
Abstract
Parenting and children’s development function in a family system that involves many biological factors that interact with the environment and the parent–child relationship. There has been rapid growth in parenting research that incorporates indirect and direct measures of a variety of biological factors, in the hope of better articulating the mechanisms of child and family development, as well as improving the efficacy of interventions to improve developmental and parenting outcomes. In the current chapter, we review this literature, with particular emphasis on genetics, neural structure and function, and hormones. We provide an overview of the major methodological approaches used, and findings pertaining to key dimensions of parenting and parent–child relationships (e.g., warmth, conflict, attachment), and children’s cognitive and social-emotional development. The chapter also points to gaps in knowledge and new directions that will be fruitful for basic and applied research in future.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
- Parenting
- Genetics
- Epigenetics
- Hormones
- Neural activity
- Cognitive development
- Social-emotional development
Introduction
Humans are psychological beings who also are biological beings. Much of what we experience in our own development into adulthood and as parents, as well as the processes that influence the development of our children, operates through evolved biochemical processes involving molecules, cells and cell systems, and organs and organ systems. These biological complexes have evolved to provide sustainable structures in the body that would support a vast array of phenotypes (i.e., observable characteristics)—but this arrangement need not be equated with determinism. On the contrary, these biological complexes have evolved to be responsive to information coming in from the external environment. Parenting, and its influences on children’s development, is no exception. Sexual reproduction, complex social structures of kinship (that often transcend genetic relatedness), and sustained periods of child-rearing ensure opportunities for stability and change that support adaptive plasticity in biological, psychological, behavioral, and ecological systems. This plasticity is necessary for increasing the odds of survival of individuals and of the species.
In this chapter on parenting, we examine the intersection of children’s development and biological factors. Biology is a vast concept. We have included information in this chapter on a specific set of factors that have been examined in child and adolescent development research. These factors span levels of structures and their functions with respect to psychological and behavioral outcomes: genetics, groups of neurons and neural systems, and hormones. Much of the literature has focused on monoamine neurotransmitters (e.g., dopamine, serotonin, norepinephrine), and the hypothalamus–pituitary–adrenal or HPA axis in the body. These are major domains of biological research in parenting, in part because of their central roles in healthy and maladaptive development and functioning. These domains also lend themselves to empirical inquiry in human and animal model studies, making it feasible to examine biological structures and functions in laboratory and field settings.
Child development is another broad and encompassing concept. For our purposes, while considering the three biological domains just defined above (genetics, neural systems, hormones), we address a variety of aspects of development spanning social-emotional (including aspects of emerging psychopathology), and cognitive (including processing of information). At the intersection of these three biological and two broad developmental domains, we have organized the chapter around two major foundations in parenting research and children’s development: attachment security, and harsh adverse child-rearing experiences. Because so much of the biologically informed research on parenting is on adults and parents themselves, we conclude the chapter on that topic, to set the stage for considering the role of bio-environmental processes in the intergenerational transmission of parenting behaviors. Rather than attempting to comprehensively review the vast literature in all these areas of biopsychological research, we instead provide an overview of the major methods and levels of analysis being used, and cite studies that exemplify the use of each of these approaches. At times, we also make reference to the most recent edited handbooks that are relevant for each domain, for any readers who are interested in delving more deeply into specific topics or methods.
Background on Biological Measures
The most comprehensive theory that has guided research on biological factors in parenting is the bioecological model (Bronfenbrenner & Ceci, 1994). This model was a revision of Bronfenbrenner’s earlier theory, in which the authors elaborated many premises about how biological factors develop and interact with nonbiological factors at many levels of a constantly developing system within and outside of the child (e.g., family, school/peer, neighborhood, culture). Development of the bioecological interface operates via proximal processes in the interaction between neurobiology and environment within each person, operating in a multilevel context of ecological niches that change over time. Two other theories also have played a major role in the integration of biological measures: attachment theory, and a group of parenting stress theories. These will be elaborated on later.
There are a wide variety of measurement approaches for examining the role of biological factors in child development, and these approaches are no different in that subset of the literature that focuses more specifically on parenting and home environments. There are far too many potential biological markers to consider in just one chapter. In what follows, we highlight the main domains of biological factors used in parenting and child development research: genetics, neural structure and activity, and hormones.
Genetics
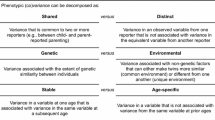
At their smallest and most molecular, biological influences in parenting and children’s development involve structural and functional variations in genes in the DNA molecule, as well as RNA molecules that transmit information from DNA into proteins. But long before our sciences had the capacity to measure and study DNA and RNA variation directly, researchers applied quasi-experimental epidemiological approaches to make inferences about the relative contributions of genetic and nongenetic (i.e., environmental) influences on observed phenotypes. These behavioral genetics methods, still in use today, utilize comparisons of genetically identical and nonidentical or fraternal twin pairs, genetically related and unrelated (i.e., adoptive and step) siblings, and genetically related and unrelated parent–offspring pairs. Genetic similarity of each pair is then used to account for variance in the phenotype similarity of each pair. This yields variance estimates representing genetic influences (i.e., heritability), and nongenetic influences that contribute either to the pairs’ similarity or not (i.e., shared and nonshared environment; for an overview see Knopik, Neiderhiser, DeFries, & Plomin, 2016).
Increasingly, scientists directly measure structural variations in DNA and RNA molecules, especially with automation, making the process fast, highly reliable, and relatively inexpensive compared to older technology (Demkow & Ploski, 2016). In family and developmental sciences, much of the work to date has focused on so-called candidate gene methods , whereby structural variations in genes thought to play a causal role in phenotypes of interest are measured and compared to estimate their statistical effects. Most common are studies of whole classes of genes involved in the production and regulation of monoamine neurotransmitters, such as dopamine, serotonin, and norepinephrine. This genetic information can also be used to test for potential interaction effects with measured environmental factors—including family and parenting variables (Deater-Deckard, Chen, & El Mallah, 2016). In addition, scientists increasingly are studying epigenetic alterations to DNA—biochemical modifications (e.g., methylation, acetylation, histone modification) that alter gene expression in response to input from outside the nuclei in the cells (Zhang & Meaney, 2010). In this chapter, we present an illustrative handful of examples of behavioral and molecular genetic studies that illustrate some of the important principles of how scientists make inferences about gene–environment transactions in parenting and children’s development. Those who wish to explore these genetics literatures more thoroughly can explore the work presented in handbooks by Hood and colleagues (2010), and Horwitz and Neiderhiser (2015).
Neural Structures and Functions
Individual nervous system cells, neurons, operate in groups as part of cell networks that connect specialized brain regions (e.g., prefrontal cortex, intraparietal sulcus) and structures (e.g., amygdala, hippocampus), as well as connections to neurons throughout the body via the spinal column. In parenting and family science, by far the most widely studied domain of the nervous system is the autonomic nervous system (ANS). The ANS is comprised of sympathetic and parasympathetic branches that interact to produce continuous responses to changes in the environment, and regulation of the body to maintain homeostasis (Sapolsky, Romero, & Munck, 2000). There are a wide variety of techniques used to measure, directly or indirectly, the neural activity of specific groups of neurons and their functional role in an observed phenotype.
By far the most common (and oldest) such approaches in parenting and family science are psychophysiological methods that record individual differences in changes in neural and cardiovascular functions. Electroencephalography (EEG) is a method of testing electrical activity in the brain, and electrocardiography (ECG) is a method of measuring the heart’s electrical activity, using sensory electrodes placed on specific locations on the body. Both methods have been used in parenting and child development research to examine individual differences and ontogeny (for an overview of psychophysiological methods in developmental and family science, see Schmidt & Segalowitz, 2008).
Most recently, scientists have begun applying neuroimaging using structural and functional magnetic resonance imaging (sMRI and fMRI, respectively) methods, to explore and test hypotheses about biosocial processes in parenting and children’s development. sMRI captures variations in the absolute and relative (to others) brain volumes in specific locations as well as the whole brain, and proportions of white matter (i.e., myelinated axons connecting brain cells) and gray matter (i.e., neuronal cell bodies, axons, glial cells, synapses between neurons, and blood vessels). fMRI assesses changes in blood flow in the brain that are thought to correspond with shifts in neural activation during and after the presentation of stimuli (Johnstone, Kim, & Whalen, 2009). Although the field is rapidly adopting MRI techniques to study aspects of parenting and children’s development and the parent’s brain (e.g., Abraham et al., 2014; Kim et al., 2010), MRI studies of parenting and the child’s brain are rare because it is difficult, if not impossible, to gather functional brain activity information with young children in a wakeful state (Johnson, Page, Williams, Wassemer, & Whitehouse, 2002). However, there have been several highly cited studies of anatomical differences in the brains of children exposed to different child-rearing environments, and we review those in this chapter.
Hormones
A third major domain of biological factors and their role in parenting and children’s development focuses on hormones—molecules produced by glands throughout the body, that convey information to nearby and distant organs and bodily systems to maintain or alter current functions. Of particular interest to developmental and family scientists are molecules involved in the stress response—cortisol and alpha amylase, in particular. The stress response is intimately tied to the functioning of the HPA axis, which is the fundamental neuroendocrine vector that prepares for and executes responses to potential and actual threats, then enables regulation so that the individual can return to a calmer, non-stressed state (i.e., homeostasis; Sapolsky et al., 2000).
Cortisol plays a key role in short-term stress responses, but this cortisol reactivity can signal dysfunction in the stress and self-regulation endocrine and physiological system if it is chronically hyperactive or hypoactive. Furthermore, HPA development is influenced by caregiving from early in life, such that warm and sensitive parenting (including secure attachment) predicts subsequent healthy HPA functioning (Francis & Meaney, 1999). In contrast, prenatal distress and postnatal maltreatment may have long-lasting deleterious effects on HPA functioning as indicated by cortisol reactivity (Hostinar & Gunnar, 2013; Matthews, 2002). In this chapter, we present illustrative correlational and experimental studies that show the critical role that cortisol and other hormones (e.g., oxytocin) play in parenting and children’s developmental outcomes.
