
Abstract
The fossil record of Late Triassic tetrapods can be organized biostratigraphically and biochronologically into five, temporally successive land-vertebrate faunachrons (LVFs) that encompass Late Triassic time (in ascending order): Berdyankian, Otischalkian, Adamanian, Revueltian and Apachean. An up-to-date review of the age constraints on Late Triassic tetrapod fossil assemblages and correlation within the framework of the LVFs is presented. This makes possible a much more accurate evaluation of the timing of biotic events of Late Triassic tetrapod evolution, including: (1) Otischalkian, HO (highest occurrence) of almasaurids and chroniosuchians?, LOs (lowest occurrences) of crocodylomorphs and dinosaurs; (2) Adamanian, HO of mastodonsaurids and trematosaurids, LO of mammals; (3) Revueltian, HOs of capitosaurids, rhynchosaurs and dicynodonts; and (4) Apachean, HOs of metoposaurids, plagiosaurids and aetosaurs. The LO of turtles is Early Triassic or older, and the HO of phytosaurs is an Early Jurassic record. There is no compelling evidence of tetrapod mass extinctions at either the Carnian-Norian or the Triassic-Jurassic boundaries.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
10.1 Introduction
The Late Triassic was a major juncture in the evolution of terrestrial tetrapods (amphibians and reptiles) marked by many evolutionary events, including the oldest records of crocodylomorphs, pterosaurs, dinosaurs and mammals. It was also an interval of extinctions of some important tetrapod taxa, notably most of the temnospondyl amphibians and all of the crurotarsan archosaurs (“thecodonts” of older terminology). Much of the recent literature on Late Triassic terrestrial tetrapods takes a phylogenetic and taxonomic approach, which means it focuses on the origin, phylogenetic relationships and evolution of a particular taxon or taxa. Such an approach takes place within the context of phylogenetic (cladistic) analysis that dominates the discussion of the evolution of Late Triassic terrestrial tetrapods.
My approach here focuses on establishing the timing of biotic events in the evolution of Late Triassic terrestrial tetrapods. First, I review the temporal succession of Late Triassic terrestrial tetrapod assemblages to provide the most precise global correlation and temporal ordering possible. Within that framework, I then discuss key biotic (origination and extinction) events in the Late Triassic history of terrestrial tetrapods.
10.2 Temporal Framework
I organize the Late Triassic fossil record of terrestrial tetrapods within the temporal framework of land-vertebrate faunachrons (LVFs) developed by Lucas and Hunt (1993a), Lucas (1998, 2010) and Lucas et al. (2007a). This framework identifies five LVFs that encompass Late Triassic time (ascending order): Berdyankian, Otischalkian, Adamanian, Revueltian and Apachean (Fig. 10.1). All Late Triassic terrestrial tetrapod assemblages can be assigned to a LVF based on the genus-level taxa present in each assemblage, so the LVFs provided a means of ordering and correlating Late Triassic tetrapod assemblages that is independent of the standard global chronostratigraphic scale (SGCS, also sometimes called the “marine timescale”). Nevertheless, the cross correlation of the LVFs to the SGCS is important because it allows the tetrapod record to be correlated to physical events well constrained by marine biostratigraphy. However, that cross correlation is complicated, in places debatable, and at several points imprecise (Lucas and Heckert 2000; Lucas 2010).

The base of the Upper Triassic Series (base of the Carnian Stage) is defined by its GSSP (global stratotype section and point) at Prati di Stuori/Stuores Wiesen in northern Italy, with the primary signal the FAD (first appearance datum) of the ammonoid Daxatina canadensis (Mietto et al. 2012). Radioisotopic ages from Ladinian-age rocks indicate that the base of the Carnian is ~237 Ma (Mundil et al. 2010; Ogg 2012; Ogg et al. 2014; Lucas 2017b). However, direct correlation of the base of the Carnian to the LVFs is only possible in the Germanic basin, where the Carnian base is located in the middle Keuper (Gipskeuper) at about the base of the upper Grabfeld Formation (Bachmann and Kozur 2004; Kozur and Bachmann 2005, 2008). The Berdyankian tetrapod assemblage from the lower Keuper (Lettenkohle = Erfurt Formation) and the oldest Otischalkian tetrapod assemblage in the Schilfsandstein (Stuttgart Formation), which is “middle” Carnian in age, indicate the base of the Carnian correlates to a point in Berdyankian time. However, no tetrapod assemblage is known from the upper part of the Gipskeuper, so the tetrapod succession across the Ladinian-Carnian boundary is not clear in the Germanic basin.
The age of the base of the Otischalkian is well constrained as being during Carnian time, co-eval with the mid-Carnian wet episode, which is late Julian, and close in age to 231 Ma (e.g., Ruffell et al. 2016). Thus, the base of the Otischalkian in the Germanic basin is in the mid-Carnian Schilfsandstein, and that is the oldest possible age of the Otischalkian type assemblage from the basal Chinle Group in the western USA.
There is no agreed on GSSP for the base of the Norian, but that base will likely be defined using conodont biostratigraphy in western Canada or Sicily. The expectation is that the definition will be close to the longstanding working definition, which equates the base of the Norian to the base of the North American Stikinoceras kerri ammonoid zone (Orchard 2010, 2014). I have long advocated the equivalence or near equivalence of the base of the Norian and the beginning of the Revueltian LVF (Lucas 1998, 2010, 2015; Lucas et al. 2012). However, the “long Norian” of Muttoni et al. (2004, 2010), based on a magnetostratigraphic correlation, makes the base of the Norian much older (about 227–228 Ma) and places it within the Adamanian LVF. Lucas et al. (2012) provided a lengthy refutation of the “long Norian,” placing the base of the Norian close to 220 Ma, and that correlation is followed here (Fig. 10.1). However, numerical ages that constrain the age of the base of the Norian are mostly from detrital zircons and are fraught with inconsistencies and contradictions discussed at length by Lucas et al. (2012) and Lucas (2017b). Recent work by Kohút et al. (2017) provides detrital zircon ages from Central Europe that indicate the base of the Norian is close to 221 Ma, but more data are needed to resolve fully the numerical age of the Carnian-Norian boundary.
Like the Norian, the Rhaetian base has no agreed on GSSP, though it will be defined based on the FAD (first appearance datum) of the conodont Misikella posthernsteini. Currently, there are two candidate GSSP sections, at Steinbergkogel in Austria, and in the Lagonegro basin of southern Italy (Krystyn et al. 2007a, b; Rigo et al. 2016; Bertinelli et al. 2016). The Apachean LVF encompasses the Norian-Rhaetian boundary, based on conchostracan biostratigraphy, magnetostratigraphic correlations and the presence of the aetosaur Aetosaurus in Apachean strata (Lucas 2010; Weems and Lucas 2015). Thus, Lucas (2010) regarded the Apachean as late Norian-Rhaetian.
Magnetostratigraphy, palynostratigraphy, conchostracan biostratigraphy and vertebrate biostratigraphy indicate the beginning of the Wassonian LVF is very close to the base of the Hettangian (Kozur and Weems 2005, 2007, 2010; Lucas and Tanner 2007b; Cirilli et al. 2009). Thus, I regard the Apachean-Wassonian boundary as a good approximation of the Triassic-Jurassic boundary (Fig. 10.1).
10.3 Biofacies and Biases
During the Early and Middle Triassic, terrestrial tetrapod assemblages can be divided into amphibian dominated and dicynodont dominated (Lucas and Hunt 1993b). These two kinds of assemblages likely are biofacies that represent aquatic (including shallow marine), amphibian-dominated communities and terrestrial, dicynodont-dominated communities. This dichotomy mostly ends in Berdyankian time, when the dicynodont dominated assemblages largely disappear, though they do persist into the Adamanian in South America (see below). The different composition of the tetrapod assemblages of each biofacies makes difficult direct comparisons and correlation of assemblages of the two different biofacies.
Late Triassic terrestrial tetrapod assemblages have been collected and studied for nearly 200 years, going back to the earliest studies in the Germanic basin during the early 1800s. Nevertheless, it is clear that the temporal coverage of Late Triassic time provided by terrestrial tetrapod assemblages is patchy at best. Lucas (1997) advocated using the Chinle Group tetrapod assemblages from the western USA as the standard succession of Late Triassic tetrapod assemblages. Indeed, the Otischalkian, Adamanian, Revueltian and Apachean LVFs are based primarily on Chinle Group assemblages (Lucas and Hunt 1993a; Lucas 1998, 2010; Lucas et al. 2007a). But, the Chinle Group is a succession of fluvial strata no more than 600 m thick that encompasses most of Late Triassic time, 30 million years or more. Therefore, a priori, the Chinle succession must be riddled with hiatuses ranging from paraconformities to substantial unconformities (Lucas 1993; Lucas and Spielmann 2013). This means the succession of Chinle Group tetrapod assemblages provides “snapshots” of the Late Triassic record, not a continuous and complete succession of assemblages.
Indeed, numerical calibration of the Late Triassic LVFs indicates that they are about 2–10 million years long (Fig. 10.1). Thus, at the level of LVF, the temporal resolution is poor. Subdivision of the Adamanian and Revueltian LVFs has been proposed in the Chinle Group section (Hunt 1991; Hunt et al. 2005), but these subdivisions cannot readily be correlated to other non-Chinle assemblages. This means that plotting vectors of tetrapod evolution within each LVF is difficult to impossible, so the comparisons are between LVFs, not within them.
10.4 Late Triassic Terrestrial Tetrapod Record
Terrestrial tetrapod fossil assemblages have a broad paleogeographic distribution over what was Late Triassic Pangea (Fig. 10.2). Here, I review the composition and correlation (Figs. 10.3 and 10.4) of these assemblages.

Map of Late Triassic Pangea showing locations of principal tetrapod assemblages discussed in the text. Localities are: A = Argentina, B = Brazil, C = Chinle basin, western USA, G = Greenland, Ge = Germanic basin, I = Northern Italy, In = India, K = Karoo basin, South Africa, M = Madagascar, N = Namibia, Ne = Newark Supergroup, eastern North America, R = Russian Urals, U = United Kingdom

Correlation chart of Berdyankian-Adamanian tetrapod assemblages

Correlation chart of Revueltian-Apachean tetrapod assemblages
10.4.1 Berdyankian Tetrapod Assemblages
Berdyankian-age tetrapod assemblages are known from Russia (the assemblage characteristic of the LVF), Germany, Argentina, Brazil and Namibia (Figs. 10.2 and 10.3). The characteristic assemblage from the Bukobay Formation in the Russian Ural foreland basin includes an anthracosaur, the amphibians Mastodonsaurus, Bukobaja, Cyclotosaurus?, Plagioscutum and Plagiosternum, an erythrosuchid, a rauisuchid, and the dicynodonts “Elephantosaurus jachimovitschi” Vyushkov (a Stahleckeria-like form) and a generically indeterminate kannemeyeriid (e.g., Shishkin et al. 1995, 2000a, b; Ivakhhnenko et al. 1997; Gower and Sennikov 2000).
The Lettenkohle (Lettenkeuper, Lower Keuper, Erfurt Formation) in Germany and the Chanarian LVF localities in Argentina and Brazil are the principal correlatives of the Berdyankian type assemblage. The Lettenkohle record is important because it establishes the Ladinian age of at least part of the Berdyankian (see above). The Lettenkohle fossils are from the Grenze bonebed, the laterally equivalent/overlying Vitriolschiefer and the Kupferzell locality, so they are above the unconformity that separates the Keuper from the underlying Muschelkalk. Lettenkohle tetrapods include a chroniosuchian, the amphibians Mastodonsaurus giganteus, Callistomordax, Plagiosternum, Plagiosuchus and Kupferzella, the rauisuchian Batrachotomus, the prolacertiform Tanystropheus and small cynodonts (e.g., Wild 1978, 1980; Schoch 1997, 2000; Lucas 1999; Schoch and Werneburg 1999; Damiani et al. 2009; Gower and Schoch 2009).
The Chañares local fauna from the Ischichuca (formerly Chañares) Formation of the Ischigualasto-Villa Unión basin of northwestern Argentina includes various archosaurs such as Lagerpeton, Marasuchus and Chanaresuchus, the dicynodont Dinodontosaurus, the traversodontid Massetognathus, the chiniquodontid Chiniquodon and the probainognathid Probainognathus (Bonaparte 1970, 1997; Romer 1973; Sereno and Arcucci 1993, 1994; Lucas and Harris 1996; Arcucci and Marsicano 1998; Hsiou et al. 2002; Mancuso et al. 2014). Bonaparte (1966, 1967, 1982) based the Chanarian “provincial age” on this assemblage.
Marsicano et al. (2016) reported U-Pb ages on detrital zircons of ~236–234 Ma for the Chanarian tetrapod assemblage, which, on face value indicate that they are early Carnian. However, as they noted, the “samples contain complex age inventories” (Marsicano et al. 2016: 511), so the reliability of the reported ages are open to question. However, if accurate, these ages indicate that the Chanarian tetrapod assemblage is likely of early Carnian age.
The lower part of the Santa Maria Formation in the Paraná basin of Rio Grande do Sul, Brazil, yields vertebrate fossil assemblages from Candelaria and Chiniquá considered by Barberena (1977) and Barberena et al. (1985) to be two different local faunas of different ages. Lucas (2002, 2010) regarded them as a single biostratigraphic assemblage that includes a procolophonid, archosaurs, including Tarjadia (=Archaeopelta Desojo et al. 2011; Lucas et al. 2013), the dicynodonts Dinodontosaurus and Stahleckeria, chiniquodontids, and the traversodontids Massetognathus, Belesodon, Traversodon, Exaeretodon, Santacruzodo, Protuberum and Probelesodon (e.g., Abdala and Ribeiro 2003; Cisneros et al. 2004; Langer et al. 2007; Reichel et al. 2009). This assemblage and the Chanarian type assemblage in Argentina are assigned a Berdyankian age based largely on their dicynodonts and traversodontids and their stratigraphic position, which places them between tetrapod assemblages of Nonesian and Adamanian age.
The upper part of the Omingonde Formation in Namibia produces a tetrapod assemblage that includes an eryopoid temnospondyl, the dicynodonts Kannemeyeria, Dolichuranus, and Rhopalorhinus, a bauriamorph, and cynodonts (Keyser 1973a, b, 1978; Pickford 1995; Smith and Swartt 2002) that was long considered Perovkan in age (e.g., Lucas 2010). However, recent work has identified the cynodont Chiniquodon and the dicynodont Stahleckeria in the upper part of the Omingonde Formation (e.g., Abdala and Smith 2009; Abdala et al. 2013). This supports correlation to Berdyankian assemblages in South America, as detailed by Abdala et al. (2013).
Global correlation of tetrapod assemblages within the Berdyankian (Fig. 10.3) is problematic in part because of the biofacies problems outlined above. Thus, the Argentinian and Brazilian tetrapod assemblages are dicynodont-cynodont dominated and readily correlated to each other. The German and Russian assemblages are amphibian dominated and also readily correlated to each other. But, correlation of the tetrapod assemblages from the two biofacies to each other is less certain, though all of these assemblage do fall into a time interval between Perovkan and Otischalkian.
10.4.2 Otischalkian Tetrapod Assemblages
Otischalkian tetrapod assemblages are broadly distributed: in western and eastern North America, Germany, India, Morocco and possibly Kyrgyzstan (Figs. 10.2 and 10.3). The characteristic tetrapod assemblage of the Otischalkian is the assemblage of vertebrate fossils from just north of the defunct town of Otis Chalk in Howard County, Texas (Fig. 10.3). Lucas et al. (1993) and Long and Murry (1995) reviewed the fauna, which is from the Colorado City Formation of the Chinle Group. The following taxa are present: the amphibians Latiscopus, Buettneria and Apachesaurus, a procolophonid, the rhynchosaur Otischalkia, the archosaurs Doswellia, Trilophosaurus (=Malerisaurus) and Poposaurus, the aetosaurs Longosuchus (=Lucasuchus) and Coahomasuchus, the phytosaurs Parasuchus and Angistorhinus and the dinosaur Lepidus (Lucas et al. 1993; Long and Murry 1995; Heckert and Lucas 1999; Spielmann et al. 2006c; Nesbitt and Ezcurra 2015).
Otischalkian tetrapod assemblages are found across a broad geographic range of Chinle Group outcrops in Wyoming, New Mexico and Texas. The most well-known principal correlative of the type Otischalkian fauna in the Chinle Group is the vertebrate-fossil assemblage from the Popo Agie Formation of Wyoming (Branson and Mehl 1928; Mehl 1928; Colbert 1957; Lucas 1994; Lucas et al. 2002) that includes the metoposaurid Koskinonodon, the phytosaurs Parasuchus and Angistorhinus, the aetosaur Desmatosuchus, the archosaurs Poposaurus and Heptasuchus, the rhynchosaur Hyperodapedon, and the dicynodont Placerias. A less well-known principal correlative is the small assemblage from the Salitral Formation in Rio Arriba County, New Mexico, that consists of a metoposaur, Longosuchus, a phytosaur, and an indeterminate dinosaur (Lucas and Hunt 1992). Heckert (2004) provided some microvertebrate basis for recognition of the Otischalkian in Chinle Group strata, such as the LO of the “dinosaur” Protecovasaurus and the archosaur Trilophosaurus buettneri (also see Spielmann et al. 2008). Outside of the Chinle Group, Otischalkian assemblages are also known from the Newark Supergroup in eastern North America, the Germanic basin, Morocco, India and, possibly, Kyrgyzstan.
In the Newark Supergroup of eastern North America, the stratigraphically lower formations of the Deep River, Gettysburg, Newark and Fundy basins contain two distinct vertebrate fossil assemblages. The older of these was used by Huber et al. (1993b) as the basis of the Sanfordian LVF, after the characteristic assemblage from the Passaic (“Sanford”) Formation in the Sanford sub-basin of the Deep River basin complex (see Weems et al. 2016 for a revised lithostratigraphy of the Newark Supergroup used throughout this article). An age-equivalent assemblage from the Passaic Formation (Fundy basin) is also assigned to this LVF. The collective Newark tetrapod fauna of the Sanfordian LVF includes a metoposaurid, the trematosauroid Calamops, procolophonids, the traversodontids Arctotraversodon and Plinthogomphodon, the dicynodont Placerias, the rhynchosaur Hyperodapedon, the archosaur Doswellia, the aetosaurs Desmatosuchus and Longosuchus, indeterminate rauisuchians (“Zamotus”), the rauisuchian Postosuchus, the “sphenosuchian” Dromicosuchus, indeterminate phytosaur fragments and fragmentary dinosaur remains (e.g., Cope 1871; Olsen et al. 1989; Hunt and Lucas 1990; Huber et al. 1993a; Hunt 1993; Sues et al. 1994, 1999, 2003; Langer et al. 2000; Lucas et al. 2002; Peyer et al. 2008; Dilkes and Sues 2009; Sues and Schoch 2013). The Sanfordian correlates with the Chinle Group Otischalkian LVF based on the shared presence of Koskinonodon, Hyperodapedon, Desmatosuchus, Longosuchus, Doswellia, and Placerias.
In Germany, the Schilfsandstein (Stuttgart Formation) produces Metoposaurus and Parasuchus but lacks Stagonolepis, so it can be assigned an Otischalkian age (Hunt and Lucas 1991; Lucas 1999; Schoch and Werneburg 1999; Hungerbühler 2001; Kimmig and Arp 2010). Note that the Schilfsandstein is the age of the Carnian wet episode, which is “middle” Carnian (late Julian) in age (e. g., Hornung et al. 2007; Ruffell et al. 2016).
