
Abstract
Amino acids are classically considered as the building blocks for the synthesis of proteins. However, amino acids are also used as precursors for the biosynthesis of numerous compounds with various physiological functions. For instance, compounds like glutathione, creatine, carnitine, taurine, and heme of hemoglobin are synthesized in the body from amino acids in different metabolic pathways. Some amino acids like glutamine, aspartate, and lysine are nitrogen and carbon precursors for the synthesis of ribo- and deoxyribonucleotides which are precursors for DNA and RNA synthesis. Furthermore, some amino acids like arginine and glutamine are precursors for amino acids which are not present in proteins like ornithine and citrulline and which play important roles in detoxification metabolism (ammonia metabolism in the urea cycle, e.g.) and in interorgan metabolism. In addition, some amino acids like glutamine, glutamate, and aspartate are precursors of the tricarboxylic acid cycle intermediates and represent oxidative substrates in numerous cell types which use them as major energy substrates. Amino acids can also be used indirectly as energy source, notably in the postprandial phase, as precursors for neoglucogenesis and ketogenesis.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others



Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Amino acids are classically considered as the building blocks for the synthesis of proteins. However, amino acids are also used as precursors for the biosynthesis of numerous compounds with various physiological functions (Wu et al. 2007; Kim et al. 2007; Li et al. 2007). For instance, compounds like glutathione, creatine, carnitine, taurine, and heme of hemoglobin are synthesized in the body from amino acids in different metabolic pathways. Some amino acids like glutamine, aspartate, and lysine are nitrogen and carbon precursors for the synthesis of ribo- and deoxyribonucleotides which are precursors for DNA and RNA synthesis. Furthermore, some amino acids like arginine and glutamine are precursors for amino acids which are not present in proteins like ornithine and citrulline and which play important roles in detoxification metabolism (ammonia metabolism in the urea cycle, e.g.) and in interorgan metabolism. In addition, some amino acids like glutamine, glutamate, and aspartate are precursors of the tricarboxylic acid cycle intermediates and represent oxidative substrates in numerous cell types which use them as major energy substrates (Yao et al. 2012). Amino acids can also be used indirectly as energy source, notably in the postprandial phase, as precursors for neoglucogenesis and ketogenesis.
In addition, and more recently, amino acids by themselves or after conversion in secondary metabolites have been associated with “signal functions” associated with metabolic and physiologic modifications. Just to take an example, L-arginine, after its enzymatic conversion by one isoform of nitric oxide synthase into nitric oxide (NO) and L-citrulline in endothelial cells, allows the action of NO which acts as a regulator of the arterial pressure. It is also feasible to consider that some amino acids, which are acting as neurotransmitter (like glutamate) or are precursor of neurotransmitter (like tryptophan which is a precursor of serotonine) or of biogenic amines (like histidine which is a precursor for histamine production), are amino acids with signaling functions.
Furthermore, an amino acid-like l-arginine is able to exert both metabolic regulatory (by activating the first step of ammonia detoxification in the urea cycle) and secretagogue functions (by stimulating for instance insulin and growth hormone secretion). Last but not least, the role of leucine as a stimulator of the protein synthesis through its action on the intracellular signaling pathways represents a very interesting example of the signaling functions of the amino acids (Yin et al. 2010; Li et al. 2011).
Proteins of all biological organisms, from the virus and bacteria to humans, are made up from the same set of 20 amino acids. Nineteen of these are α-amino acids with a primary amino group (−NH3 +) and a carboxyl group (−COO−) attached to a central carbon atom, aspartate ammonia-lyase, which is called the α-carbon atom (Cα) because it is adjacent to the carboxyl group (Fig. 7.1). The remaining groups attached to the Cα atom are a hydrogen atom (H) and a variable side chain or R′ group. The one exception to this general structure is proline, a cyclic amino acid, which has a secondary amino group and is thus an α-imino acid. The only difference between any two amino acids is in their different side chains. Each side chain has distinct properties, including charge, hydrophobicity, and polarity. It is the arrangement of amino acids, with their distinct side chains, that is mainly responsible for protein structure and related functions.

