
Abstract
Although originally proposed as functional tuners capable of influencing signaling thresholds of immune cells, emerging evidence indicates a more central and hierarchical role of glycans and glycan-binding proteins as stimulators or inhibitors of innate and adaptive immunity. An increasing number of studies reveal the relevance of glycan-containing information in host-pathogen recognition and the control of immune cell homeostasis, inflammation, and tumor immunity. In this chapter, we summarize general concepts of glycoimmunology and illustrate selected examples of the contribution of glycans and glycan-binding proteins (C-type lectins, siglecs, and galectins) to innate and adaptive immune responses in physiologic and pathologic settings.
Access provided by Autonomous University of Puebla. Download reference work entry PDF
Similar content being viewed by others


Keywords
Introduction: Glycans as Central Regulators of Immune Responses
Genetic manipulation of components of the glycosylation machinery has demonstrated essential roles of glycans and glycan-binding proteins in innate and adaptive immunity (Baum and Crocker 2009). Elegant studies over the past decade highlighted the contribution of N-glycans to the development of spontaneous T-cell-mediated autoimmunity. Deficiency in β1,6 N-glycan branching in N-acetylglucosaminyltransferase 5 (MGAT5)-deficient mice resulted in sustained T-cell activation which led to a spontaneous demyelinating and neurodegenerative disease that recapitulated multiple sclerosis (MS). Supporting these findings, recent studies in MS patients revealed that dysregulated Golgi N-glycosylation is a final common pathway in which disease-associated environmental factors and multiple genetic variants (IL-7RA, IL-2RA, and CTLA-4) converge (Dennis et al. 2009; Rabinovich and Croci 2012).
Another paradigmatic example illustrating the role of N-glycans in immunity has been reported in mice deficient in the α-mannosidase (αM)-II. Targeted deletion of αM-II led to diminished N-glycan branching and occurrence of an autoimmune disease similar to human systemic lupus erythematosus (SLE). A mechanistic analysis revealed that, under αM-II deficiency, an atypical exposure of cryptic N-glycans (high mannose-containing ligands) occurs, which enables recognition by mannose-binding lectins such as mannose receptor (MR). Because these particular glycan epitopes are typically expressed by pathogenic bacteria, lack of αM-II recapitulates bacterial infection leading to aberrant activation of innate immunity and promotes development of an autoimmune disease similar to that observed in mice lacking the MR (Marth and Grewal 2008).
Likewise, alterations in O-glycans can also promote initiation and perpetuation of inflammation and autoimmune responses. Disruption of core-3 β1,3-N-acetylglucosaminyltransferase (B3GNT6; encoded by C3GNT gene), an enzyme responsible for the synthesis of core-3-derived O-glycans, leads to aberrant exposure of naked Tn antigen in colonic epithelium and development of inflammatory colitis and colon adenocarcinoma, emphasizing the protective role of core-3-derived O-glycans as essential components of intestinal mucins. Accordingly, mutations in the molecular “chaperone” Cosmc, responsible for the correct folding of the core-1 β(1–3)-galactosyltransferase (T-synthase), generate a truncated O-glycan composed of only GalNAc, which is the cause of the rare “Tn syndrome” characterized by IgA nephropathy (Cummings and Pierce 2014; Rabinovich and Croci 2012). These studies provided categorical evidence that alterations of N- and O-glycosylation pathways, either in experimental models or clinical settings, result in abnormal activation of immune cell pathways and development of inflammatory responses.
Interestingly, differential glycosylation can also influence pathogenic or regulatory responses evoked by circulating immunoglobulins. Elegant studies demonstrated that the anti-inflammatory activity of intravenous immunoglobulins (IVIGs), commonly used for the treatment of autoimmune diseases, is triggered by a minor population of IgG Fcs expressing α2,6-linked sialic acid. The authors demonstrated that sialylated IgG targets myeloid regulatory cells expressing DC-SIGN, which triggers IL-33-mediated Th2 responses and expansion of macrophages expressing the FcγRIIB inhibitory receptor. In contrast, agalactosyl and asialoglycoforms of IgG have been linked to the pathogenesis of autoimmune diseases, including rheumatoid arthritis and IgA nephropathy (van Kooyk and Rabinovich 2008).
