
Abstract
Regional approaches to estimate the carbon budget of Italian forest ecosystems using Process-Based Models (PBMs), have been applied by several national institutions and researchers. Gross and net primary productivity (GPP and NPP) have been estimated through the PBMs simulations of carbon, water, and elemental cycles driven by remotely sensed data set and ancillary data. In particular the results of the GPP and NPP estimations provided by the implementation of two hybrid models are presented. The first modeling approach, based on the integration of two widely used models (C-fix and BIOME-BGC), has been applied to simulate monthly GPP and NPP values of all Italian forests for the decade 1999–2008. The approach, driven by remotely sensed SPOT-VEGETATION ten-day Normalized Difference Vegetation Index (NDVI) images and meteorological data, provided a NPP map of Italian forests reaching maximum values of about 900 g C m−2 year−1. The second modeling approach is based on the implementation of a modified version of the 3-PG model running on a daily time step to produce daily estimates of GPP and NPP. The model is driven by MODIS remotely sensed vegetation indexes and meteorological data, and parameterized for specific soil and land cover characteristics. Average annual GPP and NPP maps of Italian forests and average annual values for different forest types according to Corine Land Cover 2000 classification are reported.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Normalize Difference Vegetation Index
- Forest Type
- Standing Volume
- Basic Wood Density
- Normalize Difference Vegetation Index Image
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Simulation models of forest ecosystems answer two needs: first to clarify the relationship between key ecosystem components, for a deeper understanding of their functioning (Kimmins 2008), and second to predict how the state variables of a dynamic system change due to processes in a forest stand or landscape (Brang et al. 2002). In recent years, modeling has undergone significant developments especially in forestry. Modeling tools are increasingly used by both forest ecologists, who face the challenge of transferring knowledge to stakeholders and the general community, and forest managers, who benefit from the development of scenario-based supports for decision-making (Vacchiano et al. 2012). From a general point of view, modeling means trying to capture the essence of a system, deconstructing complex interactions between system components until only the most essential structures and processes remain (Haefner 2005). From stochastic and empirical models, developed over the past 50 years, the increased availability of the data has led to a significant enhancement in the knowledge of the processes that regulate the tree eco-physiology. The difficulties to apply empirical models in sites other than those they were calibrated for, which do not reflect the changes occurred in site conditions or related to management operations since they were developed, have switched to using models able to predict changes in growth and productivity of forests also subject to climate changes, often taking into consideration some factors relating to anthropogenic disturbance. Depending on the modeling purpose, in the last three decades a series of modeling approaches were developed in order to capture forest processes for a wide spatial and temporal resolution scale. The most used approaches are: gap models (Bugmann 2001), landscape models (He 2008), process-based models (PBMs) (Makela et al. 2000) and hybrid models (Zhang et al. 2008). The former of this series explicitly includes site and climate drivers for predicting forest composition, structure and biomass. Small-area or gap models reproduce the growth of single trees within forest patches (e.g., 100 m2) in relation to the prevailing growth conditions at the site level (Botkin et al. 1972; Shugart 1984; Leemans and Prentice 1989; Pacala et al. 1993). Recent modeling approaches as for the 3D-CMCC FEM (three Dimensional Forest Ecosystem Model of the euro-Mediterranean Center for Climate Change) (Collalti et al. 2014) integrates several characteristics of the functional–structural tree models, based on the light use efficiency (LUE) approach, to investigate forest growth patterns and yield processes for complex multi-layer forests.
However, physiological processes are not explicitly accounted for, requiring statistical fitting procedures between each environmental factor and observed growth (Vacchiano et al. 2012).
