
Abstract
Halophyte farming seems to be a promising alternative to conventional agriculture under marginal environments, since it does not compete with dwindling fresh water and land resources. For this purpose halophytic species need to be domesticated to serve as a “crop” plant. Field evaluation of six Salicornia bigelovii Torr. populations (LA, GA, MI, FL2, SP, FL1) was conducted in 2012–2013 in the United Arab Emirates applying two irrigation treatments: brackish groundwater (20 dSm−1) and seawater (55 dSm−1) to examine their growth performance taking into account 24 plant characteristics, biomass, seed yield parameters, and growth stages attributes. The experimental results indicated that S. bigelovii populations were highly variable for the majority of traits. Irrigation with seawater negatively influenced the agronomic performance of S. bigelovii populations compared to groundwater. SP, MI, and GA populations received higher values for almost all characteristics observed, as compared to LA, FL1, and FL2 populations especially under groundwater treatment. Seawater extended the duration of growth cycle for all Salicornia populations compared to groundwater irrigation. The outcomes clearly indicated that the salinity level had an impact on S. bigelovii populations’ performance and yield potential. It is suggested that screening should be evaluated under both optimum and full-strength saline water to optimize biomass and seed production. The existing genotypes could be further improved through breeding taking into account spike characteristics and days to flowering as revealed by path analysis. Combining suitable germplasm with proper agronomic practices, there is a big potential to develop this halophytic species plantations for economically viable production systems in hot and dry regions.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
14.1 Introduction
With growing demand for food and energy production, new resources are sought to meet population needs. The exploration of marginal environments to increase the agricultural production has raised the interest of scientists and other stakeholders. Marginal areas are mainly located in semiarid and arid climatic zones, where degraded soils and low water quality cannot support agriculture production. Due to the aforementioned limiting factors, the feasibility of seawater agriculture is explored in desert environments (Glenn et al. 1998, 2013). The use of crops that can withstand high saline conditions is extremely important in such a context, especially for marginal lands where no other crops can grow, conserving good quality water for human consumption in parallel.
Developing salt-tolerant crops constitutes a very important strategy to improve yield on salt-affected lands. However, improving salt tolerance in traditional crops through breeding is a laborious process at both plant and cellular level, because of the complexity and multigenic nature of this trait (Ashraf et al. 2008; Cassaniti et al. 2013; Fageria et al. 2012; Flowers 2004). Halophytes flourish in highly saline environments, where conventional crops cannot grow (Shabala 2013). Halophyte cultivation seems to be a promising solution, since these plants already possess the trait of salt tolerance (Rozema and Flowers 2008). Nonetheless, wild halophytic populations need to be collected and bred, so that they can be transformed into productive and profitable crops (Brown et al. 2014a; Ruan et al. 2010).
Salicornia bigelovii Torr. (dwarf glasswort), among other candidate monocots and dicotyledonous halophytic species, has received more attention because of its multifold uses and high salt tolerance (Zerai et al. 2010; Lu et al. 2010; Jaradat and Shahid 2012; Shahid et al. 2013; Glenn et al. 2013; Ventura et al. 2011a, b; Ventura and Sagi 2013; Brown et al. 2014a). S. bigelovii belongs to Chenopodiaceae, a popular family for its members tolerant-to-salinity. It is a C3 plant and an annual, succulent euhalophytic species that grows very fast. S. bigelovii is a cross-pollinating plant that can also self-pollinate to a lower extent. The fresh tips can be used as salad or pickled directed for human consumption. The biomass can be mixed with other forages and used as livestock feed (Khan and Ansari 2008; Masters et al. 2007). S. bigelovii seeds have good qualitative and quantitative oil characteristics (≈30%) and low salt content (<3%), attributes that strengthen its importance as oilseed crop with high potential for biofuel production (Brown et al. 2014b; Cybulska et al. 2014a, b; Glenn et al. 1998). The seedcake has high protein content (≈45%) and could be also used as animal feed (Glenn et al. 1991). S. bigelovii as a halophytic species of good commercial value has been proposed as a component to be incorporated in farming systems that integrate crops and fish, since the aquaculture effluents constitute a valuable source of nutrients and water for irrigation that could be recycled efficiently (Hamed et al. 2014; Bailis and Yu 2012; Hodges 2004; Brown et al. 1999).
In order to acquire highly productive cultivars with good commercial potential, wild germplasm needs to be domesticated, and selective breeding programs must be undertaken. Zerai et al. (2010), after comparing S. bigelovii lines produced in two breeding programs with wild germplasm in greenhouse trials irrigated with brackish water (16 dS−1), concluded that sufficient genetic diversity existed among wild accessions and cultivars to support a crop improvement program using wild S. bigelovii populations. Several S. bigelovii breeding populations and lines respectively were evaluated for 12 plant traits under high salinity irrigation in field nurseries (Shahid et al. 2013; Jaradat and Shahid 2012).
