
Abstract
Freshwater ecosystems provide ecosystem services essential to human well-being, such as provisioning of water and fishery resources, but they are the most vulnerable to human disturbances and concerns have been raised about their loss of biodiversity. The ancient Lake Biwa is one of valuable Asian freshwater ecosystem because of its high biodiversity and endemism but it has been exposed to various severe anthropogenic environmental changes during the past half century. Although we have great concern about the biodiversity loss in this lake, it remains highly unknown what kind of human disturbances cause deterioration of the ecosystem. Elucidation of environmental pressures for the biodiversity loss is important to illuminate subjects for conservation of biodiversity. We particularly focus on a littoral benthic macroinvertebrate fauna because its habitats, located in the interface between aquatic and terrestrial ecosystems, are those most vulnerable to human activities. We found that littoral benthic macroinvertebrate diversity was largely affected by pH, temperature, phytoplankton biomass, benthic microalgae biomass, and coverage of submerged plants. These environmental variables are largely associated with environmental problems that occurred in the Lake Biwa: eutrophication, warming, and massive submerged plant expansion. This finding suggests that past environmental problems caused serious impacts on the biodiversity of Lake Biwa.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Anthropogenic pressure
- Eutrophication
- Lake Biwa
- Littoral environment
- Submerged vegetation
- Water temperature
- Zoobenthos fauna
Introduction
Lake Biwa dates back approximately 4 million years and is one of the largest representative ancient lakes in Asia. The lake contains a high biodiversity of about 1,700 aquatic species including 61 endemics (Nishino 2012). Since the 1960s, however, the Lake Biwa ecosystem has suffered deterioration of water quality and habitat loss for native aquatic organisms as a result of human activities within its watershed, such as human population increase, land development, and agricultural modernization (Rossiter 2000). Biodiversity loss has become increasingly evident in this lake (Nakanishi and Sekino 1996). For example, the abundance of several common endemic plankton species has substantially decreased in Lake Biwa over the past 40 years (Ichise et al. 2004). A long-term analysis has also demonstrated that eutrophication and increased water temperatures have led to drastic changes in pelagic plankton communities (Hsieh et al. 2010).
However, it remains unknown which types of human disturbances and resultant environmental changes in the lake ecosystem affect the diversity of benthic macroinvertebrates inhabiting littoral waters. Littoral lake habitats serve as the interface between aquatic and terrestrial ecosystems and are thus among those most vulnerable to human disturbances, especially at a catchment scale (Karube et al. 2010). It is crucial to understand the processes and mechanisms causing the loss of diversity of littoral benthic macroinvertebrate fauna during the past half century, as these taxa account for the majority of endemism in Lake Biwa (Nishino and Watanabe 2000). Some long-term monitoring data exist for littoral zoobenthic fauna in this lake, but most data are highly scattered or unpublished. Compilation and analysis of these archival data will help to trace the historical changes in the lake under recent human disturbances and to identify the driving factors of the drastic biodiversity loss in Lake Biwa.
In this study, we first analyzed existing historical data on littoral macroinvertebrate fauna in Lake Biwa in relationship to coastal environmental changes resulting from human disturbances since the 1960s. Using multivariate statistics and ordination methods, we then focused on spatial variation in local littoral macroinvertebrate communities in the current Lake Biwa and examined which environmental characteristics of their microhabitats affect community composition.
Methods
Archival Data
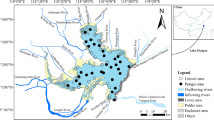
We used archival data on littoral benthic macroinvertebrate fauna in Lake Biwa to examine historical changes in these taxa since the 1960s when environmental alterations began to appear. The archival data were provided by the Shiga Prefectural Fishery Experimental Station. Monitoring surveys were conducted at 55 sites within littoral waters during the summers (July to August) of 1965, 1995, and 2003 (Fig. 8.1). At each site, 15 × 15 cm littoral sediment samples were collected from six depths (1–7 m) using an Ekman-Berge grab sampler, and benthic macroinvertebrates were sieved using a 1-mm-mesh sieve. Samples were then sorted and classified to the species level whenever possible, although some were identified to genus or a higher taxonomic group. Details of these monitoring surveys have been documented as annual reports elsewhere (Shiga Prefectural Fishery Experimental Station 1972, 1998, 2005).