We now turn to several key substantive areas of parenting research and biological factors, chief among them the literatures on attachment security, and harsh parenting and adverse environments. We also consider the smaller literature on fathering, to bring special attention to that growing literature—given that nearly all relevant research to date has focused on mothers. We complete the review by examining biological factors on parenting itself, and how this can inform us about the likely effects on child and adolescent development and the intergenerational transmission of parenting.
Attachment
One of the foundational concepts in parenting science and developmental psychology is that of attachment security and its importance in healthy development (van Bakel & Hall, 2018). The study of this key aspect of human development is rooted in attachment theory, which posits a critical causal role of the child’s developing social cognitive model of human relationships that influence feelings of safety and being loved, that have extensive strong links with variability in a wide range of social-emotional, cognitive, and physical outcomes (for an overview, see Cassidy & Shaver, 2016). According to this theory and empirical evidence, attachment security is derived from the earliest experiences in infancy based on sensitive, responsive caregiving—or, insecure attachment can arise from insensitive, nonresponsive, or harsh caregiving. There has been a long-standing interest in the potential role of biological factors in the attachment relationship, and how deficiencies in those biological factors might interfere in healthy development by altering the trajectory of attachment security from early in childhood. Some of the foundational work in this area sought to tease apart the roles of genetic and environmental factors using twin and adoption quasi-experiments (e.g., Fearon, Shmueli-Goetz, Viding, Fonagy, & Plomin, 2014; O’Connor & Croft, 2001; Roisman & Fraley, 2008).
More recently, parenting and developmental scientists interested in biological factors have focused on more direct indicators of potential genetic influences on attachment using molecular genetic methods. From this literature, several studies have shown that maternal responsive parenting—along with a secure attachment relationship with the child—mitigates a genetic risk for insecure attachment and behavioral/emotional problems thought to be associated with dysfunctional serotonin regulation (i.e., serotonin transporter gene 5-HTT; Barry, Kochanska, & Philibert, 2008; Gilissen, Bakermans-Kranenburg, van IJzendoorn, & Linting, 2008). In a more recent study, researchers found evidence to suggest that a genetic risk factor for an overreactive stress response in the child (i.e., a single nucleotide polymorphism [SNP] in a glucocorticoid receptor gene FKBP5) is enhanced within dyads in which the child has an insecure attachment relationship with the mother (Luijk et al., 2010). What is apparent from this nascent field of research in attachment security and genetic factors in children is that there are likely to be interactions between insecure versus secure attachment and specific genetic risks for disrupted stress reactivity and self-regulation, in the prediction of trajectories of healthy versus maladaptive social-emotional and behavioral outcomes.
Turning to attachment research examining psychophysiological and neural activity factors, a number of studies have examined these biological parameters during the strange situation, a separation and reunion manipulation in the laboratory that challenges the young child’s attachment system to reveal likely attachment security or insecurity. These neurophysiological studies have shown that insecurely attached infants are more likely to have higher heart rates prior to and during the strange situation; they also may show reduced left frontal lobe brain activation, a pattern implicated in growth in behavioral withdrawal and avoidance strategies that predict subsequent internalizing problems (Dawson et al., 2001). More broadly, children with an avoidant attachment style tend to show physiological hypo-arousal prior to a stressor, such as entering an unfamiliar playroom with adult and peer strangers (i.e., lower resting heart rate, higher resting baseline respiratory sinus arrhythmia (RSA; Burgess, Marshall, Rubin, & Fox, 2003)). However, these same children may show hyper-arousal during and following such social stressors, such as those experienced in the separation phase of the strange situation (i.e., greater vagal withdrawal and higher salivary alpha-amylase levels [an indicator of HPA axis activity]; Hill-Soderlund et al., 2008). Similar types of disturbances in cardiophysiology have been observed among foster children who experienced early life adversity, such as physical neglect or sexual abuse. Foster children with disorganized attachment internal working models may have a hyperactive sympathetic nervous system, as seen during the strange situation (Oosterman, De Schipper, Fisher, Dozier, & Schuengel, 2010). Perhaps not surprisingly, these biological risks associated with avoidant attachment in early childhood may combine with broader aspects of temperament, to predict maladaptive outcomes later. Illustrative of this, Burgess et al. (2003) found that avoidant attachment in infancy, when coupled with uninhibited temperament, predicted externalizing behavior problems at 4 years of age.
Psychophysiological and hormonal indicators of HPA axis activity and neural functioning more broadly have also been applied to studies of attachment at older ages. In one such study, secure attachment at 4-years of age was linked with better social skills at 8-years of age among children with more mature cortical development—itself indicated by higher EEG alpha power (Almas et al., 2012). In a more recent study of adolescents, investigators found that those with a dismissing attachment style showed greater amplitudes of negative left frontal slow-waves after experiencing rejection from peers in a virtual ball toss game; characteristically, these same individuals also underreported feeling distressed following their peer rejection (White et al., 2012).
An event-related potential (ERP) is a specific indicator from the EEG capturing a measured brain response that is the result of a specific sensory, cognitive, or motor event. ERP studies in combination with EEG readings are often used to study underlying emotional or cognitive processes in children. It allows for precise temporal measurement of early cognitive processing; for example, a certain wave component of ERP may reflect attentional resource allocation. Insecure attachment in adolescence has been linked with ERP indicators. For example, in one study, adolescents who were characterized as having an insecure attachment with their parents had longer reaction times when processing negative emotion faces and words; these behavioral biases were accompanied by neural signals—larger P1 and smaller N170 amplitudes—that indicate greater vigilance and a negativity bias when viewing faces (Escobar et al., 2013).
More broadly, insecure attachment in childhood is associated with greater cortisol reactivity in response to a stressor (Ahnert, Gunnar, Lamb, & Barthel, 2004; Bernard & Dozier, 2010; Schieche & Spangler, 2005). This effect and its link with behavior are even more pronounced when insecure attachment is coupled with high levels of behavioral inhibition (Schieche & Spangler, 2005). When taken together, the biologically informed studies in this growing literature on attachment and neurophysiological reactivity and regulation all point to the critical role that attachment security plays in the development of typical, healthy cerebral, cardiovascular and neuroendocrine responses to stressors—and the effective self-regulation of those stress responses.
Adverse Environments and Stress
In addition to the foundational work on attachment relationships and security in children’s development, biological factors have been examined in an even broader sense with respect to a wide range of adverse parenting and home environments and experiences in childhood that are well established risk factors in child and adolescent development. Chief among the multitude of potential adverse experiences are exposure to chronic parenting stress; this emerges as child maltreatment (i.e., abuse and neglect) and, far more commonly, harsh reactive parenting. According to a group of distinct yet complementary parenting stress theories (e.g., Abidin’s theory, Crnic’s theory; for an overview, see Deater-Deckard & Panneton, 2017), these early child-rearing experiences—particularly if they occur at sensitive periods in development or are chronic and severe—usually have lasting effects not only on children and adolescents but on parents and parenting behavior. According to these theories, parenting stress emerges in contexts in which the parent’s resources for managing the roles and demands of caregiving are exceeded by those demands—an imbalance that produces a sustained stress response within the parent that spills over into relationship dynamics and household environments that have a big impact on the developing child (See Box 1). As with the attachment literature reviewed above, researchers studying parenting stress and adverse child-rearing experiences have incorporated a variety of biological factors into their theories and methods, to examine the interface of these more extreme environments and underlying biological risk and resilience indicators that enhance or dampen the effects of those experiences.
Considering first the genetics research literature, studies have examined early adversity, maltreatment, and harsh parenting exposure—and how these experiences may interact with genetic factors in predicting subsequent trajectories of adaptive versus maladaptive functioning . Numerous behavioral genetic studies of parenting and children’s development have been conducted. These rely on designs assessing twins and adoptive siblings or parent–child dyads, to examine the interface of genetic and nongenetic influences on parenting and child outcomes alike (for an overview, see Horwitz & Neiderhiser, 2015). Perhaps the most important finding to emerge from this broader behavioral genetic literature is that children who are genetically at risk for more challenging behavioral and emotional problems, are more likely to experience harsher parenting—a so-called evocative gene –environment correlation or child effect on the parenting environment (for a meta-analysis, see Avinun & Knafo, 2014).
Turning to molecular genetic research that has examined actual variations in DNA structure , most of the relevant studies have focused on dopamine and serotonin neurotransmitter genes and their role in parent–child antagonistic relationship processes. The first such study of humans found that a functional polymorphism in the gene that expresses monoamine oxidase A or MAOA (a neurotransmitter-metabolizing enzyme) interacted with boys’ childhood maltreatment history to predict different antisocial behavioral outcomes years later, depending on the version of the gene (Caspi et al., 2002). Since then, the field has witnessed an explosion of studies—an exciting direction in research, though notably, few if any of the gene–environment interaction effects have been replicated consistently. Still, potentially promising findings have emerged regarding harsh versus supportive child-rearing and its interactions with dopamine receptor 2 gene (e.g., Mills-Koonce et al., 2007; Propper et al., 2008), dopamine receptor 4 gene (e.g., Bakermans-Kranenburg, van Ijzendoorn, Pijlman, Mesman, & Juffer, 2008; Berry, Deater-Deckard, McCartney, Wang, & Petrill, 2013), catechol-O-methyltransferase gene (COMT ; Voelker, Sheese, Rothbart, & Posner, 2009), and serotonin transporter gene (e.g., Fox et al., 2005; Koss, Cummings, Davies, Hetzel, & Cicchetti, 2016; Stein, Schork, & Gelernter, 2008). Although little is known currently about precisely how adverse experiences interact with gene variants in predicting developmental outcomes, the current prevailing view is that these effects are explained by proximal changes in gene expression of neurotransmitters, via epigenetic modifications to the DNA molecule (for recent reviews, see Deater-Deckard, Chen, & El Mallah, 2016). This nascent literature on epigenetic changes in children following exposure to stressors in utero and postnatally suggests that such modifications to gene expression can occur, and may ultimately help explain some of the gene–environment interaction effects already published and yet to be found. However, major challenges remain with respect to methodology and inferences, given the nature of the molecular biological mechanisms involved.