General structure of an amino acid. Chemical structure of an amino acid showing the four different groups around the central α-carbon atom (with the exception of proline). The R group or side chain (red) attached to the α-carbon (blue) is different in each amino acid
All the 20 amino acids, except glycine (Gly or G), have four different groups arranged in tetrahedra around the central Cα atom which is thus known as an asymmetric center or chiral center and has the property of chirality. We note in Fig. 7.1 that the α-carbon is asymmetric, bonded to four different substituent groups: a carboxyl group, an amino group, an R group, and a hydrogen atom. Because of the tetrahedral arrangement of the bonding orbitals around the α-carbon atom of amino acids, the four different substituent groups can occupy two different arrangements in space, which are nonsuperimposable mirror images of each other. These two forms are called enantiomers or stereoisomers. Enantiomers are physically and chemically indistinguishable by most techniques, but can be distinguished on the basis of their different optical rotation of plane-polarized light. Molecules are classified as dextrorotatory (d; Greek dextro = right) or levorotatory (l; Greek levo = left) depending on whether they rotate the plane of plane-polarized light in a clockwise or an anticlockwise manner (Fig. 7.2). Surprisingly, only the l-amino acids are found in proteins, while d-isomers have been found only in small peptides of bacterial cell walls and in some peptide antibiotics.

Stereoisomerism in amino acids. The two stereoisomers of alanine, l- and d-alanine, are nonsuperimposable mirror images of each other (enantiomers)
Amino acids joined together by peptide bonds form the primary structure of a protein. The amino group of one molecule reacts with the carboxyl group of the other in a condensation reaction resulting in the elimination of water and the formation of a dipeptide (Fig. 7.3). A short sequence of amino acids is called a peptide, with the term polypeptide applied to longer chains of amino acids, usually of known sequence and length. When joined in a series of peptide bonds, amino acids are called “residues” to distinguish between the free form and the form found in protein. The peptide bond has a partial double-bond character.

Formation of a peptide bond. The peptide bond is a chemical, covalent bond formed between the α-amino group of one amino acid and the α-carboxyl group of another. Once two amino acids are joined together via a peptide bond to form a dipeptide, there is still a free amino group at one end and a free carboxyl group at the other, each of which can in turn be linked to further amino acids
2 Classification of Amino Acids
There are reasons to believe that amino acids are the oldest nutrients that have existed on earth. They have been used as the source of life and are sometimes called as the building blocks of life. There are as many as hundred thousand kinds of proteins that constitute the body, and these proteins are made with only 20 amino acids arranged in various combinations. The combination of the amino acids in proteins depends on the genetic code. In addition to the amino acids found in proteins, some amino acids not involved in the proteosynthesis are found in living organisms.
The names of the amino acids are often abbreviated, either to three letters or to a single letter. Thus, for example, alanine is abbreviated to Ala or A (Table. 7.1).
The hydrophobicity index informs on the relative hydrophobicity of amino acids. Higher positive value indicates stronger hydrophobicity. Hydrophilic amino acids have negative values. In a protein, hydrophobic amino acids are more likely to be located in the protein interior, whereas hydrophilic amino acids are more likely to face the aqueous environment.
2.1 Common Proteinogenic Amino Acids
Some of the 20 standard amino acids are called essential amino acids, because they cannot be synthesized by the body from other compounds through available metabolic pathways in the body, but instead must be provided by dietary sources. In humans, the nine essential amino acids are lysine, leucine, isoleucine, methionine, phenylalanine, threonine, tryptophan, valine, and (in children) histidine. It is thus necessary to provide these essential amino acids from proteins in food in well-balanced, appropriate amounts and, in some cases, to provide some individual amino acids which are limiting for protein biosynthesis. Some amino acids are considered as conditionally indispensable since in some physiological situation (for instance in neonates) and in some pathological situations, the needs for some amino acids are increased to such an extent that endogenous synthesis capacity is not able to provide enough amino acids related to these needs.
Based on the chemical structure of the R groups, the 20 amino acids of proteins can be divided into aliphatic amino acids, aromatic amino acids, and heterocyclic amino acids, and among them, aliphatic amino acids are the most.
2.1.1 Aliphatic Amino acids
-
1.
Neutral amino acids (Fig. 7.4):
Fig. 7.4 
Neutral aliphatic amino acids
-
(a)
Glycine (Gly or G): The smallest amino acid with the simplest structure, has an hydrogen atom in the side-chain position, and thus does not exist as a pair of stereoisomers since there are two identical groups (hydrogen atoms) attached to the Cα atom. In addition, Gly has minimal steric hindrance and in protein structure Gly offers the most flexibility.
-
(b)
Alanine (Ala or A).
-
(c)
Valine (Val or V).