Landscape models comprise a broad class of spatially explicit models that incorporate heterogeneity in site conditions, neighborhood interactions and feedbacks between different spatial processes (Pretzsch et al. 2008). The aims of these models are to develop scenarios for the sustainability of forest or landscape functions (natural resources, habitat, hydrology, socioeconomic), to forecast their response to disturbances and potential environmental change (climate, N deposition, land use and land use change), to investigate the relationship between landscape structure and regionally distributed risks, and to assess regional-scale matter fluxes, e.g. water, carbon and nutrients. One example is the mesoscale Land Surface Model proposed by Alessandri and Navarra (2008) representing the momentum, heat and water flux at the interface between land-surface and atmosphere; it has been coupled to a general circulation model (GCM) to estimate the rate of forcing by existing vegetation on precipitation patterns. PBMs can be defined as a procedure by which the behavior of a system is derived from a set of functional components and their interactions with each other and the system environment, through physical and mechanistic processes occurring over time (Godfrey 1983; Bossel 1994). More generally, these models are part of the Soil-Vegetation-Atmosphere Transport (SVAT) models giving a representative description of land surface-atmosphere interaction, and describing the physical and biological processes in vegetation and soil, as well as physical processes within the atmospheric boundary layer. SVAT models are commonly used to estimate the exchanges of energy, mass and momentum between the atmosphere and the land surface. These types of models, which are widely applied and validated across the world, use the “big leaf” concept based on one canopy layer or multiple layer schemes, to simulate water and carbon cycles on a variety of spatial (hectare to km) and temporal (daily, monthly or annually) scales. The implementation of these models in forestry in the last decades has been having great success thanks to the availability of remotely sensed data offering a greater amount of information both during the initialization and validation phase. Also, fluxes of energy, CO2 and water vapor exchanges between the vegetation and the atmosphere measured by the FLUXNET network give the possibility to test such models over as many different circumstances as possible. The spatial scale which they generally work at (ecosystem) can describe the main features in the structure and physiognomy of the forest and they can be considered a valuable tool in the study of those eco-physiological fundamental processes, at species level but also at forest typology level, at an intermediate spatial scale between gap models and Dynamic Global Vegetation Models (DGVM). An important feature of SVAT models is that they can be used as stand-alone models (Marras et al. 2011; Staudt et al. 2011) or as the land surface scheme of a climate model (Pyles et al. 2003). However, as reported by Zhang et al. (2008), most process-based models are unable to simulate forest stand variables (e.g., height, diameter at breast height and volume) since they were not designed for forest management and do not predict forest stand attributes. Battaglia and Sands (1998), Landsberg and Coops (1999) and Makela et al. (2000) have extensively discussed the advantages and disadvantages of using empirical and mechanistic process models. Generally, as postulated by Peng et al. (2002), the weakness of one type of model is the strength of the other, and vice versa. It is almost always possible to find an empirical model providing a better fit for a given set of data due to the constraints imposed by the assumptions of process models. Nevertheless, empirical and process-based models can be combined and integrated into hybrid models in which the shortcomings of both approaches can be overcome to some extent.
According to this general framework, several ecosystem models have been applied to estimate carbon budgets of Italian forests, relying on the availability of remotely sensed data and ancillary dataset provided within the activities of specific national research projects as for the CarboItaly project.
2 The Carbon Budget Estimation of the Italian Forests: Ecosystem Models Approach
Regional approaches to estimate the carbon budget of Italian forests have been applied in the last decade by several national institutions and researchers. In particular several PBMs driven by remotely sensed and ancillary data have been applied to run simulations of carbon, water and elemental cycles in order to provide estimates of GPP and NPP and thus of NEP over a wide variety of vegetation types across Italian forest ecosystems.
Over the last decade, the availability of micrometeorological data measured within a national ground-based monitoring network of Eddy Covariance tower sites (flux sites), has been used to calibrate and validate PBMs. In general, the modeling approach is mainly based on the combination and integration of widely applied PBMs into hybrid models so to better represent the high variability of land use, climate and environmental conditions over the Italian territory.
