as well as the repression of protein synthesis to ensure energy conservation and minimise unnecessary burden on the part of the cell. Marker-assisted selection (MAS) programme may be clubbed with appropriate breeding programme involving these thermotolerant genes to produce animals which have superior adaptive capability to environmental extremes.
7 Conclusion
Climate change is seen as a major threat to the survival of many species, ecosystems and the sustainability of livestock production systems in many parts of the world. Livestock production is thought to be adversely affected by detrimental effects of extreme climatic conditions. Consequently, adaptation and mitigation of detrimental effects of extreme climates have played a major role in combating the climatic impact in livestock production. In fact, the animals can adapt to the hot climate; nevertheless, the response mechanisms are helpful for survival but are detrimental to performance. Hence, formulating mitigation strategies incorporating all requirements of livestock is the hour of need to optimise productivity in livestock farms. This chapter also elaborates on different adaptive strategies that need to be given due consideration to prevent huge economic losses incurred due to climate change impact on livestock productivity. Further, this chapter details the issues of less-than-perfect information on climate impacts and vulnerabilities and need for better informed decisions on “resilient adaptation” by merging adaptation, mitigation and amelioration strategies. It offers new perspectives for policymakers, institutions, societies and individuals on improved ways of identifying most at-risk communities and “best practices” of coping with current climate variability and extreme climate events. With the development of molecular biotechnologies, new opportunities are available to characterise gene expression and identify key cellular responses to environmental stresses that arise due to climate change. These new tools will enable to improve the accuracy and the efficiency of selection for heat tolerance in livestock.
8 Future Perspectives
Adaptation to CC is an integral part of agricultural production now and will become more important in the future as the impacts of CC become more evident. In developing a strategy for adapting to CC, one key challenge is dealing with uncertainty. Significant uncertainty relates to the nature and extent of regional CC impacts, impacts across agricultural industries and impacts over time. The challenge for governments and agricultural industry stakeholders is to deal with these uncertainties through further research and the development of policies and farm management approaches that are flexible enough to deal effectively with a range of potential CC outcomes.
Responding to the challenges of global warming necessitates a paradigm shift in the practice of agriculture and in the role of livestock within the farming system. Science and technology are lacking in thematic issues, including those related to climatic adaptation, dissemination of new understandings in rangeland ecology, and a holistic understanding of pastoral resource management. The key thematic issues on environment stress and livestock production includes: early warning system, multiple stress research, simultaneous simulation models, water experiments, exploitation of genetic potential of native breeds, suitable breeding programme and nutritional intervention research. Livestock farmers should have key roles in determining what adaptation and mitigation strategies they support if these have to sustain livestock production in changing climate. The integration of new technologies into the research and technology transfer systems potentially offers many opportunities to further the development of CC adaptation strategies.
With the development of molecular biotechnologies, new opportunities are available to characterise gene expression and identify key cellular responses to environmental stresses that arise due to CC. These new tools will enable to improve the accuracy and the efficiency of selection for heat tolerance in livestock. Substantial efforts are also needed to identify specific genes associated with tolerance and sensitivity to heat stress. Continued research evaluating genomic and proteomic approaches to improve reproductive performance and nutritional status of heat stress animals is also warranted. Also, further researches are required to quantify the genetic antagonism between adaptation and production traits to evaluate the potential selection response. Epigenetic regulation of gene expression and thermal imprinting of the genome could also be an efficient method to improve thermal tolerance.
References
Abecia L, Toral PG, Martín-García AI, Martínez G, Tomkins NW, Molina-Alcaide E, Newbold CJ, Yaňez-Ruiz DR (2012) Effect of bromochloromethane on methane emission, rumen fermentation pattern, milk yield, and fatty acid profile in lactating dairy goats. J Dairy Sci 95:2027–2036
Anderson RC, Callaway TR, Van Kessel JA, Jung YS, Edrington TS, Nisbet DJ (2003) Effect of select nitrocompounds on ruminal fermentation; an initial look at their potential to reduce economic and environmental costs associated with ruminal methanogenesis. Bioresour Technol 90:59–63
Antle JM (2010) Asymmetry, partial moments, and production risk. Am J Agric Econ 92(5):1294–1309
