
Abstract
Santa Maria is the oldest island of the Azores Archipelago and is remarkably rich in exposed marine fossiliferous sediments and submarine volcanic sequences. This chapter summarises the geological history of the island and reports on the most important palaeontological studies done on the outcrops of Santa Maria since the early studies, during the 19th century. The most important early Pliocene and late Pleistocene (Last Interglacial) fossiliferous deposits are described and palaeoecological reconstructions are presented for each sedimentary succession. The most abundant, diversified and well-studied fossil groups are also reviewed, namely the algae, vertebrates (cetaceans and selaceans), and invertebrates (molluscs, echinoderms, brachiopods, crustaceans and ostracods). We also discuss the palaeontological significance of carbonate sequences in reefless volcanic oceanic islands. Finally, we discuss the importance of applied palaeontology, with products specifically designed for tourism, such as the Museum “House of the Fossils”, the “Route of the Fossils” project, and the “PalaeoPark Santa Maria”, all of them aiming to protect and conserve fossil sites, allowing at the same time, its sustainable use by locals and tourists.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

1 Introduction
Seamounts and oceanic islands are prominent volcanic features that rise abruptly from the deep sea to shallower waters, and often constitute true oases of biodiversity in the otherwise relatively poorly diverse open ocean. Seamounts, especially when they rise above the photic zone, provide substratum to a variety of benthic organisms—such as algae, corals and sponges—that in turn provide shelter and nutrients for a series of other benthic and pelagic organisms, allowing the establishment of more complex ecosystems as time passes. Oceanic islands, as they emerge above sea-level, allow the formation of coastlines, further increasing the diversity of available shallow marine environments. Likewise, the formation of landmasses in the vast open ocean is generally followed by a selective terrestrial biological colonization which is a wonder as many naturalists and scientists have recently noted. Thus, with time, mid-ocean volcanic edifices typically host unique ecosystems that reflect coeval environmental conditions and may be the result of different evolutionary pathways concerning patterns of colonization, adaptive and non-adaptive radiation, speciation, etc. The study of such ecosystems has been at the basis of many of the ground-breaking advances in the biological sciences in the last 250 years, and is at the forefront of several disciplines such as biogeography, ecology, evolution, etc.
Ocean island volcanoes typically grow through intermittent summit eruptions, gradually building up to form prominent edifices. Volcanic activity is, generally, discontinuous in space and time, allowing for the establishment of biological communities away from active vents (see Vogt and Jung, Chapter “The “Azores Geosyndrome” and Plate Tectonics: Research History, Synthesis, and Unsolved Puzzles” for an overview on the Azores). In a similar fashion, in these areas, erosion and sedimentation gradually contribute to the formation of soils and other sedimentary bodies that further enhance the possibilities for biological colonization. However, when volcanic activity shifts back to these areas, sedimentary sequences and biological remains may be incorporated into the volcanic edifice and thus preserved from subsequent erosion. The ongoing but intermittent volcanic activity—although highly disruptive for living communities—leads to a more generalised preservation of taphonomical assemblages into the fossil record at oceanic islands, allowing the study and reconstruction of past biological communities and their surrounding environmental conditions. This is especially true for marine environments, because the rapid cooling of lava flows imposed by the subaqueous environment minimizes the destruction associated with the extrusion of these flows, leading to the generation of submarine volcano-sedimentary sequences that are very informative of the environment in which they were formed (e.g. Johnson et al. 2012, 2017; Meireles et al. 2013; Ávila et al. 2015a; Rebelo et al. 2016a; Uchman et al. 2016).
The marine fossil record at oceanic islands is, however and at a first glance, relatively poor when compared to the marine fossil record on continents; the lack of fossiliferous exposures at oceanic islands sharply contrasts with the nearest continental coasts. Nevertheless, the marine fossil record at oceanic islands is of the upmost importance for the comprehension of life’s dispersal in the oceans and adjacent coastlines, and may provide useful constraints to disciplines such as palaeo-climatology and palaeo-oceanography (e.g., Ávila et al. 2015b, 2016a). But why are marine fossiliferous exposures so rare in oceanic islands? The answer lays in the subsidence trend that most oceanic islands experience.
The growth and decay of volcanic island edifices is typically accompanied by vertical movements of diverse nature (e.g. Walcott 1970; McNutt and Menard 1978; Watts and ten Brink 1989; Ramalho et al. 2010a, b, c; Madeira et al. 2010). Commonly, island edifices subside markedly during their initial stages of growth, as the weight of the rapidly-growing edifices induce a downwards flexure of the “rigid” lithospheric plate underneath (Brotchie and Silvester 1969; Walcott 1970; Watts and ten Brink 1989; Huppert et al. 2015). Likewise, as the tectonic plate on which the island edifices rest gradually cools with age, it slowly subsides and so also the edifices with it, resulting in a longer-term subsidence response (Stein and Stein 1992; Morgan et al. 1995, see also Vogt and Jung, Chapter “The “Azores Geosyndrome” and Plate Tectonics: Research History, Synthesis, and Unsolved Puzzles”). In a similar fashion, the bathymetric swell associated with many hotspots (whose origins are still highly debated) and in which many islands rest, may experience a slow topographic decay, further contributing to long-term island subsidence (Dietz and Menard 1953; Crough 1978; Sleep 1990; Morgan et al. 1995). Thus, not surprisingly, oceanic islands are typically ephemeral features in the vastness of the geological time, subsidence and erosion being the main contributors to their relatively short-lived existence above sea-level.
The short- and long-term subsidence trends that most oceanic islands experience—together with ongoing vigorous volcanic activity during the initial stages—gradually bury and/or submerge coastlines as the edifices themselves age. As a consequence, the coastal sediments and the skeletal remains of existing marine living communities that eventually get incorporated in the subsiding volcanic edifice, become inaccessible, especially because eustatic sea level was rarely above the present level in the last 5–10 Ma, a reasonable life-time for a subsiding island (see Miller et al. 2005). Hence, the natural occurrence of exposed marine fossiliferous sequences at oceanic islands require particular conditions that are associated with the rare edifices that experienced a long-term vertical trend different than most islands—like many of the Cape Verdes and the Canaries, Porto Santo (Madeira archipelago), and Santa Maria (Azores), for example (Madeira et al. 2010; Ramalho et al. 2010a, b, c, 2015, 2017; Ramalho 2011; Menéndez et al. 2008; Klügel et al. 2005, 2015; Ávila et al. 2012). These islands all have undergone vertical movements that resulted in a net uplift, at least during part of their evolution. This uplift, coupled with marine and fluvial erosion, exposed sequences that otherwise would be deeply buried in the interior of the volcanic edifice and below present sea level.
2 The Marine Fossil Record of Santa Maria as a Result of Exposure by Uplift and Erosion
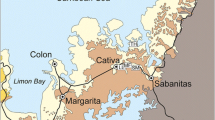
Santa Maria Island—the oldest and southeasternmost island in the Azores (Fig. 1a)—is remarkably rich in exposed marine fossiliferous sediments and submarine volcanic sequences (Fig. 1b), even within the North Atlantic context (Mitchell-Thomé 1976; Serralheiro et al. 1990; Madeira et al. 2007). This is mostly due to several factors: (1) Santa Maria’s edifice experienced a relatively long but intermittent volcanic life intercalated by periods of quiescence, erosion and sedimentation, leading to the deposition and preservation of thick marine volcano-sedimentary sequences; (2) the island experienced pronounced uplift during the Pliocene-Quaternary that, coupled with coastal and fluvial erosion, exposed numerous submarine sequences.

a Map of the Azores (insert) and geographical location of Santa Maria Island. © Secção de Geografia, Departamento de Biologia, Universidade dos Açores. b Map of Santa Maria with the location of the most important and well-studied early Pliocene and Pleistocene (MIS 5e) outcrops

The emergence of Santa Maria’s volcanic edifice above sea level occurred during the Late Miocene (Mitchell-Thomé 1976; Serralheiro et al. 1987; Serralheiro and Madeira 1990; Serralheiro 2003; Sibrant et al. 2015; Ramalho et al. 2017). The oldest volcanostratigraphic units in the island attest to this transition between submarine and subaerial volcanic activity and correspond to remains of surtseyan and strombolian monogenetic cones (Cabrestantes and Porto Formations; Serralheiro et al. 1987), dated to ~6.0 Ma (Ramalho et al. 2017).
The next stage in the development of Santa Maria was the extrusion of a basaltic subaerial shield volcano, through a number of fissural vents. The remains of this largely effusive edifice correspond to the Anjos Volcanic Complex (Serralheiro et al. 1987) extruded 5.8–5.3 Ma (Ramalho et al. 2017). The former dimensions of this edifice are difficult to constraint but it partially corresponded to the western half of present-day Santa Maria, as its eroded products do not extend to its eastern portion (Serralheiro 2003; Sibrant et al. 2015; Ramalho et al. 2017).
Subsequently to the extrusion of a basaltic shield volcano, the edifice experienced a period of general quiescence and erosion during which it was truncated by marine erosion and probably totally submerged again (Ávila et al. 2012; Meireles et al. 2013). This stage is attested to by the Touril Complex that comprises a thick (up to 130 m) sedimentary sequence of terrestrial and marine sediments with increasingly marine characteristics towards the top. On the eastern side of the island, submarine lavas and surtseyan tuffs occur intercalated within the sediments indicating that low-volume submarine volcanic activity occurred alongside erosion and sedimentation on this side of the edifice (Serralheiro et al. 1987; Serralheiro 2003; Ávila et al. 2012; Meireles et al. 2013; Ramalho et al. 2017). The Touril sequence was deposited during the Early Pliocene (5.3–4.1 Ma), as the overall volcanostratigraphy, fossil content and existing isotopic ages suggest (see Feraud et al. 1980; Serralheiro et al. 1987; Serralheiro and Madeira 1990; Serralheiro 2003; Kirby et al. 2007; Sibrant et al. 2015; Ramalho et al. 2017). The marine sediments of the Touril Complex constitute the most important and richest source of marine fossil assemblages on Santa Maria, reflecting a period during which the island represented a wide shallow and sandy submarine bank where marine life thrived (Ávila et al. 2012, 2015a, c, 2016a; Ramalho et al. 2017).
The following volcanostratigraphic unit—the Pico Alto Volcanic Complex (Serralheiro et al. 1987)—suggests that towards the beginning of the Pliocene, volcanic activity resumed with increased intensity. This new volcanic edifice started entirely submarine but as it grew upwards it breached sea-level forming a new island, elongated along a NNW-SSE direction, on the eastern portion of the old edifice (Serralheiro and Madeira 1990; Ávila et al. 2012; Ramalho et al. 2017). This renewed period of volcanic activity rapidly covered a large portion of the pre-existing shallow sandy bank with extensive submarine sheet flows, lava deltas and surtseyan deposits, effectively protecting the Touril sedimentary sequence from marine erosion and contributing to the preservation of such a rich coastal and shelf fossiliferous record. Likewise, the later onset of effusive subaerial activity and the formation of lava deltas along the existing coastlines further contributed to the preservation of more recent and discrete shelf sedimentary bodies that developed in depocentres away from active volcanic vents.
With the demise of Pico Alto volcanic activity, Santa Maria’s edifice eroded briefly during which shore platforms formed and subaerial reliefs decayed. Alluvial/fluvial deposits and remains of marine terraces that were later covered by the products of the last eruptive stage of Santa Maria’s volcanic life—corresponding to the Feteiras Formation (Serralheiro et al. 1987)—attest to this period. This brief period of volcanic activity took place during the Late Pliocene and involved low volumes of erupted material; it was restricted to a set of monogenic magmatic and hydromagmatic cones and their proximal and distal deposits, centred along the western portion of the then existing edifice, with volcanic activity seemingly ceasing at ~2.8 Ma (Serralheiro 2003; Sibrant et al. 2015; Ramalho et al. 2017).
Since the extrusion of the Feteiras volcanics, Santa Maria entered a long period of volcanic quiescence, uplift and erosion, as indicated by the presence of well-developed staircase shore platforms up the western portion of the present-day edifice. These marine terraces, found at elevations from 210–230 m down to 7–11 m above the present sea level, confirms that the edifice underwent a slow uplift from ~3.5 Ma to the Holocene, at a rate of ~60 m/Ma (Ramalho et al. 2017). This recent uplift, coupled with marine and fluvial erosion, was responsible for the exposure of the rich and diverse submarine volcanic and sedimentary sequences that otherwise would be inaccessible. These circumstances make Santa Maria one of the best places to study the Neogene marine fossil record in the North Atlantic region. Furthermore, the fossiliferous outcrops on Santa Maria are of utmost importance to gain a better understanding of how coeval living communities relate to the broader evolutionary and biogeographic history of the Atlantic basin during the late Neogene (Kirby et al. 2007; Ávila et al. 2015b, 2016a).
3 A Brief Overview About the Palaeontological Studies at Santa Maria Island
The marine fossils of Santa Maria Island have been known since the 16th century. Gaspar Frutuoso (1522–1591), a Portuguese priest, first reported on the fossils of Santa Maria in his third volume of “Saudades da Terra” (Frutuoso 1983). He described a quarry at Figueiral, where the extracted calcareous sandstones had “seafood shells glued on it” (Madeira et al. 2007). A long lapse of time passed until the late 19th century studies by Bronn (1860), Hartung (1860) and Morelet (1860). The first two authors extensively described the geomorphology of the island and, together with the work of Reiss (1862), Hartung (1864) and Mayer (1864), established the background for the palaeontological research in Santa Maria during the first half of the 20th century (Madeira et al. 2007). During the 1950s, a series of studies published by Berthois (1950, 1951, 1953a, b, c), Ferreira (1952, 1955) and Krejci-Graf et al. (1958) was followed by the important palaeontological studies of Zbyszewski et al. (1961), da Ferreira (1961) and Zbyszewski and Ferreira (1961, 1962a, b), which predominantly concentrated on the Pliocene fossils. In 1990, a paper by García-Talavera raised attention to the Pleistocene Prainha outcrop, located on the southern coast of the island. Callapez and Soares (2000) described another outcrop of the same age from Lagoinhas on the northern shores.
From 1998 on, a series of expeditions to Santa Maria undertaken by the first author of this chapter were followed by the birth of a palaeontological research group—the Marine PalaeoBiogeography Working Group (MPB)—based at the Department of Biology of the University of the Azores. Its main objectives were to achieve an understanding of the palaeoecology and palaeobiogeography of the Pleistocene and Pliocene outcrops, and to explore the legal protection of the geological legacy of Santa Maria Island. As a result, a checklist of the Pleistocene marine molluscs of Lagoinhas and Prainha (Ávila et al. 2002), and a technical report for the protection of the outcrops of Pedreira do Campo and Figueiral (Cachão et al. 2003), the first “Regional Natural Monument” of the Azores, were produced. Since 2002, the outcrops of Santa Maria became the target of the international workshops “Palaeontology in Atlantic Islands”, which have taken place on a yearly basis. During these workshops, researchers studied the geology and geomorphology of the island, the petrology, geochemistry and chronology of magmatic rocks, the volcanostratigraphy, the sedimentology and the systematic palaeontology of several phyla, the palaeoecology of the fossil assemblages and the palaeobiogeography of diverse invertebrate groups embedded within these strata. This research effort resulted in several publications covering a wide range of topics, such as the evolution of marine organisms in oceanic islands (Ávila 2013), systematics (Ávila et al. 2002, 2007, 2012; Estevens and Ávila 2007; Janssen et al. 2008; Kroh et al. 2008; Madeira et al. 2011; Meireles et al. 2012), palaeoecology and geochemistry (Kirby et al. 2007; Ávila et al. 2008b, 2009a, 2015a, c; Habermann 2011; Stöckhert 2011a, b), palaeobiogeography (Ávila et al. 2008a, 2009b, 2016a), geology (Habermann 2010; Ramalho et al. 2017), sedimentology (Meireles et al. 2013; Johnson et al. 2017), ichnology (Uchman et al. 2016), conservation and sustainable touristic management (Cachão et al. 2003; Madeira and Ávila 2006; Calado et al. 2007; Nunes et al. 2007), educational purposes (Ávila 2009; Ávila and Monteiro 2009; Ávila et al. 2010; Ávila and Rodrigues 2013), geoconservation (Ávila et al. 2016b) and historical reviews (Madeira et al. 2007). The majority of these papers dealt with the fossils of marine molluscs, but other groups of fossils also were studied, e.g.: calcareous algae (Amen et al. 2005; Rebelo et al. 2014, 2016a, b), cetaceans (Estevens and Ávila 2007; Ávila et al. 2015c), brachiopods (Kroh et al. 2008), (…) crustaceans (barnacles: Winkelmann et al. 2010), ostracods: Meireles et al. 2012), echinoderms (Madeira et al. 2011; Santos et al. 2015), and fish (sharks; Ávila et al. 2012). Two congresses entitled “Atlantic Islands Neogene” were also organised by MPB members in June 2006 and in September 2008 aiming at: (1) reviewing the accumulated scientific knowledge on the palaeontology of the Atlantic oceanic islands and (2) outlining future strategies of research in the Azores (Ávila and Martins 2007).
4 The Early Pliocene Outcrops
4.1 Pedreira do Campo and Figueiral
Pedreira do Campo (Fig. 3a) and Figueiral (Fig. 4a) are two historic quarries located in the area southwest of Pico Facho, a 254 m high tuff-cone ~2 km east of Villa do Porto close to the southern coast of the island, in which up to 20 m thick successions of the sedimentary Touril Complex and the overlying volcanic Pico Alto Complex (sensu Serralheiro et al. 1987) crop out. The outcrops are situated at altitudes between 95–115 m and comprise major deposits of fossiliferous calcarenites and limestones (Krejci-Graf et al. 1958; Zbyszewski and Ferreira 1962b; Abdel-Monem et al. 1975; Serralheiro et al. 1987, 1990; Serralheiro and Madeira 1990; Kirby et al. 2007; Madeira et al. 2007).