Map of Lake Biwa. Open circles show observation sites in the 2006 survey; closed triangles show sampling sites in the historical monitoring from 1969 to 2003 by Shiga Prefectural Fisheries Institute
Environmental Assessment of Littoral Habitats
To assess the habitat quality for littoral benthic macroinvertebrates in Lake Biwa, we conducted a field survey in July 2006. Benthic macroinvertebrates were collected from 33 sites in littoral waters shallower than 1 m in depth, using a 475-μm-mesh Surber net sampler with a 30 × 30 cm quadrat and three replicate samples (Fig. 8.1). At each sampling site, we also monitored environmental characteristics of the microhabitat in terms of water chemistry, physical habitat structure, food quality and quantity, and predation pressure, to examine how local benthic macroinvertebrate communities are colonized along environmental gradients of the littoral habitats.
For water chemistry, we measured the water temperature, pH, turbidity, and dissolved oxygen concentration using a multi-profiler (U-20; Horiba, Japan). In terms of physical habitat structure, we scored substratum size based on the dominant substrate types (rock = 1, pebble = 2, sand = 3, mud = 4) and coverage of submerged and emergent plants in rank order (no vegetation = 0, <25 % coverage = 1, 25–50 % = 2, 50–75 % = 3, and >75 % = 4).
As an indicator of food quality and quantity, we measured the abundance of particulate organic carbon (POC) and epilithic organic matter (EOM), which serve as the basal food sources for filter-feeding and grazing zoobenthos, respectively (Karube et al. 2010). The abundance of EOM was measured as the chlorophyll a concentration (mg cm−2) of epilithic microalgae, which were scraped from 6 × 6 cm areas on each of five boulders at each site and then mixed to reduce the effect of spatial variation in algal biomass at a microhabitat scale. For POC, littoral water samples were collected from each site and filtered on precombusted (450 °C for 2 h) glass-fiber filters (GF/F, pore size ~0.7 μm; Whatman) after sieving through 150-μm mesh to remove coarse particles. The carbon content of particulate samples (POC) was measured using an elemental analyzer (EA1108; Fisons, Italy; EA1112; Thermo Fisher, Germany) at the Center for Ecological Research, Kyoto University. For details of the sampling methods, see Sakai et al. (2013).
To collect data on the density of littoral fishes, which are considered potential predators of benthic macroinvertebrates, we enclosed a given area of littoral waters at each sampling site with a 1-cm-mesh beach seine net and collected all fishes within the enclosure by towing the net toward the shoreline. Fish density was calculated as the number of fishes per unit area of the enclosure.
Statistical Analysis
We analyzed historical changes in littoral benthic macroinvertebrate communities in Lake Biwa using archival data from 1965, 1995, and 2003 (Shiga Prefectural Fishery Experimental Station 1972, 1998, 2005). We calculated the Shannon–Wiener H’ as an index of littoral zoobenthic diversity for each depth at each site in each of the 3 years, and this value represented α-diversity. Interannual differences in α-diversity were tested using one-way analysis of variance (ANOVA) followed by a Sheffé’s post hoc test. We also tested interannual differences in macroinvertebrate abundance using a likelihood ratio test (LRT) for a generalized linear model (GLM) with negative binomial distribution. We set the significance level of whole pairwise tests at 0.05 by applying a sequential Bonferroni adjustment (Rice 1989). To examine how the community composition of benthic macroinvertebrates varies along environmental gradients of littoral habitat, we performed redundancy analysis (RDA) and Monte Carlo permutation analysis of variance with the “vegan” package of R statistics software (R Development Core Team 2012). For these analyses, data on spatial patterns were processed in relationship to both benthic macroinvertebrate community composition and environmental variables. Before the RDA, community composition data were standardized using the Hellinger method (Legendre and Gallagher 2001).
