
Abstract
Larvae of ant-like stone beetles are exceptionally poorly known. Scydmaeninae comprises over 5000 species, but the immature stages have been described for less than 0.5% of them; the pupa has been illustrated for four species, and the first larval instar for only one species. In several tribes, larvae still remain unknown, and many descriptions are inaccurate or poorly illustrated. Chaetotaxic structures have been coded in only six species. Our knowledge of larval Scydmaeninae is so fragmentary that even for the largest, most common, and abundant genus, Euconnus Thomson (nearly 2500 nominal species!), the immature stages have never been adequately described. Known larvae of Scydmaeninae show a great diversity of body shapes and structures, more than expected within one subfamily of Staphylinidae. Known larvae of Eutheiini, Scydmaenini, Glandulariini, Mastigini, Clidicini, and Leptomastacini have a ten-segmented abdomen, whereas those of Cephenniini have only nine abdominal segments. Larvae of Eutheiini and Mastigitae are campodeiform, subcylindrical, or flattened, resembling those of other subfamilies of Staphylinidae. However, larvae of Scydmaenus s. str. are nearly onisciform, with demarcated laterotergites of thoracic segments, densely and asymmetrically covered with setae and microtrichia. Larvae of Glandulariini are also onisciform, but without demarcated laterotergites; they are sparsely setose and have nearly smooth cuticles. Larvae of some tribes have short, unsegmented urogomphs, whereas in others the urogomphs are absent. This chapter summarizes known data on the immature stages of Scydmaeninae, including their biology, indicating major problems and future directions.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Ant-like stone beetles were treated as a separate family, Scydmaenidae, for nearly 200 years until Grebennikov and Newton (2009) placed this group within Staphylinidae. This is a large subfamily, currently comprising over 5300 species classified into 1 extinct and 3 extant supertribes, including 10 tribes and over 100 genera. Within rove beetles, scydmaenines were hypothesized to represent a sister group of a clade Steninae + Euaesthetinae (Grebennikov and Newton 2009) or Solieriinae (Thayer et al. 2012; McKenna et al. 2015), although their true relationships remain unclear. Within Scydmaeninae, the monophyly of Cephenniitae was well supported, but the largest supertribe, Scydmaenitae, is almost certainly not monophyletic (Jałoszyński 2012a, 2014). Larval morphological characteristics are expected to help solve still problematic relationships within Scydmaeninae, but our knowledge of immature ant-like stone beetles is surprisingly scarce, considering there has been two centuries of research.
The published record of immature Scydmaeninae began with the most spectacular misidentification ever made regarding beetles. Tömösvary (1882) described a family Anisosphaeridae in Zygentoma (then Thysanura) to accommodate a new genus, Anisosphaera , an odd-looking arthropod discovered in Central Europe. Silvestri (1899) did not agree that Anisosphaera belonged to the same group as the silverfish and established a new insect order, Anisosphaeridia , discussing its possible affinities to springtails (Collembola). Tömösvary’s description and illustrations were fairly accurate, and Bergroth (1899) recognized in Anisosphaera a larva belonging to a common western Palaearctic scydmaenine genus, Cephennium Müller & Kunze (Cephenniini).
Larvae of merely 13 genera (out of over 100) have been described or illustrated so far, accounting for less than 0.5% of known species. Some figures, especially those in older publications, are highly simplified and inaccurate, and some identifications require verification. Therefore, even within these 13 genera whose larvae are known, only a few have been characterized with satisfactory accuracy. There is only one description based on immature stages obtained ex ovo by rearing adults; consequently, details of the life cycle are known for one species. Moreover, all immature stages, from egg to pupa, have only been described for one species (De Marzo 1983, 1984). Larvae of four species have been unambiguously identified by rearing, including two described on the basis of the shed last larval skin, obtaining the pupa and, subsequently, the adult (Jałoszyński 2013, 2016). All remaining descriptions were based on identifications made by associating larvae with adults collected in nature. Detailed descriptions of pupae have been published for only two species (De Marzo 1984; Jałoszyński 2012b); pupae of two more species are illustrated by photographs, showing only some details (Jałoszyński 2013, 2016). Details of the chaetotaxy were coded using modern methods for only six species (Wheeler and Pakaluk 1983; Jałoszyński 2013, 2015a, b, 2016; Jałoszyński and Kilian 2016). Not surprisingly, available data on larval biology are exceptionally scarce and mostly restricted to occasional observations of feeding on soil arthropods (Jałoszyński and Kilian 2012; Jałoszyński 2016).
2 Eggs
Only one species oviposited under laboratory conditions; its eggs have also been found in nature (De Marzo 1983). Eggs of Palaeostigus pilifer (Mastigini) are orange, elongate, 1.5 × 1.0 mm, laid in clusters of 2–9, and covered with a white, granulose secretion from the female’s abdominal gland. The eggs are relatively small in relation to the female’s body, which is about 6 mm long. According to De Marzo (1983), oviposition takes place in autumn; the eggs are deposited under stones, and larvae hatch a few days later. The number of eggs laid in one egg cluster is often larger than the number of ovarioles, which was found to be only two in each ovary in South African (Jałoszyński et al. 2015) and European (Jałoszyński, unpublished observations) species of Mastigini.