Literature does not show evaluation studies of S. bigelovii germplasm that have been conducted (a) under field conditions in hot and dry climates using different salinity treatments and (b) on a large scale of traits. In the current study, we examined the differentiation in the growth performance of six S. bigelovii wild collected populations under two salinity treatments, taking into account a wide variety of agronomic parameters. Secondly, we highlighted those characteristics that are linked with high biomass and seed production under local conditions. Finally, we tested the hypothesis whether the two salinity levels should be used as selection criteria for classification of S. bigelovii germplasm which is vital for decision-making on the suitable breeding processes further adopted.
14.2 Materials and Methods
The field experiments were conducted at the facilities of International Center for Biosaline Agriculture (ICBA) (25°05′45.98″ N; 55°23′25.29 E; elevation 30 m) in Dubai, United Arab Emirates. The annual average minimum and maximum temperature were 20.9 °C and 34.8 °C, respectively. The annual mean rainfall was 38.6 mm with average daily evapotranspiration of 5.3 mm. The soil at ICBA experimental station is sandy in texture (sand 98%, silt 1%, and clay 1%). The soil salinity was 768 ppm, and the organic matter content was less than 0.5 (Shahid et al. 2009).
14.2.1 Plant Material and Field Trials
Plant material (seeds) of six wild collections of S. bigelovii populations FL1, FL2, GA, MI, LA, and SP were kindly provided by Masdar Institute of Science and Technology (MI) research team.
Field was prepared adding organic compost at 40 t/ha. The sowing date for S. bigelovii seeds was 25th of November 2012. The sowing density was 3 g/m2, and S. bigelovii seeds were lightly covered with soil. Agryl sheets were placed on top of the seeded plots in order to avoid seed losses (by wind and birds) and provide suitable microenvironment for germination. Seeds/seedlings were irrigated with brackish groundwater (GW) (20 dS m−1) and seawater (SW) (55 dS m−1). Plot size was 2 × 2 = 4 m2, and the planting distance was 25 × 25 cm. The seedlings/plants were irrigated through drip irrigation system with the aforementioned salinity treatments adjusted to the water requirement plus 30% leaching to maintain root zone salinity to a desired level. The total water quantity used for irrigation was 468 m3 and was applied from sowing until harvest.
The randomized complete block design was followed with three replications for each treatment. The salinity level of GW and SW irrigation was regularly monitored. Soil salinity was checked by taking composite soil sample from various experimental plots representing different treatments from 0 to 25 cm which corresponded to the vital root zone. Soil samples were dried, and soil extract was obtained to determine the electrical conductivity. At the end of the trial, soil samples were again collected and analyzed for salinity and pH. The levels of salinity were maintained throughout the cultivating season with very small fluctuations (Table 14.1). In particular, pH and EC values ranged from 7.5 to 7.8 and from 8.6 to 25.1 dS/m for GW treatment and from 7.8 to 8.1 and from 28.9 to 52.3 dS/m for SW, respectively, for both samplings.
14.2.2 Data Collection
When S. bigelovii plants reached the optimum vegetative stage (stage before heading), two lines from each plot were harvested (surface area = 1 m2), and fresh (green) shoot weight/m2 was estimated. At final harvest, measurements were taken from 10 plants per experimental plot, and 17 morphological characteristics were recorded. All the characters studied with their abbreviations and units are presented in Table 14.2. SWTPL was calculated as the average of the ten tagged plant values. SWTSQ was measured for each treatment plot (total area = 4 m2 − 1 m2 corresponded to green biomass harvest = 3 m2). HI was estimated as the ratio of seed yield to total above ground crop dry weight. Additionally, S. bigelovii plants were monitored for six developmental stages throughout the whole cultivating season. DTOFL were calculated when 5% of the plants were at the anthesis stage. DUFL was the period from when flowering started until all plants bloomed. DTOSP were the total number of days immediately after the end of flowering. DUSP was the period between completion of anthesis and onset of seed maturity. S. bigelovii plants mature gradually. DUM initiated when 10% of the plants were mature (light brown color) and was completed when the plants were totally mature (dark brown color). GC refers to the whole cultivating period of S. bigelovii plants.
14.2.3 Data Analysis
Data collected during the growing season were tested for homogeneity of variances before conducting univariate or multivariate statistical analysis. Descriptive statistics (mean and standard error) were estimated for all the data measured. Analysis of variance (ANOVA) was applied to evaluate the variability for the abovementioned growth attributes among S. bigelovii populations and between salinity treatments. Least significant difference (LSD) post hoc tests were implemented to detect differences among the average values. Hierarchical cluster analysis was performed in order to identify the relationships among the growth traits in S. bigelovii populations grouping in two different salinities. Ward’s method was used to classify the data. Correlation analysis was also computed using Pearson’s test. Pathway analysis was conducted, in order to examine the direct and indirect associations of various growth characteristics on SWTSQ. Path analysis differs from simple correlations in the sense that it points out the causes and their relative importance, whereas the latter simply measures the mutual association ignoring the causation. All statistical analyses were performed with JMP statistical software version 8.0 (SAS Institute, Inc., Cary, NC, USA).