In this regard, recent studies highlighted the contribution of cell surface glycosylation to cancer immunoediting. Glycan and glycan-binding proteins can influence the creation of immunosuppressive networks that thwart antitumor responses and promote tumor growth and metastasis. Overexpression of the C2GnT1 glycosyltransferase on bladder cancer cells has been shown to modulate NK cell-mediated tumor immunity. These effects were mainly due to the interactions between galectin-3 and poly-LacNAc-branched core-2 O-glycans decorating tumor-associated major histocompatibility complex class I-related chain A (MICA) which reduces the affinity of MICA for the activating NK cell receptor NKG2D, thereby impairing NK cell activation and antitumor activity. In this regard, Siglec-7 and Siglec-9 ligands can confer susceptibility to NK cell-mediated cytotoxicity. Thus, changes in the glycome occurring during tumor growth and metastasis contribute to activate regulatory circuits that profoundly affect the cancer immunoediting process (Rabinovich and Croci 2012). Moreover, recent studies highlighted the critical role of glycans in mucosal immunity. Glycans associated with MUC2 imprinted mucosal DCs with tolerogenic properties by assembling a galectin-3-Dectin-1-FcγRIIB receptor complex that activates β-catenin and contributes to inhibition of pro-inflammatory responses and generation of an immunosuppressive mucosal microenvironment (Shan et al. 2013). In addition, recent studies demonstrated that nutrients can regulate transcription of immune-related genes through dynamic cycling of O-linked N-acetylglucosamine (O-GlcNAc) on nuclear factor-κB (NF-κB) transcription factor (Hart 2013).
As many of the functions of glycans are related to their lectin-binding activity, in the next sections we will highlight biochemical and functional aspects of three families of biochemically unrelated glycan-binding proteins (C-type lectins, siglecs, and galectins) which play key roles in assembling and/or disarming immune cell networks.
C-Type Lectins
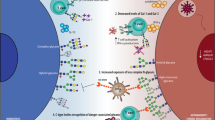
Members of the C-type lectin receptor (CLR) superfamily play key roles in tailoring innate and adaptive immune responses. This lectin family is divided into two categories and subclassified into 17 subgroups (I–XVII). Of these, the most studied are the integral membrane CLRs belonging to the subgroups II, V, and VI that are expressed prominently in monocytes, macrophages, dendritic cells (DCs), and granulocytes. Type I CLRs contain an amino-terminal cysteine-rich repeat (S–S) domain, a fibronectin type II repeat domain, followed by eight or ten carbohydrate recognition domains (CRDs), which bind ligands in a Ca2+-dependent manner. These include the MR and DEC-205. Type II CLRs contain only one CRD at their carboxy-terminal extracellular domain, and the recognition of the glycan can be either Ca2+ dependent or Ca2+ independent. Macrophage galactose lectin (MGL), Dectin-2, macrophage-inducible C-type lectin (Mincle), dendritic cell-specific ICAM3-grabbing non-integrin (DC-SIGN), langerin, and dendritic cell immunoreceptor (DCIR) belong to the type II group that displays a Ca2+-dependent glycan recognition. On the other hand, Dectin-1, C-type lectin receptor (CLEC)-1, CLEC-2, and DC NK lectin group receptor-1 (DNGR-1, also known as CLEC9A) are type II CLRs whose CRD binds glycans in a Ca2+-independent manner (Fig. 1).