-
(d)
Leucine (Leu or L).
-
(e)
Isoleucine (Ile or I).
-
(a)
-
2.
Hydroxyl-containing or sulfur-containing amino acids (Fig. 7.5):
Fig. 7.5 
Hydroxyl-containing or sulfur-containing amino acids
-
(a)
Serine (Ser or S).
-
(b)
Threonine (Thr or T).
-
(c)
Cysteine (Cys or C): Capable of reacting with another cysteine to form a disulfide bond.
-
(d)
Methionine (Met or M).
-
(a)
-
3.
Acidic amino acids and their amide derivatives (Fig. 7.6):
Fig. 7.6 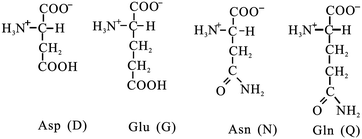
Acidic amino acids and their amide derivatives
-
(a)
Aspartate (Asp or D).
-
(b)
Glutamate (Glu or E).
Asp and Glu have carboxyl in their R groups. They have net negative charge at pH 7.0, thus usually named as aspartate and glutamate (conjugate base names, instead of aspartic acid and glutamic acid, unionized form).
-
(c)
Asparagine (Asn or N).
-
(d)
Glutamine (Gln or Q).
-
(a)
-
4.
Basic amino acids (Fig. 7.7):
Fig. 7.7 
Basic amino acids and heterocyclic amino acids
-
(a)
Lysine (Lys or K).
-
(b)
Arginine (Arg or R).
-
(a)




2.1.2 Aromatic Amino Acids (Fig. 7.8)

Aromatic amino acids
-
1.
Phenylalanine (Phe or F): Which contains a benzene ring.
-
2.
Tyrosine (Tyr or Y): Which contains a benzene ring.
-
3.
Tryptophan (Trp or W): Which contains an indole ring.
They are jointly responsible for the light absorption of proteins at 280 nm, a property used as a measure of the concentration of proteins.
2.1.3 Heterocyclic Amino Acids (Fig. 7.7)
-
1.
Histidine (His or H).
-
2.
Proline (Pro or P): Has an imino group, instead of an amino group, forming a ring structure, being rigid in conformation. It is often found in the bends of folded protein chains and often present on the surface of proteins. It offers the least flexibility.
Based on the polarity of R groups, the 20 amino acids of proteins can be classified as follows: (1) nonpolar, (2) polar but uncharged, (3) positively charged, and (4) negatively charged (Table. 7.2).
-
1.
Nonpolar amino acids
The R groups in this class of amino acids are nonpolar and hydrophobic. The bulky side chains of alanine, valine, leucine, and isoleucine, with their distinctive shapes, are important in promoting hydrophobic interactions within protein structures. Glycine has the simplest amino acid structure. Where it is present in a protein, the minimal steric hindrance of the glycine side chain allows much more structural flexibility than the other amino acids. Proline represents the opposite structural extreme. The secondary amino (imino) group is held in a rigid conformation that reduces the structural flexibility of the protein at that point. Methionine, one of the two sulfur-containing amino acids, has a nonpolar thioether group in its side chain. Phenylalanine and tryptophan, with their aromatic side chains, are relatively nonpolar (hydrophobic).
-
2.
Polar-uncharged amino acids
The R groups of these amino acids are more water soluble, or hydrophilic, than those of the nonpolar amino acids, because they contain functional groups that form hydrogen bonds with water. This class of amino acids includes serine, threonine, cysteine, asparagine, glutamine, and tyrosine. The polarity of serine, threonine, and tyrosine is contributed by their hydroxyl groups; that of cysteine and methionine by their sulfur atom; and that of asparagine and glutamine by their amide groups.
-
3.
Positively charged amino acids
The amino acids in which the R groups have significant positive charge at pH 7.0 are lysine, which has a second amino group at the e position on its aliphatic chain; arginine, which has a positively charged guanidino group; and histidine, containing an imidazole group. Histidine is the only standard amino acid having a side chain with a pK a near neutrality. This group of amino acids is sometimes refereed as cationic amino acids.
-
4.
Negatively charged amino acids
The two amino acids having R groups with a net negative charge at pH 7.0 are aspartate and glutamate, each with a second carboxyl group. These amino acids are the parent compounds of asparagine and glutamine, respectively. This group of amino acids is sometimes referred as anionic amino acids.
