
Abstract
Palau, an island archipelago located in the western Pacific warm pool, has over 500 km of outer reefs with steep slopes that host mesophotic coral ecosystems (MCEs). This low-latitude reef system is impacted by strong seasonal variations in oceanic conditions, particularly in the depth range of MCEs. El Niño brings low water temperatures with rapid variation, shallow thermoclines, phytoplankton blooms, and cold-water bleaching to MCEs, while La Niña has warm temperatures, deep thermoclines, and warm-water bleaching. MCE species diversity for Palau is relatively well-known from 20 years of collections using submersibles and mixed-gas diving. We have observed changes in MCEs through typhoon impacts and seasonal water temperature anomalies. The structure of MCEs varies around the main island group of Palau, with the east coast MCEs being dominated by rubble transported downslope due to typhoons with sparse corals, and the west/southwest MCEs are more protected, host a more diverse assemblage and higher cover of sessile invertebrates, and are periodically affected by coral bleaching. In addition, MCEs are also found at the remote oceanic islands and atolls of Palau’s southwest Islands and within the deep lagoons of the main island group. Through our observations and measurements of oceanographic conditions around Palau, we suggest temperature is an important factor limiting coral growth on MCEs, while typhoons also have major impacts on the structure of MCEs through sedimentation and debris moving downslope on the steep outer reefs.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
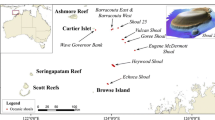
The Republic of Palau (Fig. 16.1) is at the western end of the Caroline Island chain about 1000 km east of the Philippines and 600 km north of the New Guinea Islands. The main island and reef complex, centered on 7.5°N, 134.5°E, covers over 1500 km2 (460 km2 of land and over 1000 km2 of marine area) within a barrier/fringing reef enclosing lagoon areas, the large islands of Babeldaob and Koror, and hundreds of other smaller islands. One normal atoll (Kayangel) and a sunken atoll (Velasco Reef) occur above 8°N. Five oceanic islands and one atoll (Helen Reef) make up the “Southwest (SW) Islands” lying up to 500 km southwest of the main group reaching to 3°N.

(a) Stations around the main Palau Island group where MCEs have been examined. Yellow symbols have temperature monitoring at 57 m depth. Yellow/red symbols have vertical thermograph arrays (2–90 m depth). White symbols have additional temperature and community studies. Inside the white dashed line delimits northern lagoon mesophotic area. (b) Angaur Island with stations shown. (c) Velasco Reef with Ngeruangel station and lagoon mesophotic area inside dashed white line. (d) Bathymetric image of the Palau Islands showing southwest island stations, depths of 3–8 km occur within 100 km of the islands and reefs
Mesophotic coral ecosystems (MCEs) are identified by the presence of light-dependent corals and associated communities (including sponges and algae) from depths of 30–40 m to over 150 m in tropical and subtropical regions (Hinderstein et al. 2010). Palau is within the oceanic waters of the western Pacific warm pool (Longhurst 2010). Its shallow waters are a near ideal environment for coral reefs, existing in isolation from many environmental stressors affecting continental shelf coral reefs. Being located in equatorial latitudes near the world’s areas of highest reef diversity and surrounded by a generally oligotrophic ocean, there is expectation for deep-dwelling reef corals and extensive mesophotic reefs to exist. However, our research has found that this is not necessarily the case in Palau, and we hypothesize that temperature and typhoon effects in Palau might be important limiting factors for MCEs as refuges for coral reef organisms.
In this chapter, we provide the status of MCEs throughout Palau while highlighting selected elements of the geomorphology, ocean environments, and community structure. Reviews of MCEs have included little information from the equatorial Indo-West Pacific (IWP) regions (Lesser et al. 2009; Turner et al. 2017), which includes massive reef tracts in the Philippines, Indonesia, Papua New Guinea, Solomon Islands, and Vanuatu. Colin (2016) described some aspects of the MCEs around Palau’s main island group, and we add to that information herein while including observations from Palau’s remote SW Islands.
1.1 Research History
Over the last 20 years, the Coral Reef Research Foundation (CRRF) and its collaborators have investigated the biological communities, species present, and physical environment of Palau’s MCEs. Extensive collections and observations were made via deep air and mixed-gas diving (400 dives) and research submersibles (60 dives) as part of the marine collections program sponsored by the US National Cancer Institute (NCI). Visits to the SW Islands of Palau were made in 2008, 2014, and 2017. A network of temperature monitoring stations was started in 1999, and Wolanski et al. (2004) and Colin (2009) reported that the mesophotic slope is thermally dynamic with large internal waves. The network has 73 stations with over 155 thermograph instruments at depths from 1 to 90 m. Extensive data on oceanic conditions close to reefs (i.e., depth, temperature, salinity, chlorophyll a fluorescence, and acoustic backscatter) have been obtained with Spray sea gliders since 2009 and by boat-based instruments and other surveys along reef slopes from 2011 by Scripps Institution of Oceanography. Slattery and Lesser (2012) also compared a small series of mesophotic depth stations in the west Pacific (Chuuk and to a lesser extent Palau) with the western Atlantic (Bahamas and Cayman Islands).
2 Environmental Setting
The main reef tract and all separate islands/atolls of Palau have outer slopes quickly reaching oceanic depths (Fig. 16.1). MCEs can be found along a nearly continuous band on the ocean slopes around the main group’s perimeter (330 km), Angaur (15 km), SW Islands (90 km), Kayangel (19 km), and sunken Velasco Reef/Ngeruangel (88 km) (Colin 2009, 2016). This represents a linear distance on outer reefs totaling 542 km; in addition, there are MCEs found in areas of the deep lagoon and at least two offshore pinnacles. The horizontal width of MCEs in Palau is relatively narrow, compared to those found along continental shelves, and depending on the slope can range from as little as 40–50 m on near vertical faces to 100–150 m on more gentle slopes. Colin (2009, 2016) showed slopes for the upper reaches (shallower than 50 m) of the outer reef with the steepest slopes on west and southwest facing reefs. However, the same slopes do not always continue through mesophotic depths. For example, at southwestern Peleliu (station 9 shown in Colin 2016), there is a steep to vertical wall to a depth of 70 m, which then slopes more gently to oceanic depths. The remote SW Islands (Sonsorol, Fana, Pulo Anna, Merir, and Tobi) have narrow shelves around the islands, then drop steeply to oceanic depths, and the single SW atoll, Helen Reef, has a variable outer slope with some areas near vertical.