Pedreira do Campo outcrop. a The former quarry for basalts. b Detail of the fossil assemblage rich in mollusc fragments, spines of echinoderms, bryozoans and benthic foraminifers. c Ash and lapilli tuffs of the Pico Alto Complex with planar to low-angle cross-bedding. d basanitic pillow lavas of the Pico Alto Complex

Figueiral outcrop. a The historical quarry seen from the outside. b Shells of the dominant bivalves Aequipecten macrotis (Sowerby) and Aequipecten opercularis (Linnaeus) (facies 6). c Note the abundant benthic and planktonic macroforaminifers (amphestigenids and globigerinids) (facies 5). d The interior of one of the galleries exploited for limestone. e ash and lapilli tuffs of the Pico Alto Complex (facies 8)
4.1.1 Pedreira do Campo
At Pedreira do Campo, located ~1.4 km southwest of Pico Facho, a ~20 m thick succession of basanitic pillow lavas of the Pico Alto Complex is exposed in a cross-section with a lateral extent of ~150 m (Fig. 3a). Underlying strata only crop out locally, comprising deposits of the Touril and Pico Alto Complexes (Serralheiro et al. 1987, 1990; Cachão et al. 2003; Kirby et al. 2007; Madeira et al. 2007). The base of the Touril deposits is not exposed.
Three lithological facies (1–3) are distinguished at Pedreira do Campo (Fig. 2a). The lowermost unit (facies 1) consists of a white, impure, volcaniclastic limestone, which forms the top of the Touril Complex at this locality (Serralheiro et al. 1987; Figs. 2a, 5a–c). Well-rounded volcaniclastic pebbles and cobbles are scattered within the limestone. Volcaniclastic components make up ~5% of these compact rocks and are commonly concentrically encrusted by coralline algae (rhodoliths). The basaltic cores of these rhodoliths are small, ranging from 10 to 40 mm (average = 20.8 mm; 56 rhodoliths); 32% of the 82 rhodoliths sampled had no nucleus. This rhodolith floatstone exhibits a packstone matrix, rich in mollusc fragments (mainly bivalves and a few gastropods), spines of echinoderms (Fig. 3b), bryozoans and benthic foraminifers (mostly amphestigenids that are less than 2 mm in length). The rhodolith floatstone is unconformably overlain by a ~4.5 m thick unit (facies 2; cf. Fig. 2a) of light brown, well-stratified surtseyan tuffs (previously classified as lithic arenites by Kirby et al. 2007) that were assigned to the Pico Alto Complex according to Serralheiro et al. (1987, 1990). The tuffs are moderately sorted and show planar to low-angle cross-bedding (Fig. 3c). Influx of epiclastic materials, particularly moulds of bivalves and gastropods, are restricted to the basal 10 cm (Kirby et al. 2007). The tuffs are succeeded by about 20 m of basanitic pillow lavas (facies 3; Fig. 3d), which are assigned to the Pico Alto Complex (Serralheiro et al. 1987, 1990). The basal contact is abrupt or made up of a thin discontinuous horizon of pillow breccias and hyaloclastites. The pillows are approximately 1–2 m wide and abundantly display typical concentric structures with disjunctive radial solidification fractures and altered glassy quenching rims at their surfaces.

Outcrop photographs and photomicrographs of Pedreira do Campo and Figueiral limestones. a–c Pedreira do Campo (facies 1). d–f Figueiral (facies 2). g–i Figueiral (facies 5). j–l: Figueiral (facies 7). The blue stain is due to resin impregnation. For detailed facies descriptions and stratigraphic sample localities see text and Fig. 4

4.1.2 Figueiral
Known since the 16th century (Frutuoso 1983), the Figueiral outcrop (Fig. 4a)—located on the south-western flank of Pico do Facho, ~500 m east of Pedreira do Campo—was repeatedly mentioned in literature for its fossiliferous marine strata (Hartung 1860; Reiss 1862; Mayer 1864; Ferreira 1952, 1955, 1961; Zbyszewski et al. 1961; Zbyszewski and Ferreira 1962a; Cachão et al. 2003; Madeira et al. 2007).
Eight lithological facies (1–8) are distinguished through the exposed deposits (Fig. 2b). The outcrop starts with ~5 m thick, unstratified, poorly sorted, volcaniclastic cobble breccia (facies 1). The base of this unit is not exposed. Towards the top, increasing numbers of rounded cobbles and pebbles occur, which are frequently encrusted by coralline red-algae (rhodoliths). The volcaniclastic matrix of the breccia grades into a strongly cemented bioclastic pack- to floatstone (facies 2) rich in fragments of bivalves, gastropods, bryozoans, coralline algae, echinoids, benthic and planktonic foraminifers (amphestigenids, globigerinids) and with a micritic lime-mud matrix (Figs. 2b, 5d–f). The overlying 2 m thick beds are poorly exposed on the surface. Small outcrops in this interval suggest a heterogeneous composition of volcaniclastic gravel and boulders and increased amounts of arenitic and argillaceous matrix, generally lacking lime-mud. Bioturbated sandstone lenticules, thin horizons of brown clays, as well as scattered rhodoliths and pectinid shells occur within this ‘transition zone’ (facies 3). The basal bed exposed in the Figueiral quarry consists of a rhodolith-floatstone (facies 4). Most volcaniclastic pebbles that are scattered in a white grainstone matrix are well rounded and encrusted by red algae. The abundance of volcaniclastic pebbles and rhodoliths decreases upwards, while matrix amounts increase and the floatstone grades into a ~1.5–2 m thick, unstratified, well-sorted grainstone (facies 5), mainly consisting of benthic and planktonic foraminifers (amphestigenids up to 1 mm; globigerinids), fragments of bryozoans and red algae (Fig. 5g–i). Larger fossil-fragments and volcaniclastic detritus are accessories, lime mud is absent. Towards the top, the amount of pyroclastic materials considerably increases. The overlying ~50 cm thick tuffaceous coquina (facies 6) is rich in disarticulated, well-preserved pectinid shells <5 cm in length (dominantly Aequipecten macrotis (Sowerby) and Aequipecten opercularis (Linnaeus); cf. Fig. 4b). Poorly-sorted palagonitic lapilli form the major matrix constituents besides variable accessory amounts of skeletal fragments of bivalves, gastropods, bryozoans and foraminifers. Barnacle fragments of Zullobalanus santamariaensis Buckeridge and Winkelmann also are observed (Winkelmann et al. 2010). A sharp boundary characterizes the transition to the overlying 5–10 cm thick, laterally discontinuous amphestigenid-grain- to rudstone horizon (facies 7) with rhodoliths, benthic foraminifers, bryozoans and accessory echinoid fragments (Fig. 5j–l). This unit represents the top of the Touril Complex at Figueiral, and is abruptly overlain by ash and lapilli tuffs of the Pico Alto Complex with a thickness of >10 m at this outcrop (facies 8). Fossil content and the amount of epiclastic materials rapidly decrease at the base of this unit. The pyroclastic succession abundantly shows primary sedimentary structures. Horizontally stratified, subparallel, thin-bedded subsets (up to 1–2 m thick) are alternatingly interbedded with tabular- to tangential low-angle cross-stratified channels. Diagenetic nodular limestone crusts are commonly intercalated within the tuffs.
4.1.3 Interpretation and Discussion
On Santa Maria, sediment supply to the shelf mostly comes from marine erosion during sea-level highstands, while sediment supply by river systems is negligible (Ávila et al. 2008b). This allowed the development of carbonate factories during phases of minor volcanic activities, which were preserved in small depocentres, such as at Figueiral and Pedreira do Campo during the Lower Pliocene.
At both localities, carbonate factories developed in a shallow-marine photic environment. Stratigraphic relationships and the faunal composition suggest that the Pedreira do Campo limestone (facies 1) stratigraphically corresponds to the uppermost limestone horizon (facies 7) at Figueiral (Fig. 2a, b). Facies differences between both limestones likely reflect the high facies variability due to the variability in palaeo-shelf morphology between two small depocentres or within such a small depocentre.
The sedimentary succession within these outcrops is interpreted as a retrogradational-progradational cycle that reached open-ocean conditions, but remained within the photic zone, during maximal retrogradation. While the retrogradational phase is represented by the Touril deposits, the progradational phase is formed by the tuffs of the Pico Alto Complex. The basal breccia at Figueiral (facies 1 and 2) is interpreted as a debris flow with short transport distance (no rounding of angular clasts). Clast composition indicates a provenance from the Anjos Complex basement lavas. Because the lower part of this unit lacks indicators for submarine deposition, a subaerial depositional environment is suggested. In the course of a subsequent rise in (relative) sea-level (retrogradation), the debris flow was reworked at its top (facies 2), most likely in a shallow-intertidal, high-energy marine environment, as indicated by the occurrence of rounded pebbles and rhodoliths. A carbonate factory developed and the bioclastic pack- to floatstones, which form the matrix of the volcaniclastic breccia, were deposited. During the ongoing retrogradation, a second debris flow likely formed the ‘transition zone’ sediments (facies 3). Marine sedimentation continued afterwards and the graded float- to grainstone units (facies 4 and 5), exposed in the Figueiral quarry, were deposited. These sediments represent the continuing transgression and a deeper, probably open-ocean environment, during which the input of siliciclastic sediment from emerged land was limited. The overlying tuffaceous coquina horizon was probably deposited as a debris flow, based on the lack of sedimentary structures and shell orientation. The incorporated tuffs indicate a first, short period of submarine eruptive activity. During the subsequent eruptive recreation phase, the overlying amphestigenid-grain- to rudstone horizon (facies 7) formed. The horizon represents the top of the Touril Complex and was likely deposited during the maximal retrogradation of the sequence exposed at Figueiral. Abundant phototrophic organisms indicate that deposition took place within the photic zone. According to Kirby et al. (2007), the limestone unit at Pedreira do Campo probably formed on a shallow bank or shoal far from any subaerial sources of terrigenous sediment, due to the dominance of photosynthetic organisms (rhodoliths and symbiont-bearing benthic foraminifera) and the absence of fine-grained terrigenous material. The small size of the basaltic cores of the rhodoliths, also points to some distance to the shore (Santos et al. 2012). These conclusions were supported by cathodoluminescence studies done on rhodoliths from Figueiral and Pedreira do Campo (Rebelo et al. 2016b), with both sites interpreted as being located far from the islands’s palaeo-shore (at about 2.9 and 2.8 km, respectively). At Pedreira do Campo, at least, a period of consolidation and erosion of the marine sequence took place before burial by the extrusion of the surtseyan tuffs of the Pico Alto Complex. The planar bedding at Pedreira do Campo might suggest their formation in a shoreface environment (Kirby et al. 2007). With the onset of tuff deposition, which cut off the previous carbonate factory on the shelf, sediment supply to the shelf was tremendously increased and the sedimentary system switched from retrogradation to progradation, most likely without a significant change in relative sea level. The topmost pillow lavas at Pedreira do Campo (Fig. 3d) are absent at Figueiral, which suggests that their distribution was likely morphologically controlled and/or restricted to the vicinity of the local feeder vent.
4.2 Pedra-que-pica
4.2.1 Geological Setting
The Pedra-que-pica outcrop, situated in a coastal area at the southeastern corner of Santa Maria Island (Serralheiro 2003; Kirby et al. 2007; Ávila et al. 2015a; Fig. 7), contains a 10–11 m thick succession of marine and very fossiliferous sediments, of which only the uppermost 3–4 m are presently exposed above sea level, that lies in between basalt pillow lavas and a volcano-sedimentary sequence on top. The lower part of the outcrop is presently submerged and its area was estimated to be 23,436 m2 (Ávila et al. 2015a), whereas the intermediate part of the section is exposed on a wave cut platform, extending for ~3,179 m2 in the intertidal zone, the upper part extending below the slope deposits at the base of the present steep cliff. The basal volcanic sequence and the overlying shell accumulation (coquina) are assigned to the Touril Complex, whereas the overlying tuffites and the succeeding basalt flow are assigned to the volcanic Pico Alto Complex (sensu Serralheiro et al. 1987, 1990; Ávila et al. 2015a; Fig. 6). 87Sr/86Sr isotope age estimates of three mollusc shells collected from the coquina yielded an average age of 5.51 ± 0.21 Ma (Kirby et al. 2007); the age of the deposit, however, is slightly younger given that the sediments intercalate basaltic pillow lavas yielding 40Ar/39Ar ages of 4.78 ± 0.13 Ma and 4.13 ± 0.19 Ma respectively, for the base and the top of the sequence (Ramalho et al. 2017). The base of the section is formed by limited remains of strata that predate the coquina, which can be found in spaces in between the pillow lavas (facies 1), forming sandstone pockets composed of fine-grained, light grey calcarenites, where trace fossils belonging to Macaronichnus segregatis Clifton and Thompson 1978 are abundant (Ávila et al. 2015a; Uchman et al. 2016). The pillow basalts show an erosional top. A laterally discontinuous 0.5–1 m thick conglomerate (facies 2; Fig. 6), composed of volcaniclastic pebbles and cobbles in a volcaniclastic calcarenite matrix, unconformably overlies the lavas. The conglomerate grades upwards into a 3–4 m thick unstructured coquina (facies 3; Fig. 6). The very poorly-sorted coquina-rudstone is rich in large, mainly disarticulated valves of bivalves (dominated by pectinids, ostreids and spondylids), and also contains echinoids, barnacles, brachiopods, bryozoans, calcareous algae (rhodoliths), bryoliths (nodules entirely composed by bryozoans), corals, and rare teleost fish teeth and shark teeth, rare gastropods (e.g., moulds and stomatofossils of Persististrombus coronatus; Ávila et al. 2016a), and whale bones (Kirby et al. 2007; Madeira et al. 2011; for a detailed list of species, see Ávila et al. 2015a). Volcaniclastic calcarenites, composed of moderately sorted, predominantly sand-grade skeletal bioclasts (fragments of molluscs, echinoids, coralline algae, bryozoans and planktonic foraminifers) and well-rounded volcaniclasts (coarse palagonitic ash and tachylitic to lithic clasts) form the matrix of the coquina. The majority of larger pectinid shells (e.g., Gigantopecten with shell length over 10 cm) is oriented in concave-down position, whereas shells of other smaller bivalve species showed a chaotic disposition of the disarticulated valves (Ávila et al. 2015a). Most shells are encrusted by bryozoans, balanids, oyster serpulids and coralline red algae, and bioeroded (Gastrochaenolithes isp. and Entobia isp.) at external shell surfaces. The preservation, however, can still be considered as good. The top of the coquina exhibits a fining-upward trend and is succeeded by coarse-grained, faintly bedded volcaniclastic calcarenites (Fig. 6). The arenites are ~10–60 cm thick and show a subhorizontal, very regular, bioturbated top. The most common trace fossil is Asterosoma isp. Bichordites isp. burrows occur about 1–2 cm below the top of this unit in the eastern part of the outcrop. The topmost 10 cm contain abundant spines of the echinoderm Eucidaris tribuloides (Lamarck) and Porites sp. coral fragments. The arenites are abruptly overlain by a 36 m thick unit of well-stratified, fine- to coarse-grained vitric ash to lapilli tuff, generated by a nearby surtseyan eruption (facies 4; Ávila et al. 2015a; Fig. 6). The tuffs show water-escape structures in the basal 20 cm. The sedimentary structures within the tuffs include thin planar lamination to medium-thick bedding, low-angle cross-bedding and internal erosive surfaces where the planar bedding is discordant. Multiple reverse and normal graded beds are also present. Angular lava blocks and basalt pebbles are rarely scattered in this unit. Pebble- to boulder-sized lithic clasts of sedimentary origin (bioclastic tuffaceous sandstone to almost pure limestone) are subordinately present within the lowermost layers of the tuffs. Locally, the tuffs are truncated by a Pleistocene shore platform and beach deposit, and show a thickness of ~4 m only. The erosional unconformity and the overlying 0.5–1 m thick boulder conglomerate (facies 5; Fig. 6), however, pinch out ~50 m to the east, where a wave-cut notch is developed. This conglomerate is topped by a >20 m thick, wedge-shaped talus deposit, consisting predominantly of boulder breccias (facies 6; Fig. 6). Beyond the erosional unconformity, the tuffs reach a thickness of about 32 m and show less distinct bedding and more volcaniclastic boulders and blocks (Kirby et al. 2007). The tuffs are overlain by a ~0.5–1 m thick conglomerate horizon. A faint erosive relief occurs along the contact between this conglomerate and the overlying basalt flows. The passage zone between the foreset unit of pillow lavas and hyaloclastites, and the overlying topset of flat-lying subaerial lavas occurs at an altitude of ~55 m within these basaltic flows (Ramalho et al. 2017).

