
Abstract
Geobacteraceae, a family within the order Desulfuromonadales, is in this chapter described as consisting of the genus Geobacter and the sole species Pelobacter propionicus. The genus Geobacter comprises anaerobic, non-fermenting chemoorganotrophic mesophiles. Their hallmark feature is the ability to reduce insoluble Fe(III) and Mn(IV) for which they can employ several mechanisms, of which the most notably is extracellular electron transfer via electric conductive nanowires. Geobacter species are physiological versatile. They all oxidize acetate and mainly use small organic acids and alcohols. Several species also oxidize monoaromatic hydrocarbons, such as toluene and benzene. Next to Fe(III), a range of other electron acceptors can be used, and Geobacter species can transfer electron directly to other microbial species or graphite electrodes. Sulfur is often respired, and some species respire organohalides. P. propionicus is phylogenetically located within the Geobacter clade, but several of its physiological characteristics are distinct from Geobacter properties: it can ferment but does not utilize acetate as an electron donor nor oxidizes organic compounds completely. Furthermore, it does not contain the c-type cytochromes that are involved in electron transfer to Fe(III) in Geobacter species. Members of the genus Geobacter often dominate in iron-reducing settings, in particular in environments that have been subject to anthropogenic influences. They represent a rare case of environmental dominant species that are relatively easy to enrich and isolate. Genome-scale metabolic models have strongly contributed to understanding their physiology and ecology. The physiological characteristics of Geobacter species are employed in environmental biotechnology, such as the natural attenuation of organic matter, bioremediation of aromatic hydrocarbons, heavy metals and organohalides, and generating bioenergy in microbial fuel cells and microbial electrolysis cells.
Access provided by Autonomous University of Puebla. Download reference work entry PDF
Similar content being viewed by others


Keywords
- Humic Acid
- Microbial Fuel Cell
- Electron Shuttle
- Microbial Electrolysis Cell
- Extracellular Electron Transfer
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Taxonomy, Historical and Current
Ge.o.bac.ter.a′ce.ae. M.L. masc. n. Geobacter type genus of the family; -aceae ending to denote a family; M.L. fem. Pl. n. Geobacteraceae the Geobacter family. The description is an emended version of the validated description in Bergey’s Manual (Garrity et al. 2005; Validation List 107 2006).
The family Geobacteraceae is located in the order Desulfuromonadales in the δ-subclass of the Proteobacteria. The order branches phylogenetically between the orders Desulfovibrionales and Desulfobacterales. The family Geobacteraceae is currently not consistently described, which has led to erroneous assignments of genera to this family. In this chapter an emended description of the Geobacteraceae by Garrity et al. (2005) will be presented, consisting of one genus, Geobacter, and the species Pelobacter propionicus. The family branches between the family Desulfuromonadaceae (Kuever et al. 2005) and Myxococcales (Fig. 12.1 ).


Maximum likelihood genealogy reconstruction of the 16S rRNA gene sequences of all validly described species (March 2013) in the family Geobacteraceae. Also members of the Desulfuromonadaceae are shown, to indicate that the genera Geothermobacter, Geoalkalibacter, and Geopsychrobacter fall within the Desulfuromonadaceae. The bar indicates 2 % sequence divergence. Type strains for genera are indicated in bold
The current inconsistent assignment of genera and species to the family Geobacteraceae appears to relate to the nearly parallel description of the novel family Geobacteraceae by Holmes et al. (2004a), and by Garrity et al. (2005), with differences in the genera assigned by these authors to the family. Holmes et al. (2004a) described the family Geobacteraceae as consisting of six genera divided over three subclades on basis of phylogenetic analysis of six genes: the genera Geobacter and Trichlorobacter in the Geobacter subclade; the genera Desulfuromonas, Pelobacter, and Malonomonas in the Desulfuromonas subclade; and the genera Desulfuromusa in the Desulfuromusa subclade. 16S-rRNA gene-based evolutionary distances between the Geobacter and Desulfuromonas subclades were observed to be large and generally confirmed by the additional analysis of five other marker genes (Holmes et al. 2004a). Garrity et al. (2005) and Kuever et al. (2005) assigned in Bergey’s Manual the same six genera to two different families, the Geobacteraceae and the Desulfuromonadaceae, with Geobacteraceae (Garrity et al. 2005) comprising members of the genera Geobacter and Trichlorobacter, while the other genera were included in the Desulfuromonadaceae (Kuever et al. 2005). Garrity et al. (2005) described the family Geobacteraceae in also on basis of the phylogenetic analysis of 16S rRNA gene sequences. There appear to be no unique physiological characteristics that distinguish Geobacteraceae from Desulfuromonadaceae, although members of the family Geobacteraceae have only been isolated from anoxic fresh water-related settings, while the Desulfuromonadaceae are more, but not exclusively, associated with marine settings (Garrity et al. 2005; Kuever et al. 2005).
The Geobacteraceae description in Bergey’s Manual (Garrity et al. 2005) is currently (March 2013) most adhered to (e.g., http://www.bacterio.cict.fr/classifgenerafamilies.html#Geobacteraceae; http://www.ncbi.nlm.nih.gov/taxonomy; http://www.arb-silva.de/projects/living-tree/) and largely followed here. The sole species assigned to the genus Trichlorobacter (De Wever et al. 2000) was later renamed to Geobacter thiogenes (Nevin et al. 2007), on basis of its phylogenetic and physiological similarities to Geobacter species. A few new genera, Geothermobacter, Geoalkalibacter, and Geopsychrobacter, are currently assigned to the Geobacteraceae (e.g., see abovementioned webpages); however, based on their 16S rRNA gene phylogeny (Fig. 12.1 ) and phylogeny on basis of other genes (Holmes et al. 2004a), these genera clearly belong to the family Desulfuromonadaceae. Greene et al. (2009) have proposed to amend the description of Desulfuromonadaceae to include also the genus Geoalkalibacter. In this chapter therefore the description of the Geobacteraceae comprises one genus, Geobacter, and the species Pelobacter propionicus. This description is currently also followed by http://eztaxon-e.ezbiocloud.net/. While Pelobacter propionicus belongs to the family Geobacteraceae on basis of its phylogeny of six genes (Holmes et al. 2004a), all other isolates currently described as belonging to the Pelobacter genus are in the Desulfuromonadaceae (Kuever et al. 2005), including the type strain for the Pelobacter genus, Pelobacter acidigallici (Fig. 12.1 ). Therefore, Pelobacter propionicus requires taxonomic reassignment.
All members of the Geobacteraceae are Gram-negative, nonspore-forming rods with rounded ends. They are anaerobic chemoorganotrophic mesophiles able to reduce Fe(III). They mainly use small organic acids and alcohols. They contain genes for nitrogen fixation, which are functional in the strains (G. metallireducens, G. sulfurreducens) that have been tested for nitrogen fixation (Bazylinski et al. 2000; Holmes et al. 2004b). Several characteristics of P. propionicus are distinct from Geobacter properties: it cannot utilize acetate as an electron donor (Schink 1984) or completely oxidize organic acids. Furthermore, although it can reduce Fe(III) (Lonergan et al. 1996), it does not contain the c-type cytochromes that are involved in electron transfer to Fe(III) in Geobacter species (Leang et al. 2003). It has been suggested that the fermentative members in the Geobacteraceae and Desulfuromonadaceae have developed their fermentative metabolism as a secondary evolutionary event (Stackebrandt et al. 1989). P. propionicus clusters among Geobacter species (Fig. 12.1 ).
Molecular Analyses
DNA-DNA Relatedness Studies
Conventional DNA-DNA hybridization has been infrequently used in the taxonomy of Geobacteraceae, even though species delineation based on 16S rRNA gene sequences is often difficult, as species with clearly different physiological characteristics can have more than 97 % identity in their 16S rRNA genes. Hybridization of G. bemidjiensis and G. bremensis yielded a DNA-DNA relatedness value of 63.5 %, below the species threshold of 70 %, while their 16S rRNA genes were 99 % identical (Nevin et al. 2005). Genomic DNAs of G. grbiciae and G. hydrogenophilus were, respectively, 30 and 54 % homologous to G. metallireducens, while they were more than 99 % similar at the 16S rRNA gene level (Coates et al. 2001). G. daltonii revealed 21 % relatedness to its closest relative G. uraniireducens, 16S rRNA gene sequence similarity was 98.1 %, but rpoB similarity was 81.1 % and below the suggested species threshold of 97.7 % (Prakash et al. 2010). Prakash et al. (2010) concluded that DNA-DNA hybridization data are redundant for Geobacter species delineation when molecular data other than rRNA gene sequences indicate low genome similarity.
Genome sequencing can replace DNA-DNA hybridization studies and allow for an estimate for DNA relatedness using regression analysis, which indicated a relatedness of below 20 % between G. daltonii and G. uraniireducens (Prakash et al. 2010). The average nucleotide identity (ANI) was 73.4 %, and the percentage of DNA conserved (>90 % nucleotide identity) only 0.27 % (Prakash et al. 2010). G+C content is standard reported, ranging from 50.2 to 63.8 %, P. propionicus in the middle at 57.8 %.
PCR-Based Analysis of Phylogenetic Markers
Phylogeny of the currently recognized 17 species is mainly based on 16S rRNA gene sequences. However, delineation of Geobacter species solely on basis of 16S rRNA sequences and using the 97 % identity threshold generally employed to delineate novel species (Stackebrandt and Goebel 1994) are not definitive, as several distinct species share greater than 97 % sequence identity (Coates et al. 2001; Nevin et al. 2005; Prakash et al. 2010). Phylogeny of type strains of species of the Geobacteraceae and Desulfuromonadaceae has been studied in large detail by Holmes et al. (2004a) on the basis of six conserved genes: next to 16S rRNA, also nifD, recA, gyrB, rpoB, and fusA gene sequences were obtained. This study included eight Geobacter species and Pelobacter propionicus. The phylogenies derived from the later five genes also clustered Geobacter species together, with P. propionicus falling in the Geobacter cluster. Also gltA, encoding an eukaryote-like citrate synthase, has been used as a marker, since this gene is phylogenetic distinct from those of other prokaryotes and a key enzyme in the central metabolism of Geobacteraceae and Desulfuromonadaceae (Prakash et al. 2010).
