
Abstract
This multiscalar study explores Holocene environmental changes across the Sahara, within the eastern Sahara , and along the Nile in northern Sudan . The Early Holocene saw increased moisture across most parts of northern Africa after c. 10,000 BC, with peak humid conditions 7800–7000 BC. A short but significant dry interval after c. 7000 BC was followed by wetter conditions around 6000 BC, and then a gradual aridification from 5000 BC. The latter dry phase has continued until present times. The exceptional environments near the Nile are known to have seen impressive variations as climate oscillations and flora left traces in the palaeobotanical record. Multidisciplinary archaeological studies in this area—including analysis of plant macroremains—have focused on the transitions from hunting-fishing-gathering (Khartoum Variant) to pastoralism (Abkan) and later to agro-pastoralism (Pre-Kerma). The palynological data from four Sai Island sites (8-B-10C, 8-B-76, 8-B-81, and 8-B-10A) and the nearby mainland site of Amara West (2-R-66) provide new perspectives on local environmental shifts during this time of profound economic and social change. Despite poor pollen preservation, the high number of samples enables comparisons that show both diachronic changes and synchronic variation. Since the earliest phases, pollen spectra reflect mixed flora from various habitats and some seasonal variability. During the Early Holocene and the initial part of the Middle Holocene , dramatic floods on Sai’s east side and seasonal desiccation on Sai’s west side together created an ecological mosaic that exposed people to several different habitat types within a short distance. These included swamps and marshes, wooded savannas, grasslands and desert savanna, providing access to plants used for food, medicine, and other purposes. Documenting localized patterns of vegetation variation and change can lay important groundwork for explaining changes in subsistence and social organization.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
As Saharan climates fluctuated dramatically through the past 12,000 years, Northern Sudan has seen important innovations in subsistence and social life: food producing economies emerged, use of the Nile Valley intensified, storage and mortuary practices transformed, and eventually powerful polities arose (Bonnet 1992, 2006; Reinold 2001, 2006; Honegger 2004a, 2007; Garcea and Hildebrand 2009; Garcea 2011–2012; Garcea et al. 2013, 2016a, b; Honegger and Williams 2015; Hildebrand and Schilling 2016). Interactions between humans and local habitats were crucial to processes of social and economic change; these have been clarified by geoarchaeological, zooarchaeological and macro-botanical investigations of datasets that directly reflect human activities (Chaix and Grant 1993; Lario et al. 1997; Salvatori et al. 2011). Palynological research complements these efforts by establishing ecological baselines using datasets that reflect natural regional conditions rather than specific human actions. Although poor preservation has hindered palynological studies in North Sudan (Watrin et al. 2009; Florenzano et al. 2016), archaeological deposits represent an exciting data source for examining diachronic environmental changes, and evaluating possible natural and anthropic causes in arid lands (Horowitz 1992; Mercuri 2008).
Throughout Holocene changes in climate, and their profound effects on Saharan environments and plant migrations, the Nile has always offered unusual sets of resources (Maley 2010). When arid conditions reduced inland vegetation to desert types, waterplaces were refugia, maintaining wet habitats that sustained plants, animals and people (Garcea et al. 2013; Mercuri et al. 2015). Cultural trajectories were often concurrent with climate oscillations; these changes eventually affected expansions or contractions of past human populations influencing mobility and land use in the Sahara (Kuper and Kröpelin 2006, Kröpelin et al. 2008; Mercuri et al. 2011; deMenocal and Tierney 2012; Garcea et al. 2013; Manning and Timpson 2014; Macklin and Lewin 2015; Maley and Vernet 2015). However, not all arid phases triggered crisis or abandonment; some created opportunities for new subsistence strategies and social networks (Zerboni et al. 2016).
The Northern Sudanese Nile is a fascinating setting to study human/climate interaction during dramatic environmental changes (flood to drought) and human responses (periodic environmental reassessment driven by unstable ecosystems and land degradation). We report a synthesis of pollen analyses from four sites (8-B-10C, 8-B-10A: Florenzano et al. 2018; 8-B-76, 8-B-81: Florenzano et al. 2016) on Sai Island , in Northern Sudan (Fig. 1). In this synthesis we discuss the results of interdisciplinary investigations on human adaptations and subsistence strategies under Holocene climate changes in light of paleoclimate and paleoethnobotanical information obtained from the literature on this region.

Environmental Setting: Modern Climate, Vegetation, and Land Use
Today Central and Northern Sudan encompass an incredible range of environments, from semi-arid areas near Khartoum to arid full desert farther north, crossed by a ‘green corridor’ of the riparian Nile . Near Khartoum, the rainy season (July to September) delivers an average 125 mm per year. In far northern Sudan at Abri near Sai Island , average annual rainfall is 2 mm (Climate Data n.d.). At this latitude (~20.5° N), areas away from the river have numerous sand dunes west of the Nile , or shift to a rocky substrate east of the Nile . Today, many areas >200 m from the river have scant vegetation, and some animals rely on the Nile and its floodplains for critical resources and water.
Modern Vegetation of the Sahara and Sahel
White (1986:38) discerns three main vegetation areas in northern and central Sudan . The country’s northern and northwestern extremities areas have limited vegetation of the Saharan type (XVII; see White 1986:223). A northeast-to-southwest belt from Sudan ’s coast to the Jebel Marra supports Sahelian vegetation of Khordofan type, e.g. Sahel wooded grassland and Sahel semi-desert grasslands (XVI; see White 1986:206–207). These semi-desert shrublands have Acacia wooded grassland, perennial and annual grasses, such as Panicum turgidum Forsk., and other herbs such as Indigofera hochstetteri Baker (Abuswar 2007; Mustafa 2007). Farther south, Sudanian woodland and grasslands have higher moisture requirements (III; see White 1986:105–107 for descriptions). This last environment lies outside of our study area today, but elements of it may have pushed north during wetter periods in the past.
The Nile and Its Exceptional Environments
On its route to the Mediterranean, the Nile carries runoff from several source areas with distinct seasonality of rainfall, flow gradient, and sediment transport. Bimodal (spring/autumn) rainfall in equatorial eastern Africa (Burundi, Rwanda, Uganda, western Kenya) feeds the Kagera-White Nile ; much of this water is held in the Sudd marshes of South Sudan . Long summer and short spring rains in southwest Ethiopia highlands (>2000 mm rainfall per year drained by the Baro, Gilo, and Akobo Rivers), as well as precipitation in southeastern Sudan together feed the Sobat River, which joins the White Nile at the northern edge of the Sudd. The low gradient of the White Nile and its diverse seasonal inputs result in a perennial, slow flow bearing sediment of small particle size. Strong, highly seasonal summer rains in the western Ethiopian highlands feed the Didessa, Abbay, Dinder, and Rahad (uniting in the Blue Nile ), and the northwestern Ethiopian highlands and Ethio-Eritrean borderlands (Takezze/Atbara). Due to their seasonality, steeper gradients, and passage through arid areas more vulnerable to erosion, these rivers carry higher volumes of sediment than the White Nile , and coarser particles (Garzanti et al. 2015). Today, the largest overall contributor to the main Nile is the Blue Nile . However, tributary flows have varied in the past, when the eastern Saharan drainage systems of Wadi Howar and Wadi el Melik were also active.
In Northern and Central Sudan , riparian and arable lands are richer in plant species than other habitats (Abdelkreem and Ibrahim 2016). Today, land use is practically limited to riverine environments or areas that can be irrigated. Along the Nile , farming activity is well-developed thanks to irrigation technologies: the historic shaduf (wherein a pole mounted in an upright frame and balanced with a counterweight is used to raise water from the Nile channel to field level) and modern pumps. In places where the Nile follows a deep channel, well-watered riverine habitats are separated from many mainland and island surfaces. Elsewhere, more shallow riverine margins sustain swamp communities rich in sedges and grasses (Ali 2006; Madani et al. 2015). Islands sometimes have at least one shallow margin that is periodically inundated, receiving both flood moisture and enriched waterborne silts; thus, they may sustain higher plant species diversity.
Modern conditions in northern Sudan offer insights into environmental parameters that affected people throughout the Holocene . These include strong distinctions between riparian and interior habitats, latitudinal differences in rain-fed vegetation away from the river that responded to amplified or weakened monsoons, changes in channel depth and flood level, and fluctuating contributions from different Nile tributaries that would capture rainfall at different times of the year. Together these suggest a complex mosaic of factors which interacted to produce highly variable environments along the Nile and nearby hinterlands over the last 12,000 years.
Paleoclimate and Paleovegetation
Northern Africa has witnessed several dramatic climatic changes. During the Last Glacial Maximum (24,000–17,000 BCFootnote 1; Tierney and deMenocal 2013) and for many millennia thereafter, extremely dry conditions made the Sahara hardly habitable (see Cancellieri et al. 2016) and substantially reduced the flow of the Nile , although several Late Pleistocene sites were recorded in Lower Nubia and farther north (Schild and Wendorf 2010). Beginning c. 10,000 BC, the Early Holocene saw increased moisture across northern Africa, with peak humid conditions 7800–7000 BC. A short but significant dry interval began just after 6500 BC, and affected many parts of the Sahara. Wetter conditions resumed 6000 BC, but a gradual aridification began after 5000 BC and has continued until present times.
Although enhanced moisture during the Early and Middle Holocene has been termed the ‘African Humid Period’ (deMenocal et al. 2000), the exact manifestations of this climate event varied across Africa (Costa et al. 2014; Williams et al. 2015a). This is especially true north of the equator, where different circulation systems govern different regions. For example, during periods of exceptional monsoon strength, the ITCZ extends farther north during the northern hemisphere summer, delivering more moisture to the Horn of Africa, the northern Sahel, and the southern Sahara. There is complex interplay between tropical moisture-bearing systems from the Indian Ocean (SE Africa) versus Atlantic/Gulf of Guinea (SW Africa), and between these and higher-latitude circulation systems (Gasse 2000, 2002).
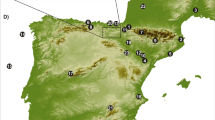
Global climatic events and their local manifestations exerted a profound impact on environments and resources, altering the relative utility of riverine versus hinterland contexts for human livelihoods. To contextualize our new data from the Middle Nile , we review current perspectives on climatic and environmental changes at three geographic scales (Fig. 1). Saharan denotes the region now encompassing the Sahara desert, which for much of the Holocene sustained Sahelian and Sudanic environments. The Eastern Sahara encompasses the plains between rain-capturing mountains farther west (Tibesti, Ennedi, and Jebel Marra) and the Nile ; it includes Saharan tributaries of the Nile that were active during moist periods (e.g. Wadi Howar) and their watersheds, as well as closed lake basins. Nile Corridor denotes areas adjoining the Nile in its current active channels and tributaries.