3 Larval Morphology
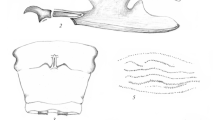
Larvae of Scydmaeninae (Figs. 14.1 and 14.2) show such a diversity of structures that to date it has not been possible to provide a clear-cut diagnosis of the subfamily based on larval morphology. The only constant structures shared by all known larvae are the labrum indistinguishably fused with frontoclypeus, broadly separated insertions of labial palps, and annuliform spiracles. The known range of structural diversity comprises onisciform (Fig. 14.1a) or campodeiform (Fig. 14.1c) body shape; head with or without demarcated “neck”; epicranial stem and frontal sutures present (Fig. 14.1e) or absent (Fig. 14.1f); antennae short and clubbed (Fig. 14.1a, b, f) or long and not thickened (Fig. 14.1c); dome-shaped, subconical, or even spatulate sensory appendage on antennomere II; nasale with or without teeth; maxillae with undivided mala or with distinct galea and lacinia; abdomen with ten (Fig. 14.1a–c) or nine segments (Fig. 14.1d); and urogomphs present or absent. Even the number of functional spiracles differs among taxa. Larval characteristics are therefore given for each tribe separately.

Scanning electron micrographs of Scydmaeninae larvae. (a) Mature larva of Scydmaenus tarsatus (Scydmaenini), ventral view. (b) Mature larva of Stenichnus sp. (Glandulariini), ventral view. (c) Mature larva of Palaeostigus sp. (Mastigini), dorsal view. (d) Mature larva of Cephennium sp. (Cephenniini), lateral view. (e) Head of Stenichnus sp. larva, anterodorsal view. (f) Head and prothorax of Cephennium sp. larva, anteroventral view. (g) Head of Scydmaenus tarsatus larva, lateral view. Abbreviations: abt1 abdominal tergite I, an1–3 antennomere I–III, est epicranial stem, frs frontal suture , gi glandular impression, lt laterotergite, msn mesonotum, mtn metanotum, mxp2–3 maxillary palpomere II–III, pnt pronotum, SA sensory appendage, st stemma

Scydmaeninae larvae and pupae, living individuals. (a–c) Larvae of Stenichnus spp. (Glandulariini) feeding on Oribatida. (d) Larva of Scydmaenus tarsatus (Scydmaenini) feeding on Ceratophysella springtail . (e) Larva of Scydmaenus rufus (Scydmaenini). (f) Larva of Palaeostigus palpalis (Mastigini) feeding on beetle pupa. (g) Larva of Stenomastigus longicornis (Mastigini). (h) Prepupa of Stenichnus godarti (Glandulariini). (i) Freshly emerged pupa of Stenichnus godarti. (j–k) Pupa of Stenichnus godarti shortly before emergence of the adult
3.1 Eutheiini
This tribe currently comprises six extant genera. Incomplete descriptions of larvae representing only two of these are available: Eutheia Stephens is illustrated for the European E. schaumi Kiesenwetter, whereas the larva of the sympatric E. linearis Mulsant & Rey was only mentioned as similar (Brown and Crowson 1980). An undetermined Nearctic species of Veraphis Casey has also been described (Newton 1991).
Larvae are strongly elongate and slender, nearly parallel-sided, and sparsely setose; pigmentation was not described. The head capsule is subrectangular or suboval. The epicranial suture in Eutheia was described by Brown and Crowson (1980) as indistinct, but the head was illustrated as entirely lacking stem and frontal arms of epicranial suture. In Veraphis , the epicranial stem is present, shorter than half the length of the head; frontal sutures are distinct but incomplete and obliterated anteriorly (Newton 1991). The anterior margin of the frontolabral region (nasale) has fine paired teeth; stemmata are absent. The antenna has short antennomere I, elongate antennomere II that broadens distally, and tiny antennomere III about as long as dome-shaped but broader than long accessory appendage of antennomere II. Mandibles are falciform, curved and pointed, with two tiny submedian mesal teeth ( Eutheia ) or several submedian mesal and dorsomesal denticles ( Veraphis ). Stipital projection of maxilla has two apical lobes, one short and the other several times longer, possibly representing galea and lacinia, both with sparse bristles. Maxillary palp for Eutheia was illustrated as two-segmented (Brown and Crowson 1980), which might have been a result of overlooking very short, annuliform palpomere I, illustrated for Veraphis (Newton 1991). In both genera, palpomere III is strongly elongate with rounded apex. Labial palp is short and composed of two palpomeres, of which II is distinctly longer than I, and has a rounded apex. Thoracic tergites were illustrated only for Eutheia (Brown and Crowson 1980) as subtrapezoidal (prothorax) or subrectangular (meso- and metathorax), transverse, and undivided. Abdomen is composed of ten segments; all abdominal segments in Eutheia are transverse; in Veraphis only tergites VIII–X were illustrated (Newton 1991), of which VIII and IX are strongly transverse and X weakly transverse. Urogomphs are present, composed of one segment fused with lateroposterior margin of abdominal segment IX, slightly upturned and pointed. Abdominal segment X is strongly elongate. Legs are short. Spiracles (presumably nine pairs) are present lateroventrally on mesothorax and abdominal segments I–VIII.
3.2 Cephenniini
Larval characters are known for 2 genera out of 19 currently recognized within this tribe. Immature stages of the European Cephennium carnicum Reitter, C. gallicum Ganglbauer, C. laticolle (Aubé), C. majus Reitter, and C. thoracicum (Müller & Kunze) have been adequately described and illustrated (Tömösvary 1882; Peyerimhoff 1899; Jeannel 1909; Scholz 1926; Ionesco 1937; Paulian 1941; Schuster 1966; Brown and Crowson 1980; Schmid 1988a, b; Jałoszyński and Beutel 2012). Additionally, illustrations of the general habitus and a few morphological details of an undetermined Japanese and Australian species of Cephennodes Reitter were published, respectively, as an unidentified genus by Morimoto and Hayashi (1986) and as Coatesia Lea, a junior synonym of Cephennodes , by Newton (1991).