14.3 Results
14.3.1 Impact of Salinity Treatments on Growth Parameters of S. bigelovii Populations
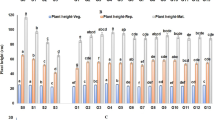
Generally, plant characteristics differentiated to a greater extent when irrigated with GW compared to SW for all S. bigelovii populations (Table 14.3). SPK and BR were the characteristics most negatively affected by SW compared to GW treatment for all halophytic genotypes, and the differences ranged from −95% to −629% and from −52% to −124%, respectively. PH was also reduced after SW in contrast with GW application for all populations; however, the differences that varied between −3% and −105% were not all statistically significant. FIRSTSPKD, SPL, SPWT, and RL were either positively, negatively, or neutrally influenced by the higher salinity treatment among S. bigelovii populations. In contrast, an increase in SPW was observed for most S. bigelovii populations irrigated with SW except from SP and GA genotypes, which they did not demonstrate statistically significant differences between salinity treatments for the specific characteristic. Under GW irrigation, MI, SP, and GA plants were characterized as taller (59.8, 60.6, and 50.9 cm, respectively), having the highest number of BR (36.9, 37.0 and 38.8) and SPK (311.5, 333.8 and 373.0), increased RL (18.6, 19.6 and 17.9 cm), the FIRSTSPKD was developed in a higher level (11.2, 13.0 and 10.9 cm), and their spikes were longer (109.4, 115.0 and 85.3 mm) and heavier (495, 533 and 254 mg) compared to LA, FL1, and FL2 plants. The former group of S. bigelovii populations maintained their preponderance for most of the characteristics even after SW application.
DSHWT, DRWT, and ASH measurements were significantly reduced for all S. bigelovii genotypes after SW compared to GW irrigation, and the differences ranged from −30% to −498%, 31% to −532%, and − 9% to −35%, respectively (Table 14.4). FRSHWT decreased dramatically for MI (−476%), SP (−407%), and GA (−618%) plants and to a lower extent for FL2 (−78%) and LA (−8%) populations after high salinity treatment. In contrast, green biomass slightly increased for FL1 (6%). Both SWTPL and SWTSQ were severely affected by SW treatment for MI (−191% and −250%, respectively), SP (−133% and −388%), and GA (−165% and −201%) genotypes. In contrast, high salinity provoked an increase in SWTSQ for LA (38%), FL1 (52%), and FL2 (21%) genotypes, although it was not statistically significant. Indifferent results were obtained for LA and FL1 populations for SWTPL between the two treatments; however, a slight decrease was observed for FL2 (−9%). The SSG is related to the size of the seed; hence the higher the number, the smaller the size of the seeds. Generally, more, but smaller and lighter seeds were produced after GW compared to SW treatments for all populations, except for MI which SSG remained almost similar in both irrigation treatments. A number of seeds per spike of MI and GA plants were significantly decreased after SW irrigation. For the rest of S. bigelovii populations, no statistical significant differences were spotted between the treatments, although an increase was observed in genotypes LA and FL2 for this spike parameter. None statistical significant difference was observed for SWTSP for all S. bigelovii genotypes, although a considerable decrease was observed for MI and GA populations under high salinity treatment.
Finally, HI increased because of higher salinity impact for all S. bigelovii genotypes, and the increase varied between 13% and 59%. However, HI values were statistically significant only for MI, SP, and GA populations. High HI values do not necessarily reflect higher seed yield but significantly less biomass due to high salinity levels, which finally increased the ratio [seed weight/(seed weight + biomass)]. Overall, MI and SP populations excelled for almost all seed and biomass yield measurements in both salinity treatments. GA also received comparable satisfying values for most yield characteristics under GW treatment; however, it did not maintain its good performance at higher salinity level. FL1 and FL2 populations irrigated with SW outperformed for several yield parameters such as FRSHWT (3.5 and 3.2 kg/m2) even surpassing MI and SP at the same treatment.
Early flowering was observed for MI, SP, and GA populations and ranged from 134.7 (MI) to 142.3 (SP) and from 124.7 (MI) to 151.0 (SP) days for GW and SW treatments, respectively, whereas LA, FL1, and FL2 populations exhibited late flowering after several weeks in both applications (Table 14.5). SW irrigation extended DUFL for all S. bigelovii genotypes from 12% to 25%, except from LA (−30%). After anthesis stage, SW compared to GW extended the duration of growth stages of S. bigelovii plants. Ten to twenty two more days were needed for all S. bigelovii genotypes to reach seed production stage. DUSP was also prolonged from 4 to 42 days for LA, MI, FL1, and FL2, whereas the opposite, although not significant effect, was observed for SP and GA populations at higher salinity application. DUM was also significantly extended after SW treatment for all S. bigelovii populations, apart from GA which salinity effect was neutralized. Finally, S. bigelovii plants completed their GC 17 to 42 days later after irrigation with SW compared to GW. MI was characterized by the shortest GC at high salinity rate (287.7 days).
