
Abstract
Tarsal morphology of spiders differs from species to species, reflecting adaptations to habitat demands. Jumping spiders (Salticidae) rely on the hierarchically structured claw tuft (scopula) to firmly attach to smooth surfaces. This scopula consists of single hairs (setae) covered by numerous cuticular processes (setules). These setules are the direct contacting elements of the attachment system. Adhesion does not depend on secreted fluids but employs short-ranging van der Waals forces. The estimated 624,000 setules in the spider Evarcha arcuata allow for a theoretical safety factor of 160 times the spider´s average weight. Strong adhesion is balanced by easy detachment through an asymmetric design of the setules’ distal ends. Detachment is controlled in an integrated manner via leg kinematics. The combination of dry adhesion, functional also in vacuum, substrate compliant hierarchical structure, and switchable adhesive properties, perfectly qualifies for biomimetic transfer. To be able to transfer the adhesive performance like the one of the salticid spider E. arcuata to a technical design, it is necessary to analyze the individual contributions of adhesion, compliance, and detachment.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction: Spider Ecology and Tarsus Morphology
Similar to other animals, spiders possess structures for handling their surrounding environment. These are mostly claws, located on the pretarsus of all spiders, and—in only a number of taxa—adhesive hairs, the so-called setules. These setules are either distributed over the whole length of the tarsus as in lycosid spiders (Rovner 1978) or located ventrally of the claws as a tuft or the so-called scopula (Hill 1977). While the claws serve for handling threads or rough structured substrate, the scopula is used for attachment to smooth surfaces such as leafs or glass slides. In contrast to insect adhesion, the active secretion of fluids for adhesive purpose is not common and still subject of discussion (see Niederegger 2013).
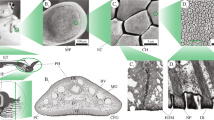
Spiders follow different strategies to gather food, ranging from web building (and waiting for the prey to get trapped) to freely hunting by approaching and subsequently grabbing their prey. Due to its free-ranging lifestyle, the salticid Evarcha arcuata does not have any specialized tool for the handling of spider silk as, for example, the orb weaver Araneus diadematus. The hierarchically structured scopula with its single hairs and abundant number of setules is an optimal adaptation toward the ground dwelling, predatory lifestyle of this species. Jumping spiders hunt down their prey in the vegetation moving on slippery plant surfaces, and thereby a firm grip to the substrate is vital, as is the firm attachment to captured prey. Tight attachment to smooth surfaces is ensured by the claw tuft apparatus, the scopula (Fig. 35.1a). The scopula is formed by a number of long hair-like appendages in a parallel configuration—the setae. Each seta is covered by tiny pointed hairs on the dorsal side and by a dense population of hairs on the ventral side, called setules. The ventral hairs are flattened at the free end, showing an inclined triangular-shaped spatula (Fig. 35.1b–d). These spatulae form the point of contact when adhering to the substrate. E. arcuata has an estimate of 624,000 contact elements, a high number in comparison to other arthropods (Martin et al. 2002b).

Hierarchical structure of Evarcha arcuata’s attachment devices. (a) Tarsus with scopula (Sc) and claws (Cl). (b) The scopula consists of setae, while each seta is covered by many setules. (c) Setules on the ventral side end in a flattened and tilted triangular structures, called spatula. (d) Each spatula is about 500 nm wide (SEM micrographs: Andrew Martin, taken from Kesel et al. 2003)
In contrast to the contacting setules on the ventral side, the function of the tapered processes on the setae’s dorsal surface remains unclear. Their size and distribution along the dorsal surface might play a role in the prevention of any adherence of one seta to another. The scopulae on the two frontal leg pairs are considerably smaller in size than those on the hind leg pairs (Martin et al. 2002a). This may be due to the individual role of legs during locomotion and prey capture (Betz and Kölsch 2004). While it appears rather a trivial aspect at first sight, easy detachment from the substrate is crucial for highly motile organisms and indeed a central requisite for the design of any tarsal attachment device be it insect, spider, frog, or gecko (Arzt et al. 2003).