Due to their high abundances in iron-reducing environments, members of the Geobacteraceae are already easily encountered by culturing-independent approaches using general, bacteria-specific 16S rRNA gene-directed primers. In order to specifically target and quantify Geobacter species, a number of specific 16S rRNA gene primers have been developed (Cummings et al. 2003; Holmes et al. 2002; Snoeyenbos-West et al. 2000). This work dates from before the family Desulfuromonadaceae was separated from the Geobacteraceae, thus the primers also target the Desulfuromonadaceae. All reported primer sets have their drawbacks, by either also amplifying 16S rRNA genes from other Deltaproteobacteria, such as Syntrophus species (Cummings et al. 2003; Holmes et al. 2002; Snoeyenbos-West et al. 2000), or being too specific and missing dominant members of the Geobacteraceae/Desulfuromonadaceae (Lin et al. 2005). Functional genes are therefore more preferred. Of these, in particular gltA has been applied to samples from metal-reducing environments (Holmes et al. 2005). Published primers do not distinguish fermentative Pelobacter propionicus from iron-reducing Geobacter species. Quantification of functional genes unique to Geobacter species, such as the gene encoding the multicopper protein OmpB which is required for Fe(III) oxide reduction, may allow for specific targeting of iron-reducing Geobacter species (Holmes et al. 2008).
Rep-PCR has been reported for G. lovleyi and G. thiogenes (Sung et al. 2006).
Genome Comparison
The complete genome sequences of five validly described Geobacter species and Pelobacter propionicus have been reported. Key characteristics are summarized in Table 12.1 . Genome length ranges from 3.8 to 4.6 Mb. The number of 16S rRNA genes varies from one to four but is generally two. Between 3,434 and 4,023, coding genes have been deduced. A phylogeny of the family was constructed using 697 protein families that had a single ortholog in all sequenced genomes. The resulting phylogeny was in line with the 16S rRNA gene phylogeny (Butler et al. 2010).
The genes for acetate transport and its oxidation via the TCA cycle and for proton transport and energy generation across the inner membrane are well conserved in Geobacter species (Butler et al. 2010). Predicted pathways for use of other electron donors can vary between species, as for instance for propionate (Aklujkar et al. 2010). Orthologs to the four subunits of the uptake hydrogenase were only found in those species capable of hydrogen oxidation (Butler et al. 2010). Unexpected capacity for the use of carbon sources has been revealed: transporters for glucose were found in the genome of G. bemidjiensis, and its ability to grow on glucose was subsequently confirmed experimentally (Aklujkar et al. 2010). The central metabolism of G. metallireducens contains several energy-inefficient reactions that are not present in G. sulfurreducens and may aid it to improve flux for rapid energy generation when growing with complex substrates such as benzoate (Sun et al. 2009).
Although the reduction of insoluble Fe(III) is an ubiquitous feature of Geobacter species and occurs on the outer membrane, intriguingly, most (86 %) of the cytochromes for Fe(III) reduction are not conserved, including many of the outer-membrane cytochromes (Butler et al. 2010). While poorly conserved, cytochromes are very abundant in all Geobacter genomes, comprising about 2 % of all genes. Also their diversity is large, with 472 different predicted cytochromes encountered so far. Cytochrome duplication and divergence appear to play a role in these genotypes, while lateral gene transfer has a minor role at most as only 19 of the 472 predicted cytochromes have been identified as lateral gene transfer candidates. This abundance and diversity of weakly conserved cytochromes is postulated to indicate that they are important in their heme-bearing capacity, as sinks for electrons between the inner-membrane transport chain and the extracellular acceptor, and they may not be specific terminal reductases (Butler et al. 2010). Deletion studies confirmed that the outer c-type cytochromes involved in Fe(III) oxide reduction are different between G. sulfurreducens and G. metallireducens (Smith et al. 2013).
The genome of G. sulfurreducens provides evidence for aerobic metabolism (Methe et al. 2003), in line with its reported capability to withstand and even grow with oxygen (Lin et al. 2004). Several genomic features indicate that also G. bemidjiensis has enhanced abilities to respire, detoxify, and avoid oxygen (Aklujkar et al. 2010). Key genes related to oxygen tolerance and reactive oxygen species detoxification are absent from G. lovleyi, which does not tolerate oxygen (Wagner et al. 2012).
Geobacter species encode enzymes for the acetyl-coA pathway, which is employed to assimilate carbon dioxide (Methe et al. 2003). G. bemidjiensis is possibly capable of carbon dioxide fixation via citrate lyase (Aklujkar et al. 2010). Whereas most Geobacter species are expected to obtain reducing equivalents for biosynthesis from electron transfer pathways via a ferredoxin oxidoreductase, G. metallireducens derives them from the oxidative pentose phosphate pathway (Aklujkar et al. 2009). Genome-scale models combined with experiments revealed that Geobacteraceae have an uncommon isoleucine biosynthesis route, via citramalate as intermediate and with acetyl-coA and pyruvate are precursors (Tang et al. 2007).
Geobacter species encode a remarkable large number of genes for two-component signaling proteins (Aklujkar et al. 2009, 2010; Methe et al. 2003), suggesting a large adaptive potential (Lovley et al. 2011). They also contain multiple chemotaxis systems or homologs of the chemotaxis system and an unusually large number of chemoreceptor genes (Tran et al. 2008). One chemotaxis(−like) system is predicted to be involved in flagellar motility and appears to be unique for Geobacter species. The large numbers of methyl-accepting chemotaxis proteins homologs may enable the sensing of a great variety of environmental signals.
Comparative genomic analysis of P. propionicus, G. metallireducens, and G. sulfurreducens revealed several common features. All tricarboxylic acid cycle reactions are present, even though P. propionicus does not completely oxidize organic electron acceptors to carbon dioxide while Geobacter species can (Sun et al. 2010). Unique reactions for P. propionicus are methylmalonyl-coA-dependent propionate formation, corresponding to its propionate formation, and a citrate lyase reaction.
Several features that are unique for individual Geobacter species have been obtained by lateral acquisition and are located on genomic islands. G. metallireducens contains a 300 kb island predicted to encode enzymes for the degradation of many aromatic compounds except toluene; the toluene degradation genes are encoded by a separate island (Butler et al. 2007). G. lovleyi is the only Geobacter species known to respire the organohalide perchloroethene (PCE). Based on GC content and the codon adaptation index, its duplicated reductive dehalogenase (RDase) genes are on a genomic island that has recently been obtained from an ancestral gene cluster (Wagner et al. 2012). Gram-positive Desulfitobacterium also appear to have obtained its RDase genes from this cluster. A second genomic island in the G. lovleyi genome contains a predicted F-factor conjugative pilus tra-gen cluster, which does not have homologs in other sequenced Geobacter genomes. Cobalamin is required for organohalide respiration. Surprisingly, 15 of the 24 genes needed for de novo cobalamin biosynthesis are located on a 77 kb plasmid. However, newly isolated PCE-respiring strains did not carry a similar plasmid, indicating that this plasmid is not a requirement for PCE respiration in this species (Wagner et al. 2012).
The genomes of the G. sulfurreducens strains PCA and KN400 have been compared (Butler et al. 2012). The faster reduction of insoluble Fe(III) and higher current production by strain KN400 does not appear to be due to novel gene acquisition but to changes in the common metabolic network. There is significant enrichment for single-nucleotide polymorphisms in noncoding or synonymous amino acid sites, suggesting selective pressure since the divergence of the two strains. Substantial sequences differences occur especially in cytochromes and integral membrane proteins (Butler et al. 2012).
Phenotypic Analyses
The main features of members of the Geobacteraceae are shown in Tables 12.2 and 12.3 .
Geobacter (Lovley et al. 1993)
The type strain of Geobacter (Ge.o.bac′ter; Gr n. ge the earth; masc. bacter equivalent of Gr. Nl. Bacterion a small rod; M.L masc. n. Geobacter a rod from the earth) is Geobacter metallireducens. Sixteen species have so far been validly described. Geobacter cells are rod shaped with rounded ends and grow as single cells, in pairs or in chains. They are catalase and carotenoid negative. Cells contain menaquinone, mainly the MK8 type, and c cytochromes. Cytochromes cause pink-colored colonies on plate. They are diverse in their use of electron acceptors and donors (Table 12.3 ). Geobacter species are mesophiles, although G. psychrophilus can grow at 4 °C, and prefer circumneutral pH and low salt concentrations. Physiology is influenced by culturing conditions, with for instance the type of Fe(III) used affecting substrate utilization and motility (Childers et al. 2002; Straub et al. 1998). A key phenotypic characteristic of Geobacter species is the conservation of energy from the oxidation of organic compounds to carbon dioxide with Fe(III), Mn(IV), and graphite electrodes as the electron acceptor. All Geobacter species contain the nifH gene (Holmes et al. 2004a), and nitrogen fixation has been reported for G. metallireducens (Bazylinski et al. 2000) and G. sulfurreducens (Holmes et al. 2004b).
Metal Reduction and Other Electron Acceptors
A hallmark physiological feature of Geobacter species is their capability to directly reduce insoluble Fe(III) forms, with the accessibility relating to the surface area (Yan et al. 2008). Poorly crystalline Fe(III) hydroxides and structural Fe(III) of phyllosilicates are well reducible, while crystalline Fe(III) are poorly accessible (Shelobolina et al. 2004). Geobacter species produce electric, metallic-like conductive pili (Malvankar et al. 2011; Reguera et al. 2005) that extracellular transfer electrons to insoluble Fe(III) forms. C-cytochromes are likely important in the transfer of electrons from the outer membrane to the pili and from the pili to Fe(III) but not for electron transport along the pili (Leang et al. 2010; Malvankar et al. 2011; Shi et al. 2007). G. metallireducens can access insoluble Fe(III) and Mn(IV) by chemotaxis towards their reduced products (Childers et al. 2002), for which it uses flagella and pili.
Most Geobacter species also reduce quinone moieties in natural and artificial humic acids (AQDS), which function as extracellular electron shuttles and transfer abiotically their electrons to insoluble Fe(III) compounds, including crystalline Fe(III) oxides (Bauer and Kappler 2009; Scott et al. 1998). The small size of humic acids enables the reduction of Fe(III) that is not physically accessible by the microbes. Cysteine can also be utilized as electron shuttle (Doong and Schink 2002). Geobacter species grow faster with dissolved, chelated Fe(III), or the availability of electron shuttles; however, they have not been found to produce metabolic energy-expensive chelators or electron shuttles themselves, in contrast to other iron reducers (Nevin and Lovley 2000). Extracellular magnetite can be produced during iron reduction, as well as other Fe(II)-containing minerals.