14.3.2 Grouping of S. bigelovii Populations Based on GW and SW Treatments
Hierarchical clustering revealed three main clusters based on plant growth (A, B, C) (Fig. 14.1). MI, SP, and GA populations excelled for the majority of the plant attributes under GW irrigation and were grouped in Cluster A. Cluster B contained all S. bigelovii populations treated with SW and was divided into two sub-clusters (D and E). Cluster D encompassed LA, FL1, FL2, and GA, and this cluster was placed next to cluster C, where the same populations congregated after irrigation with GW. MI and SP populations treated with SW assembled in Cluster E. Cluster analysis showed that the halophytic populations were separated to the two salinity treatments based on their overall response.

Clustering for six S. bigelovii populations (LA, GA, MI, FL2, SP, FL1) irrigated with brackish groundwater (GW) and seawater (SW) for all parameters studied
14.3.3 Correlations and Interrelationships Among the Characters Studied Comparing the Two Salinities Treatments
Out of 276 possible pairwise correlation coefficients for all 24 plant growth parameters, 176 (64%) were significant for GW and only 57 (21%) for SW treatment (Table 14.6). 94% and 88% of significant correlations for GW and SW treatment, respectively, were higher than 0.500 implying the strong association (negative or positive) among the traits. Positive associations were noted between SWTSQ and the rest of the traits except for SSG (−0.590) and DTOFL (−0.865) for S. bigelovii plants irrigated with GW. Negative correlation between SWTSQ and DTOFL (−0.524) was also observed for SW treatment. The same associations were observed between FRSHWT and other growth parameters for GW application. However, FRSHWT was negatively correlated only with harvest index for S. bigelovii populations treated with SW. ASH was not correlated with any other character for both treatments.
Pathway analysis was used to partition the observed correlation coefficient into direct and indirect effects of several components on SWTSQ, in order to provide clear picture of the interrelationships among traits. Subramanian and Subramanian (1994) determined the values for direct and indirect effects path analysis as follows: value of >1 (very high), 0.3–0.99 (high), 0.2–0.29 (moderate), 0.1–0.19 (low), and <0.1 (negligible).
SWTSP, SSSP, and SPL were characterized by the highest direct effects 0.604, 0.467, and −0.302, respectively, whereas moderate effects were observed for SPK, DUSP, and HI (−0.208, 0.250 and 0.226, respectively) for S. bigelovii populations irrigated with GW. On the other hand, SW application significantly increased the values of direct effects for several parameters such as BR (0.302), SSSP (1.537), SWTSP (−1.516), DTOFL (−0.752), and DUM (0.472). Lower direct effects were spotted for the rest of the parameters.
SSSP was characterized by high and positive direct and total effects (correlations) for both water treatments, and this meant that the correlation explained the true relationship and this trait should be considered one of the characters for S. bigelovii improvement in a breeding program. The same observation was made for SWTSP only for GW application. In contrast, for SW treatment, a very high and negative direct effect was noted, but a positive and moderate correlation was observed. When a high positive or negative correlation is observed, but the direct effects are negative and positive respectively or negligible, then the indirect effects are the cause of correlation. Such findings were observed for BR, SPK, SPL, DTOFL, DUM, and HI for S. bigelovii populations irrigated with GW and for SWTSP, DUSP, and HI for S. bigelovii populations irrigated with SW. SSSP and SWTSP were the parameters with the highest indirect effect in all cases followed by DTOFL and SPL. SPK and SPL were characterized by low and negligible direct and correlation effects in SW treatment.
14.4 Discussion
14.4.1 Differentiation of Growth Performance of S. bigelovii Populations Comparing GW and SW Treatments
Screening and evaluation of six S. bigelovii populations irrigated with GW and SW, under field conditions, revealed large differences among themselves and between the treatments. The high variation in the characteristics measured indicated that these are highly variable populations, presumably due to the outcrossing nature of the breeding system (Zerai et al. 2010; Martinez-Garcia 2010; Jaradat and Shahid 2012). Significant intraspecific variation in salt tolerance has been observed for various halophytic grasses such as Spartina patens, Spartina alterniflora (Hester et al. 2001), and Distichlis spicata (Aschenbach 2006). Under elevated salinities, the study of variation within and between species can give insight into physiological adaptations that result in morphological changes (Hester et al. 2001).