2 Hierarchical Design of the Claw Tuft
The use of van der Waals forces for attachment faces two principal challenges: these forces are small and do not range very far. Hence, many contact elements are required and need to be brought close to the substrate, that is, nanometers, since these forces are inversely proportional to the surface distance.
Since real-world substrates are seldom smooth to an industrial standard, it is necessary to employ a mechanism, which allows sufficient compliance of the scopula to bring a sufficient amount of hairs close to potential attachment sites.
The single elements of the scopula exhibit certain mechanical properties: under load, each of the long and thin hairs is subject to bending, storing energy like a cantilever beam, and, hence, can be modeled as a spring in tension or compression (Bhushan et al. 2006; Gasparetto et al. 2009). In consequence, the scopula—with seta, setule, and triangular end—can be described as a three-level spring model made of tarsus (cuticula), setae, and setules (Fig. 35.2).

Elasto-mechanical model of the hierarchical adhesive structure of spider claw tufts. sp = spatulae, se = setulae, st = setae, cl = claws. Modified from Gasparetto et al. (2009)
Let θ be the approaching angle between the fibrillar elements and the surface. By considering an F force perpendicular to the substrate, a bending (δ b) and compressive (δ c) deformation is created as
where L is the length, I the moment of inertia (πR 4/4), R the radius, E the Young modulus, and A the cross-sectional area (πR 2). The total stress results in
and the stiffness of a single element becomes
By considering the Young modulus of the fibrillar elements (materials such as β-keratin have Young modulus in the order of a few gigapascals), the equivalent stiffness of the setae and setules can be estimated. Subsequently, the optimized mechanical compliance of the spider tarsi can be reached over surfaces of different roughness with a suitable hierarchical structure with respect to single layer dry-adhesive systems simulated and demonstrated by Gasparetto et al. (2009).
3 Adhesion with a Nonadhesive Material
Quantifying the adhesive properties of single setule has become possible with the invention of atomic force microscopes and related instruments. Point spectroscopic measurements (Fig. 35.3) with calibrated cantilevers allowed to determine an average attachment force of 38.12 nN per setule in E. arcuata, resulting in a theoretically possible total adhesive force of 2.38 × 10−2 N (Kesel et al. 2003). Considering that the mean body mass of Evarcha arcuata is 15.1 × 10−3 kg, the maximal adhesion force equals 160 times the body weight and, hence, grants safe attachment even when hunting prey. These scopulae are part of the animal’s cuticle exhibiting super-hydrophobic characteristics too.

Force-distance curve of an atomic force microscope during point spectroscopy. The straight vertical line of the lower curve denotes the lift-off event of the cantilever from the specimen. The force required to break the contact is the attachment force of the specimen to the cantilever. Phases of force spectroscopy: (A) cantilever out of contact, (B) snap-in due to attractive forces, (C) loading of probe onto sample with increasing force, (D) begin retraction of cantilever, (E) unloading, (F) probe remains in contact due to adhesive forces, negative loading, (G) force exerted by cantilever spring equals and subsequently exceeds adhesive force of probe-sample contact (from Kesel et al. 2004)
As prior mentioned, the major acting physical principle is the van der Waals forces, weak and short-ranging (i.e., tens of nanometers) intermolecular forces, possibly supported by capillary forces (Kesel et al. 2003; Autumn and Peattie 2002). Concerning the adhesive force model, if the surface asperities and the adhesive element ends are modeled like spherical tips, the interaction force can be estimated by considering the interaction of two spheres as formulated by the JKR model (Johnson et al. 1971). Thus, the adhesion force results in
where r c is the reduced radius:
with r 1 the radius of the hemisphere on the tip of the adhesive element and r 2 the radius of the hemisphere that represents the asperity, here assumed as r 1 = r 2. Let the energy of adhesion E ad range from 10 to 66 mJ/m2, that is, the typical range for van der Waals surfaces, and working with the estimated radius of the spider’s setules, the adhesion force becomes very close to the experimental ones, that is, 38 nN (Gasparetto et al. 2009).
