
Abstract
Immunosenescence contributes to render older adults more susceptible to pathogens and to develop main age-related diseases. With the ever-increasing rise in the number and proportion of older persons worldwide, one of the challenges of a “long-life society” is to ensure that the years gained are not only healthy and disability-free years but also offering a good quality of life. In this perspective, recent research has concentrated on identifying the factors hindering healthy ageing, and among them, the age-related changes of the immune system have thus been particularly investigated. With respect to its central role in orchestrating the immune response, this chapter describes the main features of T cell-mediated immune senescence and the underlying mechanisms contributing to this state of increased vulnerability. Furthermore, it will explore the means by which T cell functions could be identified by using biomarkers.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
10.1 Introduction
Advances in medicine and technologies and socioeconomic development have all contributed to the well-being of mankind and an unprecedented increasing in longevity (Lang and Aspinall 2012). Consecutively, in addition to declining fertility this has also led to a continuous rise in the number and proportion of older persons worldwide (Lutz et al. 1997; Oeppen and Vaupel 2002). Presently 673 million inhabitants in the world are aged ≥60 years, among them 88 million are ≥80 years old. According to the United Nations Prospects (United Nations 2008), the expected numbers for 2050 are of 2 billion (60 and over) and 400 million (80 and over), which means a multiplication by 3 and 4.5, respectively. Today, 21 % of the European population and 17 % of the Northern American population are aged 60 and above. Those figures will increase to 35 and 27 %, respectively, by 2050. Five years from now, for the first time in the human being history, the number of people aged 65 years or older will outnumber children younger than 5 years (Shetty 2012). Moreover, one of the challenges in industrialized societies is that aging will progressively impact on every country in the world. Indeed, whether today, 50 % of the population aged ≥80 years lives in the most developed countries (29 % of them in Europe and 13 % in Northern America); tomorrow, less-developed countries will also observe this demographic transition. By 2050, 62 % of the aged ≥60 years population will live in Asia (United Nations 2008).
Although the aging of the general population is one of humanity’s greatest triumphs (Lloyd-Sherlock et al. 2012), it also confronts our societies to enormous medical challenges (Oeppen and Vaupel 2002). If aging should be considered as being a positive experience (World Health Organization 2002), lengthening lifetime is not necessarily synonymous of extending life expectancy in good health; studies demonstrated that chronic and degenerative disorders become more and more prevalent with advancing age and multimorbidity is increasing (Thorpe and Howard 2006). Thus, the optimism created by the ever-increasing life expectancy, and the expectation of many individuals that they will live longer and healthier, must be balanced by the increased number of older individuals (Lang and Aspinall 2012). Thus, one of the challenges of a “long-life society” is to ensure that the years gained with a higher life expectancy are not only healthy and disability-free years but are years offering a good quality of life. In this perspective, recent research has concentrated on identifying the factors contributing rather than hindering the healthy aging process, and among them, the age-related changes of the immune system, commonly termed immunosenescence, have been particularly investigated (Weiskopf et al. 2009; Ongrádi and Kövesdi 2010).
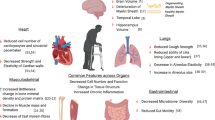
As depicted in the Fig. 10.1, immunosenescence has been implicated in the increasing state of susceptibility to pathogens previously encountered and the decline in the body’s ability to mount adequate immune responses to new antigens. It also acts as a contributing factor to the increased susceptibility of older adults to develop not only infectious diseases but cancer, Alzheimer’s diseases, osteoporosis, insulin resistance, diabetes, atherosclerosis and autoimmunity, and other main aged-related diseases (Lang et al. 2010c; Fulop et al. 2011; Giunta et al. 2008; Ginaldi et al. 2005). Although individuals’ age seems to be a major contributor of this state of vulnerability, there is no single cause of immunosenescence, which is the consequence of a compilation of events (Govind et al. 2012; Lang et al. 2013a), including thymic involution (Aspinall et al. 2010), the continuous reshaping of the immune repertoire by persistent antigenic challenges (Virgin et al. 2009), the dysregulation of Toll-like receptor (TLR) functions (Shaw et al. 2011), the reduced production of naïve B cells and the intrinsic defects arising in resident B cells (Frasca et al. 2011), and the impact of nutritional status and dysregulation of hormonal pathways (Lang et al. 2012a; Kelley et al. 2007). Moreover, human aging is, by itself, also inextricably linked with an ever-increasing incidence of chronic-comorbid conditions which contribute to increase the age-related chronic low-grade inflammation (i.e., inflammaging) that further impinge the immune system (Fulop et al. 2010). Effects of immunosenescence are now evident in both arms of the immune system: the innate and adaptive immune system (Lang et al. 2010b). With respect to its central role in orchestrating the immune response, this chapter will focus on the main features of T cell-mediated immunity senescence and the underlying mechanisms contributing to the age-related state of increased vulnerability. Furthermore, it will explore the means by which T cell functions could be identified and predicted by using biomarkers.

The immunosenescence process with advancing age. This figure depicts the concept of accumulation of deficits applied to immunosenescence. Thus, an accumulation of immune deficits could be used to predict immune status of a given individual. In complex systems such as the immune system, reliability of functions being undertaken is dependent in part of the quality of the component and also on any functional overlap. Thus, reliability in the face of possible component failure can be achieved by having multiple components capable of fulfilling the same task ensuring that while some components fail the system as a whole remains functional. CMV cytomegalovirus, IL interleukin, TNF tumor necrosis factor, and TREC T cell receptor excision circles
10.2 What Are the Main Features of the Senescence of T Cell Immunity?
Quantification of T cell numbers throughout the life-span shows that they are maintained in humans (Aspinall et al. 2010) even in their tenth decades at levels which are comparable to those found in younger individuals (Mitchell et al. 2010). This would imply that there is no decline in the homeostatic mechanisms which preserve the size of the peripheral T cell pool within defined boundaries. As shown in Fig. 10.1, the age-related changes in cell-mediated immunity are characterized by two major patterns: the reduction in thymic output (i.e., decrease in naïve T cells) and the increase in the number of antigen-experienced memory and in particular effector cells (i.e., increase in senescent cells) (Weiskopf et al. 2009). In addition, but not further detailed thereafter, thymic involution also leads to a decreased output of T regulatory cells (Treg) which have been reported to decline after the age of 50 and could contribute to age-related phenomena such as autoimmunity and increased inflammation as well (Tsaknaridis et al. 2003; Weiskopf et al. 2009).
10.2.1 Decreased Number of Naïve T Cells
Production and maintenance of the peripheral naïve T cell repertoire are critical to the normal function of the adaptive immune system as a whole (Ongrádi and Kövesdi 2010; Weiskopf et al. 2009). As a result of thymic involution, the output of peripheral naïve T cells is dramatically reduced with advancing age. Indeed, at birth the thymus is fully developed, its involution and the replacement of the active areas of thymopoiesis related to fat accumulation start soon after birth and continue throughout life, and it is almost complete at the age of 50 years (Aspinall et al. 2010). This leads to a reduced ability of the aged host to respond adequately to new antigens (Naylor et al. 2005), including not only pathogen but also vaccine antigens (Lang et al. 2010a). Low naïve T cell numbers have been described in the periphery as well as lymphoid tissues (Aspinall et al. 2010). The thymic involution appears to be one of the major features of human immunosenescence (Ostan et al. 2008; Aspinall et al. 2010) because this is the single preceding event in all cases.