MI, SP, and GA were characterized by higher values for the majority of characteristics, compared to LA, FL1, and FL2 populations for both treatments. They were also characterized by early-flowering habit, a desirable characteristic especially for hot and arid environments as observed in Eritrea region by Zerai et al. (2010). All wild accessions, breeding populations, and lines of S. bigelovii differentiated for most of the parameters measured, when irrigated with water which salinity was 17 dS m−1 (Zerai et al. 2010) and >40 dS m−1 (Jaradat and Shahid 2012). The maximum seed yield was obtained by MI (1.27 t/ha), SP (0.85 t/ha), and GA (0.82 t/ha) populations treated with GW. However, seed production was less than 1.39–2.46 t/ha for Salicornia plants irrigated with SW as reported in Glenn et al. (1999). The seed production of LA, FL1, and FL2 populations was slightly higher after SW irrigation, although it remained very low and did not exceed 0.2 t/ha. However, attention should be drawn to the fact that FL1 population produced two times more seed after SW (0.2 t/ha) compared to GW (0.1 t/ha) treatment. The mechanism that is involved to overcome increased salinity levels and produce more seeds could be further investigated. MI, SP, and GA populations also received the maximum values for FRSHWT and DSHWT after irrigation with GW. Green biomass values were the highest ever reported in the literature ranging from 120 to 140 t/ha. It should be mentioned that the values extrapolated from 1 m2 to a hectare and that the experimental plots were small in size (4 m2) and well-managed and sufficient water was provided to each plant through drip irrigation system. DSHWT and DRWT were significantly reduced after SW treatment compared to root length values. It has been observed that shoot growth was more sensitive than root growth to elevated salinities (Munns and Tester 2008; Läuchli and Grattan 2007). This is due to the fast root recovery compared to shoots (Munns 2002). Differences in shoot growth could result from increased production of nodes or increased size of internodes, as a result of increased cell division and/or elongation, when plants are grown in optimal salinity (Ayala and O’Leary 1995). SPL (>11 cm) measured for the current populations exceeded the ones observed for Salicornia accessions from other studies (Jaradat and Shahid 2012; Shahid et al. 2013). Glenn et al. (1997) reported an average SPL of 3–5 cm as compared to 10–15 cm spike length of native Salicornia. The increase in SPW, which was monitored after SW irrigation, constituted a physiological response often observed in plants grown under high saline conditions, which is attributed to Na and Cl accumulation (Karabourniotis et al. 2012).
SW compared to GW extended the duration of the growth stages after flowering and the whole growth cycle as a consequence. A common feature of plant response to salinity is the inhibition of growth and productivity. Slower growth is an adaptive characteristic for plant survival under stress, since it allows plants to rely on multiple resources (e.g., building blocks and energy) to combat the stress (Zhu 2001). According to Ventura et al. (2011b), cultivation based on full-strength seawater (≈48 dS/m) had no effect on the general flowering pattern of the different Salicornieae types.
The assemblage of MI, SP, and GA populations through cluster analysis confirmed their supremacy under GW and SW irrigation and could be characterized as high-yielding populations, compared to LA, FL1, and FL2 which could be denoted as low-yielding populations. Although GA plants were characterized by slower growth compared to MI and SP plants after SW treatment, they still performed better compared to LA, FL1, and FL2 populations.
14.4.2 Traits Targeted for Improving High-Yielding S. bigelovii Populations
Correlation analyses showed that most of the studied parameters were highly correlated among themselves, implying that salt tolerance is a multigenic trait which is reflected on morphological, physiological, and biochemical aspects (Munns and Tester 2008; Flowers 2004). In addition, yield is a very complex feature resulting from the process of plant growth. For the halophytic species, S. bigelovii positive and significant associations were observed between yield (seed or biomass) and some other traits especially for GW irrigation. This observation is favorable for breeding, since selection for one trait may produce correlated response for improvement of other traits, which are positively associated with it (Pandey et al. 2012; Yadav et al. 2013; Subramanian and Subramanian 1994). In particular, spike characteristics maintained their high association among themselves even after SW treatment. Similar results were obtained by Jaradat and Shahid (2012), who observed that, under SW treatment, seed morphometric characteristics were highly correlated (>0.5) among themselves and with seed yield.
Pathway analysis revealed that selection for SPWT and SSSP may lead to improved germplasm for seed yield (Table 14.7). SPWT is highly correlated with SPL which is also correlated with the SSSP it encloses (Shahid et al. 2013). DTOFL was another desirable trait revealed from the current study that should be selected as criterion for further genotypes’ improvement, in order to compress the GC so that summer months could be avoided (Zerai et al. 2010). Although HI was not as high as reported from Zerai et al. (2010) and Glenn et al. (1991), it can also be improved through reproductive traits. Desirable traits that also need to be taken into account in a breeding program would be shorter plants, in order to avoid lodging, with considerable BR and SPK that should be placed in a higher level so that plucking could be facilitated (Zerai et al. 2010). Path analysis implemented for eleven S. bigelovii genotypes irrigated with SW (55 dS m−1) validated the findings of the current study since similar spike parameters and growth stages were highlighted for selection to improve seed production (Lyra et al. 2016).