Appuhamy JADRN, Strathe AB, Jayasundara S, Wagner-Riddle C, Dijkstra J, France J, Kebreab E (2013) Antimethanogenic effects of monensin in dairy and beef cattle: a meta-analysis. J Dairy Sci 96:5161–5173
Arieli A, Adin G, Bruckental I (2006) The effect of protein intake on performance of cows in hot environmental temperatures. J Dairy Sci 87:620–629
Asa R, Tanaka A, Uehara A, Shinzato I, Toride Y, Usui N, Hirakawa K, Takahashi J (2010) Effects of protease-resistant antimicrobial substances produced by lactic acid bacteria on rumen methanogenesis. Asian Aust J Anim Sci 23:700–707
Baumgard LH, Rhoads RP (2007) The effects of hyperthermia on nutrient partitioning. Proc Cornell Nutr Conf 93–104
Beauchemin KA, McGinn SM (2006) Methane emissions from beef cattle: effects of fumaric acid, essential oil, and canola oil. J Anim Sci 84:1489–1496
Beauchemin KA, McGinn SM, Martinez TF, McAllister TA (2007) Use of condensed tannin extract from quebracho trees to reduce methane emissions from cattle. J Anim Sci 85:1990–1996
Beauchemin KA, Kreuzer M, O’Mara F, McAllister TA (2008) Nutritional management for enteric methane abatement: a review. Aust J Exp Agric 48:21–27
Beever DE, Cammell SB, Sutto JD, Spooner MC, Haines MJ, Harland JI (1989) Effects of concentrate type on energy utilization in lactating dairy cows. In: Van der Honing Y, Close WH (eds) Energy metabolism of farm animals, EAAP Publication no. 43. Pudoc, Wageningen, pp 33–36
Benchaar C, Pomar C, Chiquette J (2001) Evaluation of dietary strategies to reduce methane production in ruminants: a modelling approach. Can J Anim Sci 81:563–574
Bergquist R, Rinaldi L (2010) Health research based on geospatial tools: a timely approach in a changing environment. J Helminthol 84:1–11
Bhatta R, Saravanan M, Baruah L, Sampath KT (2012) Nutrient content, in vitro ruminal fermentation characteristics and methane reduction potential of tropical tannin-containing leaves. J Sci Food Agric 92:2929–2935
Bhatta R, Enishi O, Yabumoto Y, Nonaka I, Takusari N, Higuchi K, Tajima K, Takenaka A, Kurihara M (2013a) Methane reduction and energy partitioning in goats fed two concentrations of tannin from Mimosa spp. J Agric Sci (Cambridge) 151:119–128
Bhatta R, Saravanan M, Baruah L, Dhali A, Kolte A, Prasad CS (2013b) Effect of graded levels of tropical leaves containing-secondary metabolites on rumen fermentation pattern protozoa population and methanogenesis in vitro. Adv Anim Biosci 4(2):307
Blaxter KL, Clapperton JL (1965) Prediction of the amount of methane produced by ruminants. Br J Nutr 19:511–522
Block J, Hansen PJ (2007) Interaction between season and culture with insulin-like growth factor-1 on survival of in vitro produced embryos following transfer to lactating dairy cows. Theriogenology 67:1518–1529
Boadi D, Wittenburg KM, Scott SL, Burton D, Buckley K, Small JA, Ominski KH (2004) Effect of low and high forage diets on enteric and manure pack GHG emissions from a feedlot. Can J Anim Sci 84:445
Bodas R, Prieto N, García-González R, Andrés S, Giráldez FJ, López S (2012) Manipulation of rumen fermentation and methane production with plant secondary metabolites. Anim Feed Sci Technol 176:78–93
Bowman RL, Croucher JC, Picard MT (1992) Assessment of the prefeasibility of strategic supplementation as an opportunity for reducing methane emissions in Gujarat, India, A.T, International, prepared for the Global change Division, U.S. Environmental protection agency, Washington, DC
Brask M, Lund P, Weisbjerg MR, Hellwing AL, Poulsen M, Larsen MK, Hvelplund T (2013) Methane production and digestion of different physical forms of rapeseed as fat supplement in dairy cows. J Dairy Sci 96:2356–2365
Brown EG, Anderson RC, Carstens GE, Gutierrez-Baňuelos H, McReynolds JL, Slay LJ, Callaway TR, Nisbet DJ (2011) Effects of oral nitroethane administration on enteric methane emissions and ruminal fermentation in cattle. Anim Feed Sci Technol 166–167:275–281
Buddle BM, Denis M, Attwood GT, Altermann E, Janssen PH, Ronimus RS, Pinares-Patiño CS, Muetzel S, Wedlock DN (2011) Strategies to reduce methane emissions from farmed ruminants grazing on pasture. Vet J 188:11–17
Busquet M, Calsamiglia S, Ferret S, Kamel C (2005) Screening for the effects of natural plant extracts and secondary plant metabolites on rumen microbial fermentation in continuous culture. Anim Feed Sci Technol 123:597–613
Callaway TR, Alexandra MS, Carneiro De Melo, Russell JB (1997) The effect of nisin and monensin on ruminal fermentations in vitro. Curr Microbiol 35:90–96
Carro MD, Ranilla MJ (2003) Effect of the addition of malate on in vitro rumen fermentation of cereal grains. Br J Nutr 89:181–188
Carro MD, Lebzien P, Rohr K (1992) Influence of yeast culture on the in vitro fermentation (Rusitec) of diets containing variable portions of concentrates. Anim Feed Sci Technol 37:209–220
Castillo C, Benedito JL, Méndez J, Pereira V, López-Alonso M, Miranda M, Hernández J (2004) Organic acids as a substitute for monensin in diets for beef cattle. Anim Feed Sci Technol 115:101–116
Chaucheyras F, Fonty G, Bertin G, Gouet P (1995) In vitro H2 utilization by a ruminal acetogenic bacterium cultivated alone or in association with an archaea methanogen is stimulated by a probiotic strain of Saccharomyces cerevisiae. Appl Environ Microbiol 61:3466–3469