When comparing the adhesives properties of cuticle with other materials that were tested in the same setup, it comes very clear that by its material properties, the cuticle does not show extremely high adhesive properties (Table 35.1). Apparently this weakness is compensated for by the structural arrangement of the material in the scopula.
4 Adhesion and Adhesive Properties
The previously described physical principle is only one element contributing to the design of biomimetic adhesive devices based on the spider attachment performance. In the following we introduce the most important features:
The triangular-shaped distal end on each setule—the so-called spatula—shows anisotropic adhesion to the substrate, meaning that the adhesive force is strongly correlated with the spatula’s orientation towards the substrate. This allows the animal to actively control adhesion and, for example, to produce low detachment force when required, as demonstrated nicely by Gao et al. (2005) using finite element modeling techniques. Here, adhesion maximizes at angles around 30°; at smaller angles, sliding occurs and detachment at higher angles. And indeed, the spider’s scopula attachment system seems to work completely passively, in contrast to the muscularly mediated digital hyperextension in geckos (Autumn and Peattie 2002). In the latter, an active distal peeling motion allows easy detachment of the adhesive system from the substrate, while E. arcuata’s attachment is only controlled via the movement of the spider’s legs. To this point there were no active detachment systems observed in living spiders, and since the scopula lies on the tarsus tip, both the pulling of the front and the pushing of the rear legs create a sort of natural and passive rotation motion with a pivot of the touching area. This mechanism allows to gradually detaching the fibrillar elements in contact with the surface without a significant effort.
Two important properties keep these hierarchical structures functional during their lifetime: self-cleaning and anti-bunching. The former prevents loose particles from sticking to the tarsus and consequently blocking adhesion sites and reducing attachment properties (Hansen and Autumn 2005). The latter prevents the densely packed setules from sticking to each other rather than to the substrate. In short, this is achieved by (1) the anisotropic adhesive properties of the spatula and (2) the optimized geometrical conditions and spring constants of the setules preventing their terminal ends to meet in such an alignment that would allow bunching (see, for instance, Sitti and Fearing 2003). Indeed, the anti-bunching effect is a result of a careful choice of geometrical parameters of the fibrillar elements at each level, that is, radius, length, and distance, to create a particle-repellant and bunching-resistant hairy attachment system. These parameters can and need to be taken into account when designing biomimetic attachment devices.
5 Technical Application
Spiders are skillful locomotors and, together with E. arcuata’s powerful passively controlled attachment and detachment system, great models for biomimetic transfer. The two most appealing approaches will be described in the following.
5.1 Bioadhesives
The first attempt is to create bio-inspired dry-adhesive structures for universal application. Such attachment devices would employ the same physical principle and the same observed statistical adhesion with a summed attachment force correlating with the dynamics during attaching. Such “stickers” could function in many environments, even liquid or vacuum, but, contrary to, for example, Velcro, would not need a predefined substrate. They would have no power required to maintain attachment, no dependence upon gravitational forces to realize attachment, and availability on any surface regardless of issues related to surface roughness or friction coefficient. It is easy to imagine that space research is highly interested in creating a bio-inspired artificial adhesive able to mimic the most versatile used repairing and attaching technique in orbit.
Early attempts tried to duplicate the natural dry-adhesive performance by designing and fabricating extremely small hairs (Sitti and Fearing 2003). This resulted in an ineffective adhesion on the macroscale. After that, as reported in Sameoto and Menon (2010) and Zhang et al. (2010), a significant effort has been given to this topic, and effective adhesive systems, even if not yet as powerful as the natural ones, have been realized. In particular, the evolution of synthetic dry-adhesives has taken two main directions: the first focuses on the development of relatively soft elastomer fibers with mushroom-shaped tips, which typically exhibit high normal adhesion strengths, while the second focuses on stiff, very high aspect ratio fibers, which exhibit good friction forces, but either low or zero normal adhesion strength.