Considering next the relevant neural activity research, scalp electrophysiology and neuroimaging methods have been used to examine the potential impact of maltreatment and other early adversities on children’s developmental outcomes. In one such study (Hanson et al., 2010), children who had experienced physical abuse had smaller brain volumes in the right orbitofrontal cortex (a region of the prefrontal cortex that has been implicated in many aspects of emotion and decision making; see Stalnaker, Cooch, & Schoenbaum, 2015), compared to those who had not been abused. This variation in brain volume was, in turn, associated with poorer social relationships and social behaviors. In another study (De Bellis et al., 2002), researchers found that maltreated children had larger gray matter and smaller white matter volumes in the superior temporal gyrus. Anomalies in this structure have been implicated in language and social cognitive deficits in children and adolescents (e.g., Bigler et al., 2007).
In a longitudinal study of infants from institutions (McLaughlin, Fox, Zeanah, & Nelson, 2011), investigators found that these children were more likely to have longer periods of right-lateralized EEG alpha power and a blunted rebound in left-hemisphere alpha power. This right-lateralized pattern was associated with subsequent internalizing symptoms at 54 months of age—a finding that is consistent with other studies showing that right-lateralized alpha power is indicative of poorer emotion regulation and higher levels of fear and sadness (Bell, Schwartz, Hardin, Baldwin, & Kline, 1998; Gotlib, Ranganath, & Rosenfeld, 1998). Interestingly, McLaughlin et al. found that the children who were placed into foster care prior to 24-months of age generally fared better, suggesting that earlier removal from institutional care and placement in an enriched, loving home can ameliorate or mitigate these effects.
Psychophysiological studies of parenting and children’s development also have incorporated ECG indicators of heart rate variability (HRV) , respiratory sinus arrhythmia (RSA) , or vagal tone (so named because it represents the activity of the vagus nerve, which inhibits heart beats). These are three closely related concepts that all reflect the degree to which heart rate varies from beat to beat. Lower levels indicate relatively poor parasympathetic regulation of heart rate, whereas higher levels indicate better parasympathetic regulation in children, adolescents and adults (Porges, 2003; Thayer & Lane, 2009). Better cardiac parasympathetic regulation has been linked to a host of cognitive, social-emotional and behavioral indicators of social competence, efficient cognitive processing, and self-regulation (Bridgett, Burt, Edwards, & Deater-Deckard, 2015; Calkins, Graziano, & Keane, 2007).
Higher HRV or vagal tone are indicative of resilience and well-regulated stress reactivity, and may be particularly important for children living in chronically stressful homes and parenting environments (Obradović, Bush, Stamperdahl, Adler, & Boyce, 2010). For example, children with higher vagal tone may be protected from the negative effects of parental marital conflict on internalizing and externalizing symptoms, as well as physical health problems (El-Sheikh, Harger, & Whitson, 2001; Katz & Gottman, 1997). Similarly, children with higher vagal suppression, which is associated with fewer behavior problems and better social skills, may be protected from the negative effects of mothers’ poorer emotion socialization behaviors (Perry, Calkins, Nelson, Leerkes, & Marcovitch, 2012).
Turning to brain-imaged neural factors, children with a history of parental maltreatment may develop less gray matter volume in the right temporal lobe, especially if coupled with posttraumatic stress disorder (De Bellis et al., 2002). This population of children may also have smaller total volume in the superior posterior cellular lobes (Bauer, Hanson, Pierson, Davidson, & Pollak, 2009) and hippocampus (Bremner et al., 2003), yet greater amygdala volume (Tottenham et al., 2010). Maltreated children tend to show less white matter connectivity (Eluvathingal et al., 2006), and less efficient neural activity in the orbital frontal cortex, prefrontal infralimbic cortex, lateral temporal cortex, medial temporal structures, and brain stem (Chugani et al., 2001). In part because of these structural changes, maltreated children do not perform as well as non-maltreated peers on a wide range of cognitive tasks involving memory, attention, and executive functions—even though maltreated children show greater brain activation during these tasks because they must allocate more resources to do them (Carrion, Garrett, Menon, Weems, & Reiss, 2008; Mueller et al., 2010).
In addition to MRI variables, ERP signals have been used to study neural activity and maltreatment. Institutionalized children have been shown to display larger N170 amplitude (a general signal indicated during face processing) when viewing fearful faces (Parker & Nelson, 2005). In several studies, physically abused children have been found to display a larger “P3b” amplitude—a component reflecting allocation of resources for attention—while viewing their mothers’ angry faces (Pollak & Sinha, 2002; Shackman, Shackman, & Pollak, 2007). In a similar study of behavior, children who had been physically maltreated displayed more negative affect and allocated more attention to social threat (as measured by higher P3b amplitudes) when they had to control their frustration with a virtual child who was performing poorly on a task and losing points on the game for the target child (Shackman & Pollak, 2014). Another ERP signal, the error-related negativity or ERN, has been studied because it is an indicator of the neural response to errors. Previous research established that higher error related brain activity is associated with internalizing symptoms, but lower error related activity is linked to externalizing symptoms (Olvet & Hajcak, 2008). Two studies have shown that greater ERN amplitudes are found during inhibitory control or attention task performance, among children who experienced harsher, more punitive parenting when they were toddlers (Brooker & Buss, 2014; Meyer, Hajcak, Torpey-Newman, Kujawa, & Klein, 2015).
Biologically informed research of adverse child-rearing effects has also incorporated neuroendocrine assessment methods to examine stress hormones as indicators of HPA axis dysfunction among high-risk youth. This approach is well illustrated in a recent longitudinal study spanning the first 6 years of life, which showed that children of depressed mothers had higher cortisol levels and less cortisol variability compared to children of non-depressed mothers (Apter-Levi et al., 2016). The investigators also found that lower levels of cortisol variability were associated with higher levels of child behavioral and emotional problems, as well as social withdrawal. Even more robust effects on stress hormones are seen among previously institutionalized children placed in foster care or adoptive homes. Children from the most severe neglectful early environments, often show the highest cortisol levels prior to a laboratory task, as well as following interactions with their foster or adoptive mothers (Fries, Shirtcliff, & Pollak, 2008).
There have been several relevant hormone studies that have focused specifically on fathering —a very neglected domain of parenting research in the larger field of biopsychological studies of children’s development. In one study, fathers who received external oxytocin displayed warmer parenting behavior and higher RSA during observed parent–child interaction. In turn, infants of these fathers displayed elevations in oxytocin and RSA, as well as greater social reciprocity during interaction with their fathers (Weisman, Zagoory-Sharon, & Feldman, 2012). In another study, higher levels of paternal negativity toward his infant were associated with greater increases in cortisol reactivity in response to emotionally arousing tasks at 2-years of age; this finding suggests that harsher fathering behavior may increase stress reactivity and impede emotion and stress regulation, as these systems develop rapidly in very early childhood (Mills-Koonce et al., 2011). Conversely, these investigators also found that positive fathering behavior, such as high levels of involvement in infancy, can serve as a buffer against mental health problems when the children are older—especially among those infants who had hyperreactive cortisol increases in response to social stressors.
Overall, the human research literature on hormones, parenting and children’s development has relied on correlational studies. However, experiments have shown the key role that hormones play. Family and parenting interventions with foster children (a population that is at risk for insecure attachment relationships) have shown that effective interventions result in developmentally typical and healthy cortisol patterns, along with reductions in behavioral and emotional problems and improvements in the attachment relationship (Dozier, Peloso, Lewis, Laurenceau, & Levine, 2008; Fisher, Gunnar, Dozier, Bruce, & Pears, 2006). Together, the correlational and experimental studies reviewed above suggest that deprivation in the social caregiving environment early in life can have a lasting impact on the stress reactivity and self-regulation of children, even after exposure has ended.
Parents’ Biological Factors
Parents are biological beings too. Up to this point in the chapter, we have focused on studies examining parenting and children’s development that have assessed biological indicators in the children. However, there is a large and growing literature on the same biopsychological methods, applied to parents—and a much larger literature when one includes studies of adults more generally, that have examined parenting-relevant aspects of adult functioning. In this final section, we consider some of the key findings and their implications for how developmental and family scientists think about intergenerational transmission of biologically based risk and resiliency in development.
Attachment theory is a good place to return to at this point in our review. It was one of the first theories in developmental and social psychology to stipulate cognitive constructs that develop in childhood but remain active throughout the lifespan—and more importantly for the present chapter, constructs that influence adults’ romantic and platonic relationships as well as their relationships with their own children (Cassidy & Shaver, 2016). The literature on adult attachment in romantic partnerships is also relevant to understanding parenting, given that these adults are also responsible for coparenting when they have children together—and both partners’ attachment security have been implicated in coparenting and parent–child relationship quality (Roisman, Madsen, Hennighausen, Sroufe, & Collins, 2001).
Genetically informed research has documented the evidence of heritable variance, as well as substantial nongenetic variance, in adult attachment style as well as its link to adult psychopathology (Brussoni, Jang, Livesley, & Macbeth, 2000; Crawford et al., 2007). One molecular genetic study found preliminary evidence for a potential risk allele in the serotonin neurotransmitter system for insecure attachment in early adulthood—the effect of which may be modulated by the patterns of maternal sensitivity experienced earlier in childhood (Fraley, Roisman, Booth-LaForce, Owen, & Holland, 2013). The adult attachment research literature also includes several studies that have incorporated electrophysiology methods, showing distinct patterns of arousal and alertness via EEG being associated with anxious and dismissive attachment styles (e.g., Roisman, 2007; Roisman, Tsai, & Chiang, 2004; Verbeke, Pozharliev, Van Strien, Belschak, & Bagozzi, 2014).