The oceanic water parameters near the reefs of Palau have been sampled by Spray sea gliders and have given us considerable insight into the variation of temperature, salinity, chlorophyll a fluorescence, and acoustic backscatter to depths of 1000 m (Data courtesy of Dan Rudnick, Scripps Institution of Oceanography). The data indicate that during the El Niño Southern Oscillation (ENSO) neutral periods, the oceanic water column has slight thermal stratification in the upper 100 m, surface salinity of about 34.1 ppt with a maximum of about 34.9 ppt at 100–120 m, and a chlorophyll a maximum (0.2–0.4 μgL−1) at near 100 m depth. However, water column conditions within mesophotic depths shift significantly between El Niño and La Niña periods (see Fig. 9 in Colin 2016). El Niño periods have shallow thermoclines, cooler temperatures at a given depth, increased primary production at mesophotic depths, and more chance of upwelling, while La Niña periods have deep thermoclines, warm temperatures at depth, reduced primary production, and little upwelling. The shifts in oceanic conditions directly impact the entire outer reef, but MCEs are where the parameters are most dynamic.
Through vertical arrays of thermographs (Fig. 16.2a) deployed on the reefs around Palau (Fig. 16.1), we have documented major shifts in water temperatures on MCEs over different time frames. Monthly values show ENSO-related overall shifts in a few months (Fig. 16.2). The strong El Niño periods (2002–2003 and 2015–2016) had weekly mean water temperatures near 20 °C at 57–60 m (Fig. 16.2b). Examining the data at shorter time intervals indicates temperatures often change 6–8 °C in minutes to hours and at their extremes can change as much as 14 °C in 2 min at 57 m depth (Fig. 16.2c). At a single site, massive internal waves can produce changes from 16–18 to 27–28 °C twice in 1 day (Fig. 16.2d). Such waves around Palau at times show a positive correlation with tides for several days, while at other times the relationship is negative or nonexistent (Wolanski et al. 2004). La Niña temperatures at those same depths can reach 29.5–30.0 °C (Fig. 16.2e) without short-term variation (2010 and 2016). The temperature shifts during El Niño, due to internal waves and tides, alongside monthly/yearly El Niño-La Niña changes in mean temperatures, is likely the most important physical factor controlling the lower depth limits of MCEs in Palau (Wolanski et al. 2004; Colin 2009, 2016).

(a) Location of thermographs in a vertical array, Short Drop Off, station17, eastern barrier reef. Bathymetry image courtesy E. Terrill. (b) Weekly mean water temperatures, 1999–early 2017, showing large changes at mesophotic depths. A nominal 30 °C threshold is shown for coral bleaching. (c) Temperature dynamics during 1 day at 57 m depth, southern end Angaur Island (station10). (d) Temperature dynamics at 10, 57 and 90 m depth over 1 day at Ulong Rock, western barrier (station 16) with temporary tidal coupling. (e) Daily mean temperatures for 2010 at station17showing El Niño (blue arrow) and La Niña (red arrow) extremes
The current systems around the main Palau group are complex, a mass of eddying water driven by the interactions of the westerly moving North Equatorial Current (NEC) above 8°N and the North Equatorial Counter Current (NECC) flowing east at 3–6°N through the SW Islands. The NECC is the strongest current system affecting Palau. Its northerly loop is near the longitude of Palau and moves further north seasonally (late spring) to more directly impact the main Palau group, particularly the southern islands of Angaur and Peleliu. During El Niño, its impact may become more extensive, often moving sufficiently north to encompass the entire main island group in its eastward flow. The currents along most MCEs are similar to those found on shallower reefs, but near topographic features, due to turbulence and friction, the speed decreases with increasing depth (e.g., up to a 30% decrease in current speed was recorded from the surface to 45 m at the southern end of Peleliu) with considerable boundary layer turbulence. Deep ridges and promontories along the outer reefs also produce high turbulence with upwelling water visible as surface boils.
Tides in Palau are semidiurnal with amplitudes up to 2.3 m. On the barrier reef, they drive ocean-lagoon exchange through deep channels and across shallow barrier reef tops. The West Channel (station 25, Figs. 16.1 and 16.3) is 80–90 m deep where it bisects the shallow barrier reef and on rising tides carries stratified ocean water into the lagoon. During El Niño, the deeper water ingressed is much cooler with higher nutrients and is mixed in the channel as it is being advected lagoonward (Colin 2009). A series of pH profiles made adjacent to eastern reefs of Palau by Dr. Rebecca Albright in December 2016 indicated that surface waters were at about pH 8.2, decreasing to pH 8.0 by 20 m depth and remaining constant to 75 m depth.