Schematic representation of the structure of glycan-binding proteins (CLRs, siglecs, and galectins) involved in immune cell homeostasis. (a) Most CLRs are glycan-binding receptors that contain one or more carbohydrate recognition domains (CRDs). Selected members (MR, DEC-205, DC-SIGN, MGL, langerin, Dectin-1, and DCIR) that play key roles in innate regulatory programs are shown. (b) Siglecs are grouped into the most distantly interrelated members such as sialoadhesin (Sn; Siglec-1), CD22 (Siglec-2), and myelin-associated glycoprotein (MAG; Siglec-4) and CD33-related siglecs that share sequence similarity. (c) Galectins are subdivided into “prototype” galectins, which contain one carbohydrate recognition domain (CRD) and can form homodimers; “tandem repeat” galectins, which contain two distinct CRDs in tandem connected by a linker of up to 70 amino acids; and the unique “chimera-type” galectin-3, which consists of unusual tandem repeats of proline- and glycine-rich short stretches fused onto the CRD. The most relevant and well-established functions of different lectin families are listed
In the CLR family, a Ca2+-dependent CRD with a Glu-Pro-Asn (EPN) motif confers specificity for mannose- or fucose-containing glycans (Lewisa,b,x,y). These include DC-SIGN, MR, and langerin, whereas a Gln-Pro-Asp (QPD) motif is typical of galactose-specific CRDs (e.g., MGL). In this way, some CLRs bind pathogen-associated molecular patterns (PAMPs), others bind self or altered self ligands, and a number of them can bind both motifs. In either case, myeloid CLRs recognize and deliver antigens to intracellular compartments and trigger inflammatory or tolerogenic signaling programs that tailor adaptive immunity (van Kooyk and Rabinovich 2008).
Evidence demonstrating the role of CLRs in immune tolerance came from experiments in which glycosylated antigens targeted to DEC-205 on DCs induced peripheral CD8+ T-cell tolerance. Similarly, delivery of mannosylated myelin-derived antigens suppressed experimental autoimmune encephalomyelitis (EAE) by interfering with peripheral T-cell reactivity. Mannosylated antigens also stimulate the differentiation of IL-10-producing T regulatory type 1 (Tr1) cells in inflamed intestinal mucosa by SIGNR1+ lamina propria DCs, a mouse homologue of human DC-SIGN. Likewise, commensal flora glycoantigens promote the differentiation of Tr1 cells with gut-homing specialization (Johnson et al. 2013). Also in mice, DCIR binding inhibits DC activation suppressing spontaneous autoimmune sialadenitis. In addition, macrophages and tolerogenic DCs express MGL that binds to GalNAc-decorated CD45 and triggers the phosphatase activity of CD45 which in turn leads to T-cell apoptosis and inhibition of pro-inflammatory cytokines (Sancho and Reis e Sousa 2012; van Kooyk and Rabinovich 2008).
CLRs are also involved in the protective response against bacteria, viruses, helminths, protozoa, and fungi, either by inducing an efficient immune response and clearing of the pathogen or by suppressing an exuberant inflammatory response. As a downside of the latter, many pathogens use CLRs as an immune escape mechanism to subvert immune responses. For example, recognition of HIV by DC-SIGN expressed on DCs facilitates viral entry and induces IL-10 production, impairing T-cell proliferation. Furthermore, HIV gp120 accelerates DC apoptosis, negatively affecting DC maturation and leading to limited pathogen recognition by the immune system (van Kooyk and Rabinovich 2008).
Remarkably, malignant transformation generates abnormally glycosylated antigens, including carcinoembryonic antigen (CEA), which are specifically recognized and internalized by CLRs on DCs. Accordingly, tumor antigens that target MGL or MR induce a protective antitumor response mediated by CD8+ T cells. In contrast, CLRs might be used as an evasion mechanism by tumors as tumor-associated Lewis glycans expressed on CEA or CEA-related cell adhesion molecule-1 (CEACAM-1) can result in impaired DC function and inhibition of antitumor responses.