Pedra-que-pica outcrop. a Aerial view of the southeastern shores of Santa Maria Island (photo by Paulo Henrique Silva/SRAM). b View from the sea off Pedra-que-pica outcrop area, just before loading areas
4.2.2 Interpretation and Discussion
The Pedra-que-pica sedimentary succession is interpreted as a retrogradational-progradational cycle, of which the retrogradational phase is represented by carbonates that were deposited during a transgressional period of rising relative sea-level, while the progradational phase is documented by tuffs. During the late retrogradation, a coquina was formed, which is interpreted as the result of a succession of several debris-fall deposits (sensu Titschack et al. 2005) whose redeposition was triggered by major storm events that removed the sediments from its original nearshore setting to a local depocenter below fair-weather wave base. The retrogradation phase was prematurely ended by the rapid progradational deposition of the tuffs, which filled up the available accommodation space on the shelf.
The basal pillow basalt (facies 1) indicates submarine volcanism. The overlying pockets of bioturbated calcarenite that include the trace fossil Macaronichnus segregatis indicate intertidal to shallow subtidal conditions (i.e., upper foreshore). With rising sea level, a carbonate factory development and a coquina with a volcaniclastic calcarenite matrix was deposited in a local depocenter. The coquina exclusively comprises allochthonous, poorly-sorted components, which exhibit a low degree of fragmentation/reworking. Thus, the fossil assemblage within this deposit indicates a proximal shallow-marine provenance and deposition of the coquina below fair-weather wave base, where low-energy conditions prevented reworking and shell fragmentation (cf. Ávila et al. 2015a). The occurrence of few articulated Gigantopecten shells, together with the low degree of shell fragmentation and sorting, indicate a proximal provenance of the coquina constituents. The volcaniclastic calcarenite matrix might suggest that Pedra-que-pica was close to emerged land (Kirby et al. 2007). Increased bioturbation towards the top of the overlying calcarenite corroborates a retrogradational (deepening-upward) trend in that the calcarenites represent the deepest environment of the Touril sequence. The sharply developed transition to the overlying tuffs (facies 4) indicates an abrupt change in sediment supply, which is related to the onset of a new volcanic phase in this area (Ávila et al. 2015a). The limestone clasts within the tuffs are classified as “accidental clasts”, which were produced by the disruption of pre-existing and already consolidated subvolcanic rocks on the seafloor during the explosive hydromagmatic volcanic eruption. A continuous and voluminous supply of pyroclastic material in a relatively short period of time likely caused a switch in the sedimentary system from retrogradational to progradational, which filled up the available accommodation space. Most likely, no significant changes in relative sea level occurred during that period. The absence of ballistic emplacement structures of volcanic bombs (e.g., impact pits, bomb sags) suggests a submarine origin of these pyroclastics, classifying them as surtseyan tuffs or water-settled tuffs, formed by hydromagmatic volcanic activity in shallow waters (Fisher and Schmincke 1984).
The subaquatic deposition of this unit is further corroborated by the common occurrence of cross-bedding and small cross-cutting channels, as these sedimentary structures are interpreted as secondary structures related to the marine environment and the resedimentation of the tuffs. The unconformity at the top of the tuffs at an altitude of ~35 m and the overlying conglomerate indicate erosion (although minor) and reworking in shallow waters and thus formation in a high-energy environment. The tuffs, therefore, describes a progradational trend, caused by an extreme enhancement in sediment supply and the rapid fill up of the local depocentres. Following the formation of the conglomerate on top of the tuffs, another period of volcanism emplaced the overlying basaltic lava flows that occur at altitudes between ~32–60 m. Low relief, which indicates a hiatal surface due to sediment reworking, characterizes the transition between the latter units. Pillows and palagonitised hyaloclastites are developed in the basal part of these lavas, indicating submarine volcanism. The submarine lavas are immediately topped by flat-lying massive subaerial flows. The transition between submarine and subaerial morphologies occurs at an altitude of ~55 m above present sea level and represents a palaeo-sea-level marker (Meireles et al. 2013; Ramalho et al. 2017). The conglomerate (facies 5) overlying the erosional unconformity (wave cut bench) carved on the lowermost tuffs (facies 4) is interpreted as a Pleistocene boulder-beach conglomerate, most likely formed during the Marine Isotopic Stage 5e (MIS 5e) interglacial period (Ávila et al. 2008a, 2015b). Breccias above the conglomerate are interpreted as multi-phase slope deposits (facies 6). Modern (Holocene) counterparts to both the latter Quaternary deposits (facies 5 and 6) occur at the footslope of the cliff east of Pedra-que-pica.
4.3 Ponta do Castelo
4.3.1 Geological Setting
Ponta do Castelo constitutes the southeastern-most headland on Santa Maria Island and comprises a typical lava delta sequence, whose bottom-set unit comprises marine fossiliferous sediments. The overall sequence is thought to be Early Pliocene in age and is attributed to the Pico Alto Volcanic Complex (Serralheiro et al. 1987). It is comprised from the base to the top (see Meireles et al. 2013) by: (1) fossiliferous marine sediments with a high volcaniclastic content that partially correspond to the remobilization of tuffs and water-settled tuffs (the same that cover the Pedra-que-pica outcrop) from the surtseyan cone whose remains can still be seen farther west at Rocha Alta; (2) a steeply dipping foreset unit of pillow lavas and hyaloclastites, prograding to the E; (3) a topset unit of flat-lying subaerial lava flows; and (4) pillow lavas of another lava delta stacked on top of the previous, but whose topset unit was locally removed by erosion. The passage zone between the forests of submarine lavas and the overlying subaerial lava flows of the main lava delta is presently located at ~55 m a.s.l and marks the contemporaneous sea level. It is thus possible to infer very accurately the palaeo-water depth for the deposition of the bottomset sediments. The exposed sedimentary body at the base of the sequence exhibits a sigmoidal or wedge-shaped geometry, and comprises a set of 4 or 5 sandstone units amounting to a total thickness of 9 m. A complete and detailed description for this sequence can be found in Meireles et al. (2013), from where the following summary was extracted. The first 1 m thick unit is composed of coarse- to medium-grained sandstones that show large-scale cross-stratification with centimetre- to decimetre-thick, slightly wavy sets, forming low but wide swales and possibly hummocks. This unit is eroded at the top and exhibits a centimetre- to decimetre-high palaeo-relief. These sediments are unconformably overlain by a wedge-shaped sedimentary unit that gradually thins towards the ENE, from about 1.2 to about 0.5 m. The sediments grade from very coarse-grained sandstones at the base, into coarse- to medium-grained, chaotic-bedded sandstones in the middle (both very rich in shell debris), and medium- to fine-grained sandstones with plane-parallel bedding and small-scale bioturbation at the top. A shallow but wide erosional channel unconformably cuts through the topmost plane-parallel beds and marks the top of this unit.
The subsequent up to 1.5 m thick sedimentary unit comprises medium-grained, poorly-sorted fossiliferous sandstones that exhibit diffuse bedding. These sediments fill the above-mentioned erosional channel. Rip-up clasts that originate from the previous unit occur within the channel-fill sandstone. Fossils and shell debris are generally dispersed in random orientation and remains of solitary corals and rock-encrusting bryozoan colonies that were ripped from their hard-substrate occur “floating” within the sandstones (Björn Berning, pers. comm.).
A pronounced erosive unconformity deeply cuts through all previous units, forming an irregular palaeo-relief characterised by side-by-side channels up to 3–4 m deep and about as wide. This palaeo-relief was filled and overlain by a 4–5 m thick unit of fossiliferous, medium-grained sandstones exhibiting 1–3 m wide, 0.3–1.5 m high slightly asymmetric hummocks and swales that exhibit onlapping contacts along the steep channel sides; beds typically fan out from conformable almost-parallel fine beds, internal truncation surfaces are rare, and hummocks are almost ubiquitously preserved. Towards the top bedding becomes almost plane-parallel, with occasional smaller-scale ripples. Fossil content is composed of bivalves, gastropods, echinoids, fragments of endemic barnacles, and remains of rock-encrusting bryozoan colonies that were ripped from their hard-substrate; shells and shell fragments are generally dispersed within the sediment and show no preferred orientation.
Above the previous sediments and bounded by two faint erosive surfaces, a 0.5 m thick unit of unstratified/massive, fossil-rich, medium- to coarse-grained sandstones occurs. This unit is very rich in microgastropods, bivalves, fragments of barnacles, bryozoans, echinoids and rhodoliths. Shell debris is randomly/chaotically distributed within these sediments, and exhibits no preferential orientation.
The topmost sedimentary unit is about 3.7 m thick and shows vertical fining upward from medium- to fine-grained fossiliferous sandstones. These exhibit a faint cross-stratification at the base that gradually passes to an almost imperceptible plane-parallel sparse stratification. The uppermost ~1.5 m show abundant bioturbation by burrowing organisms (Thalassinoides isp., Diplocraterion isp., Crossopodia isp., Ophiomorpha isp. and Rhizocorallium isp.; Meireles et al. 2013). Basaltic pillow lavas of the overlying lava delta cap the sequence, imprinting load casts in the once soft sediments. These were dated to 4.13 ± 0.19 Ma (Ramalho et al. 2017).
4.3.2 Interpretation and Discussion
The sediments at Ponta do Castelo are interpreted as tempestite deposits that represent a rapid succession of 4 or 5 individual energetic events. Thereby, each event caused massive sediment transport from shallow waters to greater depths with intense bottom erosion and rapid redeposition along the steep shelf of Santa Maria by unidirectional and/or combined flows. Final deposition took place above or close to storm wave base (Meireles et al. 2013). The first unit is interpreted as a tempestite formed under wave oscillations (or combined flows) above or close to storm wave base, as suggested by the presence of swaley and hummocky stratification. The second unit is thought to represent a density-induced flow that brought sediments from shallower levels (as suggested by the presence of allochthonous littoral faunas), initially eroding the sea-bed and eventually settling as it gradually lost energy. After a short period of fair weather—attested to by the presence of bioturbation—another tempestite was deposited with strong basal erosion, creating a channel, which was subsequently filled. The existence of well-preserved bryozoan colonies in the sediment, which were ripped from their shallow-water rocky substrate, attests the vigorous energy of the event and the short transport distance from the shallow-water/littoral environment. Another even more massive event followed the previous one, cutting deeply into the existing sequence and resulting in the very rapid deposition of the fourth unit. The resulting deeply irregular palaeo-relief (with 3–4 m deep channels) attests the vigorous erosion necessarily associated to very energetic bottom currents. In contrast, the distinct mega-hummocky stratification with preserved metric, slightly asymmetrical hummocks with rare internal truncations, attests to the subsequent deposition under extremely high aggradation rates—perhaps corresponding to a deposition time of just a few hours—as a result of combined flow conditions above or at storm wave base (see Dumas and Arnott 2006; Meireles et al. 2013). The generation of such a deep basal erosional unconformity and the deposition of sediments under extremely high aggradation rates is interpreted as the result of strong downwelling currents (combining wave oscillation with unidirectional flow dynamics) associated with a very violent storm event that remobilised, transported and deposited large amounts of littoral sediments onto a deeper part of the shelf (Meireles et al. 2013). Finally, the last sedimentary unit represents the transition to the fair-weather suite. The intense bioturbation by burrowing organisms in the uppermost part of the sequence is related to the biological colonization that typically occurs at such depths (~50 m). Sedimentation was subsequently interrupted by the rapid progradation of a costal lava-fed delta sustained by an eruption on land. The prograding lavas thus preserved the underlying strata from subsequent erosion (Meireles et al. 2013). As the basaltic pillow lavas of the overlying lava delta imprinted load casts in the once soft sediments, the volcanic sequence is penecontemporaneous of the underlying sediments.
The sequence at Ponta do Castelo thus suggests that storm wave base in steep and narrow insular shelves may reach down to depths of 50–60 m and that deposition in this setting is mainly controlled by storm-events, during which sediments are being remobilised across the shelf, from the nearshore to the offshore, by strong ebb-return currents (Meireles et al. 2013).
4.4 The Palaeontological Significance of Carbonate Sequences in Reefless Volcanic Oceanic Islands
Sedimentary systems on reefless volcanic oceanic islands are subject to multiple controls, which differ considerably from other open shelf systems (e.g. Reuter and Piller 2011 and references therein), even when they are also influenced by volcanic activity (Wilson and Lokier 2002). The general steep relief, the limited sediment supply by river systems (Ávila et al. 2008b), the windward/leeward asymmetry in energy conditions, the influence of volcano-tectonic processes (uplift/subsidence; cf. Ramalho et al. 2013, 2017), frequent eruptive events and their related processes with localised ash-tuff-lava flow input result in small depocentres with a high facies variability and potentially even with an individual depositional evolution controlled by the input of volcanic material. Furthermore, volcanic events might influence carbonate systems by: (1) covering partly or completely the carbonate producing organisms, which results in physical stress, tissue necrosis and/or death of these organisms (Fortes 1991; Heikoop et al. 1996a, b; (2) decreasing light penetration when suspended in the water, thus affecting photosynthetic communities (Short and Wyllie-Echeverria 1996; Wilson and Lokier 2002); and (3) changing the water chemistry (Frogner et al. 2001; Ralph et al. 2006; Duggen et al. 2010). Additionally, shelves at reefless oceanic islands are frequently exposed to the full force of recurrent storms that inevitably disrupt the sedimentary cycle and frequently force the remobilization, transport and redeposition of existing sediments (see Meireles et al. 2013 and references therein). All these processes should be taken into account when interpreting carbonate sequences at volcanic islands.
In conclusion, the early Pliocene carbonate deposits of Santa Maria highlight the importance of the interaction of different processes for the development and evolution of a carbonate system on reefless volcanic oceanic islands (the present-day dominant calcitic mineralogy of the carbonates at Pedra-que-pica, Pedreira do Campo and Figueiral is regarded as a taphonomical artefact and is attributed to the diagenesis of the aragonitic shells). Besides volcanic quiescence, as already suggested by Wilson and Lokier (2002), a rise in sea-level that provides accommodation space on the steep island shelf is generally necessary for the development and preservation of carbonate factories. Hence, marine deposits on reefless volcanic oceanic islands should be predominantly formed and preserved during transgressions and sea-level highstands, and their structure and distribution frequently reflect the highly energetic environment in which they were formed. Comparable to other structured island shelves, the high variability in volcanic island shelf morphology results in a complex pattern of small local depocentres (patchy outcrop patterns in sedimentary prisms), enhances the number of microhabitats and biocoenoses, and provokes a highly discontinuous facies variability (see also Titschack and Freiwald 2005; Titschack et al. 2005, 2013). In reefless insular shelves, sedimentary bodies typically have small residence times as these almost invariably get to be remobilised by storm-induced processes such ebb-return currents, onto deeper waters (Quartau et al. 2012, 2014, 2015; Meireles et al. 2013) or across the shelf (Johnson et al. 2017). Combined with the island’s narrow and steep shelf, which provides little space for the deposition of sediments, the remobilisation of the transgressive/highstand deposits during subsequent regressive trends over the shelf edge into adjacent deep-sea basins, seems to represent their most likely fate (Ávila et al. 2008b; Ávila 2013). However, eruptive events, which generally cut off marine carbonate factories and interrupt marine sedimentary processes, might locally prevent the erosion of these marine carbonate deposits by their rapid and abrupt burial, as in the cases of Ponta do Castelo, Malbusca (see Rebelo et al. 2016a), and of the other above-mentioned outcrops. Consequently, volcanic events might limit carbonate production but might on the other hand be responsible for the effective fossilization and preservation of these deposits on reefless volcanic oceanic island shelves.
5 The Pleistocene MIS 5e Outcrops
Scattered around the shores of the island of Santa Maria, wave-cut platforms, wave-cut notches, and the remains of former boulder and sandy beaches (Fig. 8) are preserved at altitudes ranging from 3–4 up to 9–10 m above present mean sea level (Ávila et al. 2015b). These outcrops are interpreted as a series of sea-level stands and their formation during the last interglacial maximum sea level, hence the MIS 5e (Marine Isotopic Stage 5e, ~120–130 ka in age) is indicated (Ávila et al. 2008a, 2009a, 2010). Some of these outcrops are very rich in marine fossils. The best studied outcrops are Prainha and Praia do Calhau (both on the southern shores; Zbyszewski and Ferreira 1961; García-Talavera 1990; Ávila et al. 2002, 2007, 2008a, 2009a, 2015b; Amen et al. 2005; Ávila 2005), Vinha Velha (also located on the south; Ávila et al. 2015b) and Lagoinhas (northern shores of Santa Maria; Callapez and Soares 2000; Ávila et al. 2002, 2007, 2009a, 2015b; Ávila 2005; cf. Fig. 1b). Other known Pleistocene outcrops, some of them not yet studied nor dated are: a small outcrop west of Pedra-que-pica (see Ávila et al. 2015b), Ponta do Castelo, and Ponta do Cedro.