Multi-beam sonar image of station 25 on north side of West Channel, 7°32.560′ N, 134°28.059′ E, showing depths on wall. Bathymetric image courtesy E. Terrill. The reef structure of the channel wall has slumped with collapsed material flowing onto the channel bottom
3 Habitat Description
Numerous stations (Fig. 16.1 and Tables 16.1 and 16.2) were examined in a qualitative assessment of MCE communities. Twenty-six stations include thermographs placed on the reef at 57 m and other depths to record temperature fluctuations. These thermographs are retrieved and deployed at least annually by divers and provide a time to observe, document, and photograph the mesophotic habitat. The MCE slopes on the eastern side of Palau (stations 02, 15, 14, 13, 12, and 11 moving north to south) have large amounts of reef rubble extending downslope with a small number of corals and gorgonians, usually transitioning to finer sediment bottoms below about 45–55 m depth. Western stations (03, 04, 05, 06, and 18) have richer coral communities but still appear to have limited numbers of corals and gorgonians growing upon unconsolidated rubble and talus. The southwestern facing MCEs (stations 23, 16, 07, 08, and 09) have the steepest slopes and some of the highest coral cover of MCEs around the main group. Some stations (26 and 28), which due to their orientation had protection from effects of recent storms, appear to have the highest coral cover of any of Palau’s MCEs (see Fig. 3 in Colin 2016). An offshore pinnacle (station 33), surrounded by deep water, reaches up to 97 m depth with an unusual benthic community, with few corals. Two recent typhoons (Bopha, December 2012, and Haiyan, November 2013) had great impact through mechanical damage on MCEs and shallow reefs. Typhoon Mike (1990) probably had similar effects, but they were not documented.
The outer perimeter of Palau’s reef is irregular (Fig. 16.1) with major projections as well as indentations, which have been suggested as larval retention zones (Hamner and Largier 2012). The mesophotic band of outer slopes has a limited width due to steep profiles, and reef rubble from the shallows is easily transported down such slopes. Steep to vertical reef faces have areas scoured by sediments forming reentrants and sediment chutes (Fig. 16.4). A variety of sub-habitats occur on the outer slope, including buttresses, promontories, caverns (Fig. 16.4b), and caves, often at mesophotic depths and likely increase biodiversity through more diverse environmental conditions. Very steep mesophotic slopes can become unstable with increasing reef growth and eventually fail with reef rock sliding downward creating a talus slope at the base of the rocky face (Fig. 16.4).

(a) Composite photograph showing erosion channel (reentrant) on the vertical reef face at station21, Angaur Island with the base of the vertical escarpment at 65 m. (b) Composite photograph showing downslope sediment transport along the Angaur reef face 20–60 m depth
4 Biodiversity
4.1 Macroalgae
Macroalgae on outer reef MCEs are generally members of the genus Halimeda (Fig. 16.5a), with some coralline algae occurring on rocky surfaces. The northern lagoon and Velasco Reef have algal flats (30–50 m depth) with several genera of calcareous algae (their plates making up most lagoon sediments) and fleshy members of Caulerpa. No seagrasses occur on MCEs in Palau.

Mesophotic corals in the northern Short Drop Off basin (station 28) are in an area largely protected from waves generated by typhoons over the last several decades. (a) Plates of Porites lichen at 45 m depth with abundant Halimeda calcareous algae among them. (b) Deep water Acropora sp. (possibly A. tenella) from 45 m depth
4.2 Anthozoans
Palau has over 330 species of scleractinian corals (DeVantier and Turak 2017). Those occurring on MCEs have not been well characterized, but a number do occur below 45 m depth. Several species of Acropora are found with regularity below 30 m; Acropora tenella appears to be the most common (Fig. 16.5b). The flattened A. pichoni, common at mesophotic depths elsewhere in Micronesia (Chuuk), is uncommon within Palau. Madracis asanoi, the only branching coral common below 60 m depth, is zooxanthellate to 70–90 m depth and becomes azooxanthellate >90 m to ~120 m (see Fig. 4 in Colin 2016). Flattened Leptoseris sp. are found up to 50–70 m deep (see Fig. 6.4 in Baker et al. 2016) and are highly adapted for light capture, but vulnerable to sediment accumulation. Numerous ahermatypic scleractinians occur in the 60–150 m depth range (Veron 2000).
Extensive efforts were made in Palau to obtain a complete sampling of the octocorals for the US National Cancer Institute collections, and all species encountered were identified to genus/species to elucidate the entire fauna (although many species remain undescribed). Fabricius et al. (2007) recorded approximately 150 species of octocorals from Palau, 90 would be considered traditional “soft corals,” while the remainder are largely gorgonians. Of 45 genera of IWP octocorals (Fabricius and Alderslade 2001), 16 are likely to have mesophotic members and 14 of those (roughly 30 species) occur in Palau. Of all octocorals in Palau, 25 appear restricted to depths below 60 m. Some shallow genera also range into the upper mesophotic, including Annella, Melithaea, Dendronephthya, and Umbellulifera. Others were recorded from <10 to 150 m, including Acanthogorgia, Muricella, Eleutherobia, Paraminabea, Chironephthea, Euplexaura, and Carijoa. On many MCE slopes, gorgonians are the most visible benthic organisms, more so than stony corals. Three stations (21, 22, and 31; Fig. 16.1) were dominated by the delicate Stephanogorgia faulkneri from 35 to 60 m depth in numbers sufficient to give the bottom a yellowish hue. Primnoid gorgonians and species of Nicella, Viminella, Keroeides, Paracis, Bebryce, and Parisis first occur at 90–100 m depth and are useful indicators of the lower limit of MCEs. One station (27; Fig. 16.1) had a forest of Umbellulifera sp. on a sandy slope starting at 70 m depth and extending down to at least 90 m (Fig. 16.6a). Additional information on the octocoral and gorgonian faunas of Palau is included in Benayahu et al. (2019) and Sanchez et al. (2019).

(a) Forest of soft corals, Umbellulifera sp. at 75 m depth (scale bar approximate) on the northern coast of Angaur Island (station 27). (b) Vertical wall at 100 m depth (scale bar approximate) with globular lithistid (stony) sponges (1), areas of encrusting sponges (2), encrusting ascidians (3) and slope areas with sediment (4). (c) Overhanging area on vertical wall at 110 m depth (scale bar approximate), with gorgonians (1), solitary coral (2), encrusting sponges (3), and sediment covered bottom (4)
Other cnidarians are less well-known. About 10–15 species of antipatharians occur on MCEs (some of which occur shallower) and some zoanthids (particularly the genus Epizoanthus).