Larvae (Fig. 14.1d, f) are onisciform; creamy white, yellowish, or brownish, suboval; and typically capable of curling into a compact ball (illustrated by Ionesco 1937). Setae are sparse, unmodified, and often very short. The head (Fig. 14.1f) is prognathous, strongly declined, and lacking stemmata. The epicranial stem, if present, is very short with frontal sutures variously distinct; nasale lacks teeth. The antenna has short antennomere I, long antennomere II that is broadest near the distal third or middle; and tiny antennomere III that is typically shorter (sometimes only slightly) than the asymmetrical, subconical, and pointed sensory appendage of antennomere II. Mandibles are short and subtriangular, with strongly curved distal portion and numerous denticles along outer margin. Maxillary mala forms a single protruding apical lobe with sparse bristles, maxillary palp with short palpomeres I and II, and elongate palpomere III. Prementum has one large median adhesive pad and a pair of small lateral suckers; labial palps are short, with elongate palpomere II. Ecdysial line on thoracic tergites is variously distinct (in Cephennium typically faint, barely discernible). Pronotum is extremely enlarged in Cephennium (Fig. 14.1d), usually much longer than meso- and metanotum combined, but less enlarged in Cephennodes ; meso- and metanotum are very short and strongly transverse.
Nine abdominal segments are visible. The abdomen has greatly enlarged tergite I in Cephennium (usually longer than all remaining tergites combined); it is unmodified in Cephennodes ; urogomphs absent; abdominal sternites are undivided. Legs are short and robust. Spiracles are annular, one pair ventrally on mesothorax and an unclear number on proximal abdominal segments. Ionesco (1937) listed four abdominal pairs of spiracles in Cephennium , but there are spiracles in small impressions located lateroventrally on segment I and laterodorsally on segments II–IV. At least on segments V–VII, in the same places, there are smaller, presumably rudimentary, spiracles lacking the surrounding impression (Jałoszyński, unpublished observations).
3.3 Glandulariini
Glandulariini is the largest tribe of Scydmaeninae, currently comprising 71 extant genera. Larval characters have been illustrated for only four genera: Stenichnus Thomson, Scydmoraphes Reitter, Neuraphes Thomson, and Euconnus Thomson. However, some species were identified only putatively.
The first truly modern description of scydmaenine larvae was given for a species of this tribe, with detailed mapping and coding of chaetotaxic characteristics (Wheeler and Pakaluk 1983). The study was focused on the larva of Nearcic Stenichnus turbatus (Casey) and became a standard description for comparative studies published later, with detailed redescriptions of the immature western Palaearctic St. collaris (Müller & Kunze) and St. godarti (Latreille) (Jałoszyński 2013, 2016). In all these papers, modern methods of coding chaetotaxic structures were applied and, for the first time, provided data on the serial and interspecific homology within Scydmaeninae.
Paulian (1941) was the first to present data on selected structures of an immature glandulariine species, St. collaris. Larvae of the same Stenichnus were also studied to a limited extent by Kasule (1966), Brown and Crowson (1980), and Schmid (1988a, b). Franz (1965) gave simplified illustrations of the immature St. gomerae Franz from the Canary Islands. Wheeler and Pakaluk (1983) described some characters of the Nearctic St. conjux (Casey), and Schmid (1988a) illustrated various structures of another European species, St. godarti (redescribed in detail by Jałoszyński 2016) and a putative larva of St. scutellaris (Müller & Kunze). Moreover, Brown and Crowson (1980) mentioned, but did not illustrate, larval mandibular structures similar to those of St. collaris but collected in Great Britain in association with adults of St. bicolor (Denny), and others collected with adults of St. pusillus (Müller & Kunze).
Other glandulariine species with larvae adequately described or illustrated include the western Palaearctic Scydmoraphes sparshalli Denny (Brown and Crowson 1980; Jałoszyński 2015b) and Sc. helvolus (Schaum) (Schmid 1988a). Brown and Crowson (1980) described a putative larva of European Neuraphes elongatulus (Müller & Kunze) and mentioned that it was similar to that of an undescribed but studied larva of N. angulatus (Müller & Kunze) (misspelled as angularis). Schmid (1988a) illustrated two different larvae putatively attributed to Neuraphes (s. str.) and Neuraphes ( Pararaphes ); Jałoszyński (2015b) demonstrated that the latter (“Typ 2-Larve” of Schmid) was in fact a misidentified Scydmoraphes . Finally, a putative larva of an Iberian Euconnus , presumably E. eppelsheimi Croissandeau, was partly described and illustrated by Brown and Crowson (1980). Structures similar to those found in the latter larva were also illustrated by Newton (1991) and putatively attributed to a Nearctic Euconnus sp. It should be noted, however, that “Euconnus” is a conglomerate of various genera; some subgenera have been recently removed from Euconnus and, consequently, older identifications without subgeneric placement should be treated as uncertain.
In the larvae of Stenichnus , it was recently found that a similar pattern of dorsal and lateral cephalic setae was present in taxa with very large or very small frontolabral region. Consequently, some setae located anteriorly to frontal sutures (i.e., on the frontolabral area) in one species may be homologous with setae located behind the sutures (i.e., on the epicranial halves) in another species. A hypothesis of “shifted” epicranial suture was proposed in order to draw homologies between groups of cephalic setae, an alternative approach to the traditional comparisons made strictly within the limits of head regions defined by the epicranial stem and frontal arms of the epicranial suture (Jałoszyński 2016).