Regional Context: Environmental Changes Across the Sahara
Onset of Wet Conditions
The onset of the moist conditions occurred in two episodes within the Sahara and Sahel: water levels rose somewhat 13,000–12,500 BC, retreated during the Younger Dryas (10,900–9600 BC), and then resurged 9500–9000 BC (Gasse 2000). High water tables fostered the development of lakes, ponds and wetland areas in many parts of the Sahara by c. 9000 BC, with watercourses linking vast areas (Fig. 2; see also Drake et al. 2011). Many parts of the Sahara show multiple lines of geomorphological, climatic and environmental evidence for increased moisture (Fontes et al. 1985; Haynes et al. 1989; Schulz 1991; Cremaschi and di Lernia 1998, 1999, 2001; Cremaschi and Trombino 1998; Cremaschi 2002; Gasse 2000; Kröpelin et al. 2008; Mercuri 2008; Cole et al. 2009; Cremaschi and Zerboni 2009; Cremaschi et al. 2010, 2014; Zerboni et al. 2011; Giraudi et al. 2013). In the southern Saharan massifs, floods from upland areas created permanent lakes in lower depressions c. 9000–8500 BC (Gasse 2000).

Major geographical features and paleoenvironmental study localities in the Sahara and the Nile . 1–Atlas Mountains. 2–Jebel Gharbi. 3–Tadrart Acacus. 4–Erg Uan Kasa. 5–Edeyen of Murzuq. 6–Ahaggar Mountains. 7–Tibesti Mountains. 8–Ennedi Mountains. 9–Jebel Marra. 10–Northwestern Ethiopian Highlands. 11–Lake Tana. 12–Lake Victoria (probable early Holocene extent). 13–(Paleo) Lake Chad (probable early Holocene extent). Approximate catchment boundaries and lake maxima from Drake et al. 2011.
Vegetation responded: tropical taxa probably followed riverine corridors north into greening parts of the Sahara, reaching their northernmost extent by c. 8500 BC. Because species responded individually to climate oscillations, and riverine and hinterland areas existed side by side, pollen spectra record a mosaic of plant taxa that does not have any analogues in modern environments (Watrin et al. 2009). Many animals—including fish and molluscs—also recolonized the Sahara, but some species requiring deep water (hippo, Nile perch) or other particular conditions stayed south of the Ahaggar (Drake et al. 2011).
Peak Moisture Delivery
By c. 7800 BC the InterTropical Convergence Zone (ITCZ) shifted as far north as 24°N in some parts of Africa, bringing tropical rainfall to much of the Sahara (Gasse and Roberts 2004). The exact timing and duration of this “hydrological optimum” varied geographically (Gasse 2000; Lézine et al. 2011; see also Costa et al. 2014; Williams et al. 2015a). In the Libyan Sahara’s Tadrart Acacus, tufa development shows that heavy monsoon rains saturated the Tanezzuft Formation and created freshwater springs 7600–5600 BC (U-series and 14C dates; Cremaschi et al. 2010). Lacustrine sedimentation indicates rainfall sufficient to sustain open bodies of water in the erg Uan Kasa 7200–6100 BC and the Edeyen of Murzuq 7500–5600 BC (Cremaschi 1998; Cremaschi and Zerboni 2009). The southern Sahara saw highest lake levels 7300–6600 BC (Gasse 2000), but the Chad Basin had a short regression c. 7500–7000 BC (Maley 1981, 2010).
Pioneer tropical taxa that pushed north during the earliest Holocene continued growing in Saharan areas during this time of peak moisture (Watrin et al. 2009). In addition, Sahelian plants such as Acacia migrated north into central Saharan mountain ranges sometime between 6300–5800 BC, where plants associated with permanent freshwater environments (e.g., Juncus, Potamogeton) peaked early in this phase (Mercuri 2008).
Brief Arid Event, and Renewed Moisture
Northern Saharan records show an arid episode c. 6000–5500 BC (Gasse 2000). Reduced tufa sedimentation in the Tadrart Acacus (c. 6100 BC) suggests decreased local rainfall, though spring activity continued until c. 5600 BC (Cremaschi et al. 2010).
Although regional records suggest aridity 6340–6210 BC (Cremaschi and di Lernia 1999), pollen spectra do not intercept the arid event in the Tadrart Acacus (Mercuri 2008). Local effects may have varied. Archaeological surveys suggest people were more mobile and had more ephemeral sites (Cremaschi and di Lernia 1998; Cremaschi and Zerboni 2009). The last major transgression of lakes in the southern Sahara occurred c. 5000 BC (Gasse 2000).
Late Holocene Arid Phase
By 4000–3000 BC, lower summer insolation weakened monsoons (Lézine 1989; Williams 2009). Progressively, the 100-mm isohyet moved 250 km south and rainfall declined as the ITCZ retreated south. Many lakes regressed; those fed by groundwater persisted longer than rain-fed playas (Nicoll 2004). The northern Sahara saw lacustrine regressions and fluctuations and increasing salinity 5400–3400 BC; lakes dropped to present levels after 3000 BC (Fontes et al. 1985). The southern Sahara saw regressions after 5000 BC, with a brief, modest increase in moisture 1800 BC (Gasse 2000).
Diminished moisture eventually affected flora. Dendroclimatic research in Wadi Tannezuft, SW Libya , shows the growth of Cupressus dupreziana was affected by droughts 3900–3760 BC and again at 3120 BC, and extremely arid conditions that prevailed during the past 500 years (Cremaschi et al. 2006). In the central Sahara , tropical plant communities were replaced by herbaceous-dominated desert ecosystems (Damnati 2000; Lézine et al. 2011; Mercuri et al. 2011). Some central Saharan areas had fairly sudden vegetation changes: a shift toward permanent aridity began 4000 BC (Mercuri et al. 2011; Cremaschi et al. 2014), and by 3400 BC hyperarid habitats were present, as indicated by psammophilous vegetation (Mercuri et al. 2015). In the northern Sahara aridity increased gradually in the Middle Atlas, Morocco and in Libya (Nourelbait et al. 2015; Giraudi et al. 2013). There, at Jebel Gharbi, Zygnemataceae (algae) HdV-Type E indicate fresh but stagnant water, possibly due to drier conditions. As aridity increased between 3400 and 2500 BC grassland-like vegetation first expanded and then contracted in favor of even more xeric-adapted plants (Giraudi et al. 2013).
The Eastern Sahara
The Eastern Sahara is commonly regarded as more arid than other parts of northern Africa, and encompasses inland environments closest to the Nile (Fig. 1). Few data sources exist for the initial stages of the AHP, but several lines of evidence become available starting c. 8500 BC. Here we review environmental inferences drawn from sediment and pollen sequences from paleolakes in this part of the Sahara, and additional terrestrial evidence. Our review begins with Lake Gureinat on the southern edge of the eastern Sahara , then follows a series of archives eastward across northern Sudan from El Atrun to Selima, and finally concludes with Lake Yoa in the northwest corner of the eastern Sahara . We then explore spatial variation in precipitation and vegetation across the eastern Sahara region.
Gureinat Paleolake
The southernmost well-studied paleolake in the eastern Sahara is Gureinat, south of Wadi Howar in western Sudan . Gureinat is thought to have been mainly rain-fed, and not linked to other paleodrainage or groundwater systems, although the presence of hippos suggest connections to other deep-water habitats existed at some point. Still, Gureinat deposits are regarded as excellent indicators of local precipitation, providing an important benchmark of monsoon strength. Its sequence begins before 9000 BC when aeolian sands underwent fluvial redeposition and groundwater levels rose and fluctuated. Around 9000 BC, the water table reached the basin’s lowest point and a shallow lake formed. Sediments, diatoms, and chemical records suggest lake levels and conditions continued to fluctuate, with regressions likely at c. 8500 and 7000 BC and high stands likely at c. 8100 and 6700 BC. Then Gureinat’s levels became more stable. After a brief (possibly century-long) major regression between 6200 and 5600 BC, the lake refilled to its maximum. Although the latest clear evidence for Lake Gureinat occurs at 5100 BC, hippos as late as 2000 BC indicate continuation of local conditions wetter than today (Hoelzman et al. 2010).
El Atrun
Lying c. 70 km north of Wadi Howar, the large El Atrun depression contains a sequence of lacustrine sediments covered by sands. Pollen data show tropical arboreal taxa as continuously present c. 9300–7500 BC, increasing 7500 BC, and declining c. 5700 BC (Ritchie and Haynes 1987).
Jahns (1995) divided El Atrun’s sequence into four local pollen assemblage zones (LPAZ). Zone A (9100–8900 BC) sediments show initial lake formation and high biogenic activity; pollen suggests abundant sedge and steppic vegetation and few trees , probably due to the underrepresentation of pollen of Sudanian arboreal taxa in spectra. Zone B’s (8900–8000 BC) carbonate mud sediments point to decreased salinity and alkalinity and a more humid climate; pollen contains more Sudano-Sahelian and Saharo-Sahelian elements suggestive of wooded savanna. Zone C (8000–7400 BC) has multiple lines of evidence for more arid conditions: higher sedimentation rates, chemical indicators of increased alkalinity and reduced open water, overall reduction in pollen concentration, diminished representation of pollen from moist environments, and heightened frequency of pollen from steppic vegetation. Zone D (7400–5900 BC) records a further increase in salinity and alkalinity, but other chemical indicators suggest open water returns. Overall pollen concentration increases again, and suggests a wooded savanna with the sequence’s highest representation of Sudanian pollen such as Celtis, Piliostygma, Hyphaene, Phyllanthus, and Salvadora persica (Jahns 1995).
The Western Nubian Paleolake
A large paleolake area ~100 km NW of El Atrun has yielded five stratigraphic sequences from four profiles. These suggest lake formation began by 8900 BC. A large, coherent freshwater lake existed by 6300 BC, becoming segmented or discontinuous by 4000 BC. Progressive desiccation reduced lakes, but water may have lingered in some areas as late as c. 2300 BC. Human occupation of the area is documented between c. 6000 and 2000 BC, when the lake dried up (Hoelzman et al. 2001).
Oyo
About 50 km NE of the Western Nubian Paleolake’s eastern shore, Paleolake Oyo began forming before 7500 BC. Oyo received seasonal input of freshwater until c. 5000 BC. Pollen from these fully lacustrine deposits include tropical Sudano-Sahelian taxa, so the area probably supported a savanna woodland/scrub mosaic similar to that found today in Darfur and Ennedi uplands, and savanna-woodlands in Khordofan ~500 km south of Oyo (Ritchie 1994).
At c. 5000 BC, Oyo sediments show downslope resedimentation and slumping, possible indicators of regression. Increasing amounts of windblown sands suggest lake levels dropped further by c. 4750 BC. Vegetation incorporated more Sahelian (savanna) and Saharan (thorn scrub) taxa and fewer deciduous savanna-woodland elements. Final desiccation occurred by 3700 BC, and vegetation was reduced to desert scrubs and grasses near other remaining oases (Ritchie et al. 1985; Ritchie 1994). After 3500 BC, remnants of Sudanian and Sahelian savannas around the lake were replaced by Acacia-thorn savanna and shrub-grasslands with increasing Cyperaceae until 2500 BC, when aeolian sands covered the lacustrine sediments (Ritchie 1994).