Indicators of the last interglacial maximum sea-level (MIS 5e). a Wave-cut platform at Prainha (+4 m). b Boulder conglomerates lined up at Lagoinhas (+6 to +7 m). c The MIS 5e sea-level nearby Vinha Velha (note the boulder conglomerates, and the wave-cut notch at the base of the palaeocliff (bottom right). d The MIS 5e sea-level at Ponta do Castelo (+4 to +5 m). e Wave-cut notch at Baía da Rocha Alta (+8 m). f Wave-cut notches at Ponta do Pesqueiro Alto (+7 to +10 m). g The MIS 5e deposit at Ponta do Cedro (+6 to +7 m). h The MIS 5e deposit at Vinhas do Sul (+3 to +4 m)
The Prainha outcrop was first reported by Zbyszewski and Ferreira (1961) and later by García-Talavera (1990), with all authors paying special attention to the marine molluscs. Callapez and Soares (2000) discovered the Lagoinhas deposit and published a palaeoecological interpretation of this outcrop. Ávila et al. (2002) redescribed the stratigraphic sequence of both Lagoinhas and Prainha and revised the marine molluscs’ checklist, whereas Amen et al. (2005) wrote a detailed account of the fossil coralline–algal framework of Prainha. Ávila (2005) and Ávila et al. (2007, 2009a, b, 2015b) reviewed the Pleistocene mollusc fossils of Prainha and Lagoinhas. Based on U/Th ages on shells of Patella spp., and on faunal, facies and geomorphological considerations, Ávila et al. (2008a) assigned the basal conglomerate of the Prainha outcrop to Marine Isotopic Substage 5e (MIS 5e). Due to taphonomic problems and the wet climatic conditions prevailing on the island, the U-series measurements from their fossilised remains can only help constrain the age of the uranium taken up diagenetically (Ávila et al. 2008a, 2009a). U-series data from Santa Maria Island indicate an open system, providing mean 230Th ages that are much younger than the true age of the unit. They suggest relatively steady, more or less continuous, diagenetic U-uptake by the fossils since their deposition in the Pleistocene (Ávila et al. 2009a). Therefore, accepting an age of 120–130 ka for the deposits, Ávila et al. (2008a) estimated that within the embedded biogenic carbonates U-uptake rates varied from c. 2.5 to 5 μg gCaCO −13 kyr−1. This was based on 230Th-ages in the 60–70 ka range, for shells that would have started accumulating diagenetic U some 130–120 ka ago (i.e. during the MIS 5e; Ávila et al. 2008a).
5.1 Prainha and Praia do Calhau
These sections consist of unconsolidated shallow-water deposits overlying a basement formed by basaltic lavas (facies 1; Fig. 9). The bases of the outcrops are formed by irregular shore platforms on top of the ankaramitic basalts of the Anjos Complex, which occur at an altitude of 3–4 m (cf. Figs. 9 and 10; Serralheiro et al. 1987; Serralheiro 2003), and exhibit a lateral extent of ~800 m. Roughly circular depressions, 2.7–7.0 cm in diameter (mean = 4.26 cm, SD = 1.01, n = 48), were observed at this surface (Fig. 10e). Ávila et al. (2009a) attributed these structures to bioerosion traces of epilithic sea urchins and Ávila et al. (2015b) assigned them to the ichnofossil Circolites kotocensis Mikulás. The traces were recently destroyed, after Hurricane Gordon passed the Azores, in August 2012. The unconformity is overlain by a 40 cm thick, strongly cemented conglomerate (facies 2; Figs. 9 and 10a). A calcareous coralline algal biostrome (facies 3; Fig. 9) with a maximum thickness of ~50 cm covers the conglomerate or, locally, directly on the unconformity on top of the basalts. This biostrome is built by encrusting and warty to fruticose non-geniculate Corallinales (Rhodophyta) growing one over the other. Fragments of mollusc shells, bryozoans, and echinoderms, are accessory components. Amen et al. (2005) reported four species of Corallinaceae (Rhodophyta): Spongites fruticulosus Kützing (the main builder of the algal framework), Lithophyllum incrustans Philippi, Neogoniolithon brassica-florida (Harvey) Setchell and Mason, and Titanoderma pustulatum (Lamouroux) Nägeli. Of these, only T. pustulatum is recorded for the recent algal flora of the Azores (Neto 1994). The algal crust abundantly shows macrobioerosion structures, mostly clavate borings, assigned to the ichnogenus Gastrochaenolites Leymarie. Remains of the producer of these borings, the endolithic bivalve Myoforceps aristatus (Dillwyn), may still be found in situ inside most of the borings (Fig. 11a). At least two species of Corallinaceae described in this facies appear with a superimposed growth in thin sections (Fig. 11b). The inter-skeletal spaces in the algal framework and bioerosion structures were later filled by two internal sediments (Fig. 11c). The basal one consists of angular, poorly sorted rock fragments, marphic mineral grains’, and bioclasts (foraminifers, echinoids, coralline algae and scarce molluscs) of medium- to coarse-grained size in a micritic matrix. The succeeding internal sediment is richer in micrite. The conceptacles of the red algae are filled with spherulitic aragonite cement. These cements are slightly weathered, with limited superficial dissolution of aragonite. Locally, towards the top of the biostrome, accumulations of iron oxide micro-nodules occur between coralline thalli. Traces of iron oxides and interruptions in algal growth are observed in thin sections. In some places, where the thickness of the biostrome is enhanced, 3–4 discontinuity surfaces are macroscopically observable by the increased presence of bioerosive structures at these surfaces (Ávila et al. 2009a). Ávila et al. (2002) reported fractures in the algal crust (facies 3) that were initially attributed to “subaerial exposure and desiccation”. Ávila et al. (2009a) reinterpreted these fractures on the upper surface of the algal framework and along the entire vertical section of the fossil algae as the result of an extensional process fracturing the algal framework after its formation. Fracturing by local extension can take place underwater with no subaerial exposure. In fact, formation of neptunian dykes (open fractures filled by marine deposits; cf. Fig. 10c) by extension is a common process in submarine settings (Winterer and Sarti 1994).


The Pleistocene deposits at Praia do Calhau and Prainha. a One of the most representative sections studied at Praia do Calhau: over an irregular shore platform carved on top of the ankaramitic basalts of the Anjos Complex, a strongly cemented conglomerate was stabilised by a calcareous coralline algal biostrome. b Partly cross-lamination sands that overlain the calcareous coralline algal biostrome, covered by colluvial-alluvial deposits. c Fossil coralline algal crust with abundant signs of bioerosion by endolithic bivalves (clavate borings), and neptunian dykes (large fractures affecting the algal framework), filled with sand; both structures are present along the entire vertical section of the fossil algae. d The algal framework at Prainha. e Bioerosion traces of epilithic sea urchins, probably made by Paracentrotus lividus (Lamarck) and assigned to the ichnofossil Circolites kotocensis Mikulás