2.1 Estimation of Italian Forest NPP. C-Fix and BIOME-BGC Integration Model
Maselli et al. (2009a) and Chiesi et al. (2011) proposed the estimation of forest NPP in Italy based on the integration of a parametric model, C-Fix, and of a bio-geochemical model, BIOME-BGC. C-Fix is a Monteith type parametric model (Veroustraete et al. 2002) which combines satellite-derived estimates of the fraction of Photosynthetically Active Radiation absorbed by forest (fAPAR) with field based estimates of incoming solar radiation and air temperature to simulate total photosynthesis. The annual GPP (g C m−2 year−1) of a forest can be computed as:
where ε is the maximum radiation use efficiency, Tcor i is a factor accounting for the dependence of photosynthesis on air temperature, Cws i is the water stress index, fAPAR i is the fraction of absorbed PAR, and Rad i is the solar incident PAR, all referred to the i-th month. fAPAR can be derived from the top of canopy NDVI according to the linear equation proposed by Myneni and Williams (1994). Cws was introduced by Maselli et al. (2009a) to optimize the model application in Mediterranean environments, which are characterized by a long and dry summer season when vegetation growth is constrained by water availability. This modification is completed by the use of the MODIS temperature correction factors and the maximum radiation use efficiency equal to 1.2 [(g C MJ−1(APAR))] (Chiesi et al. 2011).
Modified C-Fix was applied to simulate monthly GPP values of all Italian forests for the past decade (1999–2008) following the multi-step methodology described in Maselli et al. (2009a). In summary, a 1-km2 dataset of monthly minimum and maximum temperatures, precipitation and solar radiation was derived from the available meteorological maps. These maps were further processed to compute the temperature and water stress correction factors which are needed to drive Modified C-Fix. The Spot-VGT ten-day NDVI images of the ten study years were corrected for residual disturbances, composed over monthly periods and processed to obtain fAPAR maps. All these maps were used to apply Modified C-Fix and yield monthly GPP images over the study years. These images were aggregated to compute an annual average GPP image of Italy, from which average values were extracted for all forest types and Italian Regions.
The ecosystem respirations needed for the prediction of NPP in the Italian forest types were then simulated by BIOME-BGC. This model was developed at the University of Montana to estimate the storage and fluxes of carbon, nitrogen and water within terrestrial ecosystems (Running and Hunt 1993). It requires daily weather data, general information on the environment (i.e. soil, vegetation and site conditions) and on parameters describing the ecophysiological characteristics of vegetation. The model works by searching for a quasi-climax equilibrium (homeostatic condition) with local eco-climatic conditions through the spin-up phase: this means that the sum of simulated respirations become nearly equivalent to GPP, which makes annual NPP approach heterotrophic respiration (Rhet) and NEE tend to zero. Also, such modeling makes the obtained GPP estimates similar to those produced by C-Fix, which are descriptive of all ecosystem components (Maselli et al. 2009b). The version of the model currently used includes complete parameter settings for all main biome types (White et al. 2000). These settings were modified for six forest types to adapt to Mediterranean environments, which show eco-climatic features markedly different from those the model was originally developed for (see Chiesi et al. 2007 for details).
The application of BIOME-BGC in the Italian context required the transformation of the quasi-climax GPP, respiration and allocation estimates into estimates of real forest ecosystems, which are generally far from climax due to the occurred disturbances. The modeling strategy of Maselli et al. (2009b) considers the ratio between actual and potential forest standing volume as an indicator of ecosystem proximity to climax. This ratio can therefore be used to correct the photosynthesis and respiration estimates obtained by the model simulations. Accordingly, actual forest NPP (NPPA, g C m−2 year−1) can be approximated as:
where GPP, Rgr and Rmn correspond to the GPP, growth and maintenance respiration estimated by BIOME-BGC (g C m−2 year−1), and the two terms FC A (actual forest cover) and NV A (actual normalized standing volume), both dimensionless, are derived from the ratio between actual and potential tree volume.
Due to the previously described functional equivalence of C-Fix and BIOME-BGC GPP estimates, the outputs of the two models can be integrated by multiplying BIOME-BGC photosynthesis and respiration estimates for a ratio between C-Fix and BIOME-BGC GPP. In the current case, BIOME-BGC was applied only to the Tuscany territory, due to the lack of daily meteorological data for the rest of Italy. This required the application of an approximation methodology based on the use of two further assumptions. First, respiration simulated by BIOME-BGC was assumed to vary linearly following photosynthesis, which allowed the calculation of growth and maintenance respiration as constant fractions of GPP for each forest type. Second, a similar assumption was applied to simulate spatial variations of maximum standing volume and LAI, which were needed to compute FC A and NV A (Maselli et al. 2009a). Both these assumptions are in reasonable accordance with BIOME-BGC logic, which simulates ecosystems whose all main properties and functions are descriptive of a quasi-climax equilibrium.