14.4.3 Water Salinity Levels as Selection Criteria for Screening S. bigelovii Germplasm
The threshold of the water salinity and suitable selection criteria used to evaluate ecotypes within a species for salinity tolerance are of highly importance for breeding purposes and have been discussed in several studies (Ruan et al. 2010; Lee et al. 2005, 2004a, b; Ashraf 2004; Zeng et al. 2002). Lee et al. (2004a) emphasized that evaluation of seashore paspalum ecotypes, at water salinity <20 dS m−1, was not sufficient for assessing salinity tolerance. Previous studies only focused on one salinity regime assessment for screening S. bigelovii lines and accessions. In the current study, S. bigelovii plants growth was significantly stimulated by salinity of EC = 20 dS m−1, compared to salinity of EC = 55 dS m−1. Similar findings were obtained for the same halophytic species (Ayala and O’Leary 1995) and for the perennial halophyte Suaeda fruticosa (Khan et al. 2000). Salinity around 20 dS m−1 constitutes the optimum concentration for S. bigelovii and other halophytes growth (Panta et al. 2014; Katschnig et al. 2013; Flowers and Colmer 2008; Ayala and O’Leary 1995). In addition, the highest salinity level to which S. bigelovii would be expected to be exposed for screening ecotypes within a species is needed, in order to examine the highest degree of salinity tolerance (Ruan et al. 2010). Besides, SW is the only available water resource to irrigate S. bigelovii or any other halophytic species along coastal deserts, so the plant material that is going to be used must be characterized by high growth rates under such elevated saline conditions (Brown et al. 2014a). As a result, GW and SW used in the present study could be considered as optimum and maximum salinity levels, respectively, for S. bigelovii growth. High-yielding populations of MI and SP outperformed under GW irrigation, and they maintained their preponderance even at SW salinity , although population growth was significantly hampered. In addition, high salinity affected GA population to a greater extent, which performance resembled the growth of low-yielding populations. Under high saline conditions, FL1 population produced twice more seed compared to low salinity level. As a result, decisions on plant breeding tactics based only on screening results conducted under optimum salinity (≈20 dS m−1) would not guarantee a consistent result. Since considerable variability existed among S. bigelovii populations for both salinity levels, the two salt concentrations could be used as evaluation criteria for the specific halophytic species.
14.5 Conclusions
Seawater agriculture could be more feasible as long as suitable germplasm of halophytic species of high yield potential is identified (Brown et al. 2014a). The current study was executed under field conditions in a desert environment, and germplasm with desired characteristics was distinguished. MI, SP, and GA received higher values for almost all characteristics observed, compared to LA, FL1, and FL2 populations especially under GW treatment. The first three populations constitute valuable germplasm that can go through the breeding process, in order to expand in similar hot and arid areas like UAE. However, it is worthwhile to explore the physiological functions involved in the last three populations, since they overcame the hindrance of increased salinity and led to double seed yield. The substantial genetic-based variation that existed within S. bigelovii wild populations facilitates the effective selection of diverse genotypes. S. bigelovii seems to have the potential to become a successful crop since it is tested in a climate similar to the one of its natural occurrence (Hamed et al. 2014). Several production trials have been conducted with S. bigelovii in Eritrea, Saudi Arabia, Kuwait, and Mexico, in order to compare the yields to other conventional crops and examine the adaptation to local climatic conditions (Brown et al. 2014a; Abdal 2009).
The study demonstrated that by targeting few traits such as spike characteristics and DTOFL, considerable improvement could be achieved in a short period. Additionally, mass selection through collection of bigger seeds could also contribute in accelerating its domestication (Brown et al. 2014a; Zerai et al. 2010). Other desirable traits that need to be taken into consideration for S. bigelovii cultivars are short growth cycle to avoid adverse climatic conditions, low saponin levels in seeds and vegetable tips, more uniform flowering and ripening, nematode resistance, better retention of seeds on spikes, and shorter plants that can resist lodging (Zerai et al. 2010). Finally, inbreeding would also help to receive purified lines of S. bigelovii.