a At Prainha, the endolithic bivalve Myoforceps aristatus (Dillwyn) is sometimes found in situ inside some of its borings. b, c Thin section microphotographs of the algal framework collected at Prainha. d, e Thin section microphotographs of the bioclastic sand sediments collected at Prainha
The biostrome (facies 3) is overlain by 1.3 m thick, yellowish, partly cross-laminated, volcaniclastic to bioclastic, uncemented sands (facies 4; Figs. 9 and 10b) that also fill most of the fractures penetrating facies 3 (cf. Fig. 10c). Carbonate contents range from 56.2 to 67.3% (in the samples investigated) (Fig. 10b). The grain-size distribution is dominated by the 125–250 μm fraction. The bioclasts consists almost exclusively of small mollusc fragments. Lenses with ripple marks, trace-fossils and/or root casts are preserved locally (Ávila et al. 2002) and are better lithified. The sediments correspond to a beach foreshore (intertidal) facies and show cross-lamination (cf. Fig. 10b). A thin carbonate crust of pedogenic origin occurs at the top of the bioclastic sandy sediments (facies 5, Fig. 9). The crust mostly consists of micrite that precipitated together with clay and other silt-sized impurities and exhibits a clotted texture (Fig. 11d). Sediment below the carbonate crust (Fig. 11e) includes bioclasts (molluscs, echinoid spines, branching coralline algae), poorly-sorted volcanic grains and very poorly-sorted rock fragments. Aeolian dunes consisting of a mixture of biogenic sand particles derived from molluscs, echinoderms, bryozoans and coralline algae with black, basaltic sand (facies 5; Fig. 9) and colluvial–alluvial deposits (facies 6 and 7; Fig. 9) cover the carbonate crust (Ávila et al. 2009a).
5.2 Lagoinhas
The Lagoinhas outcrop is about 100 m wide and its base lies ~7.4 m above present sea-level (Ávila et al. 2002, 2009a). The base is formed by an irregular shore platform on top of the ankaramitic basalts of the Touril Complex (facies 1; Fig. 9; Serralheiro et al. 1987; Serralheiro 2003). A ~10 cm thick, strongly cemented basal-conglomerate (facies 2) passes laterally and upward into a ~30 cm thick calcareous red algal biostrome (facies 3). The coralline algal framework is similar to the one found at Prainha, but was not yet studied in detail. The faunal assemblage is dominated by molluscs, with bryozoans and echinoderms as accessory components. As at Prainha, macrobioerosion structures in the biostrome are assigned to the ichnogenus Gastrochaenolites Leymarie and were produced by the endolithic bivalve Myoforceps aristatus (Dillwyn) (Ávila et al. 2009a). The overlying white-yellowish unconsolidated sands (facies 4) exhibit thicknesses of up to ~0.7 m and are rich in very-well preserved fossil assemblages, which are dominated by molluscs. The grain-size distribution of these sands is dominated by the 250–500 μm fractions. A thin carbonate crust of pedogenic origin occurs at the top of these bioclastic sands (cf. Fig. 9). This crust is covered by colluvial deposits (facies 5; Fig. 9).
5.3 Interpretation and Discussion
Although structures similar to the Prainha and Lagoinhas coralline algal biostromes are not known from other islands of the Azores, their occurrence is well documented from shallow-water rocky substrates in the Mediterranean Sea (e.g., Pérès and Picard 1964; Hofrichter 2001) and Western Atlantic, where they form flat calcareous patches, stabilizing shore conglomerates (Thorton et al. 1978; Bosence 1983, 1984). These formations develop primarily on shores with a narrow tidal amplitude (Adey 1986), as is the case for all the Azores. No known direct analogues of the Prainha and Lagoinhas shallow-water fossil algal frameworks exist in other regions of the world (Ávila et al. 2009a). According to Amen et al. (2005), the algal biostromes were formed in shallow water depths, probably less than 10 m deep. Today, Lithophyllum incrustans, L. pustulatum (= Titanoderma pustulatum) and Neogoniolithon brassica-florida are most common in the shallowest infralittoral zone within the Mediterranean, while Spongites fructiculosus shows a wider depth range (Bressan and Babbini 2003). The mastophoroid genera Spongites and Neogoniolithon predominate in the Neogene algal assemblages, when the western Mediterranean experienced tropical/subtropical climatic conditions (Braga and Aguirre 2001). The Mediterranean and western Atlantic “trottoirs” are more similar as they are structures a few decimetres thick and locally developed on abrasion terraces (Thorton et al. 1978; Adey 1986). Neogoniolithon brassica-florida and Lithophyllum incrustans, abundant in the Prainha framework (Amen et al. 2005), can be main components of the Mediterranean “trottoirs”, together with Lithophyllum byssoides (frequently mentioned as Lithophyllum tortuosum) (Adey 1986).
Based on stratigraphic, petrologic, and faunal analysis of the Lagoinhas and Prainha outcrops, Ávila et al. (2009a, 2015b) suggested that during a marine transgression, an irregular shore platform on volcanic rocks developed. Subsequently, during the ongoing transgression, a conglomerate (facies 2) and the overlying red algal biostrome (facies 3) formed in a shallow-marine, high-energy (shoreface) environment (Fig. 9). The hydrodynamic setting is further supported by the presence of sea urchin burrows likely being attributable to the activity of the epilithic grazer Paracentrotus lividus (Lamarck), of which one test fragment and numerous primary spine fragments were found at Lagoinhas (Madeira et al. 2011). In the present-day, this sea urchin is extremely common on the shores of the islands of the Azores, at depths of 1–2 m, always on rocky substrate, where it produces similar shallow excavations (Ávila et al. 2009a). The bioerosion structures—Gastrochaenolites isp.—that characterize the algal reef in both outcrops are also indicative of shallow water depths (less than 5 m). The endolithic bivalve Myoforceps aristatus, found in life-position, is the producer of these structures. Today, the congeneric M. aristatus lives in shallow-marine surf benches (called “trottoirs” in the Mediterranean). Maximal transgression with minimal sediment input was reached on top of the coralline algal biostrome (top of facies 3). At the start of the subsequent regression, the poorly consolidated yellow fossiliferous sands (facies 4) were deposited. Sedimentary structures such as wave-ripple cross-bedding and cross-lamination indicate a shoreface to foreshore depositional setting. The palaeoecological evidence derived from the analysis of the bioerosion structures, the characteristics of the coralline algal assemblages, the taxonomic composition of the fossil assemblage and the bathymetrical zonation of the species represented in it, indicate a change from a transgressive marine depositional setting in shallow infralittoral (facies 2 and 3) to a regressive trend (foreshore/beach; facies 4). The regressive, shallowing upwards trend proceeds with dune sediments, palaeosols (micritic crusts) and colluvial–alluvial deposits (facies 5, 6 and 7; Fig. 9). The position of the Pleistocene sediments, not taking the apparent uplift trend of Santa Maria Island and glacio-isostatic adjustments into account, suggests a palaeo-sea-level highstand about 4–6 m above the modern sea-level. This suggests the formation of these deposits during the interglacial sea-level highstand of MIS 5e (Ávila et al. 2009a, 2015b).
6 The Fossils
Zbyszewski and Ferreira (1962b) reported 188 taxa (179 invertebrates and 9 vertebrates) from the early Pliocene deposits of Santa Maria Island. Some groups were recently revised, e.g., the Brachiopoda, the Echinodermata, the Chordata (sharks and cetaceans), and the Ostracoda (Kroh et al. 2008; Madeira et al. 2011; Ávila et al. 2012, 2015c; Meireles et al. 2012). Other groups are presently under review, such as the Bryozoa, the Cnidaria (Anthozoa) and also the Mollusca, the latter representing the most abundant fossil group. The MPB group has reviewed the Zbyszewski and Ferreira checklist and over 40 taxa of fossil molluscs need revision. Table 1 provides a revised list of species with a conservative number of 191 taxa (13 of which are considered as endemic species) from the early Pliocene outcrops of Santa Maria. With 99 taxa accepted as valid, the Mollusca exhibit the highest number of species. Of these 99 taxa, 7 are considered as endemic to Santa Maria.
Regarding the Pleistocene (MIS 5e) outcrops, Ávila et al. (2015b) reported 136 species of marine molluscs. Twenty-two of the gastropods are endemic, and all of them still live in the waters of the Azores. Considering all groups, this chapter updates the latest checklists and increases the total number of Pleistocene species to 146 (Table 1).
6.1 Fossil Algae
The Pleistocene layer of calcareous algae described in detail in Sect. 5.1 was first reported by Berthois (1953b, c) and by Zbyszewski and Ferreira (1961) who named this specific layer as “Lithothamnium”. The first phycological study of the algal components of the fossil framework of Prainha was made by Amen et al. (2005) who, based on morphological and anatomical characters, as well as reproductive structures, reported 4 species: Spongites fruticulosus Kützing (the most abundant algal species), Lithophyllum incrustans Philippi, Neogoniolithon brassica-florida (Harvey) Setchell and Mason, and the rare Titanoderma pustulatum (J.V. Lamouroux) Nägeli. No Lithothamnion species were found at Prainha and structures similar to these algal frameworks are not known in the present times on the Azores (Amen et al. 2005).
The calcareous fossil algae present on rhodoliths were recently studied by Rebelo et al. (2014, 2016a, b), who reported 6 taxa. Her fieldwork reports fossil rhodolith forming coralline algae from all outcrops but Cré (Ana Rebelo, pers. comm).
6.2 Fossil Vertebrates
The Azores are well-known as one of the best spots for whale-watching nowadays, with 28 species of cetaceans reported around the archipelago (Ávila et al. 2011). In Santa Maria, the fossils of whales have been known for a long time, being called “ossos de gigante” (bones of giants) by the inhabitants of the island. However, the first published information on this matter was done only in the 19th century by Boid (1835) who wrote “In a part (…) (of the) N.W. side scarcely accessible, is to be seen an immense fossil thighbone”. This bone, located at Ponta do Pesqueiro, was later identified by Bedemar (1837) as a whale bone (Estevens and Ávila 2007). Other fossil fragments of cetacean vertebrae and ribs, reported by subsequent authors (Cotter 1888–1892; Teixeira 1950; da Ferreira 1955; Zbyszewski and Ferreira 1962b) were never the subject of a proper study in order to identify these remains.
6.2.1 Cetaceans
The first thoroughly systematic study on the fossil whales of Santa Maria Island was done by Estevens and Ávila (2007). These authors reported 10 occurrences of fossil cetaceans belonging to two suborders: Odontoceti (Ziphiidae), with one Mesoplodon species collected at Assumada outcrop; and several bone fragments of ribs, vertebrae and other skeletal parts, assigned to undetermined Mysticeti (?Balaenopteridae). All of these samples were collected in sediments from the Touril Complex (Estevens and Ávila 2007). Albeit meager, the fossil Pliocene whales of Santa Maria were compared with contemporaneous faunas (Messinian-Zanclean) from the western Atlantic—Eastover (7.2–6.1 Ma) and Yorktown (4.8–3.0 Ma) formations from the Middle Atlantic Coastal Plain, the Palmetto Fauna (5.2–4.5 Ma) from Florida—and from the eastern Atlantic—the Kattendijk (5.0–4.4 Ma) and Lillo (4.2–2.6 Ma) formations from Belgium (see references in Estevens and Ávila 2007: 157). Remarkably, all of these faunistic associations share a noticeable modern character, being dominated by living groups such as the beaked whales (Ziphiidae) and true dolphins (Delphinidae) among the odontocetes, and the rorquals (Balaenopteridae) and right whales (Balaenidae) among the mysticetes, similar to the Azorean fossil fauna described by Estevens and Ávila (2007) (Fig. 12a–d).

Neogene fossil cetaceans from Santa Maria (Estevens and Ávila 2007; Ávila et al. 2015c). a Small rib fragment assigned to Cetacea indet (Ponta Negra). b Small rib fragment assigned to Cetacea indet (Cré). c, d Fragment of a large left rib assigned to ?Balaenopteridae indet. (DBUA-F 401) in lateral view (c) and cross-section at intermediate break (d) (Cré). e, f Large fragment (~1 m) of the mandible of a medium- to large-sized baleen-bearing mysticete (Chaeomysticeti) (MIS 5e of Prainha)
Recently, in September 2012, after the passage of the Hurricane Gordon, a large fragment (~1 m in length) from the mandible of a medium- to large-sized baleen-bearing mysticete (Chaeomysticeti) was collected at Praia do Calhau, in the Pleistocene (MIS 5e) sandy layer (Fig. 12e, f). This finding was reported by Ávila et al. (2015c) as the first world record of fossil whales for the last interglacial (MIS 5e), in outcrops located in oceanic islands, thus increasing the relevance of the fossil record of Santa Maria Island for the palaeobiogeography of the North Atlantic.
6.2.2 Selaceans
Reports of fossil shark teeth from oceanic islands are few to date. In the Atlantic, shark teeth are reported from fossiliferous deposits of Jamaica (Donovan and Gunter 2001), Puerto Rico (Nieves-Rivera et al. 2003), Cape Verde (Serralheiro 1976), and the Azores (Zbyszewski and Ferreira 1962b; Ávila et al. 2012). In the Caribbean, Cuba has 15 taxa of fossil sharks from the Miocene-Pliocene (Iturralde-Vinent et al. 1996) and 5 species are known from the Miocene of Carriacou Island (the Grenadines, Lesser Antilles; Portell et al. 2008).
Zbyszewski and d’Almeida (1950) and Zbyszewski and Ferreira (1962b) were the first to report fossil sharks from Santa Maria. Ávila et al. (2012) added three new records, increasing the number of fossil sharks to seven species: Notorynchus primigenius (Agassiz), Carcharias acutissima (Agassiz), Cosmopolitodus hastalis (Agassiz), Paratodus benedenii (Le Hon), Isurus oxyrinchus Rafinesque, Megaselachus megalodon (Agassiz in Charlesworth), and Carcharhinus cf. leucas (Valenciennes in Müller and Henle) (Fig. 13a–q). The Azorean Pliocene selachian fauna clearly differs from those described from sediments deposited on continental shelves, in which batoids (i.e., electric rays, sawfishes, guitarfishes, skates and stingrays) and small benthic sharks (e.g., scyliorhinids) are usually well represented. Several attempts were made to recover small selachian teeth by screen-washing (sieve sizes: 0.5 and 1 mm) from bulk sediment samples from most of the studied localities, but that effort yielded no teeth of batoids or small sharks (Ávila et al. 2012). The absence of batoids in the Pliocene coastal environments of Santa Maria Island is not in accordance with the diversity of the recent batoid fauna from the Azores, including about 15 species of Torpediniformes, Rajiformes and Myliobatiformes (Santos et al. 1997; Barreiros and Gadig 2011).

Neogene fossil sharks from Santa Maria (Ávila et al. 2012). a–c Carcharias acutissima (Agassiz). Upper-anterior tooth in labial (a), lingual (b) and lateral view (c) (H = 27.0 mm, W = 9.3 mm). d–f Paratodus benedenii (Le Hon) in labial (d), lingual (e), and distal view (f) (H = 52.0 mm, W = 37.0 mm). g–k Cosmopolitodus hastalis (Agassiz). g–i: labial (g), mesial (h), and lingual (i) view (H = 55.0 mm, W = 35.0 mm). j, k Lingual (J) and labial view (k) (H = 50.0 mm, W = 36.0 mm). l–n Megaselachus megalodon (Agassiz) in lingual (l), lateral (m), and labial view (n) (H = 88.3 mm, W = 74.5 mm). o, p Carcharhinus cf. leucas (Valenciennes) in lingual (o) and labial view (p). q Notorynchus primigenius (Agassiz). Lower anterolateral tooth in labial view (H = 9.8 mm, W = 23.5 mm)
During the early Pliocene, subtropical to warm-temperate seas were prevalent in the area of the Azores, as deduced from palaeontological, geological and isotopic data, all indicating a warmer climate than today (Kirby et al. 2007; Ávila et al. 2015b). Five species are extinct nowadays: C. acutissima, C. hastalis, N. primigenius, M. megalodon and P. benedenii. All these species disappeared by the end of the Pliocene, including N. primigenius, formerly thought having went extinct during the Miocene (Ávila et al. 2012).
6.3 Fossil Invertebrates
6.3.1 Mollusca
Molluscs are by far the most diverse and abundant fossil group from the outcrops of Santa Maria (see Table 1). The particularly good fossil record of molluscs can be explained by a combination of initial abundance, taphonomic biases and selective investigations (most palaeontological studies on Santa Maria during the 19th and 20th centuries focused on molluscs).
Notwithstanding the many studies on the early Pliocene deposits (Bronn 1860; Hartung 1860; Reiss 1862; Mayer 1864; Cotter 1888–1892; Ferreira 1952, 1955; Zbyszewski and Ferreira 1962b), the fossil molluscs have not been recently studied systematically/taxonomically, with the exceptions of Janssen et al.’s (2008) paper on the early Pliocene heteropods and pteropods, and Habermann’s (2011) study on the bivalve Gigantopecten latissimus (Brocchi). Much material has been collected since the beginning of the international workshops “Palaeontology in Atlantic Islands”, in 2002, with a special emphasis on the micromolluscs. The revision of the 105 historically reported taxa and the formal description of many new species (Ávila, unpublished data) are of utmost importance and present the major research task of the MPB for the next years.
For the Pleistocene (MIS 5e) malacofauna, Ávila et al. (2002) reviewed the old literature (Berthois 1951, 1953b; Zbyszewski and Ferreira 1961; García-Talavera 1990; Callapez and Soares 2000) and added new records, reporting 89 taxa, including 75 gastropods and 14 bivalves. Further work by Ávila (2005) and Ávila et al. (2007, 2009b, 2010, 2015b) increased the number of Pleistocene mollusc species to 138 taxa (114 gastropods and 24 bivalves). During the course of the last glaciation, 23 of these species locally disappeared from the Azores, of which 14 were thermophilic species that were not able to cope with the drop of the sea temperatures, and 7 species were associated with shallow sandy substrates that disappeared from the island when sea-level dropped below the shelf-edge (cf. Ávila et al. 2008b, 2015b for further information). Of the 138 taxa of molluscs, 22 are endemic gastropods, all of them still living along the coastlines of the Azores (Table 1).
6.3.2 Echinodermata
The most recent review on the fossil echinoid material collected from the fossiliferous deposits of Santa Maria (both the early Pliocene deposits as well as the Pleistocene deposits) was published by Madeira et al. (2011). These fossils are predominantly disarticulated skeletal material (primary spines and coronal fragments). Only subordinate complete tests are preserved. At least 7 taxa are reported by Madeira et al. (2011) from the Upper Miocene to lower Pliocene beds: Eucidaris tribuloides (Lamarck), Echinoneus cf. cyclostomus Leske, Clypeaster altus (Leske), Echinocyamus pusillus (Müller), Echinocardium sp. 1, Echinocardium sp. 2, and Schizobrissus sp (cf. Fig. 14a–m). Undetermined spatangoids were also found, but the poor preservation of the fossils did not allow their identification. Irregular echinoids such as spatangoids, clypeasteroids and Echinoneus usually live in soft-bottom environments, whereas the regular echinoid Eucidaris tribuloides preferentially lives epifaunally on hard substrates.