4.3 Sponges
Mesophotic sponges in Palau are relatively well-known and diverse (72 species, 19 orders within 4 Porifera classes). From a total of 450 species recorded by CRRF in Palau (represented by museum specimens), 377 (84%) were found only in the shallow-water zone (0–45 m), 42 (9%) were found only in the mesophotic zone (defined as 45–150 m), and just over 30 (7%) straddled both zones. There are some clear patterns of taxonomic occurrence in MCEs, and these are summarized in Pomponi et al. (2019).
In Palau, vertical reef walls deeper than about 90 m have numerous “lithistid” (or stony) sponges, typified by Microscleroderma herdmani (Fig. 16.6b). Alongside these sponges, areas with vertical or overhanging faces have a spotty, thin film of encrusting sponges, but where sediment can accumulate sponges are lacking (Fig. 16.6c). A surprising geographic difference exists for the barrel sponge Xestospongia testudinaria, which in Palau occurs only on hard bottom slopes at >50 m, while elsewhere in the IWP, it is common in shallow-water reefs. Colin (2016) provides some additional information on Palau MCE sponges.
4.4 Fishes
Although quantitative surveys of mesophotic fish fauna are still underway (through diving stereo-video transects), observations of mesophotic fishes in Palau reveal a diverse range of smaller species, which typically start below 40 m and are indicative of the mesophotic zone. The angelfishes (Centropyge colini and Genicanthus spp.), basslets (Pseudanthias parvirostris and Serranocirrhitus latus), butterflyfishes (Chaetodon burgessi), gobies (Glossogobius colini, Nemateleotris helfrichi, and N. decora), and tilefishes (Hoplolatilus spp.) are some of the more common species. Other rarer species can be found in the lower mesophotic zone, such as the angelfish Centropyge abei (Allen et al. 2006) and the damselfish Chromis abyssus (Pyle et al. 2008). Caverns in the Palau MCE are the only known habitat of the world’s most primitive eel, Protoanguilla palau (Johnson et al. 2011). Some fishes associated with an invertebrate “host” found most often in deep water may mirror the depth distribution of the host. The small seahorse, Hippocampus denise, is an example that occurs at 80 m where its preferred gorgonian host, Muricella sp., occurs in Palau (Lourie and Randall 2003; see Fig. 6c in Colin 2016). Other larger fish species and their ecology are described subsequently in the Ecology section. Quantifying the biodiversity of mesophotic fishes is an area of active study.
4.5 Other Biotic Components
Through 20 years of deep diving and submersible collections for the US National Cancer Institute, there is a reasonable knowledge of the biodiversity of other invertebrates found in the mesophotic zone of Palau. Colin and Arneson (1996) and Colin (2009, 2016) have numerous photographs and descriptions of mesophotic invertebrates from Palau. The large benthic ctenophore, Lyrocteis imperatoris, is found at the lower limit of the mesophotic and appears to move its vertical range up (to as shallow as 90 m) and down (to 200 m) with ENSO temperature shifts. It was collected as shallow as 90 m during El Niño cold-water events (see Fig. 5e in Colin 2016) but was found between 120 and 290 m in 2001 at the end of a weak La Niña. Among mollusks, the large oyster, Empressostrea kostini, occurs in overhanging caverns below 60 m and is an excellent indicator species for the mesophotic (see Fig. 5f in Colin 2016). The gastropod, Cypraea leucodon, is also found in the lower mesophotic zone (and deeper). A variety of shrimps, crabs, and other crustaceans occur on Palau MCEs. One species of MCE squat lobster, Chirostylus sp., has proven to be a parasite cleaner for fishes (Colin 2009). One spectacular sea star, Astrosarkus idipi, first noted in Palau from 67 m, occurs widely throughout the IWP (Mah 2003). Several species of encrusting colonial ascidians have been noted at depth (Fig. 16.6b), but solitary ascidians are fewer in number. Colonies of the sand-dwelling genus Plurella, as well as Eudistoma spp., have been collected and photographed between 60 and 90 m.
5 Ecology
The many MCE stations that have been examined (Fig. 16.1, Supplemental Fig. 16.S1) through diving allow some generalizations despite a lack of quantitative ecological data. The east side stations show much evidence of damage from typhoon Bopha, largely consisting of rubble slopes below about 30–45 m, with little coral growth and scattered gorgonians, and transition to sediment bottoms at 45–60 m depth. Stations on the western side of Palau have more intact reef communities at depth but still are not lush coral environments. Stations on the southwest margin appear to have more corals and less rubble than others. The richest coral communities (Figs. 16.1 and 16.5; Table 16.1; stations 22, 26, and 28) occurred in areas protected from wave action associated with the last three major typhoons hitting Palau (Mike in 1990, Bopha in 2012, and Haiyan in 2013).
A few areas of outer slope MCEs have been examined for lengthy periods. Station 17 (Fig. 16.2) at Short Drop Off has been monitored for coral bleaching (major events in 1998 and 2010), vertical temperature structure (starting 1999), and sediment deposition and benthic algal blooms (1997 and 2015). This is also where the most MCE submersible dives have been made in Palau. On the western barrier reef, Ulong Rock (station 16) has a comparative thermograph array (since 2000), with coral bleaching and photo transect monitoring. As these results are included in Colin (2009), they are not further reported here.