Chaucheyras F, Walker ND, Bach A (2008) Effects of active dry yeasts on the rumen microbial ecosystem: past, present and future. Anim Feed Sci Technol 145:5–26
Chaucheyras-Durand F, Walker ND, Bach A (2008) Effects of active dry yeasts on the rumen microbial ecosystem: past, present and future. Anim Feed Sci Technol 145:5–26
Chen H, Hoocver DG (2003) Bacteriocins and their food applications. CRFSFS 12:82–99
Chianese DS, Rotz CA, Richard TL (2009) Whole farm greenhouse gas emissions. A review with application to a Pennsylvania dairy farm. Appl Eng Agric 25(3):431–442
Clark H, Wright AD, Joblin K, Molano G, Chavanagh A, Peters J (2004) Field testing and Australian developed antimethanogen vaccine in growing ewe lambs. In: Proceedings of the workshop on the science of atmospheric trace gases, Wellington, 18–19 Mar 2004, pp 107–108
Clemens J, Ahlgrimm HJ (2001) Greenhouse gases from animal husbandry and mitigation options. Nutr Cycl Agroecosyst 60:287–300
Cook SR, Maiti PK, Chaves AV, Benchaar C, Beauchemin KA, McAllister TA (2008) Avian (IgY) anti-methanogen antibodies for reducing ruminal methane production: in vitro assessment of their effects. Aust J Exp Agric 48:260–264
De Mazancourt C, Loreau M, Abbadie L (1998) Grazing optimization and nutrient cycling: when do herbivores enhance plant production? Ecology 79:2242–2252
Demeyer D, Fievez V (2000) Ruminants et environment: la méthanogénèse. Ann Zootech 49:95–112
Deressa T, Hassan R, Poonyth D (2005) Measuring the economic impact of climate change on South Africa’s sugarcane growing regions. Agrekon 44(4):524–542
Ding X, Long R, Zhang Q, Huang X, Guo X, Mi J (2012) Reducing methane emissions and the methanogen population in the rumen of Tibetan sheep by dietary supplementation with coconut oil. Trop Anim Health Prod 44:1541–1545
Dohme F, Machmüller A, Wasserfallen A, Kreuzer M (2000) Comparative efficiency of various fats rich in medium-chain fatty acids to suppress ruminal methanogenesis as measured with RUSITEC. Can J Anim Sci 80:473–484
Doreau M, Ferlay A (1995) Effect of dietary lipids on nitrogen metabolism in the lumen: a review. Livest Prod Sci 43:97–110
Doreau M, Jouany JP (1998) Effect of a Saccharomyces cerevisiae culture on nutrient digestion in lactating dairy cows. J Dairy Sci 81:3214–3221
Eckard RJ, Grainger C, De Klein CAM (2010) Options for the abatement of methane and nitrous oxide from ruminant production – a review. Livest Sci 130:47–56
Ellis JL, Dijkstra J, Bannink A, Kebreab E, Hook SE, Archibeque S, France J (2012) Quantifying the effect of monensin dose on the rumen volatile fatty acid profile in high-grain fed beef cattle. J Anim Sci 90:2717–2726
FAO (2012) Balanced feeding for improving livestock productivity – increase in milk production and nutrient use efficiency and decrease in methane emission. In: Garg MR (ed) FAO Anim Prod Health Pap No. 173. Rome. 1080/00330124.2014.921017 (2014)
Foley PA, Kenny DA, Callan JJ, Boland TM, O’Mara FP (2009) Effect of dl-malic acid supplementation on feed intake, methane emission, and rumen fermentation in beef cattle. J Anim Sci 87:1048–1057
Fox NJ, Marion G, Davidson RS, White PCL, Hutchings MR (2012) Livestock helminths in a changing climate: approaches and restrictions to meaningful predictions (review). Animals 2:93–107
Francis G, Kerem Z, Makkar HSP, Becker K (2002) The biological action of saponins in animal systems: a review. Br J Nutr 88:687–695
Friedman E, Voet H, Reznikov D, Dagoni I, Roth Z (2011) Induction of successive follicular waves by gonadotropin-releasing hormone and prostaglandin F2α to improve fertility of high-producing cows during the summer and autumn. J Dairy Sci 94:2393–2402
Gerber PJ, Steinfeld H, Henderson B, Mottet A, Opio C, Dijkman J, Falcucci A, Tempio G (2013) Tackling climate change through livestock – a global assessment of emissions and mitigation opportunities. Food and Agriculture Organization of the United Nations (FAO), Rome
Goel G, Makkar HPS (2012) Methane mitigation from ruminants using tannins and saponins. Trop Anim Health Prod 44:729–739
Goel G, Makkar HPS, Becker K (2008) Changes in microbial community, structure, methanogenesis and rumen fermentation in response to saponin-rich fractions from different plant materials. J Appl Microbiol 105:770–777
Gómez JA, Tejido ML, Carro MD (2005) Mixed rumen microorganisms growth and rumen fermentation of two diets in RUSITEC fermenters: influence of disodium malate supplementation. Br J Nutr 93:479–484
Gutierrez-Banuelos H, Anderson RC, Carstens GE, Slay LJ, Ramlachan N, Horrocks SM, Callaway TR, Edrington TS, Nisbet DJ (2007) Zoonotic bacterial populations, gut fermentation characteristics and methane production in feedlot steers during oral nitroethane treatment and after the feeding of an experimental chlorate product. Anaerobe 13:21–31
Hahn GL (1981) Housing and management to reduce climatic impacts on livestock. J Anim Sci 52:175–186
Haisan J, Sun Y, Beauchemin K, Guan L, Duval S, Barreda DR, Oba M (2013) Effect of feeding 3-nitrooxypropanol on methane emissions and productivity of lactating dairy cows. Adv Anim Biosci 4(2):260
Hamano Y (2012) Alleviative effects of a-lipoic acid supplementation on acute heat stress-induced thermal panting and the level of plasma nonesterified fatty acids in hypothyroid broiler chickens. Br Poult Sci 53:125–133
