
Abstract
Biological diversity has long been a source of wonderment and scientific curiosity, but it is now increasingly a source of concern. We attempted to classify, map, and characterize vegetation and its biodiversity content using satellite imagery and field enumeration in the cold desert of the Kargil-Ladakh region located in the Western Himalaya, India. Fair-weather, multi-temporal IRS-P6 LISS III data were subjected to visual interpretation, which estimated rocky barren land as the dominant land cover, followed by vegetation (alpine pasture, alpine scrub, agroforest, and subalpine forest) and snow cover. Asteraceae was reported as the most speciose family, numbering more species than genera. Artemisia emerged as the most speciose genus. Species diversity decreased with increasing altitude, but the maximum was found at transition zones of agroforest and alpine scrub. The highest species similarity was observed between alpine scrub and alpine pastures, indicating pronounced intermixing and interaction. The detailed updated map (at a scale of 1:50,000) with extensive field observation would overcome a long-pending gap in ecological study in the region and may also be used as baseline data for conservation and planning, and the region may serve as a natural ecological laboratory for climate change studies.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
Biodiversity, the variety of life, is distributed heterogeneously across the Earth. The most striking feature of the Earth is the existence of life; and the most striking feature of life is its diversity. This biological diversity has long been a source of wonderment and scientific curiosity, but it is now increasingly a source of concern. Some areas teem with biological variation (e.g., moist tropical forests and coral reefs), whereas others are virtually devoid of life (e.g., deserts and polar region), and others are somewhere in between. Determining why these differences occur has long been a core objective for ecologists and biogeographers. The reasons for this interest are twofold. First, it reflects increased opportunity provided by improvements in available data and analytical tools, the former resulting mostly from extensive collation of existing specimen and species occurrence records, the establishment of dedicated distribution mapping schemes, and the use of remote sensing technology (to measure vegetation and other environmental variables). Second, it reflects concern over the future of biodiversity and the resultant need to determine its current status, to predict its likely response to global environmental change, and to identify the most effective schemes for in situ conservation and sustainable use. Biodiversity changes across environmental gradients such as latitude, altitude, and moisture, and hence inventorying and monitoring of biodiversity, is also done at different organizational levels from genes to ecological systems (landscapes), and at different spatial scales from a few square meters to continents.
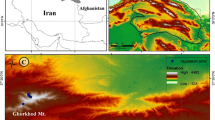
Here, we have attempted to characterize the biodiversity of the Kargil district through in situ enumeration and utilization of geoinformatics techniques, in part of the Ladakh cold desert in Jammu and Kashmir state in India. Kargil is a mountainous region situated in the Himalayas along the northernmost extreme of India. The district covers an area of 17,303 km2 with a population of 1.25 lakhs. It is situated between 30° and 35° N latitude and 75° and 70°E longitude. The district is referred to as a “cold desert” because it combines features of arctic and desert climates, including wide diurnal and seasonal fluctuations in temperature, from −40 °C in winter to +35 °C in summer, and extremely low precipitation, with an annual 10 to 30 cm, primarily from snow (LAHDC 2005). The high altitude and low humidity mean the radiation level is among the highest in the world (up to 6–7 kWh/mm). Soil is thin, sandy, and porous. Because of these peculiar physical and climatic conditions, the Kargil region shows a unique floral assemblage. The whole region is composed of high rocky mountains (Fig. 13.1) and arid desert, snowbound and devoid of natural vegetation. It occupies a unique position because of its high altitude, ranging from 2,500 to 7,000 m above sea level (Fig. 13.2). The region is an almost treeless expanse. The scarcity of precipitation causes plants to be generally found growing along moist river margins or in moist rock crevices. The aspect characteristics of the landscape have a vital role in vegetation growth. Many slopes on the eastern side show a higher degree of vegetation cover than on western slopes (Fig. 13.3). These forests also support the lives of various ecologically important wild animal species such as Aconitum heterophyllum, Uncia uncia, Ovis vignei vignei, Cuon alpinus laniger, Bos grunniens, and Otocolobus manul (Table 13.1).