Slope is critical to the types of MCEs that can occur in an area. The multi-beam sonar image (Fig. 16.7) of Croc Head (station 18; Fig. 16.1 and Table 16.1) on the western barrier reef is an excellent example of how MCE geomorphology can change over short distances, affecting the substrate available for benthic communities. The named feature is a promontory which juts out from the slope as a giant buttress with vertical reentrants on its perimeter, undercut caverns, and overhangs. At its outer end is a vertical escarpment from 25 to 60 m depth with the many corals on its upper edge, becoming much fewer on the vertical surfaces (Fig. 16.7b–d). Vertical faces are heavily shaded and lack substrates for corals making them far from ideal habitats. Areas a few 100 m north or south of this structure have slopes (30–40°) with richer MCE coral populations. The general profile of the sloping reef in this area is typical of the west and southwest facing MCE barrier reef slope. Their exposure to the west and northwest ocean exposes them to heavy surf with storm swells from the north and monsoon seas from the SW.

Croc Head MCE. (a) Perspective view multi-beam sonar image of the Croc Head promontory (station 18 in Fig. 16.1a) centered at 7°24.75′ N, 134°20.27′ E. Depths are indicated, and the image has no vertical exaggeration. Bathymetric image courtesy E. Terrill. (b) Upper edge at 25–30 m depth. (c) Vertical area of promontory. (d) Corals at 40 m depth
A few slope MCEs have interesting sub-habitats associated with them. Extensive rubble mounds of tilefishes of the genus Hoplolatilus (Clark et al. 1998) were found in a few areas just below the downslope zone of reef rubble (Fig. 16.8a). Also one area (station 05) was found to have high numbers of solitary corals, Cycloseris sp., at 50 m depth (Fig. 16.8b), a genus found on some lagoon mesophotic reefs. MCE caverns have many environmental characteristics of deeper areas, and several such systems (Blue Holes and Saies Tunnel) have fishes at 15–30 m depths that do not typically occur shallower than 50–60 m on open reef faces.

Unusual deep slope communities, station 5. (a) Hoplolatilus tilefish mounds at 60 m depth. (b) Button corals (Cycloseris sp.) at 50 m depth
North of the main Palau group are two atolls, Kayangel and the sunken Velasco Reef/Ngeruangel. Kayangel Atoll has steep slopes all around with one area (Fig. 16.1; Sta. 21) on its southwest side with high coral cover to 50 m depth. Velasco Reef measures nearly 30 km north to south and has its southern portion (Ngeruangel) at the surface, while the northern (sunken atoll) extremes of the shallow rim are at least 18–23 m deep (Fig. 16.1c). The western side of Ngeruangel (station 26) is different from other MCE stations in Palau, with a rich community of nearly 100% coral cover and high diversity from 30 to 60 m depths on a 30–45° slope. This area was protected from typhoons Mike (which went south of Ngeruangel) and Haiyan (which went almost directly over Ngeruangel), being in the lee of the shallow reef.
5.1 MCEs Found in Lagoon Areas
The main group’s Northern Lagoon (Fig. 16.1a) is connected to oceanic water via three wide passages (Fig. 16.1 indicated as WE, NE, and EE) 15–25 m deep, while the lagoon at Velasco Reef is open across its 15–25 m deep sunken barrier reef. The Northern lagoon is 36–50 m deep with sediment bottoms and has low-relief patch reefs (approaching 250 in number) with high coral cover and diversity (station 30; Fig. 16.9a). Some reach near the surface, while many others are elevated only 3–10 m above the lagoon bottom (Fig. 16.9b) and represent an unrecognized, but significant, portion of reef habitat in Palau. The soft sediments around the patches have grazing halos surrounding them, and time-lapse cameras at 38 m depth have verified the activities of herbivorous acanthurid fishes there. At Velasco Reef, the water is generally clearer than other lagoon areas, often having deep algal flats at 30–50 m depth visible from the surface (Figs. 16.1a, c), with scattered unattached corals, such as Trachyphyllia geoffroyi. Like the Northern Lagoon, it has high cover/diversity patch reefs rising from the lagoon bottom, but the shallowest of these are truncated at 15 m depth, due to the historical subsidence of Velasco Reef (Colin 2009).

(a) Aerial photo mosaic of north lagoon deep bottom (36–40 m depth) with algal flat (AF), low-profile patch reefs (R) and grazing halos (GH) indicated. (b) Detail view of highly diverse coral community at 36 m depth on one of the low-profile patch reefs (Photo credit: Phillip Dustan)
The large lagoon areas close to Babeldaob and the Rock Islands, further south, have no comparable MCEs. The water visibility is more limited (generally 20–25 m as seen from the air), and although there are large numbers of shallow patch reefs, their coral communities decrease with depth, with few observed deeper than 30–35 m. The deeper lagoon is muddy with low circulation, and not a favorable environment for MCEs. An exception to the general lack of southern lagoon MCEs are found west of Ngeruktabel Island at 30–36 m depth (station 29; Supplemental Fig. 16.S2). Protected from wind and waves, the area has a combination of normal lagoon water visibility overlying a persistent nepheloid layer a few meters thick above the mud bottom, resulting in low light and high sediment loading on the bottom. Covering an area of several km2, delicate colonies of Leptoseris gardineri and Seriatopora hystrix exist in an area where it was previously believed no reefs could survive. Discovered during side-scan sonar surveys, their extent has been mapped, and they appear to survive due to extreme adaptations of the individual coral colonies to low light and high sediment loads.
5.2 SW Islands MCEs
The reefs of the SW Islands, including MCEs, are dominated by the strong currents of the NECC, generally moving from west to east through the area. These are of sufficient strength to produce down-current island wakes, a pattern described by Johannes (1981), changing seasonally and with ENSO conditions. Considering their species diversity, the SW Islands are “stepping-stones” by which elements of Coral Triangle fauna reach the main Palau group. But as the islands lack numerous habitats present in the main group, and when combined with considerable distances between islands and generally unfavorable currents, larval transport between the SW Islands and the main group is likely limited.