The subtle differences in glycan recognition and divergent immune responses triggered by CLRs could be explained by their different signaling capacity, as different CLRs possess different cytoplasmic domains. While some Syk-coupled CLRs bear ITAM domains and inhibitory CLRs carry ITIM domains, other CLRs have no clear ITAM or ITIM domains such as MR, DEC-205, and DC-SIGN. Thus, an integrated picture has emerged in which the intensity of the transmitted signal and the combination of CLRs activated by a given stimulus can synergize with, antagonize, or simply modulate signals from other receptors, thereby tuning responses to infection or tissue damage (Geijtenbeek and Gringhuis 2009).
Siglecs
Sialic acid-binding immunoglobulin-like lectins (siglecs ) constitute a family of 15 human and 9 murine cell surface receptors that are primarily expressed by cells of the immune system, with the exception of most T cells (Fig. 1). Structurally, this family has an N-terminal Ig domain with a close sequence homology to Ig V regions (a V-set domain) that binds sialic acid-containing ligands followed by 1–16 Ig domains. Siglecs are conventionally divided into two subsets: those that are conserved structurally between rodents, humans, and other vertebrates and share about 25–30 % identity, which includes sialoadhesin (Sn; Siglec-1), CD22 (Siglec-2), myelin-associated glycoprotein (MAG; Siglec-4), Siglec-15, and the CD33-related siglecs comprised by Siglec-3, Siglec-5, Siglec-6, Siglec-7, Siglec-8, Siglec-9, Siglec-10, Siglec-11, and Siglec-14 in humans and Siglec-E, Siglec-F, Siglec-G, and Siglec-H in mouse, which have high homology to CD33 in their extracellular domains (50–85 % identity) (Fig. 1).
The majority of siglecs are endocytic coreceptors capable of mediating cis and trans interactions with sialylated glycans, whereas their key role is to assist immune cells in sensing self in both innate and adaptive immune responses. Engagement of most siglecs triggers inhibitory signals which maintain immune cell homeostasis and temper inflammation. To this end, siglecs bear ITIM and ITIM-like motifs in their cytoplasmic tails. Thus, it is not surprising that some pathogens and tumors shield themselves with sialic acid to downregulate immune response via siglec-dependent mechanisms (Pillai et al. 2012).
Many siglecs are expressed in a cell type-specific manner. For example, Sn is mainly expressed by macrophages, CD22 and Siglec-10 (Siglec-G in mice) by B cells, Siglec-8 and Siglec-F by eosinophils, and Siglec-5 and Siglec-H are expressed on pDCs. Both CD22 and Siglec-G mediate B-cell tolerance induced by sialylated self-associated antigens, and their absence induces autoimmune glomerulonephritis because of breakdown of B-cell tolerance in mice. Moreover, engagement of CD22 with sialylated IVIGs induces inhibition of BCR signaling and promotion of B-cell apoptosis. Furthermore, Siglec-F modulates eosinophil apoptosis, and allergen-challenged Siglec-F-deficient mice show increased lung eosinophil infiltration. Similarly, Siglec-G selectively controls expansion and survival of B1 cells, and Siglec-H controls the ability of plasmacytoid DCs to generate CD4+ effector T cells and CD4+Foxp3+ Treg cells. Thus, siglecs are sialic acid-binding proteins that modulate signaling pathways in B cells and innate immune cells (Paulson et al. 2012).
Galectins
While CLRs and siglecs are cell surface-associated carbohydrate-binding proteins connected to the cell signaling machinery, galectins are soluble β-galactoside-binding lectins that are present in the nucleus, cytoplasm, and extracellular compartment. These glycan-binding proteins can exert their functions either inside or outside the cells. The molecular architecture of galectins allows their classification into three groups: (a) “prototype” galectins (galectin-1, galectin-2, galectin-5, galectin-7, galectin-10, galectin-11, galectin-13, galectin-14, and galectin-15) which share one carbohydrate recognition domain (CRD) that can dimerize; (b) the “tandem repeat” subgroup (galectin-4, galectin-6, galectin-8, galectin-9, and galectin-12) whose members contain two homologous CRDs in tandem in a single polypeptide chain; and (c) the “chimera-type” lectin, galectin-3, which contains one CRD connected to a non-lectin N-terminal region that is responsible for oligomerization (Fig. 1).