Image chips and corresponding field photographs

Contours at 500-m interval prepared from SRTM DEM

Slope map shows absence of tablelands except in valleys
Biodiversity Assessment in Himalayan Cold Deserts at Different Physiognomic Gradients
Species diversity and its distribution along the altitudinal gradient has been a subject of ecosystem analysis. Community composition, structure, diversity, and dynamics are related to gradients in obvious ecological factors (e.g., elevation, slope, aspect). Direct gradient analyses have provided many insights into the nature of ecological communities, the factors influencing the distribution of individual species, the adaptive significance of particular growth forms and their contribution to competitive success in different environments, and potential controls on productivity, biomass, and biological diversity (Whittaker 1956, 1960; Peet 1978; Gentry 1982, 1988, 1992). Gradient analyses in alpine and subalpine forest can be particularly challenging, given the large number of species involved, the frequent absence of local taxonomic treatments, and the difficulty of access to the diagnostic parts of many canopy trees and epiphytes. Rahbek (1995) commented that approximately half the studies detected a mid-altitude peak in species richness in a critical literature review on species richness patterns in relationship to altitude. Vetaas and Grytnes (2002) have also reviewed these aspects in the Nepalese Himalaya. Although the plant community of a region is a function of time, nevertheless altitude, latitude, slope, aspect, rainfall, and humidity have roles in the formation of community composition. Very few researchers have explored the region of western Himalaya through biodiversity studies, and most of them were reliant on field surveys. Klimeš (2003) studied life forms and clonality of vascular plants along an altitudinal gradient in the eastern cold deserts of Ladakh and explored special habitats and threatened plants of Ladakh and Kashmir Himalaya, respectively. Geneletti and Dawa (2009) carried out environmental impact assessment of mountain tourism in Ladakh; Fox (1983) studied wildlife conservation and land-use changes in the trans-Himalayan region of Ladakh; and Humbert-Droz and Dawa (2004) studied the biodiversity of Ladakh and suggested a strategic action plan. There have been a few remote sensing-based biodiversity characterization studies in the western Himalaya. Joshi et al. (2006) studied biodiversity characterization in Ladakh, with a focus on the Nubra Valley. Porwal et al. (2003) carried out stratification and mapping of Ephedra gerardiana Wall. in the cold deserts of Himachal Pradesh. Matin et al. (2012) demonstrated a method to integrate the fauna component with existing flora data for complete diversity analysis, utilizing the GPS-based location information on Medicago sativa and Plantago annua to simulate their potential distribution in the year 2020 (SRES A1B scenario, IPCC) using the Maxent model in part of Ladakh Himalaya.
Mountain Alpine Ecosystems and Kargil Ladakh
Alpine ecosystems occur at a range of air density, water availability, and seasonality in the treeless high terrain of mountains worldwide. Mountainous regions have a diversity of landforms, topography, climate, soils, slope exposure, geology, and other abiotic factors. Each combination of these factors affect site temperature and moisture status. Because plant distributions are influenced primarily by temperature and moisture (as controlled by their ecological amplitude), any significant change in abiotic factors causes a change in plant composition. Composition varies along geographic scales: boreal dwarf-shrub heaths, temperate sedge heaths, subtropical dwarf shrubs and tussock grasslands, and tropical giant forest lands. Along local topographic gradients plant cover changes from windswept dwarf-shrub heath, to dense grass-sedge heath, to snow bank vegetation. These cold and relatively little exploited alpine ecosystems, nonetheless, are among those where climate warming impacts are forecast to be pronounced and detectable early on. The variability of alpine environments is predominantly dependent upon altitude (air density), water availability, and seasonality, which are specific to each mountain region. These factors determine, besides the available flora and fauna, the altitudinal zonation and the structure and functioning of the ecosystems. In Kargil, three vegetation belts are observed. The first belt begins at the valley foothills and climbs a few meters in which introduced agroforest is associated with planted tree species of Populas and Salix. The second belt of scrubland ranging between lower foothills and the medium slopes is mostly associated with the medicinally important species of Hippophae rhamnoides, Hordeum vulgare, Juglans regia, and Brassica napus. The third belt appears after 3,500 m, where many alpine grasses grow. The climatic conditions of the upper part of the Kargil mountains resemble the cold-arid climate prevailing in the Southern Hemisphere. For this reason, most of the species belongs to the grass family (Poaceae) with species such as Poa annua, Saussurea ceratocarpa, and Melica persica.
Ecological Gradient
The terrain being hilly and sloped, the available land for agriculture is meager. Many valleys are dotted with formations of alluvial fans, marsh meadows, boulder-strewn riverbeds, and small sandy meanders. Remaining areas, which are not inhibited by humans, are characterized by seasonal to perennial vegetation. These areas are either in the narrow valleys or at higher altitudes, but altogether represent very scanty vegetation. The altitude increases toward the south. The alpine and subalpine flora of the high-altitude desert region of Ladakh, Jammu, and Kashmir has the greatest affinity toward the Central Asiatic region (Dhar and Kachroo 1983). More than 20 threatened species have been identified in the region (Table 13.1), of great medicinal and ecological importance (Anonymous 2011). The vegetation of these zones is characterized by scattered low bushes, with extensive coverage of tussock grasslands, herbaceous formations, sedge meadows, and stony deserts with cushion-like growing forms. Three major vegetation types have been reported in the region. (I) Subalpine forests have been found in very few areas in Kargil region. The altitude range for these areas is 3,000 to 4,000 m, and domination is mainly Pinus roxburghii. (II) Alpine scrub occurs mostly in drier areas and can also be found in areas near river streams. The major vegetation association is Hippophae and Myricaria scrub. (III) Alpine pasture is mainly moist alpine grasslands or pastures occurring at higher altitudes and moister areas near the snow line. Poa annua, Medicago sativa, and Plantago lanceolata are the dominant species, forming the major land cover of the region. (IV) Agroforest comprises clusters of Populas and Salix plantations in the areas near rivers and streams. These species are mostly planted adjacent to the agriculture fields by local people and the forest department. The presence of the three natural vegetation types, that is, moist alpine forest, scrubland, and pastureland, almost corresponds to the dominance of the three life forms, trees, shrubs, and herbs, in altitudinal gradients that offer prospects for various ecological analyses. Because the landscape has scanty anthropogenic influence, which is restricted to valleys, the majority of the landscape is suitable for basic spatial analysis of patch dynamics studies. The complex physiognomy leading to varied microclimates provides a basis to analyze the vegetation distribution and other structural and functional behavior. The influence of snow is the major dominant factor triggering other ecological processes that need to be understood as a potential linkage to biodiversity characterization. In the changing climatic scenario, while there is much hue and cry about glacier melting, increase in tropical areas, vegetation shifts, species tolerance and adaptation, etc., the region offers a perfect laboratory for ecological studies and global change research.