Investigation of the MCEs in the SW Islands has been limited to a few sites (Table 16.2), but all have narrow reef flats and steep slopes around their perimeters. The five oceanic islands (Fig. 16.1d; Sonsorol, Fana, Pulo Anna, Merir, and Tobi) are similar, built upon mountains rising from deep ocean, with island dimensions of only a few km and narrow reef flats. Helen Reef is an atoll an order of magnitude larger in area than any of the islands, with a deep lagoon and 50 m deep ocean/lagoon channel (but fewer habitats than the main island group). Starting at the surf zone, the outer reef slopes at the islands and atoll become increasingly steep, typically 45° or more at 50–60 m depth. The examined sites where thermographs were deployed were all on the SW quadrant of the islands. Most had low to medium coral coverage, abundant gorgonians with large barrel sponges, Xestospongia testudinaria, common, particularly at Sonsorol. Based on 2 years of data from Tobi, water temperatures of MCEs were about 2 °C higher at any given depth than in the main Palau group, and therefore, stony corals might occur deeper there.
5.3 Phytoplankton Blooms
ENSO shifts not only affect temperature but also ocean productivity which may also influence MCE growth. During the spring 2016 El Niño, the chlorophyll a maximum moved upward to 20–40 m depth, much shallower than its normal depth of >100 m. High light levels combined with presumed enhanced nutrients from the cool, deep water produced phytoplankton blooms (1.4–5.4 μgL−1 chlorophyll a) which reduced water visibility considerably (at times as little as 6 m horizontal). These conditions may negatively affect MCE growth by reducing light available to deeper zooxanthellate corals coinciding with cool <20 °C temperatures. However, as a food source, this productivity could be beneficial for MCEs, especially as common organisms, such as octocorals and sponges, are known to feed on phytoplankton on shallower reefs (Fabricius et al. 1998; Yahel et al. 1998). The impact of this type of productivity and energetics on MCEs is likely significant but requires further study.
5.4 Temperature Limitation on the Outer Reef Slope
The water temperatures on the mesophotic slope vary in both the short (min to hrs to days, Fig. 16.2c, d) and medium term (yearly, Fig. 16.2b). During El Niño periods, weekly mean temperatures at 57 m depth can approach the lower limits of reef growth (Kleypas et al. 1999) and at 90 m lengthy periods have even lower values (15–18 °C). It is at these times when the ocean is cooler with shallow thermoclines and cold-water bleaching likely occurs on MCEs. With short-term variation, temperatures at 57 m can drop into the mid-teens quickly (5–10 °C change has been observed in 5 min or less) and remain there for hours (Fig. 16.2c, d). During La Niña, water temperatures at MCEs are high (>28 °C), similar to shallow reefs, and can induce warm-water bleaching. The two types of bleaching conditions can occur within a 6-month period and is noteworthy, as cold-water bleaching is considered largely a phenomenon of shallow continental shelf waters (Hoegh-Guldberg et al. 2007; Paz-Garcia et al. 2012).
These thermal dynamics beg the question if the observed temperature extremes produce a depauperate mesophotic fauna in Palau as suggested in Wolanski et al. (2004). Would corals occur deeper if it was warmer? In some tropical Pacific areas, reef corals occur to 100–160 m (Baker et al. 2016), but in Palau, they have not been observed deeper than 100 m. Slattery and Lesser (2012), based on two areas on the western barrier reef, indicated sponges dominated the MCE community with decreasing coral coverage with increasing depth, from 60–70% at 30 m to 15–22% at 61 m. Their next sampling depth (76 m) had less than 5% coral cover, while at the deepest sampling depth (91 m), corals barely occurred. This major change in coral cover between the 61 and 76 m stations and the near lack of corals at 91 m tends to support our hypothesis that temperatures at those depths are not favorable to maintenance of reef coral populations. Where there is predominately clear oceanic water, temperatures at 60–90 m depth likely limit reef growth to a greater extent than light penetration. Temperature records from other MCEs in the IWP comparable to those for Palau are needed to address the important questions of temperature and/or light depth limitation of MCEs.
We find the thermal conditions on MCEs surrounding Palau to be more extreme compared to the few locations in the Pacific with any comparable datasets. Hawaiʻi, for example, has warmer conditions at a given depth (seldom falling below 21–22 °C at depths of 80–120 m) than Palau with higher stability (Kahng et al. 2012; Pyle et al. 2016). The lower temperature limits of reef corals have been examined to some extent (Kleypas et al. 1999; Hoegh-Guldberg et al. 2007) and evidence points to this being the factor limiting depth on MCEs in Palau. We think similar temperature limitation will be found in other areas, particularly the low-latitude MCEs that are so poorly known. We recommend the establishment and maintenance of thermograph arrays throughout MCEs to better understand temperature limitations.
5.5 Typhoons
Strong storms (typhoons in particular) are capable of devastating the entire vertical extent of outer reefs in Palau via wave energy breaking loose reef materials and transporting it into both shallow and mesophotic depth water. After temperature extremes, it is suggested that the physical effects of typhoons have the most influence on MCEs. Typhoons suspend vast amounts of fine sediments that can be transported 10s to 100s of km by currents and settle elsewhere, as witnessed through aerial surveys after typhoons in Palau. After typhoon Bopha, the rubble berms produced in shallow water continued to wash out sediments into the water column for 2 years. The death of flattened mesophotic corals through deposition of thick layers of sediment in areas protected from storm wave effects was one of the more surprising outcomes of Bopha (Fig. 16.10).