Larvae (Figs. 14.1b and 14.2a–c) are typically onisciform or elongate suboval, yellowish to dark brown and glossy, sparsely to densely setose, and with setae unmodified. Head (Fig. 14.1e) is prognathous, weakly declined in living larvae, with one or three stemmata in a close group; epicranial stem and frontal sutures are distinct; nasale lacks teeth. The antenna has short antennomere I, long and clubbed antennomere II, which is usually broadest in the distal half, and small but not rudimentary antennomere III that is comparable in length to the long, subconical, typically slightly curved and pointed sensory appendage of antennomere II. Mandibles are falciform, slender, and pointed, with mesal margins often serrated. Maxillary mala is undivided and elongate, apically with bristles or spatulate setae; maxillary palp is long and usually with all palpomeres elongate, palpomere III being the longest. The prementum in some species has a pair of membranous adhesive pads or suckers that are difficult to notice, labial palps with short palpomere I, and long, slender palpomere II. The ecdysial line is typically distinct on all thoracic tergites; the pronotum is usually the longest. The abdomen is composed of ten segments; tergites I–IX are transverse and X usually elongate; abdominal sternites are undivided; urogomphs are absent. Legs are variously long; the pretarsus sometimes has ventral barbs. Spiracles are annular, ventral, or ventrolateral. There are nine pairs: one on the mesothorax and eight pairs on abdominal segments I–VIII, typically located on small protuberances. The spiracles on abdominal segment VIII are much smaller than the others, barely discernible, and nonfunctional .
3.4 Scydmaenini
Seven genera are known, but the larvae have been described only for Scydmaenus Latreille. Meinert (1888) was the first to describe immature Scydmaenus (s. str.) tarsatus Müller & Kunze (Europe). Additional (but partly misinterpreted) details were given by Brown and Crowson (1980), and larvae of the same species were redescribed in detail by Jałoszyński and Kilian (2012). Jałoszyński (2012b) later described and illustrated the pupa. Jeannel and Paulian (1945) gave some morphological details of the Afrotropical Scydmaenus (Pseudeumicrus) tachyoryctidis (Jeannel & Paulian), and Böving and Craighead (1931) illustrated larva of the Nearctic Scydmaenus (presumably s. str.) longicollis (Casey). Some structures of an undetermined Scydmaenus (presumably s. str.) from Panama were illustrated by Newton (1991). Beutel and Molenda (1997) illustrated the head of Scydmaenus sp. of unknown origin. Larvae of the European Scydmaenus ( Cholerus ) hellwigii (Herbst) and Scydmaenus ( Parallomicrus ) rufus (Müller & Kunze) were described and illustrated in detail by Jałoszyński and Kilian (2012) and Jałoszyński (2015a), respectively.
Larvae are nearly onisciform, broad and suboval ( Scydmaenus s. str.; Figs. 14.1a and 14.2d) or campodeiform, strongly elongate and nearly parallel-sided (subgenera Cholerus and Parallomicrus ; Fig. 14.2e), creamy white to yellowish brown, typically with dense and often asymmetrically distributed setae inserted on small papillae and with dense fine microtrichia on the head, thorax, and abdomen (especially in Scydmaenus s. str.). The head (Fig. 14.1g) is prognathous, with one pair of stemmata; epicranial stem and frontal sutures are distinct. The nasale has a row of several small teeth, often of unequal length and slightly asymmetrically distributed; sometimes teeth are located under the anterior margin of the labrum and not projecting in dorsal view. The antenna has short antennomere I; long and only slightly broadened apically antennomere II that is typically broadest near the apex (sometimes thickened indistinctly); and very small but distinct antennomere III that is shorter than the slightly asymmetrical sensory appendage of antennomere II, which can be subconical with blunt apex or barrel-shaped in the proximal half with a subconical apex. Mandibles are falciform and slender, pointed or blunt, with smooth mesal margin. Stipital projection of maxilla with two small apical lobes can be identified as lacinia and galea, both densely setose. The maxillary palp is long, with all palpomeres elongate; palpomere III is typically longest; prementum is distinctly emarginate anteriorly; labial palps have elongate palpomere I and II of various lengths. Thoracic tergites in onisciform larvae have fleshy laterotergites, each demarcated by a longitudinal groove; tergites are undivided in campodeiform larvae; ecdysial lines are not visible. Ten abdominal segments are present, with all except IX and X being transverse; segment X is elongate; abdominal sternites are undivided; urogomphi are absent. Legs are relatively short. Spiracles are annular and ventrolateral. One of the nine pairs is on the mesothorax and eight pairs are on abdominal segments I–VIII.
3.5 Clidicini
Three extant genera are known; the larva is described only for the Australian Clidicus abbotensis O’Keefe (O’Keefe and Monteith 2000).
The larva is campodeiform, strongly elongate, subparallel-sided and slightly flattened, whitish but with reddish brown head and tergal plates (however, O’Keefe and Monteith (2000) stated that “in life the larvae were the same bright color as the adults,” which are orange-reddish brown), moderately densely setose, and with unmodified setae. Head is prognathous, weakly declined, with demarcated short, annuliform “neck” and one pair of stemmata. Epicranial stem and frontal sutures are distinct; nasale lacks teeth. The antenna is longer than the head and very slender, not clubbed; long antennomeres I and II are of similar diameter; antennomere III is small but not rudimentary, strongly elongate, longer than strongly elongate, spatulate accessory appendage of antennomere II. Mandibles are falciform, moderately slender and pointed, each with one submedian mesal tooth. The maxillary mala is undivided and densely setose at apex; the maxillary palp is long, with all palpomeres elongate; palpomeres II and III are comparable in length. For labial palps, palpomere I is longer than II, inserted on a long palpifer resembling an additional palpomere. Thoracic tergites have distinct ecdysial lines. Ten abdominal segments are present; all except X are transverse; segment X is elongate; urogomphi are present, composed of one elongate segment fused with posterolateral margin of abdominal segment IX; abdominal sternites are not described. Legs are very long and slender. Spiracles are annular and ventrolateral; one of the nine pairs is on the mesothorax and eight pairs are on abdominal segments I–VIII.