Vegetation Versus Elevation and Seasonality
Exposure is one of the key factors responsible for biological richness in mountainous areas. There is a considerable difference in the distribution of the vegetation communities between the north-facing and south-facing slopes of the mountains, which is related to both solar radiation and precipitation. As a general rule, the south-facing slopes of the mountains receive more sun radiation than do the northern slopes, and that is why the density (number of species) is greater in the southern aspect. North-facing slopes of the mountains receive a lesser amount of sun time, and hence snow cover time increases accordingly, resulting in less ground coverage of vegetation. On the southern slopes of the Kargil, sun radiation intensity is higher on the slopes facing south than the other slopes. For that reason, high occurrence of dry scrub is common on the south-facing slopes. For example, spiny species of Hippophae are found mostly in south-facing plots.
With the increase of altitude, temperature, relative humidity, and air pressure decrease continuously, but solar radiation intensifies. Depending on these circumstances, the temperature of southern slopes is a few degrees higher than the northern slopes. In Kargil, the vegetation and land cover show drastic changes with increasing altitude. At lower altitudes, mainly along the valleys, human-induced vegetation and land-use classes such as agroforestry and settlements show considerable spread in the region. As altitude increases, however, the distribution of agroforests decreases, taken over by moist alpine pastures and moist alpine scrub. The landscape is dominated by barren land and snow. The distribution of snow starts at around 4,500 m and continues to the highest altitudinal point in the region. Mountains with an alpine zone occur at all latitudes, from the wet tropics to the polar regions. Apart from a steady decrease of temperature with increasing elevation, at an average rate of 0.60 °C⁄100 m, air pressure also decreases (Nagy and Grabherr 2009). The latter becomes particularly relevant in mountains such as the Himalayas, where the highest peaks reach more than 8,000 m. Low CO2 might be one of the causes for the absence of diversity in families from the high ground or their generally low diversity compared to the lowlands (Nagy and Grabherr 2009). In contrast, low carbon dioxide pressure seems to have no limiting effect on plants: other factors such as low temperature set the limits (Korne 2003). The macroclimate of the life zone to which a mountain region belongs determines the climatic conditions in its alpine zone. Species that are sensitive to frost require permanent snow protection and as a result have developed remarkable snow tolerance. For example, various species of Ranunculus in alpine areas are known to be able to survive up to 33 months permanently under snow (Moser et al. 1977). This freezing tolerance may be related to adaptation to low temperatures (Sømme 1997).
Landform Characteristics Versus Flora
The vegetation cover in mountain areas is of immense importance as it affects local and regional climate and reduces erosion. Understanding the distribution and patterns of vegetation distribution along with the local influencing factors is important, and these factors have been studied by many researchers (Turner and Gardner 1992; Henebry 1993). Topography is the principal controlling factor in vegetation growth; climatological factors play a secondary role. Elevation, aspect, and slope are the three main topographic factors that control the distribution and patterns of vegetation in mountain areas. Among these three factors, elevation is the most important (Day and Monk 1974). Elevation (Fig. 13.2), along with slope (Fig. 13.3) and aspect (Fig. 13.4), in many respects determines the microclimate and thus the large-scale spatial distribution and patterns of vegetation (Day and Monk 1974; Marks and Harcombe 1981). Several observations can be mentioned regarding the effects of elevation and aspect on vegetation growth in mountainous areas. Vegetation distribution in the region exhibits a vertical gradient from the climatic changes with elevation. Second, vegetation growth in such regions can be significantly affected by aspect, where most occurs in the northern aspect (Fig. 13.4), but the highest diversity and equitability are observed in the southwestern aspect. The impact of aspect on the vegetation growth is most significant in the vertical zone of 3,500 to 6,000 m. The best vegetation in this zone is distributed between eastern and northeastern aspects. In other words, the best vegetation growth is on the shady side of the mountain, where much less evapotranspiration (ET) is expected. The reduced ET on the shaded side is important for vegetation growth in the Kargil region because it is located in a cold desert region. It is also observed that better vegetation growth occurs over a larger elevation range on the side facing southwest and south. The much wider vertical zone with better vegetation growth on the shady side may significantly affect the local water cycle and climate.