The 500-m-thick Irohalene Member of the Timesgadiouine Formation (interval T-5 of Dutuit 1966; Tixeront 1971) has produced most of the Late Triassic vertebrate fauna from Morocco. It contains the majority of vertebrate fossil localities described by Dutuit (1972, 1976, 1977, 1988, 1989a, b). Most of these occur in the lower part of the member, and they have produced a moderately diverse assemblage that includes the amphibians Almasaurus and Dutuitosaurus, the phytosaur Parasuchus, the aetosaur Longosuchus, the dicynodont Placerias (=Moghreberia, =Azarifeneria: Cox 1991; Lucas and Wild 1995) and the archosauromorph Azendohsaurus (Gauffre 1993; Lucas 1998; Jalil 1999; Flynn et al. 2010). Several of Dutuit’s (1976) localities are in the upper part of the Irohalene Member, which is a distinct faunal horizon that includes the amphibian Arganasaurus, the phytosaur Angistorhinus, and the dicynodont Placerias. The presence of Parasuchus, Angistorhinus, Longosuchus and Placerias supports assigning the Irohalene Member tetrapod assemblage(s) an Otischalkian age.
In the Pranhita-Godavari Valley of India, the basal Maleri Formation produces a tetrapod assemblage that includes the amphibian Metoposaurus, the rhynchosaur Paradapedon, the phytosaur Parasuchus, the archosaur “Malerisaurus,” an aetosaur, the theropod dinosaur Alwalkeria, a prosauropod (“cf. Massospondylus” of Kutty and Sengupta 1989), a large dicynodont, and the cynodont Exeraetodon (e.g., von Huene 1940; Jain et al. 1964; Roychowdhury 1965; Chatterjee 1967, 1974, 1978, 1980, 1982, 1987; Chatterjee and Roychowdhury 1974; Jain and Roychowdhury 1987; Bandyopadyhay and Sengupta 2006; Spielmann et al. 2006c; Kammerer et al. 2016). This is the only well-described Upper Triassic tetrapod assemblage from the Pranhita-Godavari Valley. It includes Parasuchus and Metoposaurus, taxa indicative of an Otischalkian age.
Northeast of the Pranhita-Godavari valley, in the Rewa basin of India, the Tiki Formation yields an Otischalkian tetrapod assemblage. This includes Metoposaurus, Parasuchus, the rhynchosaur Hyperodapedon, the supposed agamid lizard Tikiguana, the cynodont Rewacodon and the supposed mammals Tikitherium and Gondwanodon (e.g., Datta and Das 1996; Datta 2005; Mukherjee et al. 2012).
In the Fergana basin of Kyrgyzstan, the upper part of the Madygen Formation yields a tetrapod assemblage usually assigned a Ladinian-Carnian age based on the associated paleoflora (e.g., Dobruskina 1995a). Voigt et al. (2017) recently reported a U-Pb age of 237 ± 2 Ma on six concordant zircons from a pyroclastic bed stratigraphically below the tetrapod assemblage, which suggests the tetrapods may be of Carnian age. However, the entire tetrapod assemblage from the Madygen Formation is endemic and of no value to biostratigraphy. It consists of the amphibian Triassurus, the reptiliomorph Madygenerpeton, the unusual diapsids Sharovipteryx, Kyrgyzsaurus and Longisquama and the cynodont Madygenia (Sharov 1970, 1971; Ivakhnenko 1978; Tatarinov 2005; Unwin et al. 2000; Schoch et al. 2010; Alifanov and Kurochkin 2011). The assemblage may be of Otischalkian age, but more precise data are needed to confirm this.
10.4.3 Adamanian Tetrapod Assemblages
The characteristic tetrapod assemblage of the Adamanian is the assemblage of vertebrate fossils found in the Blue Mesa Member of the Petrified Forest Formation in the Petrified Forest National Park, near the defunct railroad siding of Adamana, Arizona (Fig. 10.3). Recent faunal lists have been provided by Murry and Long (1989), Long and Murry (1995), Heckert et al. (2005) and Parker et al. (2006). The fauna includes the following tetrapods: the amphibians Apachesaurus and Koskinonodon, the aetosaurs Desmatosuchus (=Acaenasuchus), Stagonolepis, Adamanasuchus and Paratypothorax, Rutiodon-grade phytosaurs (including Leptosuchus and Smilosuchus), the rauisuchian Postosuchus, the archosaurs Hesperosuchus, Acallosuchus, Parrishea and Vancleavea, and the dicynodont Placerias, as well as many microvertebrate taxa.
Besides the Chinle Group correlatives, major Adamanian faunas are those of the Conewagian interval of the Newark Supergroup basins of eastern North America; Lossiemouth Sandstone Formation, Scotland; Lehrberg Schichten-Obere Bunte Mergel interval of the German Keuper; the Krasiejów locality in Poland; Ischigualasto Formation, Argentina; and upper Santa Maria Formation and Caturitta Formation, Brazil (Fig. 10.3).
In the Chinle Group, Adamanian vertebrates are widespread and include the vertebrate fossil assemblages of the Placerias and Downs’ quarries, Bluewater Creek Formation, Arizona (Camp and Welles 1956; Kaye and Padian 1994; Long and Murry 1995; Lucas et al. 1997; Heckert 2004; Heckert et al. 2005); the Bluewater Creek Formation and Blue Mesa Member of the Petrified Forest Formation in the Blue Hills, Arizona; the Bluewater Creek Formation and Blue Mesa Member of the Petrified Forest Formation, McKinley and Cibola counties, New Mexico (Heckert 1997); the Los Esteros and Tres Lagunas members, Santa Rosa Formation, and the Garita Creek Formation in the vicinity of Lamy, Santa Fe County, New Mexico (Hunt et al. 2005; Lucas et al. 2010); Garita Creek Formation, Santa Rosa and vicinity, Guadalupe County, New Mexico (Hunt and Lucas 1993a); and Tecovas Formation, West Texas (Murry 1986, 1989; Long and Murry 1995; Lucas et al. 2016).
The fauna at the Placerias and Downs’ quarries has been discussed by Kaye and Padian (1994), Long and Murry (1995), Lucas et al. (1997) and Heckert (2004). It includes the amphibians Koskinonodon and Apachesaurus, the prolacertiform Gwyneddosaurus (=Tanytrachelos), the phytosaurs Parasuchus and Rutiodon/Leptosuchus, the aetosaurs Stagonolepis and Desmatosuchus (=Acaenasuchus), the rauisuchid Postosuchus, the archosaurs Trilophosaurus, Acallosaurus, Poposaurus, Chatterjeea, Hesperosuchus, Tecovasaurus and cf. Uatchitodon, an indeterminate ceratosaur and the dicynodont Placerias.
A U-Pb age recently reported from Chinle Group strata at the Placerias/Downs’ quarries by Ramezani et al. (2014) is not consistent with earlier published ages. This is an age of 219.39 ± 0.16 Ma from near the base of the Chinle Group at the Placerias fossil locality in Arizona. Stratigraphic position puts this age well below a series of ages in the 220–227 Ma range reported by Ramezani et al. (2011) and Atchley et al. (2013). To explain this contradiction, Ramezani et al. (2014) claim massive lateral facies changes in the lower Chinle lithosome, and even conclude that “geochronological correlation independent of conventional stratigraphic methods [lithostratigraphy, biostratigraphy] is the only viable means for deciphering the depositional history of rock similar to the Chinle Formation” (p. 995). I prefer instead to rely on a century of geologic mapping, detailed lithostratigraphic analysis and the biostratigraphy of palynomorphs, conchostracans and vertebrates (e.g., Heckert and Lucas 2002 and references cited therein) that demonstrates that the Placerias quarry numerical age of Ramezani et al. (2014) is stratigraphically below many older numerical ages. Thus, the Placerias quarry age published by Ramezani et al. (2014) is anomalously young, likely due to postcrystallization lead loss, and should be ignored.
The following tetrapod taxa are known from the Los Esteros Member, Santa Rosa Formation, near Lamy, New Mexico: the amphibian Apachesaurus, the phytosaurs Rutiodon and Angistorhinus, the aetosaurs Desmatosuchus, Tecovasuchus and Stagonolepis and the dicynodont cf. Ischigualastia (Hunt and Lucas 1993a, 1994; Hunt et al. 2005; Heckert et al. 2007). The overlying Garita Creek Formation contains the following taxa: the amphibian Koskinonodon, phytosaurs, rauisuchians, and the aetosaurs Desmatosuchus, Stagonolepis and Paratypothorax (Hunt et al. 2005; Lucas et al. 2010).
The Tecovas Formation of West Texas yields the following tetrapod taxa: the amphibians Koskinonodon and Apachesaurus, the probable tetrapod Colognathus, the archosauromorphs Trilophosaurus, Spinosuchus, Parrishea, Tecovasaurus, and Crosbysaurus, the phytosaurs Rutiodon, Leptosuchus and Smilosuchus, the aetosaurs Desmatosuchus and Stagonolepis, the rauisuchian Postosuchus, and the oldest known mammal, Adelobasileus (Lucas and Luo 1993; Lucas et al. 1994, 2016; Long and Murry 1995; Spielmann et al. 2008, 2013).
In the Deep River basin of North Carolina, an assemblage of the Conewagian LVF from the Cumnock Member of the Lockatong Formation (cf. Weems et al., 2016) is superposed on the characteristic Sanfordian assemblage. Conewagian assemblages are characterized by the tetrapod assemblage in the basal Lockatong (=“Gettysburg”) Formation (Kozur and Weems 2010) along Little Conewago Creek in south-central Pennsylvania (Gettysburg basin: Huber et al. 1993b; Sullivan et al. 1995; Lucas and Sullivan 1997) and also are known from the Lockatong (=“Cow Branch”) Formation (Dan River basin), and upper Stockton and Lockatong formations (Newark basin). The most widespread and characteristic Conewagian tetrapod is the phytosaur Rutiodon, which co-occurs with the amphibian Koskinonodon, archosaurs of uncertain affinity, the problematic reptile Colognathus, an aetosaur (Desmatosuchus), one or more “ornithischian dinosaurs” (e.g., Pekinosaurus, Crosbysaurus, Revueltosaurus and Galtonia), the archosaurs Uatchitodon and Gwyneddosaurus (=Tanytrachelos), lepidosaurs, including the single record of the gliding lepidosauromorph Icarosaurus and cynodonts, including aff. Boreogomphodon and Microconodon (e.g., Emmons 1856; Olsen 1980, 1988; Olsen et al. 1989; Sues 1992; Huber et al. 1993a; Hunt 1993; Hunt and Lucas 1994; Doyle and Sues 1995; Lucas and Huber 2003; Heckert et al. 2012). Conewagian assemblages correlate with the Adamanian LVF of the Chinle Group, based on the shared presence of Koskinonodon, Colognathus, Uatchitodon, Rutiodon and other Rutiodon-grade phytosaurs (Smilosuchus of Long and Murry 1995), Desmatosuchus and broadly similar “ornithischian dinosaurs” (e.g., Murry and Long 1989; Lucas et al. 1992, 1997, 2016; Huber et al. 1993b; Hunt 1993; Hunt and Lucas 1994; Heckert 2004; Heckert et al. 2012).
The tetrapod assemblage of the Lossiemouth Sandstone Formation of Grampian (Elgin) Scotland comes from small quarries and the coastal section at Lossiemouth. Benton and Spencer (1995; also see Fraser 2006) provided a detailed summary that indicates that all sites come from a narrow stratigraphic interval, so they are a single biostratigraphic assemblage. It includes the procolophonid Leptopleuron, the sphenodontid Brachyrhinodon, the rhynchosaur Hyperodapedon, the aetosaur Stagonolepis, the ornithosuchid Ornithosuchus, the crocodylomorph Erpetosuchus, the probable ornithodiran Scleromochlus and the “dinosaur” Saltopus. The presence of Hyperodapedon and Stagonolepis supports assigning this assemblage an Adamanian age.
In Germany, strata in the interval between the Schilfsandstein and the Stubensandstein (Lehrberg Schichten, Blasensandstein and Kieselsandandstein) produce Stagonolepis, Parasuchus and Metoposaurus (e.g., Lucas 1999), and are assigned an Adamanian age (Kozur and Weems 2005).
Kear et al. (2016) recently identified a temnospondyl skull fragment as Cyclotosaurus from the DeGeerdalen Formation in Svalbard. These are marine strata assigned a middle-late Carnian age based on ammonoids. Thus, if correctly identified, this would be the oldest record of Cyclotosaurus, which is otherwise known from Revueltian (Norian) strata.
The Polish fossil record of Late Triassic tetrapods advanced greatly during the 1990s, when scientific study of the extensive bonebed in the Krasiejów clay pity near Ople began, and much has been published since (see especially the reviews by Dzik and Sulej 2007; Szulc et al. 2015). The Krasiejów tetrapod assemblage includes the amphibians Cyclotosaurus and Metoposaurus, the phytosaur Parasuchus, the aetosaur Stagonolepis, the rauisuchian Teratosaurus and the dinosauriform Silesaurus (Dzik 2001, 2003; Sulej 2002, 2005, 2007, 2010; Sulej and Majer 2005; Dzik and Sulej 2007; Lucas et al. 2007b ; Szulc et al. 2015; Lucas 2015). This assemblage is from strata ~80 m above the Reed Sandstone (a Schilfsandstein equivalent) that are homotaxial to the German Lehrberg Schichten and is of Adamanian age (Lucas 2015).
In Argentina, the Ischigualasto Formation is 500–900 m thick and consists of drab mudstones, tuffs and sandstones that produce an extensive tetrapod assemblage, including: the amphibian Promastodonsaurus, the archosaurs Saurosuchus, Sillosuchus, and Proterochampsa, the ornithosuchid Venaticosuchus, the aetosaur Stagonolepis (=Aetosauroides), the rhynchosaur Hyperodapedon, the dinosaurs Herrerasaurus (=Ischisaurus? =Frenguellisaurus), Eoraptor and Pisanosaurus, the chiniquodontid cynodont Chiniquodon, the gomphodont cynodonts Exeraetodon, Proexaraetodon, and Ischignathus and the dicynodont Ischigualastia (e.g., Cabrera 1944; Reig 1959, 1961, 1963; Casamiquela 1960, 1962; Cox 1965; Bonaparte 1976, 1997; Rogers et al. 1993; Sereno et al. 1993; Alcober and Parrish 1997; Heckert and Lucas 2002; Baczko and Ezcurra 2013). This assemblage was the basis of the Ischigualastian land-vertebrate “age” of Bonaparte (1966, 1967, 1982).
Martínez et al. (2013) reviewed the stratigraphic distribution of the tetrapod fossils in the Ischigualasto Formation to show that most are from the lower 300 m of the formation. They also reviewed the radioisotopic ages associated with the Ischigualasto Formation tetrapods (e. g., Valencio et al. 1975; Rogers et al. 1993; Currie et al. 2009) to assign them an age range of ~226–231 Ma. This suggests a late Carnian (Tuvalian) age, which fits well with the conclusion that the Ischigualasto tetrapods are of Adamanian age (Lucas 2010; Lucas et al. 2012).
In northwestern Argentina, the Puesto Viejo Group contains a tetrapod assemblage that includes Kannemeyeria and Cynognathus and has long been considered to be of Nonesian (late Olenekian) age (e. g., Lucas 2010). However, Ottone et al. (2014) reported a SHRIMP 238 U/206Pb age of 235.8 ± 2.0 Ma from a rhyolitic tuff in the approximate middle of the Puesto Viejo succession. They accepted this numerical age as evidence of the Carnian age of the Puesto Viejo tetrapod assemblage. Instead, it is much more likely that the age reported by Ottone et al. (2014) is simply incorrect (too young) and does not indicate that Nonesian tetrapods (including the classic Cynognathus Assemblage Zone of the South African Karoo) are of Late Triassic age.
In Brazil, the principal Upper Triassic vertebrate assemblage from the Santa Maria Formation is from the vicinity of Santa Maria City. This is the Rhynchocephalia assemblage zone of Barberena (1977) or the Scaphonyx assemblage of Barberena et al. (1985), from the upper part of the Santa Maria Formation. The assemblage consists of abundant fossils of the rhynchosaur Hyperodapedon and the aetosaur Stagonolepis (=Aetosauroides); traversodontids, proterochampsids; the archetypal rauisuchian Rauisuchus and the primitive dinosaur Staurikosaurus (Barberena et al. 1985; Lucas 2002; Lucas and Heckert 2001; Abdala et al. 2001; Langer et al. 2007; Raugust et al. 2013; Melo et al. 2015). Clearly, the presence of Scaphonyx and Stagonolepis supports correlation with the vertebrates of the Ischigualasto Formation in Argentina, and therefore an Adamanian (=Ischigualastian) age (Lucas and Heckert 2001; Heckert and Lucas 2002; Lucas 2002, 2010).
Abdala et al. (2001) identified a “traversodontid biozone” (later termed the Santacruzodon Assemblage Zone) intermediate between the Dinodontosaurus and Hyperodapedon assemblages of the Santa Maria Formation. This biozone yielded the cynodont Menadon, also found in the Isalo II strata of Madagascar (Melo et al. 2015). Abdala et al. (2001) and Melo et al. (2015) consider the Santacruzodon Assemblage Zone to be of Ladinian or early Carnian age. However, correlation to Isalo II suggests an age of Adamanian, which is late Carnian. Furthermore, there is no lithostratigraphic basis for placing the so-called Santacruzodon Assemblage Zone between the Dinodontosaurus and Hyperodapedon assemblages (cf. Langer et al. 2007). In effect, the Santacruzodon Assemblage Zone is a hypothetical biostratigraphic construct based on a single locality that is the same age as the Hyperodapedon assemblage of the Santa Maria Formation. The Santacruzodon Assemblage Zone should be abandoned.
The tetrapod assemblage of the Caturrita Formation, which overlies the Santa Maria Formation, includes a mastodonsauroid amphibian, the procolophonid Soturnia, the sphenodont Clevosaurus, the lepidosaur Cargninia, the rhynchosaur Hyperodapedon, the proterochampsid Proterochampsa, the supposed pterosaur Faxinalipterus (but see Dalla Vecchia 2013), the dinosaurs Unaysaurus and Guaibisaurus, the dinosauriform Saccasaurus, a phytosaur, the cynodonts Exaeretodon and Riograndia, the dicynodont Ischigualastia (=Jachaleria) and diverse cynodonts (Araújo and Gonzaga 1980; Barberena et al. 1985; Dornelles 1990; Bonaparte et al. 1999, 2001, 2007, 2010a, b; Cisneros and Schultz 2003; Kischlat and Lucas 2003; Leal et al. 2003; Ferigolo and Langer 2006; Bonaparte and Sues 2006; Langer et al. 2007; Dias-da-Silva et al. 2009; Soares et al. 2011).
Philipp et al. (2013), in an extended abstract, reported U-Pb ages on 13 zircons from an ash bed (though Schultz et al. 2016 referred to these as “detrital zircons”) that form an isochron of 236 ± 3.5 Ma. This ash bed is approximately at the stratigraphic level of the “Santacruzodon assemblage,” so it suggests an age close to the Ladinian-Carnian boundary between the Berdyankian and Adamanian tetrapod assemblages of the Santa Maria Formation. Further publication of the analytical data associated with these ages is needed to assess their accuracy.