Galectins recognize specific carbohydrate structures decorating cell surface or extracellular matrix glycoproteins. The minimal structure recognized by galectins is the disaccharide N-acetyllactosamine (LacNAc) that is attached to polypeptides either as an N-glycan or an O-glycan branching structure and is usually present in multiple units. Although galectins typically recognize this common ligand, they have different glycan-binding preferences, which could explain differences in their biological activity. To illustrate this concept, galectin-1 preferentially binds to complex N-glycans and nonreducing terminal N-acetyllactosamine, while sialylation of terminal N-acetyllactosamine in α2-6 position abrogates galectin-1 binding. On the other hand, galectin-3 does bind to α2,6-linked sialic acid as a result of internal LacNAc recognition, while galectin-12 binds weakly to lactose but recognizes fucosylated and sialylated structures. On the other hand, galectin-4 can recognize sulfatides, and galectin-10 binds with higher affinity to mannose than to LacNAc saccharides. While some members of the galectin family (such as galectin-1 and galectin-3) have broad tissue distribution, others have a more restricted localization (e.g., galectin-7 is preferentially expressed in stratified epithelia, galectin-12 is abundant in adipose tissue, and galectin-10 is mainly expressed in eosinophils and Treg cells). These glycan-binding proteins are involved in several cellular processes that control the balance between immune cell activation and tolerance such as proliferation, differentiation, trafficking, cytokine secretion, and apoptosis (Rabinovich and Croci 2012).
Galectins play key roles during thymocyte development. Galectin-1, galectin-3, and galectin-9 recognize a set of cell surface glycoprotein receptors in double-negative and double-positive thymocytes, leading to different intracellular signals that modulate positive and negative selection. For example, galectin-1 induces rapid and transient activation of extracellular signal regulated kinase (ERK) in developing thymocytes thus promoting negative selection, whereas it antagonizes positive selection by inhibiting sustained ERK activation. On the other hand, galectin-1 and galectin-10 have emerged as markers of human natural Tregs (nTregs). Targeted inhibition of galectin-1 or galectin-10 in human nTregs reverses their anergic state and counteracts their suppressive function.
Galectins can also modulate T-cell differentiation. Galectin-1, galectin-2, and galectin-9 suppress Th1 and Th17 cytokine production and promote polarization toward Th2 and Treg cell phenotypes. Galectin-9 binds to the Tim-3 inhibitory receptor, which controls Th1 cell fate and contributes to suppression of immune responses. Recent studies showed that myeloid-derived suppressor cells (MDSCs) can control T-cell responses through galectin-9-Tim-3 interactions. Interestingly, T helper subsets express a distinct repertoire of cell surface glycans (particularly differential sialylation of cell surface glycoproteins) which contributes to selectively regulate their viability. In addition, galectin-1 promotes the differentiation of IL-27-producing DCs, which in turn contribute to the generation of IL-10-producing T regulatory type 1 (Tr1) cells. Moreover, this lectin deactivates microglia cells through interaction with CD45 O-glycans, retention of this glycoprotein on the cell surface, and modulation of the phosphatase activity of this glycoprotein (Mendez-Huergo et al. 2014).