3.6 Leptomastacini
Three extant genera are known; only larva of the European (Mediterranean) Leptomastax hypogaea Pirazzoli has been described (Vít and De Marzo 1989).
The larva is campodeiform, with subparallel and flattened body, whitish with testaceus head, and moderately densely setose; setae were illustrated and described as “rigid, spine-shaped.” The head is prognathous, unusually shaped, rhomboidal, strongly transverse, strongly narrowing anteriorly and posteriorly from the broadest place located near the middle, with demarcated annuliform “neck” and one pair of stemmata; epicranial stem and frontal sutures are distinct; nasale has two long robust teeth separated by deep U-shaped emargination . The antenna is only slightly longer than the head; antennomeres I and II are long and of similar length, but antennomere II is slightly broadened distally, being broadest near the apex; antennomere III is small but not rudimentary, strongly elongate, shorter than the very long, slightly asymmetrical, subconical accessory appendage, which is slightly bent near the middle. Mandibles are falciform and extremely long and slender, longer than the head or antennae, pointed, with smooth mesal margins. Stipital projection of maxilla has two lobes: subapical lobe is elongate and densely setose; apical lobe is broad and with two modified, thick setae. The maxillary palp is long, with all palpomeres elongate; palpomeres II and III are comparable in length; in labial palps palpomere II is longer than I. Thoracic tergites have distinct ecdysial lines, at least on pronotum and mesonotum. Ten abdominal segments are present, all except X are transverse; segment X is elongate. Urogomphi are present, composed of one elongate segment fused with the posterolateral margin of abdominal segment IX. Legs are long and slender, densely covered with relatively short, spiny setae. Spiracles are annular and lateral; one of the nine pairs is on the mesothorax and eight pairs are on abdominal segments I–VIII.
3.7 Mastigini
This tribe includes three extant genera; larvae are known for Palaeostigus Newton and Stenomastigus Leleup. De Marzo (1983, 1984) described eggs, all larval instars, and the pupa of the European Palaeostigus pilifer (Kraatz). Newton (1991) illustrated some structures of the Turkish Palaeostigus ruficornis schimitscheki (Machulka). Grebennikov and Newton (2009) illustrated some larval structures of the South African Palaeostigus bifoveolatus (Boheman). Most recently, Jałoszyński and Kilian (2016) gave a detailed description of the second- and third-instar larvae of the South African Stenomastigus longicornis (Boheman).
Mature larvae (Figs. 14.1c and 14.2f, g) are campodeiform, subparallel or with a strongly narrowing abdomen, flattened or subcylindrical . Membranous areas are whitish or yellowish, heavily sclerotized regions (head, tergal, and sternal plates) are variously dark brown to nearly black (Fig. 14.2f). Larvae of Stenomastigus are orange (Fig. 14.2g). The body is covered with sparse, long setae and dense asperities, typically granulose or pointed and forming patterns among smooth areas of tergal plates. Most setae are unmodified; additionally, short, flat, leaf-like setae with fine elongate ribs are distributed along anterior and posterior margins of tergal plates, often partly covered by folds of intersegmental membranes. The head is prognathous and slightly declined, lacking a demarcated “neck,” with one pair of stemmata. Epicranial stem and frontal sutures are distinct but short, together with antennal insertions shifted to the posterior half of the head capsule; nasale has a row of several short setae inserted on papillate protuberances. The head has a large glandular impression at the junction of the epicranial stem and frontal sutures, filled with or surrounded by modified setae to form a presumed evaporation apparatus. The antenna is much longer (often several times longer) than the head and very slender, not broadened; long antennomeres I and II are of similar diameter; antennomere II is subdivided into three sections; antennomere III is rudimentary, developed as a tiny, often barely discernible, papilla adjacent to the base of strongly elongate, slightly asymmetrical, subconical, and pointed accessory appendage. Mandibles are falciform, moderately slender, pointed, each with one submedian mesal tooth. The stipital projection of the maxilla is divided into two very short and broad, densely setose lobes, but it is unclear whether they represent lacinia and galea or one slightly emarginate mala. The maxillary palp is long, with palpomere I being short and II and III strongly elongate; labial palps have palpomere I longer than II. Thoracic tergites are in transparent mounts with a distinct ecdysial line , which in SEM images is visible as a smooth longitudinal median line among lateral fields of dense asperities. Ten abdominal segments are present, all except X (or IX and X) being transverse; segment X is elongate; urogomphi are absent. Sternal plates on the thorax and abdomen are reduced to small, paired (2–4), and setose sclerites. Legs are very long and slender , with particularly densely setose tibiotarsi. Spiracles are annular and lateral; one of the nine pairs is on the mesothorax and eight pairs are on abdominal segments I–VIII.
First instar larvae are known for Palaeostigus ; they differ from older instars in having a short, stout, and orange body with strikingly thick legs and antennae; setae on all body parts are very short and the frons lacks glandular impression.