Most South American workers (e.g., Bonaparte 1982; Barberena et al. 1985; Langer 2005a, b; Rubert and Schultz 2004; Dias-da-Silva et al. 2007; Langer et al. 2007; Soares et al. 2011) advocate dividing the Brazilian Upper Triassic tetrapod succession into two biostratigraphically distinct assemblages largely based on their judgment that the dicynodonts Jachaleria and Ischigualastia are not the same taxon. They, therefore, correlate the Brazilian Caturrita Formation to the Argentinian Los Colorados Formation. Langer (2005b) also claimed that the Ischigualastian = Otischalkian + Adamanian, largely based on not recognizing the temporal range of Hyperodapedon as longer than the temporal range of the Ischigualastian. I do not accept either evaluation of the Brazilian Upper Triassic tetrapod biostratigraphy (Lucas 2002, 2010). Thus, I regard the Caturitta Formation as a correlative of the Ischigualasto Formation, so it is Adamanian, of late Carnian age.
In the Pranhita-Godavari Valley of India, the upper vertebrate fossil assemblage from the Maleri Formation is stratigraphically above the lower assemblage, but its stratigraphic range is not clear. This upper assemblage includes an aetosaur, prosauropods and a large dicynodont. Chigutisaurid amphibians (Compsocerops and Kuttycephalus: Sengupta 1995) and a “Rutiodon-like” phytosaur are also present (Bandyopadyhay and Sengupta 2006). Therefore, this assemblage may be Adamanian, but needs further documentation.
In western Madagascar, the Isalo group (“Groupe d l’Isalo” of Besarie 1930; also see Besarie and Collignon 1960, 1971) has long been divided into Isalo I, Isalo II and Isalo III based on perceived geologic age. The Isalo II strata yield Late Triassic tetrapods, including metoposaurs, sphenodontids, phytosaurs, dinosaurs, the rhynchosaur Hyperodapedon, the aetosaur Desmatosuchus, the archosaur Azendohsaurus, various cynodonts (including the traversodontids Dadadon and Menadon) and a dicynodont (Guth 1963; Westphal 1970; Dutuit 1978; Buffetaut 1983; Flynn et al. 1999, 2000, 2008; Lucas et al. 2002; Flynn and Wyss 2003; Burmeister et al. 2006; Kammerer et al. 2010; Ranovharimanana et al. 2011). The stratigraphic range of the Isalo II tetrapods is about 1200 m, but the rhynchosaur Hyperodapedon is one of the stratigraphically lowest taxa in the assemblage. This means the Isalo II assemblage is no older than Otischalkian and, based on the Desmatosuchus record, likely to be Adamanian. Flynn and collaborators (e.g., Flynn and Wyss 2003) advocate an older age, perhaps as old as Ladinian, for the Isalo II assemblage, but no data support that conclusion.
10.4.4 Revueltian Tetrapod Assemblages
The characteristic tetrapod assemblage of the Revueltian is that of the Bull Canyon Formation in east-central New Mexico (Fig. 10.4), and the following taxa are present: the amphibian Apachesaurus, the turtle Chinlechelys, the phytosaur Pseudopalatus and other Pseudopalatus-grade phytosaurs, the aetosaurs Rioarribasuchus, Paratypothorax, Typothorax coccinarum and Aetosaurus, the suchian Revueltosaurus, the “dinosaur” Lucianosaurus, the rauisuchian Postosuchus, the chatterjeeids Shuvosaurus (=Effigia) and Chatterjeea, the sphenosuchian Hesperosuchus and the cynodont Pseudotriconodon (e.g., Hunt 1994, 2001; Lucas et al. 2001; Joyce et al. 2009).
In the Chama basin of north-central New Mexico, the Petrified Forest Formation of the Chinle Group also yields Revueltian tetrapods, especially from the Snyder, Hayden and Canjilon phytosaur-dominated bonebeds (Sullivan and Lucas 1999; Zeigler et al. 2003; Heckert et al. 2005; Ezcurra 2006; Nesbitt and Stocker 2008). In northern Arizona, two Chinle Group units, the Painted Desert Member of the Petrified Forest Formation and the overlying Owl Rock Formation, have produced numerous Revueltian fossils, especially from the Petrified Forest National Park and from localities on Ward’s Terrace north of Flagstaff (e.g., Kirby 1989, 1991, 1993; Heckert et al. 2005; Spielmann et al. 2007).
Outside of the Chinle Group, Revueltian tetrapod assemblages are known from the Newark Supergroup in eastern North America, Greenland, the Germanic basin, (including eastern Poland), northern Italy, India, Argentina and South Africa (Fig. 10.4). In eastern North America, the provincial Neshanician LVF is based on a limited fossil assemblage typified by the aetosaur Aetosaurus arcuatus (Lucas et al. 1998; Lucas and Huber 2003). This taxon is present in the Passaic Formation in the Durham sub-basin of the Deep River basin, the Newark Basin (range zone: Warford through Neshanic members of the lower Passaic Formation), and the middle Sugarlof Member of the Passaic Formation of central Connecticut. Other vertebrates from the Neshanician LVF include indeterminate metoposaurid and phytosaur teeth, skull and scute fragments (e.g. “Belodon validus”), a rauisuchian, a crocodylomorph, a traversodontid and a sphenodontid. The dominance of the primitive neopterygian Semionotus sp. over other fish taxa is a trend also apparent in age-equivalent strata of the Chinle Group and German Keuper (Huber et al. 1993c; Lucas and Huber 2003).
The Cliftonian LVF is based on a low-diversity assemblage defined by the distribution of the procolophonid Hypsognathus fenneri. This taxon is common in the type area, from the middle (?Mettlars Member) to the upper (?Member TT) Passaic Formation of the northern Newark basin (e.g., Baird 1986). It is also known from the upper Passaic Formation of the Hartford basin, central Connecticut, and the basal Blomidon Formation in the Fundy basin, Nova Scotia (Sues et al. 1997). The Fundy basin specimen of Hypsognathus was obtained from pebble conglomerate at the base of the Blomidon Formation, which unconformably overlies the Wolfville Formation. The only other vertebrates that occur in the interval of Cliftonian age are indeterminate phytosaur remains (including the holotype of “Clepsysaurus pennsylvanicus” Lea 1851) from the Ukrainian Member of the Passaic Formation in the Newark basin, moderately diverse tetrapod footprint assemblages at many horizons in the Passaic Formation (e.g., Szajna and Silvestri 1996; Lucas and Sullivan 2006), and an indeterminate sphenodontid from the upper Passaic Formation (Olsen 1980; Sues and Baird 1993; Lucas and Huber 2003).
The Malmros Klint and overlying Ørsted Dal members of the Fleming Fjord Formation in eastern Greenland yield tetrapod fossils of Revueltian age (Jenkins et al. 1994, 1997, 2001, 2008; Clemmensen et al. 2016). The Malmros Klint Member has produced fragmentary fossils of plagiosaurid amphibians, the amphibian Cyclotosaurus, phytosaur bones and the prosauropod dinosaur Plateosaurus. The Ørsted Dal Member assemblage is much more diverse: the amphibians Gerrothorax and Cyclotosaurus, the turtle cf. Proganochelys, unidentified sphenodontians, the aetosaurs Aetosaurus and Paratypothorax, the pterosaur Eudimorphodon, the prosauropod dinosaur “Plateosaurus,” a theropod dinosaur, theropod dinosaur footprints (Grallator), and the mammals Kuehneotherium, cf. Brachyzostrodon? and Haramiyavia. As Jenkins et al. (1994) argued, this assemblage shares many taxa with the German Stubensandstein. More specifically, other than Plateosaurus, most taxa from the Ørsted Dal Member are known in the Lower Stubensandstein, to which I correlate the Greenland assemblage (Fig. 10.4).
In Germany, the best known and most diverse Keuper tetrapod assemblage is that of the Lower Stubensandstein (Löwenstein Formation). This assemblage includes the amphibians Cyclotosaurus and Gerrothorax, the turtles Proganochelys and Proterochersis, Pseudopalatus-grade phytosaurs (Nicrosaurus), the aetosaurs Aetosaurus and Paratypothorax, rauisuchians (Teratosaurus), theropod dinosaurs, and the prosauropod dinosaurs Sellosaurus and Thecodontosaurus (e.g., Benton 1993; Hungerbühler 1998; Lucas 1999; Schoch and Werneburg 1999; Schoch 2007; Kimmig and Arp 2010). The phytosaurs, aetosaurs, and rauisuchians provide a strong basis for assigning a Revueltian age to the Lower Stubensandstein (Lucas and Hunt 1993a; Hunt 1994; Lucas 1999). The younger, Middle and Upper Stubensandstein, produce a similar, but less diverse assemblage, so I also assign them a Revueltian age (Lucas 1999). Whether or not the lowest occurrence of Mystriosuchus in the Middle Stubensandstein is of biochronologic significance is not clear.
The tetrapod assemblages of the Upper Stubensandstein and Knollenmergel (Trossingen Formation) are almost entirely dinosaurian—95% or more of the fossils are of dinosaurs (Benton 1986, 1991). This contrasts sharply with the Lower and Middle Stubensandstein assemblages, in which dinosaurs are a much smaller percentage of the fossils collected. However, I regard this change to dinosaur domination as largely a local facies/taphonomic effect, not a biochronologically significant event (Hunt 1991). It seems likely but not certain that the Knollenmergel assemblage is of Apachean age (see below).
In eastern Poland, Dzik et al. (2008) announced the discovery of an Upper Triassic bonebed at the Lipie Slaski clay pit near Lubliniec. This bonebed yields an assemblage dominated by dicynodonts and archosaurs. There are three other correlative fossil vertebrate localities in Silesia. Another bonebed in the Woźniki clay pit yield vertebrates similar to those from Lipie (Sulej et al. 2011). Sulej et al. (2011) correlated the Woźniki assemblage with the Krasiejów locality, but Szulc et al. (2015) argue convincingly that the Woźniki assemblage and the Lipie assemblage are stratigraphically equivalent and subsume it under what they call the Lisowice level. Bones from Poręba were discovered in 2008 and include amphibians, turtles, and aetosaurs, among others, and have in part been described (Sulej et al. 2012; Niedźwiedzki et al. 2014). This locality, and very recently discovered bones from another locality at Zawiercie, are stratigraphically equated to Lipie (Szulc et al. 2015). Thus, the Lisowice level comprises the fossil vertebrate localities at Lipie, Woźniki, Poręba and Zawiercie.
Biochronologically significant tetrapod taxa reported and/or documented from the Lisowice level include the amphibians Cyclotosaurus and Gerrothorax, the turtle cf. Proterochersis, an aetosaur I judge to be Paratypothorax (Lucas 2015) and a large dicynodont (Dzik et al. 2008; Sulej et al. 2012; Niedźwiedzki et al. 2012, 2014; Świło et al. 2014). Szulc et al. (2015) represents the first explicit correlation of the Lipie bonebed to the Triassic LVFs, assigning it a Revueltian age.
In the Lombardian Alps of northern Italy, after the regional progradation of platform carbonates during the early-middle Norian (Dolomia Principale), extensional tectonism produced intraplatform depressions occupied by patch reefs, turbiditic debris flows and lagoonal to freshwater facies (Jadoul 1985; Jadoul et al. 1994). Tetrapods from these intraplatform strata, the Zorzino Limestone at the Cene and Endenna quarries in Lombardy, are the diapsids Endennasaurus and Vallesaurus, the prolacertiform Longobardisaurus, the rhynchocephalian Diphydontosaurus, the drepanosaurids Drepanosaurus and Megalancosaurus, the phytosaur Mystriosuchus, the aetosaur Aetosaurus, the pterosaurs Eudimorphodon and Peteinosaurus and the placodont Psephoderma (e.g., Wild 1989; Pinna 1993; Renesto 2006; Renesto et al. 2010). In Germany, Mystriosuchus is well known from the Middle Stubensandstein (Hungerbühler 2002) and Aetosaurus from the Lower-Middle Stubensandstein, so a Revueltian age of the Zorzino Limestone is certain. The Calcare di Zorzino also crops out in Austria, where it yields specimens of Langobardisaurus and the pterosaur Austriadactylus (Dalla Vecchia 2009, 2013). Also, in Austria, unpublished specimens of Mystriosuchus are known from Totes Gebirge (possibly Dachstein) (Buffetaut 1993).
The other Italian Late Triassic tetrapod sites are in the Forni Dolomite (Dolomia di Forni) in the Veneto Prealps of northeastern Italy. They yield the drepanosaurids Drepanosaurus and Megalancosaurus, the pterosaurs Eudimorphodon and Preondactylus (Dalla Vecchia 1995, 2003, 2006) and a specimen of Langobardisaurus. The presence of Eudimorphodon supports a Revueltian age assignment.
Upper Triassic tetrapod assemblages from the Indian Subcontinent come from the Pranhita-Godavari Valley of south-central India. Several summaries (Jain et al. 1964; Kutty 1969; Kutty and Roychowdhury 1970; Sengupta 1970; Jain and Roychowdhury 1987; Yadagiri and Rao 1987; Kutty et al. 1988; Kutty and Sengupta 1989; Bandyopadhyay and Roychowdhury 1996; Bandyopadyhay and Sengupta 2006; Kammerer et al. 2016) have been published, but other than the lower Maleri assemblage (see above), relatively few of the fossils have been adequately documented in print, forcing me to rely largely on unsubstantiated genus-level identifications to evaluate the ages of the tetrapod assemblages. A case in point is the Dharmaram Formation, which yields two stratigraphically discrete vertebrate fossil assemblages (lower and upper). The stratigraphic range of the lower assemblage has not been published, and it includes a phytosaur that Kutty and Sengupta (1989: table 2) list as Nicrosaurus, aetosaurs, including a so-called “Paratypothorax-like” form, and prosauropod dinosaurs. Based primarily on the supposed Nicrosaurus record, I consider the lower assemblage of the Dharmaram Formation a possible Revueltian correlative.
I formerly and tentatively regarded the Coloradan LVF of Argentina and the tetrapod assemblage of the Lower Elliot Formation in South Africa as of Apachean age. In Argentina, the Los Colorados Formation consists of siliciclastic red beds approximately 800 m thick. Near its base, a single tetrapod fossil—a dicynodont skull, the holotype of “Jachaleria” colorata Bonaparte 1970—was collected. I regard Jachaleria as a synonym of Ischigualastia, so it is likely the lower part of the Los Colorados Formation is of Adamanian age.
The remainder of the tetrapod fossils from the Los Colorados Formation are from its middle and upper parts but have not been stratigraphically organized. The assemblage includes the turtle Palaeochersis, the ornithosuchid Riojasuchus, the aetosaur “Neoaetosauroides” (see below), the rauisuchid Fasolasuchus, the crocodylomorphs Hemiprotosuchus and Pseudhesperosuchus, the prosauropod dinosaurs Riojasaurus and Coloradisaurus, the theropod dinosaur Zupaysaurus and the tritheledontid cynodont Chaliminia (e.g., Bonaparte 1970, 1971, 1978, 1980, 1997; Lucas and Hunt 1994; Rougier et al. 1995; Arcucci et al. 2004). The correlative Quebrada del Barro and El Tranquilo formations also produce prosauropods (e.g., Riojasaurus, “Mussaurus”) (Bonaparte and Vince 1979; Casamiquela 1980; Bonaparte and Pumares 1995). The Los Colorados assemblage clearly is of Late Triassic age (Arcucci et al. 2004) and must be post-Ischigualastian. However, its endemism makes it difficult to correlate precisely. I tentatively considered it an Apachean correlative based primarily on its abundant prosauropods. However, the possibility that it is Revueltian needs to be considered, especially given the similarity of Neoaetosauroides to Aetosaurus.
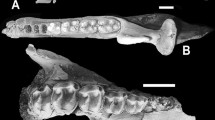
Indeed, having now had the opportunity to study the type material of Neoaetosauroides engaeus firsthand, it is abundantly clear that Neoaetosauroides is a junior synonym of Aetosaurus. Thus, the armor plates of Neoaetosauroides (Fig. 10.5) display all of the diagnostic features of the armor of Aetosaurus (cf. Fraas 1877; Heckert and Lucas 2000; Schoch 2007). To wit, the dorsal paramedian plates are moderately wide (width/length = 2.4–3.1) and possess a radial pattern of elongate ridges and pits and a low dorsal boss near the posterior margin of the plate (Fig. 10.5). The work of Desojo and Baéz (2007: text-fig. 5) makes it clear that the skulls of Neoaetosauroides and Aetosaurus have virtually identical suture patterns and proportions. This synonymy will be documented in greater detail elsewhere. But, I draw attention to it here because Aetosaurus is an index taxon of the Revuletian LVF. Thus, its presence in the middle-upper Los Colorados Formation indicates a Revueltian age.

Part of the holotype of Aetosaurus (“Neoaetosauroides”) engaeus, Instituto Miguel Lillo, Tucuman, Argentina, PVL 3525. On left is a dorsal view of the dorsal armor, and on the right is a detail of part of the armor showing paramedian and lateral plates of the posterior dorsal to sacral region
Kent et al. (2014) presented a magnetostratigraphy of the Los Colorados Formation based on limited sampling of only 52 geomagnetic sites with a stratigraphic spacing of 10 m or more. The 15 polarity intervals identified were pattern matched to the E8-E15 interval of the Newark section, which, if correctly correlated, means the Los Colorados Formation crosses the Carnian-Norian boundary (is of Adamanian-Revueltian age). This is consistent with my interpretation that the Los Colorados is of Adamanian-Revueltian age. Nevertheless, I have little confidence in the pattern matching to the Newark magnetostratigraphy of so few and such widely spaced samples from the Los Colorados Formation.
The age of the tetrapod assemblage from the Lower Elliott Formation in South Africa has long been considered Late Triassic. Lucas and Hancox (2001) reviewed the age of this assemblage, which is dominated by sauropodomorph dinosaurs, but also has rare amphibians (a large chigutisaurid), a possible rauisuchian (Basutodon), the ornithischian dinosaur Eocursor, a traversodontid (Scalenodontoides) and the characteristic Late Triassic footprint ichnogenus Brachychirotherium (Kitching and Raath 1984; Lucas and Hancox 2001; Butler et al. 2007). This is the “Euskelosaurus range zone” of Kitching and Raath (1984), the youngest Triassic tetrapod assemblage in the Karoo basin. Yates (2003) re-evaluated the prosauropods of the Lower Elliott Formation and concluded that most are indeterminate sauropodomorphs or basal sauropods. He noted similarities of indeterminate prosauropods from the Lower Elliott Formation to Riojasaurus from the Los Colorados Formation of Argentina, and similarities between the basal sauropod Antetonitrus from South Africa and Lessemsaurus from Argentina (Yates and Kitching 2003). These conclusions suggest a Lower Elliott-Los Colorados correlation, and thus a tentative Revueltian age assignment.
10.4.5 Apachean Tetrapod Assemblages
The characteristic tetrapod assemblage of the Apachean LVF is from the Redonda Formation of the Chinle Group in Guadalupe and Quay Counties, New Mexico, USA. The following taxa are present: the amphibian Apachesaurus, a sphenodontid, a procolophonid, the phytosaur Redondasaurus, the aetosaurs Redondasuchus and Apachesuchus, the rauisuchian Redondavenator, the archosauriform Vancleavea, theropod dinosaurs and the cynodont Redondagnathus (e.g., Hunt 1994; Hunt and Lucas 1993b, 1997; Heckert et al. 2001; Hunt et al. 2005; Spielmann et al. 2006a, b; Spielmann and Lucas 2012).