ERP studies have shown that insecure adults show distinct features of information processing of visual and auditory stimuli involving positive and negative emotions and social contexts (Chavis & Kisley, 2012; Cohen & Shaver, 2004; Dan & Raz, 2012; Rognoni, Galati, Costa, & Crini, 2008; Zilber, Goldstein, & Mikulincer, 2007). Numerous fMRI studies have also been conducted, showing that various types of insecure adult attachment are associated with distinct hyperactivation or hypoactivation of brain regions involved in emotion processing and regulation such as anterior temporal pole, orbitofrontal and prefrontal cortex, amygdala, somatosensory cortex, and structures in dopaminergic reward circuitry (Buchheim et al., 2006; DeWall et al., 2012; Donges et al., 2012; Gillath, Bunge, Shaver, Wendelken, & Mikulincer, 2005; Lemche et al., 2006; Suslow et al., 2009; Vrtička, Andersson, Grandjean, Sander, & Vuilleumier, 2008; Zhang, Li, & Zhou, 2008).
These neural features of adult attachment security overlap with known regions associated with social emotional responding (including amygdala, frontal cortex, insula, and medial temporal cortex) that are linked with variation in sensitive caregiving (for a review, see Swain, 2011).
Turning to studies of caregiving of children, EEG studies of mothers at various time points in their children’s lives have yielded evidence for a neural basis for parenting behaviors that have an influence on subsequent child development outcomes. Mothers appear to be perceptually sensitive (as evidenced by N100, N170 and left positive potential amplitudes) to infant-specific auditory and visual stimuli (Peltola et al., 2014; Proverbio, Brignone, Matarazzo, Del Zotto, & Zani, 2006; Purhonen et al., 2001). The amplitude of this response seems to be strongest when viewing or hearing distress cues from infants, suggesting this aspect of affective processing may be unique and important to enabling mothers to respond sensitively to their infants. These neural processes may not operate as well or efficiently, however, for mothers who are prone to neglecting or abusing their young children (Rodrigo et al., 2011).
Most recently, a special issue on neurobiological factors in parenting was published in which investigators presented physiological and neuroimaged indicators of attentional and emotional processing, stress reactivity, and self-regulation processes (Deater-Deckard & Sturge-Apple, 2017). Three papers utilized ECG or EEG to operationalize individual differences in parental physiological and neurological reactivity and regulation. In one, investigators studied the association between parents’ physiological reactivity to experimentally induced positive and negative moods, and their observed positive and negative affect when engaging with their adolescents in a variety of discussion tasks. They found that RSA suppression in response to a sad movie clip was associated with less anger during a conflict discussion task, with the opposite effect found for parents who displayed RSA augmentation. In response to the amusing movie, there was an interaction effect with parental depression; parents with low levels of depression who also displayed greater RSA augmentation to the amusing film were observed to show greater positive affect when discussing a recent disagreement with their child (Connell, Dawson, Danzo, & McKillop, 2017).
In the second relevant study from the special issue, researchers examined parental heart rate dynamics and their links with observed parent–child (6- to 12-year-olds) interactions in a conflict resolution task. Parents who displayed a heart rate increase followed by a decrease in response to the conflict task, were more likely to be emotionally sensitive and responsive during the interaction (Han, Zhang, Cui, & Yan, 2017). In the third study, the harshest parenting was found among mothers who showed a combination of deficits in executive function and the least well-regulated physiological response (based on EEG and ECG reactivity measures) to cognitive challenge (Deater-Deckard & Bell, 2017)—a finding that builds on prior evidence that poorer parasympathetic regulation (as indicated by lower vagal tone) is part of a broader matrix of biological and cognitive deficits in emotion regulation that are particularly deleterious in their effects on parenting under chronic stress conditions (Deater-Deckard, Li, & Bell, 2016).
Attachment and parenting research has also examined the role of adults’ hormones. Women with avoidant attachment styles show greater cortisol reactivity and slower recovery following a conflict with their partners. In contrast, for men, it is an anxious attachment style that is associated with greater cortisol activity (Laurent & Powers, 2007; Powers, Pietromonaco, Gunlicks, & Sayer, 2006). In addition to cortisol, oxytocin has been studied in the adult’s attachment relationships with partners and children. Oxytocin is a hormone that is associated with greater affiliation and feelings of warmth and that operates as a key component of the parent–child and parent–parent couple bond and relationship (Neumann, 2008). Oxytocin levels are higher in both mothers and fathers after contact with infants (Feldman, Gordon, Schneiderman, Weisman, & Zagoory-Sharon, 2010; Ross & Young, 2009) and this response seems to modulate warm and supportive parenting in conjunction with increased activation in the amygdala and the frontal cortex (Kim et al., 2010).
Oxytocin has been manipulated experimentally to show changes in parent–child closeness and decreased stress reactivity. This is demonstrated in several studies examining adversity in early childhood (i.e., beyond attachment security), in which externally administered oxytocin has been shown to reduce cortisol (a stress hormone) in adults, but only among those who had not had childhood adverse experiences (e.g., parental divorce, abuse history; Meinlschmidt & Heim, 2007). The impact of earlier child-rearing experiences on oxytocin may extend to how adults process faces and emotions. For instance, an experimental study of college undergraduates found that it was the combination of externally administered oxytocin and a history of maternal love withdrawal that predicted enhanced processing of certain faces and emotions (Huffmeijer et al., 2013). In another experimental study, women were randomly assigned to receive external oxytocin or not, and then were given a handgrip dynamometer (to measure grip strength) as they listened to infants crying. External administration of oxytocin led to weaker grip responses, if the participants reported positive experiences with parental discipline in their childhoods. Those who had experienced harsh discipline had a strong grip response to infant cries, regardless of oxytocin exposure (Bakermans-Kranenburg, van IJzendoorn, Riem, Tops, & Alink, 2012). In sum, as the growing experimental literature demonstrates, increasingly researchers will be using experimental designs to manipulate hormones and other biological factors (including neural activity and gene expression), which will permit much stronger causal inferences that will inform prevention and intervention efforts.
Caveats and Future Directions
Humans have evolved as part of an exquisitely complex set of biological systems that work to ensure survival and reproductive success. These systems span hormones, neurons and neural systems, and genes within our cells. Parenting behavior, and the effects of parenting on children’s developmental trajectories, all involve biological influences across levels of these systems. These influences do not determine outcomes, but they do reflect meaningful information about individuals’ acute/phasic and chronic/tonic responses to their environments.
In the long history of parenting and developmental science, scientists have incorporated direct measures of biological factors only relatively recently. The scope and depth of this recent growth in biopsychological methods has been astonishing, and its impact on our understanding of the causes and consequences of parenting behaviors is hard to refute. Instead of relying on assumptions about genetic factors in traditional behavioral genetic predictive models of parenting and children’s outcomes, now scientists are directly measuring genetic and epigenetic variation in specific regions of the genome, to test competing theories of gene–environment transactions in development. In the past, researchers relied on precise measurement of specific behaviors that were thought to be tied to underlying neural factors (based largely on animal models), but today they are relying on assessments of physiological changes in neurological and neuroendocrine chemical and hematic information throughout the central and peripheral nervous system.
As important and rigorous as much of the advances in this area of research may be, it comes with new limitations—some of which can be addressed through further advances in technology, but some of which cannot. The most fundamental limitation in most of the human parenting research remains, regardless of measurement techniques and indicators—that the vast majority of the empirical base has used correlational designs. This is because for many (and perhaps most) of the questions parenting scientists and practitioners seek to answer, it would be unethical to conduct rigorous experiments with random assignment (e.g., randomly assigning children to parents), let alone executing such designs that involve direct manipulation of biological parameters (e.g., using drugs to alter gene expression).
There are two major exceptions to this fundamental limitation. The first is that rigorous experiments on caregiving are conducted with animal models—but even this exception raises another limitation regarding whether such experiments generalize to humans. The second is that it is feasible to incorporate biological measures of relevant biological processes into human experiments when done as part of clinical trials to measure efficacy of parenting interventions. It is already apparent that this second exception is the main route through which parenting science will be most successful in incorporating biological measures into true experiments.
Implications for Policy and Practice
With every technological advancement in genotyping, electrophysiology, and neuroimaging, parenting and developmental scientists will have even greater access to reliable methods for measuring species-typical change and individual differences in developmental trajectories across many levels of these biological systems. This is exciting; in our view, there is no better way to demonstrate the power of improving the environments and lives of children and their parents, than to show how biological markers of stress and health can be changed as a result of such environmental enrichment. Therein lies much of the future of biopsychological research in parenting science: examining the biosocial interface through quasi experiments and experiments that are part of the broader effort to create parenting intervention tools that are evidence-based.
What we do with the mounting information on biological factors will need to be informed by the next generation of biopsychological theories of human development that will evolve from attachment theory (Cassidy & Shaver, 2016), parenting stress theories (Deater-Deckard & Panneton, 2017), and the bioecological model of Bronfenbrenner and Ceci (1994). These future-generation theories will drive the hypothesis testing that will inform innovations in policy and practice regarding children and parenting. These innovations will more fully integrate biological factors into prevention and intervention tools and delivery methods.
There are two broad implications already known, and others will emerge as theory and empirical work evolve. First, there is great potential for assessment and understanding of specific biological parameters to create new prevention and intervention targets and tools—ways to directly or indirectly manipulate a biological process in ways that directly alter the cognitive or behavioral outcome of concern. This has always been the premise of pharmacological interventions. More recent examples of new intervention tools include transcranial direct current stimulation (tDCS) for treatment-resistant chronic depression and anxiety (Lefaucheur et al., 2017), and gene therapies for a host of diseases (as seen in any issue of The Journal of Gene Medicine, Gene Therapy, and many others).
Second, there also is potential that our understanding of biological pathways will help interventionists deliver tools that are individualized to each person, in a way that is more likely to be effective and have the fewest side effects—so-called personalized intervention (e.g., Ng & Weisz, 2016). The potential of this premise is that individualizing prevention and intervention that best fits each parent’s or child’s biological and cognitive-behavioral profile, will yield the most effective and longest lasting changes that benefit the family. Both implications are only beginning to be realized in actual practice.