Aspect map revealed that North and South-East aspects occupy maximum and minimum areas respectively
Community Analysis and Alpine Ecosystem
The plant community data imply knowledge of structure and composition of the component species. The biophysical components of a plant’s environment interact to form a temperature and moisture regime based primarily on gradients of elevation, slope, and aspect. These components can either balance or offset each other: an example is the situation wherein a species switches aspect with changes in elevation. A plant species whose distribution switches from one aspect to another may seem unusual, but it really is not, because both situations may provide similar temperatures and moisture. Some ecologists have referred to this concept as an effective environment because it demonstrates that one set of site factors can be ecologically equivalent to another. Differing combinations of site factors can have a similar influence on an ecosystem consequent to the ecological principle of compensating effects (Allen and Hoekstra 1992). Community analysis was carried out to understand the floristic and vegetation pattern in the region. To carry out ground-level phytosociological studies, a sampling intensity of 0.002 % of forest area and 0.001 % for scrub/grasslands was envisaged following the species–area curve principle. Thus, 154 sample plots proportionally covered 30, 44, and 80 plots for agroforest, moist alpine scrub, and moist alpine pastures, respectively. In each plot, names of trees (GBH and height measurement), herbs, shrubs, climbers, seedlings, and saplings were recorded for all species. A phytosociological database was created, and the basic structural parameters were computed, including relative frequency, relative dominance, and relative density.
Spatial heterogeneity of the environment increases the number of different habitats, permitting a greater number of different resource use strategies, preventing competition equilibrium and exclusion, and hence may result in higher diversity. The scale of this heterogeneity is different for different organisms. Disturbance is also widely regarded as one of the major factors for species diversity (Connell 1978; Noss 1996). The disturbance may result from forest fragmentation, fires, grazing, trampling, or windblow. From different disturbance factors, larger areas of habitat may transform into a series of smaller ones, each of which will support fewer selected species. Therefore, the natural heterogeneity coupled with spatially distinct disturbance regimes brings complexity in habitats, which leads to the formation of different species assemblages. Biodiversity assessment at the landscape level requires a phytosociological database on different aspects such as species type, species richness, diversity, abundance, and basal cover. The assessment of variability of such a wide range of parameters and fixing sample size for large heterogeneous habitats is difficult. A certain sample size could be fixed and accuracy checked against samples size after the sampling exercise. This method provides an open-ended design whereby sample size could be enhanced, and the exercise can be further continued to determine accuracies within the framework of time and cost limits. In distribution of sample size in studies conducted throughout the world, the plot sizes of 0.04 ha (20 × 20 m size within a vegetation type depends on area of each sample unit), 0.1 ha (32 × 32 m), and 1 ha as belt transect or square plots could be considered. To study biodiversity at stratification based on remotely sensed data, 0.01 ha as plot size for diverse evergreen forest types, 0.04 ha for deciduous forests, belt transects for fragmented habitats, and a modified Whittaker’s plot for shrubs/scrub/grasslands/sparse trees are preferred. The random distribution of small size plots (0.04/0.01 ha) across various habitats will be helpful in statistically analyzing the diversity patterns across given vegetation types. Alternatively, a selected number of large-size plots of 1 ha across major microhabitats of given vegetation types will be helpful in understanding the role of microlevel topographic and biotic factors in species diversity and richness.
According to the discrete community theory, plant communities have distinct boundaries within which species composition is homogeneous, and the transition between them is relatively rapid. As such, the number of species would be underestimated when extrapolated from a small-scale species–area curve to a larger scale because of the occurrence of new species across community boundaries (Wilson and Chiarucci 2000). Alpha diversity can be adequately described by the coefficient c, which itself can be accurately predicted from the total number of species; and the coefficient z can be used to predict the number of species per individual and vice versa. Thus, these two coefficients are strong and complementary measures of species diversity. The higher the z, greater would be the impact of further harvest of individuals; in other words, the areas with high z would be more susceptible to selective or random felling of trees even if there is no contraction of area.
The sample plots of 0.04 ha/0.1 ha were randomly distributed across each stratum. The sample plot was reached on the ground based on GPS locations and also important ground bearings available from the survey of India (SOI) map. As the location was reached, the sample plot of desired dimension was laid. For sampling shrub species, two plots of 5 × 5 m in two opposite corners were laid. For herbaceous plants, five plots of 1 × 1 m (four in corners and one in center) were laid. A nested sampling design was followed within each sample point to record shrubs, herbs, climbers, epiphytes, and lianas. At each sample point, the circumference at breast height (CBH) of all tree species was recorded and marked with chalk to avoid duplication. To ascertain that measurements were taken at proper height, a 1.37-m stick was used. For shrubs, the CBH was measured about 30 cm above ground. The total number of seedling of various species was counted, and average girth of each species was recorded. For shrubs, total number of tillers for each species was counted and for each species an average circumference at ground height level was estimated. For herbaceous layer or ground flora, the nested quadrat method with 1 × 1 m plot size was taken in two (north and south) corners. Only the number of individuals of each life form was recorded. Based on spatial variability, a belt transect or a square plot of large size was laid on a case-by-case basis. The transect or square plot was gridded at 20 × 20 m, and all the information on trees, shrubs, herbs, climbers, and lianas was recorded independently for each subgrid. This method facilitated understanding the impact of microlevel spatial gradients, structure, and diversity patterns.