Results
Long-Term Variation in Littoral Zoobenthic Communities
In total, 25 benthic macroinvertebrate taxa were recorded during the three monitoring surveys between 1969 and 2003 (Table 8.1). Oligochaetes and chironomid larvae always dominated the littoral zoobenthic communities, accounting for 57.9–84.6 % (in abundance) of the entire community, respectively, although chironomid abundance was exceptionally low in 2003. For seven of ten taxa for which the relative abundance was more than 1 %, we observed significant historical changes in their abundance. From 1969 to 1995, some taxa exhibited a significant increase in abundance (Planorbidae spp. and Corbiculidae spp.), whereas the abundance of other species substantially decreased (Radix onchia, Oligochaeta spp., and Gammaridae spp.; 1). In contrast, from 1995 to 2003, two annelid taxa significantly increased in abundance (Oligochaeta spp. and Pharyngobdellida spp.), whereas chironomids and two gastropods (Planorbidae spp. and Lymnaeidae spp.) showed marked decreases in abundance (Table 8.1).
The littoral zoobenthic communities exhibited long-term variation in α-diversity (Fig. 8.2). These long-term patterns varied among monitoring depths (Table 8.2). At monitoring depths of 1 and 2 m, α-diversity tended to steadily decrease as time elapsed. In contrast, the α-diversity of zoobenthic communities at a depth of 3 m significantly increased from 1969 to 1995. In waters deeper than 3 m, no clear historical changes were observed across the three monitoring years.

Historical changes in the biodiversity of the littoral benthos from 1969 to 2003 at each water depth. Error bars show standard deviations
Spatial Variation in Littoral Zoobenthic Communities
In 2006, 25 macroinvertebrate taxa were found in the littoral waters of Lake Biwa. Although Oligochaeta and Chironomidae were among the most dominant within the littoral zoobenthos, community composition exhibited large spatial variation among monitoring sites along environmental gradients of littoral habitats (Tables 8.3, 8.4). According to the RDA, five of ten environmental variables were selected as significant factors explaining a large portion of the spatial variation in littoral zoobenthic community composition (Table 8.5). Two annelid taxa, the most dominant Oligochaeta spp. (Oli) and the common Pharyngobdellidae spp. (Pha), tended to appear at sites with high coverage of submerged vegetation, whereas the second most dominant Chironomidae spp. (Chi) preferred sites with low coverage of submerged vegetation (Fig. 8.3). The filter-feeding Corbicula (Cor) and water pollution-tolerant snails (Ple) were more abundant in eutrophic waters with high POC concentration. In contrast, the abundance of some grazing taxa, such as gammarids (Gam), Ephemeroptera (Eph), and planorbid snails (Cho), was higher in littoral habitats rich in epilithic algae (Fig. 8.3). Temperature and pH were also primary environmental factors affecting zoobenthic communities (Table 8.5, Fig. 8.3).

Ordination diagram with the first two redundancy analysis (RDA) axes for the littoral benthic macroinvertebrate community structures from 2006. Plots show sites; abbreviations indicate 25 taxa defined in Table 8.1. Arrows show effects of environmental variables selected in the RDA variable selection on community structure: COV-SUB, coverage of submerged vegetation; TEMP, water temperature; pH, pH in water column; POC, concentration of suspended particulate organic carbon; EPIPHYTE, chlorophyll a abundance of epiphytes on the littoral bottom
Discussion
Our long-term analysis based on archival data revealed that the community composition and biodiversity of benthic macroinvertebrates substantially varied in the littoral waters of Lake Biwa during the past half century. Particularly in shallower waters, the α-diversity of the zoobenthic communities exhibited significant, steady decreases as time elapsed. Most common taxa showed significant changes in abundance, although the temporal patterns differed greatly among these taxa. Our results support a commonly accepted, but previously untested, notion that biodiversity has declined in Lake Biwa since the 1960s when environmental alterations caused by anthropogenic disturbances became apparent (Rossiter 2000).