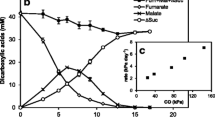
Geobacter species are versatile in the use of electron acceptors (Table 12.3 ). Most species can also reduce fumarate, malate, sulfur, and heavy metals, such as U(VI), and are electricigens, able to transfer electrons to electrodes. Nitrate, when used, is reduced to ammonia. Nitrite, sulfate, sulfite, thiosulfite, selenate, or selenite reduction has never been observed. Organohalide respiration has been observed for two species: G. thiogenes reduces trichloroacetate (TCA) while G. lovleyi respires per- and trichloroethene (PCE, TCE). Dechlorination can occur in the presence of alternative electron acceptors (Sung et al. 2006). De Wever et al. (2000) postulated a sulfur-sulfide cycle to be involved in TCA reduction, but this mechanism was not observed for PCE reduction by G. lovleyi (Sung et al. 2006).
While Geobacter species were considered to be strict anaerobes, at least G. sulfurreducens is capable of growth in the presence of 10 % O2, and it tolerates atmospheric oxygen for at least 24 h (Lin et al. 2004). The aerobic metabolism is in line with genomic information (Methe et al. 2003).
Organic and Inorganic Electron Donors
Acetate is the only carbon source found to be used by all Geobacter species, but a large number of carbon sources can be mineralized to carbon dioxide. Differentiating substrate utilization characteristics are shown in Table 12.3 . C1-C5 organic acids and C2-C4 alcohols are frequently utilized, with some species using nearly all of these substrates (e.g., G. bremensis) while others consume only a few (e.g., G. sulfurreducens, G. thiogenes). Species that used iso-propionate or/and iso-butyrate also use propionate or/and butyrate, respectively.
Also aromatic compounds are used by several species, with benzoate utilization most frequently observed. Toluene degradation has been observed for four species, two of these species are also able to degrade benzene: G. metallireducens (Zhang et al. 2012) and G. daltonii (Kostka, personal communication). Other environmental pollutants that are oxidized by some Geobacter species are phenol, p-cresol, and m-cresol; no isolates are known to degrade ethylbenzene or xylene. Species that use toluene also degrade benzoate, benzaldehyde, and benzyl alcohol and have been found to use 4-hydroxy benzaldehyde, 4-hydroxy benzyl alcohol, and 4-hydroxybenzoate.
G. bemidjiensis is the sole Geobacter species known to degrade the carbohydrate glucose; a property predicted from genome analysis and then experimentally confirmed (Aklujkar et al. 2010). Geobacter species do not ferment but can disproportionate fumarate (Aklujkar et al. 2010).
Geobacter species have not been found to use as sole source of carbon: amino acids, polyaromatic hydrocarbons, alkanes, palmitate, citrate, caproate, caprylate, and intermediates of aerobic aromate degradation such as ferulate, syringate and salicylic acid, and the sugars fructose and mannitol.
Geobacter species can also utilize inorganic electron donors. Many species grow with hydrogen, in the presence of organic carbon (Table 12.3 ). G. metallireducens can also oxidize Fe(II) and U(IV) or reduced humic acids with nitrate or fumarate as an electron acceptor, although it is unclear if energy conservation to support growth occurs with these electron donors (Finneran et al. 2002; Lovley and Blunt-Harris 1999; Weber et al. 2006). Sulfur oxidation has not been observed.
Chemotaxonomic Properties
G. metallireducens, G. sulfurreducens, and G. bemidjiensis have been investigated in a single study for their polar lipid fatty acids (PFLAs), lipopolysaccharide (LPS) hydroxyl fatty acids, and respiratory quinones. Lipid profiles varied with strains and less with electron acceptor (Hedrick et al. 2009). Variation in culturing conditions influences PFLA profiles, complicating comparisons between different studies for the same species (Hedrick et al. 2009; Lovley et al. 1993; Shelobolina et al. 2008). The most abundant PFLAs are 14:0, i15:0, 16:1ϖ7c, 16:1ϖ5c, and 16:0. LPS hydroxyl fatty acids were more variable between species and electron acceptors than PFLAs. They were dominated by 3oh14:0, unusual high levels of 3oh16:0, and the uncommon fatty acids 9oh16:0 and 10oh16:0; the latter may be used as biomarkers for Geobacter. G. metallireducens and G. sulfurreducens contain a range of hopanoid lipids, in particular squalene, diploptene, and bishomohopan-32-ol acetate. Their quantities were highest in G. metallireducens. These biomarkers were long used as indicators for ancient oxic environments (Hartner et al. 2005).
Menaquinone-8 is the most abundant respiratory quinone, constituting about 85 % of the total. Ubiquinones have not been detected (Hedrick et al. 2009). Reduced cytochrome c spectra show absorption maxima at 419–424, 521–524 and 551–555 nm.
Pelobacter propionicus (Schink 1984)
Three Gram-negative, nonspore-forming, rod-shaped Pelobacter propionicus strains were isolated by Schink (1984). Several features are contrasting those of Geobacter species (Table 12.2 ). P. propionicus is capable of fermentation of 2,3-butanediol, acetoin, and acetate and variable in the fermentation of diacetyl, pyruvate, ethanol, propanol, and butanol. Propanol and butanol are fermented to the corresponding fatty acid with concomitant reduction of acetate and carbonate to propionate (Schink 1984). In contrast to Geobacter species and despite the possession of the tricarboxylic acid cycle (Sun et al. 2010), it produces acetate and propionate as fermentation products. Its growth requires carbonate and a reductant. It does not express cytochrome c (Schink 1984). Besides fermentation, it can use chelated Fe(III) and sulfur as electron acceptor with lactate as electron donor (Lonergan et al. 1996) but not nitrate. Electron acceptors that are also not used by Geobacter are also not used by P. propionicus. The temperature range (4–45 °C) over which P. propionicus can grow is wider than reported for any Geobacter species.
Enrichment, Isolation and Maintenance Procedures
Culturing Geobacteraceae
Geobacter species are relatively easy to enrich and isolate, although it should be noted that some in situ dominant Geobacter phylotypes have resisted isolation (Botton et al. 2007; Lin et al. 2007b; Rooney-Varga et al. 1999). Most studies describing the enrichment and/or isolation of Geobacter species were not specifically aiming for Geobacteraceae but for iron-reducing microorganisms in general. For a more specific enrichment and isolation of Geobacter species, an anaerobic, liquid mineral medium containing acetate as sole source of carbon and amorphous Fe(III) as electron acceptor is recommended. Acetate is a substrate used by all Geobacter members, and only a few other iron reducers (e.g., Rhodoferax metallireducens) (Finneran et al. 2003) apart from members of the Desulfuromonadaceae. It should be noted, however, that P. propionicus does not grow with acetate. The use of amorphous or poorly crystalline Fe(III) as electron acceptor is advised, even though the preparation of amorphous Fe(III) is more laborious and faster growth is observed when chelated Fe(III) is used. The use of iron chelators, such as citrate or nitrilotriacetic acid (NTA), bears the risk that microorganisms that ferment the chelator may be enriched and use Fe(III) non-dissimilatory (Caccavo et al. 1994). If chelators are to be used, Fe(III)NTA is preferred. A dilution-to-extinction approach (comparable to a most probable number approach) is recommended to enrich for the most dominant iron reducers. Sample dilution and to a lesser extent the type of Fe(III) affects the type of iron-reducing consortia that will grow (Lin et al. 2007a, b).
Although iron reducers grow in liquid cultures to cell densities that are too low to be seen by eye, iron reduction itself can easily be observed in enrichments. Upon reduction of insoluble Fe(III), its brown color will start to disappear. A black color, due to magnetite formation, may appear over time. A color change from orange/brownish to colorless will be observed in media with soluble, chelated Fe(III). Frequently, the formation of Fe(II)-containing precipitates will occur. This formation depends on the type of medium, especially on the buffering agent employed. For instance, white precipitates (FeCO3) may occur in carbonate buffered media. The color change depends also on the amount of Fe(III) and acetate employed. Complete oxidation of one molecule of acetate requires eight molecules of Fe(III). An excess of acetate in the medium will enable complete iron reduction but also bears the risk of growth of acetoclastic methanogens on the remaining acetate in enrichments.
Soon after the observation of iron reduction, positive enrichments must be transferred a couple of times to fresh medium at 1–10 % (v/v), before strain isolation is attempted. The transfers are needed to remove compounds (e.g., sediment particles, organics) associated with the original inoculum. For strain isolation, the enrichments need to be tenfold serially diluted into a (semi)solid medium; after incubation, single colonies can be picked with sterile glass Pasteur pipettes in an anaerobic glove box and subcultured on solid or liquid medium. Insoluble Fe(III) can easily be employed in liquid media but is more difficult to use in (semi)solid media where chelated Fe(III) is preferred over the use of other soluble electron acceptors, such as nitrate, which are not used by all Geobacteraceae.
Strict anaerobic conditions must be employed in the preparation of media and handling of Geobacteraceae. The addition of oxygen scavengers aids growth. Commonly used oxygen scavengers for anaerobic culturing, such as sulfide, can react abiotically with Fe(III). Fe(II), added to a final concentration of 0.5 mM FeCl2, is a more appropriate oxygen scavenger. Anaerobic conditions are more easily maintained in liquid cultures than in solid media. Incubations should be performed in the dark. Vitamins are generally not required for Geobacter growth but enhance growth (Coates and Lovley 2005). Genome analysis and subsequent experimental validation revealed that Geobacter bemidjiensis is deficient in 4-aminobenzoate synthesis (Aklujkar et al. 2010).
A frequently employed medium to enrich, isolate, and study Geobacter species is a freshwater Fe(III) oxide basal medium (Lovley and Phillips 1988), containing per liter:
-
30–100 mM amorphous Fe(III)oxide or 5–10 mM Fe(III)NTA
-
1.36 g Na-acetate (10 mM)
-
1.5 g NH4Cl
-
2.5 g NaHCO3
-
0.6 g NaH2PO4.H2O
-
0.1 g KCl
-
0.1 g CaCl2.2H2O
-
10 mL vitamin solution according to DSMZ medium 141
-
10 mL trace element solution, prepared according to DSMZ medium 141
The medium is dispensed in serum bottles or tubes and sparged with 80–90 % N2 and 20–10 % CO2, capped with butyl rubber stoppers (Viton stoppers in case of PCE or monoaromatic hydrocarbons as carbon source), and sterilized. 1.5–2.0 % bacteriological agar is added for solid medium. Fe(III) is generally added after autoclaving and cooling down of the medium.