Some data from the cited literature have been used by various authors in identification keys, reviews, and summaries, and some illustrations were redrawn and reproduced several times, often losing (or even gaining) details (e.g., Arnolde et al. 1964; Kasule 1966; Klausnitzer 1978, 1997; Newton and Franz 1998; O’Keefe 2005; Jałoszyński and Kilian 2016).
4 Biology of Larvae
Very little is known about the behavioral ecology or synecology of Scydmaeninae larvae. They can be found in moist leaf litter, soil, rotten wood, decomposing plant debris (compost, flood remains, rice straw, etc.), and sometimes also in ant and termite nests, typically together with adults and during all seasons, even in temperate climates. However, larvae of Scydmaenus species that inhabit compost can be most frequently collected during April to June in Central Europe, and mature larvae of southern Iberian Palaeostigus can be found only in winter and early spring (Jałoszyński, unpublished observations). Notable exceptions to finding larvae and adults together by sifting leaf litter are some species of South African Mastigini, especially Stenomastigus , whose adults climb tall grasses, bushes, and even trees and can be efficiently collected using a sweeping net or entomological umbrella; their larvae are associated with humid leaf litter, often along water bodies (Jałoszyński and Kilian 2016). However, a single observation (made by Rafał Ruta, University of Wrocław) known to the author was documented with photos showing a larva of Stenomastigus found in a South African forest climbing the moist trunk of a living tree during the day, half a meter above the ground. Larvae of Clidicus abbotensis have also been observed to show diurnal and exposed behavior similar to that of adults, “walking in close proximity to foraging adults and presumably seeking the same prey” (O’Keefe and Monteith 2000).
Larvae of Scydmaeninae kept under laboratory conditions typically hide among substrate particles, but rapid exposure to direct light (even a camera flash) does not disturb feeding in larvae of Glandulariini, Scydmaenini, and Mastigini (Jałoszyński, unpublished observation). All scydmaenine larvae observed alive by the author use their pygopod (i.e., abdominal segment X) extensively during walking to adhere to particles of substrate with the anal membrane. The pygopod is often used to stabilize the position of larva during feeding, by adhering to a substrate, leaving front and middle leg pairs free to manipulate prey (Fig. 14.2b).
Females of Palaeostigus pilifer cover their eggs with a glandular secretion during oviposition; larvae stay at the secretion deposit, and the second instars have been observed feeding on it (De Marzo 1983). Mature larvae of this species under laboratory conditions feed on pieces of dead caterpillars (De Marzo 1983); mature larvae of other European species of Palaeostigus kept in captivity readily accept beetle pupae (Fig. 14.2f), dead soft-bodied arthropods, and pieces of raw or boiled meat but were not observed hunting any living prey (Jałoszyński, unpublished observations).
Mature larvae of Stenichnus feed on armored mites (Oribatida) (Fig. 14.2a–c). They attack the gnathosoma, remove mite’s mouthparts, and feed through the resulting opening (Jałoszyński 2016). Mature larvae of Scydmaenus (Fig. 14.2d) show strong feeding preference for springtails, especially slow-moving Hypogastruridae, but were also observed feeding on Tomoceridae (Jałoszyński and Kilian 2012). Larvae of Cephennium were observed feeding on Oribatida or similarly heavily sclerotized Uropodina (Mesostigmata) by using their labial suckers to immobilize prey and short mandibles to slowly scrape through the mite’s cuticle (Schuster 1966; Schmid 1988b). The entire feeding process takes place through the resulting tiny hole, by injecting digestive juices and then ingesting liquefied tissues (Jałoszyński and Beutel 2012). In the larva of Cephennium galllicum , Jałoszyński and Beutel (2012) found a system of pharyngeal valves, presumably facilitating the flow of digestive juices and liquefied tissue between the predator’s mouth opening and the interior of prey.
When feeding, onisciform larvae of Cephenniini and Glandulariini (Fig. 14.2b) often curl around their prey; the unusual body shape may have evolved as an adaptation for such a feeding posture. Alternatively, the onisciform body of Cephennium larva e, their enlarged pronotum, and the first abdominal tergite enable them to curl into a tight ball, hiding the head, appendages, and ventrites inside (Ionesco 1937), a clearly defensive behavior that helps protect them against small predators .
5 Pupation and Morphological Structures of Pupae
Under laboratory conditions, pupation takes place either between substrate particles or on any available surface (e.g., on a plaster of Paris surface). In Stenichnus , the prepupa (Fig. 14.2h) shows a darker pigmentation than that in active larva. Pupae are initially lightly pigmented (Fig. 14.2i). They have very long, paired spines on the head, pronotum, and sides of abdomen; mature pupae are typically brownish (Fig. 14.2j, k). In Stenichnus godarti, the prepupal stage lasts 2 days and the pupal stage 7 days (Jałoszyński 2016).
6 Discussion
The larval stages of Scydmaeninae are exceptionally poorly studied, and most older descriptions and illustrations are inaccurate, lacking important details, or based on putatively identified material. Nevertheless, larval characters are now known for all large tribes. Only immature stages of small and monogeneric Chevrolatiini, Marcepaniini, and Leptoscydmini have yet to be found or obtained by rearing. Rearing, however, proved successful for only one species. A more promising strategy seems to be obtaining identifiable adults by rearing mature larvae collected in nature and describing morphological structures on the basis of shed larval skins (Jałoszyński 2013, 2016). Alternatively, larvae can be associated with adults using molecular markers.