Within large basins in Lake Biwa, the littoral benthic macroinvertebrates also exhibited large spatial variation in community composition along environmental gradients of littoral habitat. The RDA revealed that five environmental variables, namely, submerged vegetation coverage, POC, EOM abundance, water temperature, and pH, all of which are considered critical for the habitat quality of littoral zoobenthos, accounted for a large fraction of the spatial variation in community composition. These variables are closely related to environmental problems that have appeared in the lake during the past half century. For example, a massive expansion of submerged vegetation has occurred in the shallow south basin during recent years (Hamabata and Kobayashi 2002). Furthermore, as is common in lakes worldwide, the surface water temperature of Lake Biwa has been gradually increasing as a result of global warming during the past half century (Endoh et al. 1999; Hayami and Fujiwara 1999; Hsieh et al. 2010). Lake eutrophication has increased POC concentrations (as a proxy for phytoplankton biomass; Hsieh et al. 2010) as well as pH (as a proxy for phytoplankton photosynthetic activity; Brönmark and Hansson 2005), but has negatively affected EOM abundance through reduced light availability from increased turbidity (Vadeboncoeur et al. 2001, 2003; Kagatsume 2012).
In Lake Biwa, eutrophication progressed in the 1960s and peaked during the 1970s. However, temporal changes in the distribution of eutrophic and oligotrophic taxa along an eutrophication–oligotrophication gradient (the POC-EPIPHYTE axis in the RDA bi-plots) became unclear between the two earlier monitoring years, 1969 and 1995, when eutrophication occurred. This inconsistency may have resulted from a periodic mismatch between the incidence and monitoring of eutrophication, because reoligotrophication has gradually progressed since the mid-1980s as a result of increasing efforts by local governments to reduce nutrient loading to the Lake Biwa watershed (Hsieh et al. 2010; Kagatsume 2012).
In Lake Biwa, surface water temperature, which is an important environmental determinant of littoral zoobenthic fauna, has increased at a rate of 0.028 °C/year over the past half century, with an accelerating trend since 1990 (Hsieh et al. 2010). However, warm-water taxa, such as chironomids and lymnaeids, have decreased, rather than increased, during this warming period. Assuming a temperature increase rate of 0.028 °C/year, we estimate that lake water temperature increased by approximately 1 °C from 1969 to 2003. This long-term change in water temperature is small relative to the range (6.3 °C) of water temperatures measured in a variety of the current littoral habitats. For the warm-water taxa, long-term changes in their abundance might be attributed to other non-climate factors. However, we must take precautions against ongoing climate changes and better understand the effects of global warming on the population dynamics of littoral benthic macroinvertebrates and thus biodiversity as a whole in the future.
In the south basin of Lake Biwa, submerged macrophytes began to drastically increase in biomass in 1995. This phenomenon is well known as a catastrophic shift or regime shift between two stable states of phytoplankton-dominated turbid waters and macrophyte-dominated clear waters (Scheffer et al. 2001). During this period, two annelid taxa, which tend to dominate submerged vegetation habitats (Table 8.5, Fig. 8.3), markedly increased in abundance, whereas chironomids and lymnaeids, which tend to dominate areas with less submerged vegetation habitat (Table 8.5, Fig. 8.3), significantly decreased in abundance. This consistency between historical changes in abundance associated with the expansion of submerged vegetation and the patterns expected from distributional changes along submerged vegetation gradients in the spatial analysis support that the regime shift to a macrophyte-dominant system has had a large effect on littoral zoobenthic diversity in Lake Biwa. In this ecosystem, the regime shift is believed to have occurred in 1995 and was promoted by severe drought in 1994. The resultant drawdown facilitated macrophyte growth through increased light penetration to the lake bottom in the shallow south basin (Hamabata and Kobayashi 2002; Nishino et al. 2012). The lake water level can also be artificially manipulated by the government to prevent flooding (Nakamura et al. 2012). Ongoing climate changes and corresponding water regime management have strong potential to affect the community composition and diversity of benthic macroinvertebrates in the littoral waters of Lake Biwa.