Hanson RS, Hanson TE (1996) Methanotropic bacteria. Microbiol Rev 60(2):439–471
Hatayama T, Asai Y, Wakatsuki T, Kitamura T, Imahara H (1993) Regulation of hsp70 synthesis induced by cupric sulfate and zinc sulfate in thermotolerant HeLa cells. J Biochem (Tokyo) 114:592–597
Hegarty RS (2001) Strategies for mitigating methane emissions from livestock – Australian options and opportunities. In: 1st international conference on GHGes and animal agriculture, Obihiro, pp 31–34
Hollmann M, Beede DK (2012) Comparison of effects of dietary coconut oil and animal fat blend on lactational performance of Holstein cows fed a high-starch diet. J Dairy Sci 95:1484–1499
Hook SE, Northwood KS, Wright AD, McBride BW (2009) Long-term monensin supplementation does not significantly affect the quantity or diversity of methanogens in the rumen of the lactating dairy cow. Appl Environ Microbiol 75:374–380
Hristov AN, Ivan M, Neill L, McAllister TA (2003) A survey of potential bioactive agents for reducing protozoal activity in vitro. Anim Feed Sci Technol 105:163–184
Hristov AN, Vander Pol M, Agle M, Zaman S, Schneider C, Ndegwa P, Vaddella VK, Johnson K, Shingfield KJ, Karnati SKR (2009) Effect of lauric acid and coconut oil on ruminal fermentation, digestion, ammonia losses from manure, and milk fatty acid composition in lactating cows. J Dairy Sci 92:5561–5582
Hristov AN, Lee C, Cassidy T, Long M, Heyler K, Corl B, Forster R (2011) Effects of lauric and myristic acids on ruminal fermentation, production, and milk fatty acid composition in lactating dairy cows. J Dairy Sci 94:382–395
Hristov AN, Oh J, Lee C, Meinen R, Montes F, Ott T, Firkins J, Rotz CA, Dell C, Adesogan A, Yang WZ, Tricarico J, Kebreab E, Waghorn G, Dijkstra J, Oosting S (2013) Mitigation of greenhouse gas emissions in livestock production – a review of technical options for non-CO2 emissions. In: Gerber P, Henderson B, Makkar H (eds) FAO animal production and health. FAO, Rome, p 177
Hulshof RBA, Berndt A, Gerrits WJJ, Dijkstra J, van Zijderveld SM, Newbold JR, Perdok HB (2012) Dietary nitrate supplementation reduces methane emission in beef cattle fed sugarcane-based diets. J Anim Sci 90:2317–2323
IPCC (2001) Technical summary: contribution of Working Group I to the Third Assessment Report. Inter governmental panel on climate change, Jan 2001
IPCC (2007) Climate change 2007: the physical science basis, summary for policy makers. Contribution of working group I to the fourth assessment report of intergovernmental panel on climate change. http://www.ipcc.ch
IPCC (Intergovernmental Panel on Climate Change) (2013) Impacts, adaptation and vulnerability. http://ipcc-wg2.gov/AR5/images/uploads/WGIIAR5-TS_FGDall.pdf
Itabashi H, Kobayashi T, Matsumoto M (1984) The effects of rumen ciliate protozoa on energy metabolism in some constituents in rumen fluid and blood plasma of goats. Jpn J Zoo Tech Sci 55:248
Jayanegara A, Leiber F, Kreuzer M (2012) Meta-analysis of the relationship between dietary tannin level and methane formation in ruminants from in vivo and in vitro experiments. J Anim Physiol Anim Nutr 96:356–375
Johnson KA, Johnson DE (1995b) Methane emissions from cattle methane emissions from cattle. J Anim Sci 73:2483–2492
Johnson DE, Ward GW, Ramsey JJ (1996) Livestock methane: current emissions and mitigation potential. In: Kornegay ET (ed) Nutrient management of food animals to enhance and protect the environment. Lewis Publishers, New York, pp 219–234
Jordan E, Lovett DK, Monahan FJ, Callan J, Flynn B, O’Mara FP (2006) Effect of refined coconut oil or copra meal on methane output and on intake and performance of beef heifers. J Anim Sci 84:162–170
Jousan FD, Hansen PJ (2004) Insulin-like growth factor-I as a survival factor for the bovine preimplantation embryo exposed to heat shock. Biol Reprod 71:1665–1670
Jousan FD, Hansen PJ (2007) Insulin-like growth factor-I promotes resistance of bovine preimplantation embryos to heat shock through actions independent of its anti-apoptotic actions requiring PI3K signaling. Mol Reprod Dev 74:189–196
Kabubo-Mariara J (2005) Herders response to acute land pressure under changing property rights: some insights from Kenya. Environ Dev Econ 10(1):67–85
Kabubo-Mariara J (2007) Poverty and rural livelihoods in Kenya: evidence from a semi-arid region. In: Tisdell C (ed) Poverty, poverty alleviation and social disadvantage: analysis, case studies and policies. Serials Publications, Vol III, Part VII, Chapter 56
Kabubo-Mariara J (2008) Climate change adaptation and livestock activity choices in Kenya: an economic analysis. Nat Res Forum 32:131–141
Kaim M, Bloch A, Wolfenson D, Braw-Tal R, Rosenberg M, Voet H, Folman Y (2003) Effects of GnRH administered to cows at the onset of estrus on timing of ovulation, endocrine responses and conception rates. J Dairy Sci 86:2012–2021
Kingeston-Smith AH, Edwards JE, Huws SA, Kim EJ, Abberton M (2010) Plant based strategies towards minimizing live stocks long shadow. Proc Nutr Soc 69:613–620
Kirchgessner M, Windisch W, Muller HL (1995) Nutritional factors for quantification of methane production. Ruminant physiology, digestion metabolism growth and reproduction. In: Proceedings of the 8th international symposium on ruminant physiology. Ferdinand Enke Verlag, Stuttgart, pp 333–348