Selima
Located >400 km NE of Oyo and El Atrun, Selima lies in the heart of the eastern Sahara ’s driest areas, ~100 km west of the Nile ’s Second Cataract. Around 7700 BC the water table rose, creating a 14-m deep lake at Selima and other lakes in the area (Haynes et al. 1989). Pollen data from a Selima core c. 9200–5100 BC indicate a drier set of surrounding vegetation than contemporary deposits at Oyo and El Atrun. After initial development of Typha marshes, fully lacustrine conditions emerged by 9200 BC and Poaceae and Cyperaceae dominate the pollen spectrum. A sparsely wooded savanna was comparable to modern Sahelian-Sudanian vegetation, and featured Acacia, Commiphora and Maerua, Chenopodiaceae/Amaranthaceae shrubs, and perennial herbs (Tribulus, Blepharis) (Ritchie et al. 1985; Ritchie and Haynes 1987; Haynes et al. 1989; Ritchie 1994).
Pollen of Sudanian taxa typically associated with moister areas were never abundant at Selima, but extrapolations from core dates suggest slight peaks c. 9200, 6900, and 6500 BC (Ritchie and Haynes 1987). Later research suggested a narrower moist interval: c. 7500–5900 BC (Haynes et al. 1989). At c. 6100 BC (2.4 m core depth), Chenopodiaceae/Amaranthaceae increased, and Saharo-Sahelian taxa present throughout Selima’s sequence reached their peak frequency c. 6100–5550 BC. Vegetation changed to a semi-arid steppe or desert with chenopods 5000–4000 BC (Ritchie 1994), and the lake dried up by 2000 BC (Hoelzman et al. 2001).
Charcoal from contexts 5250–4560 BC complements pollen data, recording Sahelian taxa such as Acacia sp., Acacia albida, Maerua crassifolia, Leptadenia pyrotechnica, Ziziphus sp., Boscia senegalensis, Balanites aegyptica, cf. Cassia senna, and Chenopodiaceae (Neumann 1989).
Lake Yoa
In the NW corner of the eastern Sahara region, northern Chad’s Ounianga lakes (including Yoa) drain the Tibesti mountains. Lake Yoa deposits cover the last 6000 years. Low surface water conductivity and magnetic susceptibility, coupled with high organic content, suggest a productive freshwater lake that formed c. 3600 BC. Yoa changed to a more shallow, saline water body 2200–1900 BC (Kröpelin et al. 2008).
Wooded communities shifted gradually to desert in this large basin. Tropical and montane trees and shrubs and fern spores were important elements of pollen assemblages 4000–2300 BC, reflecting local humid conditions and continuing freshwater runoff from the Tibesti mountains. Xeric taxa, such as Acacia, Boerhavia, and Tribulus, increased c. 2800 BC. Commiphora and Acacia became significant locally c. 2700–2100 BC. Steppic vegetation (Chenopodiaceae/Amaranthaceae, Artemisia, Ephedra, Salvadora persica) became dominant after 700 BC, along with Typha (Lézine 2009; Lézine et al. 2011; see also Kröpelin et al. 2008).
Other Terrestrial Environmental Archives
Wood charcoal from terrestrial sources records environmental changes in Egyptian portions of the eastern Sahara (Neumann 1989). Areas north of 25°N had contracted desert vegetation of the northern type (dwarf shrubs, likely sustained by winter rainfall) 8150–5100 BC.
At Abu Ballas (~24° N), flora was still limited c. 7200 BC, with acacias and tamarisks dominating and isolated examples of Maerua, Leptadenia, and Chenopodiaceae. By 5900–5500 BC the floral assemblage was more diverse, adding Grewia, Calotropis, Ziziphus and cf. Cassia senna. These probably represent the limits of expansion for these Sahelian taxa, and Neumann (1989) notes that runoff capture made the Ballas Mudpans exceptionally moist for that latitude.
The Gilf Kebir, slightly farther south and 1000 m asl., has charcoal assemblages dominated by Tamarix and (to a lesser degree) Ziziphus 6500–2900 BC. Slightly wetter conditions may have occurred at 5550, 4530, and 3900–3600 BC given the appearance of Acacia, Maerua, and/or Balanites, and 5000 BC (Acacia albida).
Wadi Howar (the ‘Yellow Nile ’ drainage system, North Sudan ) drained portions of the Ennedi Mountains and Jebal Marra. Analogies with other parts of the Sahara (based on pollen comparisons) have led Neumann (1989) to propose that Sudan Zone vegetation would have been present c. 5900 BC in the dunes near Wadi Howar. Such vegetation might have included taxa such as Terminalia cf. macroptera, Annona senegalensis, cf. Rhus sp., Crateva adansonii, Celtis integrifolia, Ximenia americana, Ficus sp., Boscia cf. salicifolia, Balanites aegyptiaca, Cadaba farinosa, Ziziphus sp., and Acacia albida. By 4150 BC more arid conditions fostered a more restricted set of Sahelian taxa, recovered in charcoal assemblages around Wadi Howar itself: besides the dominant Acacia sp., charcoal samples have only Acacia nilotica, Ziziphus sp., cf. Grewia tenax and Capparidaceae (Neumann 1989).
In central Sudan , charcoal analysis suggests a thorn savanna vegetation 4900–3800 BC near the archaeological site of Kadero (Barakat 1995). Various Acacia species (including A. nilotica), Commiphora sp., Capparidaceae (Boscia senegalensis), Grewia sp., Balanites aegyptiaca, and Cadaba sp. are all present. Barakat notes that Kadero’s charcoal assemblage may reflect proximity to the Nile , human selection of taxa for burning and use, and environmental modification by humans such as landscape burning and grazing .
Regional Patterns Within the Eastern Sahara
Several eastern Saharan records belie the notion of sudden climate changes during the Middle Holocene , or quick, dramatic oscillations between arid and wet conditions (Kröpelin et al. 2008). Tropical taxa may have diminished gradually, depending on local sediments’ propensity to trap or shed runoff, and the dependability of precipitation entering local drainage systems.
A west-east transect of paleoenvironmental data sources at ~23° N shows that freshwater lakes existed to the west (closer to the central Sahara ) at the same time as mudpans existed to the east (closer to the Nile ), thereby suggesting that western areas received more moisture and sustained a more generous complement of vegetation. Exceptions to this trend are found in two settings. First, where uplifted basement rocks created higher groundwater pressure, freshwater surface ponds could form (e.g., near Bir Tarfawi). Second, montane locations experiencing orographic effects (e.g., Meidob Hills) may have captured moisture earlier and in greater quantity, and contributed to lowland parts of the eastern Sahara through distributaries (Pachur and Hoelzmann 2000).
A south-north transect reveals a similar gradient of moisture from high (south) to low (north). As one moves north, freshwater lakes give way to playas and pans, Mg and Sr ions become more concentrated in lake carbonates, and 18O values become more depleted (Pachur and Hoelzmann 2000).
The Nile Corridor
The Nile and areas immediately adjacent are environments that vary independently from inland areas. Because fluctuations in Nile flow are mainly influenced by precipitation in the western Ethiopian highlands, Uganda, and South Sudan (Fig. 2), they are not necessarily in step with broader environmental changes seen in the eastern Sahara or the greater Saharan region. This is partly due to the fact the SE African monsoons from the Indian Ocean govern precipitation in the Nile headwaters, Nile Valley and eastern Sahara , whereas SW African monsoons from the Atlantic/Gulf of Guinea govern moisture delivery in central and western parts of the Sahara. It also reflects the topography, gradient, and physiographic contexts of the various Nile tributaries (reviewed above in “The Nile and Its Exceptional Environments”).
Here we review major changes in Nile flow as seen through deposits along the Blue Nile , White Nile , and sapropel activity indicative of outflow into the Mediterranean Sea. We also consider pollen archives from areas near to Nile sources and from riparian environments adjacent to the middle Nile (from the confluence of the Blue and White Niles at Khartoum, to Aswan near the First Cataract).
The AHP Onset: Resumption of Constant Flow c. 13,000–10,000 BC
Nile flow was intermittent and seasonal during the Late Glacial. Returning monsoons raised the levels of Lake Victoria and Lake Albert to overflow c. 13,000 BC (OSL dating), restoring White Nile flow and pushing it to extremely high levels c. 12,700–11,100 BC (Williams et al. 2006; Williams 2009). Alluvial plains near Ed Dueim (Fig. 3) show elevated White Nile flow c. 11,300 BC (OSL dating) (Williams et al. 2010). Lake Tana overflowed c. 12,750 BC and restored heavy flow to the Blue Nile 11,900–11,200 BC, but underwent a minor, temporary regression at 10,000 BC (Lamb et al. 2007; Williams 2009).

Archaeological sites along the Nile mentioned in the text.
Nile Flow Patterns During Peak Saharan Moisture c. 10,000–7000 BC
Alluvation resumed along the White Nile at Ed Dueim 7800 BC (OSL) (Williams et al. 2010). C. 7700–7000 BC, White Nile floods were higher (3 m above present levels, but not as high as during the initial overflow), and other western Nile tributaries became active (Gatto and Zerboni 2015). The Blue Nile does not show a high river level during this time interval, but its sedimentation record is more fragmentary than the White Nile ’s (Williams 2009).
Wadi Howar (the “Yellow Nile ”) was an important Nile tributary by 7300 BC. For the remainder of the Early Holocene , it brought water from the Ennedi and northern Jebel Marra to join the main Nile at Dongola Reach (Nicoll 2004). Downstream near Kerma on the Middle Nile ’s eastern bank, a large, swampy plain existed 8300–5300 BC (Honegger and Williams 2015).
A sharp increase in Nile freshwater outflow caused the deposition of Mediterranean sapropel S1. Starting dates for sapropel deposition range from c. 11,000 BC to 7000 BC, and ending dates range from c. 4800 to 4000 BC (Aritzegui et al. 2000; deMenocal and Tierney 2012; reviewed by Williams et al. 2015b:104). Depleted 18O signatures in sapropel S1 point to strengthened contributions from the Indian Ocean monsoon system, rather than Atlantic monsoons (Hennekam et al. 2014). Thus, Indian Ocean-sourced Early Holocene Nile flow may not always have correlated with changes in Atlantic Ocean-sourced moisture delivery in central and western parts of the Sahara.
Nile Behavior During the Saharan Arid Event c. 6200 BC and After Resumption of Saharan Moisture c. 6000–5000 BC
Although many parts of the Sahara show reduced moisture, Blue Nile sediments record high river levels as late as 6600 BC (Williams 2009). Slightly thereafter, Lake Victoria and Lake Tana dropped 6400–5800 BC, and Nile floodplain and channel systems contracted (c. 6150–5750 BC) (Macklin et al. 2015). Still, elevated flood levels are evident along the White Nile (5900–5600 BC) and Blue Nile (c. 5700 BC) (Williams 2009). Thus, the flow of upper portions of the Nile does not decline in step with the Saharan aridity event but slightly later—perhaps after monsoonal retreat was felt farther south in Nile headwaters—and temporarily. Aquifer-fed environments in the Sahara responded more quickly to decreases in rainfall than the Nile rivers.
