
Abstract
Advanced magnetic resonance imaging (MRI) techniques are improving our understanding of cerebrovascular diseases. Elucidation of the critical pathways in post-stroke recovery would not only provide important fundamental insight in brain function and plasticity but could also lead the way toward development of new rehabilitation strategies for recovering stroke patients. Improvements in hardware and pulse sequences that decrease scan time while maintaining resolution will continue to impact the field. Post-processing strategies must evolve to encompass these increasingly complicated data sets. It also seems clear that multimodal imaging strategies are necessary to develop more detailed patient profiles that can be used for precision medicine.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
- Functional magnetic resonance imaging
- Stroke
- Structural connectivity
- Blood oxygenation level dependent
- Diffusion tensor imaging
Introduction
Advanced magnetic resonance imaging (MRI) techniques such as diffusion tensor imaging (DTI), functional magnetic resonance imaging (fMRI), magnetic resonance perfusion, magnetic resonance spectroscopy, and volumetric imaging are improving our understanding of normal brain development up to pathophysiology of several adverse processes [1], including cerebrovascular diseases . The application of these techniques for research offers the opportunity to ask questions about where, when, and how the abnormal pattern of anatomical connectivity and plastic changes occurs in stroke. Indeed, neuroimaging techniques, especially the multimodality MRI, have significantly contributed to the understanding of the mechanisms of stroke recovery by characterizing brain structural and functional changes after stroke [2]. Stroke lesions trigger several brain-wide processes to accommodate for tissue loss. MRI has been extensively used to investigate brain activation changes during recovery and has provided important information on monitoring of therapeutic strategies that promote brain repair and functional reorganization after stroke [3].
This review focuses on the advances brought to the understanding of stroke mechanisms by MRI-based neuroimaging techniques, particularly those related to connectivity assessment among brain areas. Stroke is the major cause of long-term disability throughout the world [4], leaving more than half of the patients dependent on daily assistance. Nonetheless, most patients exhibit a certain degree of recovery in the weeks, months, and sometimes even years following stroke, which may be directly related to structural and functional modifications in surviving brain tissue. Several animal and human stroke studies have reported vicarious function of ipsilesional and contralesional brain regions [5], which may contribute to restoration of functions, although the exact mechanisms that lead to functional recovery remain largely unclear. Elucidation of the critical pathways in post-stroke recovery would not only provide important fundamental insight in brain function and plasticity but could also lead the way toward development of new rehabilitation strategies for recovering stroke patients [2].
Brain Connectivity from MRI Data
Connectivity models are based on the concept that the brain is organized by segregation of specialized and anatomically distinct brain regions that are functionally integrated in networks mediating cognitive, sensory, or motor processing [6]. Structural connectivity describes how spatially separated brain regions are physically linked, for example, as demonstrated by invasive tracing of single axons or by noninvasively measuring diffusion along major fiber bundles as in DTI . In contrast, functional and effective connectivity describe how anatomically connected areas interact with each other and can be estimated from noninvasive techniques such as electroencephalography, magnetoencephalography, near-infrared spectroscopy, or fMRI. The last two connectivity approaches, however, fundamentally differ in the way of how these interactions are estimated. Functional connectivity is defined as temporal correlation between spatially remote neurophysiological events. In contrast to this nondirectional, correlative nature of functional connectivity, effective connectivity refers to the causal influences that brain areas exert over another under the assumptions of a given mechanistic model [7].
Structural connectivity is obtained from MRI data through the aforementioned DTI technique [8]. This is a noninvasive MRI method that measures the random motion of water molecules in brain tissue and enables examination of white matter microstructure in vivo. Since white matter tracts are composed of highly oriented fibers, which cause relatively high anisotropy of diffusing tissue water, DTI is very suitable to measure effects on white matter integrity. DTI data consists of mathematical entities, called tensors , that give information on the amount of diffusion happening in every direction within a given voxel of the image. From these data, simpler, scalar measures, such as fractional anisotropy (FA) , can be obtained. FA quantifies the extent to which water diffusion is directionally restricted and is influenced by a number of factors including axonal myelination, diameter, density, and orientational coherence [9].