The reference values of GPP, respirations, stem carbon and LAI were recovered for each forest type from a BIOME-BGC simulation performed in Tuscany over a 12-year time period (Chiesi et al. 2011). Stem carbon was converted into maximum standing volume using the coefficients given by Federici et al. (2008). BIOME-BGC estimates were then rescaled for each forest type following relevant Modified C-Fix GPP outputs. The regional values of actual forest standing volume needed to compute FC A and NV A were extracted for each forest type and Region from the map of Gallaun et al. (2010). All these data were combined within Eq. 5.2 to compute NPP A for each forest type and Region. CAI values (m3 ha−1 year−1) were then computed through Eq. 5.3:
where SCA is the Stem C Allocation ratio, BEF the volume of above ground biomass/standing volume Biomass Expansion Factor (both dimensionless), and BWD is the Basic Wood Density (Mg m−3). The SCAs of the six forest types are those of BIOME-BGC, while BEFs and BWDs are taken again from Federici et al. (2008). The multiplication by 2 accounts for the transformation from carbon to dry matter, and that by 100 for the change in magnitude from g m−2 to Mg ha−1.
The CAI modeled values were finally validated through comparison with the CAI measurements taken during the INFC, considering only the Regions where the presence of each forest type was significant (at least 10 1-km2 pixels). The comparison was carried out considering all six forest types and summarizing the results by the correlation coefficient (r), the root mean square error (RMSE) and the percentage mean bias error (%MBE, i.e. MBE/measured average*100).
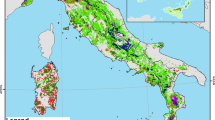
The NPP map of Italian forests simulated by the described modeling approach is shown in Fig. 5.1. The maximum NPP is around 900 g C m−2 year−1, and is prevalently found on the lowest Alpine and intermediate Apennines zones. As regards the forest types, the highest productions are obtained for species distributed over hilly-low mountain areas (i.e. deciduous oaks and chestnut), which are less affected by thermal and water limitations.

Map of estimated NPP for Italian forests
Measured (INFC) and estimated forest CAIs are shown in the scatter plot of Fig. 5.2. A moderate accordance is observable (r = 0.617; RMSE = 2.04 m3 ha−1) and there is a tendency to underestimation (%MBE = −19.5). Most of this underestimation derives from Eq. 5.2, where FC A and NV A are computed using standing volumes which are significantly lower than those of INFC (%MBE = −23.6). It can therefore be concluded that the applied modeling strategy is capable of providing realistic regional CAI estimates using information completely independent of INFC measurements.

Measured versus estimated forest CAI for all forest types and regions considered (n = 69; ** = highly significant correlation, P < 0.01)
2.2 Estimation of Italian Forest NPP. the 3-PG Model
Within the CarboItaly project, the NPP of the Italian forests has also been estimated through the application of a modified version of the widely used 3-PG model by Landsberg and Waring (1997). The 3-PG model as proposed by Nolè et al. (2013) is based on the 3-PGS (Spatial) model (Coops et al. 1998, 2005, 2007; Coops and Waring 2001; Nolè et al. 2009; Tickle et al. 2001) modified to run on a daily time step and produce estimates of GPP and NPP improving model reliability and maintaining the original simplified modeling approach at the same time.