2.2 Uncommon Proteinogenic Amino Acids
In addition to the 20 usual amino acids that are common in all proteins, other amino acids have been found as components of only certain types of proteins (Fig. 7.9). Each of these is derived from one of the 20 usual amino acids, in a modification reaction that occurs after the standard amino acid has been inserted into a protein. For example, 4-hydroxyproline, a derivative of proline, is found in the fibrous protein collagen of connective tissues and 4-hydroxylysine is found in plant cell-wall proteins. N-methyllysine is found in myosin, a contractile protein of muscle. Another important unusual amino acid is γ-carboxyglutamate, found in the blood-clotting protein prothrombin as well as in certain other proteins that bind Ca2+ for their biological function. More complicated is the nonstandard amino acid desmosine, a derivative of lysine residues, found in the fibrous protein elastin. Selenocysteine contains selenium rather than the oxygen of serine, and is found in glutathione peroxidase and a few other proteins.

Some of the uncommon proteinogenic amino acids
2.3 Nonprotein Amino Acids
Some 300 additional amino acids have been found in cells and have a variety of functions but are not substituents of proteins (Fig. 7.10). Ornithine and citrulline which are also present in proteins deserve special attention because they are key intermediates in the biosynthesis of arginine and in the urea cycle.

Some of the amino acids and nitrogenous metabolites not found in proteins
3 Function of Amino Acids
The aim of this paragraph is to briefly recapitulate the metabolism of each amino acid together with their main biological functions.
3.1 Alanine
Alanine transaminase catalyzes the reversible conversion of l-alanine and alpha-ketoglutarate into pyruvate and glutamate. Alanine is a substrate for the neoglucogenesis.
3.2 Arginine
l-arginine is a precursor for biosynthesis of other amino acids present in proteins (glutamate and proline) or not present in proteins (ornithine, citrulline). Arginine is a precursor of urea, polyamines, creatine, and nitric oxide. Polyamines are involved in cell mitosis. Creatine is synthesized from arginine, glycine, and methionine but can also be provided by the diet. Creatine and creatine phosphate are mainly found in skeletal muscles. Phosphocreatine plays a central role in muscle contraction. Nitric oxide has been implicated in numerous functions, but has been initially shown to be produced in vascular endothelial cells and to be implicated in vasorelaxation. Arginine is used in urea cycle for ammonium detoxification.
3.3 Asparagine
l-asparagine can be converted into aspartate and ammonium.
3.4 Aspartic Acid
Aspartate transaminase is responsible for the reversible conversion of aspartate and alpha-ketoglutarate into oxaloacetate and glutamate. Aspartate is involved in the urea cycle for the conversion of citrulline into argininosuccinate. Aspartate is a precursor for pyrimidine synthesis.
3.5 Cysteine
Cysteine can be produced from the metabolic conversion of methionine and serine. Cysteine is a precursor for taurine biosynthesis and is one of the amino acids which are precursors for glutathione and hydrogen sulfide (H2S). Cysteine is a precursor for pyruvate synthesis and for the synthesis of the nucleotidic coenzyme CoA. Taurine is a beta-amino sulfonic acid which is not incorporated into proteins. The circulating concentrations of taurine are much dependent on the dietary intake (Laidlaw et al. 1998). Taurine functions are numerous. This amino acid is involved in the synthesis of conjugated biliary acids. Taurine has been implicated as a regulator of the cellular volume, and as such, as an agent with cytoprotective effect. A protective role of taurine against deleterious effects of the “oxidative stress” has been proposed and may be due to the reaction between taurine and hypochlorous acid (Schuller-Levis and Park, 2004). Glutathione is a tripeptide which is synthesized in cells from glutamate, cysteine, and glycine. Circulating glutathione is usually micromolar (Jones et al. 2000). This compound from alimentary origin can be absorbed and the jejunum represents the main site of absorption (Hagen et al. 1990). Glutathione can exist in cells both in reduced (GSH) and oxidized form (GSSG) with reduced glutathione being much prominent. The GSH/GSSG couple represents the main component fixing the intracellular redox state. In mammals, the amount of cysteine used for glutathione synthesis is very high since it represents as much as 30–50% of the total body utilization (Fukagawa et al. 1996). Glutathione is involved in several physiological functions including the elimination of electrophilic compounds, control of intracellular reactive oxygen species, etc.