Biological diversity can be quantified in many ways. The two main factors taken into account when measuring diversity are richness, the number of species, and evenness, the relative abundance of individuals among the species. Species richness is described as the number of species present in a sample or habitat per unit area. The simplest species richness index is based on the total number of species and the total number of individuals in a given sample or habitat; it indicates that higher the value the greater is the species richness. Maximum diversity is observed in agroforest, based on the Shannon diversity index (1948), as 1.82, followed by moist alpine scrubs at 1.51 and moist alpine pastures with the value 0.62 (Fig. 13.5a). Equitability and dominance have been observed at different aspects and correlated with Shannon diversity. Although Pielou equitability (1969) depends upon the Shannon diversity index (J′), the J′ found was high in South-West aspect (SW), followed by Eastern (E) and North-Eastern aspect (NE) (Fig. 13.5b). The Simpson index (1948) is the most reliable and easy way to understand dominance in species occurrence. Its maximum value goes up to one, observed in case of single species. Each individual represents a small fraction of the total value. For Kargil, the maximum dominance was observed in Lamium album for agroforest (0.080), Crepis multicaulis (0.09) in scrubland, and Saussurea ceratocarpa (0.4) in pastureland.

Shannon diversity index for different vegetation classes (a) and Shannon diversity, Simpson dominance, and Pielou equitability at different aspects (b). Maximum diversity and equitability were observed at the eastern aspect and maximum dominance at the northern aspect. (c) Area estimates of forest vegetation and land cover map of Kargil. (d) Life form distribution pattern in three vegetation cover types as per field sampling
Vegetation Type Mapping and Geoinformatics Technology
Classification is a relevant procedure in studying the vegetation mosaic and the interaction between them and the landscape. The information contained in vegetation types can be studied using community analysis and other ecological measurements. Development of simple, user-friendly vegetation classification based on ecological principles is a prerequisite for vegetation type mapping. Vegetation in general and (forest in particular) is an extremely complex system (Anonymous 2008), and the orderly arrangement of vegetation comes under classification, which is nothing but a process of sorting out similar classes into a few logical classes. Here the scheme primarily considers physiognomy, structure (vertical stratification, height, and spacing), phenology (green and brown wavelength), function or life forms, flora, climate, topography, geomorphology, soil, and biotic factors for classifying vegetation entities. Mapping vegetation through remotely sensed images involves various considerations, processes, and techniques. The increasing availability of remotely sensed images from the rapid advances in remote sensing technology expands the horizon of our choices of imagery sources. Various sources of imagery are known for their differences in spectral, spatial, radioactive, and temporal characteristics and thus are suitable for different purposes of vegetation mapping. Generally, it is necessary to first develop a vegetation classification for classifying and mapping vegetation cover from remotely sensed images at either a community level or species level. Then, correlations of the vegetation types (communities or species) within this classification system with discernible spectral characteristics of remotely sensed imagery are established. These spectral classes of the imagery are finally translated into the vegetation types in the process of image interpretation.
Remote sensing is the science of acquiring information about the Earth’s surface without actually being in direct contact with it (Lillesand et al. 2007). It began with aerial photography, but technological development has enabled the acquisition of information at other wavelengths including near-infrared (NIR), thermal infrared (TIR), and microwave regions of the spectra, known as spectral bands. Vegetation has a unique spectral signature that enables it to be distinguished readily from other types of land cover in an optical/near-infrared image. Ideally, reflectance is low in visible spectra (blue and green regions) as a result of absorption by chlorophyll, and a sudden increase is called the red edge. The red edge refers to the region of rapid change in reflectance of chlorophyll in the near-infrared range, which strongly reflects at wavelengths greater than 700 nm. The change can be from 5 % to 50 % reflectance between 680 to 730 nm. In the near-infrared (NIR) region, the reflectance is much higher than that in the visible band because of cellular structure in leaves. Hence, vegetation can be identified by high NIR but generally low visible light reflectance. Of four vegetation classes in Kargil (Table 13.2), only agroforest and Chir/Pine forest consists of tree species that have compact intercellular structures and lesser amounts of leaf chlorophyll content; hence, the reflectance is medium in the NIR band and less in green and red bands. Shortwave infrared (SWIR), which is a water absorption band, shows higher reflectance only for scrubs and much less for agroforest. Scrub, which mostly consists of species such as Bromus japonicas, Hyppophae rhamnoides, and Myricari germanica, exhibits maximum reflection in the SWIR band because of the lesser water content. Species such as Poa annua, Phleum alpinum, and Artemisia brevifolia have a huge amount of fresh chlorophyll and water in their leaves and hence show higher reflectance in the red region and less in the NIR region because of the compact intercellular arrangements of leaves.