In the older adults, both the diversity and functional integrity of the naïve CD4+ and CD8+ T cells subsets are decreased, but in slightly different ways (Naylor et al. 2005; Aspinall and Andrew 2000; Weinberger et al. 2007; Effros et al. 2003; Pfister et al. 2006; Lang et al. 2013a). Although the diversity and number of naïve CD4+ T cell compartment are maintained stable for a long time, a dramatic and sudden collapse of diversity occurs after the age of 70 years, considerably shrinking the repertoire. Similar changes occur, but earlier and more gradually when aged, in the naïve CD8+ T cells subset (Arnold et al. 2011). In contrast with CD4+, naïve CD8+ T cells are more susceptible to apoptosis in aged individuals (Gupta and Gollapudi 2008). The reduced thymic output of newly generated naïve T cells seems to be compensated by different mechanisms, and among them homeostatic proliferation has been identified as playing a key role for the maintenance and restoration of the size of the naïve T cell pool (Hazenberg et al. 2003). Indeed, naive T cells are also readily detectable in elderly people (Douek et al. 1998; Chen et al. 2010), and adult thymectomy does not lead to a rapid decline in naïve T cell number. Similar data have been reported for juvenile rhesus macaques, where thymectomy did not accelerate age-related naïve T cell decline (Hazenberg et al. 2003). Thus, T cell can be produced at extra-thymic sites, such as peripheral lymph nodes and the gut.
Thymic atrophy and decreased thymopoiesis are active processes mediated by the upregulation of cytokines, i.e., interleukin (IL)-6, leukemia inhibitory factor (LIF), and oncostatin M (OSM), in aged human being and mice thymus tissue (Ongrádi and Kövesdi 2010; Sempowski et al. 2000), while IL-7 production by stromal cell is significantly decreased (Ortman et al. 2002; Andrew and Aspinall 2002). IL-7 is necessary for thymopoiesis (Surh and Sprent 2002), promoting cell survival by maintaining the antiapoptotic protein B cell lymphoma 2 (Bcl-2) and inducing VDJ recombination (Fig. 10.2) (Kim et al. 1998; Jiang et al. 2005; Aspinall and Andrew 2000). The above changes result in decreased thymic output, in diminished number of circulating naïve T cells (i.e., CD45RA+CD28+ and CD45RA+CD28+CD26L) in the blood stream and lymph nodes (Aspinall et al. 2010; Ongrádi and Kövesdi 2010). Naïve T cells from aged individuals exhibit numerous functional defects which are accelerated according to the increasing homeostatic proliferation (Arnold et al. 2011). Indeed, if the naïve T cell count drops below 4 % of total T cells, homeostatic proliferation increases exponentially. This leads, for example, to accelerated telomere shortening and may lead to a memory-like phenotype (Kilpatrick et al. 2008; Cicin-Sain et al. 2007).

Schematic representation of the somatic rearrangement process undergoing in every immature T cell TCR loci during the development from hematopoietic stem cell to mature naïve T cells. During the rearrangement process, the intervening DNA sequences, both for α- and β-chain, are deleted and circularized into episomal DNA molecules, called TCR excision circles (TRECs) (Adapted from Lang et al. (2013a))
Restricted T cell receptor (TCR) repertoire, reduced cytokine production, and impaired expansion and differentiation into effector cells following antigen stimulation are also described (Weiskopf et al. 2009; Ferrando-Martinez et al. 2011). Thus, CD45RA+CD28+CD8+ in aged individuals produce larger amounts of proinflammatory cytokines such as interferon gamma (IFN-γ) and IL-2 and have a highly restricted TCR repertoire compared to younger adults (Aspinall et al. 2010; Pfister and Savino 2008). It has been also demonstrated that the CD4+ subtype does not form immunologic synapses upon stimulation with peptide antigen and antigen presenting cell and this partly through age-associated defects in TCR signaling (Arnold et al. 2011). In addition, naïve T cells from aged individuals exhibit numerous functional defects in their activation, expansion, and differentiation that may also considerably affect their cognate helper function to B cells hence leading to reduced antibody-mediated immunity following antigen stimulation (Weiskopf et al. 2009; Ferrando-Martinez et al. 2011; Haynes 2005).
10.2.2 The Expansion of Dysfunctional Terminally Differentiated T Cells
Consequently to decreasing thymopoiesis, a shift in the ratio of naïve to memory T cells with an increasing number of the memory compartment in order to maintain peripheral T cell homeostasis is observed with advancing age. In contrast to naïve T cells, memory cells rely on IL-7 in concert with IL-15, cycle and self-renew in vivo three- to fourfold faster than naïve cells, and thereby are capable of vigorous proliferation (Arnold et al. 2011; Surh and Sprent 2002). Homeostatic turnover of naïve CD8+ T cells that induces a memory-like phenotype further contributes in a dramatically reduced diversity of the memory T cell pool in the elderly individuals (Weinberger et al. 2007; Naylor et al. 2005).
Repeated antigenic stimulation by certain pathogens shapes the T cell pool and directly contributes to immunosenescence (Virgin et al. 2009; Ongrádi and Kövesdi 2010) by accumulating clones of certain specificities (Karrer et al. 2003; Virgin et al. 2009). Scientific evidence has indeed accumulated that persistent viral infections play a major role in driving clonal expansion, contraction, and homeostasis of the T cell compartment leading to the age-dependent accumulation of dysfunctional terminally differentiated T cells (CD8+CD28−) commonly named senescent cells (Brunner et al. 2011; Virgin et al. 2009). While some reports suggest that localized, niche limited, latent herpes virus including Epstein-Barr virus (EBV), varicella-zoster virus (VZV), herpes simplex viruses (HVS) 1, and HVS2 may not have any impact, persistent viral infection with HCV (Gruener et al. 2001), HIV (Pantaleo et al. 1997; Sewell et al. 2000; Shankar et al. 2000), and CMV (Pawelec et al. 2009) has been shown to cause chronic inflammation of exhausted T cells, even already early in life. Indeed, T cells are thus repeatedly stimulated by viral antigens thereby contributing to the massive accumulation of virus-specific CD4+ and CD8+ T cell clone. This has been observed in both mice and humans (Arnold et al. 2011). Particularly in the elderly individuals, the lifelong exposure to CMV severely impairs the T cell-mediated immune system by increasing the number of highly differentiated, exhausted CMV-specific CD4+ and CD8+ T cells (Pawelec et al. 2009).
One of the most robust markers in describing these exhausted T cell is the loss of the costimulatory molecule CD28 which has been furthermore reported as key predictor of immune incompetence in older individuals (Vallejo 2005; Frasca et al. 2011). CD28 marker is expressed constitutively on >99 % of human T cells at birth. With advancing age a progressive increase in the proportion of CD28− T cells is observed and particularly within the CD8+ T cell subset (Lang et al. 2010a). CD28-mediated costimulation is needed for effective primary T cell expansion and for the generation and activation of Treg cells (Hünig et al. 2010). CD28 signal transduction results in IL-2 gene transcription, expression of IL-2 receptor, and the stabilization of a variety of cytokine messenger RNAs. Consequently, memory CD8+CD28− T cells generated from aged naïve T cells, compared to memory cells produced from young naïve cells, produced much less cytokine (IL-2 from T helper (Th) 1 and IL-4 and IL-5 from Th2) (Ongrádi and Kövesdi 2010). Aged CD4+CD28− produced from aged naïve cells also expressed decreased CD40-ligand (CD40L or CD154) marker. The CD154 ligand has been shown to induce cytokine production and costimulate proliferation of activated T cells, and this is accompanied by production of IFN-γ, tumor necrosis factor alpha (TNF-α), and IL-2. Hence, the capacity of these cells to help in B cell proliferation and antibody production is considerably reduced contributing to the impairment of humoral response in the aged (Haynes 2005; Frasca et al. 2011).