Middle Holocene Desiccation and Nile Response (after 5000 BC)
By c. 5000 BC, the Nile River had irregular flood events. Dune activity at Khor Abu Habl by the White Nile (Fig. 3) attests to diminished flow c. 4600 BC (OSL) (Williams et al. 2010), although the White Nile and Blue Nile both show high floods at 4300 BC (Williams 2009). The western Nile tributaries, Wadi Howar and Wadi Melik, lowered and eventually stopped flowing into the Nile c. 4000–2800 BC (Nicoll 2004), but local wadis near the Dongola Reach (Fig. 3, between Kawa and Debba) continued to be active (Woodward et al. 2015). Drier conditions favoured human occupation of the Kerma plain, however: An extensive channel network supported animal and vegetal life, not only along the palaeochannels, but also throughout the plain 5300–3500 BC (Macklin et al. 2013). Major changes in river flow and dynamics in the Nile catchment included phases of channel and floodplain contraction 4400–4150, 3700–3450, 2700–2250, and 1350–900 BC (Macklin et al. 2015).
Desiccation effects grew more pronounced thereafter. Dunes reactivated at Khor Abu Habl 2800 BC (OSL) and aggradation stopped by 2500 BC (Williams et al. 2006; Williams 2009). Although high floods took place along the White Nile at 1200–800 BC (Williams 2009), dunes were also forming c. 900 BC (OSL) (Williams et al. 2010), suggesting that local conditions were deteriorating even as some Nile flow continued. In the Northern Dongola Reach, major paleochannel systems east of the Nile dried out c. 1320–1290 BC (Macklin et al. 2013). Today, however, their remnants still represent the largest alluvial plain in Nubia , extending >13 km east-west and 100 km north-south, with several arms of the Nile running parallel to the main river (Honegger and Williams 2015).
Pollen Archives from Nile Tributaries and Riparian Areas
Few locations along the Nile have yielded pollen assemblages. Along the Blue Nile in central Sudan , El Malahab and Sheikh Mustafa (early Holocene ) and Sheik el Amin (middle Holocene ) have disturbed contexts that hinder assessment of precise diachronic changes. Pollen from all three sites suggests open savanna, with Acacia only present at El Malahab. Arid elements such as Chenopodiaceae/Amaranthaceae, Capparis, Boscia, and Maerua are present. Sheikh Mustafa has evidence for periodic landscape clearance via burning, and Sheikh el Amin has coprophilous ascospores suggestive of pastoral land use (López-Sáez and López-García 2003).
In northern Sudan several archaeological sites on and near Sai Island have yielded pollen assemblages (Florenzano et al. 2016, 2018). These spectra, which together comprise a sequence covering much of the Holocene , will be discussed in greater detail below; see “Pollen records from archaeological sites on Sai Island”
Implications for Human Behavior and Resource Access Issues
Reviewing paleoenvironmental data at these three scales suggests a few important points and unresolved issues. The general Saharan and eastern Saharan records are distinct in two ways that have important implications for resource distributions and human economic activities.
First, some sudden trends in the main Sahara seem muted in the eastern Sahara : vegetation shifts, and consequent changes in human behavior, may have been more gradual. Thus, although the eastern Sahara has the reputation of being the driest part of the Sahara, its fluctuations may have been less extreme.
Second, within the eastern Sahara , aridity increases in an easterly direction. Areas near the Nile , but away from its floodplain and riparian edge, would have been among the most arid environments in the Sahara at any given time. They may have offered significantly less vegetation and resources than the northwest Sudan paleolakes reviewed above, and more closely resembled environments documented at Selima. Eastern Saharan data support broader impressions that “greening” of the Sahara was neither universal nor homogeneous; rather, Saharan environments changed in a patchy fashion depending on localized conditions of circulation, topography, and substrate.
However, the Nile and its riparian environments fluctuated in distinct ways. While the Sahara depended on Atlantic circulation for monsoon rains, Nile flow depended on low-latitude precipitation influenced by Indian Ocean monsoons. Changes in Nile flow and floodplain size may therefore have been out of step—delayed—in comparison to Saharan patterns. In addition, the Nile would have served as a linear refugium, maintaining taxa that could no longer survive farther west during arid phases, and potentially allowing their more speedy introduction inland when moist conditions resumed.
Holocene Plant Use Along the Nile
The foregoing multiscalar review demonstrates the diverse forces that influenced environmental changes throughout the Holocene in the Sahara and Nile . To understand their localized effects on resources and human livelihoods requires explicit examination of archaeobotanical records from sites near the Nile itself. In this section, we review prior studies of plant use at archaeological sites near the Sudanese and Egyptian Nile (Fig. 3).
Plant Use Along the Nile During Peak Moisture Delivery
Early Holocene sites in Sudan (c. 8000–5000 BC) have many material cultural commonalities, such as pottery decorated with ‘wavy-line’ incisions and ‘dotted-wavy line’ and rocker-stamp impressions. Slight differences in decorative techniques and motifs distinguish those in central Sudan (‘Early Khartoum’) from those downstream from Debba (‘Khartoum Variant’) (Garcea and Hildebrand 2009). However, archaeobotanical records from northern and central Sudan are not comparable.
While northern Sudanese sites from this period have yielded little botanical evidence, macrobotanical remains and plant impressions in pottery at Early Khartoum sites suggest that fisher-gatherer-hunters ate plants that did not need special processing and/or preparation (e.g. fruits and berries such as Celtis integrifolia, Ziziphus spina-christi and Grewia tenax), as well as grains of wild cereals that do (e.g., Sorghum , Setaria, Echinochloa and Panicum species) (Magid 2003). This type of edible plant exploitation indicates an economic strategy based on a complex and broad spectrum of vegetal foods.
Farther south in Central Sudan , consumption of wild tubers, including Cyperus rotundus L., was documented via dental calculus from foraging and farming/herding individuals buried in the cemetery of Al Khiday, on the White Nile in central Sudan (Buckley et al. 2014). This sedge is appreciated for its high carbohydrate content and other non-dietary values (as a perfume and/or medicine).
Plant Use Along the Nile During the Onset of Food Production
Herding preceded the onset of village agriculture in most areas. In Sudan broad spectrum exploitation of plant foods continued without significant changes even after the arrival of domestic livestock at Shaheinab, Sai Island , and other sites dating to around 5000–4000 BC. Macro-botanical evidence includes fruits of Celtis and Ziziphus , as well as cereals such as Sorghum , Setaria, Echinochloa and Panicum . The primary activities continued to be focused on the consumption of plants that did not require processing and preparation (Magid 2003).
Currently, the earliest evidence for domestic plants along the Nile is at cemetery R12 in northern Sudan (Salvatori and Usai 2008), where domestic cereals of southwest Asian origin are present by c. 5311–5066 BC (Out et al. 2016). Phytoliths in human burial deposits are attributed to Hordeum sp. and/or Triticum sp., which are thought to be domesticated because there are no wild ancestors in the region (Madella et al. 2014). It has been suggested that these plants were used in the burials for ritual purposes as they represent rare or exotic (luxury?) plants. They may have been used as pillows or mats (Out et al. 2016) or as funerary offerings (Garcea et al. 2016a). In addition, dental calculus with starch of Hordeum sp./Triticum sp. from human burials farther south at El Ghaba (4750–3650 cal BC) may suggest that these plants were also used as food (Out et al. 2016).
Farther south up the Nile , phytoliths and starch in human dental calculus at the cemetery of El Ghaba (4750–3650 cal BC) show that Hordeum sp. and/or Triticum sp. were used for both ritual purposes and food (Madella et al. 2014; Out et al. 2016). On the other hand, wild C4 grasses were much more common than domestic C3 cereals, indicating continuing regular wild plant exploitation and even management for centuries. Bucrania (domestic cattle skulls) were found together with phytoliths of wild panicoid grasses. While these finds document consumption of crops, it is not yet clear whether wheat and barley were locally farmed at this time.
The earliest large-scale evidence for domestic plants is at Kom W and K in the Fayum, where the southwest Asian domesticates emmer wheat (Triticum turgidum L. ssp. dicoccum (Desf.) Husn.) and hulled barley (Hordeum vulgare L.) are stored in bulk by 4650–4350 BC (Wetterstrom 1993; Wendrich et al. 2010). Away from the Nile , emmer has been found in deposits dating to 4450 BC at site KS043 at Kharga Oasis (Briois et al. 2012; Wuttmann et al. 2012). Because only eight grains of emmer were found, chaff was absent, and grains had been cleaned before entering the site, one cannot build a systematic case for local farming. It is possible that grains were brought in from elsewhere: KS043 also at this time has large catfish, and bivalves such as Spathosis rubens and Corbicula consobrina. Because these taxa could not have lived in the oasis, but only in the Nile River, they argue for long-distance exchange networks between Kharga and the Nile (Briois et al. 2012).
Full agriculture was practiced in the Egyptian Nile Valley at Hierakonpolis c. 3750–3300 BC (Fahmy 2003). Emmer wheat provided food and drink, but bread wheat (Triticum durum/aestivum) was also present. Plants were also grown for non-alimentary purposes. Six-rowed barley (H. vulgare) and two-rowed barley (H. distichon L.), which appeared in higher quantities than wheat, were seemingly grown as fodder for livestock, indicating that herding was important. Linen bandages in tombs show that flax (Linum usitatissimum L.) was cultivated for weaving. Wild plants such as Cyperus esculentus L., Ziziphus spina-christi (L.) Desf., and Balanites aegyptiaca (L.) Del. continued to provide food (Fahmy 2003).
Between the 1st and 2nd Cataracts near the Egypt /Sudan border, A-Group sites (c. 3700–2400 BC) have evidence for farming of domestic wheat, barley, and legumes; herding cattle, sheep, and goats; and hunting and fishing. Sites occur both by the Nile and away from the river, and inland herding may have yielded to farming near the Nile banks as rainfall diminished. Some A-Group sites have evidence for small- or large-scale storage, and exchange with Egypt 3700–3300 BC; A-Group pottery is also found as an exotic ceramic at Pre-Kerma sites farther south. Food production, physical storage, and long-distance trade all appear to take hold together (Lal 1967; Nordström 1972; Gatto 2006; Lange 2007; Garcea and Hildebrand 2009).
Farther upstream in northern Sudan , despite early dates for (presumably imported) domestic cereals at R12 and Ghaba cemeteries, regular crop cultivation is not documented until c. 2700 BC at site 8-B-52A on Sai Island (Hildebrand 2006–2007; Hildebrand and Schilling 2016). Agriculture was well-developed during the Kerma period, dating from 2500 BC (Iacumin et al. 1998).
In central Sudan , farming of wheat and barley is recorded at Meroe, which is only dated between around 1000 BC and 1000 AD (Out et al. 2016). The earliest clear evidence for farming of indigenous crops ( Sorghum ) occurs c. 780–400 BC at Kawa (Fuller and Smith 2004), and by 0 AD sorghum was more widely cultivated (Rowley-Conwy 1989). Delayed adoption of southwest Asian taxa may relate to local environmental conditions. Consequently, native wild species continued to play a greater weight in the diet than in Egypt . In fact, the intensive consumption of wild grains adopted during the Early Khartoum period became widespread and lasted a long period of time (Stemler 1990; Fuller and Smith 2004; Iacumin et al. 1998).