(a) Flat mesophotic corals at Short Drop Off, 35 days post typhoon Bopha, with thick layers of fine sediment deposited on them, 45 m depth. (b) Same colony 11 months later after sediment had been swept away with skeletal structure starting to disintegrate
The present-day MCE rubble slopes with low populations of corals on the eastern slopes of Palau are almost certainly due to effects from typhoons Bopha (2012) and Haiyan (2013). Bopha’s track south of Angaur generated massive seas in its NE quadrant, which hit the eastern side of Palau from Angaur to Kayangel. Corals and other hard structures above 30–40 m depth were broken loose, forming berms on reef tops or swept downslope to accumulate as rubble at mesophotic depths. Western-facing barrier reefs were sheltered from the wave energy of Bopha but were likely damaged in 1990 by typhoon Mike, evidenced by storm berms visible in 1992 aerial photos. The impacts of typhoon Mike were poorly documented and later ignored in the context of typhoons in Palau (Gouezo et al. 2015). Typhoon Haiyan passed over Kayangel and devastated MCEs on its eastern and northern aspects while largely sparing those on the west and southeast sides. Ngeruangel/Velasco Reef was hard hit on its east side, but the western side of Ngeruangel (station 26; Fig. 16.1), protected from storm seas, still has the most coral-rich MCE observed in Palau. Localized protection from storm seas also allowed station 28, on the east side near Koror, to survive intact. We have started monitoring of MCE communities in Palau through benthic stereo-video transects at permanent sites but have yet to document any obvious recovery.
5.6 Fishes and Fisheries
Unlike relatively sedentary invertebrates, there is a higher potential for depth refuge and replenishment effects for fishes from mesophotic depths to restore shallow reefs. Fishery targeted reef fishes in Palau are predominately caught from the upper 20 m depths by hook and line or spear fishing. Many of these range into the mesophotic zone, including snappers (Lutjanidae), emperor fishes (Lethrinidae), the herbivorous parrotfish (Scarine labrids), and the unicorn and surgeonfishes (Acanthuridae). Many large predatory reef fishes are observed to be more abundant on MCEs, for example, the mangrove snapper (Lutjanus argentimaculatus), black jack (Caranx lugubris), large groupers (Epinephelus malabaricus and E. fuscoguttatus), and the dogtooth tuna (Gymnosarda unicolor).
Some large conspicuous species such as humphead wrasse (Cheilinus undulatus) and reef sharks are attractions to divers in the shallow depths and are regularly seen on MCEs. Although protected in Palau, in the nearby Micronesian island of Guam, C. undulatus have suffered population declines in shallow depths but were recorded in greater abundance on mesophotic reefs (Lindfield et al. 2014). The gray reef shark (Carcharhinus amblyrhynchos) is known to vary its vertical distribution in relation to seasonal water temperatures, with mean depths between 50 and 60 m during La Niña-like conditions when water temperatures are well mixed and warm (29 °C) but move shallower when waters are cooler at those depths (Vianna et al. 2013). Other conspicuous sharks, the silvertip (Carcharhinus albimarginatus), thresher (Alopias pelagicus), and tiger (Galeocerdo cuvier), are more frequently seen on MCEs than shallow reefs.
6 Threats and Conservation Issues
Palau has a relatively low human population compared to its reef area, moderate fishing pressure, low development pressure, good tidal flushing, high diversity of marine species, and upwelling and moderate inputs of terrestrial nutrients to shallow environments. These positive aspects promote a relatively robust recovery from bleaching events and prevent deterioration of reefs in general. Despite these positive aspects, the damage to MCEs from typhoons and coral bleaching do not support the assumption that Palau’s MCEs provide refuge from factors degrading shallow coral reefs. Typhoons have had an outsized effect on MCEs through mechanical destruction by waves and the physical effects of movement of broken reef materials on both shallow and deep reefs. Even in locations that escaped the direct damage from physical forces, sediments produced by typhoon Bopha blanketed corals leading to death in the mesophotic zone (Fig. 16.10). In addition, MCEs were subject to cold-water bleaching during El Niño periods in 2015–2016. Fish populations may at present benefit from some refuge effects, as many of the larger food fishes range widely in depth and deep populations are not caught by spear fishing. However, many hook and line fishers are now targeting deep populations of valuable food fishes as shallow populations are depleted. In the SW Islands of Palau, derelict fishing gear (nets and longlines) are found at all depths, an unsightly, but perhaps not critical threat to benthic communities.
References
Allen GR, Young F, Colin PL (2006) Centropyge abei, a new species of deep-dwelling angelfish (Pomacanthidae) from Sulawesi, Indonesia. Aqua Int J Ichthyol 11:13–18
Baker EK, Puglise KA, Harris PT (2016) Mesophotic coral ecosystems—a lifeboat for coral reefs? United Nations Environment Programme and GRID-Arendal, Nairobi/Arendal
Benayahu Y, Bridge TCL, Colin PL, Liberman R, McFadden C, Pizarro O, Schleyer MH, Shoham E, Reijnen B, Weis M, Tanaka J (2019) Octocorals of the Indo-Pacific. In: Loya Y, Puglise KA, Bridge TCL (eds) Mesophotic coral ecosystems. Springer, New York, pp 709–728
Clark E, Pohle JF, Halstead B (1998) Ecology and behavior of tilefishes, Hoplolatilus starcki, H. fronticinctus and related species (Malacanthidae): non-mound and mound builders. Environ Biol Fish 52:395–417
Colin PL (2009) Marine environments of Palau. Indo-Pacific Press, San Diego