Globally, the proliferative capacity of CD28− T cells is also limited; these cells have shortened telomeres and show increased resistance to apoptosis and restricted T cell diversity and are named senescent cells (Vallejo 2005). These cells are also able to secrete proinflammatory cytokines such as TNF-α and INF-γ through a switch from Th1- to Th2-like cytokine response that contributes to the ongoing age-related proinflammatory background observed in elderly persons (i.e., inflammaging) (Franceschi et al. 2007). Senescent cells also exert regulatory roles in vivo that further impinge the immune system capacities such as poorer immune responses to influenza vaccination (Goronzy et al. 2001; Saurwein-Teissl et al. 2002) and higher autoreactive immunologic memory (Weiskopf et al. 2009). Recently, it has been demonstrated that senescent CD8+ T cells are specially enriched in niches such as the bone marrow, where they resided in a state of pre-activation and can produce cytokine upon stimulation (Herndler-Brandstetter et al. 2011). In general, the CD8+ subset is more affected by the accumulation of terminally differentiated T cell than the CD4+ compartments with advancing age (Arnold et al. 2011).
10.2.3 The Decreased Output of Regulatory T Cells
Treg, formerly known as suppressor T cells, are a special subset of T lymphocytes of which the phenotype is CD4+CD25+FOXP3+. They are generated from the thymus or from anergized peripheral CD4+ T cells under particular conditions of suboptimal antigen exposure and/or costimulation. Natural Treg cells are positively selected in the cortex through their TCR interactions with self-peptides presented by thymic stromal cells. It is likely that this high-affinity recognition results in signals rendering them anergic and able to produce antiapoptotic molecules which protect them from negative selection and results in an endogenous long-lived population of self-antigen-specific T cells in the periphery (Maggi et al. 2005).
These cells have demonstrated to mediate self-tolerance, modulate the immune response, and abrogate autoimmune disease (Korn et al. 2007).They represent the link between the two arms of the adaptive immune system by regulating the switch between Th1-like cytokine responses (IL-2, INF-γ, and TNF-α) and Th2-like cytokine responses (IL-3, IL-4, IL-5, and IL-10) that, respectively, support T and B cell-mediated immunity (CMI) (i.e., pro- and anti-inflammatory responses). Moreover, these cells seem to also play a crucial role in defense against certain pathogens including C. albicans, C. neoformans, H. pylori, K. pneumoniae, M. tuberculosis, and Staphylococcus (Peck and Mellins 2010; Lages et al. 2008). Whether Treg cell number or functions are altered with age is still controversial (Fulop et al. 2010). Some reports highlighted however that their proportion and activity were increased with advancing age, also contributing to the reduced proliferative capacity of T cells from older adults (Tsaknaridis et al. 2003; Weiskopf et al. 2009). As this is an important issue with still rather few data available, Treg cells activity and effects on other T cell subsets activities in the elderly would be worth addressing (Fulop et al. 2010).
10.3 Progress in Understanding Underlying Mechanisms
10.3.1 Defects in Some Signaling Pathways in CD4+ T Cells
The decrease ability of aged individuals to mount adequate specific antibody response to influenza vaccination (i.e., anti-hemagglutination activity inhibition or HAI) partly results from decrease in naïve T cells (Lang et al. 2010a, b). Concomitant increase in memory/effector T cells (CD8+) (Effros 2007; Herndler-Brandstetter et al. 2011) and loss in CD28 expression (Vallejo 2005; Effros 2007), cytokine production, and T cell proliferation seem also to be affected by signal transduction defects (Sadighi Akha and Miller 2005) particularly in CD4+ (Yu et al. 2012). Recently, Yu et al. have observed that CD4+ memory cells from individuals ≥65 years displayed significant increased and sustained transcription of the dual-specific phosphatase 4 (DUSP4) that shortened expression of CD40L (Yu et al. 2012). The CD40L has been shown to induce cytokine production and costimulate proliferation of activated T cells, and this is accompanied by production of IFN-γ, TNF-α, and IL-2 (Lang et al. 2010a). The capacity of CD4+CD40L− to help B cell proliferation and anti-HA production is reduced (Haynes 2005). Moreover, sustained transcription of DUSP4 also shortened inducible T cell costimulator (ICOS) and decreased production of IL-4, IL-17A, and IL-2 after in vitro activation (Yu et al. 2012). In vivo after influenza vaccination, activated CD4+ T cells from older adults had increased DUSP4 transcription, which inversely correlated with the expression of CD40L, ICOS, and IL-4 (Yu et al. 2012). These findings therefore suggest that increased DUSP4 expression in activated T cells in part accounts for defective adaptive immune responses to influenza vaccination in the elderly. Furthermore, silencing of DUPS4 expression in elderly CD4+ T cells restores their ability to provide helper activity for B cell differentiation and antibody production (Yu et al. 2012).
10.3.2 The Role of the Aged Bone Marrow in Regulating Memory T Cell Functions
The homeostatic maintenance of memory T cells throughout lifetime is tightly regulated and preserves T cell repertoire diversity to combat common and emerging pathogens as well as recall response to booster vaccination (Nikolich-Zugich 2008). It is only recently that researcher has begun to understand how and where memory T cells are maintained and sheltered. In this respect, the bone marrow (BM) and its mesenchymal stromal cells (MSCs) have been paid attention (Herndler-Brandstetter et al. 2011, 2012). MSCs express proteoglycan ligands to CD44 which is present on memory T cells and mediates their local retention. They also produce IL-7 and IL-15 for the homeostatic maintenance of memory T cells (Tokoyoda et al. 2010). IL-7 has been shown to be important for the survival of memory CD4+ T cells (Guimond et al. 2009; Kondrack et al. 2003), and in the BM, memory CD4+ T cells with a Ly6ChiCD62 ligand– phenotype are in close contact with IL-7-producing VCAM1+ stromal cells (Tokoyoda et al. 2009). In contrast, memory CD8+ T cell survival is mostly dependent on IL-15-mediated signaling (Zhang et al. 1998), and the BM has been shown to be a preferred site for IL-15-driven activation and proliferation of memory CD8+ T cells (Becker et al. 2005). In contrast to IL-7, IL-15 is presented by IL-15Rα+ BM cell leading to sustained IL-15-mediated signaling (Schluns et al. 2004). However, lifelong homeostatic turnover of memory T cells may lead to the accumulation of highly differentiated memory T cells in elderly persons (Weinberger et al. 2007; Nikolich-Zugich 2008). In vitro studies indicate that common γ-chain signaling in CD8+CD28+ T cells, in particular, mediated by IL-15, downregulates the expression of the important costimulatory molecule CD28, thereby facilitating the accumulation of CD8+CD28− senescent T cells (Borthwick et al. 2000; Chiu et al. 2006). Thus, it has been proposed that memory T cell, when in contact with MSCs, is suppressed and displays reduced allogenic and mitogenic proliferation, a state of T cell anergy and reduced apoptosis as well as modulated cytokine production (Tokoyoda et al. 2010; Herndler-Brandstetter et al. 2011). Little is however known about the aged BM and its role in regulating the survival and function of memory T cells (Arnold et al. 2011).
In a recent report, Herndler-Brandstetter et al. have observed that the number of CD4+ and CD8+ T cells was maintained during aging into the BM (Herndler-Brandstetter et al. 2011). Similarly to what happens into the blood stream, the composition of this pool is altered with a decline of naïve and an increase in memory T cells. However, in contrast to the peripheral blood, a highly activated CD8+CD28−T cell population, which lacks the late differentiation marker CD57, accumulates in the BM of elderly persons. IL-6 and IL-15, which are both increased in the aged BM, efficiently induce the activation, proliferation, and differentiation of CD8+ T cells in vitro, highlighting a role of these cytokines in the age-dependent accumulation of highly activated CD8+CD28− T cells in the BM (Herndler-Brandstetter et al. 2012). However, these age-related changes do not impair nevertheless the maintenance of a high number of polyfunctional memory CD4+ and CD8+T cells. Effector CD8+ T cells that reside in the BM are in a state of pre-activation and can rapidly express cytokines and CD40L upon stimulations and efficiently induce the production of high-affinity antibodies by B cells (Tokoyoda et al. 2009; Na et al. 2009). Taken together these findings demonstrate that with advancing age a highly activated CD8+CD28− T cell population accumulates in the BM, which is driven by the age-related increase of IL-6 and IL-15. Terminally differentiated effector CD8+ T cells may therefore represent an interesting and important line of defense to pathogens in aging and old adult population and can still fulfil important functions and compensate for the loss of regenerative capacity (Herndler-Brandstetter et al. 2012).