Finally, at the level of broad family and child policies in communities, states/regions, and nations, some policy makers will always take evidence of biological contributions as proof that social and economic interventions will not be effective—a biological determinism that has been the root of policy that either neglects families and their needs, or attempts to decide who gets to become a parent (e.g., eugenics; Berryessa & Cho, 2013). Today, scientists and policy makers alike increasingly realize that the information about how biology contributes to child development, in part through sexual reproduction and parenting environments, informs and does not negate the need for relevant social and economic policy (for an overview see Hatemi & McDermott, 2011). The challenge for parenting scientists and policy makers is to build consensus about how to utilize the empirical evidence when changing legislation and regulations, in ways that acknowledge the complex, transactional interface of biology and environment.
Box 1 The Importance of Parental Leave Policies
A wealth of research, some of which is highlighted in this chapter, demonstrates the essential nature of developing healthy and nurturing relationships with caregivers early in life. Secure attachment to a caregiver has been shown to contribute to social and academic competencies (Drake, Belsky, & Fearon, 2014) and decreased risk for behavioral problems (Kochanska & Kim, 2013). Of particular relevance, positive caregiver relationships can serve as a protective factor even in the context of early adversity (Fisher et al., 2006; Luby et al., 2013). Positive behavioral and academic associations like these are not only limited to parental caregiving bonds, but also are found in high-quality childcare environments (NICHD Early Child Care Research Network, 2002).
The USA is one of the only countries in the world where employers are not required to provide paid time off for parents following the birth or adoption of a child (OECD, 2017). Current parental and family leave policies are insufficient, and parents of infants and young children often struggle with access to high quality childcare options. Given the importance of early healthy relationships for later success, nations should continually review and improve equity in access to, and extent and quality of, their parental leave and childcare policies and programs (Nomaguchi & Milkie, 2017). Examples of new or long-standing policies can be found in many countries; these policies are particularly prominent in the Nordic and Baltic nations (e.g., Sweden, Denmark, Estonia). Policy analysis has shown that national differences in these policies help explain some of the nation-level variation in caregiving and family division of labor, work-family role conflict and stress, and children’s developmental outcomes (e.g., Altintas & Sullivan, 2017; Cooke & Baxter, 2010; Patton, Costich, & Lidströmer, 2017). The positive effects of such policies are seen in behavior and health (Hahn, 2015). It remains to be seen in future research whether such effects also are observed on underlying biological pathways and structures throughout the nervous and endocrine systems, through reductions in parental stress.
References
Abraham, E., Hendler, T., Shapira-Lichter, I., Kanat-Maymon, Y., Zagoory-Sharon, O., & Feldman, R. (2014). Father’s brain is sensitive to childcare experiences. Proceedings of the National Academy of Sciences, 111(27), 9792–9797. https://doi.org/10.1073/pnas.1402569111
Ahnert, L., Gunnar, M. R., Lamb, M. E., & Barthel, M. (2004). Transition to child care: Associations with infant–mother attachment, infant negative emotion, and cortisol elevations. Child Development, 75(3), 639–650. https://doi.org/10.1111/j.1467-8624.2004.00698.x
Almas, A. N., Degnan, K. A., Radulescu, A., Nelson, C. A., Zeanah, C. H., & Fox, N. A. (2012). Effects of early intervention and the moderating effects of brain activity on institutionalized children’s social skills at age 8. Proceedings of the National Academy of Sciences, 109(Suppl 2), 17228–17231. https://doi.org/10.1073/pnas.1121256109/-/DCSupplemental
Altintas, E., & Sullivan, O. (2017). Trends in fathers’ contribution to housework and childcare under different welfare policy regimes. Social Politics: International Studies in Gender, State and Society, 24(1), 81–108. https://doi.org/10.1093/sp/jxw007
Apter-Levi, Y., Pratt, M., Vakart, A., Feldman, M., Zagoory-Sharon, O., & Feldman, R. (2016). Maternal depression across the first years of life compromises child psychosocial adjustment; relations to child HPA-axis functioning. Psychoneuroendocrinology, 64, 47–56. https://doi.org/10.1016/j.psyneuen.2015.11.006
Avinun, R., & Knafo, A. (2014). Parenting as a reaction evoked by children’s genotype: A meta analysis of children-as-twins studies. Personality and Social Psychology Review, 14, 87–102. https://doi.org/10.1177/1088868313498308
Bakermans-Kranenburg, M. J., van Ijzendoorn, M. H., Pijlman, F. T., Mesman, J., & Juffer, F. (2008). Experimental evidence for differential susceptibility: Dopamine D4 receptor polymorphism (DRD4 VNTR) moderates intervention effects on toddlers’ externalizing behavior in a randomized controlled trial. Developmental Psychology, 44, 293–300. https://doi.org/10.1037/0012-1649.44.1.293
Bakermans-Kranenburg, M. J., van IJzendoorn, M. H., Riem, M. M., Tops, M., & Alink, L. R. (2012). Oxytocin decreases handgrip force in reaction to infant crying in females without harsh parenting experiences. Social Cognitive and Affective Neuroscience, 7(8), 951–957. https://doi.org/10.1093/scan/nsr067
Barry, R. A., Kochanska, G., & Philibert, R. A. (2008). G× E interaction in the organization of attachment: Mothers’ responsiveness as a moderator of children’s genotypes. Journal of Child Psychology and Psychiatry, 49(12), 1313–1320. https://doi.org/10.1111/j.1469-7610.2008.01935.x
Bauer, P. M., Hanson, J. L., Pierson, R. K., Davidson, R. J., & Pollak, S. D. (2009). Cerebellar volume and cognitive functioning in children who experienced early deprivation. Biological Psychiatry, 66(12), 1100–1106. https://doi.org/10.1016/j.biopsych.2009.06.014
Bell, I. R., Schwartz, G. E., Hardin, E. E., Baldwin, C. M., & Kline, J. P. (1998). Differential resting quantitative electroencephalographic alpha patterns in women with environmental chemical intolerance, depressives, and normals. Biological Psychiatry, 43(5), 376–388. https://doi.org/10.1016/S0006-3223(97)00245-X
Bernard, K., & Dozier, M. (2010). Examining infants’ cortisol responses to laboratory tasks among children varying in attachment disorganization: Stress reactivity or return to baseline? Developmental Psychology, 46(6), 1771–1778. https://doi.org/10.1037/a0020660
Berry, D., Deater-Deckard, K., McCartney, K., Wang, Z., & Petrill, S. A. (2013). Gene–environment interaction between DRD4 7-repeat VNTR and early maternal sensitivity predicts inattention trajectories across middle childhood. Development and Psychopathology, 25, 291–306. https://doi.org/10.1002/dev.21105
Berryessa, C. M., & Cho, M. K. (2013). Ethical, legal, social, and policy implications of behavioral genetics. Annual Review of Genomics and Human Genetics, 14, 515–534. https://doi.org/10.1146/annurev-genom-090711-163743
Bigler, E. D., Mortensen, S., Neeley, E. S., Ozonoff, S., Krasny, L., Johnson, M., … Lainhart, J. E. (2007). Superior temporal gyrus, language function, and autism. Developmental Neuropsychology, 31(2), 217–238. https://doi.org/10.1080/87565640701190841
Bremner, J. D., Vythilingam, M., Vermetten, E., Southwick, S. M., McGlashan, T., Nazeer, A., & Ng, C. K. (2003). MRI and PET study of deficits in hippocampal structure and function in women with childhood sexual abuse and posttraumatic stress disorder. American Journal of Psychiatry, 160(5), 924–932. https://doi.org/10.1176/appi.ajp.160.5.924
Bridgett, D. J., Burt, N. M., Edwards, E. S., & Deater-Deckard, K. (2015). Intergenerational transmission of self-regulation: A multidisciplinary review and integrative conceptual framework. Psychological Bulletin, 141, 602–654. https://doi.org/10.1037/a0038662
Bronfenbrenner, U., & Ceci, S. J. (1994). Nature-nurture reconceptualized in developmental perspective: A bioecological model. Psychological Review, 101(4), 568–586. https://doi.org/10.1037/0033-295X.101.4.568
Brooker, R. J., & Buss, K. A. (2014). Toddler fearfulness is linked to individual differences in error-related negativity during preschool. Developmental Neuropsychology, 39(1), 1–8. https://doi.org/10.1080/87565641.2013.826661
Brussoni, M. J., Jang, K. L., Livesley, W., & Macbeth, T. M. (2000). Genetic and environmental influences on adult attachment styles. Personal Relationships, 7(3), 283–289. https://doi.org/10.1111/j.1475-6811.2000.tb00017.x
Buchheim, A., Erk, S., George, C., Kächele, H., Ruchsow, M., Spitzer, M., & Walter, H. (2006). Measuring attachment representation in an fMRI environment: A pilot study. Psychopathology, 39(3), 144–152. https://doi.org/10.1159/000091800
Burgess, K. B., Marshall, P. J., Rubin, K. H., & Fox, N. A. (2003). Infant attachment and temperament as predictors of subsequent externalizing problems and cardiac physiology. Journal of Child Psychology and Psychiatry, 44(6), 819–831. https://doi.org/10.1111/1469-7610.00167
Calkins, S. D., Graziano, P. A., & Keane, S. P. (2007). Cardiac vagal regulation differentiates among children at risk for behavior problems. Biological Psychology, 74(2), 144–153. https://doi.org/10.1016/j.biopsycho.2006.09.005
Carrion, V. G., Garrett, A., Menon, V., Weems, C. F., & Reiss, A. L. (2008). Posttraumatic stress symptoms and brain function during a response-inhibition task: An fMRI study in youth. Depression and Anxiety, 25(6), 514–526. https://doi.org/10.1002/da.20346
Caspi, A., McClay, J., Moffitt, T. E., Mill, J., Martin, J., Craig, I. W., & Poulton, R. (2002). Role of genotype in the cycle of violence in maltreated children. Science, 297(5582), 851–854. https://doi.org/10.1126/science.1072290
Cassidy, J., & Shaver, P. (Eds.). (2016). Handbook of attachment (3rd ed.). New York, NY: Guilford Press.