Amorphous, poorly crystalline Fe(III) is prepared by dissolving FeCl3·6H2O in 400 mL water to a concentration of 0.4 M. Subsequently, the pH must be carefully adjusted to pH 7.0 by slowly and dropwise adding a 10 M NaOH solution to the stirred Fe(III) solution. A (temporarily) too high pH may result in the formation of less bioavailable Fe(III) forms. Once pH 7.0 is reached, the solution must be stirred for an additional 30 min and pH further adjusted, when needed. The suspension is then centrifuged for 15 min at 5,000 rpm, and the supernatant is discarded. The Fe(III)oxide is resuspended in water, and the procedure of centrifugation and resuspension is repeated six times, to obtain chloride-free Fe(III) oxide. In the final step, the Fe(III)oxide is dissolved in 100 mL, to a concentration of ∼1 mol per liter. It is advisable to determine the Fe(III) content, using ferrozine (Viollier et al. 2000). Autoclaving this solution is not recommended, and filter sterilization is not possible: an alternative is to prepare an acidic 6-line ferrihydrite solution (Schwertmann and Cornell 1991) which can be filter sterilized at concentrations of 25–75 mM.
A 100 mM stock solution of Fe(III)NTA is prepared by dissolving 1.64 g NaHCO3 in 80 mL water, followed by adding 2.56 g sodium nitrilotriacetic acid and then 2.7 g FeCl3.6H2O. After bringing the volume to 100 mL, the solution must be sparged with an 80–90 % N2 and 20–10 % CO2 gas mixture and filter sterilized into an anaerobic, sterile serum bottle.
The addition of tungsten (0.00025 g/L NaWO4 · 2H2O) to the medium is recommended to aid the enrichment and isolation of aromatic compound degrading Geobacter species: obligate anaerobic microorganisms employ a tungsten-containing class of benzoyl-coenzyme A reductases to open the ring of benzoyl-coA, a central intermediate in aromatic degradation (Kung et al. 2009).
A second medium for culturing iron reducers is based on the medium used for the cultivation of sulfate-reducing bacteria (Widdel and Pfennig 1982), with Fe(III) replacing NaSO4. The medium of Widdel and Pfennig is also used to enrich for fermentative Pelobacter, in which case the NaSO4 is omitted.
Maintenance
Geobacter cultures can be maintained as frozen stocks at −70 °C, after growth to mid-exponential phase. The stocks can be prepared in by transferring aliquots (1–5 mL) to small anaerobic serum vials (10 mL), to which an anaerobic solution of glycerol is added to final concentrations of 10 %. After mixing, the vial is immediately frozen at −70 °C. The frozen stocks should be checked regularly for viability. The cultures can also be stored at 4 °C but then need to be transferred every 4 weeks (Coates and Lovley 2005).
Ecology
A large number of culturing-based studies, and in particular molecular analyses, which avoid cultivation bias, have revealed that Geobacteraceae are of large global biogeochemical significance (reviewed in Lovley et al. 2011). They frequently dominate in soils and sediments in which Fe(III) reduction is an important process, such as aquatic sediments, wetlands, rice paddies, and the subsurface. Geobacter species are especially present in relative high numbers in iron-reducing environments subjected to anthropogenic increases in organic matter but are certainly not only limited to these environments (reviewed by Lovley et al. 2011). In wetland sediments, Geobacter species may contribute significantly to anaerobic redox cycling of iron by performing both dissimilatory Fe(III) reduction and oxidation of Fe(II) with nitrate (Weber et al. 2006), although other microorganisms might be more important in the Fe(II) oxidation (Coby et al. 2011). A culturing-based study indicated that Geobacteraceae might also be important in the reduction of electron-shuttling humic acids (Coates et al. 1998). Geobacter species have mainly been observed in environments with circumneutral pH but also have been encountered in acidic springs, peat or sediments (Adams et al. 2007; Kusel et al. 2008; Percent et al. 2008). Geobacter species are also frequently abundant on the surface of electrodes harvesting electricity from organic compounds in water or sediments (reviewed in Lovley et al. 2011).
Geobacter species have a number of properties that contribute to their high abundances in iron-reducing settings: their ability to converse energy from the oxidation of the key organic intermediate in anaerobic fermentation (acetate) coupled to Fe(III) or Mn(IV), chemotaxis towards iron (Childers et al. 2002), the use of nanowires to access iron via extracellular electron transfer (Reguera et al. 2005), capacity to fix nitrogen (Bazylinski et al. 2000), and a very low-maintenance energy (Lin et al. 2009). Furthermore, the high number of genes for the two-component signal proteins suggest a very large adaptive potential (Lovley et al. 2011). The relative importance of these characteristics varies with the type of environment in which they thrive. Geobacter species may become dominant in acetate-amended, nitrogen-limited environments due to their ability to fix nitrogen (Zhuang et al. 2011) but are also dominant in ammonium-rich landfill leachate plumes (Röling et al. 2001), where more likely the low-maintenance energy is of importance (Lin et al. 2009).
Geobacter species can have substantial geochemical impacts in anaerobic soils and sediments. These include the release of soluble Fe(II) from insoluble Fe(III) into groundwater, accompanied by trace metals, metalloids, and phosphates, which may have secondary effects such as stimulation of microbial growth and generation of groundwater that is unsuitable as drinking water. Also changes in soil porosity can occur, and iron reduction may lead to the precipitation of Fe(II)-containing minerals.
Besides their importance in metal reduction, Geobacteraceae may also be important as fermenting microorganisms in interspecies electron transfer under nonmetal-reducing conditions. Syntrophic interactions of Geobacter species with other microbial species via extracellular transfer of electrons to them via metabolites or with humic acid, cysteine, or sulfide as electron shuttles have been shown in laboratory cocultures with acetate or toluene as the source of carbon (Cord-Ruwisch et al. 1998; Kaden et al. 2002; Meckenstock 1999). Surprisingly, while syntrophic interactions would be expected for the fermenting P. propionicus, when this species was cocultured with organisms that could potentially scavenge metabolites excreted by P. propionicus, its biomass concentration did not increase (Schink 1984). Recently, Geobacter species were found to be also capable to transfer electrons directly to syntrophic partners (Summers et al. 2010). Geobacter species can also receive electrons directly from fermenting partners (Summers et al. 2010), although this ability also depends on the partner microorganism (Rotaru et al. 2012). The environmental significance of direct interspecies electron transfer still has to be shown. Molecular analyses revealed that Geobacter and closely related species can constitute 20 % or more of cell numbers in methanogenic aggregates in anaerobic digesters that treat organic brewery waste water (Morita et al. 2011). This suggests that direct interspecies electron transfer can be important in such methanogenic settings.
The availability of genome sequences has enabled an iterative genome-scale modeling and experimental approach that resulted in an in-depth understanding of the central metabolism and ecology of Geobacter species (Mahadevan et al. 2011).
Pathogenicity, Clinical Relevance
While pathogenic Geobacteraceae have not been described, the activity of Geobacteraceae may still have major impacts on human health. Upon reduction of subsurface Fe(III) oxides by Geobacter species, also toxic heavy metals can be released into groundwater, which is in many countries a major source of drinking water. A major groundwater pollutant is arsenic, constituting a major toxic hazard to millions of people worldwide, especially East Asia. Microbial communities in microcosms containing As-mobilizing sediments from West Bengal were dominated by Geobacter species. Arsenic release occurred after iron reduction and was limited by the availability of electron donor (Islam et al. 2004).
Growth of G. sulfurreducens is inhibited by chloramphenicol (10 mg/mL), nalidixic acid (10 mg/mL), tetracycline (10 mg/mL), kanamycin (200 mg/mL), spectinomycin (50 mg/mL), streptomycin (400 mg/mL), and ampicillin (400 mg/mL) (Coppi et al. 2001).
Application
Besides being of global ecological significance, members of the Geobacteraceae have found utility in a large variety of environmental biotechnological applications. They play major roles in bioremediation and in recent years are also employed to harvest electricity from organic wastes and aquatic sediments. Geobacter species are generally not purposely applied in environmental biotechnology but become dominant in microbial communities due to the combination of the applied treatment and the physiological characteristics of Geobacteraceae, in particular their ability to use acetate and to make electrical contact with extracellular electron acceptors.
Natural Attenuation of Hydrocarbons and Organic Compounds
Iron is the fourth element on Earth and an important component of subsurface sediments. When subsurface becomes polluted with organic compounds and hydrocarbons, e.g., due to petroleum spills, highway runoff of acetate-containing deicing compounds or after disposal on unlined landfills, these pollutants are often disappearing under iron-reducing conditions, without requiring human intervention (e.g., van Breukelen et al. 2003). Geobacter species are frequently dominant in these settings and present at relative higher abundances than in control samples (Holmes et al. 2007; Lin et al. 2005; Röling et al. 2001; Rooney-Varga et al. 1999; Winderl et al. 2008).
Important groundwater pollutants are the monoaromates benzene, toluene, ethylbenzene, and xylene (BTEX), as they dissolve relatively well and are carcinogenic. BTEX were long considered to be degradable under iron-reducing conditions solely by Geobacter species, based on enrichments (e.g., Botton et al. 2007), and properties of isolated species (Table 12.3 ). In recent years, several species outside the Geobacteraceae family have also been found to be capable of degradation of toluene and benzene coupled to iron reduction (Kunapuli et al. 2007, 2010; Weelink et al. 2009). Analysis of marker genes of anaerobic BTEX degradation has suggested that these non-Geobacter species may play specific, important roles in anaerobic monoaromatic degradation in some iron-reducing settings, next to Geobacter species (Pilloni et al. 2011; Staats et al. 2011).
Active Bioremediation of Hydrocarbons
Several strategies aiming to stimulate aromatic hydrocarbon degradation by Geobacter species have been designed and successfully tested. These approaches are especially directed at enhancing the availability of Fe(III) present in solid Fe(III) forms for reduction, by the addition of chelators (Lovley et al. 1994, 1996b), or introduction of humic acids as electron shuttles (Lovley et al. 1996a). Nanoparticles of iron oxides are better reducible than their larger counter partners. Their small size may allow their injection into polluted aquifers, to stimulate iron reduction in aquifers with low amounts of bioavailable iron (Bosch et al. 2010). Inserting electrodes as in situ electron acceptor provides an interesting alternative (Zhang et al. 2010). Lovley (2011) proposed the concept of “subsurface snorkels” with graphite rods functioning in contaminated anaerobic sediments, extending into aerobic soil or water. The part of the rod in the anaerobic soil functions as the anode, while the portion of the rod in the aerobic environments above comprises the cathode.