10.3.3 Decreasing of Memory T Cells Reactogenicity
Some interesting information about the reactogenicity of memory T cells in the aged individuals can be learnt from the experience of the herpes zoster (HZ) vaccine. Indeed, the specific VZV CMI is naturally amounted following the primary contact with the virus responsible for chickenpox. While VZV becomes permanently latent in the dorsal-root sensory nerves, the specific CMI contributes to prevent VZV reactivating and acute HZ occurrence (Lang and Michel 2011). It was, however, observed that the age-related reduction in CMI to VZV was the main cause of VZV reactivation in older individuals and in individuals who are immunocompromised as a result of diseases or its treatments (Weinberg et al. 2009; Kimberlin and Whitley 2007). CMI is, however, expected to be restored across the life-span either by subclinical VZV infections occurring periodically and a similar boost in immunocompromised hosts who experience asymptomatic VZV viremia (Lang and Michel 2011) or vaccinating individuals with the licensed HZ vaccine (Levin and Hayward 1996).
Although Oxman et al. in a large randomized controlled trial has demonstrated that the HZ vaccine markedly reduced morbidity from HZ among older adults (Oxman et al. 2005), subsequent analyses have demonstrated that the vaccine-induced boost in CMI was also a function of the age of the vaccine (Weinberg et al. 2009). Furthermore, a loss of CD4+ and CD8+ early effectors and CD4+ effector memory cells was particularly observed among poorer responders (Weinberg et al. 2012). This demonstrates that the vaccine cannot reactivate VZV CMI efficiently enough in older adults. This is moreover reinforced by the recent demonstration that risk of HZ recurrences after a first episode that was initially considered being negligible appears as frequent as rates of the first occurrence in immunocompetent individuals after the age of 50 years (Yawn et al. 2011). Thus, also the natural infection cannot properly reactivate CMI to elicit durable protection in older adult population either.
10.4 Is the Senescence of T Cell-Mediated Immunity a Quantifiable Disorder?
Predicting individual immune responsiveness using biological markers that easily distinguish between healthy and immunosenescent states is a great but desirable challenge. Since the single preceding event in all cases of immunosenescence is thymic involution (Aspinall et al. 2010), the question is: can we identify specific T cell immunity makers linked to a state of immunosenescence? The pioneering OCTO and NONA studies have resulted in the emerging concept of an Immune Risk Profile (IRP) (Strindhall et al. 2007; Wikby et al. 2005, 2008). This immune condition consists of (1) a depleted number of naïve T cells, (2) a high CD8+ and low CD4+ numbers characterized by an inverted CD4+:CD8+ ratio, (3) a poor mitogen response to concanavalin (ConA) stimulation, and (4) the expansion of dysfunctional terminally differentiated CD8+CD28− T cells (i.e., senescent cells) (Brunner et al. 2011; Pawelec et al. 2009). This IRP was identified from healthy octogenarians and nonagenarians and 2-, 4-, and 6-year mortality predicted. Hirokawa et al. have thus proposed a T cell immune score expressing the immune status as a simple score combining five T cell-related parameters (Hirokawa et al. 2009): total number of T cells, CD4+:CD8+ ratio, number of naïve T cells (CD4+CD45RA+), ratio of naïve to memory (CD4+CDRO+) T cells, and T cell proliferative index (TCPI). In patients with colorectal cancer compared to healthy age-matched controls, this T cell immune score of patients in stages I–IV was significantly decreased. Furthermore, the complex remodeling of immune system observed during aging also includes profound modifications within the cytokine network (Larbi et al. 2011). The typical feature of this phenomenon is a general increase in plasma cytokine levels and cell capability to produce proinflammatory cytokines, including a chronic, low-grade, proinflammatory condition usually termed inflamm-aging (Franceschi et al. 2007; Franceschi 2007; Macaulay et al. 2012). This results from a shift from a CD4+ Th cells, Th1-like cytokine response to a Th2-like response, and, furthermore, an increase in levels of proinflammatory cytokines (i.e., IL-6, TNF-α, as well as IL-1β, IL-18, and IL-12). While a wide range of factors have been claimed to contribute to this state (i.e., increased amount of adiposity, decreased production of sex steroid, and chronic health comorbid disorders) (Ostan et al. 2008; Fulop et al. 2010), this altered inflammatory response has also been attributed to the continuous exposure to CMV antigen stimulation and/or reactive oxygen species (ROS) (Pawelec et al. 2009; Larbi et al. 2011; Brunner et al. 2011). However, whether these parameters could provide a robust set of criteria for the determination of an individual’s immunological status in the older old adults, further studies are still required in order to identify biomarkers that are identifiable earlier in life so that intervention strategies can be administered sooner rather than later (Govind et al. 2012).
With this aim, genomic not only may help to identify factors usable as a measure of biological aging but that may also be useful as a tool for predicting immune capabilities within the population (Ostan et al. 2008). Studies that tracked the changes in thymic output have attempted to establish the number of naïve cells and thereby provide an assessment of immune status by using an excisional by-product of TCR genes rearrangement (Govind et al. 2012; Douek et al. 1998, 2000; Mitchell et al. 2010; Hazenberg et al. 2002, 2003). These products are termed TCR-rearrangement excision circles (TRECs) (Kong et al. 1998; Livak and Schatz 1996; Takeshita et al. 1989).
10.5 Could sj-TREC Be Considered as a Biomarker of Effective Aging?
10.5.1 sj-TREC: Episomal DNA Sequences Generated During the TCR Gene Rearrangement
The ability of T lymphocytes to recognize a specific region of a particular antigen is driven by the presence of antigen receptors on the surface of each cell. The TCR is a heterodimer that consists in 95 % of T cells of an alpha (α) and beta (β) chain, whereas in 5 % of T cells this consists of gamma and delta (γ/δ) chains. In order to create a border repertoire of TCR, an intricate process of cutting and splicing undergoes during the complex transition from hematopoietic stem cell to mature naïve T lymphocyte that leads to random joining of DNA segments from the TRC locus (Chain et al. 2005). In T cells expressing TCR-αβ, rearrangements of both TCR-α and TCR-β genes produce TRECs, as depicted in Fig. 10.2, by VJ gene recombination and by V(D)J gene recombination, respectively (Bogue and Roth 1996). Both involve a somewhat random joining of gene segments to generate the complete TCR chain, and the two rearrangement events that occur during this process are identical in 70 % of αβ T cells (Verschuren et al. 1997). The α-chain rearrangement produces a signal-joint TREC (sj-TREC) and the β-chain a coding-joint TREC (Douek et al. 1998). Thus, the TRECs generated are common to most αβ T lymphocytes and are detectable exclusively in phenotypically naïve T cells (i.e., undetectable in memory/effector T cells, B cells, and other peripheral mononuclear cells) (Aspinall and Andrew 2000; Kohler et al. 2005; Hazenberg et al. 2003). Because of the enormous diversity of TCR-α VJ and TCR-β VDJ recombination events (Siu et al. 1984; Arden et al. 1985) and thus the number of TRECs produced, no single TREC can be used as a marker to assess the overall thymic function (Douek et al. 1998; Hazenberg et al. 2003). While α- and β-TRECs possess an identical DNA sequences, respectively, and are both stable (Livak and Schatz 1996), not duplicated during subsequent mitosis (Takeshita et al. 1989), TRECs generated during α-chain rearrangement are generally preferred (Aspinall et al. 2000). Indeed they are generated after β-TRECs and are therefore less diluted out with each subsequent cellular division. Moreover, a common requirement for productive rearrangement of the TCR-α locus is the deletion of the TCR-δ locus (Fig. 10.2). Sj-TREC generated during the α-chain rearrangement can be easily quantified in clinical samples (Douek et al. 2000; Hazenberg et al. 2000, 2002, 2003; Lang et al 2011; Zubakov et al. 2010; Patel et al. 2000; Aspinall et al. 2000; Murray et al. 2003; Kohler et al. 2005).