References
Abdal MS (2009) Salicornia production in Kuwait. J World Appl Sci 6:1033–1038
Aschenbach TA (2006) Variation in growth rates under saline conditions of Pascopyrum smithii (Western Wheatgrass) and Distichlis spicata (Inland Saltgrass) from different source populations in Kansas and Nebraska: implications for the restoration of salt-affected plant communities. Restor Ecol 14:21–27
Ashraf M (2004) Some important physiological selection criteria for salt tolerance in plants. Flora 199:361–376
Ashraf M, Athar HR, Harris PJC, Kwon TR (2008) Some prospective strategies for improving crop salt tolerance. Adv Agron 97:45–110
Ayala F, O’Leary JW (1995) Growth and physiology of Salicornia bigelovii Torr. at suboptimal salinity. Int J Plant Sci 156:197–205
Bailis R, Yu E (2012) Environmental and social implications of integrated seawater agriculture systems producing Salicornia bigelovii for biofuel. Biofuels 3:555–574
Brown JJ, Glenn EP, Fitzsimmons KM, Smith SE (1999) Halophytes for the treatment of saline aquaculture effluent. Aquaculture 75:255–268
Brown JJ, Glenn EP, Smith SE (2014a) Feasibility of halophyte domestication for high-salinity agriculture. In: Khan MA et al (eds) Sabkha ecosystems: volume IV: cash crop halophyte and biodiversity conservation. Springer, Dordrecht, pp 73–80
Brown JJ, Cybulska I, Chaturvedi T, Thomsen MH (2014b) Halophytes for the production of liquid biofuels. In: Khan MA et al (eds) Sabkha ecosystems: volume IV: cash crop halophyte and biodiversity conservation. Springer, Dordrecht, pp 67–72
Cassaniti C, Romano D, Hop MECM, Flowers TJ (2013) Growing floricultural crops with brackish water. Environ Exp Bot 92:165–175
Cybulska I, Chaturvedi T, Alassali A, Brudecki GP, Brown JJ, Sgouridis S, Thomsen MH (2014a) Characterization of the chemical composition of the halophyte Salicornia bigelovii under cultivation. Energy Fuel 28:3873–3883
Cybulska I, Chaturvedi T, Brudecki GP, Kádár Z, Meyer AS, Baldwin RM, Thomsen MH (2014b) Chemical characterization and hydrothermal pretreatment of Salicornia bigelovii straw for enhanced enzymatic hydrolysis and bioethanol potential. Bioresour Technol 153:165–172
Fageria NK, Stone LF, dos Santos AB (2012) Breeding for salinity tolerance. In: Fageria NK, Stone LF, Santos ABD (eds) Plant breeding for abiotic stress tolerance. Springer, Berlin, pp 103–122
Flowers TJ (2004) Improving crop salt tolerance. J Exp Bot 55:307–319
Flowers TJ, Colmer TD (2008) Salinity tolerance in halophytes. New Phytol 179:945–963
Glenn EP, O’Leary JW, Watson MC, Thompson TL, Kuehl RO (1991) Salicornia bigelovii Torr.: an oilseed halophyte for seawater irrigation. Science 251:1065–1067
Glenn E, Miyamoto S, Moore D, Brown JJ, Thompson TL, Brown P (1997) Water requirements for cultivating Salicornia bigelovii Torr. With sea water on sand in a coastal desert environment. J Arid Environ 36:711–730
Glenn E, Brown JJ, O’Leary J (1998) Irrigating crops with sea water. Sci Am 279:76–81
Glenn E, Brown JJ, Blumwald E (1999) Salt tolerance and crop potential of halophytes. Crit Rev Plant Sci 18:227–255
Glenn EP, Anday T, Chaturvedi R, Martinez-Garcia R, Pearlstein S, Soliz D, Nelson SG, Felger RS (2013) Three halophytes for saline-water agriculture: an oilseed, a forage and a grain crop. Environ Exp Bot 92:110–121
Hamed KB, Magné C, Abdelly C (2014) From halophyte research to halophytes farming. In: Khan MA et al (eds) Sabkha ecosystems: volume IV: cash crop halophyte and biodiversity conservation. Springer, Dordrecht, pp 135–142
Hester MW, Mendelssohn IA, McKee KL (2001) Species and population variation to salinity stress in Panicum hemitomon, Spartina patens, and Spartina alterniflora: morphological and physiological constraints. Environ Exp Bot 46:277–297
Hodges CN (2004) The seawater foundation: lessons from Africa about the world. Seawater Foundation, Tucson, AZ
Jaradat AA, Shahid M (2012) The dwarf saltwort (Salicornia bigelovii Torr.): evaluation of breeding populations. Int Sch Res Netw. https://doi.org/10.5402/2012/151537
Karabourniotis GA, Liakopoulos G, Nikolopoulos D (2012) Plant physiology under stress conditions. Chapter 3 salt tolerance mechanisms. Embryo, Athens, pp 57–72
Katschnig D, Broekman R, Rozema J (2013) Salt tolerance in the halophyte Salicornia dolichostachya Moss: growth, morphology and physiology. Environ Exp Bot 92:32–42
Khan MA, Ansari R (2008) Potential use of halophytes with emphasis on fodder production in coastal areas of Pakistan. In: Abdelly C, Ozturk M, Ashraf M, Grignon C (eds) Biosaline agriculture and high salinity tolerance. Birkhäuser, Basel, pp 157–162
Khan MA, Ungar IA, Showalter AM (2000) The effect of salinity on the growth, water status, and ion content of a leaf succulent perennial halophyte, Suaeda fruticosa (L.) Forssk. J Arid Environ 45:73–84
Läuchli A, Grattan SR (2007) Plant growth and development under salinity stress. In: Jenks MA, Hasegawa PM, Jain SM (eds) Advances in molecular breeding toward drought and salt tolerant crops. Springer, Dordrecht, pp 1–32