Principal correlatives of the type Apachean assemblage are the Whitaker quarry in the Rock Point Formation of the Chinle Group at Ghost Ranch, New Mexico, the Cliftonian LVF assemblages (in part) of the Newark Supergroup, the Knollenmergel (Trossingen Formation), time-equivalent upper Arnstadt Formation and the “Rhaetian Bonebed” (Exter Formation) of the Germanic Basin (Fig. 10.4). Some of the fissure-fill assemblages in the uppermost Mercia Mudstone Group and/or lowermost Penarth Group of the United Kingdom (Fraser 1994; Benton and Spencer 1995; Whiteside and Marshall 2008) may be Apachean correlatives. Some of the so-called Rhaetian vertebrate sites in France, such as Saint-Nicolas-de-Port, may be Apachean correlatives as well (Lucas and Huber 2003).
At Ghost Ranch in New Mexico, the Whitaker quarry bone bed is dominated by skeletons of the theropod dinosaur Coelophysis bauri (Colbert 1989; Rinehart et al. 2009). Nevertheless, it also includes the sphenodont Whitakersaurus, at least one drepanosaur, a rauisuchian skeleton (cf. Postosuchus), the sphenosuchians Hesperosuchus and Vancleavea, the chatterjeeid Shuvosaurus (=Effigia), the phytosaur Redondasaurus and the theropod dinosaur Daemonosaurus (e.g., Hunt and Lucas 1993b; Clark et al. 2000; Harris and Downs 2002; Hungerbühler 2002; Hunt et al. 2002; Lucas et al. 2003; Nesbitt 2007; Lucas et al. 2005; Heckert et al. 2008; Renesto et al. 2009; Rinehart et al. 2009; Sues et al. 2011).
A recently discovered bonebed (the “Saints and Sinners Lagerstätte”) in the Wingate Sandstone (which is the lower part of the “Nugget Sandstone” of Sprinkle et al., 2011) of northeastern Utah includes fossils of sphenodonts, sphenosuchians, drepanosaurs, theropod dinosaurs and a pterosaur, yet to be described (Britt et al. 2016: table 1). Given its stratigraphic position, this bonebed is likely of Apachean age.
The uppermost Triassic strata of the Newark Supergroup in eastern North America yield a low diversity tetrapod assemblage of mostly fragmentary material that defies precise identification. The tetrapods present include the procolophonid Hypsognathus, sphenodontids, indeterminate phytosaurs and the synapsid Oligokyphus (e.g., Gilmore 1928; Baird 1986; Huber et al. 1993b; Fedak et al. 2015). These are the tetrapods of the Cliftonian LVF of Huber et al. (1993b) and at least some are of likely Apachean age.
In the United Kingdom, fissure fills such as Durdham Down in Clifton yield fossils that include phytosaurs, aetosaurs, dinosauriforms and dinosaurs (e.g., Fraser 1994, 2006; Fraser et al. 2002; Galton 2005, 2007a, b; Whiteside and Marshall 2008). Unfortunately, other than a tentative record of Aetosaurus based on a single osteoderm (Lucas et al. 1999), the fissure fill tetrapods are mostly endemic taxa of no biochronological significance or cosmopolitan taxa with long age ranges, such as the sphenodontian Clevosaurus.
Whiteside and Marshall (2008), based primarily on the palynoflora, assigned the Tytherington fissure fill a Rhaetian age, and extrapolated this age to the other fissures. If this Rhaetian age is correct, then the fissure fill tetrapods are of Apachean age. However, as Lucas and Hunt (1994: 340) noted, “a single age should not necessarily be assigned to the fossils from one fissure and…individual fossils from the fissures may range in age from middle Carnian to Sinemurian.” Therefore, I continue to regard as problematic the precise age of the Triassic tetrapod assemblages from the British fissure fills.
10.4.6 Wassonian LVF
Lucas and Huber (2003) introduced the Wassonian LVF, and Lucas and Tanner (2007a) defined is as the time interval between the FAD of the crocodylomorph Protosuchus, and the beginning of the Dawan LVF, which is defined by the FAD of the theropod dinosaur Megapnosaurus (Lucas 2008). The FAD of Protosuchus in Arizona, Nova Scotia and South Africa appears to be lowermost Jurassic, so I take the beginning of the Wassonian LVF to approximate the Triassic-Jurassic boundary (Fig. 10.1). Most Early Jurassic tetrapod assemblages, however, are younger, of Dawan (~Sinemurian) age (Lucas 2008).
10.5 Late Triassic Tetrapod Footprints and Bromalites
The Upper Triassic record of tetrapod footprints was reviewed by Klein and Lucas (2010a) and is also reviewed by Hunt et al. (2017), so I do not review it here. It only resolves into one or two biochronological units, and is thus of much less significance to Late Triassic tetrapod biochronology than is the body-fossil record (e.g., Lucas 2007; Hunt and Lucas 2007; Klein and Lucas 2010a). Similarly, the Late Triassic bromalite (principally coprolite) record has some biochronological utility, but, like the footprint record, it resolves Late Triassic time rather poorly (see Hunt et al. 2017).
Brachychirotherium (sensu stricto), the footprint of aetosaurs (Fig. 10.6; Lucas and Heckert 2011), appears at the beginning of the Otischalkian. It is a characteristic ichnotaxon of the Late Triassic, together with Atreipus-Grallator (quadrupedal to bipedal trackways), Grallator and Eubrontes (bipedal trackways), the latter all attributed to theropods, except for Atreipus, which may also have had an ornithischian trackmaker. The stratigraphic upper limit of Brachychirotherium is the Triassic-Jurassic boundary (end of the Apachean); there is no evidence of Brachychirotherium in post-Triassic strata (Rainforth 2003; Lucas and Tanner 2007a, b, 2015).

The skeleton of the aetosaur Typothorax coccinarum as the trackmaker of Brachychirotherium, in lateral (a, b), anterior (c), posterior (d), dorsal (e) and ventral (f) views. After Lucas and Heckert (2011)
Late Triassic tetrapod footprint assemblages are characteristically archosaur-dominated, particularly by chirothere and theropod tracks, but also feature the oldest records of sauropodomorph tracks. They thus mirror the overall pattern of the body fossil record but provide much less detailed insight into Late Triassic tetrapod distribution and evolution.
10.6 Biotic Events
10.6.1 Introduction
The Late Triassic has long been recognized as a critical juncture in the evolution of terrestrial tetrapods because it was an interval of both important originations and extinctions (e.g., Fraser 2006; Sues and Fraser 2010). Here, I discuss the nature and timing of some of these events (Figs. 10.7, 10.8, 10.9, 10.10, 10.11, 10.12, and 10.13).

Restoration of the Late Triassic metoposaurid amphibian Koskinonodon (artwork by Matt Celeskey)

Restorations of the female (above) and male (below) heads of the phytosaur Pseudopalatus, showing the cranial sexual dimorphism evident in population samples of this phytosaur (artwork by Matt Celeskey)

The Late Triassic dinosauromorph Silesaurus has a foot structure that would make tracks like those of early theropod dinosaurs (artwork by Matt Celeskey)

One of the last dicynodonts, Adamanian Placerias, attacked by a phytosaur (artwork by Matt Celeskey)

Restoration of the oldest mammal, Adelobasileus (artwork by Matt Celeskey)

The drepanosaur Hypuronector (artwork by Matt Celeskey)

Timing of the records of some important biotic events in the Late Triassic evolution of terrestrial tetrapods
.
10.6.2 Archosaurs Rise to Dominance
The Berdyankian is the time of most of the youngest, dicynodont-dominated tetrapod assemblages. These assemblages are primarily from Argentina and Brazil and are dominated by fossils of dicynodonts (mostly the genus Dinodontosaurus), rhynchosaurs and traversodontid cynodonts. Fossils of amphibians and archosaurs are much less common constituents of these assemblages.
In contrast, the amphibian biofacies of Berdyankian time is dominated by mastodonsaurids and plagiosaurs, with much lesser numbers of prolacertiforms, archosaurs and small cynodonts. These assemblages are best known from the Germanic basin and from Russia. An acme in plagiosaur diversity and abundance in these assemblages characterizes Berdyankian time. No procolophonids are known from Berdyankian strata, but this must be due to a lack of discovery, not a real absence, as both pre- and post-Berdyankian procolophonids are known.
Otischalkian tetrapod assemblages are different from Berdyankian assemblages in being dominated by archosaur fossils, particularly of phytosaurs and aetosaurs. Indeed, it is during Otischalkian time, about 15 million years after the end-Permian extinctions that archosaur-dominated tetrapod fossil assemblages first appear. Thus, the old idea (e.g., Benton 1991; McGhee et al. 2004) that the Permo-Triassic extinctions were characterized by the takeover of many terrestrial tetrapod niches by archosauromorphs is erroneous (also see Lucas 2017a).
A new group of temnospondyls, the metoposaurs, is also abundant during Otischalkian time. It is striking how uniform in overall composition the Otischalkian tetrapod assemblages are, from North America, Western Europe, Morocco and India. This implies a degree of cosmopolitanism and no major biofacies differences.
The beginning of the Otischalkian essentially corresponds or overlaps the onset of the Carnian humid episode (e. g., Ruffell et al. 2016). Two groups of dominantly amphibious/aquatic tetrapods are abundant at this time, the metoposaurs and the phytosaurs. This may be a real acme as a response to wetter climates, but that conclusion may be confounded by taphonomy—preferential preservation of aquatic tetrapods by widespread river systems of the Carnian “pluvial.” That caveat aside, metoposaurs and phytosaurs are the dominant components of almost all Otischalkian and younger Late Triassic tetrapod assemblages.
The overall character of many Adamanian tetrapod assemblages is very similar to Otischalkian assemblages—domination by fossils of metoposaurs, phytosaurs and aetosaurs. However, Adamanian tetrapod assemblages from Argentina and Brazil are dominated by dicynodonts, cynodonts and rhynchosaurs, with few to no phytosaurs and metoposaurs. Nevertheless, aetosaurs are common in these assemblages.
Revueltian and Apachean tetrapod assemblages are also dominated mostly by archosaurs and metoposaurs. In some assemblages, dinosaurs are very abundant, and it is clear that the diversification of the dinosaurs had already begun by Revueltian time (Hunt 1991).
10.6.3 Temnospondyl Diminishment and Extinctions
The Late Triassic was the last interval of any substantial temnospondyl diversity (Milner 1990, 1993; Schoch and Milner 2000; Schoch 2008, 2014) (Fig. 10.7). It is also clear that this diversity collapsed stepwise throughout the Late Triassic, so that by Jurassic time few temnospondyl clades remained (Fig. 10.13). The lissamphibian radiation had begun in the Early Triassic (e.g., Rage and Roček 1989), though no definite lissamphibian fossils are known from the Upper Triassic, with the possible exception of Triassurus from Madygen (Ivakhnenko 1978).
The Late Triassic temnospondyl families are the Brachyopidae, Capitosauridae, Chigutisauridae, Almasauridae, Mastodonsauridae, Metoposauridae, Plagiosauridae and Trematosauridae. All are families of low diversity following much greater temnospondyl diversity earlier in the Permian and Triassic (Milner 1990; Ruta and Benton 2008). The record of almasaurids ends in the Otischalkian, and that of mastodonsaurids and trematosaurids ends in the Adamanian. Capitosaurid records are no younger than Revueltian, and metoposaurids and plagiosaurids have records that end in the Apachean. Brachyopids continued into the Jurassic, whereas chigutisaurids continued to the Cretaceous. Chroniosuchians have their last record at Madygen during the Otischalkian(?). Clearly, old ideas of a major extinction of temnospondyls at the end of the Triassic can be abandoned (Lucas and Tanner 2015).
10.6.4 Turtle Origins
The oldest turtles were long considered to be Revueltian in age. Best known is Proganochelys from Germany (Gaffney 1990), and Revueltian or younger turtle records are also known from Poland, Thailand, Greenland, the USA and South America (e.g., Lucas et al. 2000; Joyce et al. 2009; Joyce 2017). Recently added to this record is Odontochelys from marine lower Carnian strata in China (Li et al. 2008) and a supposed Ladinian turtle, Pappochelys from Germany (Schoch and Sues 2015). Furthermore, the middle Permian Eunotosaurus, with expanded ribs, originally proposed as a close turtle relative by Watson (1914), is now being placed back into that position (Lyson et al. 2010, 2012, 2013, 2016).
Much overlooked in the discussion of turtle origins is their footprint record, which begins in the late Early Triassic (Spathian) (Rühle v. Lilienstern 1939; Haubold 1971; Lovelace and Lovelace 2012; Thomson and Lovelace 2014; Lichtig et al. 2017). Thus, recent claims that Odontochelys or Pappochelys are the oldest turtles are incorrect—the oldest turtle fossils (footprints) are of Early Triassic (Nonesian) age. They thus predate the oldest body fossils of turtles (and I do not consider Pappochelys to be a turtle; it is a placodont) by at least 10 million years. The origin of turtles thus is not, as long thought, a Late Triassic event, but took place much earlier (Fig. 10.13).
10.6.5 Rhynchosaurs
Rhynchosaurs are a Triassic group of herbivorous archosauromorphs that were most diverse during the Middle Triassic. Indeed, Romer (1966) long argued that together with gomphodont cynodonts they were dominant elements of Middle Triassic tetrapod assemblages. However, rhynchosaurs were of low diversity during the Late Triassic, known from only a few genera. Most interesting is the very cosmopolitan Hyperodapedon found in Otischalkian-Adamanian assemblages in North America, western Europe, India, South America, Africa and Madagascar (Lucas et al. 2002).
The rhynchosaur genus Otischalkia is known from Otischalkian-Revueltian strata of the Chinle Group in the western USA (Hunt and Lucas 1991; Long and Murry 1995; Spielmann et al. 2013; Lucas et al. 2016). The Revueltian record of Otischalkia is the only bona fide Norian record of a rhynchosaur, and demonstrates that they did not go extinct at the Carnian-Norian boundary, as long thought (e.g., Benton 1991).
Schultz et al. (2016: fig. 10) recently presented what I regard as an erroneous time calibration of Late Triassic rhynchosaurs. Thus, they show “Isalorhynchus” (a synonym of Hyperodapedon) predating Hyperodapedon during the Carnian. They show Teyumbatia (a cladotaxon named and described by Montefeltro et al. 2010, 2013, likely synonymous with Hyperodapedon; cf. Azevedo and Schultz 1987) as Norian, though the fossil is from the Adamanian Caturitta Formation and therefore of Carnian age (see above). Schultz et al. (2016) also uniquely claim that Otischalkia is not diagnostically a rhynchosaur. Clearly, my view of Late Triassic rhynchosaur distribution differs markedly from that of Schultz et al. (2016), though all agree that rhynchosaur diversity was low during the Late Triassic.
10.6.6 Phytosaurs and Aetosaurs
Recent analysis identifies Middle Triassic Diandongosuchus as the oldest phytosaur and cladotaxonomically splits many long-recognized phytosaur genera into multiple genera (e. g., Stocker 2012; Stocker and Butler 2013; Butler et al. 2014; Stocker et al. 2017). This work ignores important factors of biological variation in phytosaurs (e.g., Fig. 10.8; Kimmig and Spielmann 2011) and will need substantial revision. The peak of phytosaur diversity was during the Adamanian-Revueltian, with perhaps no more than one genus surviving into Apachean time. A phytosaur snout fragment from the Lower Jurassic of the UK is the stratigraphically highest phytosaur fossil not clearly reworked, so it suggests phytosaur survival across the Triassic-Jurassic boundary (Maisch and Kapitzke 2010) (Fig. 10.13).
Aetosaur taxonomy is similarly being overspilt by cladotaxonomy (Desojo et al. 2013) and will need revision. The oldest aetosaur records are Otischalkian, with an acme during the Adamanian-Revueltian. Most of the evidence of Apachean aetosaurs is based on the footprint ichnogenus Brachychirotherium (see above).
Assemblages dominated by metoposaurs, phytosaurs and/or aetosaurs characterize much of the Late Triassic tetrapod record. Clearly, a taphonomic overprint is at least in part responsible for this. Metoposaurs and phytosaurs were amphibious tetrapods that would have been preferentially preserved/fossilized in the fluvial sediments from which comes most of the Late Triassic tetrapod fossil record. Nevertheless, before the Otischalkian no such metoposaur/phytosaur/aetosaur assemblages are known, though these groups likely had a Middle Triassic ancestry. The abundance and ubiquity of metoposaurs and phytosaurs is a Late Triassic phenomenon across Pangea, with the exception of South America.
10.6.7 Crocodylomorphs
The oldest crocodylomorphs are sphenosuchans, with a record that begins in the Otischalkian (Lucas et al. 1998). Among them are taxa well-known from comprehensive cranial and postcranial material such as Hesperosuchus, Sphenosuchus, Saltoposuchus, Dibothrosuchus and Protosuchus (e. g., Colbert and Mook 1951; Colbert 1952; Crush 1984; Parrish 1991; Sereno and Wild 1992; Wu and Chatterjee 1993; Lucas et al. 1998; Clark et al. 2004). These early crocodiles have long been interpreted as lightly built cursorial and fully terrestrial forms.
No substantial change takes place in the Sphenosuchia across the Triassic-Jurassic boundary (Klein and Lucas 2015). What does occur much later, definitely in the Late Jurassic, perhaps as early as the Middle Jurassic, is the appearance of aquatic/amphibious crocodylians. The body fossil and footprint records document this (e.g., Lockley et al. 2010; Klein and Lucas 2010b, 2015). Clearly, the aquatic predator niches now associated with crocodylians were not occupied by crocodylomorphs during the Late Triassic-Early Jurassic. Phytosaurs occupied at least some of those niches during the Late Triassic (Hunt 1989).
10.6.8 Pterosaur Origins
Pterosaurs appear suddenly and completely adapted to flight during the Late Triassic. The lack of antecedents has made the origin and precise phylogenetic placement of pterosaurs somewhat unclear. Certainly pterosaurs are archosaurs, and most (but not all) analyses place them close to dinosaurs (see Hone and Benton 2008; Dalla Vecchia 2013 for reviews). Scleromochlus from the Adamanian of Scotland has been considered to be related to the ancestry of pterosaurs by some (e.g., Padian 1984), but others have dismissed that idea (e.g., Sereno 1991).
The oldest complete and reliable skeletal records of pterosaurs are Revueltian, from Italy, but isolated teeth and bones of Adamanian age have also been identified (with less certainty) as pterosaur (see Barrett et al. 2008; Dalla Vecchia 2013). Faxinalipterus from the Adamanian of Brazil, a supposed pterosaur (Bonaparte et al. 2010b), is not demonstrably a pterosaur according to Dalla Vecchia (2013). Given the phylogenetic analysis and their temporal distribution it seems that, as with dinosaur origins (see below), pterosaur origins was likely a Middle Triassic event for which we need a fossil record. However, on face value, the pterosaur first appearance is a sudden one during the Revuletian of forms very similar to the long-tailed, non-pterodactyloid pterosaurs of the Jurassic.