Galectins can also modulate the viability of T cells. Extracellularly, galectin-1 binds and cross-links a preferred set of glycosylated receptors and triggers signaling pathways that control apoptosis through caspase-dependent or caspase-independent mechanisms. Intracellularly, this lectin acts by interfering with other apoptotic signaling pathways. Although previous assumptions suggested that galectins had redundant roles for inducing common cell death pathways, evidence indicating that each galectin binds to a specific set of glycoproteins and triggers distinct apoptotic profiles is increasing. In this sense, galectin-1 binds and clusters CD45, CD43, and CD7 to induce apoptosis of immature thymocyte and activated T cells. However, galectin-3 influences T-cell survival through binding to CD45, CD71, and CD29 (Clark and Baum 2012). Also, galectin-1 can sensitize T cells to the canonical Fas-mediated death pathway, whereas intracellular galectin-3 has been reported to interfere with Fas signaling by inhibiting cytochrome c release from the mitochondria. On the other hand, galectin-1 controls B-cell apoptosis through caspase-8-dependent mechanisms as galectin-1-deficient plasma cells are much more susceptible to apoptosis. However, other studies suggested that galectins do not elicit a full apoptotic program, but instead favor the exposure of phosphatidylserine residues without inducing T-cell apoptosis (Rabinovich and Croci 2012).
Through modulation of different immune cell pathways, galectins have emerged as central regulators of inflammatory responses. Galectin-3 acts as a threat-associated molecular pattern that mediates pathogen recognition, phagocytosis, chemoattraction, and cytokine production (Chen et al. 2014; Sato et al. 2009). On the other hand, galectin-4 and galectin-8 are endowed with potent bactericidal activity, and galectin-1 contributes to resolution of inflammation by favoring a macrophage pro-resolving phenotype. Interestingly, expression of galectin-1 is upregulated during first trimester human and mouse pregnancies and plays a pivotal role in maintaining tolerance at the feto-maternal interface. Because of their immunoregulatory activity, galectins have been proposed to control exuberant inflammation during the course of autoimmune responses. Galectin-1, galectin-2, and galectin-9 showed a protective role in autoimmune models of arthritis, hepatitis, encephalomyelitis, uveitis, and inflammatory bowel disease through their ability to dampen Th1 and Th17 pathogenic responses and promoting the expansion of Treg cells. On the other hand, galectin-3 and galectin-4 demonstrated pro-inflammatory activity in Con A-induced hepatitis, diabetes, and EAE. Recently, a unique set of cell surface glycans has been identified through the binding of galectin-4 in local memory CD4+ T cells during intestinal inflammation. Thus, some members of the galectin family can display pro-inflammatory activity, while other members contribute to the resolution of inflammatory diseases (Mendez-Huergo et al. 2014).
Expression of galectins is often dysregulated in the tumor microenvironment and greatly influences tumor immunity by inducing apoptosis of effector T cells, facilitating anergy of CD8+ T cells, promoting the differentiation of tolerogenic DCs, and favoring the expansion of Th2 and FoxP3+ Treg cells in tumors such as melanoma, Hodgkin’s lymphoma, lung carcinoma, and neuroblastoma. However, galectins may also contribute to tumor growth and metastasis through alternative mechanisms including the modulation of cell migration, adhesion, and angiogenesis. Recently, galectin-1 and N-glycans decorating the surface of endothelial cells have been reported as molecular links that connect vascular to immune signaling programs and preserve angiogenesis in tumors refractory to anti-vascular endothelial growth factor (VEGF) treatment (Croci et al. 2014). Thus, galectins have emerged as immunoregulatory and angioregulatory proteins that play their central roles in tumor microenvironments (Rabinovich and Croci 2012).
Conclusions
The field of immunoglycobiology is just beginning to find firm grounding as a field through the emergence of more accessible carbohydrate analytical tools, the generation of mice deficient in components of the glycosylation machinery, and the creation of more rational paradigms to analyze the information encoded by the cellular glycome (Angata et al. 2012; Cummings and Pierce 2014). This brief chapter aims to highlight some of the best characterized areas of glycan-focused immunology and emphasizes the role of carbohydrates as integral components of all immunological pathways. Understanding the information encrypted in glycan structures and decoded by endogenous lectins will aid to the design of more rational therapeutic strategies in immune-related disorders.