Pollen Records from Archaeological Sites on Sai Island
Four archaeological sites together span economic transitions from fishing-hunting-gathering to herding to farming at Sai. They also reflect regional changes in material culture, from pre-pastoral Khartoum Variant (c. 7600–4800 BC), pastoral Abkan (c. 5000–4300 BC) and agricultural pre-Kerma/Kerma (c. 1800–1700 BC). Ceramic assemblages reveal significant cultural shifts at Sai during this time interval, involving cross-cultural interactions (D’Ercole et al. 2015; Garcea 2006, 2006–2007, 2011–2012, 2016a, b, in press; Garcea and Hildebrand 2009; Garcea et al. 2016a, b; D’Ercole et al. 2017a, b).
Pollen from Sai sites can clarify the environmental contexts of these economic and social changes. The following sites and contexts have yielded pollen samples: 8-B-10C Level 2 (7600–7200 BC, 2 samples); 8-B-10C Level 1 (5000–4800 BC, 10 samples); 8-B-76 (6400–6250 BC, 5 samples), 8-B-81 (5000–4300 BC, 5 samples) and 8-B-10A (1700 BC, 6 samples) (Fig. 4). Florenzano et al. (2016, 2018) describe pollen recovery, preparation, analysis methods and results for all Sai samples. Florenzano et al. (2016) also report on pollen analyses from the Amara West district (not described here). Pollen samples show poor preservation and low concentrations, presenting taxa from local plant cover or long-distance arrival of pollen . State of preservation and phytogeographical attributions help to distinguish local from regional or extra-regional pollen composition. Steady alteration of seasonal wet/dry conditions may have impeded the full preservation of exines throughout the Holocene sequence, and Nile flood events may have further diluted representation of organic matter.

Topographic map of Sai Island with archaeological sites mentioned in text. Base map by T. Schilling, with contour data from Despagne n.d. Topographic data are not available for mainland areas east of the main channel and west of the minor channel; these areas are the lightest shade of grey.
Site 8-B-10C
This Khartoum Variant site consists of gravel bars with archaeological deposits atop an early Holocene fluvial terrace. Seasonal Nile floods created a marshy environment with stagnating water pools; gravel bars occasionally became separated islets (Hesse 1996; Geus 1998, 2002). The 105-m2 excavation area had two superimposed Khartoum Variant occupation phases: Level 2 (7600–7200 BC), an occupational phase with 75 post holes; and Level 1 (5000–4800 BC), an occupational phase c. 2500 years after Level 2, consisting of a large (semi-) permanent settlement with seven hut floors, 100 post holes, three hearths, and other structural features such as a rubbish pit.
Level 2 has a tiny concentration of <260 pollen grains per gram (p/g) probably reflecting the increased inputs of sediments with poor organic content from Wadi Howar, and the White and Blue Niles (Williams 2009; Gatto and Zerboni 2015). Pollen spectra show low diversity with prevalence of Poaceae (31%) and low percentage of woody plants (8%). Different vegetation types are represented: desert savanna, just out of the green borders of the river (Capparis 3%, Salvadora persica 1%, Chenopodiaceae 4%, Asteraceae 4% excl. Artemisia), wet environments (Cyperaceae 4%), and tropical forest (Celtis, Myrica). The latter type of forest probably did not grow locally but likely lived farther north than its modern range. Considering that some tropical plants are not high-pollen producers, possibly their pollen arrived to the site via water with the river flow. Pollen from the river shore belt is relatively limited suggesting that flood events may have swept away riverbank areas. The mixed spectra from various habitats may also reflect some seasonal variability, also seen in earlier phases studied in the nearby site of Amara West (Florenzano et al. 2016).
Level 1’s pollen concentration is even weaker than in Level 2 (<150 p/g), probably due to floods diluting pollen and other organic matter and depositing large quantities of silt or gravels. Pollen percentages of Poaceae (44%) dominate, suggesting grasslands. Woody plants are similar in percentage (7% on average) to Level 2, but differ in taxonomic composition. Pollen of Acacia (3% on average) indicates desert wooded savanna with Capparis and Tamarix. Also other tropical genera (Cadaba, Barleria, Indigofera) are evident in these spectra. Several Holoarctic pollen grains (broadleaved Quercus, Ostrya, Pinus) attest to long-distance wind transport from a Mediterranean source area. Cyperaceae (12%) are more frequent than in Level 2, and indicate that remnants of the vegetation typical of the riverine belt and marshy environments remain visible, together with tamarisks, reeds and floating plants living in shallow water.
Site 8-B-76
The site lies near a currently inactive floodplain on the western side of Sai Island . Excavations (a 27-m transect and a 2 × 1-m test unit) revealed two occupations: early Khartoum Variant hunter-gatherer occupations (6400–6250 BC), and then Abkan herders (5400–3700 cal BC). Pollen samples were only taken from Khartoum Variant levels, and two samples were sterile. Pollen concentration was c. 300 p/g. Poaceae (18% on average) and Fabaceae (17%) prevail, suggesting their presence near the site (Florenzano et al. 2018). They probably represent shrubby desert savanna where these plants lived on sandy soils with other xerophilous species. Woody plants (7% on average) have similar overall representation to that at 8-B-10C with Acacia (0.6%). Together with other trees like Celtis, Grewia, and shrubs/small trees like Capparis and Myrica, they represent a good contingent of tropical pollen taxa at the site, probably produced by a wooded savanna. Cyperaceae (3%) and other plants from wet habitats are not well represented, but microscopic freshwater algal remains (HdV-181 cf., Fritschiella) suggest wet substrates and freshwater. Fritschiella, which today blooms between August and October near Khartoum (Brook 1956), suggests seasonal dry conditions around 8-B-76. Quercus and Pinus pollen grains witness long-distance transport from Mediterranean regions.
Site 8-B-81
A 4 × 1-m trench revealed Abkan artefacts, dated to 5000–4300 BC. Pollen concentration was poor (<150 p/g). High values of Poaceae (52% on average) and low Cyperaceae (2%) likely represent local grassland or wetland plant cover. According to the phytogeographical affinity (Watrin et al. 2009; Lézine et al. 2011; Hely et al. 2014), pollen from woody plants (11%) represents both long-distance wind transport (e.g., Quercus and Pinus) and a complex indicating local wooded grassland or savanna woodland along rivers (this is, for example, the plant association suggested by pollen by Acacia, Grewia, Morus cf. and Myrica, 5% altogether). These values are consistent with northern penetration of tropical flora seen elsewhere (Lézine 2007).
Comparing our two c. 5000 BC contexts shows differences between the west (8-B-81) and east (8-B-10C, Level 1) sides of Sai Island . Fritschiella, seen at 8-B-81 and earlier at the west side site of 8-B-76, indicates that for millennia, this side of Sai experienced a distinct set of seasonal oscillations: less availability of water overall, and probable drought crises during dry seasons. Less extreme flooding on the eastern side may have allowed more permanent vegetation to grow, such as the riverine wooded grassland mentioned above.
Site 8-B-10A
This stratified site is located in the eastern part of the island about 2 km south of 8-B-10C (Geus 1998; Garcea and Hildebrand 2009). Its ceramic assemblage parallels Pre-Kerma sites elsewhere (D’Ercole et al. 2015, 2017a, b), but its chronology appears to overlap with the Kerma period (2500–1500 BC; Hildebrand and Schilling 2016). Archaeological deposits yielded numerous animal remains, the majority of which were domestic livestock (Chaix 2011). Caprines were much more numerous than cattle (270 vs. 14 remains, or 93.1% vs. 4.8%), but the weight of cattle bones was considerable (94.3 g, or 19.3% of assemblage weight). Small livestock that could be identified as sheep versus goat exhibited a prevalence of sheep (Ovis aries L.), including lambs and young individuals, over goats (96.7% vs. 3.3%). Birds and fish were insignificant. High representation of sheep contrasts with the contemporary site of Kerma, where goats make up 21.8% of the faunal assemblage. Sai sheep appear to have been much smaller in size than Kerma sheep, whereas goats did not show a size difference between the two sites. Because vertebrae and ribs were underrepresented with respect to the other bone segments, Chaix (2011) has suggested slaughtering probably occurred elsewhere, away from 8-B-10A’s habitation area. In pollen spectra, the desert environment is evident from the presence of Capparis (3%), Salvadora persica (0.2%), Chenopodiaceae (3%) and Asteraceae (6%). Celtis, cf. Hagenia abyssinica and Rhynchosia are tropical taxa that probably arrived via river flow from southeastern regions. Poaceae (20%) and Cyperaceae (5%), indeed, represent grassland and riverine vegetation. Plantago, Cichorieae and other Asteraceae may be also connected to browsing and trampling by domestic animals. Although no cereal pollen was found, this is the only site where pollen of palms (Phoenix, and possibly Elaeis?) are recorded. This indicates palms were growing either along the river or inland at oases.
Relations Between Environmental and Economic Changes
Since the beginning of the Holocene , areas in and near to Sai Island have showed a mosaic of habitats with open-vegetation and wet environments (Fig. 5). Palynology reveals that vegetative changes along the northern Sudanese Nile paralleled those documented in the eastern Sahara , albeit with some interesting variations and details.

Percentage pollen diagram of the Sai Island sites (redrawn from Florenzano et al. 2018)
Local Environmental Changes
8700–8500 BC: The palaeoenvironmental picture emerges from site 2-R-66, in the Amara West district dated to the Arkinian period (Florenzano et al. 2016; see also Woodward et al. 2015). The mixed pollen flora including species from southeastern regions testifies that some plant displacement near this area had occurred during the Early Holocene , paralleling Watrin et al. (2009)’s documentation that tropical trees extended their northernmost expansion from 12° to 19° N at c. 9800–5600 BC. Grasses were part of the ephemeral or perennial desert grassland vegetation, as well as the savanna vegetation. The environment was characterised by an open vegetation with a Saharan gravel desert or shrubby vegetation and swamp communities with hygrophilous trees .
7600–7200 BC: Grass cover was again distributed in several habitats including desert savanna and swamp or riverine communities. The xerophilous communities and dry habitats would likely have been positioned just beyond the riparian belt. Even at this time of maximum moisture delivery to the Sahara, we do not see evidence for the local growth of tropical forest species at this time. Vegetation signalling different habitats suggests that there may have been seasonal vegetative growth in different locations—some in response to Nile floods, and some in response to seasonal rainfall.
6400–6250 BC: Wooded savannas appear to have shifted north, such that a good contingent of tropical plants could grow near Sai. A steppe-like environment rich of xerophilous taxa spread during the dry oscillation that characterised this phase.