The model fundamental assumption is the canopy LUE (light use efficiency) approach, considering the GPP as the product of the absorbed photosynthetically active radiation (aPAR) and ε max, which is assumed to be a biome-specific constant for potential LUE (g C m−2 MJ−1), and reduced by the effect of environmental constraints (f (x)). Daily GPP (g C m−2 MJ−1) has then been computed as follows:
The model reduces daily potential GPP by the effect of environmental constraints represented by four modifiers, ranging between 0 (system “shutdown”) and 1 (no constraint). Main environmental modifiers are daily average temperature (T) modifier (f T ), daily VPD modifier (f D ), soil water modifier (f θ ) and light modifier (f L ). Effects of daily average temperature on daily GPP have been modeled as a function of cardinal temperatures, minimum (Tmin), maximum (Tmax) and optimum (Topt) temperature for net photosynthetic production, as proposed by Sands and Landsberg (2002). Other environmental modifiers have been calculated according to the original model routine as proposed by Landsberg and Waring (1997). A new environmental modifier introduced in this new version is the light modifier f L , to describe the nonlinearity light response of forest ecosystem (Grace et al. 1995; Baldocchi and Harley 1995). The light modifier describes, with a hyperbolic function, the gradual saturation of GPP with increasing irradiance, as proposed by Makela et al. (2008):
where γ (m2 mol−1) is an empirical parameter.
The input dataset, provided by the partners of the CarboItaly Project, is composed by daily maps of meteorological variables derived from NCEP/NCAR (Reanalysis) and MSG (Meteosat 2nd generation), remotely sensed vegetation indexes from MOD15A2 LAI–fPAR, soil characteristics from SPADE-2 European Soil Database and land use-land cover maps from Corine Land Cover 2000. Model estimates of GPP have been validated against daily measurements from two Mediterranean Eddy Covariance sites of the CarboItaly Project (Arca di Noè–Le Prigionette (Sardinia) and Castelporziano (Lazio). In particular, model results show a significant correlation (Fig. 5.3) for the Mediterranean sites and a tendency to overestimate GPP during the summer season.

EC measured and 3PG estimated GPP daily patterns for: a Arca di Noè–Le Prigionette (2005–2007); b Castelporziano (2005–2007) (Nolè et al. 2013)
The map of estimated annual GPP and NPP for Italian forests is shown in Figs. 5.4 and 5.5 respectively, with maximum values of forest production mainly distributed in the Apennines sub-alpine areas. In Table 5.1 average annual GPP and NPP for different forest types according to Corine Land Cover 2000 classification is reported, showing the highest values of estimated NPP for chestnut woods, holm oak and evergreen woods, and more generally for low and middle mountain forest ecosystems.

Map of estimated annual GPP for Italian forests

Map of estimated annual NPP for Italian forests
3 Conclusions
The implementation of hybrid models, based on the integration of different process-based and empirical models, represents one of the most important tools for the understanding of forest ecosystem processes and to estimate forest ecosystem productivity at regional scale. These models have been applied on a wide range of Italian forest types within several research projects, as for the national specific CarboItaly project.
The availability of both high resolution remotely sensed dataset and micrometeorological data for model parameterization and validation, contributed to the development of new methodological approaches for the estimation of carbon budgets of Italian forests. The converging results provided by the two different hybrid models previously presented, show the reliability of these models in predicting national forest productivity at regional scale. A significant contribution to models reliability is provided by the availability of ground-based data set measured at the national flux network of Eddy Covariance sites covering main national ecosystem typologies.
References
Alessandri A, Navarra A (2008) On the coupling between vegetation and rainfall inter-annual anomalies: possible contributions to seasonal rain fall predictability over land areas. Geophys Res Lett 35:L02718