Using archival data, we demonstrated that the diversity of benthic macroinvertebrates has actually declined within the littoral waters of Lake Biwa during the past half century. In addition, several environmental characteristics closely related to past environmental problems (eutrophication, warming, and regime shift) have strongly affected the composition of the littoral benthic macroinvertebrate community. To conserve biodiversity of littoral zoobenthos in this lake, the anthropogenic drivers causing environmental deterioration in local habitats should be identified, as mitigating environmental stressors is easier and more effective on a catchment scale than on a global scale (e.g., water warming). The most likely cause of freshwater environmental deterioration is human land use within catchments (Agarwal et al. 2005; Light and Marchetti 2007), which in turn affects material inputs to littoral waters through tributary rivers (Sakai et al. 2013) and subsequent trophic flows within littoral benthic communities. Land-use changes may also alter habitat quality for littoral zoobenthos in terms of physicochemical environments (Karube et al. 2010). At present, we have limited knowledge of the underlying mechanisms by which specific land uses alter the environments of littoral habitats and how such environmental alterations can cause community compositional changes. Path analysis is a powerful and promising tool to disentangle the complexity of causal relationships among human-derived forces and pressures, environmental states, resultant impacts on biota, and biological responses within a watershed, including terrestrial, river, and lake ecosystems (Sakai et al. 2013). This future challenge will be useful for the restoration of littoral habitats and the conservation of lake biodiversity.
References
Agarwal DK, Silander J, Gelfand AE, Dewar RE, Mickelson JG (2005) Tropical deforestation in Madagascar: analysis using hierarchical, spatially explicit, Bayesian regression models. Ecol Modell 185:105–131
Brönmark C, Hansson L (2005) The biology of lakes and ponds. Oxford University Press, New York
Endoh S, Yamashita S, Kawakami M, Okumura Y (1999) Recent warming of Lake Biwa water (in Japanese). Jpn J Limnol 60:223–228
Hamabata E, Kobayashi Y (2002) Present status of submerged macrophyte growth in Lake Biwa: recent recovery following a summer decline in the water level. Lake Reserv Res Manag 7:331–338
Hayami Y, Fujiwara T (1999) Recent warming of the deep water in Lake Biwa (in Japanese). Res Ocean 8:197–202
Hsieh CH, Ishikawa K, Sakai Y, Ishikawa T, Ichise S, Yamamoto Y, Kuo TC, Park HD, Yamamura N, Kumagai M (2010) Phytoplankton community reorganization driven by eutrophication and warming in Lake Biwa. Aquat Sci 72:467–483
Ichise S, Morita T, Wakabayashi T, Kusuoka Y, Nishino M (2004) Disappearance and its past distribution of the endemic protista Difflugia biwae Kawamur, 1918 in Lake Biwa, central Japan (in Japanese). Rep Shiga Pref Inst Public Health Environ Sci 39:57–63
Kagatsume T (2012) Water conservation policy of Shiga prefectural government. In: Kawanabe H, Nishino M, Maehata M (eds) Lake Biwa: interactions between nature and people. Springer, New York, pp 423–427
Karube Z, Sakai Y, Takeyama T, Okuda N, Kohzu A, Yoshimizu C, Nagata T, Tayasu I (2010) Carbon and nitrogen stable isotope ratios of macroinvertebrates in the littoral zone of Lake Biwa as indicators of anthropogenic activities in the watershed. Ecol Res 25:847–855
Legendre P, Gallagher E (2001) Ecologically meaningful transformations for ordination of species data. Oecologia (Berl) 129:271–280