5000–4800 BC: A desert wooded savanna spread locally while grass cover expanded in grasslands and in the communities typical of the riverine belt and marshy habitats. Floods became less dramatic and even irregular, but still supported well-developed wet environments. The eastern banks of Sai Island had a more open environment, while the western side of the island sustained a wooded grassland or savanna woodland.
2500–1500 BC: At Sai, the spreading of date palms 1800–1700 BC is the first clear evidence of the establishment of anthropogenic environments. Although the desert environment continued to spread as rainfall diminished, the local riverine vegetation continued to grow along the Nile .
Push and Pull: Hinterland and Refugium
Sai area pollen spectra, but especially those from Amara West, suggest that tropical species spread quickly to this area during the Early Holocene . This is in contrast with other archives from the eastern Sahara (e.g., charcoal from Abu Ballas) that suggest gradual ecological changes. Spatial trends of greater aridity to the east and north do not hold for the Nile ; the riverine corridor may have abetted the extension of tropical taxa north from their rain-fed distribution areas. Although Sai Island ’s Early Holocene record has not yielded well-dated evidence for sustained human use until 7600–7200 BC (8-B-10C Level 2), it is possible that such sites underlie the more recent deposits on Sai. Also, Amara West has evidence for human occupation 8700–8500 BC, suggesting that hunter-gatherer-fishers used this stretch of the Nile during the wettest phases of the AHP, even though Saharan environments away from the Nile may have offered attractive sets of resources which people may have seasonally exploited. Subsequent Khartoum Variant horizons (8-B-10C Level 1 and 8-B-76) suggest occupation of Sai intensified as time went on.
Around 5000 BC, the wooded grasslands and grassy woodland environments suggested by the Sai pollen spectra are evidence for increased aridity (Fig. 5), but not as marked as effects seen in other parts of the Eastern Sahara , from the Selima Sandsheet west to Lake Oyo, where lakes shrank dramatically and desert vegetation expanded. Continuing activity of Nile tributaries near Dongola suggests that some eastern Saharan areas still received rainfall or were sustained by large aquifers whose Early Holocene recharges allowed them to continue contributing water even after local precipitation decline; it also suggests that Nile headwaters were still capturing abundant rainfall from the Indian Ocean monsoon. The delayed deterioration of the Sai flora may be due to either of these factors, or simple maintenance of mesic vegetation near a still-active river. Distinguishing among these possibilities would be important to understanding how the last of the Khartoum Variant hunter-gatherers, and the early Abkan pastoral populations, might have juggled their use of riverine versus inland areas.
By 2500 BC, desert environments are clearly expanding at Sai, although grassland and riparian habitats remain. With most Eastern Saharan lake basins drying up by c. 2000 BC, focusing on intensive exploitation of resources near the Nile is the only viable strategy. Sai shows the use of subterranean pits by 3600 BC and the start of an agricultural economy as early as 2700 BC at the specialized storage facility of 8-B-52A (Fig. 4), and presumably human-made stands of date palms by 1800 BC via pollen at 8-B-10A. However, we are lacking knowledge of a critical chronological interval (4500–2800 BC) that might show the exact nature of local environmental changes and human responses leading up to farming and a full focus on Nile resources. Analysis of macroremains from 8-B-52A (dated to 3600–2800 BC) can fill part of this gap, but sites covering earlier parts of this interval (4500–3600 BC), such as 8-B-76 and 8-B-81 have so far not yielded sufficient macroremains for a secure analysis.
Final Remarks
Our knowledge of changing plant uses along the Nile is far from complete, but a few interesting trends and avenues for further research are clear. First, generalized exploitation of grasses and riparian resources continues even after herding takes hold. Second, there seems to be a clear contrast between the Egyptian Nile —where domestic plants occur in contexts and quantities that argue for increasing emphasis on agricultural activities—and the Sudanese Nile , where southwest Asian cereals arrive early in mortuary contexts but economic shifts from mobile herding to sedentary farming and village life take place much later (Hildebrand and Schilling 2016). The first appearances of Hordeum and Triticum at R12 and El-Ghaba are not accompanied by clear signs of an agricultural economy, such as well-defined storage facilities (subterranean storage pits , above-ground granaries or storage bins) and more permanent village architecture. Although scholars have noted strong deflation in northern Sudan (Reinold 2001), such features do appear in the archaeological record in Pre-Kerma times, where they are correctly interpreted as harbingers of a more sedentary lifestyle with a new economic focus on farming (Hildebrand and Schilling 2016). Other material indications of local agricultural production, such as the presence of chaff impressions from domestic plants in pottery, are so far not known from R12 or El-Ghaba, again suggesting that long-distance exchange may have brought wheat and/or barley up the Nile for local consumption at these sites.
What might have caused the delay from the presence of domestic plants to a commitment to agriculture in Sudan ? The paleoenvironmental contexts outlined here can help lay a firmer foundation for future practice-based considerations of agricultural adoption: knowledge of a resource or crop, or occasional consumption, does not necessarily guarantee immediate sweeping economic changes (e.g., Denham and Haberle 2004). One set of events may stimulate initial introduction of a domesticate, followed by decades, centuries, or millennia of occasional, sporadic, or small-scale use; and a completely different set of environmental or social factors may subsequently trigger a profound economic reorientation in which local production and consumption of a crop becomes pervasive. Non-economic factors should also be taken into consideration with regard to the delayed shift to village-based farming along the Sudanese Nile , namely environmental and technical ones. Unlike Egypt , Sudan is located in the summer rain belt, which is not naturally favorable to the cultivation of southwest Asian crops; this would have hindered the adoption of rain-fed (non-irrigated) farming. Well-dated benchmarks for specific environmental changes along the Nile can help evaluate whether environmental events contributed to (or delayed) economic changes.
Multi-scalar examination of paleoenvironmental data from the eastern Sahara and Nile gives important insights about changes in environments, resources, and likely strategies for human livelihoods across the Holocene . As more and more stratigraphic archives yield more precise data, it becomes clear that specific parts of the Sahara experienced distinct changes that were influenced by localized patterns of rainfall and runoff, and substrates and groundwater. Moving beyond the generalized concept of a “greening” and “browning” Sahara to more nuanced readings of local circumstances is essential to providing environmental contexts for specific archaeological sequences, and explaining particular changes in economy and society.
Establishing a detailed palynological record for Sai Island , comparing it to records from other eastern Saharan localities, and exploring its implications for local economic practices is one small step in a massive endeavour. Ideally, such pollen sampling should be applied throughout the Middle Nile to achieve coverage of numerous sites. Riverine sites at different latitudes could be compared to track the advances—and limits—of tropical taxa, and eventual resurgence of desert flora. Such data could provide a much more nuanced picture of the challenges faced by past peoples of the Nile and Eastern Sahara , and the motivations that spurred important innovations in subsistence and social life.
Notes
- 1.
All dates are given in cal BC. In cases where dates were originally published in uncalibrated form, we have calibrated them using Calpal (www.calpal-online.de). Please note that authors of some dates kept them uncalibrated in recognition of potential reservoir effects that may have varied over time, making calibration less accurate.
References
Abdelkreem MIM, Ibrahim DA (2016) Check list of Flora and vegetation of an archeological habitat in North Sudan. P J B C 1:1–9
Abuswar AO (2007) Range management. UNESCO chair on desertification. University of Khartoum, Sudan
Ali MM (2006) Shoreline vegetation of Lake Nubia, Sudan. Hydrobiol 570:101–105
Aritzegui D, Asioli A, Lowe JJ et al (2000) Paleoclimate and the formation of sapropel S1: inferences from Late Quaternary lacustrine and marine sequences in the central Mediterranean region. Palaeogeogr Palaeoclimatol Palaeoecol 158:215–240
Barakat HN (1995) Middle Holocene vegetation and human impact in central Sudan: charcoal from the Neolithic site at Kadero. Veg Hist Archaeobot 4:101–108
Bonnet C (1992) Excavations at the Nubian royal town of Kerma: 1975–91. Antiquity 66:611–625
Bonnet C (2006) Les établisments des cultures Kerma. In: Caneva I, Roccati A (eds) Acta Nubica: proceedings of the X international conference of Nubian studies. Istituto Polygrafico e Zecca dello Stato, Roma, pp 15–20
Briois F, Midant-Reynes B, Marchand S et al (2012) Neolithic occupation of an artesian spring: KS043 in the Kharga Oasis. Egypt J Field Archaeol 37(3):178–191
Brook AJ (1956) A note on the ecology of the terrestrial alga Fritschiella tuberosa in the Sudan. New Phytol 55:130–132
Buckley S, Usai D, Jakob T et al (2014) Dental calculus reveals unique insights into food items, cooking and plant processing in prehistoric Central Sudan. PLoS ONE 9(7):e100808
Cancellieri E, Cremaschi M, Zerboni A et al (2016) Climate, environment, and population dynamics in Pleistocene Sahara. In: Jones S, Stewart B (eds) Africa from 6-2: population dynamics and palaeoenvironments. Springer, Dordrecht, pp 123–145
Chaix L (2011) Contribution à l’étude de l’économie de la période Pré-Kerma. Premiers résultats sur la faune du site 8-B-10A sur l’Ile de Saï (nord Soudan). In: Rondot V, Alpi F, Villeneuve F (eds) La pioche et la plume–Autour du Soudan, du Liban et de la Jordanie: Hommages archéologiques à Patrice Lenoble. Presses de l’université Paris-Sorbonne, Paris, pp 207–223