Chavis, J. M., & Kisley, M. A. (2012). Adult attachment and motivated attention to social images: Attachment-based differences in event-related brain potentials to emotional images. Journal of Research in Personality, 46(1), 55–62. https://doi.org/10.1016/j.jrp.2011.12.004
Chugani, H. T., Behen, M. E., Muzik, O., Juhász, C., Nagy, F., & Chugani, D. C. (2001). Local brain functional activity following early deprivation: A study of postinstitutionalized Romanian orphans. Neuroimage, 14(6), 1290–1301. https://doi.org/10.1006/nimg.2001.0917
Cohen, M., & Shaver, P. (2004). Avoidant attachment and hemispheric lateralisation of the processing of attachment- and emotion-related words. Cognition and Emotion, 18(6), 799–813. https://doi.org/10.1080/02699930341000266
Connell, A. M., Dawson, G., Danzo, S., & McKillop, H. (2017). The psychophysiology of parenting: Individual differences in autonomic reactivity to positive and negative mood inductions and observed parental affect during dyadic interactions with children. Journal of Family Psychology, 31(1), 30–40. https://doi.org/10.1037/fam0000278
Cooke, L. P., & Baxter, J. (2010). “Families” in international context: Comparing institutional effects across Western societies. Journal of Marriage and Family, 72(3), 516–536. https://doi.org/10.1111/j.1741-3737.2010.00716.x
Crawford, T. N., Livesley, W. J., Jang, K. L., Shaver, P. R., Cohen, P., & Ganiban, J. (2007). Insecure attachment and personality disorder: A twin study of adults. European Journal of Personality, 21(2), 191–208. https://doi.org/10.1002/per.602
Dan, O., & Raz, S. (2012). Adult attachment and emotional processing biases: An event-related potentials (ERPs) study. Biological Psychology, 91(2), 212–220. https://doi.org/10.1016/j.biopsycho.2012.06.003
Dawson, G., Ashman, S. B., Hessl, D., Spieker, S., Frey, K., Panagiotides, H., & Embry, L. (2001). Autonomic and brain electrical activity in securely- and insecurely-attached infants of depressed mothers. Infant Behavior and Development, 24(2), 135–149. https://doi.org/10.1016/S0163-6383(01)00075-3
De Bellis, M. D., Keshavan, M. S., Shifflett, H., Iyengar, S., Beers, S. R., Hall, J., & Moritz, G. (2002). Brain structures in pediatric maltreatment-related posttraumatic stress disorder: A sociodemographically matched study. Biological Psychiatry, 52(11), 1066–1078. https://doi.org/10.1016/S0006-3223(02)01459-2
Deater-Deckard, K., & Bell, M. A. (2017). Maternal executive function, heart rate, and EEG alpha reactivity interact in the prediction of harsh parenting. Journal of Family Psychology, 31(1), 41–50. https://doi.org/10.1037/fam0000286
Deater-Deckard, K., Chen, N., & El Mallah, S. (2016). Gene-environment interplay in coercion. In T. Dishion & J. Snyder (Eds.), Oxford handbook of coercive dynamics in close relationships (pp. 23–38). New York, NY: Oxford University Press.
Deater-Deckard, K., Li, M., & Bell, M. A. (2016). Multi-faceted emotion regulation, stress, and affect in mothers of young children. Cognition and Emotion, 30(3-4), 444–457. https://doi.org/10.1080/02699931.2015.1013087
Deater-Deckard, K., & Panneton, R. (2017). Parental stress and early child development: Adaptive and maladaptive outcomes. New York, NY: Springer.
Deater-Deckard, K., & Sturge-Apple, M. (2017). Mind and matter: New insights on the role of parental cognitive and neurobiological functioning in process models of parenting. Journal of Family Psychology, 31(1), 5–7. https://doi.org/10.1037/fam0000300
Demkow, U., & Ploski, R. (2016). Clinical applications for next-generation sequencing. New York, NY: Academic Press.
DeWall, C. N., Masten, C. L., Powell, C., Combs, D., Schurtz, D. R., & Eisenberger, N. I. (2012). Do neural responses to rejection depend on attachment style? An fMRI study. Social Cognitive and Affective Neuroscience, 7(2), 184–192. https://doi.org/10.1093/scan/nsq107
Donges, U. S., Kugel, H., Stuhrmann, A., Grotegerd, D., Redlich, R., Lichev, V., & Dannlowski, U. (2012). Adult attachment anxiety is associated with enhanced automatic neural response to positive facial expression. Neuroscience, 220, 149–157. https://doi.org/10.1016/j.neuroscience.2012.06.036
Dozier, M., Peloso, E., Lewis, E., Laurenceau, J. P., & Levine, S. (2008). Effects of an attachment-based intervention on the cortisol production of infants and toddlers in foster care. Development and Psychopathology, 20(03), 845–859. https://doi.org/10.1017/S0954579408000400
Drake, K., Belsky, J., & Fearon, R. M. (2014). From early attachment to engagement with learning in school: The role of self-regulation and persistence. Developmental Psychology, 50(5), 1350. https://doi.org/10.1037/a0032779
El-Sheikh, M., Harger, J., & Whitson, S. M. (2001). Exposure to interparental conflict and children’s adjustment and physical health: The moderating role of vagal tone. Child Development, 72(6), 1617–1636. https://doi.org/10.1111/1467-8624.00369
Eluvathingal, T. J., Chugani, H. T., Behen, M. E., Juhász, C., Muzik, O., Maqbool, M., & Makki, M. (2006). Abnormal brain connectivity in children after early severe socioemotional deprivation: A diffusion tensor imaging study. Pediatrics, 117(6), 2093–2100. https://doi.org/10.1542/peds.2005-1727
Escobar, M. J., Rivera-Rei, A., Decety, J., Huepe, D., Cardona, J. F., Canales-Johnson, A., & Manes, F. (2013). Attachment patterns trigger differential neural signature of emotional processing in adolescents. PLoS One, 8(8), e70247. https://doi.org/10.1371/journal.pone.0070247
Fearon, P., Shmueli-Goetz, Y., Viding, E., Fonagy, P., & Plomin, R. (2014). Genetic and environmental influences on adolescent attachment. Journal of Child Psychology and Psychiatry, 55(9), 1033–1041. https://doi.org/10.1111/jcpp.12171
Feldman, R., Gordon, I., Schneiderman, I., Weisman, O., & Zagoory-Sharon, O. (2010). Natural variations in maternal and paternal care are associated with systematic changes in oxytocin following parent–infant contact. Psychoneuroendocrinology, 35(8), 1133–1141. https://doi.org/10.1016/j.psyneuen.2010.01.013
Fisher, P. A., Gunnar, M. R., Dozier, M., Bruce, J., & Pears, K. C. (2006). Effects of therapeutic interventions for foster children on behavioral problems, caregiver attachment, and stress regulatory neural systems. Annals of the New York Academy of Sciences, 1094(1), 215–225. https://doi.org/10.1196/annals.1376.023
Fox, N. A., Nichols, K. E., Henderson, H. A., Rubin, K., Schmidt, L., Hamer, D., & Pine, D. S. (2005). Evidence for a gene-environment interaction in predicting behavioral inhibition in middle childhood. Psychological Science, 16, 921–926. https://doi.org/10.1111/j.1467-9280.2005.01637.x
Fraley, R. C., Roisman, G. I., Booth-LaForce, C., Owen, M. T., & Holland, A. S. (2013). Interpersonal and genetic origins of adult attachment styles: A longitudinal study from infancy to early adulthood. Journal of Personality and Social Psychology, 104(5), 817–838. https://doi.org/10.1037/a0031435
Francis, D. D., & Meaney, M. J. (1999). Maternal care and the development of stress responses. Current Opinion in Neurobiology, 9(1), 128–134. https://doi.org/10.1016/S0959-4388(99)80016-6
Fries, A. B. W., Shirtcliff, E. A., & Pollak, S. D. (2008). Neuroendocrine dysregulation following early social deprivation in children. Developmental Psychobiology, 50(6), 588–599. https://doi.org/10.1002/dev.20319
Gilissen, R., Bakermans-Kranenburg, M. J., van IJzendoorn, M. H., & Linting, M. (2008). Electrodermal reactivity during the Trier Social Stress Test for children: Interaction between the serotonin transporter polymorphism and children’s attachment representation. Developmental Psychobiology, 50(6), 615–625. https://doi.org/10.1002/dev.20314
Gillath, O., Bunge, S. A., Shaver, P. R., Wendelken, C., & Mikulincer, M. (2005). Attachment-style differences in the ability to suppress negative thoughts: Exploring the neural correlates. Neuroimage, 28(4), 835–847. https://doi.org/10.1016/j.neuroimage.2005.06.048
Gotlib, I. H., Ranganath, C., & Rosenfeld, J. P. (1998). Frontal EEG alpha asymmetry, depression and cognitive functioning. Cognition and Emotion, 12, 449–478. https://doi.org/10.1080/026999398379673
Hahn, H. (2015). Federal expenditures on children: What budget policy means for children’s policy. Society for Research in. Child Development Social Policy Report, 29(1), 1–17.
Han, Z. R., Zhang, X., Cui, L., & Yan, J. (2017). The heart of parenting: Parent HR dynamics and negative parenting while resolving conflict with child. Journal of Family Psychology, 31(1), 129–138. https://doi.org/10.1037/fam0000285
Hanson, J. L., Chung, M. K., Avants, B. B., Shirtcliff, E. A., Gee, J. C., Davidson, R. J., & Pollak, S. D. (2010). Early stress is associated with alterations in the orbitofrontal cortex: A tensor-based morphometry investigation of brain structure and behavioral risk. Journal of Neuroscience, 30(22), 7466–7472. https://doi.org/10.1523/JNEUROSCI.0859-10.2010
Hatemi, P. K., & McDermott, R. (2011). Man is by nature a political animal: Evolution, biology, and politics. Chicago, IL: University of Chicago Press.