Light T, Marchetti MP (2007) Distinguishing between invasions and habitat changes as drivers of diversity loss among California’s freshwater fishes. Conserv Biol 21:434–446
Nakamura M, Ogino Y, Akiyama M, Moriyasu K (2012) Evolving history of Lake Biwa and Yodo River basin management. In: Kawanabe H, Nishino M, Maehata M (eds) Lake Biwa: interactions between nature and people. Springer, New York, pp 371–418
Nakanishi M, Sekino T (1996) Recent drastic changes in Lake Biwa bio-communities, with special attention to exploitation of the littoral zone. GeoJournal 40:63–67
Nishino M (2012) Biodiversity of Lake Biwa. In: Kawanabe H, Nishino M, Maehata M (eds) Lake Biwa: interactions between nature and people. Springer, New York, pp 31–153
Nishino M, Watanabe NC (2000) Evolution and endemism in Lake Biwa, with special reference to its gastropod mollusc fauna. Adv Ecol Res 31:151–180
Nishino M, Azuma Y, Tatumi M, Kaneko Y (2012) Ecological changes in the littoral of Lake Biwa and its adjacent areas. In: Kawanabe H, Nishino M, Maehata M (eds) Lake Biwa: interactions between nature and people. Springer, New York, pp 157–160
R Development Core Team (2012) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/
Rossiter A (2000) Lake Biwa as a topical ancient lake. Adv Ecol Res 31:571–598
Rice R (1989) Analyzing tables of statistical tests. Evolution 43:223–225
Sakai Y, Karube Z, Takeyama T, Kohzu A, Yoshimizu C, Nagata T, Tayasu I, Okuda N (2013) Seasonal and site-specific variability in terrigenous particulate organic carbon concentration in near-shore waters of Lake Biwa, Japan. Limnology 14:167–177
Scheffer M, Carpenter S, Foley J, Folke C, Walker B (2001) Catastrophic shifts in ecosystems. Nature (Lond) 413:591–596
Shiga Prefectural Fishery Experimental Station (1972) Reports of the survey of littoral zone in Lake Biwa in 1969 (in Japanese). Shiga Prefectural Fishery Experimental Station, Shiga
Shiga Prefectural Fishery Experimental Station (1998) Reports of the survey of littoral zone in Lake Biwa in 1995 (in Japanese). Shiga Prefectural Fishery Experimental Station, Shiga
Shiga Prefectural Fishery Experimental Station (2005) Reports of the survey of littoral zone in Lake Biwa in 2002–2003 (in Japanese). Shiga Prefectural Fishery Experimental Station, Shiga
Vadeboncoeur Y, Lodge DM, Carpenter SR (2001) Whole-lake fertilization effects on distribution of primary production between benthic and pelagic habitats. Ecology 82:1065–1077
Vadeboncoeur Y, Jeppesen E, Vander Zanden MJ, Schierup H, Christoffersen K, Lodge DM (2003) From Greenland to green lakes: cultural eutrophication and the loss of benthic pathways in lakes. Limonol Oceanogr 48:1408–1418
Acknowledgments
This study was supported by The Environment Research and Technology Development Fund (S-9) of the Ministry of the Environment, Japan and by the JSPS Grant-in Aid (No.24370010). We greatly appreciate the Shiga Prefectural Fishery Institute for providing monitoring data of the Lake Biwa benthos fauna. The present study was conducted using Cooperative Research Facilities (EA-IRMS) of Center for Ecological Research, Kyoto University.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Japan
About this chapter
Cite this chapter
Shibata, Jy. et al. (2014). Long-Term and Spatial Variation in the Diversity of Littoral Benthic Macroinvertebrate Fauna in Lake Biwa, Japan. In: Nakano, Si., Yahara, T., Nakashizuka, T. (eds) Integrative Observations and Assessments. Ecological Research Monographs(). Springer, Tokyo. https://doi.org/10.1007/978-4-431-54783-9_8
Download citation
DOI: https://doi.org/10.1007/978-4-431-54783-9_8
Published:
Publisher Name: Springer, Tokyo
Print ISBN: 978-4-431-54782-2
Online ISBN: 978-4-431-54783-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)