Functional and effective connectivity are obtained from MRI data by means of the fMRI technique. Functional MRI methods have been traditionally sensitized to changes in cerebral hemodynamics in response to task or stimulus-triggered neuronal activity. The most frequently used type of fMRI data is that obtained by means of the blood oxygenation level-dependent (BOLD) contrast, measured through a T2*-weighted gradient-echo MRI sequence [10]. Although effective connectivity has been, indeed, obtained from task-based fMRI data, functional connectivity has been mostly obtained through resting-state fMRI (rs-fMRI) data.
Rs-fMRI data is obtained without the need of a stimulation paradigm, during wakeful rest, but in the absence of an active task performance [11, 12]. Spontaneous fluctuations in baseline (“resting-state”) neuronal signaling are reflected in low-frequency fluctuations (<0.1 Hz) of the BOLD signal and show temporal coherence between anatomically connected brain regions within a particular neuronal network, such as the sensorimotor system [11]. Throughout the gray matter, the extent of synchronization between these low-frequency BOLD fluctuations has been related to functional connectivity. Correlation of these signals with electroencephalographic brain activity has indicated that these slow hemodynamic fluctuations are associated with neuronal function [13].
There are many approaches to estimate functional connectivity from rs-fMRI data. One of the most used has been the seed-based approach , where a representative time series is extracted from a region of interest (the seed), and this series is correlated to all voxel time series of the brain [14] (Fig. 4.1). Another widely used approach has been independent component analysis (ICA) that aims at retrieving the brain networks that produce similarly varying time series [15]. And more recently, functional connectivity from rs-fMRI has been modeled with the aid of graphs, by parcellating the brain in several (anatomical or functional) regions and computing correlations among all the representative time series of these regions. This allows building a mathematical entity, the graph, composed of nodes (the regions) and edges (the connections), from which many topological properties may be extracted that in turn serve to characterize the underlying brain network [16].

Example of functional connectivity (rs-fMRI) data between cortical motor areas in stroke patients
Some of the advantages of the rs-fMRI technique are that many different cortical systems may be studied with a single acquisition (as opposed to task-based fMRI, where only the system associated to the given task is studied) and that it allows increasing the number of subjects for a given study or performing studies with task-impaired populations, who would be otherwise excluded from a task-based study [17]. Rs-fMRI has been increasingly applied as a tool to study alterations in the brain’s intrinsic functional architecture as potential physiological correlates of neurological disorders [18].
Differently from functional connectivity , effective connectivity , as already mentioned, is estimated from task-based fMRI data. Also, effective connectivity requires an a priori model of the brain regions involved in the task being studied, as well as their interaction. It is, therefore, a model-driven technique, as opposed to the functional connectivity technique, which is data-driven [19]. From the proposed model, effective connectivity approaches infer the strength and direction of the connections, giving some idea of temporal relations among the regions (e.g., which region acted before the others).
Approaches to estimate effective connectivity include psychophysiological interactions [20], structural equation modeling (SEM) [21], Granger causality [22], and dynamic causal modeling (DCM) [23]. SEM is based on the translation of a network model with predefined regions linked by a set of directional paths into a linear regression model. The path coefficients are subsequently estimated using an iterative maximum likelihood algorithm to minimize the difference between observed and predicted covariance matrices [21]. DCM relies on a deterministic model that treats the brain as an input-output system of hidden neural dynamics [23]. This neural model describes changes in the system over time as a function of interactions between regional activity, known experimental inputs, and neuronal parameters [12].
All types of MRI data-derived connectivities (structural, functional, and effective) have been used to shed light on stroke mechanisms, and some of the main results are discussed below.