10.5.2 sj-TREC: A Marker of the Resting Naïve T Cell Pool
Phenotypic analyses have confirmed that the exhaustion of thymic output with advancing age was at the basis of the deficient replacing of naïve T cells lost in the periphery (i.e., by death or conversion to memory/effector cells) (Weiskopf et al. 2009; Haines et al. 2009; Ostan et al. 2008; Kohler et al. 2005). Whether this contributes to the inability of maintaining the T cell repertoire breadth in older adults, TREC values could not be immediately interpreted to reflect continuous thymic output of naïve T cells (Hazenberg et al. 2003). While, as showed in Fig. 10.3, some reports have shown age-associated decline in the sj-TREC values (Mitchell et al. 2010; Zubakov et al. 2010), Chen et al. have demonstrated that TRECs were still readily detectable in healthy nonagenarians (Chen et al. 2010). This suggests, as demonstrated by Hazenberg, that TREC T cells content should be finally more considered as a biomarker of the resting naïve T cell pools rather than a record of thymic output (Hazenberg et al. 2003). This is well illustrated by findings from studies performed in individuals suffering from different health conditions (Douek et al. 1998, 2000; Markert et al. 1999; Patel et al. 2000). Two major biological parameters that complicate the interpretation of TREC data explain this assertion: longevity of naïve T cells and TREC dilution within the two daughter cells after each round of cell division (Hazenberg et al. 2003). Indeed, estimating that healthy adult has a steady state of 1011 naïve T cells and a thymic output of 107–108 naïve cells per day, it was estimated that naïve T cells have a life-span of 1,000–10,000 days (Sprent and Tough 1994). Consistently, thymectomy should not lead to rapid decline in naïve T cell numbers, and in a group of adults thymectomized three to 39 years prior to analysis, TRECs were still clearly present (Douek et al. 1998). It was thus assumed that naïve T cell division would be too low to significantly affect the TREC content (Douek et al. 1998). Whether that is true in healthy adults, it is not the case in human immunodeficiency virus (HIV)-infected individuals or in lymphopenic cancer adults (Hazenberg et al. 2000, 2002). In these two populations TREC values are significantly lower compared to healthy age-matched control, but TREC increased rapidly with highly active antiretroviral therapy (HAART) and during T cell reconstitution with stem cell transplantation, respectively, and even TREC values reached supranormal levels (Hazenberg et al. 2000, 2002). In individuals with severe combined immunodeficiency (SCID) or in congenitally athymic patients (i.e., Di George syndrome), TRECs became detectable after either haematopoietic stem cell transplantation or transplantation of cultured postnatal thymic tissue (Markert et al. 1999; Patel et al. 2000). Finally, in any case, in clinical conditions involving or influencing the cell-mediated immune system or with advancing age, the number of TREC and the T cell TREC content are not only determined by thymic output but also by peripheral events such as homeostatic proliferation of existing naïve T cells which replace those cells lost by death or conversion to memory/effector cells (Hazenberg et al. 2003). Thus, analyzing TREC numbers in healthy individuals, Murray et al. found a marked change in the source of naive T cells before and after 20 years of age (Murray et al. 2003). The bulk of the naive T cell pool was sustained primarily from thymic output for individuals younger than 20 years of age, whereas proliferation within the naïve phenotype was dominant for older individuals. Over 90 % of phenotypically naïve T cells in middle age were not of direct thymic origin.

Schematic representation of the age-related changes in TREC values across the life-span based on Zubakov et al. (2010) and Mitchell et al. (2010) study results. The red line depicts the decline in TREC value in healthy individuals, and the two dashed lines on either side the upper and lower TREC values for a given age observed within this population. Thus, the whole of the figure gives (Adapted from Lang et al. (2013a))
10.6 Could We Identify Different Trends of Aging When Analyzing sj-TREC Values?
A possibly clearer picture the TREC decline in the oldest old was recently shown in a study analyzing blood samples from 215 healthy individuals ranging in age from 60 to 104 years (Mitchell et al. 2010). The number of donors aged ≥70 years were 66 %, and ≥80 years were 27 %. Changes in thymic output were quantified using TREC: 105 T cells ratio. TREC measurements were obtained by quantitative polymerase chain reaction (QPCR), and the number of T cells was determined using fluorescence-activated cell sorter (FACS) analysis. Thus, while the absolute number of leukocytes and T lymphocytes did not change significantly across the age range studied, the authors demonstrated a slowly accelerated curvilinear decline of the TREC ratio between sixth and nineth decade of life. As shown with Fig. 10.4, the most pronounced decline was seen in those individuals more than 90 years of age. Moreover, samples from earlier decades showed a wide range of TREC values with a convergence of the sample heterogeneity observed in the TREC levels with increasing age (Fig. 10.4 –Panel a). These findings contribute to speculate for a number of interesting hypotheses presented in Fig. 10.4 –Panel b. First, are low TREC measurements reflective of an individual’s immunosenescence status? If so, are the individuals in the lower left (LL) quadrant (low TREC level at younger age) at a more advanced stage of immunosenescence? The converse argument could also be inferred for individuals with the highest TREC levels (upper left (UL) quadrant). These individuals may therefore be more likely to progress to become the long-lived healthy individuals observed in the lower right (LR) quadrant. This concept lends itself to the argument that immunosenescence is not merely a measurement of chronological age but points towards immune exhaustion arising at different ages (i.e., physiological age) (Mitchell et al. 2010; Lang et al. 2010c). The downward trajectory of an individual’s thymic output profile over time has been demonstrated previously by Kilpatrick et al. (Kilpatrick et al. 2008) and could be considered as part of longitudinal studies similar to the OCTA and NONA studies to investigate further the potential role of sj-TREC as predictive marker of aging (Strindhall et al. 2007; Wikby et al. 2005, 2008). Interestingly, in one study carried out in old female rhesus macaques, which were vaccinated with inactivated influenza vaccine (strain A/PR/8/34), animals with the higher HAI and best specific T cell proliferation titres against influenza antigen were among those with the highest TREC ratio levels (Aspinall et al. 2007).

Graphic representation of the age-related changes in TREC:105 T cells ratio. (a) (1) The slow decline in the ratio values between the sixth and nineth decades of life with a more pronounced decline seen in individuals more than 90 years of age and (2) a convergence of the sample heterogeneity observed in the TREC levels with increasing age. (b) An annotated diagram of the age-related changes observed in TREC measurement. The dashed horizontal line indicates the median TREC:105 T cell ratios in the sample population and the dashed vertical line the average life expectancy across the study population (79.0 years). UL, LL, UR, and LR quadrants refer to different quadrants formed by the bisection of the data horizontal and vertical lines (Adapted from Lang et al (2013a))
Thus, whether predicting human phenotypes from genotypes is relevant both for personalized medicine and applying preventive strategies (Janssens and van Duijn 2008), additional clinical and translational studies at population, clinical, cellular, and molecular levels are still needed in order to elucidate the exact implications of the TREC values on the age-related senescence of the cell-mediated immune response (Lang et al. 2011). With these perspectives, we have recently developed and optimized a quantitative real-time polymerase chain reaction (qPCR) mono-assay measuring the TREC ratio in dried blood spot (DBS) samples (Lang et al. 2011, 2012b, 2013b). This technology (Fig. 10.5) will be applied on the DBS collected during the 2002–2011 World Health Organization’s (WHO) Study on global AGing and adult health (SAGE) project (World Health Organization 2011). The SAGE project is a longitudinal study conducted in six countries (China, Ghana, India, Mexico, Russia Federation, and South Africa) with a total sample size of nearly 50,000 respondents aged 18 years or over with an special emphasis on the population aged ≥50 years. The objective of the study is to obtain reliable, valid, and comparable health-related and well-being data over a range of key domains for younger and older adult populations in nationally representative samples. The collected data examines health, health-related outcomes and well-being, and their determinants over time. With a first whole data collection completed, a follow up will be conducted every 2 years. SAGE intends to generate large cohorts of older adult populations to be compared with cohorts of younger populations to follow up intermediate outcomes, monitoring trends, examining transitions and life events, and addressing relationships between determinants and health, well-being, and health-related outcomes (http://www.who.int/healthinfo/systems/sage/en/).