Klieve AV, Joblin K (2007) Comparison in hydrogen utilisation of ruminal and marsupial reductive acetogens. In: Kennedy R (ed) 5 year research progress report 2002–2007. The pastoral greenhouse gas research consortium, Wellington, pp 34–35
Knight T, Ronimus RS, Dey D, Tootill C, Naylor G, Evans P, Molano G, Smith A, Tavendale M, Pinares-Patino CS, Clark H (2011) Chloroform decreases rumen methanogenesis and methanogen populations without altering rumen function in cattle. Anim Feed Sci Technol 166:101–112
Kreuzer M, Kirchgessner M, Müller HL (1986) Effect of defaunation on the loss of energy in wethers fed different quantities of cellulose and normal or steam-flaked maize starch. Anim Feed Sci Technol 16:233–241
Lascano CE, Cárdenas E (2010) Alternatives for methane emission mitigation in livestock systems. Rev Bras Zootec 39:175–182
Le-Liboux S, Peyraud JL (1999) Effect of forage particle size and feeding frequency on fermentation patterns and sites and extent of digestion in dairy cows fed mixed diets. Anim Feed Sci Technol 76:297–319
Leahy SC, Kelly WJ, Altermann E, Ronimus RS, Yeoman CJ, Pacheco M, Li D, Kong Z, McTavish S, Sang C, Lambie SC, Janssen PH, Dey D, Attwood GT (2010) The genome sequence of the rumen Methanogen Methanobrevibacter reveals new possibilities for controlling ruminant methane emissions. PLoS ONE 5:e8926
Lee SS, Hsu JT, Mantovani HC, Russell JB (2002) The effect of bovicin HC5, a bacteriocin from Streptococcus bovis HC5, on ruminal methane production in vitro. FEMS Microbiol Lett 217:51–55
Leng RA (2008) The potential of feeding nitrate to reduce enteric methane production in ruminants. http://www.penambulbooks.com/Papers%20&%20Presentations.htm
Lopez S, McIntosh FM, Wallace RJ, Newbold CJ (1999) Effect of adding acetogenic bacteria on methane production by mixed rumen microorganisms. Anim Feed Sci Technol 78:1–9
Lovett D, Bortolozzo A, Conaghan P, O’Kiely P, O’Mara FP (2004) In vitro total and methane gas production as influenced by rate of nitrogen application, season of harvest and perennial ryegrass cultivar. Grass Forage Sci 59:227–232
Lovett DK, Stack LJ, Lovell S, Callan J, Flynn B, Hawkins M, O’Mara FP (2005) Manipulating enteric methane emissions and animal performance of late-lactation dairy cows through concentrate supplementation at pasture. J Dairy Sci 88:2836–2842
Luo Y, Pfister P, Leisinger T, Wasserfallen A (2001) The genome of archaeal prophage ψm100 encodes the lytic enzyme responsible for autolysis of Methanothermobacter wolfeii. J Bacteriol 183:5788–5792
Mamuad LL, Kim SH, Lee SS, Cho KK, Jeon CO, Lee SS (2012) Characterization, metabolites and gas formation of fumarate reducing bacteria isolated from Korean native goat (Capra hircus coreanae). J Microbiol 50:925–931
Mantovani HC, Russell JB (2002) The ability of a bacteriocin of streptococcus bovis HC5 (bovicin HC5) to inhibit clostridium aminophilum, an obligate amino acid fermenting bacterium from the rumen. Anaerobe 8:247–252
Martin C, Dubroeucq H, Micol D, Agabriel J, Doreau M (2007) Methane output from beef cattle fed different high-concentrate diets. In: Proceedings of the British Society of Animal Science, Southport, 2–4 Apr 2007, p 46
Martinez-Fernandez G, Arco A, Abecia L, Cantalapiedra-Hijar G, Molina-Alcaide E, Martin-Garcia AI, Kindermann M, Duval S, Yanez-Ruiz DR (2013) The addition of ethyl-3-nitrooxy propionate and 3-nitrooxypropanol in the diet of sheep sustainably reduces methane emissions and the effect persists over a month. Adv Anim Biosci 4(2):368
Masike S (2007) The impacts of CC on cattle water demand and supply in Khurutshe, Botswana. PhD thesis, University of Waikato, New Zealand
Mathison GW, Okine EK, McAllister TA, Dong Y, Galbraith J, Dmytruk OIN (1998) Reducing methane emissions from ruminant animals. J Appl Anim Res 14:1–28
McAllister TA, Newbold CJ (2008) Redirecting rumen fermentation to reduce methanogenesis. Aust J Exper Agri 48:7–13
McAllister TA, Okine EK, Mathison GW, Cheng KJ (1996) Dietary, environmental and microbiological aspects of methane production in ruminants. Can J Anim Sci 76:231–243
McCaughey WP, Wittenberg KM, Corrigan D (1997) Methane production by steers on pasture. Can J Anim Sci 77:519–524
McCaughey WP, Wittenberg K, Corrigan D (1999) Impact of pasture type on methane production by lactating cows. Can J Anim Sci 79:221–226
McGinn SM, Beauchemin KA, Coates T, Colombatto D (2004) Methane emissions from beef cattle: effects of monensin, sunflower oil, enzymes, yeast and fumaric acid. J Anim Sci 82:3346–3356
MEA (2005) Ecosystems and human well-being: our human planet. Summary for decision makers. The Millennium Ecosystem Assessment, online at http://www.millenniumassessment.org
Mills JA, Dijkstra J, Bannink A, Cammel SB, Kebreab E, France J (2001) A mechanistic model of whole-tract digestion and methanogenesis in the lactating dairy cow: model development, evaluation, and application. J Anim Sci 79:1584–1597
Mitsumori M, Shinkai T, Takenaka A, Enishi O, Higuchi K, Kobayashi Y, Nonaka I, Asanuma N, Denman SE, McSweeney CS (2011) Responses in digestion, rumen fermentation and microbial populations to inhibition of methane formation by a halogenated methane analogue. Br J Nutr 8:1–10
Molano G, Knight TW, Clark H (2008) Fumaric acid supplements have no effect on methane emissions per unit of feed intake in wether lambs. Aust J Exp Agric 48:165–168
Morgavi DP, Forano E, Martin C, Newbold CJ (2010) Microbial ecosystem and methanogenesis in ruminants. Animal 4:1024–1036