3.6 Glutamic Acid
Glutamic acid is metabolized in transamination pathways. This amino acid can be converted in the presence of pyruvate into alpha-ketoglutarate and alanine, and in the presence of oxaloacetate into aspartate and alpha-ketoglutarate. Glutamic acid can also undergo conversion to alpha-ketoglutarate and ammonium. As such, glutamate is an oxidative substrate in several cell types. Glutamic acid can be synthesized from other amino acids like arginine, proline, and histidine. Glutamine is a precursor of glutamate and ammonium. Inversely, glutamate is a precursor for glutamine synthesis. Lastly, glutamate is one of the amino acid precursors for glutathione synthesis (Wang et al. 2008).
3.7 Glutamine
Glutamine is metabolized into glutamate, ammonium, alanine, aspartate, citrulline and proline. Glutamine is a major oxidative substrate in numerous cell types. Glutamine is a nitrogen donor for the biosynthesis of nucleotides.
3.8 Glycine
Glycine is a precursor for the biosynthesis of serine and for the synthesis of glutathione, creatine, purines, and heme in hemoglobin.
3.9 Histidine
Histidine is abundant in hemoglobin. Histidine is a precursor of glutamate and histamine. This latter compound is involved in allergy and in inflammatory reactions.
3.10 Isoleucine, Leucine, and Valine
The amino acids with branched chain undergo the transamination in the presence of alpha-ketoglutarate allowing the synthesis of glutamate and of the corresponding alpha-ketoacids (alpha-ketoisocaproate from leucine, alpha-keto-beta-methyl-valerate from isoleucine, and alpha-ketoisovalerate from valine). Each of these ketoacids undergoes then an oxidative decarboxylation by a ketoacid dehydrogenase (BCKAD). The products of decarboxylation undergo then several metabolic steps leading to the synthesis of acetyl-CoA, acetoacetic acid, propionyl-CoA, and succinyl CoA. It is well known that branched-chain amino acids can be in competition with other amino acids (notably tryptophan and tyrosine) for transport through cellular membranes (Anderson and Johnston 1983). Although branched-chain amino acids are not direct precursors for neurotransmitter synthesis, they can affect the transport of other amino acids across the blood–brain barrier, and then the concentrations of some neurotransmitters in the central nervous system (Fernstrom, 1973).
3.11 Lysine
Lysine is a precursor for acetoacetyl-CoA, carnitine, and cadaverine. Carnitine is a metabolite with important functions in the intermediary metabolism. Carnitine is involved in the transport of fatty acids with long chain from the cytosol to the mitochondrial matrix where the beta-oxidation occurs. In the body, carnitine is originating mainly from dietary source, with endogenous synthesis playing a minor role (Vaz and Wanders, 2002). Carnitine is mainly synthesized in the liver from lysine and methionine. Cadaverine is a polyamine produced by the intestinal microbiota.
3.12 Methionine
Methionine is a precursor of succinyl-CoA, homocysteine, cysteine, creatine, and carnitine. In addition, methionine is a precursor for S-adenosyl-methionine synthesis which is involved in polyamine, creatine, and phosphatidylcholine (one of the membrane phospholipids) metabolism.
3.13 Phenylalanine
This amino acid is a precursor for the synthesis of tyrosine and acetoacetyl-CoA.
3.14 Proline
Proline is a precursor for glutamate synthesis in several cell types. Proline can undergo hydroxylation leading to the production of hydroxyproline. Proline and hydroxyproline are abundant in collagens.
3.15 Serine
Serine is one of the precursors for the biosynthesis of cysteine and for the synthesis of the sphingolipid sphingosine. This amino acid can also be metabolized into pyruvate.
3.16 Threonine
This amino acid is a precursor for the synthesis of glycine and acetyl-CoA. This amino acid is very abundant in mucins (Wang et al. 2007).
3.17 Tryptophan
Tryptophan is a precursor of numerous metabolites including the neurotransmitter serotonine (5-hydroxytryptamine), tryptamine, the hormone melatonin, and niacin (vitamin B3). In addition, tryptophan is a precursor for the synthesis of acetyl-CoA (Yao et al. 2011).
3.18 Tyrosine
Tyrosine is a precursor for active substances including the neurotransmitters epinephrine and norepinephrine, the pigment melanin, and the hormone thyroxine. Tyrosine is also a precursor for the production of acetoacetyl-CoA.