Schematic representation of the novel QPCR mono-assay measuring the TREC:T cell ratio in blood samples including DBS samples. This QPCR mono-assay (1) considering DNA samples purified either from PBMCs isolated by a Ficoll-Hypaque density gradient centrifugation or from DBS samples. The prior method combined QPCR measurement for TREC measurement (2) and FACS analysis for the T cell count (3). This figure depicts how the approximation of the VD-J to T cell count is derived with the novel QPCR mono-assay (1). From either whole blood samples or DBS samples, the total cellular DNA is quantified by using the count for ALB2 (albumin gene) considered as a housekeeping gene (A). Similarly, unrearranged TCR-β count is quantified according to the VD-J level; unrearranged TCR-β cells correspond to the total germline cell number (B). Therefore, the unrearranged TCR-β PBMC count subtracted from the total PBMC count gives us the final rearranged TCR-β count and hence the T cell count ((A) − (B)) (Adapted from Lang et al (2011))
10.7 Conclusion
This chapter clearly has presented the main features of T cell-mediated immunity both related to intrinsic defects and its reduced capacity to help B cells proliferation, maturation, and specific antibody production. It has also explored some new insights in the understanding of the mechanisms underlying immunosenescence. Ongoing research in this field is very active (Govind et al. 2012), and this is not only in exploring the process itself but also more and more growing regarding how best to rejuvenate T cell-mediated immunity (Lang et al. 2013a). Nevertheless some robust markers for identifying and grading immunosenescent states and finally distinguishing between healthy and immunosenescent individuals are still and profoundly lacking. Furthermore, although age clearly imposes drastic changes in the immune physiology and contributes to render individuals more prone to develop main age-related diseases and being vulnerable facing of many pathogens, older adults also demonstrate a broad heterogeneity in their health and/or immune phenotypes (Yao et al. 2011; Mitchell et al. 2010) (Fig. 10.1). This finally poses new challenges to scientists. Research on the immunology of aging has to go beyond the simple identification of age-associated immune features.
References
Andrew D, Aspinall R (2002) Age-associated thymic atrophy is linked to a decline in IL-7 production. Exp Gerontol 37:455–463
Arden B, Klotz JL, Siu G et al (1985) Diversity and structure of genes of the alpha family of mouse T-cell antigen receptor. Nature 316:783–787
Arnold CR, Wolf J, Brunner S et al (2011) Gain and loss of T cell subsets in old age–age-related reshaping of the T cell repertoire. J Clin Immunol 31:137–146
Aspinall R, Andrew D (2000) Thymic involution in aging. J Clin Immunol 20:250–256
Aspinall R, Pido J, Andrew D (2000) A simple method for the measurement of sjTREC levels in blood. Mech Ageing Dev 121:59–67
Aspinall R, Pido-Lopez J, Imami N et al (2007) Old rhesus macaques treated with interleukin-7 show increased TREC levels and respond well to influenza vaccination. Rejuvenation Res 10(1):5–17
Aspinall R, Pitts D, Lapenna A et al (2010) Immunity in the elderly: the role of the thymus. J Comp Pathol 142(Suppl 1):S111–S115
Becker TC, Coley SM, Wherry EJ et al (2005) Bone marrow is a preferred site for homeostatic proliferation of memory CD8 T cells. J Immunol 174:1269–1273
Bogue M, Roth DB (1996) Mechanism of V(D)J recombination. Curr Opin Immunol 8:175–180
Borthwick NJ, Lowdell M, Salmon M et al (2000) Loss of CD28 expression on CD8(+) T cells is induced by IL-2 receptor γ-chain signaling cytokines and type I IFN, and increases susceptibility to activation-induced apoptosis. Int Immunol 12:1005–1013
Brunner S, Herndler-Brandstetter D, Weinberger B et al (2011) Persistent viral infections and immune aging. Ageing Res Rev 10:362–369
Chain JL, Joachims ML, Hooker SW et al (2005) Real-time PCR method for the quantitative analysis of human T-cell receptor gamma and beta gene rearrangements. J Immunol Methods 300:12–23
Chen JC, Lim FC, Wu Q et al (2010) Maintenance of naïve CD8 T-cells in nonagerians by leptine, IGFBP3 and T3. Mech Ageing Dev 131:37–38
Chiu WK, Fann M, Weng NP (2006) Generation and growth of CD28nullCD8+ memory T cells mediated by IL-15 and its induced cytokines. J Immunol 177:7802–7810
Cicin-Sain L, Messaoudi I, Park B et al (2007) Dramatic increase in naive T cell turnover is linked to loss of naive T cells from old primates. Proc Natl Acad Sci U S A 104:19960–19965
Douek DC, McFarland RD, Keiser PH et al (1998) Changes in thymic function with age and during the treatment of HIV infection. Nature 396:690–695
Douek DC, Vescio RA, Betts MR et al (2000) Assessment of thymic output in adults after haematopoietic stem-cell transplantation and prediction of T-cell reconstruction. Lancet 355:1875–1878
Effros RB (2007) Role of T lymphocyte replicative senescence in vaccine efficacy. Vaccine 25:599–604
Effros RB, Cai Z, Linton PJ (2003) CD8 T cells and aging. Crit Rev Immunol 23:45–64
Ferrando-Martinez S, Ruiz-Mateos E, Hernandez A et al (2011) Age-related deregulation of naive T cell homeostasis in elderly humans. Age 33:197–207
Franceschi C (2007) Inflammaging as a major characteristic of old people: can it be prevented or cured? Nutr Rev 65:S173–S176
Franceschi C, Capri M, Monti D et al (2007) Inflammaging and anti-inflammaging: a systemic perspective on aging and longevity emerged from studies in humans. Mech Ageing Dev 128:92–105
Frasca D, Diaz A, Romero M et al (2011) Age effects on B cells and humoral immunity in humans. Ageing Res Rev 10:330–335
Fulop T, Larbi A, Witkowski JM et al (2010) Aging, frailty and age-related diseases. Biogerontology 11:547–563
Fulop T, Larbi A, Kotb R et al (2011) Aging, immunity, and cancer. Discov Med 11:537–550
Ginaldi L, Di Benedetto MC, De Martinis M (2005) Osteoporosis, inflammation and ageing. Immun Ageing 2:14
Giunta B, Fernandez F, Nikolic WV et al (2008) Inflammaging as a prodrome to Alzheimer's disease. J Neuroinflammation 5:51
Goronzy JJ, Fulbright JW, Crowson CS et al (2001) Value of immunological markers in predicting responsiveness to influenza vaccination in elderly individuals. J Virol 75:12182–12187
Govind S, Lapenna A, Lang PO et al (2012) Immunotherapy of immunosenescence: who, how and when? Open Longev Sci 6:56–63
Gruener NH, Lechner F, Jung MC et al (2001) Sustained dysfunction of antiviral CD8+ T lymphocytes after infection with hepatitis C virus. J Virol 75:5550–5558
Guimond M, Veenstra RG, Grindler DJ et al (2009) Interleukin 7 signaling in dendritic cells regulates the homeostatic proliferation and niche size of CD4 T cells. Nat Immunol 10:149–157
Gupta S, Gollapudi S (2008) CD95-mediated apoptosis in naïve, central and effector memory subsets of CD4+ and CD8+ T cells in aged humans. Exp Gerontol 43:266–274
Haines CJ, Giffon TD, Lu LS et al (2009) Human CD4+ T cells recent thymic emigrants are identified by protein tyrosine kinase 7 and have reduced immune function. J Exp Med 206:275–285