Morgavi DP, Martin C, Boudra H (2013) Fungal secondary metabolites from Monascus spp. reduce rumen methane production in vitro and in vivo. J Anim Sci 91:848–860
Moss AR, Jouany JP, Newbold J (2000) Methane production by ruminants: its contribution to global warming. Ann Zootech 49:231–253
Mwenya B, Santoso B, Sar C, Gamo Y, Kobayashi T, Arai I, Takahashi J (2004) Effects of including β1-4 galacto-oligosaccharides lactic acid bacteria or yeast culture on methanogenesis as well as energy and nitrogen metabolism in sheep. Anim Feed Sci Technol 115:313–326
Newbold CJ, Rode LM (2005) Dietary additives to control methanogenesis in the rumen. In: Soliva CR, Takahashi J, Kreuzer M (eds) Second international conference on greenhouse gases and animal agriculture, Working Papers, ETH, Zurich, pp 60–70
Newbold CJ, Rode LM (2006) Dietary additives to control methanogenesis in the rumen. In: Soliva CR, Takahashi J, Kreuzer M (eds) Greenhouse gases and animal agriculture: an update, vol 1293, Elsevier International Congress Series. Elsevier, Amsterdam, pp 138–147
Nolan JV, Hegarty RS, Hegarty J, Godwin I, Woodgate R (2010) Effects of dietary nitrate on rumen fermentation, methane production and water kinetics in sheep. Anim Prod Sci 50:801–806
Nollet L, Mbanzamihigo L, Demeyer D, Verstraete W (1998) Effect of the addition of Peptostreptococcus productus ATCC 35244 on reductive acetogenesis in the ruminal ecosystem after inhibition of methanogenesis by cell-free supernatant of Lactobacillus plantarum 80. Anim Feed Sci Technol 71:49–66
O’Mara F (2004) GHG production from dairying: reducing methane production. Adv Dairy Technol 16:295–309
Odongo NE, Bagg R, Vessie G, Dick P, Or-Rashid MM, Hook SE, Gray JT, Kebreab E, France J, McBride BW (2007) Long-term effects of feeding monensin on methane production in lactating dairy cows. J Dairy Sci 90:1781–1788
Ohene-Adjei S, Chaves AV, McAllister TA, Benchaar C, Teather RM, Forster RJ (2008) Evidence of increased diversity of methanogenic archaea with plant extract supplementation. Microbial Ecol 56:234–242
Paaijmans KP, Blanford S, Bell AS, Blanford JI, Read AF, Thomas MB (2010) Influence of climate on malaria transmission depends on daily temperature variation. Proc Natl Acad Sci U S A 107:15135–15139
Patra AK, Yu Z (2012) Effects of essential oils on methane production and fermentation by and abundance and diversity of rumen microbial populations. Appl Environ Microbiol 78:4271–4280
Pfister P, Wasserfallen A, Stettler R, Leisinger T (1998) Molecular analysis of Methanobacterium phage ΨM2. Mol Microbiol 30:233–244
Pinares-Patiño CS, Hour PD, Jouany JP, Martin C (2007) Effects of stocking rate on methane and carbon dioxide emissions from grazing cattle. Agric Ecosyst Environ 121:30–46
Popova M, Martin C, Eugène M, Mialon MM, Doreau M, Morgavi DP (2011) Effect of fibre- and starch-rich finishing diets on methanogenic Archaea diversity and activity in the rumen of feedlot bulls. Anim Feed Sci Technol 166:113–121
Quere CL, Peters GP, Andrew RM, Boden TA, Ciais P, Friedlingstein P, Houghton RA et al (2014) Global carbon budget 2013. Earth Syst Sci Data 6:235–263
Roth Z, Arav A, Braw-Tal R, Bor A, Wolfenson D (2002) Effect of treatment with follicle-stimulating hormone or bovine somatotropin on the quality of oocytes aspirated in the autumn from previously heat-stressed cows. J Dairy Sci 85:1398–1405
Rotz CA, Corson MS, Chianese DS, Coiner CU (2009) The integrated farm system model: reference manual. University Park, Pa: USDAARS Pasture Systems and Watershed Management research unit: www.ars.usda.gov/SP2UserFiles/Place/19020000/ifsmreference
Rowlinson P, Steele M, Nefzaoui A (2008) Livestock and global climate change. Proceedings of the international conference in Hammamet. Cambridge University Press, pp 216–216
Rutledge JJ (2001) Use of embryo transfer and IVF to bypass effects of heat stress. Theriogenology 55:105–111
Sakatani M, Kobayashi S, Takahashi M (2004) Effects of heat shock on in vitro development and intracellular oxidative state of bovine preimplantation embryos. Mol Reprod Dev 67:77–82
Sang SL, Mantovani HC, Russell JB (2002) The binding and degradation of nisin by mixed ruminal bacteria. FEMS Microbiol Ecol 42:339–345
Santoso B, Mwenyaa B, Sara C, Gamoa Y, Kobayashia T, Morikawaa R, Kimurab K, Mizukoshib H, Takahashi J (2004) Effects of supplementing galacto-oligosaccharides, Yucca schidigera or nisin on rumen methanogenesis, nitrogen and energy metabolism in sheep. Livest Prod Sci 91:209–217
Sar C, Santoso B, Gamo Y, Kobayashi T, Shiozaki S, Kimura K, Mizukoshi H, Arai I, Takahashi J (2004) Effects of combination of nitrate with β1-4 galacto-oligosaccharides and yeast (Candida kefyr) on methane emission from sheep. Asian Austr J Anim Sci 17(1):73–79
Sar C, Mwenya B, Santoso B, Takaura K, Morikawa R, Isogai N, Asakura Y, Toride Y, Takahashi J (2005) Effect of Escherichia coli wild type or its derivative with high nitrite reductase activity on in vitro ruminal methanogenesis and nitrate/nitrite reduction. J Anim Sci 83:644–652
Sejian V (2013) Climate change: impact on production and reproduction, adaptation mechanisms and mitigation strategies in small ruminants: a review. Indian J Small Rumin 19(1):1–21