Neogene fossil echinoderms from Santa Maria (Madeira et al. 2011). a–d Clypeaster altus (Leske) in aboral (a), oral (b), and left lateral view (c). d Cross view of a specimen photographed at Pedra-que-pica. e–i Eucidaris tribuloides (Lamarck) in oral (e) and lateral view (f, g). h Fragment of a specimen photographed at Pedra-que-pica. i Spines. j–m Echinoneus cf. cyclostomus Leske in aboral (j), lateral (k) and oral view (l). m specimen photographed at Pedra-que-pica. n–p Cross section of spines from the Pleistocene of Lagoinhas. Arbacia lixula Linnaeus (n), Sphaerechinus granularis (Lamarck) (o), and Paracentrotus lividus (Lamarck) (p). q, r, v Arbacia lixula in aboral (q), oral (r), and lateral view (v). s–u Echinocyamus pusillus (Müller) in aboral (s), oral (t) and left lateral view (u) (Madeira et al. 2011)
The Upper Pleistocene outcrops (MIS 5e) yielded three regular echinoid species: Sphaerechinus granularis (Lamarck), Arbacia lixula Linnaeus and Paracentrotus lividus (Lamarck) (Fig. 14n–v). The presence of taxa typical of tropical seas in the Mio-Pliocene sediments contrasts with the Pleistocene and modern echinoid fauna, which is warm-temperate in composition (Madeira et al. 2011). The diversity and specific composition of echinoids preserved in the Pliocene deposits of Santa Maria differs markedly from that of the island’s Pleistocene fossil record. Worldwide, Pleistocene faunas tend to be closely related to the living biota of any area (Madeira et al. 2011), and the Azorean Pleistocene is no exception.
6.3.3 Brachiopoda
Although brachiopods are common members of shallow-water communities in Miocene and Pliocene sediments of the Mediterranean or Central Paratethys (Ruggiero 1994; Bitner and Dulai 2004), only two species are known from the Pliocene sediments of Santa Maria: Terebratulina retusa (Linnaeus), reported by Bronn (1860) and by Ferreira (1955) as T. caput-serpentis; and Novocrania turbinata (Poli), reported by Kroh et al. (2008; Fig. 15a–c). Today, none of these species occurs in the Azores. N. turbinata presently lives in warmer waters (the Cape Verde Islands, off the north-western African coast and the Mediterranean), and is a non-molluscan example of the local disappearance of thermophilic taxa from the Azores since the Pliocene (Kroh et al. 2008).