Baldocchi DD, Harley PC (1995) Scaling carbon dioxide and water vapour exchange form leaf to canopy in a deciduous forest. II. Model testing and application. Plant Cell Environ 18:1157–1173
Battaglia M, Sands P (1998) Process-based forest productivity models and their applications in forest management. For Ecol Manage 102:13–32
Bossel H (1994) Modeling and simulation. AK Peters Ltd, Wellesley
Botkin D, Janak J, Wallis J (1972) Some ecological consequences of a computer model of forest growth. J Ecol 60:849–873
Brang P, Courbaud B, Fischer A, Kissling-Naf I, Pettenella D, Schoneberger W et al (2002) Developing indicators for the sustainable management of mountain forests using a modelling approach. For Policy Econ 4:113–123
Bugmann H (2001) A review of forest gap models. Clim Change 51:259–305
Chiesi M, Maselli F, Moriondo M, Fibbi L, Bindi M, Running SW (2007) Application of BIOME-BGC to simulate Mediterranean forest processes. Ecol Model 206:179–190
Chiesi M, Fibbi L, Genesio L, Gioli B, Magno R, Maselli F, Moriondo M, Vaccari F (2011) Integration of ground and satellite data to model Mediterranean forest processes. Int J Appl Earth Obs Geoinf 13:504–515
Collalti A, Perugini L, Santini M, Chiti T, Nolè A, Matteucci G, Valentini R (2014) A process-based model to simulate growth in forests with complex structure: evaluation and use of 3D-CMCC Forest Ecosystem Model in a deciduous forest in Central Italy. Ecol Model 272(24):362–378
Coops NC, Waring RH (2001) Estimating forest productivity in the eastern Siskiyou Mountains of southwestern Oregon using a satellite driven process model, 3-PGS. Can J For Res 31:143–154. doi:10.1139/cjfr-31-1-143
Coops NC, Waring RH, Landsberg JJ (1998) Assessing forest productivity in Australia and New Zealand using a physiologically-based model driven with averaged monthly weather data and satellite-derived estimates of canopy photosynthetic capacity. For Ecol Manage 104(1–3):113–127. doi:10.1016/S0378-1127(97)00248-X
Coops NC, Waring RH, Law BE (2005) Assessing the past and future distribution and productivity of ponderosa pine in the Pacific Northwest using a process model, 3-PG. Ecol Model 183(1):107–124. doi:10.1016/j.ecolmodel.2004.08.002
Coops NC, Black TA, Jassal RS, Trofymow JA, Morgenstern K (2007) Comparison of MODIS, eddy covariance determined and physiologically modeled gross primary production (GPP) in a Douglas-fir forest stand. Remote Sens Environ 107:385–401
Federici S, Vitullo M, Tulipano S, De Lauretis R, Seufert G (2008) An approach to estimate carbon stocks change in forest carbon pools under the UNFCCC: the Italian case. iForest 1:86–95
Gallaun H, Zanchi G, Nabuurs GJ, Hengeveld G, Schardt M, Verkerk PJ (2010) EU-wide maps of growing stock and above-ground biomass in forests based on remote sensing and field measurements. For Ecol Manag 260:252–261
Godfrey K (1983) Compartmental models and their applications. Academic Press, New York
Grace J, Lloyd J, McIntyre J, Miranda A, Meir P, Miranda H, Moncrieff J, Massheder J, Wright I, Gash J (1995) Fluxes of carbon dioxide and water vapour over an undisturbed tropical forest in south-west Amazonia. Glob Change Biol 1:1–12
Haefner J (2005) Modeling biological systems: principles and applications. Springer, New York
He (2008) Forest landscape models: definitions, characterization, and classification. For Ecol Manage 254:484–498
Kimmins J (2008) From science to stewardship: harnessing forest ecology in the service of society. For Ecol Manage 256:1625–1635
Landsberg J, Coops N (1999) Modeling forest productivity across large areas and long periods. Nat Res Model 12:383–411
Landsberg JJ, Waring RH (1997) A generalized model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning. For Ecol Manage 95(3):209–228. doi:10.1016/S0378-1127(97)00026-1
Leemans R, Prentice I (1989) FORSKA, a general forest succession model. Institute of Ecological Botany, Uppsala
Makela A, Landsberg J, Ek A, Burk T, Ter-Mikaelian M, Agren G et al (2000) Process-based models for forest ecosystem management: current state of the art and challenges for practical implementation. Tree Physiol 20(289):298