Hill-Soderlund, A. L., Mills-Koonce, W. R., Propper, C., Calkins, S. D., Granger, D. A., Moore, G. A., ... & Cox, M. J. (2008). Parasympathetic and sympathetic responses to the strange situation in infants and mothers from avoidant and securely attached dyads. Developmental Psychobiology, 50(4), 361–376.
Hood, K., Halpern, C., Greenberg, G., & Lerner, R. (2010). Handbook of developmental science, behavior, and genetics. Malden, MA: Wiley-Blackwell.
Horwitz, B., & Neiderhiser, J. (Eds.). (2015). Gene-environment interplay in interpersonal relationships over the lifespan. New York, NY: Springer.
Hostinar, C. E., & Gunnar, M. R. (2013). The developmental effects of early life stress: An overview of current theoretical frameworks. Current Directions in Psychological Science, 22(5), 400–406.
Huffmeijer, R., Alink, L. R., Tops, M., Grewen, K. M., Light, K. C., Bakermans-Kranenburg, M. J., & van IJzendoorn, M. H. (2013). The impact of oxytocin administration and maternal love withdrawal on event-related potential (ERP) responses to emotional faces with performance feedback. Hormones and Behavior, 63(3), 399–410. https://doi.org/10.1016/j.yhbeh.2012.11.008
Johnson, K., Page, A., Williams, H., Wassemer, E., & Whitehouse, W. (2002). The use of melatonin as an alternative to sedation in uncooperative children undergoing an MRI examination. Clinical Radiology, 57(6), 502–506. https://doi.org/10.1053/crad.2001.0923
Johnstone, T., Kim, M. J., & Whalen, P. J. (2009). Functional magnetic resonance imaging in the affective and social neurosciences. In E. Harmon-Jones & J. Beer (Eds.), Methods in social neuroscience (pp. 313–335). New York, NY: Guilford Press.
Katz, L. F., & Gottman, J. M. (1997). Buffering children from marital conflict and dissolution. Journal of Clinical Child Psychology, 26(2), 157–171. https://doi.org/10.1207/s15374424jccp2602_4
Kim, P., Leckman, J., Mayes, L., Feldman, R., Wang, X., & Swain, J. (2010). The plasticity of human maternal brain: Longitudinal changes in brain anatomy during the early postpartum period. Behavioral Neuroscience, 124(5), 695–700. https://doi.org/10.1037/a0020884
Knopik, A., Neiderhiser, J., DeFries, J., & Plomin, R. (2016). Behavioral genetics (7th ed.). New York, NY: Worth Publishers.
Kochanska, G., & Kim, S. (2013). Early attachment organization with both parents and future behavior problems: From infancy to middle childhood. Child Development, 84(1), 283–296. https://doi.org/10.1111/j.1467-8624.2012.01852.x
Koss, K. J., Cummings, E. M., Davies, P. T., Hetzel, S., & Cicchetti, D. (2016). Harsh parenting and serotonin transporter and BDNF Val66Met polymorphisms as predictors of adolescent depressive symptoms. Journal of Clinical Child and Adolescent Psychology, 1–14. https://doi.org/10.1080/15374416.2016.1220311
Laurent, H., & Powers, S. I. (2007). Emotion regulation in emerging adult couples: Temperament, attachment, and HPA response to conflict. Biological Psychology, 76, 61–71. https://doi.org/10.1016/j.biopsycho.2007.06.002
Lefaucheur, J. P., Antal, A., Ayache, S. S., Benninger, D. H., Brunelin, J., Cogiamanian, F., & Marangolo, P. (2017). Evidence-based guidelines on the therapeutic use of transcranial direct current stimulation (tDCS). Clinical Neurophysiology, 128(1), 56–92.
Lemche, E., Giampietro, V. P., Surguladze, S. A., Amaro, E. J., Andrew, C. M., Williams, S. C., … Simmons, A. (2006). Human attachment security is mediated by the amygdala: Evidence from combined fMRI and psychophysiological measures. Human Brain Mapping, 27(8), 623–635. https://doi.org/10.1002/hbm.20206
Luby, J., Belden, A., Botteron, K., Marrus, N., Harms, M. P., Babb, C., & Barch, D. (2013). The effects of poverty on childhood brain development: The mediating effect of caregiving and stressful life events. JAMA Pediatrics, 167(12), 1135–1142. https://doi.org/10.1001/jamapediatrics.2013.3139
Luijk, M. P., Velders, F. P., Tharner, A., van Ijzendoorn, M. H., Bakermans-Kranenburg, M. J., Jaddoe, V. W., … Tiemeier, H. (2010). FKBP5 and resistant attachment predict cortisol reactivity in infants: Gene–environment interaction. Psychoneuroendocrinology, 35(10), 1454–1461. https://doi.org/10.1016/j.psyneuen.2010.04.012
Matthews, S. G. (2002). Early programming of the hypothalamo–pituitary–adrenal axis. Trends in Endocrinology & Metabolism, 13(9), 373–380. https://doi.org/10.1016/S1043-2760(02)00690-2
McLaughlin, K. A., Fox, N. A., Zeanah, C. H., & Nelson, C. A. (2011). Adverse rearing environments and neural development in children: The development of frontal electroencephalogram asymmetry. Biological Psychiatry, 70(11), 1008–1015. https://doi.org/10.1016/j.biopsych.2011.08.006
Meinlschmidt, G., & Heim, C. (2007). Sensitivity to intranasal oxytocin in adult men with early parental separation. Biological Psychiatry, 61(9), 1109–1111. https://doi.org/10.1016/j.biopsych.2006.09.007
Meyer, A., Hajcak, G., Torpey-Newman, D. C., Kujawa, A., & Klein, D. N. (2015). Enhanced error-related brain activity in children predicts the onset of anxiety disorders between the ages of 6 and 9. Journal of Abnormal Psychology, 124(2), 266–274. https://doi.org/10.1037/abn0000044
Mills-Koonce, W. R., Garrett-Peters, P., Barnett, M., Granger, D. A., Blair, C., & Cox, M. J. (2011). Father contributions to cortisol responses in infancy and toddlerhood. Developmental Psychology, 47(2), 388–395. https://doi.org/10.1037/a0021066
Mills-Koonce, W. R., Propper, C. B., Gariepy, J. L., Blair, C., Garrett-Peters, P., & Cox, M. J. (2007). Bidirectional genetic and environmental influences on mother and child behavior: The family system as the unit of analyses. Development and Psychopathology, 19, 1073–1087. https://doi.org/10.1017/S0954579407000545
Mueller, S. C., Maheu, F. S., Dozier, M., Peloso, E., Mandell, D., Leibenluft, E., & Ernst, M. (2010). Early-life stress is associated with impairment in cognitive control in adolescence: An fMRI study. Neuropsychologia, 48(10), 3037–3044. https://doi.org/10.1016/j.neuropsychologia.2010.06.013
Neumann, I. D. (2008). Brain oxytocin: A key regulator of emotional and social behaviors in both females and males. Journal of Neuroendocrinology, 20(6), 858–865. https://doi.org/10.1111/j.1365-2826.2008.01726.x
Ng, M. Y., & Weisz, J. R. (2016). Annual research review: Building a science of personalized intervention for youth mental health. Journal of Child Psychology and Psychiatry, 57(3), 216–236. https://doi.org/10.1111/jcpp.12470
NICHD Early Child Care Research Network. (2002). Early child care and children’s development prior to school entry: Results from the NICHD Study of Early Child Care. American Educational Research Journal, 39(1), 133–164.
Nomaguchi, K., & Milkie, M. (2017). Sociological perspectives on parenting stress: How social structure and culture shape parental strain and the well-being of parents and children. In K. Deater-Deckard & R. Panneton (Eds.), Parental stress and early child development: Adaptive and maladaptive outcomes (pp. 47–73). New York, NY: Springer. https://doi.org/10.1007/978-3-319-55376-4_3
O’Connor, T. G., & Croft, C. M. (2001). A twin study of attachment in preschool children. Child Development, 72, 1501–1511. https://doi.org/10.1111/1467-8624.00362
Obradović, J., Bush, N. R., Stamperdahl, J., Adler, N. E., & Boyce, W. T. (2010). Biological sensitivity to context: The interactive effects of stress reactivity and family adversity on socioemotional behavior and school readiness. Child Development, 81(1), 270–289. https://doi.org/10.1111/j.1467-8624.2009.01394.x
Olvet, D. M., & Hajcak, G. (2008). The error-related negativity (ERN) and psychopathology: Toward an endophenotype. Clinical Psychology Review, 28(8), 1343–1354. https://doi.org/10.1016/j.cpr.2008.07.003
Oosterman, M., De Schipper, J. C., Fisher, P., Dozier, M., & Schuengel, C. (2010). Autonomic reactivity in relation to attachment and early adversity among foster children. Development and Psychopathology, 22(01), 109–118. https://doi.org/10.1017/S0954579409990290
Organization for Economic Co-operation and Development/OECD. (2017). Key characteristics of parental leave systems (PF2.1). Retrieved May 13, 2017, from https://www.oecd.org/els/soc/PF2_1_Parental_leave_systems.pdf
Parker, S. W., & Nelson, C. A. (2005). The impact of early institutional rearing on the ability to discriminate facial expressions of emotion: An event related potential study. Child Development, 76(1), 54–72. https://doi.org/10.1111/j.1467-8624.2005.00829.x
Patton, D., Costich, J. F., & Lidströmer, N. (2017). Paid parental leave policies and infant mortality rates in OECD countries: Policy implications for the United States. World Medical and Health Policy, 9(1), 6–23. https://doi.org/10.1002/wmh3.214
Peltola, M. J., Yrttiaho, S., Puura, K., Proverbio, A. M., Mononen, N., Lehtimäki, T., & Leppänen, J. M. (2014). Motherhood and oxytocin receptor genetic variation are associated with selective changes in electrocortical responses to infant facial expressions. Emotion, 14(3), 469–477. https://doi.org/10.1037/a0035959
Perry, N. B., Calkins, S. D., Nelson, J. A., Leerkes, E. M., & Marcovitch, S. (2012). Mothers’ responses to children’s negative emotions and child emotion regulation: The moderating role of vagal suppression. Developmental Psychobiology, 54(5), 503–513. https://doi.org/10.1002/dev.20608
Pollak, S. D., & Sinha, P. (2002). Effects of early experience on children’s recognition of facial displays of emotion. Developmental Psychology, 38(5), 784. https://doi.org/10.1037/0012-1649.38.5.784
Porges, S. W. (2003). The polyvagal theory: Phylogenetic contributions to social behavior. Physiology and Behavior, 79(3), 503–513. https://doi.org/10.1016/S0031-9384(03)00156-2
Powers, S. I., Pietromonaco, P. R., Gunlicks, M., & Sayer, A. (2006). Dating couples’ attachment styles and patterns of cortisol reactivity and recovery in response to a relationship conflict. Journal of Personality and Social Psychology, 90(4), 613–628. https://doi.org/10.1037/0022-3514.90.4.613
Propper, C., Moore, G. A., Mills-Koonce, W. R., Halpern, T., Hill-Soderlund, A. L., Calkins, S. D., & Cox, M. (2008). Gene-environment contributions to the development of infant vagal reactivity: The interaction of dopamine and maternal sensitivity. Child Development, 79, 1377–1394. https://doi.org/10.1111/j.1467-8624.2008.01194.x
Proverbio, A. M., Brignone, V., Matarazzo, S., Del Zotto, M., & Zani, A. (2006). Gender and parental status affect the visual cortical response to infant facial expression. Neuropsychologia, 44(14), 2987–2999. https://doi.org/10.1016/j.neuropsychologia.2006.06.015
Purhonen, M., Kilpeläinen-Lees, R., Pääkkönen, A., Yppärilä, H., Lehtonen, J., & Karhu, J. (2001). Effects of maternity on auditory event-related potentials to human sound. Neuroreport, 12(13), 2975–2979.