10.6.9 Dinosaur Origins
Understanding dinosaur origins has been beset by three problems: (1) semantic, namely what is and what is not a dinosaur; (2) cladistic—disagreements over the phylogeny of dinosaur origins, confounded by an inability to sort out convergence; and (3) biochronologic—disagreements over the actual age of the “oldest dinosaur.” Despite these problems, all agree that the oldest dinosaur body fossils are Carnian (e.g. Langer et al. 2009; Lucas 2010; Benton 2012; Nesbitt and Ezcurra 2015). More precisely, they are Otischalkian, from the Chinle Group in the western USA (Lepidus) and the Maleri Formation of India (Alwalkeria). Otischalkian or Adamanian records of “dinosaurs” (Azendohsaurus) from Morocco and Madagascar have been discounted as not being based on dinosaurs (see Langer 2014 for discussion). Records from South America are Adamanian, so they are not the oldest records of dinosaur body fossils (contra Langer et al. 2009; Langer 2014).
The footprint record of the earliest dinosaurs also does not demonstrate dinosaurs older than Carnian. This, despite diverse claims (one of the most recent being by Marsicano et al. 2007) that tridactyl, apparently mesaxonic footprints from Middle Triassic strata are dinosaurian. Most of these are tridactyl remnants of chirothere tracks (see review by Klein and Lucas 2010a). Also, some could be the tracks of dinosauromorphs (Fig. 10.9) or of ornithodirans close to dinosaur origins, as discussed by Marsicano et al. (2007).
Particularly misleading, however, was the claim of dinosauromorph tracks in the Lower Triassic of Poland (Brusatte et al. 2011). This claim was based on the ichnogenus Protorodactylus, which is barely, if at all distinguishable from Rhynchosauroides (Klein and Niedzwiedźki 2012). The so-called “dinosauromorph” tracks of Brusatte et al. (2011) thus are most likely the tracks of lepidosauromorphs.
To conclude, the oldest dinosaur fossils are Carnian, more specifically of Otischalkian age. Importantly, they are not from South America, as many have erroneously claimed, though, clearly, much more remains to be discovered of dinosaur origins.
10.6.10 Dicynodont Diminishment and Extinction
The dinocephalian extinction event near the end of the middle Permian opened up the tetrapod herbivore niches to dicynodonts (Lucas 2017a). During the late Permian and Early Triassic, successive tetrapod assemblages were dominated by a single dicynodont genus, most famously Lystrosaurus of the earliest Triassic. However, other than in South America, by Berdyankian time dicynodont domination of the tetrapod herbivore niches had diminished (Fig. 10.10). Dicynodont diversity was low in the Late Triassic, with only one-two genera per LVF, and dicynodonts are only abundant in some South American assemblages. Late Triassic dicynodonts are also known from mass death assemblages in Arizona, Morocco and Poland.
The tetrapod herbivore niches occupied by dicynodonts may have been taken over by herbivorous aetosaurs, rhynchosaurs, prosauropod dinosaurs and cynodonts. However, the feeding mechanism of these herbivores are very different than those of dicynodonts, so replacement seems unlikely.
The HO (highest occurrence) of dicynodonts was long thought to be Adamanian. However, there is a putative Cretaceous record from Australia (Thulborn and Turner 2003), though it seems extremely doubtful. Dzik et al. (2008) recently reported a Triassic dicynodont from Poland in strata they deemed Rhaetian based on paleobotany, but that are Revueltian (Lucas 2015). These are the youngest known dicynodonts (Fig. 10.13).
10.6.11 Cynodont Evolution and Mammal Origins
The first cynodonts appear at about the end of the middle Permian and gradually increased in generic diversity through the Permo-Triassic boundary, the prelude to a much greater diversification during the Middle-Late Triassic (e. g., Abdala and Ribeiro 2010; Smith et al. 2012; Ruta et al. 2013; Abdala and Gaetano 2017). Late Triassic cynodont families are primarily the Traversodontidae, Trithelodontidae and Tritylodontidae. Traversodontids were most diverse (~ 17 genera), mostly of Adamanian age and mostly known from Gondwana (Liu and Abdala 2014). The only well-documented, post-Adamanian traversodontid is Scalenodontoides from the Revueltian Lower Elliott Formation of South Africa.
Tritylodontids were very mammal-like, especially in their postcranial anatomy. Their first appearance is during the Revueltian. However, most of their diversity is Early Jurassic. Trithelodontids (including the “dromatheres”) were also not very diverse during the Late Triassic, but they most likely include the ancestors of mammals.
For decades, the oldest mammals were considered to be of Late Triassic age, notably Morganucodon from the British fissure fills. Cladistic analysis of the earliest mammals by Rowe (1988), however, changed that by redefining Mammalia as a crown clade consisting of the common ancestor of living monotremes and therians.
Cladistic analysis also “made” mammals monophyletic, even though such authorities as Olson and Simpson had entertained the possibility of a polyphyletic Mammalia. Questions of mammal origins had long been deeply emeshed in functional-adaptive analyses of the biology of the transition, particularly changes in the auditory and masticatory apparatuses. In the abiological world of numerical cladistics, these questions were trivialized as “essentialist” and not of scientific interest (Rowe and Gauthier, 1992; DeQuieroz 1994).
Miao (1991) provided a pointed discussion of these issues, rejecting cladistic redefinition of the Mammalia (also see Lucas 1992). Following Lucas and Luo (1993), I define Mammalia as the monophyletic group that includes Adelobasileus (Fig. 10.11), Sinoconodon and Morganucodon in the Mammalia.
Thus, I recognize a monophyletic Mammalia defined primarily by the biologically significant novelty of a petrosal promontorium, likely with a coiled cochlea. The oldest mammal thus is Adelobasileus from the Adamanian of Texas, USA (Lucas and Luo 1993). Potentially older mammals may be known from teeth from the Tiki Formation of India (Datta and Das 1996; Datta 2005). Mammal origins from a trithelodontid cynodont seem likely, and this was a Late Triassic event.
During the Revueltian-Apachean there was a diversification of morganucodontids, haramiyids and “symmetrodonts,” particularly in Europe (Kielan-Jaworowska et al. 2004; Luo 2011; Debuysschere et al. 2015). Moganucodonts achieved a Pangean distribution and were small (less than 100 g estimated body weight) insectivores represented by 16 genera. All of the well known morganucodontid genera cross the Triassic-Jurassic boundary.
10.6.12 Oddities
There are a variety of highly specialized, ergo bizarre, Late Triassic diapsids. These include drepanosaurs (Fig. 10.12), trilophosaurs and the possible gliders Icarosaurus, Sharovipteryx and Longisquama. Most or all of these taxa defy precise classification and lack antecedents that would connect them morphologically to more generalized ancestors. They demonstrate just how incomplete our knowledge is of the Late Triassic tetrapod record. In effect, they are end members of clades that have been little sampled.
10.6.13 Provinciality
There have been at least two views of tetrapod paleobiogeography across Late Triassic Pangea. The traditional view is one of cosmopolitanism, and whatever provinciality existed was the difference between Laurussian archosaur-dominated assemblages and Gondwanan therapsid-dominated assemblages (e. g., Romer 1966; Cox 1973). More recent studies argue for a distinction between Laurussian and Gondwanan tetrapod assemblages largely predicated on floral differences or for provinciality driven by paleloatitude (zonal climate belts) (Ezcurra 2010; Whiteside et al. 2011).
Shubin and Sues (1991) inferred tetrapod cosmopolitanism during the Middle Triassic followed by latitudinal variation that correlated to long-recognized paleofloral differences (e.g., Dobruskina 1995b; Artabe et al. 2003). However, their correlation of tetrapod assemblages (see especially Shubin and Sues 1991: fig. 3) is fraught with inaccuracies. Furthermore, their analysis identifies India and Madagascar as having affinities with the tetrapod assemblages of Europe and North America, even though the floras of India and Madagascar are Gondwanan (Dicroidium dominated).
Whiteside et al. (2011) claimed latitudinal differences between tetrapod faunas as due to the differences between areas in which sedimentation was driven by ~10 kyr cycles (cynodont-dominated) and those driven by ~20 kyr cycles (procolophonid-dominated). Their analysis was based on the tetrapod record of the Newark Supergroup in which there is essentially no temporal overlap between older, cynodont-dominated assemblages and younger procolophonid-dominated assemblages. Clearly, the differences between these assemblages could be due to their different geological ages, not to paleolatituidinal differences.
Ezcurra (2010) analyzed the paleobiogeography of Triassic tetrapods for three time slices: Middle Triassic, “Ischigualastian” and “Coloradan.” Like Shubin and Sues (1991), his analysis identified Middle Triassic cosmopolitanism. Interestingly, his analysis of the “Ischigualastian” linked India and Europe but not North America, despite the great similarity between North American and European tetrapod assemblages. In general, his analysis employs too coarse of temporal resolution to identify meaningful paleobiogeographic differences.
The correlations advocated here indicate that the strikingly distinct Late Triassic tetrapod assemblages are those from the Berdyankian-Adamanian of South America. Whether or not these assemblages, dominated by dicynodonts, represent a distinct province or a distinct facies, however, is difficult to determine. Given the inland nature of deposition in the South American basins that produce these assemblages, the possibility that they are just representatives of the dicynodont-dominated biofacies seen earlier in the Triassic (see discussion above) needs to be entertained. Otherwise, the Late Triassic tetrapod record seems to demonstrate much cosmopolitanism.
10.7 Conclusion: Late Triassic Extinctions
Colbert (1958) long ago drew attention to the striking differences between Late Triassic and Early Jurassic tetrapod faunas. Late Triassic tetrapod faunas are populated by many “thecodonts,” notably phytosaurs, aetosaurs and rauisuchians. Dinosaurs were not major components of most Late Triassic tetrapod faunas, and metoposaurs dominated the amphibians. In strong contrast, Early Jurassic tetrapod faunas are dinosaur dominated, with various crocodylomorphs, and totally lack “thecodonts” and metoposaurs, taxa that suffered extinction across the Triassic-Jurassic boundary. The question has been when and how did these extinctions take place.
Lucas and Tanner (2015) recently analyzed the Late Triassic tetrapod extinctions to conclude that they were not a single extinction at the Triassic-Jurassic boundary. There is also no evidence of a tetrapod extinction across the Carnian-Norian boundary (e.g. Lucas 1994; Sues and Fraser 2010).
Using the best temporal resolution, the Late Triassic looks like a prolonged interval of stepwise tetrapod extinctions and low origination rates. However, none of these is a mass extinction, and no reliable data support continued claims of a tetrapod extinction just before or at the end of the Triassic (contra Kent et al. 2017). With new and more detailed stratigraphic data, the perceived Triassic-Jurassic boundary tetrapod extinction is mostly an artifact of coarse temporal resolution, the compiled correlation effect. The amphibian, archosaur and synapsid extinctions of the Late Triassic were not concentrated at the Triassic-Jurassic boundary, but instead occurred stepwise, beginning in the Norian and extending into the Hettangian (Fig. 10.13; Lucas and Tanner 2015).
References
Abdala F, Gaetano LC (2017) Late Triassic cynodont life: time of innovations in the mammal lineage. In: Tanner LH (ed) The Late Triassic world: earth in a time of transition. Topics in geobiology. Springer, New York (this volume)
Abdala F, Ribeiro AM (2003) A new traversodontid cynodont from the Santa Maria Formation (Ladinian-Carnian) of southern Brazil, with a phylogenetic analysis of Gondwanan traversodontids. Zool J Linnean Soc 139:529–545
Abdala F, Ribeiro AM (2010) Distribution and diversity patterns of Triassic cynodonts (Therapsida, Cynodontia) in Gondwana. Palaeogeogr Palaeoclimatol Palaeoecol 286:202–217
Abdala F, Smith RMH (2009) A Middle Triassic cynodont fauna from Namibia and its implications for the biogeography of Gondwana. J Vert Paleont 29:837–851
Abdala F, Ribeiro AM, Schultz CL (2001) A rich cynodont fauna of Santa Cruz do Sul, Santa Maria Formation (Middle-Upper Triassic), in southern Brazil. Neues Jahrb Geol Paläont Monat 2001:669–687
Abdala F, Marsicano CA, Smith RHM, Swart R (2013) Strengthening western Gondwanan correlations: a Brazilian dicynodont (Synapsida, Anomodontia) in the Middle Triassic of Namibia. Gondwana Res 23:1151–1162
Alcober O, Parrish JM (1997) A new poposaurid from the Upper Triassic of Argentina. J Vert Paleont 17:548–556
Alifanov VR, Kurochkin EN (2011) Kyrgyzsaurus bukhanchenkoi gen. et. sp. nov., a new reptile from the Triassic of southwestern Kyrgyzstan. Paleont J 45:639–647
Araújo DC, Gonzaga TD (1980) Uma nova especie de Jachaleria (Therapsida, Dicynodonta) do Triassico de Brasil. Actas II Congr Paleontol Bioestrat I Congr Latinoamer Paleont Buenos Aires 1:159–174
Arcucci AB, Marsicano CA (1998) A distinctive new archosaur from the Middle Triassic (Los Chañares Formation) of Argentina. J Vert Paleont 18:222–232
Arcucci AB, Marsicano CA, Caselli AT (2004) Tetrapod association and palaeoenvironment of the Los Colorados Formation (Argentina): a significant sample from western Gondwana at the end of the Triassic. Geobios 37:557–568
Artabe AE, Morel EM, Spalletti LA (2003) Caracterización de las provincias fitogeográficas triásicas del Gondwana extratropical. Ameghin 40:387–405
Atchley SC, Nordt LC, Dworkin SJ, Ramezani J, Parker WG, Ash SR, Bowring SA (2013) A linkage among Pangean tectonism, cyclic alluviation, climatic change, and biologic turnover in the Late Triassic: the record from the Chinle Formation, southwestern United States. J Sed Res 83:1147–1161
Azevedo SAK, Schultz CL (1987) Scaphonyx sulcognathus sp. nov., um novo rincossaurídeo Neotriássico do Rio Grande do Sul, Brasil. Anais do X Congr Brasil Paleont, Rio de Janeiro 1987:99–113
Bachmann GH, Kozur HW (2004) The Germanic Triassic: correlations with the international scale, numerical ages and Milankovitch cyclicity. Hall Jahrb Geowissen B26:17–62
Baczko MB, Ezcurra MD (2013) Ornithosuchidae: a group of Triassic archosaurs with a unique ankle joint. In: Nesbitt SJ, Desojo JB, Irmis RB (eds) Anatomy, phylogeny and paleobiology of early archosaurs and their kin. Geol Soc London Spec Publ 379: 187–202
Baird D (1986) Some Upper Triassic reptiles, footprints, and an amphibian from New Jersey. Mosasaur 3:125–153
Bandyopadhyay S, Roychowdhury TK (1996) Beginning of the continental Jurassic in India: a paleontological approach. Mus North Ariz Bull 60:371–378
Bandyopadyhay S, Sengupta DP (2006) Vertebrate faunal turnover during the Triassic-Jurassic transition: an Indian scenario. New Mex Mus Nat Hist Sci Bull 37:77–85
Barberena MC (1977) Bioestratigrafia preliminar da Formacâo Santa Maria. Pesquisas Porto Alegre 7:111–129
Barberena MC, Araújo DC, Lavina EL (1985) Late Permian and Triassic tetrapods of southern Brazil. Nat Geog Res 1:5–20
Barrett PM, Butler RJ, Edwards NP, Milner AR (2008) Pterosaur distribution in time and space. Zitteliana B28:35–60
Benton MJ (1986) The Late Triassic tetrapod extinction events. In: Padian K (ed) The beginning of the age of dinosaurs. Cambridge University Press, New York, pp 303–320
Benton MJ (1991) What really happened in the Late Triassic? Hist Biol 5:263–278
Benton MJ (1993) Late Triassic terrestrial vertebrate extinctions: stratigraphic aspects and the record of the Germanic basin. Paleont Lomb Nu Ser 2:19–38
Benton MJ (2012) Origin and early evolution of dinosaurs. In: Brett-Surman MK, Holtz TR Jr, Darlow J (eds) The complete dinosaur. Indiana University Press, Bloomington, pp 331–345
Benton MJ, Spencer PS (1995) Fossil reptiles of Great Britain. Chapman and Hall, London
Bertinelli A, Casacci M, Concheri G, Gattolin G, Godfrey L, Katz ME, Maron M, Mazza M, Mietto P, Muttoni G, Rigo M, Sprovieri M, Stellin F, Zaffani M (2016) The Norian/Rhaetian boundary interval at Pignola-Abriola section (Southern Apennines, Italy) as a GSSP candidate for the Rhaetian stage: an update. Albertiana 43:5–18
Besarie H (1930) Recherches géologiques à Madagascar. Contribution à l’étude des resources minérales. Bull Soc Hist Nat Toul 59:345–616
Besarie H, Collignon M (1960) Madagascar (Supplément). Lex Strat Internat Paris 4(2):1–190
Besarie H, Collignon M (1971) Géologie de Madagascar I. Les Terrains Sedimentaires. Ann Geol Madagascar 35:1–461