Two fabrication methods are preferred for prototyping attachment devices: micro/nanocasting and gas-phase growth or etching (Sameoto and Menon 2010). In the former, the majority of developed adhesives have been realized using materials such as polydimethylsiloxane (PDMS) and polyurethanes (PU), while, in the latter, aligned arrays of carbon nanotubes are usually grown.
Such perfect adhesives can answer to different technical challenges or create new effective and efficient systems for a broad range of applications. Indeed, they could be used for biomedical applications such as safe and sterilized endoscopy and tissue adhesives; micro-electro-mechanical systems (MEMS) and wafer alignment and manipulation; structures to substitute glues, screws, traditional adhesives, reversible adhesive gloves, or rock climbing aids; and robotic systems.
5.2 Bio-inspired Spider Robots
Taking the complexity of biomimetic transfer some level higher, the entire subsystem of locomotion and adhesion, that is, the spider’s leg, gets into focus. From the locomotion point of view, a spider-inspired legged robot is not a new idea, but the availability of new mechatronic systems and the discovery and study of the spiders’ reversible passive adhesive principles have drawn new attention and interest to these arthropods.
Indeed, the idea of using biomimetic, asymmetric adhesives to control attachment to various types of surfaces merely via the kinematics of the legs can be exploited in order to create legged robots that can both overcome obstacles and cope with different kinds of surfaces by means of these synthetic dry-adhesive systems and with a minimal effort.
The European Space Agency has encouraged works in this direction (compare Gasparetto et al. 2008, 2009, 2010; Menon et al. 2008; Li et al. 2012). The first approach deals with modeling and simulating a climbing spider-inspired robot by studying the anatomy, the adhesive and locomotion capabilities of the spider (i.e., an eight-legged system), and the kinematic and dynamic behavior in different operative and slope conditions (Fig. 35.4a), while the second approach deals with the macro-, micro-, and nano-structural design and realization of a miniaturized spider-inspired legged robot capable to negotiate different roughness and slope surfaces and of operating in a space environment (Fig. 35.4b). The realized prototype, called Abigaille II, is a lightweight robot actuated by eighteen miniaturized motors and equipped on the tip of the legs with bio-inspired adhesive patches and is able to climb effectively vertical Plexiglas surfaces.

Bio-inspired spider robots. Above: spider robot simulator implementing the cooperation between locomotion and adhesion (Vidoni and Gasparetto 2011). Blue elements represent spiders’ leg joints. The tip of the tarsus of each leg is visualized with different colors due to the angle of adhesion and to the condition of the leg: red—in adhesion; green—in detachment or in flight. Arrows represent the body reference frame. Below: spider robot prototype with dry-adhesive pads developed at Simon Fraser University (Li et al. 2012). The prototype shows six legs in order to reduce the number of actuators and, in the meantime, allow a biomimetic gait
6 Conclusions
The attempt to design autonomous devices—commonly known as robots—has led many people to draw inspiration from the broad choice of functional autonomous mobile organisms—commonly known as animals. Among these, hunting spiders have caught the attention of the researchers since they can move fast over a broad range of surfaces and slopes and easily overcome obstacles. Walking on vertical and inverted surfaces is as possible as dynamically hunting down prey. In contrast to insects, spiders employ a dry-adhesive system using van der Waals interactions between their numerous tiny terminal ends of the hairy scopula and hence supply a model for adhesion also in extreme environments like vacuum. Indeed, effective attachment is achieved through elastic and damping properties of both material and hierarchical structure, while detachment is achieved via the angle variation between tarsus and substrate thanks to the macroscopic legs motion (Gasparetto et al. 2010) and hence comes “free of charge” by control of leg posture. Mimicking the adhesive system as well as locomotory capabilities will enable engineers to create effective and innovative technological devices in a broad range of application fields, starting from universally usable reversible adhesives to autonomous space and hazardous environments exploring robots and manipulators.