Rodrigo, M. J., León, I., Quiñones, I., Lage, A., Byrne, S., & Bobes, M. A. (2011). Brain and personality bases of insensitivity to infant cues in neglectful mothers: An event-related potential study. Development and Psychopathology, 23(1), 163–176. https://doi.org/10.1017/S0954579410000714
Rognoni, E., Galati, D., Costa, T., & Crini, M. (2008). Relationship between adult attachment patterns, emotional experience and EEG frontal asymmetry. Personality and Individual Differences, 44(4), 909–920. https://doi.org/10.1016/j.paid.2007.10.021
Roisman, G. I. (2007). The psychophysiology of adult attachment relationships: Autonomic reactivity in marital and premarital interactions. Developmental Psychology, 43(1), 39–53. https://doi.org/10.1037/0012-1649.43.1.39
Roisman, G. I., & Fraley, R. C. (2008). A behavior genetic study of parenting quality, infant attachment security, and their covariation in a nationally representative sample. Developmental Psychology, 44(3), 831–839. https://doi.org/10.1037/0012-1649.44.3.831
Roisman, G. I., Madsen, S. D., Hennighausen, K. H., Sroufe, L. A., & Collins, W. A. (2001). The coherence of dyadic behavior across parent-child and romantic relationships as mediated by the internalized representation of experience. Attachment and Human Development, 3(2), 156–172. https://doi.org/10.1080/14616730126483
Roisman, G. I., Tsai, J. L., & Chiang, K. H. S. (2004). The emotional integration of childhood experience: Physiological, facial expressive, and self-reported emotional response during the adult attachment interview. Developmental Psychology, 40(5), 776–789. https://doi.org/10.1037/00121649.40.5.776
Ross, H. E., & Young, L. J. (2009). Oxytocin and the neural mechanisms regulating social cognition and affiliative behavior. Frontiers in Neuroendocrinology, 30(4), 534–547. https://doi.org/10.1016/j.yfrne.2009.05.004
Sapolsky, R. M., Romero, L. M., & Munck, A. U. (2000). How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocrine Reviews, 21(1), 55–89. https://doi.org/10.1210/edrv.21.1.0389
Schieche, M., & Spangler, G. (2005). Individual differences in biobehavioral organization during problem-solving in toddlers: The influence of maternal behavior, infant–mother attachment, and behavioral inhibition on the attachment-exploration balance. Developmental Psychobiology, 46(4), 293–306. https://doi.org/10.1002/dev.20065
Schmidt, L. A., & Segalowitz, S. J. (Eds.). (2008). Developmental psychophysiology: Theory, systems, and methods. New York, NY: Cambridge University Press.
Shackman, J. E., & Pollak, S. D. (2014). Impact of physical maltreatment on the regulation of negative affect and aggression. Development and Psychopathology, 26(4pt1), 1021–1033. https://doi.org/10.1017/S0954579414000546
Shackman, J. E., Shackman, A. J., & Pollak, S. D. (2007). Physical abuse amplifies attention to threat and increases anxiety in children. Emotion, 7(4), 838–852. https://doi.org/10.1037/15283542.7.4.838
Stalnaker, T. A., Cooch, N. K., & Schoenbaum, G. (2015). What the orbitofrontal cortex does not do. Nature Neuroscience, 18(5), 620–627. https://doi.org/10.1038/nn.3982
Stein, M. B., Schork, N. J., & Gelernter, J. (2008). Gene-by-environment (serotonin transporter and childhood maltreatment) interaction for anxiety sensitivity, an intermediate phenotype for anxiety disorders. Neuropsychopharmacology, 33, 312–319. https://doi.org/10.1038/sj.npp.1301422
Suslow, T., Kugel, H., Rauch, A. V., Dannlowski, U., Bauer, J., Konrad, C., & Ohrmann, P. (2009). Attachment avoidance modulates neural response to masked facial emotion. Human Brain Mapping, 30(11), 3553–3562. https://doi.org/10.1002/hbm.20778
Swain, J. E. (2011). The human parental brain: In vivo neuroimaging. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 35(5), 1242–1254. https://doi.org/10.1016/j.pnpbp.2010.10.017
Thayer, J. F., & Lane, R. D. (2009). Claude Bernard and the heart–brain connection: Further elaboration of a model of neurovisceral integration. Neuroscience and Biobehavioral Reviews, 33(2), 81–88. https://doi.org/10.1016/j.neubiorev.2008.08.004
Tottenham, N., Hare, T. A., Quinn, B. T., McCarry, T. W., Nurse, M., Gilhooly, T., & Thomas, K. M. (2010). Prolonged institutional rearing is associated with atypically large amygdala volume and difficulties in emotion regulation. Developmental Science, 13(1), 46–61. https://doi.org/10.1111/j.1467-7687.2009.00852.x
van Bakel, H. J. A., & Hall, R. A. S. (2018). Parent-child relationships and attachment. In M. R. Sanders & A. Morawska (Eds.), Handbook of parenting and child development across the lifespan (pp. xx–xx). New York: Springer.
Verbeke, W. J., Pozharliev, R., Van Strien, J. W., Belschak, F., & Bagozzi, R. P. (2014). “I am resting but rest less well with you.” The moderating effect of anxious attachment style on alpha power during EEG resting state in a social context. Frontiers in Human Neuroscience, 8, 1–20. https://doi.org/10.3389/fnhum.2014.00486
Voelker, P., Sheese, B. E., Rothbart, M. K., & Posner, M. I. (2009). Variations in catechol-o-methyltransferase gene interact with parenting to influence attention in early development. Neuroscience, 164, 121–130. https://doi.org/10.1016/j.neuroscience.2009.05.059
Vrtička, P., Andersson, F., Grandjean, D., Sander, D., & Vuilleumier, P. (2008). Individual attachment style modulates human amygdala and striatum activation during social appraisal. PLoS One, 3(8), 231–245. https://doi.org/10.1371/journal.pone.0002868
Weisman, O., Zagoory-Sharon, O., & Feldman, R. (2012). Oxytocin administration to parent enhances infant physiological and behavioral readiness for social engagement. Biological Psychiatry, 72(12), 982–989. https://doi.org/10.1016/j.biopsych.2012.06.011
White, L. O., Wu, J., Borelli, J. L., Rutherford, H. J., David, D. H., Kim–Cohen, J., … Crowley, M. J. (2012). Attachment dismissal predicts frontal slow-wave ERPs during rejection by unfamiliar peers. Emotion, 12(4), 690–700. https://doi.org/10.1037/a0026750
Zhang, T. Y., & Meaney, M. J. (2010). Epigenetics and the environmental regulation of the genome and its function. Annual Review of Psychology, 61, 439–466. https://doi.org/10.1146/annualrev.psych.60.110707.163235
Zhang, X., Li, T., & Zhou, X. (2008). Brain responses to facial expressions by adults with different attachment–orientations. Neuroreport, 19(4), 437–441. https://doi.org/10.1097/WNR.0b013e3282f55728
Zilber, A., Goldstein, A., & Mikulincer, M. (2007). Adult attachment orientations and the processing of emotional pictures–ERP correlates. Personality and Individual Differences, 43(7), 1898–1907. https://doi.org/10.1016/j.paid.2007.06.015
Disclosure
The authors declare that they have no disclosure.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Deater-Deckard, K., Chary, M., McCormick, S. (2018). Biological Factors in Parenting and Child Development. In: Sanders, M., Morawska, A. (eds) Handbook of Parenting and Child Development Across the Lifespan . Springer, Cham. https://doi.org/10.1007/978-3-319-94598-9_2
Download citation
DOI: https://doi.org/10.1007/978-3-319-94598-9_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-94597-2
Online ISBN: 978-3-319-94598-9
eBook Packages: Behavioral Science and PsychologyBehavioral Science and Psychology (R0)