Colin PL (2016) Spotlight on the Palau Island group. In: Baker EK, Puglise KA, Harris PT (eds) Mesophotic coral ecosystems—a lifeboat for coral reefs? United Nations Environment Programme and GRID-Arendal, Nairobi, pp 31–36
Colin PL, Arneson C (1996) Tropical Pacific invertebrates. Coral Reef Press, San Diego
DeVantier L, Turak E (2017) Species richness and relative abundance of reef-building corals in the Indo-West Pacific. Diversity 9(3):25
Fabricius KK, Alderslade PP (2001) Soft corals and sea fans: a comprehensive guide to the tropical shallow water genera of the central-west Pacific, the Indian Ocean and the Red Sea. Australian Institute of Marine Science, Townsville
Fabricius KE, Yahel G, Genin A (1998) In situ depletion of phytoplankton by an azooxanthellate soft coral. Limnol Oceanogr 43:354–356
Fabricius KE, Alderslade P, Williams GC et al (2007) Octocorallia in Palau, Micronesia: effects of biogeography and coastal influences on local and regional biodiversity. In: Kayanne H, Omori M, Fabricius K et al (eds) Coral reefs of Palau. Palau Int Coral Reef Cent, Koror, pp 79–92
Gouezo M, Golbuu Y, van Woesik R et al (2015) Impact of two sequential super typhoons on coral reef communities in Palau. Mar Ecol Prog Ser 540:73–85
Hamner WM, Largier JL (2012) Oceanography of the planktonic stages of aggregation spawning reef fishes. In: Sadovy de Mitcheson Y, Colin PL (eds) Reef fish spawning aggregations: biology, research and management, Fish & Fisheries Series, vol 35. Springer, Dordrecht, pp 159–190
Hinderstein LM, Marr JCA, Martinez FA et al (2010) Theme section on “Mesophotic coral ecosystems: characterization, ecology, and management.” Coral Reefs 29(2):247–251
Hoegh-Guldberg O, Mumby PJ, Hooten AJ et al (2007) Coral reefs under rapid climate change and ocean acidification. Science 318:1737–1742
Johannes RE (1981) Words of the lagoon: fishing and marine lore in the Palau district of Micronesia. University of California Press, Los Angeles
Johnson GD, Ida H, Sakaue J et al (2011) A ‘living fossil’ eel (Anguilliformes: Protanguillidae, fam. nov.) from an undersea cave in Palau. Proc R Soc B Biol Sci 279:934–943
Kahng SE, Wagner D, Lantz C et al (2012) Temperature related depth limits of warm-water corals. Proc 12th Int Coral Reef Symp C 9
Kleypas JA, McManus JW, Meñez LAB (1999) Environmental limits to coral reef development: where do we draw the line? Am Zool 39:146–159
Lesser MP, Slattery M, Leichter JJ (2009) Ecology of mesophotic coral reefs. J Exp Mar Biol Ecol 375:1–8
Lindfield SJ, McIlwain JL, Harvey ES (2014) Depth refuge and the impacts of SCUBA spearfishing on coral reef fishes. PLoS ONE 9(3):e92628
Longhurst AR (2010) Ecological geography of the sea. Academic, Burlington
Lourie S, Randall JE (2003) A new pygmy seahorse Hippocampus denise (Teleostei: Syngnathidae) from the Indo-Pacific. Zool Stud 42:284–291
Mah CL (2003) Astrosarkus idipi, a new Indo-Pacific genus and species of Oreasteridae (Valvatida: Asteroidea) displaying extreme endoskeletal reduction. Bull Mar Sci 73:685–698
Paz-García DA, Balart EF, García-de-Léon FJ (2012) Cold water bleaching of Pocillopora in the Gulf of California. Proc 12th Int Coral Reef Symp 9:10
Pomponi SA, Diaz MC, Van Soest RWM et al (2019) Sponges. In: Loya Y, Puglise KA, Bridge TCL (eds) Mesophotic coral ecosystems. Springer, New York, pp 563–588
Pyle RL, Earle JL, Greene BD (2008) Five new species of the damselfish genus Chromis (Perciformes: Labroidei: Pomacentridae) from deep coral reefs in the tropical western Pacific. Zootaxa 1671:3–31
Pyle RL, Boland R, Bolick H et al (2016) A comprehensive investigation of mesophotic coral ecosystems in the Hawaiian Archipelago. PeerJ 4:e2475
Sánchez JA, Dueñas LF, Rowley SJ et al (2019) Gorgonian corals. In: Loya Y, Puglise KA, Bridge TCL (eds) Mesophotic coral ecosystems. Springer, New York, pp 729–747
Slattery M, Lesser MP (2012) Mesophotic coral reefs: a global model of community structure and function. Proc 12th Intl Coral Reef Symp 1:9–13
Turner JA, Babcock RC, Hovey R et al (2017) Deep thinking: a systematic review of mesophotic coral ecosystems. ICES J Mar Sci 74(9):2309–2320
Veron JEN (2000) Coral of the world, vol 3. Australian Institute of Marine Science, Townsville
Vianna GMS, Meekan MG, Meeuwig JJ et al (2013) Environmental influences on patterns of vertical movement and site fidelity of grey reef sharks (Carcharhinus amblyrhynchos) at aggregation sites. PLoS ONE 8:e60331
Wolanski E, Colin P, Naithani J et al (2004) Large amplitude, leaky, island-generated, internal waves around Palau, Micronesia. Estuar Coast Shelf S60:705–716
Yahel G, Post AF, Fabricius K et al (1998) Phytoplankton distribution and grazing near coral reefs. Limnol Oceanogr 43:551–563
Acknowledgments
We thank the staff of CRRF, particularly L. Bell, M. Mesubed, E. Basilius, and V. Jaiteh for their assistance. The Palau National and Koror State Governments provided permits for this work. Aspects were supported by funding from the US Office of Naval Research, and deep-water collections were funded by the US National Cancer Institute. We thank scientists Drs. Eric Terrill, Dan Rudnick, Jennifer MacKinnon, and Shaun Johnston of Scripps Institution of Oceanography for allowing us to describe their unpublished results and use sonar images. Dr. Rebecca Albright of the California Academy of Sciences provided unpublished data on water quality. We thank Drs. Bruce Carlson and William Hamner, and two anonymous reviewers for comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
1 Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Colin, P.L., Lindfield, S.J. (2019). Palau. In: Loya, Y., Puglise, K., Bridge, T. (eds) Mesophotic Coral Ecosystems. Coral Reefs of the World, vol 12. Springer, Cham. https://doi.org/10.1007/978-3-319-92735-0_16
Download citation
DOI: https://doi.org/10.1007/978-3-319-92735-0_16
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-92734-3
Online ISBN: 978-3-319-92735-0
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)