Chaix L, Grant A (1993) Palaeoenvironment and economy at Kerma, Northern Sudan, suring the third millennium B.C.: archaeozoological and botanical evidence. In: Krzyżaniak L, Kobusiewicz M, Alexander J (eds) Environmental change and human culture in the Nile Basin and Northern Africa. Poznan, Poznan Archaeological Museum, pp 399–404
Cole JM, Goldstein SL, deMenocal PB et al (2009) Contrasting compositions of Saharan dust in the eastern Atlantic Ocean during the last deglaciation and African Humid Period. Earth Planet Sci Lett 278:257–266
Costa K, Russell J, Konecky B et al (2014) Isotopic reconstruction of the African Humid Period and Congo Air Boundary migration at Lake Tana, Ethiopia. Quat Sci Rev 83:58–67
Cremaschi M (1998) Late Quaternary geological evidence for environmental changes in south-western Fezzan (Libyan Sahara). In: Cremaschi M, di Lernia S (eds) Wadi Teshuinat—palaeoenvironment and prehistory in South-western Fezzan (Libyan Sahara). Quaderni di Geodinamica Alpina e Quaternaria, vol 7. CNR, Roma-Milano, pp 13–47
Cremaschi M (2002) Late Pleistocene and Holocene climatic changes in the central Sahara: the case study of the southwestern Fezzan Libya. In: Hassan, FA (ed) Droughts. Food and culture. Kluwer Academic, New York, pp 65–82
Cremaschi M, di Lernia S (1998) Thegeoarchaeological survey in central Tadrart Acacus and surroundings (Libyan Sahara): environment and cultures. In: Cremaschi M, di Lernia S (eds) Wadi Teshuinat—palaeoenvironment and prehistory in South-western Fezzan (Libyan Sahara). Quaderni di Geodinamica Alpina e Quaternaria, vol 7. CNR, Roma-Milano, pp 243–296
Cremaschi M, di Lernia S (1999) Holocene climatic changes and cultural dynamics in the Libyan Sahara. Afr Archaeol Rev 16:211–238
Cremaschi M, di Lernia S (2001) Environment and settlements in the Mid-Holocene palaeo-oasis of Wadi Tanezzuft (Libyan Sahara). Antiquity 75:815–825
Cremaschi M, Trombino L (1998) The palaeoclimatic significance of paleosols in Southern Fezzan (Libyan Sahara): morphological and micromorphological aspects. CATENA 34:131–156
Cremaschi M, Zerboni A (2009) Early to Middle Holocene landscape exploitation in a drying environment: two case studies compared from the central Sahara (SW Fezzan, Libya). C R Géosci 341:689–702
Cremaschi M, Pelfini M, Santilli M (2006) Cupressus dupreziana: a dendroclimatic record for the middle-late Holocene in the central Sahara. Holocene 16(2):293–303
Cremaschi M, Zerboni A, Spötl C et al (2010) The calcareous tufa in the Tadrart Acacus Mt. (SW Fezzan, Libya): An early Holocene palaeoclimate archive in the central Sahara. Palaeogeogr Palaeoclim Palaeoecol 287:81–94
Cremaschi M, Zerboni A, Mercuri AM, Olmi L, Biagetti S, di Lernia S (2014) Takarkori rock shelter (SW Libya): an archive of Holocene climate and environmental changes in the central Sahara. Quat Sci Rev 101:36–60
Damnati B (2000) Holocene lake records in the Northern Hemisphere of Africa. J Afr Earth Sci 31:253–262
deMenocal PB, Tierney JE (2012) Green Sahara: African humid periods paced by Earth’s orbital changes. Nat Educ Knowl 3(10):12
deMenocal P, Ortiz J, Guilderson T et al (2000) Abrupt onset and termination of the African humid period: rapid climate response to gradual insolation forcing. Quat Sci Rev 19:347–361
Denham T, Haberle S (2004) Agricultural emergence and transformation in the Upper Wahgi valley, Papua New Guinea, during the Holocene: theory, method and practice. Holocene 2008; 18:481–496
D’Ercole G, Budka J, Sterba JH et al (2017a) The successful ‘recipe’ for a long-lasting tradition: Nubian ceramic assemblages from Sai Island (northern Sudan) from prehistoric times to the New Kingdom period. Antiquity 91:24–42
D’Ercole G, Garcea EAA, Eramo G et al. (2017b) Variability and continuity of ceramic manufacturing of ethnographic productions since prehistory in Upper Nubia, Sudan. J Archaeol Sci Rep 16:553–563. http://dx.doi.org/10.1016/j.jasrep.2017.04.012
D’Ercole G, Eramo G, Garcea EAA et al (2015) Raw material and technological changes in ceramic productions at Sai Island, Northern Sudan from the 7th to the 3rd Millennium BC. Archaeometry 57(4):597–616
Despagne R (nd) Topographic data files for Sai Island. Geus archives, Lille
Drake NA, Blench RM, Armitage SJ et al (2011) Ancient watercourses and biogeography of the Sahara explain the peopling of the desert. Proc Nat Acad Sci 108:458–462
Fahmy AG (2003) Palaeoethnobotanical studies of Egyptian Predynastic cemeteries: new dimensions and contributions. In: Neumann K, Butler A, Kahlheber S (eds) Food, fuel and fields: progress in African archaeobotany. Heinrich-Barth-Institut, Köln, pp 95–106
Florenzano A, Mercuri AM, Altunoz M et al (2016) Palynological evidence of cultural and environmental connections in Sudanese Nubia during the Early and Middle Holocene. Quatern Int 412B:65–80. https://doi.org/10.1016/j.quatint.2016.01.001
Florenzano A, Mercuri AM, Fornaciari R et al (2018) Plants, water and humans: pollen analysis from Holocene archaeological sites on Sai Island, northern Sudan. Palynology (online). https://doi.org/10.1080/01916122.2017.1384411
Fontes JCh, Gasse F, Callot Y et al (1985) Freshwater to marine-like environments from Holocene lakes in northern Sahara. Nature 317:608–610
Fuller DQ, Smith L (2004) The prehistory of the Bayuda: new evidence from the Wadi Muqaddam. In: T Kendall (ed) Nubian studies 1998: proceedings of the ninth conference of the international society of Nubian Studies Northwestern University, Boston, pp 265–281
Garcea EAA (2006a) Semi-permanent foragers in semi-arid environments of North Africa. World Archaeol 38(2):197–219
Garcea EAA (2006–2007) The Holocene prehistory at Sai Island, Sudan. In: Gratien, B (ed) Mélanges offerts à Francis Geus. Cahiers de Recherches de l’Institut de Papyrologie et d’Egyptologie de Lille 26:107–113
Garcea EAA (2011–2012) Revisiting the Khartoum Variant in its landscape. Cahiers de Recherches de l’Institut de Papyrologie et d’Egyptologie de Lille 29:139–150
Garcea EAA (2016a) Multi-stage dispersal of Southwest Asian domestic livestock and the path of pastoralism in the Middle Nile Valley. Quatern Int 412B:54–64
Garcea EAA (2016b) Semi-permanent foragers in North Africa–An archaeological perspective. In: Tvedt T, Oestigaard T (eds) A history of water, Series 3. Water and Food From Hunter-Gatherers to Global Production in Africa, vol 3. I.B. Tauris, London, pp 29–54
Garcea EAA (in press) The Southern periphery of Egypt in the Predynastic period: Nubia in the 5th and 4th Millennium BC. In: Knabb K, Boswell A (eds) Life at the margins of the state: comparative landscapes from the Old and New World. University Press of Colorado, Boulder
Garcea EAA, Hildebrand EA (2009) Shifting social networks along the Nile: middle Holocene ceramic assemblages from Sai Island, Sudan. J Anthropol Archaeol 28:304–322
Garcea EAA, Karul N, D’Ercole G (2016a) Southwest Asian domestic animals and plants in Africa: routes, timing and cultural implications. Quatern Int 412B:1–10
Garcea EAA, Mercuri AM, Giraudi C (2013) Archaeological and environmental changes between 9500 BP and 4500 BP: a contribution from the Sahara for the understanding of expanding droughts in the “Great Mediterranean”. Annali di Botanica (Roma) 3:115–120
Garcea EAA, Wang H, Chaix L (2016b) High-precision radiocarbon dating application to multi-proxy organic materials from late foraging to early pastoral sites in Upper Nubia. Sudan J Afr Archaeol 14(1):83–98
Garzanti E, Andò S, Padoan M et al (2015) The modern Nile sediment system: processes and products. Quat Sci Rev 130:9–56
Gasse F (2000) Hydrological changes in the African tropics since the last glacial maximum. Quat Sci Rev 19:189–211
Gasse F (2002) Diatom-inferred salinity and carbonate oxygen isotopes in Holocene waterbodies of the western Sahara and Sahel. Quat Sci Revi 21:737–767
Gasse F, Roberts CN (2004) Late Quaternary hydrologic changes in the arid and semiarid belt of Northern Africa. Implications for past atmospheric circulation. In: Diaz HF, Bradley RS (eds) The Hadley circulation: present, past and future. Kluwer Academic, Dordrecht, pp 313–345
Gatto MC (2006) The Nubian A-Group: a reassessment. Archéo-Nil 16:61–76
Gatto MC, Zerboni A (2015) Holocene supra-regional environmental changes as trigger for major socio-cultural processes in northeastern Africa and the Sahara. Afr Archaeol Rev 32:301–333
Geus F (1998) Saï 1996-1997. Archéologie du Nil Moyen 8:85–126
Geus F (2002) Saï 1998–1999. Archéologie du Nil Moyen 9:95–134
Giraudi C, Mercuri AM, Esu D (2013) Holocene palaeoclimate in the northern Sahara margin (Jefara Plain, Northwestern Libya). Holocene 23:339–352
Haynes CV, Eyles CH, Pavlish LA et al (1989) Holocene palaeoecology of the Eastern Sahara; Selima Oasis. Quat Sci Rev 8:109–136
Hely C, Lézine A-M, APD contributors (2014) Holocene changes in African vegetation: tradeoff between climate and water availability. Climate of the Past 10:681–686
Hennekam R, Jilbert T, Schnetger B et al (2014) Solar forcing of Nile discharge and sapropel S1 formation in the early to middle Holocene eastern Mediterranean. Paleoceanography 29:343–356
Hesse A (1996) Reconnaissance archéologique sur les rives du Nil: le cas du site SKP1 de l’île de Saï. Préhistoire Archéologie Méditerranéennes 5:105–110
Hildebrand EA (2006–2007) The significance of Sai Island for early food production in Sudan. Cahiers de Recherches de L’Institut de Papyrologie et d’Egyptologie de Lille 26:173–182
Hildebrand EA, Schilling TM (2016) Storage amidst early agriculture along the Nile: perspectives from Sai Island, Sudan. Quatern Int 412B:81–95
Hoelzman P, Keding B, Berke H et al (2001) Environmental change and archaeology: lake evolution and human occupation in the Eastern Sahara during the Holocene. Palaeogeogr Palaeoclimatol Palaeoecol 169:193–217
Hoelzman P, Schwald A, Roberts N et al (2010) Hydrological response of an east-Saharan palaeolake (NW Sudan) to early-Holocene climate. Holocene 20:537–549
Honegger M (2004a) The Pre-Kerma: a cultural group from Upper Nubia prior to the Kerma civilisation. Sudan Nubia 8:38–46
Honegger M (2007) Aux origines de Kerma. Genava, n.s. 55:201–212
Honegger M, Williams M (2015) Human occupations and environmental changes in the Nile valley during the Holocene: the case of Kerma in Upper Nubia (northern Sudan). Quat Sci Rev 130:141–154
Horowitz A (1992) Palynology of Arid Lands. Elsevier, Amsterdam
Iacumin P, Bocherens H, Chaix L et al (1998) Stable carbon and nitrogen isotopes as dietary indicators of ancient Nubian populations (Northern Sudan). J Archaeol Sci 25:293–301