Haynes LES (2005) The effect of age on the cognate function of CD4+ T cells. Immunol Rev 205:220–228
Hazenberg MD, Stuart JW, Otto SA et al (2000) T-cell division in human immunodeficiency virus (HIV)-1 infection is mainly due to immune activation: a longitudinal analysis in patients before and during highly active antiretroviral therapy (HAART). Blood 95:249–255
Hazenberg MD, Otto SA, de Pauw ES et al (2002) T-cell receptor excision circle and T-cell dynamics after allogeneic stem cell transplantation are related to clinical events. Blood 99:3449–3453
Hazenberg MD, Borghans JAM, Boer RJ et al (2003) Thymic output: a bad TREC record. Nat Immunol 4:97–99
Herndler-Brandstetter D, Landgraf K, Jenewein B et al (2011) Human bone marrow hosts polyfunctional memory CD4+ and CD8+ T cells with close contact to IL-15-producing cells. J Immunol 186:6965–6971
Herndler-Brandstetter D, Landgraf K, Tzankov A et al (2012) The impact of aging on memory T cell phenotype and function in the human bone marrow. J Leukoc Biol 91:197–205
Hirokawa K, Utsuyama M, Ishikawa T et al (2009) Decline of T cell-related immune functions in cancer patients and an attempt to restore then through infusion of activated autologous T cells. Mech Ageing Dev 130:86–91
Hünig T, Lücher F, Elfein K et al (2010) CD28 and IL-4: two heavyweights controlling the balance between immunity and inflammation. Med Microbiol Immunol 199:239–246
Janssens ACJW, van Duijn CM (2008) Genome-based prediction of common diseases: advances and prospects. Hum Mol Genet 17:R166–R173
Jiang Q, Li WQ, Aiello FB et al (2005) Cell biology of IL-7, a key lymphotrophin. Cytokine Growth Factor Rev 16:513–533
Karrer U, Sierro S, Wagner M et al (2003) Memory inflation: continuous accumulation of antiviral CD8+ T cells over time. J Immunol 170:2022–2029
Kelley KW, Weigent DA, Kooijman R (2007) Protein hormones and immunity. Brain Behav Immun 21:384–392
Kilpatrick RD, Rickabaugh T, Hultin LE et al (2008) Homeostasis of the naive CD4+ T cell compartment during aging. J Immunol 180:1499–1507
Kim K, Le CK, Sayers TJ et al (1998) The trophic action of IL-7 on pro-T cells: inhibition of apoptosis of pro-T1, -T2 and -T3 cells correlates with Bcl-2 and Bax levels and is independent of Fas and p53 pathways. J Immunol 160:5735–5741
Kimberlin DW, Whitley RJ (2007) Varicella-zoster vaccine for the prevention of herpes zoster. N Eng J Med 356:1338–1343
Kohler S, Wagner U, Pierer M et al (2005) Post-thymic in vivo proliferation of naive CD4+ T cells constrains the TCR repertoire in healthy human adults. Eur J Immunol 35:1987–1994
Kondrack RM, Harbertson J, Tan JT et al (2003) Interleukin 7 regulates the survival and generation of memory CD4 cells. J Exp Med 198:1797–1806
Kong FK, Chen CL, Cooper M (1998) Thymic function can be accurately monitored by the level of recent T cell emigrants in the circulation. Immunity 8:97–104
Korn T, Oukka M, Kuchroo V et al (2007) Th17 cells: effector cells with inflammatory properties. Semin Immunol 19:362.371
Lages CS, Suffia I, Velilla PA et al (2008) Functional regulatory T cells accumulate in aged hosts and promote chronic infectious disease reactivation. J Immunol 181:1835–1848
Lang PO, Aspinall R (2012) Immunosenescence and herd immunity: with an ever increasing aging population do we need to rethink vaccine schedules? Expert Rev Vaccines 11:167–176
Lang PO, Michel JP (2011) Herpes Zoster vaccine: what are the potential benefits for the ageing and older adults? Eur Geriatr Med 2:134–139
Lang PO, Govind S, Mitchell WA et al (2010a) Influenza vaccine effectiveness in aged individuals: the role played by cell-mediated immunity. Eur Geriatr Med 1:233–238
Lang PO, Govind S, Mitchell WA et al (2010b) Vaccine effectiveness in older individuals: what has been learned from the influenza-vaccine experience. Ageing Res Rev 10:389–395
Lang PO, Mitchell WA, Lapenna A et al (2010c) Immunological pathogenesis of main age-related diseases and frailty: role of immunosenescence. Eur Geriatric Med 1:112–121
Lang PO, Mitchell WA, Govind S et al (2011) Real time-PCR assay estimating the naive T-cell pool in whole blood and dried blood spot samples: Pilot study in young adults. J Immunol Methods 369:133–140
Lang PO, Samaras D, Aspinall R et al (2012a) How important is Vitamin D in preventing infections? Osteoporos Int 24(5):1537–1553
Lang PO, Govind S, Dramé M et al (2012b) Comparison of manual and automated DNA purification for measuring TREC in dried blood spot (DBS) samples with qPCR. J Immunol Methods 384:118–127
Lang PO, Govind S, Aspinall R (2013a) Reversing T cell immunosenescence: why, who, and how. Age 35(3):609–620
Lang PO, Govind S, Dramé M et al (2013b) Measuring the TREC ratio in dried blood spot samples: intra- and inter-filter paper cards reproducibility. J Immunol Methods 389:1–8
Larbi A, Pawelec G, Wong SC et al (2011) Impact of age on T-cell signaling: a general defect or specific alteration? Ageing Res Rev 10:370–378
Levin MJ, Hayward AR (1996) The varicella vaccine. Prevention of herpes zoster. Infect Dis Clin North Am 10:657–675
Livak F, Schatz D (1996) T-cell receptor α locus V(D)J recombination by-products are abundant in thymocytes and mature T-cells. Mol Cell Biol 16:609–618
Lloyd-Sherlock P, McKee M, Ebrahim S et al (2012) Population ageing and health. Lancet 379:1295–1296
Lutz W, Sanderson W, Scherbov S (1997) Doubling of world population unlikely. Nature 387:803–805
Macaulay R, Akbar AN, Henson SM (2012) The role of the T-cell in a aged-related inflammation. Age 35(3):563–572
Maggi E, Cosmi L, Liotta F et al (2005) Thymic regulatory T cells. Autoimmun Rev 4:579–586
Markert ML, Boeck A, Hale LP et al (1999) Transplantation of thymus tissue in complete DiGeorge syndrome. N Eng J Med 341:1180–1189
Mitchell WA, Lang PO, Aspinall R (2010) Tracing thymic output in older individuals. Clin Exp Immunol 161:497–503
Murray JM, Kaufmann GR, Hodgkin PD et al (2003) Naive T cells are maintained by thymic output in early ages but by proliferation without phenotype change after age twenty. Immunol Cell Biol 81:487–495
Na IK, Letsch A, Guerreiro M et al (2009) Human bone marrow as a source to generate CMV-specific CD4+ T cells with multifunctional capacity. J Immunother 32:907–913
Naylor K, Li G, Vallejo AN et al (2005) The influence of age on T cell generation and TCR diversity. J Immunol 174:7446–7452
Nikolich-Zugich J (2008) Ageing and life-long maintenance of T-cell subsets in the face of latent persistent infections. Nat Rev Immunol 8:512–522
Oeppen J, Vaupel JW (2002) Demography. Broken limits to life expectancy. Science 296:1029–1031
Ongrádi J, Kövesdi V (2010) Factors that may impact on immunosenescence: a appraisal. Immun Ageing 7:7
Ortman CL, Dittmar KA, Witte PL et al (2002) Molecular characterization of the mouse involuted thymus: aberrations in expression of transcription regulators in thymocyte and epithelial compartments. Int Immunol 14:813–822