Insights into Stroke Mechanisms from Functional and Effective Connectivity Studies
Findings from functional connectivity obtained from rs-fMRI can be summarized by two major patterns of changes after stroke: reduced interhemispheric functional connectivity between cortical motor areas, which correlates with the severity of motor deficits, and reduced global network efficiency even in patients with good clinical recovery [12]. Studies in animals demonstrate that interhemispheric resting-state connectivity between ipsilesional primary sensorimotor cortex and its contralesional homologue significantly diminish in the first few days and subsequently increase while sensorimotor functions recover. However, interhemispheric connectivity remains reduced compared with assessments obtained prior to stroke [24].
Carter and colleagues [25] found correlations between rs-fMRI time series in a sensorimotor network consisting of M1, SMA, secondary somatosensory cortex, cerebellum, putamen, and thalamus in both hemispheres. The authors reported that particularly interhemispheric M1 connectivity positively correlates with motor performance at the subacute stage after stroke. In addition, stronger interhemispheric connectivity of ipsilesional M1 and contralesional areas such as thalamus, SMA, and middle prefrontal cortex within the first few days of onset predicts better motor recovery in the next 6 months post-stroke [26]. Also, patients with attention deficits had reduced interhemispheric connectivity between attention-related areas in parietal cortex or language areas in inferior frontal cortex [25].
In patients with sufficient integrity of ipsilesional sensorimotor cortex and corticospinal tract, motor recovery may occur rapidly after stroke and be mediated by reacquisition of normal dominance by ipsilesional sensorimotor cortex. However, in patients in whom the integrity of this sensorimotor cortex is insufficient to support good recovery, increased recruitment of contralesional sensorimotor cortices may be utilized to achieve motor recovery [27]. Nonetheless, the role of contralesional M1 for reorganization after stroke and mechanisms associated with shifts in interhemispheric functional connectivity motor cortices is still debatable.
Xu and colleagues [28] demonstrated decrease of the contralesional primary sensorimotor connectivity in the acute state; Rehme and colleagues [29] and Golestani and colleagues [30] showed the opposite. Greater activation of the contralesional hemisphere may occur to compensate for loss of connectivity in the ischemic region. Functional neuroimaging studies suggest that activity within the sensorimotor network, or ipsilesional motor cortex, is most abnormal early after hemiparetic stroke and motor recovery is related to normalization of its activity [27].
Animal models of stroke have provided evidence for a complex cascade of events enabling changes in structural connections and synaptic transmission [31]. These changes occur not only in the vicinity of the lesion but also in remote brain regions. For example, studies in rats demonstrated reduced interhemispheric resting-state connectivity with secondary myelin degeneration of transcallosal fibers within M1 [31]. In humans, it is possible to identify reduced integrity of transcallosal fiber tracts between motor areas that might result from a secondary degeneration of fibers connected to the lesion zone [32]. Lu and colleagues [33] reported reduced functional connectivity between M1 and the contralateral cerebellum in patients with pontine lesions.
We examined motor activation patterns intra- and interhemispheric in patients with stroke compared to healthy controls and the involvement of other functional networks besides the motor network. We observed that among patients with and without preserved function, functional connectivity between the primary motor region and the contralateral hemisphere was increased compared to controls. Nonetheless, only patients with decreased function exhibited decreased functional connectivity between executive control, sensorimotor, and visuospatial networks [34]. Possibly, functional recovery after stroke is associated with preserved functional connectivity of motor to nonmotor networks [34]. A recent study found that stroke patients with such greater capacity for global information integration achieved better performance in a sensorimotor skill training [35]. Thus, one hypothesis is that faster global information exchange may facilitate new functional networks’ configurations [36] (Fig. 4.2).

Example of functional connectivity between the primary motor region and others networks in stroke patients, in longitudinal analysis – time 1 (T1) and time 2 (T2)
Rehme and colleagues [12] believed that stroke lesions do not globally reduce connectivity in all functional systems of the brain but specifically alter connectivity of areas connected to that lesion. The alterations affect the communication efficiency in a given functional network which is closely related to behavioral deficits after stroke. Accordingly, the damage of hub regions has the strongest impact on local and global information transfer. In addition, more random network architectures with less local but high global efficiency seem to promote the relearning of sensorimotor skills but may also explain why performance is often less stable, even in well-recovered patients.