Bonaparte JF (1966) Chronological survey of the tetrapod-bearing Triassic of Argentina. Breviora 251:1–13
Bonaparte JF (1967) Cronología de algunas formaciones Triásicas de Argentina: Basada en restos de tetrápodos. Assoc Geol Argent Rev 21:20–38
Bonaparte JF (1970) Annotated list of the South American Triassic tetrapods. In: Second Gondw Symp Proc Pap, pp 665–682
Bonaparte JF (1971) Los tetrapodos del sector superior de la formacion Los Colorados, La Rioja, Argentina (Triásico Superior). I Parte. Op Lillo 22:1–183
Bonaparte JF (1976) Pisanosaurus mertii Casamiquela and the origin of the Ornithischia. J Paleontol 50:808–820
Bonaparte JF (1978) El Mesozoico de América del Sur y sus tetrápodos. Op Lillo 26:1–596
Bonaparte JF (1980) El primer ictidosaurio (Reptilia-Therapsida) de America del Sur, Chaliminia musteloides, del Triásico Superior de La Rioja, República Argentina. Actas II Congr Argent Paleontol Bioestrat Buenos Aires 1978(1):123–133
Bonaparte JF (1982) Faunal replacement in the Triassic of South America. J Vert Paleont 2:362–371
Bonaparte JF (1997) El Triasico de San Juan-La Rioja Argentina y sus dinosaurios. Museo Argentino de Ciencias Naturales, Buenos Aires
Bonaparte JF, Pumares JA (1995) Notas sobre el primer craneo de Riojasaurus incertus (Dinosauria, Prosauropoda, Melanorosauridae) del Triasico Superior de La Rioja, Argentina. Ameghin 32:341–349
Bonaparte JF, Sues HD (2006) A new species of Clevosaurus (Lepidosauria: Rhynchocephalia) from the Upper Triassic of Rio Grande do Sul, Brazil. Palaeont 49:917–923
Bonaparte JF, Vince M (1979) El hallazgo del primer nido de dinosaurios Triásicos (Saurischia, Prosauropoda), Triásico superior de Patagonia, Argentina. Ameghin 16:173–182
Bonaparte JF, Ferigolo J, Ribeiro AM (1999) A new early Late Triassic saurischian dinosaur from Rio Grande do Sul State, Brazil. Nat Sci Mus Monog 15:89–109
Bonaparte JF, Ferigolo J, Ribeiro AM (2001) A primitive Late Triassic ‘ictidosaur’ from Rio Grande do Sul, Brazil. Palaeontology 44:623–635
Bonaparte JF, Brea G, Schultz CL, Martinelli AG (2007) A new specimen of Guaibasaurus candelareiensis (basal Saurischia) from the Late Triassic Caturrita Formation of southern Brazil. Hist Biol 19:73–82
Bonaparte JF, Schultz CL, Soares MB (2010a) Pterosauria from the Late Triassic of southern Brazil. In: New aspects of Mesozoic biodiversity. Springer, Berlin, pp 63–71
Bonaparte JF, Schultz CL, Soares MB, Martinelli A (2010b) La fauna local de Faxinal do Soturno, Triássico Tardío de Rio Grande do Sul, Brazil. Rev Brasil Paleont 13:1–14
Branson EB, Mehl MG (1928) Triassic vertebrate fossils from Wyoming. Science 67:325–326
Britt BB, Chure DJ, Engelmann GF, Shumway JD (2016) Rise of the erg—paleontology and paleoenvironments of the Triassic-Jurassic transition in northeastern Utah. Geol Intermount West 3:1–30
Brusatte SL, Niedźwiedzki G, Butler RJ (2011) Footprints pull origin and diversification of dinosaur stem lineage deep into Early Triassic. Proc R Soc B 278:1107–1113
Buffetaut E (1983) Isalorhynchus genovefae, n. g. n. sp. (Reptilia, Rhynchocephalia), un noveau rhynchosaure du Trias de Madagascar. Neues Jahrb Geol Paläont Monats 1983:465–480
Buffetaut E (1993) Phytosaurs in time and space. Paleontol Lomb Nuo Serie 2:39–44
Burmeister KC, Flynn JJ, Parrish JM, Wyss AR (2006) Paleogeographic and biostratigraphic implications of new early Mesozoic vertebrates from Poamay, central Morondova basin, Madagascar. New Mex Mus Nat Hist Sci Bull 37:457–475
Butler RJ, Smith RMH, Norman DB (2007) A primitive ornithischian dinosaur from the Late Triassic of South America, and the early diversification of Ornithischia. Proc R Soc B 274:2041–2046
Butler RJ, Rauhut OWM, Stocker MR, Bronowicz R (2014) Redescription of the phytosaurs Paleorhinus (Francosuchus) angustifrons and Ebrachosuchus neukami from Germany, with implications for Late Triassic biochronology. Zool J Linnean Soc 170:155–208
Cabrera A (1944) Sobre un estegocéfalo de la Provincia de Mendoza. Not Mus La Plata 9:421–429
Camp CL, Welles SP (1956) Triassic dicynodont reptiles. Mem Univ Cal 13:255–348
Casamiquela RM (1960) Noticia preliminar sobre dos nuevos estagonolepoideos Argentinos. Ameghin 2:3–9
Casamiquela RM (1962) Dos nuevos estagonolepoideos Argentinos. Rev Assoc Geol Argent 16:143–203
Casamiquela RM (1980) La presencia del genero Plateosaurus (Prosauropoda) en el Triásico Superior de la Formación El Tranquilo, Patagonia. Actas II Congr Argent Paleontol Bioestratig, I Congr Latinoamer Paleont 1:143–158
Chatterjee S (1967) New discoveries contributing to the stratigraphy of the continental Triassic sediments of the Pranhita Godavari Valley. Bull Geol Soc India 4:37–41
Chatterjee S (1974) A rhynchosaur from the Upper Triassic Maleri Formation of India. Phil Trans R Soc Lond B267:209–261
Chatterjee S (1978) A primitive parasuchid (phytosaur) reptile from the Upper Triassic Maleri Formation of India. Palaeontology 21:83–127
Chatterjee S (1980) Malerisaurus, a new eosuchian reptile from the Late Triassic of India. Phil Trans R Soc Lond B291:163–200
Chatterjee S (1982) A new cynodont reptile from the Triassic of India. J Paleontol 56:203–214
Chatterjee S (1987) A new theropod dinosaur from India with remarks on the Gondwana-Laurasia connection in the Late Triassic. In: McKenzie GD (ed) Gondwana six: stratigraphy, sedimentology and paleontology. Amer Geophys Un Monog 41: 183–189
Chatterjee S, Roychowdhury T (1974) Triassic Gondwana vertebrates from India. Ind J Earth Sci 12:96–112
Cirilli S, Marzoli A, Tanner LH, Bertrand H, Buratti N, Jourdan F, Bellieni G, Kontak D, Renne PR (2009) Late Triassic onset of the Central Atlantic Magmatic Province (CAMP) volcanism in the Fundy Basin (Nova Scotia): new stratigraphic constraints. Earth Planet Sci Lett 286:514–525
Cisneros JC, Schultz CL (2003) Soturnia caliodon n. g. n. sp., a procolophonid reptile from the Upper Triassic of southern Brazil. Neues Jahrb Geol Paläont Monats 227:365–380
Cisneros JC, Damiani R, Schultz C, de Rosa A, Schwanke C, Neto LW, Aurélio PLP (2004) A procolophonoid reptile with temporal fenestration from the Middle Triassic of Brazil. Proc R Soc Lond B271:1541–1546
Clark JM, Sues HD, Berman DS (2000) A new specimen of Hesperosuchus agilis from the Upper Triassic of New Mexico and the interrelationships of basal crocodylomorph archosaurs. J Vert Paleont 20:683–704
Clark JM, Xu X, Forster CA, Wang Y (2004) A Middle Jurassic ‘sphenosuchian’ from China and the origin of the crocodilian skull. Nature 430:1021–1024
Clemmensen LB, Milàn J, Adolfssen JS, Estrup EJ, Frobose N, Klein N, Mateus O, Wings O (2016) The vertebrate-bearing Late Triassic Fleming Fjord Formation of central Greenland revisited: stratigraphy, palaeoclimate and new palaeontological data. In: Kear PB, Lindgren J, Hurm JH, Milàn J, Vajda V (eds) Mesozoic biotas of Scandinavia and its Arctic territories. Geol Soc London Spec Publ 434: 31–47
Colbert EH (1952) A pseudosuchian reptile from Arizona. Bull Am Mus Nat Hist 99:561–592
Colbert EH (1957) Triassic vertebrates of the Wind River basin. Wyo Geol Assoc Guidebk 12:89–93
Colbert EH (1958) Triassic tetrapod extinction at the end of the Triassic Period. Proc Natl Acad Sci U S A 44:973–977
Colbert EH (1989) The Triassic dinosaur Coelophysis. Bull Mus North Ariz 57:1–174
Colbert EH, Mook CC (1951) The ancestral crocodilian Protosuchus. Bull Am Mus Nat Hist 97:143–182
Cope ED (1871) Observations on the distribution of certain extinct vertebrates in North Carolina. Proc Am Philos Soc 12:210–216
Cox CB (1965) New Triassic dicynodonts from South America, their origins and relationships. Phil Trans R Soc Lond B248:457–516
Cox CB (1973) Triassic tetrapods. In: Hallam A (ed) Atlas of palaeobiogeography. Elsevier, Amsterdam, pp 213–223
Cox CB (1991) The Pangea dicynodont Rechnisaurus and the comparative biostratigraphy of Triassic dicynodont faunas. Palaeontology 34:767–784
Crush PJ (1984) A late Upper Triassic sphenosuchid crocodilian from Wales. Palaeontology 27:131–157
Currie BS, Colombi CE, Tabor NJ, Shipman TC, Montanez IP (2009) Stratigraphy and architecture of the Upper Triassic Ischigualasto Formation, Ischigualasto Provincial Park, San Juan, Argentina. J S Am Earth Sci 27:74–87
Dalla Vecchia FM (1995) A new pterosaur (Reptilia, Pterosauria) from the Norian (Late Triassic) of Friuli (northeastern Italy). Preliminary note. Gortania 16:59–66
Dalla Vecchia FM (2003) A review of the Triassic pterosaur record. Riv Mus Civ Sci Natural “E. Caffi” 22:13–29
Dalla Vecchia FM (2006) The tetrapod fossil record from the Norian-Rhaetian of Friuli (northeastern Italy). New Mex Mus Nat Hist Sci Bull 37:432–444
Dalla Vecchia FM (2009) Anatomy and systematics of the pterosaur Carniadactylus gen. n. rosenfeldi (Dalla Vecchia, 1995). Riv Ital Paleont Stratig 115:159–198
Dalla Vecchia FM (2013) Triassic pterosaurs. In: Nesbitt SJ, Desojo JB, Irmis RB (eds) Anatomy, phylogeny and paleobiology of early archosaurs and their kin. Geol Soc London Spec Publ 379: 119–155
Damiani R, Schoch RR, Hellrung H, Werneburg R, Gastou S (2009) The plagiosaurid temnospondyl Plagiosuchus pustuliferus (Amphibia: Temnospondyli) from the Middle Triassic of Germany: anatomy and functional morphology of the skull. Zool J Linnean Soc 155:348–373
Datta PM (2005) Earliest mammal with transversely expanded upper molar from the Late Triassic (Carnian) Tiki Formation, South Rewa Gondwana basin, India. J Vert Paleont 25:200–207
Datta PM, Das DP (1996) Discovery of the oldest fossil mammal from India. India Min 50:217–222
Debuysschere M, Gheerbrant E, Ailian R (2015) Earliest known European mammals. A review of Morganucodon from Saint-Nicolas-de-Port (Upper Triassic, France). J Syst Palaeont 13:825–855
DeQuieroz K (1994) Replacement of an essentialist perspective on taxonomic definition as exemplified by the definition of “Mammalia”. Syst Biol 43:497–510
Desojo JB, Baéz AM (2007) Cranial morphology of the Late Triassic South American archosaur Neoaetosauroides engaeus: evidence for aetosaurian diversity. Palaeontology 50:267–276
Desojo JB, Ezcurra MD, Schultz CL (2011) An unusual new archosauriform from the Middle-Late Triassic of southern Brazil and the monophyly of Doswelliidae. Zool J Linnean Soc 161:839–871
Desojo JB, Heckert AB, Martz JW, Parker WG, Schoch RR, Small BJ, Sulej T (2013) Aetosauria: a clade of armoured pseudosuchians from the Upper Triassic continental beds. In: Nesbitt SJ, Desojo JB, Irmis RB (eds) Anatomy, phylogeny and palaeobiology of early archosaurs and their kin. Geol Soc London Spec Publ 379: 203–240
Dias-da-Silva S, Carvalho IC, Schwanke C (2007) Vertebrate dinoturbation from the Caturrita Formation (Late Triassic, Parana basin), Rio Grande do Sul State, Brazil. Gondwana Res 11:303–310
Dias-da-Silva S, Dias EV, Schultz CL (2009) First record of stereospondyls (Tetrapoda, Temnospondyli) in the Upper Triassic of southern Brazil. Gondwana Res 15:131–136
Dilkes D, Sues H-D (2009) Redescription and phylogenetic relationships of Doswellia kaltenbachi (Diapsida: Archosauriformes) from the Upper Triassic of Virginia. J Vert Paleont 29:58–79
Dobruskina IA (1995a) Keuper (Triassic) flora from Middle Asia (Madygen, southern Fergana). New Mex Mus Nat Hist Sci Bull 5:1–49
Dobruskina IA (1995b) Triassic plants and Pangea. Palaeobotanist 44:116–127
Dornelles JEF (1990) Registro sobre a ocorrencia de dentes de um arccossaurio para a Formacao Caturrita, Triassico Superior do Rio Grande do Sul, Brasil. Cie Natur Santa Maria 12:99–101
Doyle KD, Sues HD (1995) Phytosaurs (Reptilia: Archosauria) from the Upper Triassic New Oxford Formation of York County, Pennsylvania. J Vert Paleont 15:545–553
Dutuit JM (1966) Apport des déscovertes de vertébrès à la stratigraphie du Trias continental du coloir d’Argana (Haut-Atlas occidental, Maroc). Maroc Serv Géol Notes 26:29–31
Dutuit JM (1972) Découverte d’un dinosaure ornithischien dans le Trias Supérieur de l’Atlas occidental marocain. Compt Rend Acad Sci D 275:2841–2844
Dutuit JM (1976) Introduction à l’etude paléontologique du Trias continental marocain. Description des premiers stégocéphales recuellis dans le couloir d’Argana. Mém Mus Nat Hist Nat C 36:1–253
Dutuit JM (1977) Paleorhinus magnoculus. Phytosaure du Trias Supérieur de l’Atlas Marocain Anns de Univers Provence 4:255–268
Dutuit JM (1978) Description de quelques fragments osseux provenant de la région de Folakara (Trias supérieur Malgache). Bull Mus Hist Natur 69:79–89
Dutuit JM (1988) Ostéologie crânienne et ses enseignements apports géologique et paléoécologique de Moghreberia nmachouensis, dicynodonte (Reptilia, Therapsida) du Trias Supérieur Marocain. Bull Mus Nat Hist Nat 10:227–285
Dutuit JM (1989a) Azarifeneria barrati, un deuxième genre de dicynodonte du Trias Supérieur Marocain. Compt Rend Acad Sci 309:303–306
Dutuit JM (1989b) Confirmation des affinités entre Trias Supérieurs Marocain et Sud-Americain: Découverte d’un troisième dicynodonte (Reptilia, Therapsida), Azarifeneria robustus, n. sp., de la formation d’Argana (Atlas occidental). Compt Rend Acad Sci 309:1267–1270
Dzik J (2001) A new Paleorhinus fauna in the early Late Triassic of Poland. J Vert Paleont 21:625–627
Dzik J (2003) A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. J Vert Paleont 23:556–574
Dzik J, Sulej T (2007) A review of the Late Triassic Krasiejów biota from Silesia, Poland. Palaeont Polon 64:1–27
Dzik J, Sulej T, Niedźwiedzki G (2008) A dicynodont-theropod association in the latest Triassic of Poland. Acta Palaeont Polon 53:733–738
Emmons E (1856) Geologic report of the midland counties of North Carolina. George P. Putnam and Co., New York
Ezcurra MD (2006) A review of the systematic position of the dinosauriform archosaur Eucoelophysis baldwini Sullivan and Lucas, 1999 from the Upper Triassic of New Mexico, USA. Geodiversitas 28:649–684
Ezcurra MD (2010) Biogeography of Triassic tetrapods: evidence for provincialism and driven sympatric cladogenesis in the early evolution of modern tetrapod lineages. Proc R Soc B 277:2547–2552
Fedak TJ, Sues H-D, Olsen PE (2015) First record of the tritylodontid cynodont Oligokyphus and cynodont postcranial bones from the McCoy Brook Formation of Nova Scotia, Canada. Can J Earth Sci 52:244–249
Ferigolo J, Langer MC (2006) A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone. Hist Biol 18:1–11
Flynn JJ, Wyss AR (2003) Mesozoic terrestrial vertebrate faunas: the early history of Madagascar’s vertebrate diversity. In: Goodman SM, Benstead JP (eds) The natural history of Madagascar. University of Chicago Press, Chicago, pp 34–40
Flynn JJ, Parrish JM, Rakotosamimanana B, Simpson WF, Whatley RL, Wyss AR (1999) A Triassic fauna from Madagascar, including early dinosaurs. Science 286:763–765
Flynn JJ, Parrish JM, Rakotosamimanana B, Ranivoharimanana L, Simpson WF, Wyss AR (2000) New traversodontids (Synapsida: Eucynodontia) from the Triassic of Madagascar. J Vert Paleont 20:422–427
Flynn J, Nesbitt S, Parrish M, Ranivoharimanana L, Wyss A (2008) A new species of basal archosauromorph from the Late Triassic of Madagascar. J Vert Paleont 28(3):78A
Flynn JJ, Nesbitt SJ, Parrish JM, Ranivoharimanana L, Wyss AR (2010) A new species of Azendohsaurus (Diapsida: Archosauromorpha) from the Triassic Isalo Group of southwestern Madagascar: cranium and mandible. Palaeontology 53:669–688
Fraas O (1877) Aetosaurus ferratus Fr. Die gepanzerte Vogel-Echse aus dem Stubensandstein bei Stuttgart. Festschr Feier s vierhundertjähr Jubiläums Eberhard-Karls-Univers Tübingen, Wurttemberg naturwissene Jahresh 33:1–22
Fraser NC (1994) Assemblages of small tetrapods from British Late Triassic fissure deposits. In: Fraser NC, Sues H-D (eds) In the shadow of the dinosaurs. Cambridge University Press, Cambridge, pp 214–226
Fraser N (2006) Dawn of the dinosaurs: life in the Triassic. Indiana University Press, Bloomington
Fraser NC, Padian K, Walkden GM, Davis ALM (2002) Basal dinosauriform remains from Britain and the diagnosis of the Dinosauria. Palaeontology 45:79–95
Gaffney ES (1990) The comparative osteology of the Triassic turtle Proganochelys. Bull Am Mus Nat Hist 194:1–263
Galton PM (2005) Bones of large dinosaurs (Prosauropoda and Stegosauria) from the Rhaetic bone bed (Upper Triassic) of Aust Cliff, southwest England. Rev Paléobiol Genève 24:51–74
Galton PM (2007a) Notes on the remains of archosaurian reptiles, mostly basal sauropodomorph dinosaurs, from the 1834 fissure fill (Rhaetian, Upper Triassic) at Clifton in Bristol, southwest England. Rev Paléobiol Genève 26:505–591
Galton PM (2007b) Pantydraco n. gen. for Thecodontosaurus cauducus Yates, 2003, a basal sauropodomorph dinosaur from the Upper Triassic or Lower Jurassic of South Wales, UK. Neues Jahrb Geol Paläont Abhand 243:119–125
Gauffre F (1993) The prosauropod dinosaur Azendohsaurus laaroussii from the Upper Triassic of Morocco. Palaeontology 36:897–908
Gilmore CW (1928) A new fossil reptile from New Jersey. Proc US Nat Mus 73:1–8
Gower DJ, Schoch RR (2009) Postcranial anatomy of the rauisuchian archosaur Batrachotomus kupferzellensis. J Vert Paleont 29:103–122