References
Angata T, Fujinawa R, Kurimoto A et al (2012) Integrated approach toward the discovery of glyco-biomarkers of inflammation-related diseases. Ann N Y Acad Sci 125:159–169
Baum LG, Crocker PR (2009) Glycoimmunology: ignore at your peril! Immunol Rev 230:5–8
Chen HY, Weng IC, Hong MH, Liu FT (2014) Galectins as bacterial sensors in the host innate response. Curr Opin Microbiol 17C:75–81
Clark MC, Baum LG (2012) T cells modulate glycans on CD43 and CD45 during development and activation, signal regulation, and survival. Ann N Y Acad Sci 1253:58–67
Croci DO, Cerliani JP, Dalotto-Moreno T et al (2014) Glycosylation-dependent lectin-receptor interactions preserve angiogenesis in anti-VEGF refractory tumors. Cell 156:744–758
Cummings RD, Pierce JM (2014) The challenge and promise of glycomics. Chem Biol 21:1–15
Dennis JW, Lau KS, Demetriou M et al (2009) Adaptive regulation at the cell surface by N-glycosylation. Traffic 10:1569–1578
Geijtenbeek TB, Gringhuis SI (2009) Signaling through C-type lectin receptors: shaping immune responses. Nat Rev Immunol 9:465–479
Hart GW (2013) Nutrient regulation of immunity: O-GlcNAcylation regulates stimulus-specific NF-κB-dependent transcription. Sci Signal 6:pe26
Johnson JL, Jones MB, Ryan SO et al (2013) The regulatory power of glycans and their binding partners in immunity. Trends Immunol 34:290–298
Marth JD, Grewal PK (2008) Mammalian glycosylation in immunity. Nat Rev Immunol 8:874–887
Mendez-Huergo S, Maller SM, Farez MF et al (2014) Integration of lectin–glycan recognition systems and immune cell networks in CNS inflammation. Cytokine Growth Factor Rev. pii: S1359-6101(14)00020-3. doi: 10.1016/j.cytogfr.2014.02.003
Paulson JC, Macauley MS, Kawasaki N (2012) Siglecs as sensors of self in innate and adaptive immune responses. Ann N Y Acad Sci 1253:37–48
Pillai S, Netravali IA, Cariappa A et al (2012) Siglecs and immune regulation. Annu Rev Immunol 30:357–392
Rabinovich GA, Croci DO (2012) Regulatory circuits mediated by lectin-glycan interactions in autoimmunity and cancer. Immunity 36:322–335
Sancho D, Reis e Sousa C (2012) Signaling by myeloid C-type lectin receptors in immunity and homeostasis. Annu Rev Immunol 30:491–529
Sato S, St-Pierre C, Bhaumik P et al (2009) Galectins in innate immunity: dual functions of host soluble beta-galactoside-binding lectins as damage-associated molecular patterns and as receptors for pathogen-associated molecular patterns. Immunol Rev 230:172–187
Shan M, Gentile M, Yeiser JR et al (2013) Mucus enhances gut homeostasis and oral tolerance by delivering immunoregulatory signals. Science 342:447–453
van Kooyk Y, Rabinovich GA (2008) Protein-glycan interactions in the control of innate and adaptive immune responses. Nat Immunol 9:593–601
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer Japan
About this entry
Cite this entry
Blidner, A.G., Ilarregui, J.M., Rabinovich, G.A. (2015). Immunoglycobiology: Overview. In: Taniguchi, N., Endo, T., Hart, G., Seeberger, P., Wong, CH. (eds) Glycoscience: Biology and Medicine. Springer, Tokyo. https://doi.org/10.1007/978-4-431-54841-6_132
Download citation
DOI: https://doi.org/10.1007/978-4-431-54841-6_132
Received:
Accepted:
Published:
Publisher Name: Springer, Tokyo
Print ISBN: 978-4-431-54840-9
Online ISBN: 978-4-431-54841-6
eBook Packages: Biomedical and Life SciencesReference Module Biomedical and Life Sciences