It is worth to note that although metabolites derived from amino acids play major role in the body physiology, the metabolic pathways involved in their synthesis represent a very minor utilization of the amino acid precursors when compared with the utilization of the amino acids for protein synthesis.
References
Anderson GH, Johnston JL (1983) Nutrient control of brain neurotransmitter synthesis and function. Can J Physiol Pharmacol 61:271–281
Fernstrom JD (1973) Correlations between brain tryptophan and plasma neutral amino acid levels following food consumption in rats. Life Sci 13:517–524
Fukagawa NK, Ajami AM, Young VR (1996) Plasma methionine and cysteine kinetics in response to an intravenous glutathione infusion in adult humans. Am J Physiol 270:E209–E214
Hagen TM, Wierzbicka AH, Sillau AM, Bowmann BB et al (1990) Bioavailability of dietary glutathione: effects on plasma concentration. Am J Physiol 259:G524–G529
Jakobs BS, Wanders RJ (1995) Fatty acid beta-oxidation in peroxisomes and mitochondria: the first, unequivocal evidence for the involvement of carnitine in shuttling propionyl-CoA from peroxisomes to mitochondria. Biochem Biophys Res Commun 213:1035–1041
Jones DP, Carlson JL, Mody VC, Cai J et al (2000) Redox state of glutathione in human plasma. Free Radic Biol Med 28:625–635
Kim SW, Mateo RD, Yin YL, Wu GY (2007) Functional amino acids and fatty acids for enhancing production performance of sows and piglets. Asian-Australas J Anim Sci 20:295–306
Laidlaw SA, Shultz TD, Cecchino JT, Kopple JD (1998) Plasma and urine taurine levels in vegans. Am J Clin Nutr 47:660–663
Li P, Yin YL, Li DF, Kim SW, Wu GY (2007) Amino acids and immune function. Br J Nutr 98:237–252
Li F, Yin Y, Tan B, Kong X, Guoyao W (2011b) Leucine nutrition in animals and humans: mTOR signaling 3 and beyond. Amino Acids 41:1185–1193
Schuller-Levis GB, Park E (2004) Taurine and its chloramines: modulators of immunity. Neurochem Res 29:117–126
Vaz FM, Wanders RJ (2002) Carnitine biosynthesis in mammals. Biochem J 361:417–429
Wang X, Qiao SY, Yin YL, Yu LG, Wang ZY, Wu GY (2007) A deficiency or excess of dietary threonine reduces protein synthesis in jejunum and skeletal muscle of young pigs. J Nutr 137:1442–1446
Wang JJ, Chen LX, Li P, Li XL, Zhou HJ, Wang FL, Li DF, Yin YL, Wu GY (2008) Gene expression is altered in piglet small intestine by weaning and dietary glutamine supplementation. J Nutr 138:1025–1032
Wu GY, Bazer FW, Davis TA, Johnson GA, Kim SW, Knabe DA, Spencer TE, Yin YL (2007) Important roles for arginine-family amino acids in swine nutrition and production. Livest Sci 122:8–22
Yao K, Fang J, Yin YL, Feng ZM, Tang ZR, Wu GY (2011) Tryptophan metabolism in animals: important roles in nutrition and health. Front Biosci S3:286–297
Yao K, Wang L, Ding BY, Fu DB, Liu YL, Zhu HL, Liu J, Li YT, Kang P, Yin YL, Wu GY, Hou YQ (2012) Alpha-ketoglutarate inhibits glutamine degradation and enhances protein synthesis in intestinal porcine epithelial cells. Amino Acids 42(6):2491–2500
Yin YL, Yao K, Liu ZJ, Gong M, Ruan Z, Deng D, Tan BE, Liu ZQ, Wu GY (2010) Supplementing L-leucine to a low-protein diet increases tissue protein synthesis in weanling pigs. Amino Acids 39:1477–1486
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer-Verlag Wien
About this chapter
Cite this chapter
Ren, W., Li, Y., Yin, Y., Blachier, F. (2013). Structure, Metabolism and Functions of Amino Acids: An Overview. In: Blachier, F., Wu, G., Yin, Y. (eds) Nutritional and Physiological Functions of Amino Acids in Pigs. Springer, Vienna. https://doi.org/10.1007/978-3-7091-1328-8_7
Download citation
DOI: https://doi.org/10.1007/978-3-7091-1328-8_7
Published:
Publisher Name: Springer, Vienna
Print ISBN: 978-3-7091-1327-1
Online ISBN: 978-3-7091-1328-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)