Gower DJ, Sennikov AG (2000) Early archosaurs from Russia. In: Benton MJ, Shishkin MA, Unwin DM, Kurochkin EN (eds) The age of dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, pp 140–159
Guth C (1963) Au sujets de restes de reptiles de Madagascar. Compt Rend Acad Sci 25:2661–2663
Harris JD, Downs A (2002) A drepanosaurid pectoral girdle from the Ghost Ranch (Whitaker) Coelophysis quarry (Chinle Group, Rock Point Formation, Rhaetian), New Mexico. J Vert Paleont 22:70–75
Haubold H (1971) Ichnia Amphibiorum et Reptiliorum fossilium. Encyc Paleoherp 18:1–124
Heckert AB (1997) The tetrapod fauna of the Upper Triassic lower Chinle Group (Adamanian: latest Carnian) of the Zuni Mountains, west-central New Mexico. New Mex Mus Nat Hist Sci Bull 11:29–40
Heckert AB (2004) Late Triassic microvertebrates from the lower Chinle Group (Otischalkian-Adamanian: Carnian). New Mex Mus Nat Hist Sci Bull 27:1–170
Heckert AB, Lucas SG (1999) A new aetosaur (Reptilia: Archosauria) from the Upper Triassic of Texas and the phylogeny of aetosaurs. J Vert Paleont 19:50–68
Heckert AB, Lucas SG (2000) Taxonomy, phylogeny, biostratigraphy, biochronology, paleobiogeography, and evolution of the Late Triassic Aetosauria (Archosauria:Crurotarsi). Zentralbl Geol Paläont I 1(11–12):1539–1587
Heckert AB, Lucas SG (2002) Lower Chinle Group (Upper Triassic: Carnian) stratigraphy in the Zuni Mountains, west-central New Mexico. New Mex Mus Nat Hist Sci Bull 21:51–72
Heckert AB, Lucas SG, Hunt AP, Harris JD (2001) A giant phytosaur (Reptilia: Archosauria) skull from the Redonda Formation (Upper Triassic: Apachean) of east-central New Mexico. New Mex Geol Soc Guideb 52:171–178
Heckert AB, Lucas SG, Hunt AP (2005) Triassic vertebrate fossils in Arizona. New Mex Mus Nat Hist Sci Bull 29:16–44
Heckert AB, Spielmann JA, Lucas SG, Hunt AP (2007) Biostratigraphic utility of the Upper Triassic aetosaur Tecovasuchus (Archosauria: Stagonolepididae), an index taxon of St. Johnsian (Adamanian:Late Carnian) time. New Mex Mus Nat Hist Sci Bull 41:51–57
Heckert AB, Lucas SG, Rinehart LF, Hunt AP (2008) A new genus and species of sphenodontian from the Ghost Ranch Coelophysis quarry (Upper Triassic: Apachean), Rock Point Formation, New Mexico, USA. Palaeontology 51:827–845
Heckert AB, Mitchell JS, Schneider VP, Olsen PE (2012) Diverse new microvertebrate assemblage from the Upper Triassic Cumnock Formation, Sanford subbasin, North Carolian, USA. J Paleontol 86:368–390
Hone DWE, Benton MJ (2008) Contrasting supertree and total-evidence methods: the origin of pterosaurs. Zitteliana B28:35–60
Hornung T, Brandner R, Krystyn L, Joachimski MM, Keim L (2007) Multistratigraphic constraints on the NW Tethyan “Carnian crisis”. New Mex Mus Nat Hist Sci Bull 41:59–67
Hsiou A, Abdala F, Arcucci A (2002) Novo registro de proterocampsideo (Reptilia, Archosauriformes) do Triasico medio-superior do Brasil. Revista Brasil Paleontol 5:48–55
Huber P, Lucas SG, Hunt AP (1993a) Revised age and correlation of the Upper Triassic Chatham Group (Deep River basin, Newark Supergroup), North Carolina. Southeast Geol 33:171–193
Huber P, Lucas SG, Hunt AP (1993b) Vertebrate biochronology of the Newark Supergroup Triassic, eastern North America. New Mex Mus Nat Hist Sci Bull 3:179–186
Huber P, Lucas SG, Hunt AP (1993c) Late Triassic fish assemblages of the North American Western Interior and their biochronologic significance. Mus North Ariz Bull 59:51–66
Hungerbühler A (1998) Taphonomy of the prosauropod dinosaur Sellosaurus, and its implications for carnivore faunas and feeding habits in the Late Triassic. Palaeogeogr Palaeoclimatol Palaeoecol 143:1–29
Hungerbühler A (2001) The status and phylogenetic relationships of “Zanclodon” arenaceus: the earliest known phytosaur? Paläont Zeitschr 75:97–112
Hungerbühler A (2002) The Late Triassic phytosaur Mystriosuchus westphali, with a revision of the genus. Palaeontology 45:377–418
Hunt AP (1989) Cranial morphology and ecology among phytosaurs. In: Lucas SG, Hunt AP (eds) Dawn of the age of dinosaurs in the American Southwest. New Mexico Museum of Natural History, Albuquerque, pp 349–354
Hunt AP (1991) The early diversification of dinosaurs in the Late Triassic. Mod Geol 16:43–60
Hunt AP (1993) Revision of the Metoposauridae (Amphibia: Temnospondyli) and description of a new genus from western North America. Mus North Ariz Bull 59:67–97
Hunt AP (1994) Vertebrate paleontology and biostratigraphy of the Bull Canyon Formation (Chinle Group: Norian), east-central New Mexico with revisions of the families Metoposauridae (Amphibia: Temnospondyli) and Parasuchidae (Reptilia: Archosauria). PhD dissertation, University of New Mexico, Albuquerque
Hunt AP (2001) The vertebrate fauna, biostratigraphy and biochronology of the type Revueltian land-vertebrate faunachron, Bull Canyon Formation (Upper Triassic), east-central New Mexico. New Mex Geol Soc Guideb 52:123–151
Hunt AP, Lucas SG (1990) Re-evaluation of “Typothorax” meadei, a Late Triassic aetosaur from the United States. Paläont Zeitschr 64:317–328
Hunt AP, Lucas SG (1991) A new rhynchosaur from the Upper Triassic of West Texas, and the biochronology of Late Triassic rhynchosaurs. Palaeontology 34:927–938
Hunt AP, Lucas SG (1993a) Triassic vertebrate paleontology and biochronology of New Mexico. New Mex Mus Nat Hist Sci Bull 2:49–60
Hunt AP, Lucas SG (1993b) A new phytosaur (Reptilia: Archosauria) genus from the uppermost Triassic of the western United States and its biochronologic significance. New Mex Mus Nat Hist Sci Bull 3:193–196
Hunt AP, Lucas SG (1994) Ornithischian dinosaurs from the Upper Triassic of the United States. In: Fraser NC, Sues HD (eds) In the shadow of the dinosaurs. Cambridge University Press, New York, pp 227–241
Hunt AP, Lucas SG (1997) Stratigraphy, paleontology and biochronology of the Upper Triassic Chinle Group in east-central New Mexico. Southwest Paleont Symp Proc 1:25–40
Hunt AP, Lucas SG (2007) The Triassic tetrapod track record: ichnofaunas, ichnofacies and biochronology. New Mex Mus Nat Hist Sci Bull 41:78–78
Hunt AP, Lucas SG, Heckert AB (2002) A Revueltian (Norian) phytosaur from the Sonsela Member of the Petrified Forest Formation (Chinle Group: Upper Triassic), Petrified Forest National Park, Arizona. New Mex Mus Nat Hist Sci Bull 21:165–169
Hunt AP, Lucas SG, Heckert AB (2005) Definition and correlation of the Lamyan: a new biochronological unit for the nonmarine late Carnian (Late Triassic). New Mex Geol Soc Guideb 56:357–366
Hunt AP, Lucas SG, Klein H (2017) Late Triassic nonmarine vertebrate and invertebrate trace fossils and the pattern of the Phanerozoic record of vertebrate trace fossils. In: Tanner LH (ed) The Late Triassic world: earth in a time of transition. Topics in geobiology. Springer, New York (this volume)
Ivakhhnenko MF, Golubev VK, Gubin YM, Kalandadze NN, Novikov IV, Sennikov AG, Rautian AS (1997) Permskiye i Triasoviye tetrapodi Vostochnoi Evropi [Permian and Triassic tetrapods of Eastern Europe]. GEOS, Moscow
Ivakhnenko MF (1978) Urodelans from the Triassic and Jurassic of Soviet Central Asia. Paleont J 1978(3):84–89. [in Russian]
Jadoul F (1985) Stratigrafia e paleogeografia del Norico nelle Prealpi Bergamasche occidentali. Riv Ital Paleont Stratig 91:479–512
Jadoul F, Masetti D, Cirilli S, Berra S, Claps N, Frisia S (1994) Norian-Rhaetian stratigraphy and paleogeographic evolution of the Lombardy basin (Bergamasc Alps). In: 15th Internat Assoc Sedimentol Reg Meet, April 1994, Ischia, Italy, Field Excurs B1, Salermo, Italy, pp 5–38
Jain SL, Roychowdhury T (1987) Fossil vertebrates from the Godavari Valley (India) and their stratigraphic correlation. In: McKenzie GD (ed) Gondwana Six: stratigraphy, sedimentology and paleontology. Am Geophys Un Monogr 41: 219–228
Jain SL, Robinson PL, Roychowdhury TK (1964) A new vertebrate fauna from the Triassic of Deccan, India. J Geol Soc Lond 120:115–124
Jalil N (1999) Continental Permian and Triassic vertebrate localities from Algeria and Morocco and their stratigraphical correlations. J Afr Earth Sci 29:219–226
Jenkins FA, Shubin NH, Amaral WW, Gatesy SM, Schaff CR, Clemmensen LB, Downs WR, Davidson AR, Bonde N, Osbaeck F (1994) Late Triassic vertebrates and depositional environments of the Fleming Fjord Formation, Jameson Land, east Greenland. Meddel Grønl 32:1–25
Jenkins FA Jr, Gatesy SM, Shubin NH, Amaral WW (1997) Haramiyids and Triassic mammalian evolution. Nature 385:715–718
Jenkins FA Jr, Shubin NH, Gatesy SM, Padian K (2001) A diminutive pterosaur (Pterosauria: Eudimorphodontidae) from the Greenland Triassic. Bull Mus Comp Zool 156:151–170
Jenkins FA Jr, Shubin NH, Gatesy SM, Warren A (2008) Gerrothorax pulcherrimus from the Upper Triassic Fleming Fjord Formation of east Greenland and a reassessment of head lifting in temnospondyl feeding. J Vert Paleont 28:935–950
Joyce WG (2017) A review of the fossil record of basal Mesozoic turtles. Bull Peab Mus Nat Hist 58:65–113
Joyce WG, Lucas SG, Scheyer TM, Heckert AB, Hunt AP (2009) A thin-shelled reptile from the Late Triassic of North America and the origin of the turtle shell. Proc R Soc B276:507–513
Kammerer CF, Flynn JJ, Ranovharimanana L, Wyss AR (2010) The first record of a probainognathian (Cynodontia: Chiniquodontidae) from the Triassic of Madagascar. J Vert Paleont 30:1889–1894
Kammerer CF, Butler RJ, Bandyopadhyay S, Stocker MR (2016) Relationships of the Indian phytosaur Parasuchus hislopi Lydekker, 1885. Papers Palaeontol 2:1–23
Kaye FT, Padian K (1994) Microvertebrates from the Placerias quarry: a window on Late Triassic vertebrate diversity in the American Southwest. In: Fraser NC, Sues HD (eds) The shadow of the dinosaurs. Cambridge University Press, New York, pp 171–196
Kear BP, Poropat SF, Bazzi M (2016) Late Triassic capitosaurian remains from Svalbard and the palaeobiogeographical context of Scandinavian Arctic temnospondyls. In: Kear PB, Lindgren J, Hurm JH, Milàn J, Vajda V (eds) Mesozoic biotas of Scandinavia and its Arctic territories. Geol Soc Lond Spec Publ 434: 113–126
Kent DV, Malnis PS, Colombi CE, Alcober SA, Martinez RN (2014) Age constraints on the dispersal of dinosaurs in the Late Triassic from magnetochronology of the Los Colorados Formation (Argentina). Proc Natl Acad Sci U S A 11:7958–7963
Kent DV, Olsen PE, Muttoni G (2017) Astrochronostratigraphic polarity time scale (ATS) for the Late Triassic and Early Jurassic from continental sediments and correlation with standard marine stages. Earth-Sci Rev 166:153–180
Keyser AW (1973a) A new Triassic vertebrate fauna from South West Africa. South Afr J Sci 69:113–115
Keyser AW (1973b) A new Triassic vertebrate fauna from South West Africa. Palaeont Afr 16:1–15
Keyser AW (1978) A new bauriamorph from the Omongonde Formation (Middle Triassic) of South West Africa. Palaeont Afr 21:177
Kielan-Jaworowska Z, Cifelli RL, Luo Z (2004) Mammals from the age of dinosaurs. Columbia University Press, New York
Kimmig J, Arp G (2010) Phytosaur remains from the Norian Arnstadt Formation (Leine Valley, Germany), with reference to European phytosaur habitats. Palaeodiversity 3:215–224
Kimmig J, Spielmann JA (2011) Biologic factors influencing phytosaur (Archosauria: Phytosauridae) taxonomy. New Mex Mus Nat Hist Sci Bull 53:289–294
Kirby RE (1989) Late Triassic vertebrate localities of the Owl Rock Member (Chinle Formation) in the Ward Terrace area of northern Arizona. In: Lucas SG, Hunt AP (eds) Dawn of the age of dinosaurs in the American Southwest. New Mex Mus Nat Hist, Albuquerque, pp 12–28
Kirby RE (1991) A vertebrate fauna from the Upper Triassic Owl Rock Member of the Chinle Formation in northern Arizona. MS thesis, Northern Arizona University, Flagstaff
Kirby RE (1993) Relationships of Late Triassic basin evolution and faunal replacement events in the southwestern United States: Perspectives from the upper part of the Chinle Formation in northern Arizona. New Mex Mus Nat Hist Sci Bull 3:233–242
Kischlat E, Lucas SG (2003) A phytosaur from the Upper Triassic of Brazil. J Vert Paleont 23:464–467
Kitching JW, Raath MA (1984) Fossils from the Elliot and Clarens Formations (Karoo sequence) of the northern Cape, Orange Free State, and Lesotho, and a suggested biozonation based on tetrapods. Palaeont Afr 25:111–125
Klein H, Lucas SG (2010a) Tetrapod footprints—their use in biostratigraphy and biochronology of the Triassic. In: Lucas SG (ed) The Triassic timescale. Geol Soc Lond Spec Publ 334: 219–446
Klein H, Lucas SG (2010b) The Triassic footprint record of crocodylomorphs—a critical re-evaluation. New Mex Mus Nat Hist Sci Bull 51:55–60
Klein H, Lucas SG (2015) Evolution of the semi-aquatic lifestyle in archosaurs—evidence from the tetrapod footprint record. Geol Assoc Canada Misc Pub 9:105–111
Klein H, Niedzwiedźki G (2012) Revision of the Lower Triassic tetrapod ichnofauna from Wióry, Holy Cross Mountains, Poland. New Mex Mus Nat Hist Sci Bull 56:1–62
Kohút M, Hofmann M, Havrila M, Linnemann U, Havrila J (2017) Tracking an upper limit of the “Carnian crisis” and/or Carnian Stage in the Western Carpathians (Slovakia). Int J Earth Sci. doi: 10.10007/s00531-017-1491-8
Kozur HW, Bachmann GH (2005) Correlation of the Germanic Triassic with the international scale. Albertiana 32:21–35
Kozur HW, Bachmann GH (2008) Updated correlation of the Germanic Triassic with the Tethyan scale and assigned numeric ages. Berichte GeolBundesanstalt 76:53–58
Kozur HW, Weems RE (2005) Conchostracan evidence for a late Rhaetian to early Hettangian age for the CAMP volcanic event in the Newark Supergroup, and a Sevatian (late Norian) age for the immediately underlying beds. Hall Jahrb Geowissen B27:21–51
Kozur HW, Weems RE (2007) Upper Triassic conchostracan biostratigraphy of the continental rift basins of eastern North America: its importance for correlating Newark Supergroup events with the Germanic basin and the international geologic timescale. New Mex Mus Nat Hist Sci Bull 41:137–188
Kozur HW, Weems RE (2010) The biostratigraphic importance of conchostracans in the continental Triassic of the northern hemisphere. In: Lucas SG (ed) The Triassic timescale. Geol Soc Lond Spec Publ 334: 315–417
Krystyn L, Boquerel H, Kuerschner W, Richoz S, Gallet Y (2007a) Proposal for a candidate GSSP for the base of the Rhaetian stage. New Mex Mus Nat Hist Sci Bull 41:189–199
Krystyn L, Richoz S, Gallet Y, Boquerel H, Kurschner W, Spötl C (2007b) Updated bio- and magnetostratigraphy from Steinbergkogel (Austria), candidate GSSP for the base of the Rhaetian stage. Albertiana 36:164–173
Kutty TS (1969) Some contributions to the stratigraphy of the upper Gondwana formations of the Pranhita-Godavari Valley, central India. J Geol Soc India 10:33–48
Kutty TS, Roychowdhury T (1970) The Gondwana sequence of Pranhita-Godavari Valley, India, and its vertebrate faunas. In: Second Gond Symp Proc Pap, pp 303–308
Kutty TS, Sengupta DP (1989) The Late Triassic formations of the Pranhita-Godavari Valley and their vertebrate faunal succession—a reappraisal. Indian J Earth Sci 16:189–206
Kutty TS, Jain SL, Roychowdhury T (1988) Gondwana sequence of the northern Pranhita-Godavari Valley: its stratigraphy and vertebrate faunas. Palaeobotanist 36:263–282
Langer MC (2005a) Studies on continental late Triassic tetrapod biochronology. I. The type locality of Saturnalia tupiniquim and the faunal succession in South Brazil. J S Am Earth Sci 19:205–218
Langer MC (2005b) Studies on continental late Triassic tetrapod biochronology. II. The Ischigualastian and a Carnian global correlation. J S Am Earth Sci 19:219–239
Langer MC (2014) The origins of Dinosauria: much ado about nothing. Palaeontology 2014:1–10
Langer MC, Ferigolo J, Schultz CL (2000) Heterochrony and tooth evolution in hyperodapedontine rhynchosaurs (Reptila, Diapsida). Lethaia 33:119–128
Langer MC, Ribeiro AM, Schultz CL, Ferigolo J (2007) The continental tetrapod-bearing Triassic of southern Brazil. New Mex Mus Nat Hist Sci Bull 41:201–218
Langer MC, Ezcurra M, Bittencourt JS, Novas F (2009) The origin and early evolution of dinosaurs. Biol Rev 85:55–110
Lea I (1851) Remarks on the bones of a fossil reptilian quadruped. Proc Acad Nat Sci Phil 5:171–172
Leal LA, Azevedo SK, Kellner AW, Da Rosa AS (2003) A new early dinosaur (Sauropodomorpha) from the Caturrita Formation (Late Triassic), Paraná basin, Brazil. Zootaxa 690:1–24
Li C, Wu X, Rieppel O, Wang L, Zhao L (2008) An ancestral turtle from the Late Triassic of southwestern China. Nature 456:491–501
Lichtig AJ, Lucas SG, Klein H, Lovelace DM (2017) Triassic turtle tracks and the origin of turtles. Hist Biol. https://doi.org/10.1080/08912963.2017.1339037
Liu J, Abdala FA (2014) Phylogeny and taxonomy of the Traversodontidae. In: Kammerer CF, Angielczyk KD, Fröbisch J (eds) Early evolutionary history of the Synapsida. Springer, Dordrecht, pp 255–279
Lockley MG, Lucas SG, Milan J, Harris JD, Avanzini M, Foster JR, Spielmann JA (2010) The fossil record of crocodilian tracks and traces: an overview. New Mex Mus Nat Hist Sci Bull 51:1–13
Long RA, Murry PA (1995) Late Triassic (Carnian and Norian) tetrapods from the southwestern United States. New Mex Mus Nat Hist Sci Bull 4:1–254