Active Bioremediation of Heavy Metals
Bioremediation has been used to reductively precipitate heavy metals in the subsurface, with a main focus on U(VI). Removal of soluble U(VI) in groundwater can be achieved by acetate amendment. Injection with acetate often leads to the preferential growth of Geobacter species and concomitant precipitation of U(IV) (Holmes et al. 2002). As an attractive alternative, the insertion of electrodes to provide Geobacter cells in situ with electrons has been tested (Gregory and Lovley 2005). This approach would allow for a continuous, simple supply of electrons to Geobacter growing on the electrodes for the reduction of the soluble U(VI), with the produced U(IV) precipitating on the electrode. By periodically removing and cleaning the electrode, U(VI) can be irreversibly removed.
Laboratory studies have revealed that Geobacter species also can reduce other heavy metals to less mobile forms: Ag(I) (Law et al. 2008), Co(III) (Caccavo et al. 1994), V(V) (Ortiz-Bernad et al. 2004), Tc(VII), Np(V) (Lloyd et al. 2000), Pu(IV) (Boukhalfa et al. 2007), and Hg(II) (Wiatrowski et al. 2006). In situ removal of V(V) and Tc(VII) has been revealed, however may also be due to the abiotic reduction of these heavy metals by biological produced Fe(II) (Lloyd et al. 2000; Ortiz-Bernad et al. 2004). Although Geobacter species may contribute to arsenic mobilization (Islam et al. 2004), it was observed that G. sulfurreducens does not reduce As(V) to more soluble As(III) and that the formation of solid Fe(II)-bearing phases during Fe(III) reduction aids to capture arsenic species (Islam et al. 2005).
Bioremediation of Chlorinated Contaminants
PCE and nitrate are frequently co-contaminants at uranium-contaminated sites. G. lovleyi is so far the only Geobacter species known to be capable of PCE respiration but dechlorinates PCE incompletely, with cis-dichloroethene as a more toxic end product than the parent molecule (Sung et al. 2006). G. lovleyi is, however, frequently found in enrichments with Dehalococcoides species that completely degrade PCE to ethane (Yan et al. 2012). It has been postulated that incomplete organohalide-respiring microorganisms enhance overall organohalide detoxification. Also, G. lovleyi provides Dehalococcoides species with specific cobamides for their growth (Yan et al. 2012). Based on energetic consideration, it was expected that PCE respiration precedes nitrate and U(IV) reduction; however, G. lovleyi simultaneously reduces these compounds (Sung et al. 2006). This may provide a growth advantage over competitors in poor substrate environments with time-variant substrates. G. lovleyi is also only organohalide-respiring strain known to use hydrogen and acetate. As for heavy metals, electrode-based removal strategies for PCE have been suggested (Strycharz et al. 2008).
The formation of reactive Fe(II) minerals or reduced humic acids by Geobacter species may aid the abiotic removal of carbon tetrachloride (Cervantes et al. 2004; McCormick et al. 2002).
Bioenergy
Microbial fuel cells and microbial electrolysis cells to decompose organic wastewater are intensively investigated. Microbial fuel cells generate an electric current, while microbial electrolysis cells yield hydrogen or methane from organic matter by applying a current. Geobacter species are consistently enriched on anodes in microbial fuel cells (Lovley et al. 2011), while molecular analysis has frequently revealed sequences closest related to P. propionicus in hydrogen-producing microbial electrolysis cells (Chae et al. 2008; Selembo et al. 2010), although P. propionicus dominated also anodes in acetate-fed microbial fuel cells (Kiely et al. 2011). While Geobacter species are already efficient in the transfer of electrons to electrodes, there is scope for further optimization of Geobacter strains for improved current output by metabolic engineering (Izallalen et al. 2008).
Recent observations may lead to further application of Geobacteraceae in energy production as Geobacter species are abundant syntrophs in conductive aggregates in methanogenic digesters (Morita et al. 2011), possibly enabling the rapid conversion of organic matter to methane. In an enrichment converting coal to methane, Geobacter was one of the most abundant microorganisms (Jones et al. 2010). Cocultures of Clostridium beijerinckii and G. metallireducens on xylose in the presence of a humic acid analogue as electron shuttle enhanced biohydrogen production by C. beijerinckii by up to 52 %. Hydrogen production was enhanced due to removal of Clostridium-produced, inhibitory acetate by G. metallireducens and the cycling of humic acids between the two species (Zhang et al. 2013).
Ecological Systems Biology and Environmental Biotechnology
Understanding in situ physiological status is key to bioremediation and other environmental biotechnological applications, in order to monitor and rationally engineer the activities of Geobacter species. Methods to quantify key gene transcripts and proteins, and relating their abundances to growth rates and metabolic rates, oxidative stress, acetate availability, and limitations in iron, ammonium, and phosphate, have been developed (Chin et al. 2004; Elifantz et al. 2010; Holmes et al. 2004b, 2008; Mouser et al. 2009; N’Guessan et al. 2010; O’Neil et al. 2008). The development of these indicators of in situ physiological status took optimally advantage of the availability of pure Geobacter isolates that are closely related to those in metal-reducing environments and the ability to grow these Geobacter species in chemostats under environmental relevant conditions.
The availability of sequenced genomes has contributed to constrained-based, genome-scale models of several Geobacter species and P. propionicus (Mahadevan et al. 2011). Genome-scale metabolic models integrated into reactive transport models allowed for in silico evaluation of various strategies to stimulate in situ uranium bioremediation (Scheibe et al. 2009). Flux balance analysis based on genome-scale models also contributed to a better understanding on when and why Geobacter species become dominant iron reducers or lose to other iron-reducing microorganisms (Zhuang et al. 2011) or microorganisms that employ other electron accepting processes (Barlett et al. 2012; Zhuang et al. 2012). These models have contributed to directed rational engineering to accelerate rates of respiration and power output, by creating ATP consuming futile cycles (Izallalen et al. 2008).
Genetic systems for the manipulation of G. sulfurreducens (Coppi et al. 2001) and G. metallireducens (Tremblay et al. 2012) by gene deletion and replacement have been developed and allow for metabolic engineering. Adaptive evolution by applying selective pressure for rapid Fe(III) oxide reduction resulted in the isolation of G. sulfurreducens strains with tenfold higher rates than those of the parent strain. Genome resequencing revealed mutations in a c-type cytochrome and transcriptional regulator and when these were introduced into the parent strains they were found to be responsible for the higher rates (Tremblay et al. 2011).
Future Applications
Inspired by the unique properties of Geobacter species to interact bidirectionally with electrodes, Lovley et al. (2011) presented a number of exciting potential applications. Microbial electrosynthesis is a process of synthesizing organic compounds from carbon dioxide, with the input of electrons from electrodes. Electronically functional material consisting of living Geobacter cells would allow for cheap, self-renewable applications in bioelectronics.
References
Adams LK, Harrison JM, Lloyd JR, Langley S, Fortin D (2007) Activity and diversity of Fe(III)-reducing bacteria in a 3000-year-old acid mine drainage site analogue. Geophys J Roy Astron Soc 24:295–305
Aklujkar M, Krushkal J, DiBartolo G, Lapidus A, Land ML, Lovley DR (2009) The genome sequence of Geobacter metallireducens: features of metabolism, physiology and regulation common and dissimilar to Geobacter sulfurreducens. BMC Microbiol 9:e109
Aklujkar M, Young ND, Holmes D, Chavan M, Risso C, Kiss HE, Han CS, Land ML, Lovley DR (2010) The genome of Geobacter bemidjiensis, exemplar for the subsurface clade of Geobacter species that predominate in Fe(III)-reducing subsurface environments. BMC Genomics 11:e490
Aklujkar M, Haveman SA, DiDonato R, Chertkov O, Han CS, Land ML, Brown P, Lovley DR (2012) The genome of Pelobacter carbinolicus reveals surprising metabolic capabilities and physiological features. BMC Genomics 13:e690
Barlett M, Zhuang K, Mahadevan R, Lovley D (2012) Integrative analysis of Geobacter spp. and sulfate-reducing bacteria during uranium bioremediation. Biogeosciences 9:1033–1040
Bauer I, Kappler A (2009) Rates and extent of reduction of Fe(III) compounds and O-2 by humic substances. Environ Sci Technol 43:4902–4908
Bazylinski DA, Dean AJ, Schuler D, Phillips EJP, Lovley DR (2000) N-2-dependent growth and nitrogenase activity in the metal-metabolizing bacteria, Geobacter and Magnetospirillum species. Environ Microbiol 2:266–273
Bond DR, Lovley DR (2003) Electricity production by Geobacter sulfurreducens attached to electrodes. Appl Environ Microbiol 69:1548–1555