Ostan R, Bucci L, Capril M et al (2008) Immunosenescence and immunogenetics of human longevity. Neuroimmunomodulation 15:224–240
Oxman MN, Levin MJ, Johnson GR et al (2005) A vaccine to prevent herpes zoster and postherpetic neuralgia in older adults. N Eng J Med 352:2271–2284
Pantaleo G, Soudeyns H, Demarest JF et al (1997) Evidence for rapid disappearance of initially expanded HIV-specific CD8+ T cell clones during primary HIV infection. Proc Natl Acad Sci U S A 94:9848–9853
Patel DD, Gooding ME, Parrott RE et al (2000) Thymic function after hematopoietic stem-cell transplantation for the treatment of severe combined immunodeficiency. N Eng J Med 342:1325–1332
Pawelec G, Derhovanessian E, Larbi A et al (2009) Cytomegalovirus and human immunosenescence. Rev Med Virol 19:47–56
Peck A, Mellins ED (2010) Precarious balance: Th17 cells in host defense. Infect Immun 78:32–38
Pfister G, Savino W (2008) Can the immune system still be efficient in the elderly? An immunological and immunoendocrine therapeutic perspective. Neuroimmunomodulation 15:351–364
Pfister G, Weiskopf D, Lazuardi L et al (2006) Naive T cells in the elderly: are they still there? Ann N Y Acad Sci 1067:152–157
Sadighi Akha AA, Miller RA (2005) Signal transduction in the aging immune system. Curr Opin Immunol 17:486–491
Saurwein-Teissl M, Lung TL, Marx F et al (2002) Lack of antibody production following immunization in old age: association with CD8(+)CD28(−) T cell clonal expansions and an imbalance in the production of Th1 and Th2 cytokines. J Immunol 168:5893–5899
Schluns KS, Klonowski KD, Lefrancois L (2004) Transregulation of memory CD8 T-cell proliferation by IL-15R bone marrow-derived cells. Blood 103:988–994
Sempowski GD, Hale LP, Sundy JS et al (2000) Leukemia inhibitory factor, oncostatin M, Il-6 and stem cell factor mRNA expression in human thymus increases with age and is associated with thymic atrophy. J Immunol 164:2180–2187
Sewell AK, Price DA, Oxenius A et al (2000) Cytotoxic T lymphocyte responses to human immunodeficiency virus: control and escape. Stem Cells 18:230–244
Shankar P, Russo M, Harnisch B et al (2000) Impaired function of circulating HIV-specific CD8(+) T cells in chronic human immunodeficiency virus infection. Blood 96:3094–3101
Shaw AC, Panda A, Joshi SR et al (2011) Dysregulation of human Toll-like receptor function in aging. Ageing Res Rev 10(346–353)
Shetty P (2012) Grey matter: ageing in developing countries. Lancet 379:1285–1287
Siu G, Kronenberg M, Strauss E et al (1984) The structure, rearrangement and expression of D beta gene segments of the murine T-cell antigen receptor. Nature 311:344–350
Sprent J, Tough DF (1994) Lymphocyte life-span and memory. Science 265:1395–1400
Strindhall J, Nilsson BO, Lofgren S et al (2007) No Immune Risk Profile among individuals who reach 100 years of age: findings from the Swedish NONA immune longitudinal study. Exp Gerontol 42:753–761
Surh CD, Sprent J (2002) Regulation of naïve and memory T-cell homeostasis. Microbes Infect 4:51–56
Takeshita S, Toda M, Ymagishi H (1989) Excision products of the T cell receptor gene support a progressive rearrangement model of the α/δ locus. EMBO J 8:3261–3270
Thorpe KE, Howard DH (2006) The rise in spending among Medicare beneficiaries: the role of chronic disease prevalence and change in treatment intensity. Health Aff 25:w378–w388
Tokoyoda KZS, Hegazy AN, Albrecht I et al (2009) Professional memory CD4 T lymphocytes preferentially reside and rest in the bone marrow. Immunity 30:721–730
Tokoyoda K, Hauser AE, Nakayama T et al (2010) Organization of immunological memory by bone marrow stroma. Nat Rev Immunol 10:193–200
Tsaknaridis L, Spencer L, Culbertson N et al (2003) Functional assay for human CD4 + CD25+ Treg cells reveals an age- dependent loss of suppressive activity. J Neurosci Res 74:296–308
United Nations (UN) (2008) World population ageing: 1950–2050. http://www.un.org/esa/population/publications/worldageing19502050/
Vallejo AN (2005) CD28 extinction in human T-cells: altered functions and the program of T-cell senescence. Immunol Rev 205:158–169
Verschuren MC, Wolvers-Tettero IL, Breit TM et al (1997) Preferential rearrangements of the T cell receptor-delta-deleting elements in human T cells. J Immunol 159:4341–4349
Virgin HW, Wherry EJ, Ahmed R (2009) Redefining chronic viral infection. Cell 138:30–50
Weinberg A, Zhang JH, Oxman MN et al (2009) Varicella-zoster virus-specific immune responses to herpes zoster in elderly participants in a trial of a clinically effective zoster vaccine. J Infect Dis 200:1068–1076
Weinberg A, Huang S, Song LY et al (2012) Immune correlates of herpes zoster in HIV-infected children and youth. J Virol 86:2878–2881
Weinberger B, Lazuardi L, Weiskirchner I et al (2007) Healthy aging and latent infection with CMV lead to distinct changes in CD8 and CD4 T-cell subsets in the elderly. Hum Immunol 68:86–90
Weiskopf D, Weinberger B, Grubeck-Loebenstein B (2009) The aging of the immune system. Transpl Int 22:1041–1050
Wikby A, Ferguson F, Forsey R et al (2005) An immune risk phenotype, cognitive impairment, and survival in very late life: impact of allostatic load in Swedish octogenarian and nonagenarian humans. J Gerontol A Biol Sci Med Sci 60:556–565
Wikby A, Mansson IA, Johansson B et al (2008) The immune risk profile is associated with age and gender: findings from three Swedish population studies of individuals 20–100 years of age. Biogerontology 9:299–308
World Health Organization (WHO) (2002) Active ageing: a policy framework. http://whqlibdoc.who.int/hq/2002/who_nmh_nph_02.8.pdf
World Health Organization (WHO) (2011) Initiative of vaccine research (IVR) of the Immunization, Vaccines and Biologicals Department and the Ageing and Life Course (ALC) Department. Report on the ad-hoc Consultation on Ageing and Immunization, Geneva
Yao X, Hamilton RG, Weng NP et al (2011) Frailty is associated with impairment of vaccine-induced antibody response and increase in post-vaccination influenza infection in community-dwelling older adults. Vaccine 29:5015–5021
Yawn BP, Wollan PC, Kurland MJ et al (2011) Herpes zoster recurrences more frequent than previously reported. Mayo Clin Proc 86:88–93
Yu M, Li G, Lee WW et al (2012) Signal inhibition by the dual-specific phosphatase 4 impairs T cell-dependent B-cell responses with age. Proc Natl Acad Sci U S A 109:E879–E888
Zhang X, Sun S, Hwang I et al (1998) Potent and selective stimulation of memory-phenotype CD8 T cells in vivo by IL-15. Immunity 8:591–599
Zubakov D, Liu F, van Zelm MC et al (2010) Estimating human age from T-cell DNA rearrangements. Curr Biol 20:R970–R971
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Lang, P.O. (2014). T Cell-Mediated Immunity in the Immunosenescence Process. In: Massoud, A., Rezaei, N. (eds) Immunology of Aging. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-39495-9_10
Download citation
DOI: https://doi.org/10.1007/978-3-642-39495-9_10
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-39494-2
Online ISBN: 978-3-642-39495-9
eBook Packages: MedicineMedicine (R0)