Makela A, Pulkkinen M, Kolari P, Lagergren F, Berbigier P, Lindroth A, Loustau D, Nikinmaa E, Vesala T, Hari P (2008) An empirical model of stand GPP with the LUE approach: analysis of eddy covariance data at five contrasting conifer sites in Europe. Global Change Biol 14:92–108
Maselli F, Papale D, Puletti N, Chirici G, Corona P (2009a) Combining remote sensing and ancillary data to monitor the gross productivity of water-limited forest ecosystems. Remote Sens Environ 113:657–667
Maselli F, Chiesi M, Moriondo M, Fibbi L, Bindi M, Running SW (2009b) Modelling the forest carbon budget of a Mediterranean region through the integration of ground and satellite data. Ecol Mod 220:330–342
Marras S, Pyles RD, Sirca C, Paw UKT, Snyder RL, Duce P, Spano D (2011) Evaluation of the advanced Canopy-Atmosphere-Soil Algorithm (ACASA) model performance over Mediterranean maquis ecosystem. Agric For Meteorol 151:730–745
Myneni RB, Williams DL (1994) On the relationship between FAPAR and NDVI. Remote Sens Environ 49:200–211
Nolè A, Collalti A, Magnani F, Duce P, Ferrara A, Mancino G, Marras S, Sirca C, Spano D, Borghetti M (2013) Assessing temporal variation of primary and ecosystem production in two Mediterranean forests using a modified 3-PG model. Ann For Sci 70:729–741. doi:10.1007/s13595-013-0315-7
Nolè A, Ferrara A, Law BE, Magnani F, Matteucci G, Ripullone F, Borghetti M (2009) Application of the 3-PGS model to assess carbon accumulation in forest ecosystems at a regional level. Can J For Res 39(9):1647–1661
Pacala S, Canham C, Silander J (1993) Forest models defined by field measurements: I. The design of a northeastern forest simulator. Can J For Res 23:1980–1988
Peng C, Li J, Dang Q, Apps M, Jiang H (2002) TRIPLEX: a generic hybrid model for predicting forest growth and carbon and nitrogen dynamics. Ecol Model 153:109–130
Pyles RD, Weare BC, Paw U, Gustafso KTW (2003) Coupling between the University of California, Davis, Advanced Canopy–Atmosphere–Soil Algorithm (ACASA) and MM5: Preliminary Results for July 1998 for Western-North America. J Appl Meteorol 42:557–569
Pretzsch H, Grote R, Reineking B, Rotzer T, Seifert S (2008) Models for forest ecosystem management: a European perspective. Ann Bot 101:1065–1087
Running H, Hunt ERJ (1993) Generalization of a ecosystem process model for other biomes, BIOME-BGC, and an application for global-scale models. In: Ehleringer JR, Field CB (eds) Scaling physiological processes: leaf to globe. Academic Press, San Diego, pp 141–158
Sands PJ, Landsberg JJ (2002) Parameterization of 3-PG for plantation grown Eucalyptus globulus. For Ecol Manage 163:273–292
Shugart H (1984) A theory of forest dynamics: the ecological implications of forest succession models. Springer, New York
Staudt K, Serafimovich A, Siebicke L, Pyles RD, Falge E (2011) Vertical structure of evapotranspiration at a forest site (a case study). Agric For Meteorol 151:709–729
Tickle PK, Coops NC, Hafner SD, The Bago Science Team (2001) Assessing Forest Productivity at local scales across a native eucalypt forest using a process model, 3PG-SPATIAL. For Ecol Manage 152(1–3):275–291. doi:10.1016/S0378-1127(00)00609-5
Vacchiano G, Magnani F, Collalti A (2012) Modeling Italian forests: state of the art and future challenges. iForest, e1–e8
Veroustraete F, Sabbe H, Eerens H (2002) Estimation of carbon mass fluxes over Europe using the C-Fix model and Euroflux data. Remote Sens Environ 83:376–399
White MA, Thornton PE, Running SW, Nemani RR (2000) Parameterization and sensitivity analysis of the BIOME-BGC terrestrial ecosystem model: net primary production controls. Earth Interact 4:1–85
Zhang J, Chu Z, Ge Y, Zhou X, Jiang H, Chang J et al (2008) TRIPLEX model testing and application for predicting forest growth and biomass production in the subtropical forest zone of China’s Zhejiang Province. Ecol Model 219:264–275
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Nolè, A. et al. (2015). The Role of Managed Forest Ecosystems: A Modeling Based Approach. In: Valentini, R., Miglietta, F. (eds) The Greenhouse Gas Balance of Italy. Environmental Science and Engineering(). Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-32424-6_5
Download citation
DOI: https://doi.org/10.1007/978-3-642-32424-6_5
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-32423-9
Online ISBN: 978-3-642-32424-6
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)