Lee G, Duncan RR, Carrow RN (2004a) Salinity tolerance of seashore paspalum ecotypes: shoot growth responses and criteria. HortSci 39:1138–1142
Lee G, Carrow RN, Duncan RR (2004b) Salinity tolerance of selected seashore paspalums and bermudagrasses: root and verdure responses and criteria. HortSci 39:1143–1147
Lee G, Carrow RN, Duncan RR (2005) Criteria for assessing salinity tolerance of the halophytic turfgrass seashore paspalum. Crop Sci 45:251–258
Lu D, Zhang M, Wang S, Cai J, Zhou X, Zhu C (2010) Nutritional characterization and changes in quality of Salicornia bigelovii Torr. During storage. Food Sci Technol 43:519–524
Lyra D, Ismail S, Butt KURB, Brown J (2016) Evaluating the growth performance of eleven Salicornia bigelovii populations under full strength seawater irrigation using multivariate analyses. Aust J Crop Sci 10:1429–1441
Martinez-Garcia R (2010) Physiological studies of the halophyte Salicornia bigelovii: a potential food and biofuel crop for integrated aquaculture-agriculture systems. PhD Thesis, University of Arizona
Masters DG, Benes SE, Norman HC (2007) Biosaline agriculture for forage and livestock production. Agric Ecosyst Environ 119:234–248
Munns R (2002) Comparative physiology of salt and water stress. Plant Cell Environ 25:239–250
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59:651–681
Pandey VR, Singh PK, Verma OP, Pandey P (2012) Inter-relationship and path coefficient estimation in rice under salt stress environment. Int J Agric Res 7:169–184
Panta S, Flowers T, Lane P, Doyle R, Haros G, Shabala S (2014) Halophyte agriculture: success stories. Environ Exp Bot 107:71–83
Rozema J, Flowers T (2008) Crops for a salinized world. Science 322:1478–1480
Ruan CJ, da Silva JAT, Mopper S, Qin P, Lutts S (2010) Halophyte improvement for a salinized world. Crit Rev Plant Sci 29:329–359
Shabala S (2013) Learning from halophytes: physiological basis and strategies to improve abiotic stress tolerance in crops. Ann Bot 112:1209–1221
Shahid SA, Dakheel AJ, Mufti KA, Shabbir G (2009) Automated in-situ soil salinity logging in irrigated agriculture. Eur J Sci Res 26:288–297
Shahid M, Jaradat AA, Rao NK (2013) Use of marginal water for Salicornia bigelovii Torr. Planting in the United Arab Emirates. In: Shahid SA, Abdelfattah MA, Taha FK (eds) Developments in soil salinity assessment and reclamation. Springer, Dordrecht, pp 451–462
Subramanian S, Subramanian M (1994) Correlation studies and path coefficient analysis in sesame (Sesamum indicum L.). J Agron Crop Sci 173:241–248
Ventura Y, Sagi M (2013) Halophyte crop cultivation: the case for Salicornia and Sarcocornia. Environ Exp Bot 92:144–153
Ventura Y, Wuddineh WA, Myrzabayeva M, Alikulov Z, Khozin-Goldberg I, Shpigel M, Sagi M (2011a) Effect of seawater concentration on the productivity and nutritional value of annual Salicornia and perennial Sarcocornia halophytes as leafy vegetable crops. Sci Hortic 128:189–196
Ventura Y, Wuddineh WA, Shpigel M, Samocha TM, Klim BC, Cohen S, Shemer Z, Santos R, Sagi M (2011b) Effects of day length on flowering and yield production of Salicornia and Sarcocornia species. Sci Hortic 130:510–516
Yadav A, Kumar A, Roy C, Jaiswal JP (2013) Correlation and path-coefficient analysis for grain yield, its components and quality traits in bread wheat. Pantnagar. J Res 11:337–340
Zeng L, Shannon MC, Grieve CM (2002) Evaluation of salt tolerance in rice genotypes by multiple agronomic parameters. Euphytica 127:235–245
Zerai DB, Glenn EP, Chatervedi R, Lu Z, Mamood AN, Nelson SG, Ray DT (2010) Potential for the improvement of Salicornia bigelovii through selective breeding. Ecol Eng 36:730–739
Zhu JK (2001) Plant salt tolerance. Trends Plant Sci 6:66–71
Acknowledgments
The research was part of a joint project between ICBA and MI. MI provided ICBA with S. bigelovii seeds to run the experiments conducted at ICBA’s experimental station. MI in conjunction with Boeing, Etihad Airways, UOP Honeywell, and Safran established a joint-funded consortium (SBRC, Sustainable Bioenergy Research Consortium) in 2011 and are looking into exploring the potential of growing S. bigelovii with seawater in the United Arab Emirates mostly for biofuel production but also seeking into other uses. The authors, finally, would like to acknowledge Mr. Khalil Rahman for the experiments design and planning and Mr. Balagurusamy Santhanakrishnan for his technical assistance in the field work.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Lyra, D.A., Ismail, S., Brown, J.J. (2020). Crop Potential of Six Salicornia bigelovii Populations Under Two Salinity Water Treatments Cultivated in a Desert Environment: A Field Study. In: Hirich, A., Choukr-Allah, R., Ragab, R. (eds) Emerging Research in Alternative Crops. Environment & Policy, vol 58. Springer, Cham. https://doi.org/10.1007/978-3-319-90472-6_14
Download citation
DOI: https://doi.org/10.1007/978-3-319-90472-6_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-90471-9
Online ISBN: 978-3-319-90472-6
eBook Packages: EngineeringEngineering (R0)