The vegetation and land-cover map clearly depicts the distribution pattern of the region (Fig. 13.6). It is always difficult to find good cloud-free data for interpretation. However, we found satellite images of October 2005, which was absolutely cloud free, and hence these images were used in this study. According to mapping, the largest area is covered by barren land (44.03 %), followed by vegetation, that is, 38.59 % (Fig. 13.5c). The vegetation cover represents distinct strata, with moist alpine pastures covering an area of 30.02 % (5,194.5 km2), followed by moist alpine scrub covering 7.43 % (1,285.54 km2), agroforests 1.1 % (190.55 km2), and subalpine forests 0.04 % (6.24 km2). Subalpine forest showed a few patches along the boundary of the Baramullah district distributed between 2,700 and 3,800 m altitude with Pinus wallichiana and Juniperus semiglobosa as the dominant species. Champion and Seth (1968) put this class under subalpine forest class under section 14 (Table 13.2). Agroforests are found distributed along river streams and settlements between 2,500 and 3,600 m altitude. They showed flourishing growth of Populus and Salix species, along with Poa annua, Medicago sativa, Phleum alpinium, Bromus japonicus, Digitaria stewertiana, etc. The areas were well differentiated on the satellite data because of a peculiar bright red tone. Moist alpine scrub was found in the altitudinal range of 2,800 to 5,100 m. At some places, Hippophae and Myricaria formations were observed. Aconitum heterophyllum, Aconitum violaceum, Anaphalis brusus, Agrostis gigantea, Alisma graminium, etc., were dominantly distributed. On satellite imagery, moist alpine scrub reflected a light to bright red tone with smooth texture (Table 13.2). Champion and Seth (1968) put this class under moist alpine scrub under section 16/C1. Moist alpine pastures were found distributed over 5,194.5 km2 occurring between 2,800 and 5800 m altitude, reaching close to the snow cover at places. Similarly, moist alpine scrub reflected light red to light brown color on the satellite imagery, often causing considerable ambiguity to delineate between scrub and pastureland. Scrublands were dominated by Poa annua, Medicago sativa, Artemisia brevifolia, Plantago lanceolata, and Poa pratensis. Champion and Seth (1968) put this class under moist alpine pasture category under section 15/C3. The present study with medium-resolution satellite data has characterized vegetation/land-cover classes in the Kargil region with reasonably good accuracy. Being a cold desert region, the vegetation and land-cover map showed dominance of barren land and snow cover with lesser areas under vegetation cover. The overall classification accuracy for vegetation was observed to be 93 %, which is acceptable for further landscape analysis including extrapolation of biodiversity attributes. The reason for the 15 % inaccuracy could be delineation between similar classes at the transition that is apparent because of fuzziness. A large area of the landscape is occupied by barren land (44 %), which is found to be dominant over vegetation (39 %) and snow cover (17 %) (Fig. 13.5c).

Vegetation/land-cover map derived from IRS-P6, LISS-III satellite data
Conclusion
The vegetation type map has been prepared using four individual scenes of georeferenced IRS P6 LISS III satellite data and validated with ground visits using exact GPS locations. Various enhancement techniques such as standard deviational and contrast stretching, available in ERDAS Imagine software, were used to make data more interpretable. A hybrid approach considering NDVI classified images and false color composites (FCC) of satellite data was implemented for visual interpretation. Visual interpretation involved classification of the satellite data under various vegetation and land-cover categories with the help of interpretation keys to prepare a final vegetation and land-use map of the study area. Field data collected during two field trips were used to calculate primary vegetation analyses to understand phytosociological status in the region such as Shannon diversity (1948), Pielou equitability (1969), and Simpson dominance index (1949). The dispersed landscape of Kargil includes only one vegetation class under tree species, although a few small patches of subalpine forest (Chir/Pine) were also mapped (0.04 %) in the southwestern part. About 30 % of the geographic area is mapped as moist alpine pasture followed by moist alpine scrub, agroforest, and other land-cover classes. The ecosystem diversity ranges from subalpine forest to moist alpine pastures. The agroforest covers only 1.1 % of the total land area of the district, found mostly along the rivers in the foothills of valleys; at lower slopes moist species of scrub are prominent. At top altitudinal range, alpine meadow (as moist alpine pasture) was mapped. With the help of fusion of conventional as well as remote sensing techniques, vegetation characterization in hilly areas can be done easily, accurately, and effectively. Satellite data of the optimal vegetative period are very important because of the seasonal growing nature of many pasture and scrub species and also the lesser period of ground coverage by snow. NDVI-, VI-, and NDWI-based classification integrated with visual interpretation helps to identify and differentiate vegetation from nonvegetation classes in difficult terrain, where vegetation is either dispersed or sensor reflectance is much less.
Three hundred and twenty four species were encountered during two field visits in Kargil. Of these, only 16 are tree species. About half (157) of the total species found are economically important (Annex: Economically important species list, Anonymous 2011) in many ways, and 81 species known to be of medicinal importance (Annex: List of medicinal plants, Anonymous 2011). A total of 226, 189, and 79 species were found in agroforest, moist alpine scrub, and moist alpine pasture, respectively. Family diversity was maximum in agroforest, with 46 species, followed by 36 in moist alpine scrub and 20 in moist alpine pasture. Study of genera of all species has revealed that 137 genera have been recorded in agroforest, 108 in moist alpine scrub, and only 46 in moist alpine pasture (Fig. 13.5d). Various species of Hippophae (H. rhamnoides), locally called sea buckthorn, has a higher medicinal importance with highest total importance value. The diversity status in the region can be understand more broadly and precisely if more plotting can be done at vegetative areas of high altitude and a majority of steeper slopes (Fig. 13.7).