Sejian V, Naqvi SMK (2012) Livestock and climate change: mitigation strategies to reduce methane production. In: Liu G (ed) Greenhouse gases – capturing, utilization and reduction. Intech Publisher, Croatia, pp 254–276
Sejian V, Lal R, Lakritz J, Ezeji T (2011a) Measurement and prediction of enteric methane emission. Int J Biometeorol 55:1–16
Sejian V, Lakritz J, Ezeji T, Lal R (2011b) Forage and flax seed impact on enteric methane emission in dairy cows. Res J Vet Sci 4:1–8
Sejian V, Rotz A, Lakritz J, Ezeji T, Lal R (2011c) Modeling of green house gas emissions in dairy farms. J Anim Sci Adv 1(1):12–20
Sejian V, Indu S, Ujor V, Ezeji T, Lakritz J, Lal R (2012a) Global climate change: enteric methane reduction strategies in livestock. In: Sejian V, Naqvi SMK, Ezeji T, Lakritz J, Lal R (eds) Environmental stress and amelioration in livestock production. Springer-Verlag GMbH Publisher, Germany, pp 469–502
Sejian V, Valtorta S, Gallardo M, Singh AK (2012b) Ameliorative measures to counteract environmental stresses. In: Sejian V, Naqvi SMK, Ezeji T, Lakritz J, Lal R (eds) Environmental stress and amelioration in livestock production. Springer-Verlag GMbH Publisher, Germany, pp 153–180
Sejian V, Singh AK, Sahoo A, Naqvi SMK (2014) Effect of mineral mixture and antioxidant supplementation on growth, reproductive performance and adaptive capability of Malpura ewes subjected to heat stress. J Anim Physiol Anim Nutr 98:72–83
Seyrek K, Kargin KF, Bildik A (2004) Chronic ethanol induced oxidative alterations in the rat tissues and protective effect of vitamin E. Ind Vet J 81:1102–1104
Shibata M (1996) Factors affecting thermal balance and production of ruminants in a hot environment – a review. Memories of National Institute of Animal Industry No. 10. 60 pp
Shinde AK, Sejian V (2013) Sheep husbandry under changing climate scenario in India: an overview. Indian J Anim Sci 83(10):998–1008
Sidahmed A (2008) Livestock and CC: coping and risk management strategies for a sustainable future. In: Livestock and global CC conference proceeding, May 2008, Tunisia
Skuce PJ, Morgan ER, van Dijk J, Mitchell M (2013) Animal health aspects of adaptation to CC: beating the heat and parasites in a warming Europe. Animal 7(2):333–345
Soussana JF, Loiseau P, Vuichard N, Ceschia E, Balesdent J, Chevallier T, Arrouays D (2004) Carbon cycling and sequestration opportunities in temperate grasslands. Soil Use Manag 20:219–230
Spears JW, Whisnant CS, Huntington GB, Lloyd KE, Fry RS, Krafka K, Lamptey A, Hyda J (2012) Chromium propionate enhances insulin sensitivity in growing cattle. J Dairy Sci 95:2037–2045
Staerfl SM, Amelchanka SL, Kälber T, Soliva CR, Kreuzer M, Zeitz JO (2012) Effect of feeding dried high-sugar ryegrass (‘AberMagic’) on methane and urinary nitrogen emissions of primiparous cows. Livest Sci 150:293–301
Stefanidou M, Maravelias C, Dona A, Spiliopoulou C (2006) Zinc: a multipurpose trace element. Arch Toxicol 80:1–9
Suk JE, Semenza JC (2011) Future infectious disease threats to Europe. Am J Public Health 101:2068–2079
Sunil Kumar BV, Singh G, Meur SK (2010) Effects of addition of electrolyte and ascorbic acid in feed during heat stress in buffaloes. Asian Aust J Anim Sci 23(7):880–888
Takahashi J (2011) Some prophylactic options to mitigate methane emission from animal agriculture in Japan. Asian Aust J Anim Sci 24:285–294
Tejido ML, Ranilla MJ, García-Martínez R, Carro MD (2005) In vitro microbial growth and rumen fermentation of different diets as affected by the addition of disodium malate. Anim Sci 81:31–38
Tezel U, Pierson JA, Pavlostathis SG (2006) Fate and effect of quaternary ammonium compounds on a mixed methanogenic culture. Water Res 40(19):3660–3668
Thames JL, Cluff CB (1982) Water harvesting on surface mine spoils. Paper presented at the Wisconsin mined-land rehabilitation research workshop, Ft. Collins, Colorado, 1–11 June 1982, 4 pages
Thornton PK, Herrero M (2010) The inter-linkages between rapid growth in livestock production, CC, and the impacts on water resources, land use, and deforestation. Policy research working paper 5178, pp 1–82
Tomkins WW, Colegate SM, Hunter RA (2009) A bromochloromethane formulation reduces enteric methanogenesis in cattle fed grain-based diets. Anim Prod 49(12):1053–1058
Ulyatt MJ, Lassey KR, Shelton ID, Walker CF (2002) Seasonal variation in methane emission from dairy cows and breeding ewes grazing ryegrass/white clover pasture in New Zealand. N Z J Agric Res 45:217–226
Ungerfeld E, Forster RJ (2011) A meta-analysis of malate effects on methanogenesis in ruminal batch cultures. Anim Feed Sci Technol 166–167:282–290
Ungerfeld EM, Rust SR, Burnett R (2003) Use of some novel alternative electron sinks to inhibit ruminal methanogenesis. Reprod Nutr Dev 43:189–202
Ungerfeld EM, Kohn RA, Wallace RJ, Newbold CJ (2007) A meta-analysis of fumarate effects on methane production in ruminal batch cultures. J Anim Sci 85:2556–2563
USEPA (1993) Anthropogenic methane emissions in the United States. Estimates for 1990: report to congress. Atmospheric PolluStatestion Prevention Division, Office of Air and Radiation, US, Washington, DC
Valtorta SE, Leva PE, Gallardo MR, Scarpati OE (2002) Milk production responses during heat waves events in Argentina. In: 15th conference on biometeorology and aerobiology – 16th international congress on biometeorology. Kansas City, MO. American Meteorological Society, Boston p 98–101