While functional resting-state correlations revealed a reduction of interhemispheric connections after stroke, the most consistent finding from effective connectivity analyses belongs to reduced intrahemispheric interactions in the ipsilesional hemisphere [12]. Grefkes and colleagues [37] showed that compared with healthy subjects, stroke patients with relatively poor motor performance exhibit an enhanced inhibitory influence from contralesional to ipsilesional M1 during movements of the paretic hand [37]. The hypothesis that this inhibition might contribute to the motor deficit of the patients is further substantiated by findings from intervention studies which demonstrated that reducing inhibitory influences from contralesional M1 via repetitive transcranial magnetic stimulation (rTMS) induces significant improvements in hand motor performance [38].
One factor that seems to determine the functional role of contralesional M1 for motor performance of the stroke-affected hand is the time that has elapsed since stroke onset. A longitudinal DCM study with recovering stroke patients showed that interhemispheric inhibitory influences from ipsilesional motor areas to contralesional M1 are significantly diminished in the first few days after onset [39]. After 2 weeks, this apparent disinhibition of contralesional M1 is accompanied by a promoting influence from contralesional to ipsilesional M1, particularly in patients with severe motor deficits. Hence, in the subacute phase, contralesional M1 seems to support activity of motor areas in the lesioned hemisphere. However, after 3–6 months, this supportive influence may turn into inhibition in those patients with incomplete motor recovery. Rehme and colleagues [12] suggest that motor deficits after stroke are not only caused by direct disruption of descending motor pathways but may also depend on a less effective communication between premotor areas and M1 in the lesioned hemisphere.
Another important aspect in stroke is the distributed networks for the control of behavior. It is entirely unknown what patterns of interaction within a network are most closely associated with behavioral deficits after injury [25]. Carter and colleagues [25] showed a critical behavioral significance of interhemispheric connectivity between homologous regions of a task-relevant network. In the dorsal attention network, the breakdown of interhemispheric functional connectivity correlated with difficulty in detecting targets in the contralesional visual field. Besides, connectivity scores in the dorsal attention network also correlated with measures of upper extremity and walking function. Physiological studies indicate that the dorsal attention network is important not only for stimulus selection but also for selection of limb responses [40], especially early after injury [41]. Therefore, the broader behavioral significance of the dorsal network can correspond to its involvement in a larger range of behavioral functions [25].
Vicentini and colleagues [42] found a relationship between increased default mode network (DMN) functional connectivity and depression and anxiety symptoms after stroke. DMN plays an important role in the emotional processing and is an anatomical-functional unit engaged in the processing of self-referential stimuli [43]. According to Vicentini and colleagues [42], patients with depression and anxiety symptoms show an increased connectivity in the left inferior parietal gyrus and left basal nuclei, when compared to patients without symptoms. Specific correlation between depression and anxiety scores and DMN functional connectivity indicates that depression symptoms are correlated with increased connectivity in the left inferior parietal gyrus, while anxiety symptoms are correlated with increased connectivity in the cerebellum, brainstem, and right middle frontal gyrus.
Insights into Stroke Mechanisms from Structural Connectivity Studies
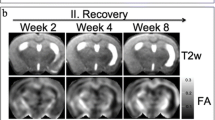
The previous items deal with functional MRI methods that can be employed to identify altered patterns of brain activity after stroke. Changes in functional brain organization, however, are often closely associated with structural modification of neuronal elements in the brain. DTI offers a MRI-based means for the assessment of neuroanatomical changes associated with brain injury and repair [2]. Studies in patients and animals have reported loss of FA in ipsilesional white matter subacutely after stroke, which has been linked to demyelination or axonal loss [44, 45], and elevated FA in ipsilesional corticospinal tracts in chronic stroke patients, which could be associated with chronic reduction in edema or improved motor function [27, 44]. Other example was shown with recovery of FA values 3 years after stroke, which was observed in the internal capsule of patients with upper limb impairments subjected to rehabilitation [46]. Structural integrity of the corticospinal pathway appears critical for a favorable outcome in sensorimotor performance after stroke [2]. Decreases [47] and increases [44, 48] in FA have been observed, and though the exact mechanism of the increase remains unclear, it is possible that perpendicular diffusion is restricted hyperacutely [4].