Biodiversity value image
Physiognomy plays a vital role in shaping the vegetation pattern in the district. Species diversity showed decreasing pattern with increasing altitude, similar to the eastern Himalaya. Trees and larger shrubs are generally rare in this region. Plantation (such as agroforest) of economically important tree species along the roads and rivers and within the arable land by the forest department has enhanced ecological stability in the region. However, pasturelands form the upper altitudinal limit of vegetation distribution, varying between 3,000 and 5,000 m under different environmental conditions. Agroforest in the mountains are a treasure trove; apart from fulfilling primary needs, they also have an important role in ecological processes such as soil conservation and nitrogen fixation, and also as windbreaks or shelterbelts to protect agricultural crops from damage caused by strong winds. Tree species such as Populas and Salix have also provided a livelihood to the local villagers. Dependence on these species for fuel, fodder, timber, and many other passive uses has increased the agricultural practices of these agroforests in the past few decades. The ability of these tree species to survive in a great range of temperature (−45 to 46 °C) and many other adverse conditions helps these to acquire native community structure in the region with the observed highest diversity value among all vegetation types. The occurrence of a large number of plant species at low altitude might result from mild climatic conditions and diverse habitats. As has been discussed, the aspect of the mountain slopes significantly affects the vegetation growth in the study area. The vegetation coverage on the sunny side in the mountain area is less developed than that on the shady side because of greater evapotranspiration and differences in their solar radiation and higher land surface temperature. The vegetation type land-cover map generated at a spatial scale of 1:50,000 integrated with ground sampling points can have a crucial role in understanding vegetation patterns and ecology at varying levels and scales.
Biodiversity and Climate Change
Climate change is a major cause of biodiversity loss (CBD 2011). Past changes in the global climate resulted in major shifts in species ranges and marked reorganization of biological communities, landscapes, and biomes. The present global biota was affected by fluctuating concentrations in the Pleistocene (the past 1.8 million years) of atmospheric carbon dioxide, temperature, and precipitation (CBD 2011), and coped through evolutionary changes, species plasticity, range movements, or the ability to survive in small patches of favorable habitat (refugia). Biodiversity and climate change are interconnected, not only through climate change effects on biodiversity, but also through changes in biodiversity that can affect climate change. Observed changes in climate have already adversely affected biodiversity at the species and ecosystem levels, and further deteriorations in biodiversity are inevitable with further changes in climate (Malhi et al. 2010). Such changes, which have resulted in major shifts in species ranges and marked reorganization of biological communities, landscapes, and biomes, occurred in a landscape that was not as fragmented as it is today and with little or no pressure from human activities in places such as Kargil. Changes in climate during the past few decades of the twentieth century have already affected biodiversity. The observed changes in the climate system (e.g., increased atmospheric concentrations of carbon dioxide, increased land and ocean temperatures, changes in precipitation, and sea level rise), particularly the warmer regional temperatures, have affected the timing of reproduction of animals and plants and migration of animals, the length of the growing season, species distributions and population sizes, and the frequency of pest and disease outbreaks in all kinds of living organisms.
References
Allen TFH, Hoekstra TW (1992) Towards a unified ecology. Columbia University Press, New York
Anonymous (2008) Biodiversity characterisation at landscape level using remote sensing & geographic information system. National Remote Sensing Centre (NRSC), DOS/DBT, Government of India, Balanagar, Hyderabad
Anonymous (2011) Biodiversity characterisation at landscape level for Kargil district using remote sensing & GIS techniques. IIT-CORAL/SAML/PR-1/(LLS-2011)
CBD (2011) Convention on biodiversity, Canada (http://www.cbd.int/)
Champion HG, Seth SK (1968) A revised survey of the forest types of India. Govt. of India Press Manager of Publications (Delhi)
Connell JH (1978) Diversity of tropical rainforest and coral reefs. Science 199:1304–1310
Daubenmire R, Daubenmire JB (1968) Forest vegetation of Eastern Washington and Northern Idaho. Washington State University Extension, Pullman (1968 as XT0060). Reprinted 2002. MISC0249
Day F, Monk CD (1974) Vegetation patterns on a southern Appalachian watershed. Ecology 55:1064–1074
Dhar U, Kachroo P (1983) The alpine flora of Kashmir. Scientific Publishers, Jodhpur