a–c The brachiopod Novocrania turbinata (Poli). d–f The crustacean Zullobalanus santamariaensis Buckeridge and Winkelmann. Lateral (d) and apical view (f) of a specimen from Malbusca outcrop. 1: carina; 2: carinolatus; 3: latus; 4: rostrum Winkelmann et al. (2010). e Zullobalanus santamariaensis photographed in situ, on a rhodolith, at Malbusca outcrop. g Chela of the crustacean Neptunus granulatus Edwards from the Pliocene of Lombo Gordo outcrop
6.3.4 Crustacea
Fossil barnacles were reported by Zbyszewski and Ferreira (1962b) from the Pliocene of Santa Maria (as Balanus laevis Bruguière) and by Callapez and Soares (2000) of the Pleistocene deposits of Lagoinhas (as Balanus sp.). However, these fossil cirripedes were not described in detail. Winkelmann et al. (2010) described a new species, Zullobalanus santamariaensis Buckeridge and Winkelmann (Fig. 15d–f), endemic from the early Pliocene outcrops of the island. This endemic barnacle lived in shallow-water, and probably, the Balanus laevis record was misidentified with Zullobalanus santamariaensis by Zbyszewski and Ferreira (1962b). The ancestral Z. santamariaensis might have reached the Azores passively attached to whale barnacles that were subsequently dislodged near Santa Maria Island, ultimately establishing a viable population (Winkelmann et al. 2010). Zbyszewski and Ferreira (1962b) also reported Neptunus granulatus Edwards, based on a chela collected at Lombo Gordo outcrop (Fig. 15g).
6.3.5 Ostracoda
The first study on the fossil ostracods from the Azores archipelago was recently published by Meireles et al. (2012). These authors reported 13 taxa from the Pliocene of the Malbusca section, representing seven families and 12 genera: Xestoleberis cf. paisi Nascimento, Loxoconcha stellifera Müeller, Loxoconcha rhomboidea (Fischer), Callistocythere oertlii Nascimento, Leptocythere azorica Meireles and Faranda, Pachycaudites cf. armilla Ciampo, Dameriacella cf. dameriacensis (Keij), Aurila sp., ?Quadracythere sp., Heliocythere magnei (Keij), Neonesidea rochae Nascimento, Paracypris sp., and Cyamocytheridea sp. All these species are typical of warm waters and epi-neritic habitats (∼10–50 m of depth) (Meireles et al. 2012). Xestoleberis cf. paisi, L. rhomboidea, L. stellifera, C. oertlii and N. rochae still occur nowadays in the shallow-water environments (down to 40 m depth) of the Azores. All other ostracod taxa recorded from Malbusca (including the new species L. azorica) disappeared from the Azores and the Atlantic domain probably at the end of the Zanclean age, around 3.6 Ma, as suggested by Ávila et al. (2016a) for marine molluscs. Meireles et al. (2012) considered these ostracods as warm-water taxa that did not survive the Plio-Pleistocene climate deterioration.
7 The Future: The PalaeoPark Santa Maria
In 1996, Richard Lane and David L. Bruton argued during the 30th International Geological Congress (IGC) in Beijing (China) that protection and conservation of fossil sites around the world should constitute a major objective of the International Palaeontological Association (IPA). This initiative was supported by four workshops that took place during the 32nd International Geological Congress (IGC—2004) in Florence (Italy), at the 2nd International Palaeontological Congress (IPC—2006) in Beijing (China), the 33rd IGC (2008) in Oslo (Norway) and the 3rd IPC (2012) in London (UK). From 2004 on, Jere Lipps promoted the concept of the “PalaeoParks” internationally, with the aim of fast-protecting and conserving fossil sites around the world. According to Lipps (2009), the preservation of fossil specimens in museums, as a rule being performed correctly, is insufficient. Instead, fossiliferous deposits themselves must be preserved because they provide data that cannot be preserved in museums, such as: the characteristics and structures of their hosting sedimentary rocks, the existence of trace fossils, data on adjacent sediment layers, and possibility of collecting new data by using techniques and methodologies not currently available or even developed. Thus, this PalaeoPark Initiative not only protects fossil-bearing sites but, perhaps more importantly, aims to “preserve the history of life on Earth to provide materials for scientific research now and in the future” (Lipps 2009: 1–2).
The Azores Geopark was created in 2010 as a non-profit association charged with the conservation and promotion of the natural heritage of the Azores with particular emphasis on the islands’ geological heritage. The challenging concept of the Azores Geopark—“9 Islands, 1 Geopark”—represents the main goal of the association. By definition, a Geopark “is a territory with well-defined boundaries, which has an important Geological Heritage, coupled with a sustainable development strategy. Thus, Geoparks include a significant number of sites of geological interest which, due to their rarity or peculiarities, have scientific, educational, cultural, economic (e.g. tourism), scenic or aesthetic (e.g. landscape) value (or importance), or can be considered as geosites. These sites may also contain other points of interest and value (e.g. ecological, historical and cultural), theme parks and other related infrastructures, which should be interconnected in a network by paths and trails” (Geoparque Açores 2013). In the Azores, 121 geosites were considered important and 57 were selected as representative of the geodiversity of the archipelago. These geosites are distributed throughout the islands, as well as on seamounts and seafloor that surround them.
On Santa Maria Island, 15 geosites were classified and 5 were selected as deserving priority status for the development of geoconservation strategies and implementation of socioeconomical development. Of these five geosites, only two (Pedreira do Campo and Ponta do Castelo) contain fossils. Moreover, no Pleistocene outcrops were selected at this stage, notwithstanding the recent inclusion of the Prainha outcrop into the first Azorean Regional Natural Monument (besides Pedreira do Campo and Figueiral; DR 47 2008). In 2012, new legislation, produced by the Azores Regional Government with the scientific support of the MPB group, has protected all fossiliferous sites on Santa Maria (DR 39 2012). Today, in the entire island, no fossils may be collected and only professionals may get licenses to sample for scientific purposes.
In order to further promote the exceptional palaeontological heritage of Santa Maria Island, the MPB group proposed to the IPA the classification of Santa Maria Island as a PalaeoPark, which was accepted in late 2012. The aim of PalaeoPark Santa Maria is to emphasize in a synergetic way the outstanding palaeontological record of such a small island within the broader “umbrella” of the Azores Geopark. Thus, in addition to the thematic routes proposed by the Azores Geopark (Volcanic Caves Route, Viewpoints Route and Hiking Trails Route), the creation of the Fossils Route was presented by the MPB group to the Azores Regional Government in 2010. This project was funded and started in July 2012. The main objective of the Fossils Route is the sustainable touristic use of the palaeontological heritage of Santa Maria Island and to showcase the relatively recent concept of biogeodiversity. For this purpose, five paths were outlined on Santa Maria, which were mapped and characterised in relation to biodiversity, geodiversity and various palaeontological aspects, such as: which fossil species are the most abundant, a detailed description of the palaeoecology of the deposits crossed by the paths and perhaps the most interesting point, the individual history of each of the deposits. One of these paths is distinguished by being entirely marine. This “pedestrian” marine path, unique in the Azores Autonomous Region, consists of a tour around the island by boat, disembarking (sea conditions permitting) at exposed deposits previously selected for this purpose. Along the way, tourists have access to scientific information provided by fully accredited guides, such as the description of the various geological units that comprise the island of Santa Maria, a description of some of the geological features visible on the coast, as well as the pinpointing of spectacular pillow lava and prismatically jointed lava sections. All this is complemented by beautiful scenery and, of course, marine fossils.
The scientific certification of the guides, who have received specialised biological, palaeontological and geological training adapted to the volcanic and sedimentary sequences of Santa Maria Island, will be mandatory for the success of this PalaeoPark. Today, with the approval of the “Parque de Ilha de Santa Maria”, more than ten guides work with school children, local inhabitants and tourists. The raising of public awareness to the value of the fossiliferous outcrops is being achieved by: (1) addressing the island’s inhabitants with a special emphasis on local schools, whose students could attend excursions to the most significant outcrops led by MPB researchers; (2) spreading the scientific knowledge and the importance of science with scientific lectures done during the annual workshops “Palaeontology in Atlantic Islands” in cafes (“Clube Naval de Santa Maria” and “Central-Pub”) and Science Centers (“Centro de Interpretação Ambiental Dalberto Pombo”); (3) writing popular science books related to the fossils of Santa Maria Island (Ávila 2009; Ávila and Monteiro 2009; Ávila et al. 2010; Ávila and Rodrigues 2013); (4) reports published in newspapers, national and regional magazines, as well as with television reports; (5) the recent “House of Fossils” Museum at Vila do Porto, a multimedia museum that also portraits the most important fossils discovered at Santa Maria; and (6) reaching tourists through the “Fossils Route” trails. These fossil-related trails have caught the interest of the maritime-touristic companies located on the island, as they provide an interesting alternative for tourists, when sea conditions do not allow diving or whale-watching activities.
On Santa Maria, fossils have always been part of the local people’s traditional life. However, with the exception of a few more educated people, this heritage was overlooked until recently. Thanks to the efforts of the scientific community, this paradigm has changed from 2000 onward. At the present time, the fossiliferous deposits have an intrinsic value that goes far beyond the scientific one, as tourist attractions, for environmental, recreational and even artistic education (see Ávila et al. 2016b). As a consequence, a genuine development strategy for tourism based on the local palaeontology and biogeodiversity is now underway on Santa Maria Island. Perhaps the most visible result of all these scientific research/publishing efforts is evident when, while landing at Santa Maria on the local aircraft company SATA, the air hostess can be heard saying: “Dear passengers: welcome to Santa Maria, where the fossil heritage is a natural story-teller”. The final surprise is reserved for the airport’s baggage claim hall, where our attention is drawn to photographs of fossils decorating the floor. Ten years ago, no one spoke about the fossils of Santa Maria… today, they are a distinguishing feature of the “Island of the Sun”… or should that be the “Island of the fossils”?
References
Abdel-Monem, A. A., Fernandez, L. A., & Boone, G. M. (1975). K-Ar ages from the eastern Azores group (Santa Maria, São Miguel and the Formigas Islands). Lithos, 8, 247–254.
Adey, W. H. (1986). Coralline algae as indicators of sea-level. In O. van de Plasche (Ed.), Sea-level research, a manual for the collection and evaluation of data (pp. 229–279). Amsterdam: Free University of Amsterdam.
Amen, R. G., Neto, A. I., & Azevedo, J. M. N. (2005). Coralline-algal framework in the Quaternary of Prainha (Santa Maria island, Azores). Revista Española de Micropaleontologia, 37, 63–70.
Ávila, S. P. (2005). Processos e Padrões de Dispersão e Colonização nos Rissoidae (Mollusca: Gastropoda) dos Açores (x + 329 pp.) (Unpublished Ph.D. Thesis). Universidade dos Açores, Ponta Delgada.
Ávila, S. P. (2009). Geologia e os fósseis. In Publiçor (Eds.), Açores. Santa Maria, a Ilha Amarela (pp. 16–23). Ponta Delgada: Publiçor Editores.
Ávila, S. P. (2013). Unravelling the patterns and processes of evolution of marine life in oceanic islands: a global framework. In J. M. Férnández-Palacios, L. Nascimento, J. C. Hernández, S. Clemente, A. González, & J. P. Díaz-González (Eds.), Climate change: past, present and future perspectives. A global synthesis from the Atlantic (pp. 85–115). La Laguna: Universidad de La Laguna, Tenerife.
Ávila, S. P., Amen, R. G., Azevedo, J. M. N., Cachão, M., & García-Talavera, F. (2002). Checklist of the Pleistocene marine molluscs of Prainha and Lagoinhas (Santa Maria Island, Azores). Açoreana, 9(4), 343–370.
Ávila, S. P., Ávila, S. P., Silva, L., Ramalho, R., Quartau, R., Hipólito, A., et al. (2015a). A review of the MIS 5e highstand deposits from Santa Maria Island (Azores, NE Atlantic): Palaeobiodiversity, palaeoecology and palaeobiogeography. Quaternary Science Reviews, 114, 126–148.
Ávila, S. P., Cachão, M., Ramalho, R. S., Botelho, A. Z., Madeira, P., Rebelo, A. C., et al. (2016a). The palaeontological heritage of Santa Maria Island (Azores: NE Atlantic): a re-evaluation of geosites in GeoPark Azores and their use in geotourism. Geoheritage, 8, 155–171.
Ávila, S. P., Cordeiro, R., Rodrigues, A. R., Rebelo, A. C., Melo, C., Madeira, P., et al. (2015c). Fossil Mysticeti from the Pleistocene of Santa Maria Island, Azores (NE Atlantic Ocean), and the prevalence of fossil cetaceans on oceanic islands. Palaeontologia Electronica, 18.2.27A, 1–12. palaeo-electronica.org/content/2015/1225-oceanic-island-fossil-cetacean.
Ávila, S. P., Madeira, P., García-Talavera, F., Silva, C. M., Cachão, M., & Martins, A. M. F. (2007). Luria lurida (mollusca: gastropoda), a new record for the Pleistocene of Santa Maria (Azores, Portugal). Arquipélago, 24, 53–56.
Ávila, S. P., Madeira, P., Mendes, N., Rebelo, A. C., Medeiros, A., Gomes, C., et al. (2008a). Mass extinctions in the Azores during the last glaciation: Fact or myth? Journal of Biogeography, 35, 1123–1129.
Ávila, S. P., Madeira, P., Silva, C. M., Cachão, M., Landau, B., Quartau, R., et al. (2008b). Local disappearance of bivalves in the Azores during the last glaciation. Journal of Quaternary Science, 23, 777–785.
Ávila, S. P., Madeira, P., Zazo, C., Kroh, A., Kirby, M., Silva, C. M., et al. (2009a). Palaeocology of the Pleistocene (MIS 5.5) outcrops of Santa Maria Island (Azores) in a complex oceanic tectonic setting. Palaeogeography, Palaeoclimatology, Palaeoecology, 274, 18–31.
Ávila, S. P., & Martins, A. M. F. (Eds.) (2007). Palaeontology in Atlantic Islands. In Proceedings of the First Atlantic Islands Neogene, International Congress, Açoreana (Suppl. 5, pp. 1–172).
Ávila, S. P., Medeiros, A., Martins, A. M. F., Silva, A., Melo, C., Gomes, C., et al. (2011). Lajes do Pico “À ban-baxe-muro” (pp. 1–345). Ponta Delgada: Editora Publiçor.
Ávila, S. P., Melo, C., Berning, B., Cordeiro, R., Landau, B., & da Silva, C. M. (2016b). Persististrombus coronatus (Mollusca: Strombidae) in the early Pliocene of Santa Maria Island (Azores: NE Atlantic): Palaeoecology, palaeoclimatology and palaeobiogeographic implications on the NE Atlantic Molluscan Biogeographical Provinces. Palaeogeography, Palaeoclimatology, Palaeoecology, 441, 912–923.
Ávila, S. P., & Monteiro, P. (2009). Açores, o Império dos Fósseis (pp. 1–272). Porto: Edições Caixotim, Lda.
Ávila, S. P., Ramalho, R., Habermann, J., Quartau, R., Kroh, A., Berning, B., et al. (2015b). Palaeoecology, taphonomy, and preservation of a lower Pliocene shell bed (coquina) froma volcanic oceanic island (Santa Maria Island, Azores). Palaeogeography, Palaeoclimatology, Palaeoecology, 430, 57–73.
Ávila, S. P., Ramalho, R., & Vullo, R. (2012). Systematics, palaeoecology and palaeobiogeography of the Neogene fossil sharks from the Azores (Northeast Atlantic). Annales de Paléontologie, 98, 167–189.
Ávila, S. P., Rebelo, A. C., Medeiros, A., Melo, C., Gomes, C., Bagaço, L., et al. (2010). Os fósseis de Santa Maria (Açores). 1. A jazida da Prainha (pp. 1–103). Lagoa: OVGA – Observatório Vulcanológico e Geotérmico dos Açores.
Ávila, S. P., & Rodrigues, J. A. (2013). PaleoPark Santa Maria (pp. 1–120). Ponta Delgada: Letras Lavadas edições.
Ávila, S. P., Silva, C. M., Schiebel, R., Cecca, F., Backeljau, T., & Martins, A. M. F. (2009b). How did they get here? Palaeobiogeography of the Pleistocene marine molluscs of the Azores. Bulletin of the Geological Society of France, 180, 295–307.
Barreiros, J. P., & Gadig, O. B. F. (2011). Catálogo ilustrado dos tubarões e raias dos Açores (pp. 1–195). Angra do Heroísmo: Instituto Açoriano de Cultura.
Berthois, L. (1950). Sur la présence d’une microfaune dans le calcaire de Santa Maria (Açores). Açoreana, 4(4), 277–287.
Berthois, L. (1951). Sur la présence de basses terrasses marines dans l’Archipel des Açores. Comptes Rendus du 76éme Congrès des Sociétés Savantes à Rennes, 101–106.
Berthois, L. (1953a). Sur la repartition granulométrique des éléments calcaires dans les sables des Açores (Portugal). Açoreana, 5(1), 54–63.
Berthois, L. (1953b). Terrasses marines d’altitude +5 à +8 m dans l’Archipel des Açores. Açoreana, 5(1), 64–70.
Berthois, L. (1953c). Contribution à l’étude lithologique de l’Archipel des Açores. Comunicações dos Serviços Geológicos de Portugal, 34, 1–198.
Bitner, M. A., & Dulai, A. (2004). Revision of Miocene brachiopods of the Hungarian Natural History Museum, with special regard to the Meznerics collection. Fragmenta Palaeontologica Hungarica, 22, 69–82.
Boid, E. (1835). A description of the Azores or Western Islands (pp. 1–373). London: Edward Churton.
Bosence, D. W. J. (1983). Coralline algal reef frameworks. Journal of the Geological Society of London, 140, 365–376.
Bosence, D. W. J. (1984). Construction and preservation of two Recent coralline algal reefs, St. Croix, Caribbean. Palaeontology, 27, 549–574.
Braga, J. C., & Aguirre, J. (2001). Coralline algal assemblages in upper Neogene reef and temperate carbonates in southern Spain. Palaeogeography, Palaeoclimatology, Palaeoecology, 175, 27–41.
Bressan, G., & Babbini, L. (2003). Biodiversità marina delle coste Italiane: Corallinales del Mar Mediterraneo: guida all deteminazione. Biologia Marina Mediterrânea, 10(Suppl. 2), 1–237.
Bronn, H. G. (1860). Die fossilen Reste von Santa Maria, der südlichsten der Azorischen Inseln. In G. Hartung (Ed.), Die Azoren in ihrer äusseren Erscheinung und nach ihrer geognostischen Natur (pp. 1–350). Leipzig: Verlag von Wilhelm Engelmann.
Brotchie, J. F., & Silvester, R. (1969). On Crustal Flexure. Journal of Geophysical Research (Solid Earth), 74(22), 5240–5252.
Cachão, M., Madeira, J., Silva, C. M., Azevedo, J. M. N., Cruz, A. P., Garcia, C., et al. (2003). Pedreira do Campo (Santa Maria, Açores): monumento natural. Actas do VI Congresso Nacional de Geologia. Ciências da Terra (UNL), 5, 120–123.
Calado, H., Ávila, S. P., & Madeira, P. (2007). The Coastal Zone Management Plan of Santa Maria as a chance for fossiliferous outcrops management. In S. P. Ávila & A. M. F. Martins (Eds.), Proceedings of the “1st Atlantic Islands Neogene”, International Congress. Ponta Delgada, 12–14 June 2006. Açoreana, Suplemento 5, 162–172.
Callapez, P., & Soares, A. F. (2000). Late Quaternary marine mollusks from Santa Maria (Azores); paleoecologic and paleobiogeographic considerations. Ciências da Terra (UNL), 14, 313–322.
Cotter, J. C. B. (1888–1892). Noticia de alguns fosseis terciarios da Ilha de Santa Maria no archipelago dos Açores. Communicações da Commissão dos Trabalhos Geológicos de Portugal, 2, 255–287.
Crough, S. (1978). Thermal origin of mid-plate hot-spot swells. Geophysical Journal International, 55(2), 451–469.
Bedemar, V. (1837). Resumo de observações geologicas feitas em uma viagem ás ilhas da Madeira Porto Santo e Açores nos annos de 1835 e 1836 (pp. 1–25). Lisboa: Impressão de Galhardo Irmãos.
Dietz, R., & Menard, H. (1953). Hawaiian swell, deep, and arch, and subsidence of the Hawaiian Islands. The Journal of Geology, 61(2), 99–113.
Donovan, S. K., & Gunter, G. C. (2001). Fossil sharks from Jamaica. Bulletin of the Mizunami Fossil Museum, 28, 211–215.
DR 47. (2008). Decreto Legislativo Regional 47/2008/A de 7 de Novembro. Parque Natural da Ilha de Santa Maria. Diário da República, 1.ª série, nº 217 – 7 de Novembro de 2008, 7787–7801.
DR 39. (2012). Decreto Legislativo Regional n.º 39/2012/A. Primeira alteração ao Decreto Legislativo Regional n.º 47/2008/A, de 7 de novembro (Parque Natural da Ilha de Santa Maria). Diário da República, 1.ª série, nº 182 – 19 de setembro de 2012, 5282–5310.
Duggen, S., Olgun, N., Croot, P., Hoffman, L., Dietze, H., Delmelle, P., et al. (2010). The role of airborne volcanic ash for the surface ocean biogeochemical iron-cycle: A review. Biogeosciences, 7, 827–844.
Dumas, S., & Arnott, R. W. C. (2006). Origin of hummocky and swaley cross-stratification—The controlling influence of unidirectional current strength and aggradation rate. Geology, 34, 1073–1076.
Estevens, M., & Ávila, S. P. (2007). Fossil whales from the Azores. In S. P. Ávila & A. M. F. Martins (Eds.), Proceedings of the “1st Atlantic Islands Neogene”, International Congress, Ponta Delgada, 12–14 June 2006. Açoreana, Suplemento, 5, 140–161.