Jahns S (1995) A Holocene pollen diagram from El Atrun, northern Sudan. Veg Hist Archaeobot 4:23–30
Kröpelin S, Verschuren D, Lézine A-M et al (2008) Climate-driven ecosystem succession in the Sahara: the past 6000 years. Science 320:765–768
Kuper R, Kröpelin S (2006) Climate-controlled holocene occupation in the Sahara: motor of Africa’s evolution. Science 313:803–807
Lal BB (1967) Indian archaeological expedition to Nubia, 1962: a preliminary report. Fouilles en Nubie 1961–1963:99–118
Lamb HF, Bates CR, Coombes PV et al (2007) Late Pleistocene desiccation of Lake Tana, source of the Blue Nile. Quat Sci Rev 26:187–299
Lange M (2007) Development of pottery production in the Laqiya region, Eastern Sahara. CRIPEL (Cahiers de Recherches de L’Institut de Papyrologie et d’Egyptologie de Lille) 26:243–251
Lario J, Sanchez-Moral S, Fernandez V et al (1997) Palaeoenvironmental evolution of the blue Nile (Central Sudan) during the early and mid-Holocene (Mesolithic-Neolithic transition). Quat Sci Rev 16:583–588
Lézine A-M (1989) Late Quaternary vegetation and climate of the Sahel. Quat Res 32:317–334
Lézine A-M (2007) Pollen records, postglacial. In Scott, EA (ed) Encyclopaedia Quaternary Science, vol. 4, pp 2682–2698
Lézine A-M (2009) Timing of vegetation changes at the end of the Holocene humid period in desert areas at the northern edge of the Atlantic and Indian monsoon systems. CR Geosci 341:750–759
Lézine A-M, Hely C, Grenier C et al (2011) Sahara and Sahel vulnerability to climate changes, lessons from Holocene hydrological data. Quat Sci Rev 30:3001–3012
López-Sáez JA, López-Garcia P (2003) Pollen analysis from Early and Middle Holocene archaeological sites in the Blue Nile area, Central Sudan. Complutum 14:397–400
Macklin MG, Lewin J (2015) The rivers of civilization. Quat Sci Rev 114:228–244
Macklin MG, Toonen WHJ, Woodward JC et al (2015) A new model of river dynamics, hydroclimatic change and human settlement in the Nile Valley derived from meta-analysis of the Holocene fluvial archive. Quat Sci Rev 130:109–123
Macklin MG, Woodward JC, Welsby DA et al (2013) Reach-scale river dynamics moderate the impact of rapid Holocene climate change on floodwater farming in the desert Nile. Geology 41:695–698
Madani I, Tahir IF, Hamdeen HM et al (2015) Vegetation ecology and taxonomy of El-Ga’ab area, north-western Sudan. Europ Acad Res 3:2927–2943
Madella M, García-Granero JJ, Out WAD et al (2014) Microbotanical evidence of domestic cereals in Africa 7000 years ago. PLoS ONE 9:e110177
Magid AA (2003) Exploitation of food-plants in the Early and Middle Holocene Blue Nile area, Sudan and neighbouring areas. Complutum 14:345–372
Maley J (1981) Etudes palynologiques dans le bassin du Tchad et paléoclimatologie de l’Afrique nord-tropicale de 30 000 ans à l’époque actuelle. Travaux et documents de l’ORSTOM 129, Paris
Maley J (2010) Climate and Palaeoenvironment evolution in north tropical Africa from the end of the Tertiary to the Upper Quaternary. Palaeoecol Afr 30:227–278
Maley J, Vernet R (2015) Populations and climatic evolution in north tropical Africa from the end of the Neolithic to the dawn of the modern era. Afr Archaeol Rev 32:179–232
Manning K, Timpson A (2014) The demographic response to Holocene climate change in the Sahara. Quat Sci Rev 101:28–35
Mercuri AM (2008) Human influence, plant landscape, evolution and climate inferences from the archaeobotanical records of the Wadi Teshuinat area (Libyan Sahara). J Arid Environ 72:1950–1967
Mercuri AM, Florenzano A, Giraudi C et al (2015) Humans and water in desert ‘refugium’ areas: palynological evidence of climate oscillations and cultural developments in early and mid-Holocene Saharan edges. IANSA–Interdisciplinaria Archaeologica 6:151–160
Mercuri AM, Sadori L, Uzquiano Ollero P (2011) Mediterranean and north-African cultural adaptations to mid-Holocene environmental and climatic change. Holocene 21:189–206
Mustafa MA (2007) Desertification processes. UNESCO chair on desertification. University of Khartoum, Sudan
Neumann K (1989) Holocene vegetation of eastern the Sahara: Charcoal from prehistoric sites. Afr Archaeol Rev 7:97–116
Nicoll K (2004) Recent environmental change and prehistoric human activity in Egypt and Northern Sudan. Quat Sci Rev 23:561–580
Nordström H-A (1972) Neolithic and A group sites. Scandinavian University, Uppsala
Nourelbait M, Rhoujjati A, Benkaddour A et al (2015) Climate changes since the mid-Holocene in the Middle Atlas, Morocco. Clim Past Discuss 11:4097–4121
Out WA, Ryan P, García-Granero JJ et al (2016) Plant exploitation in Neolithic Sudan: a review in the light of new data from the cemeteries R12 and Ghaba. Quatern Int 412B:36–53
Pachur HJ, Hoelzmann P (2000) Paleoclimatic implications of late Quaternary lacustrine sediments in Western Nubia, Sudan. Quat Res 36:257–277
Reinold J (2001) Kadruka and the Neolithic in the northern Dongola Reach. Sudan & Nubia 5:2–10
Reinold J (2006) Les cimetières préhistoriques au Soudan – cotoumes funéraires et systemes sociaux. In: Caneva I, Roccati A (eds) Acta Nubica: proceedings of the X international conference of Nubian Studies. Istituto Polygrafico e Zecca dello Stato, Roma, pp 139–162
Ritchie JC (1994) Holocene pollen spectra from Oyo, northwestern Sudan: problems of interpretation in a hyperarid environment. Holocene 4:9–15
Ritchie JC, Haynes CV (1987) Holocene vegetation zonation in the eastern Sahara. Nature 330:645–647
Ritchie JC, Eyles CH, Haynes CV (1985) Sediment and pollen evidence for an Early to Mid-Holocene humid period in the Eastern Sahara. Nature 314:352–355
Rowley-Conwy P (1989) Nubia AD 0–550 and the “Islamic” agricultural revolution: preliminary botanical evidence from Qasr Ibrim, Egyptian Nubia. Archéologie du Nil Moyen 3:131–138
Salvatori S, Usai D (eds) (2008) A Neolithic cemetery in the Northern Dongola Reach (Sudan): Excavation at Site R12. Sudan Archaeological Research Society, London
Salvatori S, Usai D, Zerboni A (2011) Mesolithic site formation and palaeoenvironment along the White Nile (Central Sudan). Afr Archaeol Rev 28(3):177–211
Schild R, Wendorf F (2010) Late Palaeolithic Hunter-Gatherers in the Nile Valley of Nubia and Upper Egypt. In: Garcea EAA (ed) South-Eastern Mediterranean peoples between 130,000 and 10,000 years ago. Oxbow Books, Oxford, pp 89–125
Schulz E (1991) Holocene environments in the central Sahara. Hydrobiol 214:359–365
Stemler ABL (1990) A Scanning Electron Microscopic Analysis of Plant Impressions in Pottery from the Sites of Kadero, El Zakiab, Um Direiwa and El Kadada. Archéologie du Nil Moyen 4:87–106
Tierney JE, deMenocal PB (2013) Abrupt shirts in the Horn of Africa hydroclimate since the last glacial maximum. Science 342:843–846
Watrin J, Lézine A-M, Hély C et al (2009) Plant migration and ecosystems at the time of the “green Sahara”. CR Geosci 341:656–670
Wendrich W, Taylor RE, Southon J (2010) Dating stratified sites at Kom K and Kom W: fifth millennium BCE radiocarbon ages for the Fayum Neolithic. Nucl Instrum Methods 268(7–8):999–1002
Wetterstrom W (1993) Foraging and farming in Egypt. In: Shaw T, Sinclair PJJ, Andah B, Okpoko A (eds) The archaeology of Africa: food, metals and towns. Routledge, London, pp 165–226
White F (1986) La végétation de l’Afrique. UNESCO, ORSTOM
Williams MAJ (2009) Late Pleistocene and Holocene environments in the Nile basin. Global Planet Change 69:1–15
Williams MAJ, Talbot M, Aharon P et al (2006) Abrupt return of the summer monsoon 15,000 years ago: new supporting evidence from the lowe White Nile valley and Lake Albert. Quat Sci Rev 25:2651–2665
Williams MAJ, Williams FM, Duller GAT et al (2010) Late Quaternary floods and droughts in the Nile valley, Sudan: new evidence from optically stimulated luminescence and AMS radiocarbon dating. Quat Sci Rev 29:1116–1137
Williams MAJ, Usai D, Salvatori S et al (2015a) Late Quaternary environments and prehistoric occupation in the lower White Nile valley, central Sudan. Quat Sci Rev 130:72–88
Williams MAJ, Duller GAT, Williams FM et al (2015b) Causal links between Nile floods and eastern Mediterranean sapropel formation during the past 125 kyr confirmed by OSL and radiocarbon dating of Blue and White Nile sediments. Quat Sci Rev 130:89–108
Woodward J, Macklin M, Fielding L et al (2015) Shifting sediment sources in the world’s longest river: a strontium isotope record for the Holocene Nile. Quat Sci Rev 130:124–140
Wuttmann M, Briois F, Midant-Reynes B et al (2012) Dating the end of the Neolithic in an Eastern Sahara Oasis: modeling absolute chronology. Radiocarbon 54(3–4):305–318
Zerboni A, Trombino L, Cremaschi M (2011) Micromorphological approach to polycyclic pedogenesis on the Messak Settafet plateau (central Sahara): Formative processes and palaeoenvironmental significance. Geomorphol 125:319–335
Zerboni A, Biagetti S, Lancelotti C, Madella M (2016) The end of the Holocene Humid Period in the central Sahara and Thar deserts: Societal collapses or new opportunities? PAGES Mag 24(2):60–61. https://doi.org/10.22498/pages.24.2.60
Acknowledgements
Archaeological excavations at Sai Island and Amara West were supported by the National Geographic Society, Committee for Research and Exploration under Grant # 9201-12 and by the University of Cassino and Southern Latium, Italy (grants to EAAG), and the US National Science Foundation (NSF BCS-0519434 to EH). EAAG and EH wish to thank the Sai Island Archaeological Mission (successively directed by Francis Geus, Didier Devauchelle, and Vincent Francigny), and Sudan’s National Corporation for Antiquities and Museums, formerly directed by Hassan Hussein Idris and currently directed by Abdelrahman Ali Mohamed, for having granted permissions to conduct fieldwork. In addition, EAAG thanks the Amara West Research Project (directed by Neal Spencer). We thank Martin Williams and an anonymous reviewer for their insights, which were extremely thoughtful and helpful. Lastly, we are grateful to the people of Sai Island and Amara West for their continuous interest and involvement in our field research.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Hildebrand, E., Garcea, E.A.A., Florenzano, A., Mercuri, A.M. (2018). Multiscalar Perspectives on Holocene Climatic and Environmental Changes in the Sahara and Nile Corridor, with Special Consideration of Archaeological Sites on Sai Island, Sudan. In: Mercuri, A., D'Andrea, A., Fornaciari, R., Höhn, A. (eds) Plants and People in the African Past. Springer, Cham. https://doi.org/10.1007/978-3-319-89839-1_12
Download citation
DOI: https://doi.org/10.1007/978-3-319-89839-1_12
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-89838-4
Online ISBN: 978-3-319-89839-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)