Van Dorland HA, Wettstein HR, Leuenberger H, Kreuzer M (2007) Effect of supplementation of fresh and ensiled clovers to ryegrass on nitrogen loss and methane emissions of dairy cows. Livest Sci 111:57–69
van Nevel CJ, Demeyer DI (1995) Feed additives and other interventions for decreasing methane emissions. Biotechnol Anim Feeds Feed 17:329–349
Van Nevel C, Demeyer D (1996) Control of rumen methanogenesis. Environ Monit Assess 42:73–97
Van Zijderveld SM, Gerrits WJJ, Apajalahti JA, Newbold JR, Dijkstra J, Leng RA, Perdok HB (2010) Nitrate and sulfate: effective alternative hydrogen sinks for mitigation of ruminal methane production in sheep. J Dairy Sci 93:5856–5866
Van Zijderveld SM, Gerrits WJJ, Dijkstra J, Newbold JR, Hulshof RBA, Perdok HB (2011) Persistency of methane mitigation by dietary nitrate supplementation in dairy cows. J Dairy Sci 94:4028–4038
Vercruysse J, Schetters T, Knox DP, Willadsen P, Claerebout E (2007) Control of parasitic disease by vaccination – an answer to drug resistance? Rev Sci Technol 26:105–115
Voellmy R (2004) On mechanisms that control heat shock transcription factor activity in metazoan cells. Cell Stress Chaperones 9:122–133
Waghorn GC, Tavendale MH, Woodfield DR (2002) Methanogenesis from forages fed to sheep. Proc N Z Grassl Assoc 64:167–171
Wang CJ, Wang SP, Zhou H (2009) Influences of flavomycin, ropadiar, and saponin on nutrient digestibility, rumen fermentation, and methane emission from sheep. Anim Feed Sci Technol 148:157–166
Wedlock DN, Pedersen G, Denis M, Dey D, Janssen PH, Buddle BM (2010) Development of a vaccine to mitigate greenhouse gas emissions in agriculture; vaccination of sheep with methanogen fractions induces antibodies that block methane production in vitro. N Z Vet J 58:29–36
Weimer PJ (1998) Manipulating ruminal fermentation: a microbial ecological perspective. J Anim Sci 76:3114–3122
Weiss RA, McMichael AJ (2004) Social and environmental risk factors in the emergence of infectious diseases. Nat Med 10:S70–S76
West JW (1999) Nutritional strategies for managing the heat stressed dairy cow. J Anim Sci 77(2):21–35
West JW (2002) Physiological effects of heat stress on production and reproduction. Proc Tri-State Nutr Conf 1–9
Whitelaw FG, Eadie WJ, Bruce LA, Shand WA (1984) Methane formation in faunated and ciliate-free cattle and its relationship with rumen volatile fatty acid proportions. Br J Nutr 52:261
Wilk J, Wittgren HB (eds) Adapting water management to CC. Swedish Water House Policy Brief Nr. 7. SIWI 2009
Williams YJ, Rea SM, Popovski S, Pimm CL, Williams AJ, Toovey AF, Skillman LC, Wright ADG (2008) Responses of sheep to a vaccination of entodinial or mixed rumen protozoal antigens to reduce rumen protozoal numbers. Br J Nutr 99:100–109
Williams YJ, Popovski S, Rea SM, Skillman LC, Toovey AF, Northwood KS, Wright ADG (2009) A vaccine against rumen methanogens can alter the composition of archaeal populations. Appl Environ Microbiol 75:1860–1866
Wina E, Muetzel S, Hoffmann E, Makkar HPS, Becker K (2005) Saponins containing methanol extract of Sapindus rarak affect microbial fermentation, microbial activity and microbial community structure in vitro. Anim Feed Sci Technol 121:159–174
Wood TA, Wallace RJ, Rowe A, Price J, Yáňez-Ruiz DR, Murray P, Newbold CJ (2009) Encapsulated fumaric acid as a feed ingredient to decrease ruminal methane emissions. Anim Feed Sci Technol 152:62–71
World Meteorological Organization (2014) Weather and climate: engaging youth. WMO Bull 63(1):1–44
Wright AD, Kennedy P, O’Neill CJ, Toovey AF, Popovski S, Rea SM, Pimm CL, Klein L (2004) Reducing methane emissions in sheep by immunization against rumen methanogens. Vaccine 22:3976–3985
Yates CM, Cammell SB, France J, Beever DE (2000) Predictions of methane emissions from dairy cows using multiple regression analysis. In: Proceedings of the British Society of Animal Science, Scarborough, UK, March 2000, pp 94
Zhang Y, Bi P, Hiller JE (2008) CC and the transmission of vector-borne diseases: a review. Asia Pac J Public Health 20:64–76
Zhou X, Sar C, Kobayashi T, Takahashi J, Santoso B, Gamo Y, Mwenya B (2004) Effects of probiotic vitacogen and ß1-4 galacto-oligosaccharides supplementation on methanogenesis and energy and nitrogen utilization in dairy cows. Asian Aust J Anim Sci 17(3):349–354
Zhou M, Chung YH, Beauchemin KA, Holtshausen L, Oba M, McAllister TA, Guan LL (2011) Relationship between rumen methanogens and methane production in dairy cows fed diets supplemented with a feed enzyme additive. J Appl Microbiol 111:1148–1158
Zhou X, Meile L, Kreuzer M, Zeitz JO (2013) The effect of lauric acid on methane production and cell viability of Methanobrevibacter ruminantium. Adv Anim Biosci 4(2):458
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer India
About this chapter
Cite this chapter
Sejian, V. et al. (2015). Overview on Adaptation, Mitigation and Amelioration Strategies to Improve Livestock Production Under the Changing Climatic Scenario. In: Sejian, V., Gaughan, J., Baumgard, L., Prasad, C. (eds) Climate Change Impact on Livestock: Adaptation and Mitigation. Springer, New Delhi. https://doi.org/10.1007/978-81-322-2265-1_22
Download citation
DOI: https://doi.org/10.1007/978-81-322-2265-1_22
Publisher Name: Springer, New Delhi
Print ISBN: 978-81-322-2264-4
Online ISBN: 978-81-322-2265-1
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)