References
Arzt E, Gorb E, Spolenak R (2003) From micro to nano contacts in biological attachment devices. Proc Natl Acad Sci 100:10603–10606
Autumn K, Peattie AM (2002) Mechanisms of adhesion in geckos. Integr Comp Biol 42: 1081–1090
Betz O, Kölsch G (2004) The role of adhesion in prey capture and predator defence in arthropods. Arthrop Struct Develop 33:3–30
Bhushan B, Peressadko TW, Kim AG (2006) Adhesion analysis of two-level hierarchical morphology in natural attachment systems for “smart adhesion”. J Adhes Sci Technol 20: 1475–1491
Gao H, Wang X, Yao H, Gorb S, Arzt E (2005) Mechanics of hierarchical adhesion structures of geckos. Mech Mater 37:275–285
Gasparetto A, Vidoni R, Seidl T (2008) Kinematic study of the spider system in a biomimetic perspective. In: Proceedings of the IEEE/RSJ international conference on intelligent robots and systems, Nice, France
Gasparetto A, Seidl T, Vidoni R (2009) A mechanical model for the adhesion of spiders to nominally flat surfaces. J Bionic Eng 6:135–142
Gasparetto A, Vidoni R, Seidl T (2010) Passive control of attachment in legged space robots. Appl Bionics Biomech 7:69–81
Hansen WR, Autumn K (2005) Evidence for self-cleaning in gecko setae. Proc Natl Acad Sci 102: 385–389
Hill DE (1977) The pretarsus of salticid spiders. Zool J Linn Soc 60:319–338
Johnson KL, Kendall K, Roberts AD (1971) Surface energy and the contact of elastic solids. Proc R Soc London A 324:301–313
Kesel AB, Martin A, Seidl T (2003) Adhesion measurements on the attachment devices of the jumping spider Evarcha arcuata. J Exp Biol 206:2733–2738. doi:10.1242/jeb.00478
Kesel AB, Martin A, Seidl T (2004) Getting a grip on spider attachment: an AFM approach to microstructure adhesion in arthropods. Smart Mater Struct 13:512–518. doi:10.1088/0964-1726/13/3/009
Li Y, Ahmed A, Sameoto D, Menon C (2012) Abigaille II: toward the development of a spider-inspired climbing robot. Robotica 30:79–89
Martin A, Dillinger SCG, Kesel AB, Nachtigall W (2002a) The structure of tarsal attachment elements in Araneae. Zoology 105(35 Suppl):95
Martin A, Seidl T, Kesel AB (2002b) Das mikro-mechanische Haftsystem der Spinne Evarcha arcuata. In: Wisser A, Nachtigall W (eds) Biona-Report 16:226–230
Menon C, Li Y, Sameoto D, Martens C (2008) Abigaille-I: towards the development of a spider-inspired climbing robot for space use. In: IEEE RAS/EMBS international conference on biomedical robotics and biomechatronics, Scottsdale
Niederegger (2013) Functional aspects of spider scopulae. In: Nentwig W (ed) Spider ecophysiology. Springer, Berlin (this volume)
Rovner JS (1978) Adhesive hairs in spiders: behavioral functions and hydraulically mediated movement. Symp Zool Soc Lond 42:99–108
Sameoto D, Menon C (2010) Recent advances in the fabrication and adhesion testing of biomimetic dry adhesives. Smart Mater Struct 19:103001
Sitti M, Fearing RS (2003) Synthetic gecko foot-hair for micro/nano structures as dry adhesives. J Adhes Sci Technol 18:1055–1074
Vidoni R, Gasparetto A (2011) Efficient force distribution and leg posture for a bio-inspired spider robot. Rob Auton Syst 59:142–150
Zhang H, Guo DJ, Dai ZD (2010) Progress on gecko-inspired micro/nano-adhesion arrays. Chin Sci Bull – Bionic Eng 55:1843–1850. doi:10.1007/s11434-010-3065-z
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Seidl, T., Vidoni, R. (2013). Adhesion to Flat Surfaces: From Spiders to Stickers. In: Nentwig, W. (eds) Spider Ecophysiology. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-33989-9_35
Download citation
DOI: https://doi.org/10.1007/978-3-642-33989-9_35
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-33988-2
Online ISBN: 978-3-642-33989-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)