Bosch J, Fritzsche A, Totsche KU, Meckenstock RU (2010) Nanosized ferrihydrite colloids facilitate microbial iron reduction under flow conditions. Geophys J Roy Astron Soc 27:123–129
Botton S, van Harmelen M, Braster M, Parsons JR, Röling WFM (2007) Dominance of Geobacteraceae in BTX-degrading enrichments from an iron-reducing aquifer. FEMS Microbiol Ecol 62:118–130
Boukhalfa H, Icopini GA, Reilly SD, Neu MP (2007) Plutonium(IV) reduction by the metal-reducing bacteria Geobacter metallireducens GS15 and Shewanella oneidensis MR1. Appl Environ Microbiol 73:5897–5903
Butler JE, He Q, Nevin KP, He ZL, Zhou JZ, Lovley DR (2007) Genomic and microarray analysis of aromatics degradation in Geobacter metallireducens and comparison to a Geobacter isolate from a contaminated field site. BMC Genomics 8
Butler JE, Young ND, Lovley DR (2010) Evolution of electron transfer out of the cell: comparative genomics of six Geobacter genomes. BMC Genomics 11:e40
Butler JE, Young ND, Aklujkar M, Lovley DR (2012) Comparative genomic analysis of Geobacter sulfurreducens KN400, a strain with enhanced capacity for extracellular electron transfer and electricity production. BMC Genomics 13:e471
Caccavo F, Lonergan DJ, Lovley DR, Davis M, Stolz JF, McInerney MJ (1994) Geobacter sulfurreducens sp. nov., a hydrogen-oxidizing and acetate-oxidizing dissimilatory metal-reducing microorganism. Appl Environ Microbiol 60:3752–3759
Cervantes FJ, Vu-Thi-Thu L, Lettinga G, Field JA (2004) Quinone-respiration improves dechlorination of carbon tetrachloride by anaerobic sludge. Appl Microbiol Biotechnol 64:702–711
Chae KJ, Choi MJ, Lee J, Ajayi FF, Kim IS (2008) Biohydrogen production via biocatalyzed electrolysis in acetate-fed bioelectrochemical cells and microbial community analysis. Int J Hydrogen Energy 33:5184–5192
Childers SE, Ciufo S, Lovley DR (2002) Geobacter metallireducens accesses insoluble Fe(III) oxide by chemotaxis. Nature 416:767–769
Chin KJ, Esteve-Nunez A, Leang C, Lovley DR (2004) Direct correlation between rates of anaerobic respiration and levels of mRNA for key respiratory genes in Geobacter sulfurreducens. Appl Environ Microbiol 70:5183–5189
Coates JD, Lovley DR (2005) Genus I. Geobacter. Pages 1017. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM (eds) Bergey’s Manual of Systematic Bacteriology. Springer, New York
Coates JD, Phillips EJP, Lonergan DJ, Jenter H, Lovley DR (1996) Isolation of Geobacter species from diverse sedimentary environments. Appl Environ Microbiol 62:1531–1536
Coates JD, Ellis DJ, Blunt-Harris EL, Gaw CV, Roden EE, Lovley DR (1998) Recovery of humic-reducing bacteria from a diversity of environments. Appl Environ Microbiol 64:1504–1509
Coates JD, Bhupathiraju VK, Achenbach LA, McInerney MJ, Lovley DR (2001) Geobacter hydrogenophilus, Geobacter chapellei and Geobacter grbiciae, three new, strictly anaerobic, dissimilatory Fe(III)-reducers. Int J Syst Evol Microbiol 51:581–588
Coby AJ, Picardal F, Shelobolina E, Xu HF, Roden EE (2011) Repeated anaerobic microbial redox cycling of iron. Appl Environ Microbiol 77:6036–6042
Coppi MV, Leang C, Sandler SJ, Lovley DR (2001) Development of a genetic system for Geobacter sulfurreducens. Appl Environ Microbiol 67:3180–3187
Cord-Ruwisch R, Lovley DR, Schink B (1998) Growth of Geobacter sulfurreducens with acetate in syntrophic cooperation with hydrogen-oxidizing anaerobic partners. Appl Environ Microbiol 64:2232–2236
Cummings DE, Snoeyenbos-West OL, Newby DT, Niggemyer AM, Lovley DR, Achenbach LA, Rosenzweig RF (2003) Diversity of Geobacteraceae species inhabiting metal-polluted freshwater lake sediments ascertained by 16S rDNA analyses. Microb Ecol 46:257–269
De Wever H, Cole JR, Fettig MR, Hogan DA, Tiedje JM (2000) Reductive dehalogenation of trichloroacetic acid by Trichlorobacter thiogenes gen. nov., sp nov. Appl Environ Microbiol 66:2297–2301
Doong RA, Schink B (2002) Cysteine-mediated reductive dissolution of poorly crystalline iron(III) oxides by Geobacter sulfurreducens. Environ Sci Technol 36:2939–2945
Elifantz H, N’Guessan LA, Mouser PJ, Williams KH, Wilkins MJ, Risso C, Holmes DE, Long PE, Lovley DR (2010) Expression of acetate permease-like (apl) genes in subsurface communities of Geobacter species under fluctuating acetate concentrations. FEMS Microbiol Ecol 73:441–449
Finneran KT, Housewright ME, Lovley DR (2002) Multiple influences of nitrate on uranium solubility during bioremediation of uranium-contaminated subsurface sediments. Environ Microbiol 4:510–516
Finneran KT, Johnsen CV, Lovley DR (2003) Rhodoferax ferrireducens sp nov., a psychrotolerant, facultatively anaerobic bacterium that oxidizes acetate with the reduction of Fe(III). Int J Syst Evol Microbiol 53:669–673
Garrity GM, Bell JA, Lilburn T (2005) Family II. Geobacteraceae fam. nov. Pages 1017. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM (eds) Bergey’s Manual of Systematic Bacteriology. Springer, New York
Greene AC, Patel BKC, Yacob S (2009) Geoalkalibacter subterraneus sp nov., an anaerobic Fe(III)- and Mn(IV)-reducing bacterium from a petroleum reservoir, and emended descriptions of the family Desulfuromonadaceae and the genus Geoalkalibacter. Int J Syst Evol Microbiol 59:781–785
Gregory KB, Lovley DR (2005) Remediation and recovery of uranium from contaminated subsurface environments with electrodes. Environ Sci Technol 39:8943–8947
Hartner T, Straub KL, Kannenberg E (2005) Occurrence of hopanoid lipids in anaerobic Geobacter species. FEMS Microbiol Lett 243:59–64
Hedrick DB, Peacock AD, Lovley DR, Woodard TL, Nevin KP, Long PE, White DC (2009) Polar lipid fatty acids, LPS-hydroxy fatty acids, and respiratory quinones of three Geobacter strains, and variation with electron acceptor. J Ind Microbiol Biotechnol 36:205–209
Holmes DE, Finneran KT, O’Neil RA, Lovley DR (2002) Enrichment of members of the family Geobacteraceae associated with stimulation of dissimilatory metal reduction in uranium-contaminated aquifer sediments. Appl Environ Microbiol 68:2300–2306
Holmes DE, Nevin KP, Lovley DR (2004a) Comparison of 16S rRNA, nifD, recA, gyrB, rpoB and fusA genes within the family Geobacteraceae fam. nov. Int J Syst Evol Microbiol 54:1591–1599
Holmes DE, Nevin KP, Lovley DR (2004b) In situ expression of nifD in Geobacteraceae in subsurface sediments. Appl Environ Microbiol 70:7251–7259
Holmes DE, Nevin KP, O’Neil RA, Ward JE, Adams LA, Woodard TL, Vrionis HA, Lovley DR (2005) Potential for quantifying expression of the Geobacteraceae citrate synthase gene to assess the activity of Geobacteraceae in the subsurface and on current-harvesting electrodes. Appl Environ Microbiol 71:6870–6877
Holmes DE, O’Neil RA, Vrionis HA, N’Guessan LA, Ortiz-Bernad I, Larrahondo MJ, Adams LA, Ward JA, Nicoll JS, Nevin KP, Chavan MA, Johnson JP, Long PE, Lovley DR (2007) Subsurface clade of Geobacteraceae that predominates in a diversity of Fe(III)-reducing subsurface environments. ISME J 1:663–677
Holmes DE, Mester T, O’Neil RA, Perpetua LA, Larrahondo MJ, Glaven R, Sharma ML, Ward JE, Nevin KP, Lovley DR (2008) Genes for two multicopper proteins required for Fe(III) oxide reduction in Geobacter sulfurreducens have different expression patterns both in the subsurface and on energy-harvesting electrodes. Microbiology 154:1422–1435
Islam FS, Gault AG, Boothman C, Polya DA, Charnock JM, Chatterjee D, Lloyd JR (2004) Role of metal-reducing bacteria in arsenic release from Bengal delta sediments. Nature 430:68–71
Islam FS, Pederick RL, Gault AG, Adams LK, Polya DA, Charnock JM, Lloyd JR (2005) Interactions between the Fe(III)-reducing bacterium Geobacter sulfurreducens and arsenate, and capture of the metalloid by biogenic Fe(II). Appl Environ Microbiol 71:8642–8648
Izallalen M, Mahadevan R, Burgard A, Postier B, Didonato R, Sun J, Schilling CH, Lovley DR (2008) Geobacter sulfurreducens strain engineered for increased rates of respiration. Metab Eng 10:267–275
Jones EJP, Voytek MA, Corum MD, Orem WH (2010) Stimulation of methane generation from nonproductive coal by addition of nutrients or a microbial consortium. Appl Environ Microbiol 76:7013–7022
Kaden J, Galushko AS, Schink B (2002) Cysteine-mediated electron transfer in syntrophic acetate oxidation by cocultures of Geobacter sulfurreducens and Wolinella succinogenes. Arch Microbiol 178:53–58
Kiely PD, Rader G, Regan JM, Logan BE (2011) Long-term cathode performance and the microbial communities that develop in microbial fuel cells fed different fermentation endproducts. Bioresour Technol 102:361–366
Kuever J, Rainey FA, Widdel F (2005) Family I. Desulfuromonaceae fam. nov. Pages 1006. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM (eds) Bergey’s Manual of Systematic Bacteriology. Springer, New York
Kunapuli U, Lueders T, Meckenstock RU (2007) The use of stable isotope probing to identify key iron-reducing microorganisms involved in anaerobic benzene degradation. ISME J 1:643–653
Kunapuli U, Jahn MK, Lueders T, Geyer R, Heipieper HJ, Meckenstock RU (2010) Desulfitobacterium aromaticivorans sp nov and Geobacter toluenoxydans sp nov., iron-reducing bacteria capable of anaerobic degradation of monoaromatic hydrocarbons. Int J Syst Evol Microbiol 60:686–695