Fox JL (1983) Constraints on winter habitat selection by the mountain goat (Oreamnos americanus) in Alaska. Ph.D. Thesis, University of Washington, Seattle
Geneletti D, Dawa D (2009) Environmental impact assessment of mountain tourism in developing regions: a study in Ladakh, Indian Himalaya. Environ Impact Assess Rev 29:229–242
Gentry AH (1982) Patterns of neotropical plant species diversity. Evol Biol 15:1–84
Gentry AH (1988) Changes in plant community diversity and floristic composition on environmental and geographical gradients. Ann Mo Bot Gard 75:1–34
Gentry AH (1992) Tropical forest biodiversity: distributional patterns and their conservational significance. Oikos 63:19–28
Henebry GM (1993) Detecting change in grasslands using measures of spatial dependence with Landsat TM data. Remote Sens Environ 46:223–234
Heywood VH (1995) Global biodiversity assessment. Cambridge University Press, New York
Humbert-Droz B, Dawa S (2004) Biodiversity of Ladakh. Strategy and action plan. LEDEG, Sampark, Kolkata
Joshi PK, Rawat GS, Padilya H, Roy PS (2006) Biodiversity characterization in Nubra valley, Ladakh with special reference to plant resource conservation and bioprospecting biodiversity and conservation. Biodivers Conserv 15:4253–4270
Klimeš L (2003) Life-forms and clonality of vascular plants along an altitudinal gradient in Eastern Ladakh (NW Himalayas). Basic Appl Ecol 4(4):317–328
Korne C (2003) Carbon limitation in trees. J Ecol 91:4–17
LAHDC (2005) Ladakh autonomous hill council, Leh. http://leh.nic.in/default.htm
Lillesand TM, Kiefer RW, Chipman JW (2007) Remote sensing and image interpretation, 5th edn. Wiley, Singapore
Malhi Y, Silman M, Salinas N, Bush M, Meir P, Saatchi S (2010) Introduction: elevation gradients in the tropics: laboratories for ecosystem ecology and global change research. Global Change Biol 16:3171–3175
Marks PL, Harcombe PA (1981) Vegetation of the Big Thicket National Preserve, Texas. Ecol Monogr 51:287–305
Matin S, Chitale VS, Behera MD, Mishra B, Roy PS (2012) Fauna data integration and species distribution modelling as two major advantages of geoinformatics based phytobiodiversity study in today’s fast changing climate. Biodivers Conserv 21:1229–1250
Moser W, Brzoska W, Zachhuber K, Larcher W (1977) Ergebnisse des IBP-Projekts Hoher Nebelkogel 3184 m. Sitzungsber Österr Akad Wiss Math-naturwiss Klasse Abt I 186:387–419
Myers N, Mittermeier RA, Mittermeier CG, Da-Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature (Lond) 403:853–858
Nagy L, Grabherr G (2009) The biology of alpine habitats. Oxford University Press, Oxford
Noss RF (1996) Ecosystems as conservation targets. Trends Ecol Evol 11:351
Paczoski J (1928) Biologiczna struktura lasu (in Polish: the biological structure of forest). Sylwan 3:193–221
Peet RK (1978) Forest vegetation of the Colorado Front Range: patterns of species diversity. J Veg 37:65–78
Pielou EC (1969) An introduction to mathematical ecology. Wiley, New York
Porwal MC, Sharma L, Roy PS (2003) Stratification and mapping of Ephedra gerardiana Wall. in Poh (Lahul and Spiti) using remote sensing and GIS. Curr Sci 84:208–212
Rahbek C (1995) The elevational gradient in species richness: a uniform pattern? Ecography 18:200–205
Shannon CE (1948) A mathematical theory of communication. Bell Syst Tech J 27:379–423
Simpson EH (1949) Measurement of diversity. Nature (Lond) 163:688
Singh E, Singh MP (2010) Biodiversity and phytosociological analysis of plants around the municipal drains in Jaunpur. Int J Biol Life Sci 6(2):77–82
Sømme L (1997) Adaptations to the alpine environment in insects and other terrestrial arthropods. In: Wielgolaski FE (ed) Polar and alpine tundra. Ecosystems of the world, vol 3. Elsevier, Amsterdam, pp 11–25
Stone WC, Frayer JRA (1935) A botanical study of pasture mixtures. Sci Aric 15:777–805
Turner MG, Gardner RH (1992) Quantitative methods in landscape ecology: the analysis and interpretation of landscape heterogeneity. Springer, New York
UNEP-WCMC (2000) Global biodiversity: Earth’s living resources in the 21st century. World Conservation Press, Cambridge
Vetaas OR, Grytnes JA (2002) Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal. Glob Ecol Biogeogr 1:291–301
Whittaker RH (1956) Vegetation of the Great Smoky Mountains. Ecol Monogr 26:1–80
Whittaker RH (1960) Vegetation of the Siskiyou Mountains, Oregon and California. Ecol Monogr 30:279–338
Wilson JB, Chiarucci A (2000) Do plant communities exist? Evidence from scaling-up local species–area relations to the regional level. J Veg Sci 11:773–775
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Japan
About this chapter
Cite this chapter
Behera, M.D., Matin, S., Roy, P.S. (2014). Biodiversity of Kargil Cold Desert in the Ladakh Himalaya. In: Nakano, Si., Yahara, T., Nakashizuka, T. (eds) Integrative Observations and Assessments. Ecological Research Monographs(). Springer, Tokyo. https://doi.org/10.1007/978-4-431-54783-9_13
Download citation
DOI: https://doi.org/10.1007/978-4-431-54783-9_13
Published:
Publisher Name: Springer, Tokyo
Print ISBN: 978-4-431-54782-2
Online ISBN: 978-4-431-54783-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)