Recent studies have provided novel data and allowed correction of some previous misidentifications or misinterpretations of morphological structures. Among the most interesting new results are those concerning the South African Stenomastigus , a genus closely related to the western Palaearctic Palaeostigus . In the previously known larvae of the European Palaeostigus pilifer, the first and second instars were orange (a pigmentation interpreted by De Marzo (1983) as coming from subcuticular deposits of an egg yolk) and spent all their time close to egg shells covered with a nutritious secretion , their only source of food. Only the third-instar larvae were capable of dispersal and feeding on other sources of food. Field observations in South Africa and morphological analysis of collected larvae of Stenomastigus longicornis demonstrated that two larval instars, presumably the second and third, were orange and capable of dispersal. Both were commonly collected by sifting leaf litter and also by finding them walking on the surface of soil particles (Jałoszyński and Kilian 2016; Jałoszyński, unpublished observations).
The known larvae, although representing merely a tiny fraction of nominal genera, show a greater diversity of body forms and structures than any other subfamily of Staphylinidae. The diversity is so great that it is not possible to give larval diagnostic characters to define Scydmaeninae, neither apomorphic nor a unique combination of synapomorphies. The most puzzling characteristic is the presence or absence of urogomphs in larvae of closely related tribes of Mastigitae, which otherwise show many similarities. The most enigmatic feature is the reduction in number of abdominal segments in Cephenniini from ten to nine. Moreover, three features of Mastigini are also unusual or unique among Staphylinidae: the antennomere II subdivided into three sections, antennae with very long spines, and the presumably glandular frontal impression (suggested by De Marzo 1983 to play a role in grooming). Future studies should be focused on the functional morphology and evolution of specialized modifications in Scydmaeninae larvae.
References
Arnolde LV, Bizova UB, Chilarov MS, Dolin VG, Gantiev RD, Zinovieva LA, Keleinikova SI, Kurcheva DF, Krivocheina NP, Mamaev BM, Medvedev LM, Tarsis VA, Chadaeva ZV, Sharova IC (1964) Opredeletelitel’ obitayushchikh v pochve lichinok nasekomykh (Key to soil-dwelling insect larvae). Izdatel’stvo ‘Nauka’, Moscow
Bergroth E (1899) Anisosphaera problematica Töm. = Cephennium, larva [Col.]. Bull Soc Ent France 295
Beutel RG, Molenda R (1997) Comparative morphology of selected larvae of Staphylinoidea (Coleoptera, Ployphaga) with phylogenetic implications. Zool Anz 41:392–394
Böving AG, Craighead FC (1931) An illustrated synopsis of the principal larval forms of the order Coleoptera. Ent Am 11:1–351
Brown C, Crowson RA (1980) Observations on Scydmaenid (Col.) larvae with a tentative key to the main British genera. Ent Mon Mag 115:49–59
De Marzo L (1983) Osservazioni sulla ovideposizione e sul ciclo larvale in Mastigus pilifer Kraatz (Coleoptera, Scydmaenidae). Entomologica (Bari) 18:125–136
De Marzo L (1984) Morfologia delle larve e pupa in Mastigus pilifer Kraatz (Coleoptera, Scydmaenidae). Entomologica (Bari) 19:61–74
Franz H (1965) Beitrag zur Bodenfauna der Kanarischen Inseln. Zur Kenntnis der Coleopterenfauna von Tenerife und La Gomera. Eos Rev Esp Ent 41:59–66
Grebennikov VV, Newton AF (2009) Good-bye Scydmaenidae, or why the ant-like stone beetles should become megadiverse Staphylinidae sensu latissimo (Coleoptera). Eur J Ent 106:275–301
Ionesco MA (1937) La larve de Cephennium carnicum Rtt, Etude morphologique. In: Hommage au Prof. E.C. Teodoresco, Bukarest, pp 101–109
Jałoszyński P (2012a) Beetles with ‘trochantelli’: phylogeny of Cephenniini (Coleoptera: Staphylinidae: Scydmaeninae) with focus on Neotropical genera. Syst Ent 37:448–477
Jałoszyński P (2012b) Morphology of pupa of Scydmaenus tarsatus Müller & Kunze (Coleoptera: Staphylinidae: Scydmaeninae). Genus 23(3):363–366
Jałoszyński P (2013) Mature larva of Stenichnus collaris (Müller & Kunze) (Coleoptera: Staphylinidae, Scydmaeninae). Zootaxa 3745(1):19–34
Jałoszyński P (2014) Phylogeny of a new supertribe Cephenniitae with generic review of Eutheiini and description of a new tribe Marcepaniini (Coleoptera: Staphylinidae: Scydmaeninae). Syst Ent 39:159–189
Jałoszyński P (2015a) Late-instar larva of Scydmaenus (Parallomicrus) rufus Müller & Kunze (Coleoptera: Staphylinidae, Scydmaeninae). Zootaxa 3973(2):579–590
Jałoszyński P (2015b) Redescription of late-instar larva of Scydmoraphes sparshalli (Denny) (Coleoptera: Staphylinidae, Scydmaeninae). Zootaxa 4032(5):582–594
Jałoszyński P (2016) Mature larva of Stenichnus godarti (Latreille) (Coleoptera: Staphylinidae, Scydmaeninae): redescription, hypothesis of displaced epicranial sutures and alternative interpretation of homology between chaetotaxic structures. Zootaxa 4196(1):77–94