Finally, other MRI-based techniques not related to brain connectivity assessment may also be used to understand the brain changes and repair mechanisms associated with stroke. For example, voxel-based morphometry (VBM) can be used to detect significant cortical gray matter volume changes in patients with stroke. Matsuoka and colleagues assessed the correlation between changes in cortical volumes and changes in neuropsychiatric symptoms during 6 months following a stroke. They found significant volume reductions in the anterior part of the posterior cingulate cortex and correlation between volume reductions and apathy scale. The delayed atrophy may reflect degeneration secondary to neuronal loss due to stroke. Such degeneration might have impaired control of goal-directed behavior, leading to the observed increase in apathy [49].
Conclusion
The field of experimental neuroimaging with MRI is rapidly expanding. Improvements in hardware and pulse sequences that decrease scan time while maintaining resolution will continue to impact the field. Post-processing strategies must evolve to encompass these increasingly complicated data sets. It also seems clear that multimodal imaging strategies are necessary to develop more detailed patient profiles that can be used to predict outcome [4]. Multicenter studies are increasingly needed to prove these technologies and their usability with stroke patients.
References
Van Horn JD, Pelphrey KA (2015) Neuroimaging of the developing brain. Brain Imaging Behav 9(1):1–4
Dijkhuizen RM, van der Marel K, Otte WM, Hoff EI, van der Zijden JP, van der Toorn A et al (2012) Functional MRI and diffusion tensor imaging of brain reorganization after experimental stroke. Transl Stroke Res 3(1):36–43
Corbetta M, Kincade MJ, Lewis C, Snyder AZ, Sapir A (2005) Neural basis and recovery of spatial attention deficits in spatial neglect. Nat Neurosci 8(11):1603–1610
Farr TD, Wegener S (2010) Use of magnetic resonance imaging to predict outcome after stroke: a review of experimental and clinical evidence. J Cereb Blood Flow Metab 30(4):703–717
Ward NS (2005) Neural plasticity and recovery of function. Prog Brain Res 150:527–535
Friston K (2002) Functional integration and inference in the brain. Prog Neurobiol 68(2):113–143
Stephan KE, Harrison LM, Kiebel SJ, David O, Penny WD, Friston KJ (2007) Dynamic causal models of neural system dynamics:current state and future extensions. J Biosci 32(1):129–144
Le Bihan D (2003) Looking into the functional architecture of the brain with diffusion MRI. Nat Rev Neurosci 4(6):469–480
Basser PJ, Jones DK (2002) Diffusion-tensor MRI: theory, experimental design and data analysis – a technical review. NMR Biomed 15(7–8):456–467
Ogawa S, Lee TM, Kay AR, Tank DW (1990) Brain magnetic resonance imaging with contrast dependent on blood oxygenation. Proc Natl Acad Sci U S A 87(24):9868–9872
Biswal B, Yetkin FZ, Haughton VM, Hyde JS (1995) Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn Reson Med 34(4):537–541
Rehme AK, Grefkes C (2013) Cerebral network disorders after stroke: evidence from imaging-based connectivity analyses of active and resting brain states in humans. J Physiol 591(1):17–31
He BJ, Snyder AZ, Zempel JM, Smyth MD, Raichle ME (2008) Electrophysiological correlates of the brain’s intrinsic large-scale functional architecture. Proc Natl Acad Sci U S A 105(41):16039–16044
Fox MD, Raichle ME (2007) Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat Rev Neurosci 8(9):700–711