Kung JW, Loffler C, Dorner K, Heintz D, Gallien S, Van Dorsselaer A, Friedrich T, Boll M (2009) Identification and characterization of the tungsten-containing class of benzoyl-coenzyme A reductases. Proc Natl Acad Sci USA 106:17687–17692
Kusel K, Blothe M, Schulz D, Reiche M, Drake HL (2008) Microbial reduction of iron and porewater biogeochemistry in acidic peatlands. Biogeosciences 5:1537–1549
Law N, Ansari S, Livens FR, Renshaw JC, Lloyd JR (2008) Formation of nanoscale elemental silver particles via enzymatic reduction by Geobacter sulfurreducens. Appl Environ Microbiol 74:7090–7093
Leang C, Coppi MV, Lovley DR (2003) OmcB, a c-type polyheme cytochrome, involved in Fe(III) reduction in Geobacter sulfurreducens. J Bacteriol 185:2096–2103
Leang C, Qian XL, Mester T, Lovley DR (2010) Alignment of the c-Type Cytochrome OmcS along Pili of Geobacter sulfurreducens. Appl Environ Microbiol 76:4080–4084
Lin WC, Coppi MV, Lovley DR (2004) Geobacter sulfurreducens can grow with oxygen as a terminal electron acceptor. Appl Environ Microbiol 70:2525–2528
Lin B, Braster M, van Breukelen BM, van Verseveld HW, Westerhoff HV, Röling WFM (2005) Geobacteraceae community composition is related to hydrochemistry and biodegradation in an iron-reducing aquifer polluted by a neighboring landfill. Appl Environ Microbiol 71:5983–5991
Lin B, Braster M, Röling WFM, van Breukelen BM (2007a) Iron-reducing microorganisms in a landfill leachate-polluted aquifer: complementing culture-independent information with enrichments and isolations. Geophys J Roy Astron Soc 24:283–294
Lin B, Hyacinthe C, Bonneville S, Braster M, Van Cappellen P, Röling WFM (2007b) Phylogenetic and physiological diversity of dissimilatory ferric iron reducers in sediments of the polluted Scheldt estuary, Northwest Europe. Environ Microbiol 9:1956–1968
Lin B, Westerhoff HV, Röling WFM (2009) How Geobacteraceae may dominate subsurface biodegradation: physiology of Geobacter metallireducens in slow-growth habitat-simulating retentostats. Environ Microbiol 11:2425–2433
Lloyd JR, Sole VA, Van Praagh CVG, Lovley DR (2000) Direct and Fe(II)-mediated reduction of technetium by Fe(III)- reducing bacteria. Appl Environ Microbiol 66:3743–3749
Lonergan DJ, Jenter HL, Coates JD, Phillips EJP, Schmidt TM, Lovley DR (1996) Phylogenetic analysis of dissimilatory Fe(III)-reducing bacteria. J Bacteriol 178:2402–2408
Lovley DR (2011) Reach out and touch someone: potential impact of DIET (direct interspecies energy transfer) on anaerobic biogeochemistry, bioremediation, and bioenergy. Rev Environ Sci Biotechnol 10:101–105
Lovley DR, Blunt-Harris EL (1999) Role of humic-bound iron as an electron transfer agent in dissimilatory Fe(III) reduction. Appl Environ Microbiol 65:4252–4254
Lovley DR, Lonergan DJ (1990) Anaerobic oxidation of toluene, phenol, and para-cresol by the dissimilatory iron-reducing organism, GS-15. Appl Environ Microbiol 56:1858–1864
Lovley DR, Phillips EJP (1988) Novel mode of microbial energy-metabolism - organic-carbon oxidation coupled to dissimilatory reduction of iron or manganese. Appl Environ Microbiol 54:1472–1480
Lovley DR, Baedecker MJ, Lonergan DJ, Cozzarelli IM, Phillips EJP, Siegel DI (1989) Oxidation of aromatic contaminants coupled to microbial iron reduction. Nature 339:297–300
Lovley DR, Giovannoni SJ, White DC, Champine JE, Phillips EJP, Gorby YA, Goodwin S (1993) Geobacter metallireducens gen. nov. sp. nov., a microorganism capable of coupling the complete oxidation of organic compounds to the reduction of iron and other metals. Arch Microbiol 159:336–344
Lovley DR, Woodward JC, Chapelle FH (1994) Stimulated anoxic biodegradation of aromatic hydrocarbons using Fe(III) ligands. Nature 370:128–131
Lovley DR, Coates JD, Blunt-Harris EL, Phillips EJP, Woodward JC (1996a) Humic substances as electron acceptors for microbial respiration. Nature 382:445–448
Lovley DR, Woodward JC, Chapelle FH (1996b) Rapid anaerobic benzene oxidation with a variety of chelated Fe(III) forms. Appl Environ Microbiol 62:288–291
Lovley DR, Ueki T, Zhang T, Malvankar NS, Shrestha PM, Flanagan KA, Aklujkar M, Butler JE, Giloteaux L, Rotaru AE, Holmes DE, Franks AE, Orellana R, Risso C, Nevin KP (2011) Geobacter: the microbe electric’s physiology, ecology, and practical applications. In: Poole RK (eds) Advances in microbial physiology, vol 59, pp 1–100. Academic Press, Amsterdam, The Netherlands
Mahadevan R, Palsson BO, Lovley DR (2011) In situ to in silico and back: elucidating the physiology and ecology of Geobacter spp. using genome-scale modelling. Nat Rev Microbiol 9:39–50
Malvankar NS, Vargas M, Nevin KP, Franks AE, Leang C, Kim BC, Inoue K, Mester T, Covalla SF, Johnson JP, Rotello VM, Tuominen MT, Lovley DR (2011) Tunable metallic-like conductivity in microbial nanowire networks. Nat Nanotechnol 6:573–579
McCormick ML, Bouwer EJ, Adriaens P (2002) Carbon tetrachloride transformation in a model iron-reducing culture: relative kinetics of biotic and abiotic reactions. Environ Sci Technol 36:403–410
Meckenstock RU (1999) Fermentative toluene degradation in anaerobic defined syntrophic cocultures. FEMS Microbiol Lett 177:67–73
Methe BA, Nelson KE, Eisen JA, Paulsen IT, Nelson W, Heidelberg JF, Wu D, Wu M, Ward N, Beanan MJ, Dodson RJ, Madupu R, Brinkac LM, Daugherty SC, DeBoy RT, Durkin AS, Gwinn M, Kolonay JF, Sullivan SA, Haft DH, Selengut J, Davidsen TM, Zafar N, White O, Tran B, Romero C, Forberger HA, Weidman J, Khouri H, Feldblyum TV, Utterback TR, Van Aken SE, Lovley DR, Fraser CM (2003) Genome of Geobacter sulfurreducens: metal reduction in subsurface environments. Science 302:1967–1969
Morita M, Malvankar NS, Franks AE, Summers ZM, Giloteaux L, Rotaru AE, Rotaru C, Lovley DR (2011) Potential for direct interspecies electron transfer in methanogenic wastewater digester aggregates. MBio 2:e00159–11
Mouser PJ, Holmes DE, Perpetua LA, DiDonato R, Postier B, Liu A, Lovley DR (2009) Quantifying expression of Geobacter spp. oxidative stress genes in pure culture and during in situ uranium bioremediation. ISME J 3:454–465
N’Guessan AL, Elifantz H, Nevin KP, Mouser PJ, Methe B, Lwoodard T, Manley K, Williams KH, Wilkins MJ, Larsen JT, Long PE, Lovley DR (2010) Molecular analysis of phosphate limitation in Geobacteraceae during the bioremediation of a uranium-contaminated aquifer. ISME J 4:253–266
Nercessian O, Parot S, Delia ML, Bergel A, Achouak W (2012) Harvesting electricity with Geobacter bremensis isolated from compost. PLoS One 7:e34216
Nevin KP, Lovley DR (2000) Lack of production of electron-shuttling compounds or solubilization of Fe(III) during reduction of insoluble Fe(III) oxide by Geobacter metallireducens. Appl Environ Microbiol 66:2248–2251
Nevin KP, Holmes DE, Woodard TL, Hinlein ES, Ostendorf DW, Lovley DR (2005) Geobacter bemidjiensis sp nov and Geobacter psychrophilus sp nov., two novel Fe(III)-reducing subsurface isolates. Int J Syst Evol Microbiol 55:1667–1674
Nevin KP, Holmes DE, Woodard TL, Covalla SF, Lovley DR (2007) Reclassification of Trichlorobacter thiogenes as Geobacter thiogenes comb. nov. J Syst Evol Microbiol 57:463–466
O’Neil RA, Holmes DE, Coppi MV, Adams LA, Larrahondo MJ, Ward JE, Nevin KP, Woodard TL, Vrionis HA, N’Guessan AL, Lovley DR (2008) Gene transcript analysis of assimilatory iron limitation in Geobacteraceae during groundwater bioremediation. Environ Microbiol 10:1218–1230
Ortiz-Bernad I, Anderson RT, Vrionis HA, Lovley DR (2004) Vanadium respiration by Geobacter metalireducens: novel strategy for in situ removal of vanadium from groundwater. Appl Environ Microbiol 70:3091–3095
Percent SF, Frischer ME, Vescio PA, Duffy EB, Milano V, McLellan M, Stevens BM, Boylen CW, Nierzwicki-Bauer SA (2008) Bacterial community structure of acid-impacted lakes: what controls diversity? Appl Environ Microbiol 74:1856–1868
Pilloni G, von Netzer F, Engel M, Lueders T (2011) Electron acceptor-dependent identification of key anaerobic toluene degraders at a tar-oil-contaminated aquifer by Pyro-SIP. FEMS Microbiol Ecol 78:165–175
Prakash O, Gihring TM, Dalton DD, Chin KJ, Green SJ, Akob DM, Wanger G, Kostka JE (2010) Geobacter daltonii sp. nov., an Fe(III)- and uranium(VI)-reducing bacterium isolated from a shallow subsurface exposed to mixed heavy metal and hydrocarbon contamination. Int J Syst Evol Microbiol 60:546–553
Reguera G, McCarthy KD, Mehta T, Nicoll JS, Tuominen MT, Lovley DR (2005) Extracellular electron transfer via microbial nanowires. Nature 435:1098–1101
Röling WFM, van Breukelen BM, Braster M, Lin B, van Verseveld HW (2001) Relationships between microbial community structure and hydrochemistry in a landfill leachate-polluted aquifer. Appl Environ Microbiol 67:4619–4629
Rooney-Varga JN, Anderson RT, Fraga JL, Ringelberg D, Lovley DR (1999) Microbial communities associated with anaerobic benzene degradation in a petroleum-contaminated aquifer. Appl Environ Microbiol 65:3056–3063
Rotaru AE, Shrestha PM, Liu FH, Ueki T, Nevin K, Summers ZM, Lovley DR (2012) Interspecies electron transfer via hydrogen and formate rather than direct electrical connections in cocultures of Pelobacter carbinolicus and Geobacter sulfurreducens. Appl Environ Microbiol 78:7645–7651