Feraud, G., Kaneoka, I., & Allegre, C. J. (1980). K-Ar ages and stress pattern in the Azores: Geodynamic implications. Earth and Planetary Science Letters, 46, 275–286.
Ferreira, O. V. (1952). Os pectinídeos do Miocénico da ilha de Santa Maria (Açores). Revista da Faculdade de Ciências de Lisboa, 2ª Série, C, 2(2), 243–258.
Ferreira, O. V. (1955). A fauna Miocénica da ilha de Santa Maria. Comunicações dos Serviços Geológicos de Portugal, 36, 9–44.
Ferreira, O. V. (1961). Afloramentos de calcário miocénico da Ilha de Santa Maria (Açores). Comunicações dos Serviços Geológicos de Portugal, 45, 467–478.
Fisher, R. V., & Schmincke, H.-U. (1984). Pyroclastic rocks (pp. 1–427). Berlin: Springer.
Fortes, M. D. (1991). Seagrass-mangrove ecosystems managmement: A key to marine coastal conservation in the ASEAN region. Marine Pollution Bulletin, 22, 113–116.
Frogner, P., Gislason, S. R., & O’Skarsson, N. (2001). Fertilizing potential of volcanic ash in ocean surface water. Geology, 29, 486–490.
Frutuoso, G. (1983). Livro Terceiro das Saudades da Terra (pp. 1–300). Ponta Delgada: Instituto Cultural de Ponta Delgada.
García-Talavera, F. (1990). Fauna tropical en el Neotirreniense de Santa Maria (I. Azores). Lavori della Società Italiana di Malacologia, 23, 439–443.
Geoparque Açores. (2013). http://www.azoresgeopark.com/. Accessed 26 February 2013.
Habermann, J. (2010). Thesis Mapping—Santa Maria, Azores. Area: Vila do Porto – Almagreira – Praia (Unpublished Diplom mapping, 84 pp). GeoZentrum Nordbayern, University of Erlangen-Nürnberg.
Habermann, J. (2011). Gigantopecten latissimus (Brocchi, 1814). Microstructure and stable isotope sclerochronology of an Upper Miocene Gigantopecten shell (Bivalvia: Pectinidae) from Santa Maria Island (Azores, Portugal) (Unpublished Diplom thesis, xi + 107 pp). GeoZentrum Nordbayern, University of Erlangen-Nürnberg.
Hartung, G. (1860). Die Azoren in ihrer äusseren Erscheinung und nach ihrer geognostischen Natur (pp. 1–350). Leipzig: Verlag von Wilhelm Engelmann.
Hartung, G. (1864). Geologische Beschreibung der Inseln Madeira und Porto Santo (pp. 1–299). Leipzig: Verlag von Wilhelm Engelmann.
Heikoop, J. M., Tsujita, C. J., Heikoop, C. E., Risk, M., & Dickin, A. P. (1996a). Effects of volcanic ashfall recorded in ancient marine benthic communities: Comparison of a nearshore and an offshore environment. Lethaia, 29, 125–139.
Heikoop, J. M., Tsujita, C. J., Risk, M. J., & Tomascik, T. (1996b). Corals as proxy recorders of volcanic activity: Evidence from Banda Api, Indonesia. Palaios, 11, 286–292.
Hofrichter, R. (2001). Das Mittelmeer: Fauna, Flora, Ökologie – Allgemeiner Teil, 1 (p. 607). Heidelberg: Spektrum.
Huppert, K. L., Royden, L. H., & Perron, J. T. (2015). Dominant influence of volcanic loading on vertical motions of the Hawaiian Islands. Earth and Planetary Science Letters, 418, 149–171.
Iturralde-Vinent, M., Hubbell, G., & Rojas, R. (1996). Catalogue of Cuban fossil Elasmobranchii (Paleocene to Pliocene) and paleogeographic implications of their Lower to Middle Miocene occurrence. Journal of the Geological Society of Jamaica, 31, 7–21.
Janssen, A. W., Kroh, A., & Ávila, S. P. (2008). Early Pliocene heteropods and pteropods (Mollusca, Gastropoda) from Santa Maria Island (Azores, Portugal): Systematics and biostratigraphic implications. Acta Geologica Polonica, 58, 355–369.
Johnson, M. E., Baarli, B. G., Santos, A., & Mayoral, E. (2012). Ichnofacies and microbial build-ups on Late Miocene rocky shores from Menorca (Balearic Islands), Spain. Facies, 57, 255–265.
Johnson, M. E., Uchman, A., Costa, P. J. M., Ramalho, R. S., & Ávila, S. P. (2017). Intense hurricane transport sand onshore: example from the Pliocene Malbusca section on Santa Maria Island (Azores, Portugal). Marine Geology, 385, 244–249.
Kirby, M. X., Jones D. S., & Ávila, S. P. (2007). Neogene shallow-marine paleoenvironments and preliminary Strontium isotope chronostratigraphy of Santa Maria Island, Azores. In S. P. Ávila & A. M. F. Martins (Eds.), Proceedings of the “1st Atlantic Islands Neogene”, International Congress, Ponta Delgada, 12–14 June 2006. Açoreana, Suplemento 5, 112–125.
Klügel, A., Hansteen, T., & Galipp, K. (2005). Magma storage and underplating beneath Cumbre Vieja volcano, La Palma (Canary Islands). Earth and Planetary Science Letters, 236(1–2), 211–226.
Klügel, A., Longpré, M.-A., García-Cañada, L., & Stix, J. (2015). Deep intrusions, lateral magma transport and related uplift at ocean island volcanoes. Earth and Planetary Science Letters, 431, 140–149.
Krejci-Graf, K., Frechen, J., Wetzel, W., & Colom, G. (1958). Gesteine und Fossilien von den Azoren. Senckenbergiana Lethaea, 39(5/6), 303–351.
Kroh, A., Bitner, M. A., & Ávila, S. P. (2008). Novocrania turbinata (Brachiopoda) from the Early Pliocene of the Azores (Portugal). Acta Geologica Polonica, 58, 473–478.
Lipps, J. H. (2009). PaleoParks: Our paleontological heritage protected and conserved in the field worldwide. In: J. H. Lipps, & B. R. C. Granier (Eds.), PaleoParks—The protection and conservation of fossil sites worldwide. Brest: Carnets de Géologie/Notebooks on Geology. http://paleopolis.rediris.es/cg/CG2009_BOOK_03/CG2009_BOOK_03_Chapter01.pdf.
Madeira, P., & Ávila, S. P. (2006). Jazidas Fósseis. In H. Calado, J. Porteiro, M. Pereira, & A. Z. Botelho (Coordenação Científica). LITOSOST: Gestão Sustentável do Desenvolvimento Social, Económico e ecológico das Áreas Litorais da Macaronésia (pp. 74–76). Plano de ordenamento da Orla Costeira – Santa Maria. Fase I – Caracterização e Diagnóstico
Madeira, P., Kroh, A., Cordeiro, R., Meireles, R., & Ávila, S. P. (2011). The fossil echinoids of Santa Maria Island, Azores (Northern Atlantic Ocean). Acta Geologica Polonica, 61, 243–264.
Madeira, P., Kroh, A., Martins, A. M. F., & Ávila, S. P. (2007). The marine fossils from Santa Maria Island (Azores, Portugal): an historical overview. In S. P. Ávila & A. M. F. Martins (Eds.), Proceedings of the “1st Atlantic Islands Neogene”, International Congress, Ponta Delgada, 12–14 June 2006. Açoreana, Suplemento 5, 59–73.
Madeira, J., Mata, J., Mourão, C., Silveira, A. B., Martins, S., Ramalho, R., et al. (2010). Volcano-stratigraphic and structural evolution of Brava Island (Cape Verde) from 40Ar/39Ar, U/Th and field constraints. Journal of Volcanology and Geothermal Research, 196, 219–235.
Mayer, K. (1864). Die Tertiär-Fauna der Azoren und Madeiren. Systematisches Verzeichniss der fossilen Reste von Madeira, Porto Santo und Santa Maria nebst Beschreibung der neuen Arten, VI + 107 pp. Zürich.
McNutt, M., & Menard, H. W. (1978). Lithospheric exure and uplifted Atolls. Journal of Geophysical Research (Solid Earth), 83(B3), 1206–1212.
Meireles, R. P., Costanza, F., Gliozzi, E., Pimentel, A., Zanon, V., & Ávila, S. P. (2012). Late Miocene marine ostracods from Santa Maria Island, Azores (NE Atlantic): palaeoecology and palaeobiogeography. Révue de Micropaléontologie, 55, 133–148.
Meireles, R. P., Quartau, R., Ramalho, R., Rebelo, A. C., Madeira, J., Zanon, V., et al. (2013). Depositional processes on oceanic island shelves—Evidence from storm-generated Neogene deposits from the Mid North Atlantic. Sedimentology (accepted article). https://doi.org/10.1111/sed.12055.
Menéndez, I., Silva, P., Martin-Betancor, M., Perez-Torrado, F., Guillou, H., & Scaillet, S. (2008). Fluvial dissection, isostatic uplift, and geomorphological evolution of volcanic islands (Gran Canaria, Canary Islands, Spain). Geomorphology, 102(1), 189–203.
Miller, K. G., Kominz, M. A., Browning, J. V., Wright, J. D., Mountain, G. S., Katz, M. E., et al. (2005). The phanerozoic record of global sea-level change. Science, 310, 1293–1298.
Mitchell-Thomé, R. C. (1976). Geology of the middle Atlantic Islands (351 pp.). Berlin: Gebrüder Borntraeger.
Morelet, A. (1860). Notice sur l’Histoire Naturelle des Açores suivie d’une description des Mollusques terrestres de cet Archipel (pp. 1–216). Paris: J.-B. Baillière et Fils.
Morgan, J., Morgan, W., & Price, E. (1995). Hotspot melting generates both hotspot volcanism and a hotspot swell. Journal of Geophysical Research (Solid Earth), 100(B5), 8045–8062.
Neto, A. I. (1994). Checklist of the benthic marine macroalgae of the Azores. Arquipélago, 12A, 15–34.
Nieves-Rivera, A. M., Ruizyantín, M., & Gottfried, M. D. (2003). New record of the lamnid shark Carcharodon megalodon from the Middle Miocene of Puerto Rico. Caribbean Journal of Science, 39, 223–227.
Nunes, J. C., Lima E. A. & Medeiros, S. (2007). Os Açores, ilhas de geodiversidade: o contributo da ilha de Santa Maria. In S. P. Ávila & A. M. F. Martins (Eds.), Proceedings of the “1st Atlantic Islands Neogene”, International Congress, Ponta Delgada, 12–14 June 2006. Açoreana, Suplemento 5, 74–111.
Pérès, J., & Picard, J. M. (1964). Nouveau manuel de bionomie benthique de la mer Méditerranée. Recueil Travaux Station Marine Endoume, 31(47), 1–131.
Portell, R. W., Hubbel, G., Donovan, S. K., Green, J. L., Harper, D. A. T., & Pickerill, R. (2008). Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles. Caribbean Journal of Science, 44, 279–286.
Quartau, R., Hipólito, A., Romagnoli, C., Casalbore, D., Madeira, J., Tempera, F., et al. (2014). The morphology of insular shelves as a key for understanding the geological evolution of volcanic islands: Insights from Terceira Island (Azores). Geochemistry, Geophysics, Geosystems, 15, 1801–1826.
Quartau, R., Madeira, J., Mitchell, N. C., Tempera, F., Silva, P. F., & Brandão, F. (2015). The insular shelves of the Faial-Pico Ridge (Azores archipelago): A morphological record of its evolution. Geochemistry, Geophysics, Geosystems, 16, 1401–1420.
Quartau, R., Tempera, F., Mitchell, N. C., Pinheiro, L. M., Duarte, H., Brito, P. O., et al. (2012). Morphology of the Faial Island shelf (Azores): The interplay between volcanic, erosional, depositional, tectonic and mass-wasting processes. Geochemistry, Geophysics, Geosystems, 13, Q04012. https://doi.org/10.1029/2011GC003987.
Ralph, P. J., Tomasko, D., Moore, K., Seddon, S., & Macinnis-Ng, C. M. O. (2006). Human impacts on seagrasses: eutrophication, sedimentation and contamination. In A. W. D. Larkum, R. J. Orth, & C. M. Duarte (Eds.), Seagrasses: Biology, ecology and conservation (pp. 567–593). Dordrecht: Springer.
Ramalho, R. (2011). Building the Cape Verde Islands. Berlin/Heidelberg: Springer.
Ramalho, R. S., Brum da Silveira, A., Fonseca, P., Madeira, J., Cosca, M., Cachão, M., et al. (2015). The emergence of volcanic oceanic islands on a slow-moving plate: the example of Madeira Island, NE Atlantic. Geochemistry, Geophysics, Geosystems, 16, 522–537.
Ramalho, R. S., Helffrich, G., Cosca, M., Vance, D., Hoffmann, D., & Schmidt, D. N. (2010a). Vertical movements of ocean island volcanoes: Insights from a stationary plate environment. Marine Geology, 275, 84–95.
Ramalho, R. S., Helffrich, G., Cosca, M., Vance, D., Hoffmann, D., & Schmidt, D. N. (2010b). Episodic swell growth inferred from variable uplift of the Cape Verde hot spot islands. Nature Geoscience, 3, 774–777.
Ramalho, R. S., Helffrich, G., Madeira, J., Cosca, M., Thomas, C., Quartau, R., et al. (2017). Emergence and evolution of Santa Maria Island (Azores)—The conundrum of uplifted islands revisited. Geological Society of America Bulletin, 129(3–4), 372–390.
Ramalho, R. S., Helffrich, G., Schmidt, D., & Vance, D. (2010c). Tracers of uplift and subsidence in the Cape Verde Archipelago. Journal of the Geological Society of London, 167, 519–538.
Ramalho, R., Quartau, R., Trenhaile, A.S., Mitchell, N.C., WOODROFFE, C.D., & Ávila, S.P. (2013). Coastal evolution in oceanic islands: A complex interplay between volcanism, erosion, sedimentation and biogenic production. Earth Science Reviews, 127, 140–170. http://doi.org/10.1016/j.earscirev.2013.10.007.
Rebelo, A. C., Meireles, R. P., Barbin, V., Neto, A. I., Melo, C., & Ávila, S. P. (2016a). Diagenetic history of lower Pliocene rhodoliths of the Azores archipelago (NE Atlantic): application of cathodoluminescence techniques. Micron, 80, 112–121.
Rebelo, A. C., Rasser, M. W., Kroh, A., Johnson, M. E., Ramalho, R. S., Melo, C., et al. (2016b). Rocking around a volcanic island shelf: Pliocene rhodolith beds from Malbusca, Santa Maria Island (Azores, NE Atlantic). Facies, 62(3), 1–31.
Rebelo, A. C., Rasser, M. W., Riosmena-Rodríguez, R., Neto, A. I., & Ávila, S. P. (2014). Rhodolith forming coralline algae in the Upper Miocene of Santa Maria Island (Azores, NE Atlantic): a critical evaluation. Phytotaxa, 190(1), 370–382.
Reiss, W. (1862). Mittheilungen über die tertiären Schichten von Santa Maria, der südlichsten der Azoren, und ihre organischen Einschlüsse (pp. 1–22). Geognosie, Geologie und Petrefactenkunde: Neues Jarhrbuch für Mineralogie.
Reuter, M., & Piller, W. E. (2011). Volcaniclastic events in coral reef and seagrass environments: Evidence for disturbance and recovery (Middle Miocene, Styrian Basin, Austria). Coral Reefs, 30, 889–899.
Ruggiero, E. T. (1994). Neogene Salento brachiopod palaeocommunities. Bollettino della Società Paleontologica Italiana, 33, 197–213.
Santos, A. G., Mayoral, E., Dumont, C. P., da Silva, C. M., Ávila, S. P., Baarli, B. G., et al. (2015). Role of environmental change in rock-boring echinoid trace fossils. Palaeogeography, Palaeoclimatology, Palaeoecology, 432, 1–14.
Santos, A. G., Mayoral, E., Johnson, M. E., Baarli, B. G., Silva, C. M., Cachão, M., et al. (2012). Basalt mounds and adjacent depressions attract contrasting biofacies on a volcanically active Middle Miocene shoreline (Porto Santo, Madeira Archipelago, Portugal). Facies, 58, 573–585.
Santos, R. S., Porteiro, F. M., & Barreiros, J. P. (1997). Marine fishes of the Azores: An annotated checklist and bibliography. Arquipélago, 1, 1–242.
Serralheiro, A. (1976). A Geologia da Ilha de Santiago (Cabo Verde) (Unpublished Ph.D. thesis). Faculdade de Ciências da Universidade de Lisboa.
Serralheiro, A. (2003). A geologia da ilha de Santa Maria, Açores. Açoreana, 10, 141–192.
Serralheiro, A., Alves, C. A. M., Forjaz, V. H., & Rodrigues, E. B. (1987). Carta Vulcanológica dos Açores - Ilha de Santa Maria. Centro de Vulcanologia do I.N.I.C.
Serralheiro, A., & Madeira, J. (1990). Stratigraphy and geochronology of Santa Maria Island (Azores). Açoreana, 7, 575–592.
Serralheiro, A., Madeira, J., Alves, C. A. M., & Fernandez-Rubio, R. (1990). Stratigraphy and geochronology of Santa Maria Island (pp. 357–376). Azores: Livro de Homenagem ao Prof. Carlos Romariz. Departamento de Geologia da Faculdade de Ciências da Universidade de Lisboa.
Short, F. T., & Wyllie-Echeverria, S. (1996). Natural and human-induced disturbance of seagrasses. Environmental Conservation, 23, 17–27.
Sibrant, A.L.R., Hildenbrand, A., Marques, F.O., & Costa, A.C.G. (2015). Volcano-tectonic evolution of the Santa Maria Island (Azores): Implications for paleostressevolution at the western Eurasia-Nubia plate boundary: Journal of Volcanology and Geothermal Research, 291, 49–62.
Sleep, N. (1990). Hotspots and mantle plumes: Some phenomenology. Journal of Geophysical Research (Solid Earth), 95(B5), 6715–6736.
Stein, C., & Stein, S. (1992). A model for the global variation in oceanic depth and heat flow with lithospheric age. Nature, 359(6391), 123–129.
Stöckhert, F. (2011a) Kartierung des Gebietes zwischen Praia und Malbusca auf Santa Maria, Azoren (Unpublished master mapping thesis). GeoZentrum Nordbayern, University of Erlangen-Nürnberg.
Stöckhert, F. (2011b) Geochemical evolution of an intraplate volcanic island using the example of Santa Maria – Azores (Unpublished master thesis). GeoZentrum Nordbayern, University of Erlangen-Nürnberg.
Teixeira, C. (1950). Notas sobre a geologia das Ilhas Atlântidas. Anais da Faculdade de Ciências do Porto, 23(3–4), 193–233.
Thorton, S. E., Pilkey, O. H., & Lynts, G. W. (1978). A lagoonal crustose coralline algal microridge: Bahiret el Bibane, Tunisia. Journal of Sedimentary Petrology, 48(3), 743–750.
Titschack, J., Bromley, R. G., & Freiwald, A. (2005). Plio-Pleistocene cliff-bound, wedge-shaped, warm-temperate carbonate deposits from Rhodes (Greece): sedimentology and facies. Sedimentary Geology, 180, 29–56.
Titschack, J., & Freiwald, A. (2005). Growth, deposition, and facies of Pleistocene bathyal coral communities from Rhodes, Greece. In A. Freiwald & J. M. Roberts (Eds.), Cold-Water Corals and Ecosystems (pp. 41–59). Heidelberg: Springer.
Titschack, J., Joseph, N., Fietzke, J., Freiwald, A., & Bromley, R. G. (2013). Record of a tectonically-controlled regression captured by changes in carbonate skeletal associations on a structured island shelf (mid-Pleistocene, Rhodes, Greece). Sedimentary Geology, 283, 15–33.
Uchman, A., Johnson, M., Rebelo, A. C., Melo, C., Cordeiro, R., Ramalho, R. S., et al. (2016). Vertically-oriented trace fossil Macaronichnus segregatis from Neogene of Santa Maria Island (Azores; NE Atlantic) records vertical fluctuations of the coastal groundwater mixing zone on a small oceanic island. Geobios, 49, 229–241.
Walcott, R. (1970). Flexure of the lithosphere at Hawaii. Tectonophysics, 9(5), 435–446.
Watts, A., & ten Brink, U. (1989). Crustal structure, exure, and subsidence history of the Hawaiian Islands. Journal of Geophysical Research (Solid Earth), 94(B8), 10473–10500.
Wilson, M. E. J., & Lokier, S. W. (2002). Siliciclastic and volcaniclastic influences on equatorial carbonates: insights from the Neogene of Indonesia. Sedimentology, 49, 583–601.
Winkelmann, K., Buckeridge, J. S., Costa, A. C., Dionísio, M. A. M., Medeiros, A., Cachão, M., et al. (2010). Zullobalanus santamariaensis sp. nov. a new late Miocene barnacle species of the family Archeobalanidae (Cirripedia: Thoracica), from the Azores. Zootaxa, 2680, 33–44.
Winterer, E. L., & Sarti, M. (1994). Neptunian dykes and associated features in southern Spain: Mechanics of formation and tectonic implications. Sedimentology, 41, 1109–1132.
Zbyszewski, G., & d’Almeida, F. M. (1950). Os peixes miocénicos portugueses. Comunicações dos Serviços Geológicos de Portugal, 31, 309–423.
Zbyszewski, G., & Ferreira, O. D. V. (1961). La faune marine des basses plages quaternaires de Praia et Prainha dans l’ile de Santa Maria (Açores). Comunicações dos Serviços Geológicos de Portugal, 45, 467–478.
Zbyszewski, G., & Ferreira, O. D. V. (1962a). Étude géologique de l’île de Santa Maria (Açores). Comunicações dos Serviços Geológicos de Portugal, 46, 209–245.
Zbyszewski, G., & Ferreira, O. D. V. (1962b). La faune Miocène de l’île de Santa Maria (Açores). Comunicações dos Serviços Geológicos de Portugal, 46, 247–289.
Zbyszewski, G., Ferreira, O. D. V., & de Assunção, C. T. (1961). Carta Geológica de Portugal na escala 1/50 000. Notícia explicativa da folha de Ilha de Santa Maria (Açores). Lisboa: Serviços Geológicos de Portugal.
Acknowledgements
We thank the Direcção Regional da Ciência, Tecnologia e Comunicações (Regional Government of the Azores), Direcção Regional do Ambiente e do Mar (Regional Government of the Azores), FCT (Fundação para a Ciência e a Tecnologia) of the Portuguese government for financial support, and we also thank the Clube Naval de Santa Maria and Câmara Municipal de Vila do Porto for field assistance. We are grateful to the organizers and participants of several editions of the International Workshop “Palaeontology in Atlantic Islands” who assisted with fieldwork (2005–2012). Sérgio P. Ávila acknowledges his IF/00465/2015 research contract funded by Fundação para a Ciência e a Tecnologia (Portugal). This research received support from the SYNTHESYS Application DE-TAF-1071 “Systematics and taxonomy of the Miocene malacofauna of Santa Maria Island (Azores)” (http://www.synthesys.info/) which is financed by European Community Research Infrastructure Action under the FP7 “Capacities” Program. Ricardo Ramalho’s funding was provided by DFG-The German Research Foundation, through the project TH1530/6-1 “Investigation of ocean island uplift in the Azores Island Region”, and later though his FP7-PEOPLE-2011-IOF Marie Curie Fellowship. The contribution of Jörg Habermann is based on his master thesis and Prof. Dr. André Freiwald and Prof. Dr. Karsten Haase are gratefully acknowledged for their supervision. Furthermore, Jörg Habermann and Jürgen Titschack benefitted from the financial support by the DFG project BE4459/1-1 “Magmatic and sedimentological evolution of Oceanic Plateaus: constraints from the Azores”. We are grateful to the referee work of Jere Lipps and Markes Johnson, whose comments helped to clarify relevant parts of the manuscript. This work was also supported by FEDER funds through the Operational Programme for Competitiveness Factors—COMPETE and by National Funds through FCT—Foundation for Science and Technology under the UID/BIA/50027/2013 and POCI-01-0145-FEDER-006821.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer-Verlag GmbH Germany
About this chapter

Cite this chapter
Ávila, S.P., Ramalho, R.S., Habermann, J.M., Titschack, J. (2018). The Marine Fossil Record at Santa Maria Island (Azores). In: Kueppers, U., Beier, C. (eds) Volcanoes of the Azores. Active Volcanoes of the World. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-32226-6_9
Download citation
DOI: https://doi.org/10.1007/978-3-642-32226-6_9
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-32225-9
Online ISBN: 978-3-642-32226-6
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)