Beckmann CF (2012) Modelling with independent components. NeuroImage 62(2):891–901
Bullmore E, Sporns O (2009) Complex brain networks: graph theoretical analysis of structural and functional systems. Nat Rev Neurosci 10(3):186–198
Fox MD, Greicius M (2010) Clinical applications of resting state functional connectivity. Front Syst Neurosci:17(4)
Auer DP (2008) Spontaneous low-frequency blood oxygenation level-dependent fluctuations and functional connectivity analysis of the ‘resting’ brain. Magn Reson Imaging 26(7):1055–1064
Friston KJ (2011) Functional and effective connectivity: a review. Brain Connect 1(1):13–36
Friston KJ, Buechel C, Fink GR, Morris J, Rolls E, Dolan RJ (1997) Psychophysiological and modulatory interactions in neuroimaging. NeuroImage 6(3):218–229
McLntosh AR, Gonzalez-Lima F (1994) Structural equation modelling and its application in network analysis in functional brain imaging. Hum Brain Mapp 2:2–22
Roebroeck A, Formisano E, Goebel R (2005) Mapping directed influence over the brain using Granger causality and fMRI. NeuroImage 25:230–242
Friston KJ, Harrison L, Penny W (2003) Dynamic causal modelling. NeuroImage 19(4):1273–1302
van Meer MP, van der Marel K, Wang K, Otte WM, El Bouazati S, Roeling TA et al (2010) Recovery of sensorimotor function after experimental stroke correlates with restoration of resting-state interhemispheric functional connectivity. J Neurosci 30(11):3964–3972
Carter AR, Astafiev SV, Lang CE, Connor LT, Rengachary J, Strube MJ et al (2010) Resting interhemispheric functional magnetic resonance imaging connectivity predicts performance after stroke. Ann Neurol 67(3):365–375
Park CH, Chang WH, Ohn SH, Kim ST, Bang OY, Pascual-Leone A et al (2011) Longitudinal changes of resting-state functional connectivity during motor recovery after stroke. Stroke 42(5):1357–1362
Schaechter JD (2004) Motor rehabilitation and brain plasticity after hemiparetic stroke. Prog Neurobiol 73(1):61–72
Xu HQW, Chen H, Jiang L, Li K, Yu C (2014) Contribution of the resting-state functional connectivity of the contralesional primary sensorimotor cortex to motor recovery after subcortical stroke. PLoS One 9:e84729
Rehme AK, Eickhoff SB, Rottschy C, Fink GR, Grefkes C (2012) Activation likelihood estimation meta-analysis of motor-related neural activity after stroke. NeuroImage 59(3):2771–2782
Golestani AM, Tymchuk S, Demchuk A, Goodyear BG, Group V-S (2013) Longitudinal evaluation of resting-state FMRI after acute stroke with hemiparesis. Neurorehabil Neural Repair 27(2):153–163
van Meer MP, Otte WM, van der Marel K, Nijboer CH, Kavelaars A, van der Sprenkel JW et al (2012) Extent of bilateral neuronal network reorganization and functional recovery in relation to stroke severity. J Neurosci 32(13):4495–4507
Wang LE, Tittgemeyer M, Imperati D, Diekhoff S, Ameli M, Fink GR et al (2012) Degeneration of corpus callosum and recovery of motor function after stroke: a multimodal magnetic resonance imaging study. Hum Brain Mapp 33(12):2941–2956
Lu J, Liu H, Zhang M, Wang D, Cao Y, Ma Q et al (2011) Focal pontine lesions provide evidence that intrinsic functional connectivity reflects polysynaptic anatomical pathways. J Neurosci 31(42):15065–15071
Almeida SR, Vicentini J, Bonilha L, De Campos BM, Casseb RF, Min LL (2016) Brain connectivity and functional recovery in patients with ischemic stroke. J Neuroimaging 27(1):65–70