Scheibe TD, Mahadevan R, Fang Y, Garg S, Long PE, Lovley DR (2009) Coupling a genome-scale metabolic model with a reactive transport model to describe in situ uranium bioremediation. Microbiol Biotechnol 2:274–286
Schink B (1984) Fermentation of 2,3-butanediol by Pelobacter carbinolicus sp. nov. and Pelobacter propionicus sp. Nov. and evidence for propionate formation from C2 compounds. Arch Microbiol 137:33–41
Schwertmann U, Cornell RM (1991) Iron oxides in the laboratory: preparation and characterization. VCh Verlagsgesellschaft mbH/VCH Publishers, Weinheim/Germany/New York
Scott DT, McKnight DM, Blunt-Harris EL, Kolesar SE, Lovley DR (1998) Quinone moieties act as electron acceptors in the reduction of humic substances by humics-reducing microorganisms. Environ Sci Technol 32:2984–2989
Selembo PA, Merrill MD, Logan BE (2010) Hydrogen production with nickel powder cathode catalysts in microbial electrolysis cells. Int J Hydrogen Energy 35:428–437
Shelobolina ES, Sullivan SA, O’Neill KR, Nevin KP, Lovley DR (2004) Isolation, characterization, and U(VI)-reducing potential of a facultatively anaerobic, acid-resistant bacterium from Low-pH, nitrate- and U(VI)-contaminated subsurface sediment and description of Salmonella subterranea sp nov. Appl Environ Microbiol 70:2959–2965
Shelobolina ES, Nevin KP, Blakeney-Hayward JD, Johnsen CV, Plaia TW, Krader P, Woodard T, Holmes DE, VanPraagh CG, Lovley DR (2007) Geobacter pickeringii sp nov., Geobacter argillaceus sp nov and Pelosinus fermentans gen. nov., sp nov., isolated from subsurface kaolin lenses. Int J Syst Evol Microbiol 57:126–135
Shelobolina ES, Vrionis HA, Findlay RH, Lovley DR (2008) Geobacter uraniireducens sp nov., isolated from subsurface sediment undergoing uranium bioremediation. Int J Syst Evol Microbiol 58:1075–1078
Shi L, Squier TC, Zachara JM, Fredrickson JK (2007) Respiration of metal (hydr)oxides by Shewanella and Geobacter: a key role for multihaem c-type cytochromes. Mol Microbiol 65:12–20
Smith JA, Lovley DR, Tremblay PL (2013) Outer cell surface components essential for Fe(III) oxide reduction by Geobacter metallireducens. Appl Environ Microbiol 79:901–907
Snoeyenbos-West OL, Nevin KP, Anderson RT, Lovley DR (2000) Enrichment of Geobacter species in response to stimulation of Fe(III) reduction in sandy aquifer sediments. Microb Ecol 39:153–167
Staats M, Braster M, Röling WFM (2011) Molecular diversity and distribution of aromatic hydrocarbon-degrading anaerobes across a landfill leachate plume. Environ Microbiol 13:1216–1227
Stackebrandt E, Goebel BM (1994) A place for DNA-DNA reassociation and 16S ribosomal RNA sequence analysis in the present species definition in bacteriology. Int J Syst Bacteriol 44:846–849
Stackebrandt E, Wehmeyer U, Schink B (1989) The phylogenetic status of Pelobacter acidigallici, Pelobacter venetianus, and Pelobacter carbinolicus. Syst Appl Microbiol 11:257–260
Straub KL, Buchholz-Cleven BEE (2001) Geobacter bremensis sp nov and Geobacter pelophilus sp nov., two dissimilatory ferric-iron-reducing bacteria. Int J Syst Evol Microbiol 51:1805–1808
Straub KL, Hanzlik M, Buchholz-Cleven BEE (1998) The use of biologically produced ferrihyduite for the isolation of novel iron-reducing bacteria. Syst Appl Microbiol 21:442–449
Strycharz SM, Woodard TL, Johnson JP, Nevin KP, Sanford RA, Loffler FE, Lovley DR (2008) Graphite electrode as a sole electron donor for reductive dechlorination of tetrachlorethene by Geobacter lovleyi. Appl Environ Microbiol 74:5943–5947
Summers ZM, Fogarty HE, Leang C, Franks AE, Malvankar NS, Lovley DR (2010) Direct exchange of electrons within aggregates of an evolved syntrophic coculture of anaerobic bacteria. Science 330:1413–1415
Sun J, Sayyar B, Butler JE, Pharkya P, Fahland TR, Famili I, Schilling CH, Lovley DR, Mahadevan R (2009) Genome-scale constraint-based modeling of Geobacter metallireducens. BMC Syst Biol 3:e15
Sun J, Haveman SA, Bui O, Fahland TR, Lovley DR (2010) Constraint-based modeling analysis of the metabolism of two Pelobacter species. BMC Syst Biol 4:e174
Sung Y, Fletcher KF, Ritalaliti KM, Apkarian RP, Ramos-Hernandez N, Sanford RA, Mesbah NM, Loffler FE (2006) Geobacter lovleyi sp nov strain SZ, a novel metal-reducing and tetrachloroethene-dechlorinating bacterium. Appl Environ Microbiol 72:2775–2782
Tang YJJ, Chakraborty R, Martin HG, Chu J, Hazen TC, Keasling JD (2007) Flux analysis of central metabolic pathways in Geobacter metallireducens during reduction of soluble Fe(III)-nitrilotriacetic acid. Appl Environ Microbiol 73:3859–3864
Tran HT, Krushkal J, Antommattei FM, Lovley DR, Weis RM (2008) Comparative genomics of Geobacter chemotaxis genes reveals diverse signaling function. BMC Genomics 9:e471
Tremblay PL, Summers ZM, Glaven RH, Nevin KP, Zengler K, Barrett CL, Qiu Y, Palsson BO, Lovley DR (2011) A c-type cytochrome and a transcriptional regulator responsible for enhanced extracellular electron transfer in Geobacter sulfurreducens revealed by adaptive evolution. Environ Microbiol 13:13–23
Tremblay PL, Aklujkar M, Leang C, Nevin KP, Lovley D (2012) A genetic system for Geobacter metallireducens: role of the flagellin and pilin in the reduction of Fe(III) oxide. Environ Microbiol Rep 4:82–88
Ueki T, Lovley DR (2010) Genome-wide gene regulation of biosynthesis and energy generation by a novel transcriptional repressor in Geobacter species. Nucleic Acids Res 38:810–821
Validation List 107 (2006) List of new names and new combinations previously effectively, but not validly, published. Int J Syst Evol Microbiol 56:1–6
van Breukelen BM, Röling WFM, Groen J, Griffioen J, van Verseveld HW (2003) Biogeochemistry and isotope geochemistry of a landfill leachate plume. J Contam Hydrol 65:245–268
Viollier E, Inglett PW, Hunter K, Roychoudhury AN, Van Cappellen P (2000) The ferrozine method revisited: Fe(II)/Fe(III) determination in natural waters. Appl Geochem 15:785–790
Wagner DD, Hug LA, Hatt JK, Spitzmiller MR, Padilla-Crespo E, Ritalahti KM, Edwards EA, Konstantinidis KT, Loffler FE (2012) Genomic determinants of organohalide-respiration in Geobacter lovleyi, an unusual member of the Geobacteraceae. BMC Genomics 13:e200
Weber KA, Urrutia MM, Churchill PF, Kukkadapu RK, Roden EE (2006) Anaerobic redox cycling of iron by freshwater sediment microorganisms. Environ Microbiol 8:100–113
Weelink SAB, van Doesburg W, Saia FT, Rijpstra WIC, Roling WFM, Smidt H, Stams AJM (2009) A strictly anaerobic betaproteobacterium Georgfuchsia toluolica gen. nov., sp nov degrades aromatic compounds with Fe(III), Mn(IV) or nitrate as an electron acceptor. FEMS Microbiol Ecol 70:575–585
Wiatrowski HA, Ward PM, Barkay T (2006) Novel reduction of mercury(II) by mercury-sensitive dissimilatory metal reducing bacteria. Environ Sci Technol 40:6690–6696
Widdel F, Pfennig N (1982) Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids. 2. Incomplete oxidation of propionate by Desulfobulbus propionicus gen nov, sp nov. Arch Microbiol 131:360–365
Winderl C, Anneser B, Griebler C, Meckenstock RU, Lueders T (2008) Depth-resolved quantification of anaerobic toluene degraders and aquifer microbial community patterns in distinct redox zones of a tar oil contaminant plume. Appl Environ Microbiol 74:792–801
Yan BZ, Wrenn BA, Basak S, Biswas P, Giammar DE (2008) Microbial reduction of Fe(III) in hematite nanoparticles by Geobacter sulfurreducens. Environ Sci Technol 42:6526–6531
Yan J, Ritalahti KM, Wagner DD, Loffler FE (2012) Unexpected specificity of interspecies cobamide transfer from Geobacter spp. to organohalide-respiring Dehalococcoides mccartyi strains. Appl Environ Microbiol 78:6630–6636
Zhang T, Gannon SM, Nevin KP, Franks AE, Lovley DR (2010) Stimulating the anaerobic degradation of aromatic hydrocarbons in contaminated sediments by providing an electrode as the electron acceptor. Environ Microbiol 12:1011–1020
Zhang T, Bain TS, Nevin KP, Barlett MA, Lovley DR (2012) Anaerobic benzene oxidation by Geobacter species. Appl Environ Microbiol 78:8304–8310
Zhang XY, Ye XF, Finneran KT, Zilles JL, Morgenroth E (2013) Interactions between Clostridium beijerinckii and Geobacter metallireducens in co-culture fermentation with anthrahydroquinone-2, 6-disulfonate (AH2QDS) for enhanced biohydrogen production from xylose. Biotechnol Bioeng 110:164–172
Zhuang K, Izallalen M, Mouser P, Richter H, Risso C, Mahadevan R, Lovley DR (2011) Genome-scale dynamic modeling of the competition between Rhodoferax and Geobacter in anoxic subsurface environments. ISME J 5:305–316
Zhuang K, Ma E, Lovley DR, Mahadevan R (2012) The design of long-term effective uranium bioremediation strategy using a community metabolic model. Biotechnol Bioeng 109:2475–2483
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer-Verlag Berlin Heidelberg
About this entry
Cite this entry
Röling, W.F.M. (2014). The Family Geobacteraceae. In: Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F. (eds) The Prokaryotes. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-39044-9_381
Download citation
DOI: https://doi.org/10.1007/978-3-642-39044-9_381
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-39043-2
Online ISBN: 978-3-642-39044-9
eBook Packages: Biomedical and Life SciencesReference Module Biomedical and Life Sciences