Jałoszyński P, Beutel R (2012) Functional morphology and evolution of specialized mouthparts of Cephenniini (Scydmaeninae, Staphylinidae). Arthr Str Dev 41:593–607
Jałoszyński P, Kilian A (2012) Larval morphology of Scydmaenus tarsatus and S. hellwigii, with notes on feeding behavior and a review of bibliography on preimaginal stages of ant-like stone beetles (Coleoptera: Staphylinidae, Scydmaeninae). Eur J Ent 109:587–601
Jałoszyński P, Kilian O (2016) Description of the second- and third-instar larva of South African Stenomastigus longicornis (Boheman) (Coleoptera: Staphylinidae, Scydmaeninae). Zootaxa 4158(2):151–182
Jałoszyński P, Matsumura Y, Beutel RG (2015) Evolution of a giant intromittent organ in Scydmaeninae (Coleoptera: Staphylinidae): functional morphology of the male postabdomen in Mastigini. Arthr Str Dev 44:77–98
Jeannel R (1909) Biospeologica. X. Coléoptères (seconde série). Arch Zool Exp Gén 1:447–532
Jeannel R, Paulian R (1945) Mission scientifique de l’Omo, VI (57). Faune des terriers des Rats-Taupes. IV. Coléoptères. Mém Mus nat d’Hist nat 19:1–214
Kasule FK (1966) The subfamilies of the larvae of Staphylinidae (Coleoptera) with keys to the larvae of the British genera of Steninae and Proteininae. Trans Ent Soc London 118:261–283
Klausnitzer B (1978) Bestimmungstabellen für die Gattungen ausgewählter Käferfamilien. 5.8 Scydmaenidae. In: Klausnitzer B (ed) Ordung Coleoptera (larven). W. Junk, Hague
Klausnitzer B (1997) 20. Familie: Scydmaenidae. In: Klausnitzer B (ed) Die Larven der Käfer Mitteleuropas. Vol 4. Polyphaga, part 3. Goecke & Evers, Krefeld; Gustav Fischer Verlag, Jena
McKenna DD, Farrell BD, Caterino MS, Farnum CW, Hawks DC, Maddison DR, Seago AE, Short AEZ, Newton AF, Thayer MK (2015) Phylogeny and evolution of Staphyliniformia and Scarabaeiformia: forest litter as a stepping stone for diversification of nonphytophagous beetles. Syst Ent 40:35–60
Meinert F (1888) Scydmaenus-Larven. Ent Medd 1:144–150
Morimoto K, Hayashi N (1986) The Coleoptera of Japan in color, vol 1. Hoikusha, Osaka
Newton AF (1991) Scydmaenidae (Staphylinoidea). In: Stehr FW (ed) Immature insects, vol 2. Kendall/Hunt, Dubuque, IO, pp 330–334
Newton AF, Franz H (1998) World catalog of the genera of Scydmaenidae (Coleoptera). Kol Rundsch 68:137–165
O’Keefe ST (2005) Scydmaenidae Leach, 1815. In: Beutel RG, RAB L (eds) Handbook of zoology. Coleoptera, Beetles, vol 1: Morphology and systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). De Gruyter, Berlin
O’Keefe ST, Monteith GB (2000) Clidicus abbotensis O’Keefe, a new species of Scydmaenidae (Coleoptera: Staphylinoidea) from Australia with description of the larva. Mem Queensland Mus 46:211–223
Paulian R (1941) Les premiers états des Staphylinoidea (Coleoptera), Étude de morphologie comparée. Mém Mus nat d’Hist nat 15:1–361
Peyerimhoff P (1899) Description de la larve de Cephennium laticolle Aub. (Col.). Bull Soc Ent France 170–174
Schmid R (1988a) Die Larven der Ameisenkäfer (Scydmaenidae, Staphylinoidea) – Neu- und Nachbeschreibung mit einem vorläufigen Bestimmungsschlüssel bis zur Gattung. Mitt Badischen Landesv Naturk Naturs 14:643–660
Schmid R (1988b) Morphologische Anpassungen in einem Räuber-Beute-System: Ameisenkäfer (Scydmaenidae, Staphylinoidea) und gepanzerte Milben (Acari). Zool Jahrb Abt Syst Ökol Geogr Tiere 115:207–228
Scholz R (1926) Die Larve von Cephennium reitteri Bris. (Col.) Ent Blätter 22:103–104
Schuster R (1966) Scydmaeniden-Larven als Milbenräuber. Naturwiss 53:439–440
Silvestri F (1899) Prima nota intorno all Anisosphaera Töm. Ann Mus Civ St Nat Genova 39:613–619
Thayer MK, Newton AF, Chatzimanolis S (2012) Prosolierius, a new mid-Cretaceous genus of Solieriinae (Coleoptera: Staphylinidae) with three new species from Burmese amber. Cret Res 34:124–134
Tömösvary Ö (1882) Adatok hazánk Thysanura-Faunájához. Matem Termész Közl, Magyar Tudom Ak 18:119–131
Vít S, De Marzo L (1989) Description of the larva of Leptomastax hypogeus Pirazzoli (Coleoptera Scydmaenidae). Arch Sci 42:569–578
Wheeler QD, Pakaluk J (1983) Descriptions of larval Stenichnus (Cyrtoscydmus): S. turbatus and S. conjux, with notes on their natural history (Coleoptera: Scydmaenidae). Proc Ent Soc Washington 85:86–97
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Crown
About this chapter

Cite this chapter
Jałoszyński, P. (2018). Morphological Diversity of Immature Scydmaeninae. In: Betz, O., Irmler, U., Klimaszewski, J. (eds) Biology of Rove Beetles (Staphylinidae). Springer, Cham. https://doi.org/10.1007/978-3-319-70257-5_14
Download citation
DOI: https://doi.org/10.1007/978-3-319-70257-5_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-70255-1
Online ISBN: 978-3-319-70257-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)