Buch ER, Modir Shanechi A, Fourkas AD, Weber C, Birbaumer N, Cohen LG (2012) Parietofrontal integrity determines neural modulation associated with grasping imagery after stroke. Brain 135(Pt 2):596–614
Beharelle AR, Kovačević N, McIntosh AR, Levine B (2012) Brain signal variability relates to stability of behavior after recovery from diffuse brain injury. NeuroImage 60:1528–1537
Grefkes C, Nowak DA, Eickhoff SB, Dafotakis M, Küst J, Karbe H et al (2008) Cortical connectivity after subcortical stroke assessed with functional magnetic resonance imaging. Ann Neurol 63(2):236–246
Grefkes C, Nowak DA, Wang LE, Dafotakis M, Eickhoff SB, Fink GR (2010) Modulating cortical connectivity in stroke patients by rTMS assessed with fMRI and dynamic causal modeling. NeuroImage 50(1):233–242
Rehme AK, Eickhoff SB, Wang LE, Fink GR, Grefkes C (2011) Dynamic causal modeling of cortical activity from the acute to the chronic stage after stroke. NeuroImage 55(3):1147–1158
Astafiev SV, Shulman GL, Stanley CM, Snyder AZ, Van Essen DC, Corbetta M (2003) Functional organization of human intraparietal and frontal cortex for attending, looking, and pointing. J Neurosci 23(11):4689–4699
Regnaux JP, David D, Daniel O, Smail DB, Combeaud M, Bussel B (2005) Evidence for cognitive processes involved in the control of steady state of walking in healthy subjects and after cerebral damage. Neurorehabil Neural Repair 19(2):125–132
Vicentini JE, Weiler M, Almeida SR, de Campos BM, Valler L, Li LM (2016) Depression and anxiety symptoms are associated to disruption of default mode network in subacute ischemic stroke. Brain Imaging Behav 11(6):1571–1580
Northoff G, Heinzel A, de Greck M, Bermpohl F, Dobrowolny H, Panksepp J (2006) Self-referential processing in our brain--a meta-analysis of imaging studies on the self. NeuroImage 31(1):440–457
Liu Y, D'Arceuil HE, Westmoreland S, He J, Duggan M, Gonzalez RG et al (2007) Serial diffusion tensor MRI after transient and permanent cerebral ischemia in nonhuman primates. Stroke 38(1):138–145
Assaf Y, Pasternak O (2008) Diffusion tensor imaging (DTI)-based white matter mapping in brain research: a review. J Mol Neurosci 34(1):51–61
Stinear CM, Barber PA, Smale PR, Coxon JP, Fleming MK, Byblow WD (2007) Functional potential in chronic stroke patients depends on corticospinal tract integrity. Brain 130(Pt 1):170–180
Morita N, Harada M, Uno M, Furutani K, Nishitani H (2006) Change of diffusion anisotropy in patients with acute cerebral infarction using statistical parametric analysis. Radiat Med 24(4):253–259
Schaechter JD, Fricker ZP, Perdue KL, Helmer KG, Vangel MG, Greve DN et al (2009) Microstructural status of ipsilesional and contralesional corticospinal tract correlates with motor skill in chronic stroke patients. Hum Brain Mapp 30(11):3461–3474
Matsuoka K, Yasuno F, Taguchi A, Yamamoto A, Kajimoto K, Kazui H et al (2015) Delayed atrophy in posterior cingulate cortex and apathy after stroke. Int J Geriatr Psychiatry 30(6):566–572
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Almeida, S.R.M., Castellano, G., Vicentini, J., Min, L.L. (2018). The Neuroimaging of Stroke: Structural and Functional Advances. In: Habas, C. (eds) The Neuroimaging of Brain Diseases. Contemporary Clinical Neuroscience. Springer, Cham. https://doi.org/10.1007/978-3-319-78926-2_4
Download citation
DOI: https://doi.org/10.1007/978-3-319-78926-2_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-78924-8
Online ISBN: 978-3-319-78926-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)