Lovelace DM, Lovelace SD (2012) Paleoenvironments and paleoecology of a Lower Triassic invertebrate and vertebrate ichnoassemblage from the Red Peak Formation (Chugwater Group), central Wyoming. PALAIOS 27:636–657
Lucas SG (1992) Extinction and the definition of the Class Mammalia. Syst Biol 41:370–371
Lucas SG (1993) The Chinle Group: revised stratigraphy and chronology of Upper Triassic nonmarine strata in the western United States. Mus North Ariz Bull 59:27–50
Lucas SG (1994) Triassic tetrapod extinctions and the compiled correlation effect. Can Soc Petrol Geol Mem 17:869–875
Lucas SG (1997) Upper Triassic Chinle Group, western United States: a nonmarine standard for Late Triassic time. In: Dickins JM et al (eds) Late Paleozoic and early Mesozoic Circum-Pacific events and their global correlation. Cambridge University Press, Cambridge, pp 209–228
Lucas SG (1998) Global Triassic tetrapod biostratigraphy and biochronology. Palaeogeogr Palaeoclimatol Palaeoecol 143:347–384
Lucas SG (1999) Tetrapod-based correlation of the nonmarine Triassic. Zentralbl r Geol Paläont Teil I 7-8:497–521
Lucas SG (2002) A new dicynodont from the Triassic of Brazil and the tetrapod biochronology of the Brazilian Triassic. New Mex Mus Nat Hist Sci Bull 21:131–141
Lucas SG (2007) Tetrapod footprint biostratigraphy and biochronology. Ichnos 14:5–38
Lucas SG (2008) Global Jurassic tetrapod biochronology. Volum Jura 6:99–108
Lucas SG (2010) The Triassic timescale based on nonmarine tetrapod biostratigraphy and biochronology. In: Lucas SG (ed) The Triassic timescale. Geol Soc Lond Spec Publ 334: 447–500
Lucas SG (2015) Age and correlation of Late Triassic tetrapods from southern Poland. Ann Soc Geol Polon 85:627–635
Lucas SG (2017a) Permian tetrapod extinction events. Earth-Sci Rev 170:31–60
Lucas SG (2017b) The Late Triassic timescale. In: Tanner LH (ed) The Late Triassic world: earth in a time of transition. Topics in geobiology. Springer, New York (this volume)
Lucas SG, Hancox J (2001) Tetrapod-based correlation of the nonmarine Upper Triassic of Southern Africa. Albertiana 25:5–9
Lucas SG, Harris SK (1996) Taxonomic and biochronological significance of specimens of the Triassic dicynodont Dinodontosaurus Romer, 1943 in the Tübingen collection. Paläont Zeitschr 70:603–622
Lucas SG, Heckert AB (2000) Biochronological significance of Triassic nonmarine tetrapod records from marine strata. Albertiana 24:27–32
Lucas SG, Heckert AB (2001) The aetosaur Stagonolepis from the Upper Triassic of Brazil and its biochronological significance. Neues Jahrb Geol Paläont Monatsh 2001:719–732
Lucas SG, Heckert AB (2011) Late Triassic aetosaurs as the trackmaker of the tetrapod footprint ichnotaxon Brachychirotherium. Ichnos 18:197–208
Lucas SG, Huber P (2003) Vertebrate biostratigraphy and biochronology of the nonmarine Triassic. In: LeTourneau PM, Olsen PE (eds) The great rift valleys of Pangea in eastern North America, vol 2. Sedimentology and paleontology. Columbia University Press, New York, pp 143–191
Lucas SG, Hunt AP (1992) Triassic stratigraphy and paleontology, Chama basin and adjacent areas, north-central New Mexico. New Mex Geol Soc Guideb 43:151–172
Lucas SG, Hunt AP (1993a) Tetrapod biochronology of the Chinle Group (Upper Triassic), western United States. New Mex Mus Nat Hist Sci Bull 3:327–329
Lucas SG, Hunt AP (1993b) A review of Triassic labyrinthodont amphibians from China. Geobios 26:121–128
Lucas SG, Hunt AP (1994) The chronology and paleobiogeography of mammalian origins. In: Fraser NC, Sues HD (eds) The shadow of the dinosaurs. Cambridge University Press, New York, pp 335–351
Lucas SG, Luo Z (1993) Adelobasileus from the Upper Triassic of West Texas: the oldest mammal. J Vert Paleont 13:309–334
Lucas SG, Spielmann JA (2013) Magnetostratigraphy of the Upper Triassic Chinle Group in New Mexico: an appraisal of 40 years of analysis. New Mex Mus Nat Hist Sci Bull 61:375–381
Lucas SG, Sullivan RM (1997) Fossils provide a Pennsylvania standard for part of Late Triassic time. Penn Geol 27(4):8–14
Lucas SG, Sullivan RM (2006) Tetrapod footprints from the Upper Triassic Passaic Formation near Graterford, Montgomery County, Pennsylvania. New Mex Mus Nat Hist Sci Bull 37:251–256
Lucas SG, Tanner LH (2007a) Tetrapod biostratigraphy and biochronology of the Triassic-Jurassic transition on the southern Colorado Plateau, USA. Palaeogeogr Palaeoclimatol Palaeoecol 244:242–256
Lucas SG, Tanner LH (2007b) The nonmarine Triassic-Jurassic boundary in the Newark Supergroup of eastern North America. Earth-Sci Rev 84:1–20
Lucas SG, Tanner LH (2015) End-Triassic nonmarine biotic events. J Palaeogeogr 4:331–348
Lucas SG, Wild R (1995) A Middle Triassic dicynodont from Germany and the biochronology of Triassic dicynodonts. Stuttgart Beitr Naturk 220:1–16
Lucas SG, Hunt AP, Long RA (1992) The oldest dinosaurs. Naturwiss 79:171–172
Lucas SG, Hunt AP, Kahle RW (1993) Late Triassic vertebrates from the Dockum Formation near Otis Chalk, Howard County, Texas. New Mex Geol Soc Guideb 44:237–244
Lucas SG, Anderson OJ, Hunt AP (1994) Triassic stratigraphy and correlations, southern High Plains of New Mexico-Texas. New Mex Bur Mines Min Res Bull 150:105–126
Lucas SG, Heckert AB, Hunt AP (1997) Stratigraphy and biochronology of the Late Triassic Placerias quarry, eastern Arizona (U.S.A.) Neues Jahrb Geol Paläont Abhand 203:23–46
Lucas SG, Wild R, Hunt AP (1998) Dyoplax O. FRAAS, a Triassic sphenosuchian from Germany. Stuttg Beitr Naturk B263:1–13
Lucas SG, Heckert B, Fraser NC, Huber P (1999) Aetosaurus from the Upper Triassic of Great Britain and its biochronological significance. Neues Jahrb Geol Paläont Monats 1999:568–576
Lucas SG, Heckert AB, Hunt AP (2000) Probable turtle from the Upper Triassic of east-central New Mexico. Neues Jahrb Geol Palaeont Monats 5:287–300
Lucas SG, Heckert AB, Hunt AP (2001) Triassic stratigraphy, biostratigraphy and correlation in east-central New Mexico. New Mex Geol Soc Guideb 52:85–101
Lucas SG, Heckert AB, Hotton N III (2002) The rhynchosaur Hyperodapedon from the Upper Triassic of Wyoming and its global biochronological significance. New Mex Mus Nat Hist Sci Bull 21:149–156
Lucas SG, Zeigler KE, Heckert AB, Hunt AP (2003) Upper Triassic stratigraphy and biostratigraphy, Chama basin, north-central New Mexico. New Mex Mus Nat Hist Sci Bull 24:15–39
Lucas SG, Tanner LH, Heckert AB (2005) Tetrapod biostratigraphy and biochronology across the Triassic-Jurassic boundary in northeastern Arizona. New Mex Mus Nat Hist Sci 29:84–94
Lucas SG, Hunt AP, Heckert AB, Spielmann JA (2007a) Global Triassic tetrapod biostratigraphy and biochronology: 2007 status. New Mex Mus Nat Hist Sci Bull 41:229–240
Lucas SG, Spielmann JA, Hunt AP (2007b) Biochronological significance of Late Triassic tetrapods from Krasiejów, Poland. New Mex Mus Nat Hist Sci Bull 41:248–258
Lucas SG, Rinehart LF, Krainer K, Spielmann JA, Heckert AB (2010) Taphonomy of the Lamy amphibian quarry: a Late Triassic bonebed in New Mexico, USA. Palaeogeogr Palaeoclimatol Palaeoecol 298:388–398
Lucas SG, Tanner LH, Kozur HW, Weems RE, Heckert AB (2012) The Late Triassic timescale: age and correlation of the Carnian-Norian boundary. Earth-Sci Rev 114:1–18
Lucas SG, Spielmann JA, Hunt AP (2013) A new doswelliid archosauromorph from the Upper Triassic of West Texas. New Mex Mus Nat Hist Sci Bull 61:382–388
Lucas SG, Rinehart LF, Heckert AB, Hunt AP, Spielmann JA (2016) Rotten Hill: a Late Triassic bonebed in the Texas Panhandle, USA. New Mex Mus Nat Hist Sci Bull 72:1–97
Luo Z (2011) Transformation and diversification in early mammal evolution. Nature 450:1011–1019
Lyson TR, Bever GS, Bhullar BS, Joyce WG, Gauthier JA (2010) Transitional fossils and the origin of turtles. Biol Lett. rsbl.20100371
Lyson TR, Sperling EA, Heimberg AM, Gauthier JA, King BL, Peterson KJ (2012) MicroRNAs support a turtle+lizard clade. Biol Lett 8:104–107
Lyson TR, Bever GS, Scheyer TM, Hsiang AY, Gauthier JA (2013) Evolutionary origin of the turtle shell. Curr Biol 23:1113–1119
Lyson TR, Rubidge BS, Scheyer TM, de Queiroz K, Schachner ER, Smith RM, Bever GS (2016) Fossorial origin of the turtle shell. Curr Biol 26:1887–1894
Maisch MW, Kapitzke M (2010) A presumably marine phytosaur (Reptilia: Archosauria) from the pre-planorbis beds (Hettangian) of England. Neues Jahrb Geol Paläont Abhand 257:373–379
Mancuso AC, Gaetano LC, Leardi JM, Abdala F, Arcucci AB (2014) The Chañares Formation: a window to a Middle Triassic tetrapod community. Lethaia 47:242–265
Marsicano CA, Domnanovich NS, Mancuso AC (2007) Dinosaur origins: evidence from the footprint record. Hist Biol 19:83–91
Marsicano CA, Irmis RB, Mancuso AC, Mundil R, Chemale F (2016) The precise temporal calibration of dinosaur origins. Proc Natl Acad Sci U S A 113:509–513
Martínez RN, Apaldetti C, Alcober OA, Colombi CE, Sereno PC, Fernandez E, Malnis PS, Correa GA, Abelin D (2013) Vertebrate succession in the Ischigualasto Formation. J Vert Paleont Mem 12:10–20
McGhee GR Jr, Sheehan PM, Bottjer DJ, Droser ML (2004) Ecological ranking of Phanerozoic biodiversity crises: ecological and taxonomic severities are decoupled. Palaeogeogr Palaeoclimatol Palaeoecol 211:289–297
Mehl MG (1928) The Phytosauria of the Wyoming Triassic. Denison Univ Bull J Sci Lab 23:141–172
Melo TP, Abadala F, Soares MB (2015) The Malagasy cynodont Menadon besairei (Cynodontia: Traversodontidae) in the Middle-Upper Triassic of Brazil. J Vert Paleont. https://doi.org/10.1080/027724634.2014.1002562
Miao D (1991) On the origins of mammals. In: Schultze H-P, Trueb L (eds) Origins of the higher groups of tetrapods: controversy and consensus. Comstock Publishing Associates, London, pp 579–597
Mietto P, Manfrin S, Preto N, Rigo M, Roghi G, Furin S, Gianolla P, Posenato R, Muttoni G, Nicora A, Buratti N, Cirilli S, Spitl C, Ramezani J, Bowring SA (2012) The global boundary stratotype section and point (GSSP) of the Carnian stage (Late Triassic) at Prati di Stuores/Stuores Wiesen section (southern Alps, NE Italy). Episodes 35:414–430
Milner AR (1990) The radiations of temnospondyl amphibians. In: Taylor PD, Larwood GP (eds) Major evolutionary radiations. System Assoc Spec Vol 42: 321–349
Milner AR (1993) Amphibian-grade Tetrapoda. In: Benton MJ (ed) The fossil record. Chapman and Hall, London, pp 665–679
Montefeltro FC, Langer MC, Schultz CL (2010) Cranial anatomy of a new genus of hyperodapedontine rhynchosaur (Diapsida, Archosauromorpha) from the Upper Triassic of southern Brazil. Earth Environ Sci Trans Royal Soc Edinb 101:27–52
Montefeltro FC, Bittencourt JS, Langer MC, Schultz CL (2013) Postcranial anatomy of the hyperodapedontine rhynchosaur Teyumbatia sulcognathus (Azevedo and Schultz, 1987) from the Late Triassic of southern Brazil. J Vert Paleont 33:67–84
Mukherjee D, Ray S, Chandra S, Pal S, Bandyopadhyay S (2012) Upper Gondwana succession of the Rewa basin, India: understanding the interrelationship of lithologic and stratigraphic variables. J Geol Soc India 79:563–575
Mundil R, Pálfy J, Renne PR, Brack P (2010) The Triassic timescale: a review of geochronological constraints. In: Lucas, SG (ed) The Triassic timescale. Geol Soc Lond Spec Publ 334: 41–60
Murry PA (1986) Vertebrate paleontology of the Dockum Group, western Texas and eastern New Mexico. In: Padian K (ed) The beginning of the age of dinosaurs. Cambridge University Press, Cambridge, pp 109–137
Murry PA (1989) Geology and paleontology of the Dockum Formation (Upper Triassic), West Texas and eastern New Mexico. In: Lucas SG, Hunt AP (eds) Dawn of the age of dinosaurs in the American Southwest. New Mex Mus Nat Hist, Albuquerque, pp 102–144
Murry PA, Long RA (1989) Geology and paleontology of the Chinle Formation, Petrified Forest National Park and vicinity, Arizona and a discussion of vertebrate fossils of the southwestern Upper Triassic. In: Lucas SG, Hunt AP (eds) Dawn of the age of dinosaurs in the American Southwest. New Mex Mus Nat Hist, Albuquerque, pp 29–64
Muttoni G, Kent DV, Olsen PE, DiStefano P, Lowrie W, Bernasconi SM, Hernández FM (2004) Tethyan magnetostratigraphy from Pizza Mondelo (Sicily) and correlation to the Late Triassic Newark astrochronological polarity time scale. Geol Soc Am Bull 116:1043–1058
Muttoni G, Kent DV, Jadoul F, Olsen PE, Rigo M, Galli MT, Nicora A (2010) Rhaetian magneto-biostratigraphy from the Southern Alps (Italy). Constraints on Triassic chronology. Palaeogeogr Palaeoclimatol Palaeoecol 285:1–16
Nesbitt SJ (2007) The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa. Bull Am Mus Nat Hist 302:1–84
Nesbitt SJ, Ezcurra MD (2015) The early fossil record of dinosaurs in North America: a new neotheropod from the base of the Upper Triassic Dockum Group of Texas. Acta Palaeont Polon 60:513–526
Nesbitt SJ, Stocker MR (2008) The vertebrate assemblage of the Late Triassic Canjilon quarry (northern New Mexico, USA), and the importance of apomorphy-based assemblage comparisons. J Vert Paleont 28:1063–1072
Niedźwiedzki G, Sulej T, Dzik J (2012) A large predatory archosaur from the Late Triassic of Poland. Acta Palaeont Polon 57:267–276
Niedźwiedzki G, Brusatte SL, Sulej T, Butler RJ (2014) Basal dinosauriform and theropod dinosaurs from the mid–late Norian (Late Triassic) of Poland: implications for Triassic dinosaur evolution and distribution. Palaeontology 57:1121–1142
Ogg JG (2012) Triassic. In: Gradstein FM, Ogg JG, Schmitz MD, Ogg GM (eds) The geologic time scale 2012. Elsevier, Amsterdam, pp 681–730
Ogg JG, Huang C, Hinnov L (2014) Triassic timescale status: a brief overview. Albertiana 41:3–30
Olsen PE (1980) A comparison of vertebrate assemblages from the Newark and Hartford basins (early Mesozoic, Newark Supergroup) of eastern North America. In: Jacobs LL (ed) Aspects of vertebrate history: essays in honor of Edwin Harris Colbert. Museum of Northern Arizona Press, Flagstaff, pp 35–53
Olsen PE (1988) Paleontology and paleoecology of the Newark Supergroup (early Mesozoic, eastern North America). In: Manspeizer W (ed) Triassic-Jurassic rifting: continental breakup and the origin of the Atlantic Ocean and passive margins, Part A. Elsevier, Amsterdam, pp 185–230
Olsen PE, Schlische RW, Gore PJW (1989) Tectonic, depositional and paleoecological history of early Mesozoic rift basins, eastern North America. In: American Geophysical Union Guidebook T351, International Geological Congress, Washington, DC
Orchard MJ (2010) Triassic conodonts and their role in stage boundary definition. In: Lucas SG (ed) The Triassic timescale. Geol Soc Lond Spec Publ 334: 139–161
Orchard MJ (2014) Conodonts from the Carnian-Norian boundary (Upper Triassic) of Black Bear Ridge, northeastern British Columbia, Canada. New Mex Mus Nat Hist Sci Bull 64:1–139
Ottone EG, Monti M, Marsicano CA, de la Fuente M, Naipauer M, Armstrong R, Mancuso AC (2014) A new Late Triassic age for the Puesto Viejo Group (San Rafael depocenter, Argentina): SHRIMP U-Pb zircon dating and biostratigraphic correlation across southern Gondwana. J S Am Earth Sci 56:186–199
Padian K (1984) The origin of the pterosaurs. In: Reif WE, Westphal F (eds) Third symposium on mesozoic terrestrial ecosystems, short papers. Attempto Verlag, Tübingen, pp 163–168
Parker WG, Ash SR, Irmis RB (2006) A century of research at Petrified Forest National Park 1906-2006. Mus North Ariz Bull 62
Parrish MJ (1991) A new specimen of an early crocodylomorph (cf. Sphenosuchus sp.) from the Upper Triassic Chinle Formation of Petrified Forest National Park. J Vert Paleont 11:198–212
Peyer K, Carter JG, Sues H-D, Novak SE, Olsen PE (2008) A new suchian archosaur from the Upper Triassic of North Carolina. J Vert Paleont 28:363–381
Philipp RP, Kloss HP, Schultz CL, Basei MAS, Horn BLD, Soares MB (2013) Provenência por U-Pb LA-ICP-MS em zircão detrítico e ipade de deposição da Formacão Santa Maria, Triassico da bacia do Paraná, RS: Evidências de estructuração do arco do Rio Grande. Anais do VIII Symp Internat Tectonics and XIV Simp Na Estud Tectón, Cuiabá, pp 154–157
Pickford M (1995) Karoo Supergroup palaeontology of Namibia and brief description of a thecodont from Omingonde. Palaeont Afr 32:51–66
Pinna G (1993) The Norian reptiles of northern Italy. Paleont Lombard Nuova Serie 2:115–124
Rage J-C, Roček Z (1989) Redescription of Triadobatrachus massinoti (Piveteau, 1936) an anuran amphibian from the Early Triassic. Palaeontograph Abt A 206:1–16
Rainforth EC (2003) Revision and re-evaluation of the Early Jurassic dinosaurian ichnogenus Otozoum. Palaeontology 46:803–838
Ramezani J, Hoke GD, Fastovsky DE, Bowring SA, Therrien F, Dworkin SI, Atchley SC, Nordt LC (2011) High-precision U-Pb zircon geochronology of the Late Triassic Chinle Formation, Petrified Forest National Park (Arizona, USA): temporal constraints on the early evolution of dinosaurs. Geol Soc Amer Bull 123:2142–2159
Ramezani J, Fastovsky DE, Bowring SA (2014) Revised chronostratigraphy of the lower Chinle Formation strata in Arizona and New Mexico (USA): high-precision U-Pb geochronological constraints on the Late Triassic evolution of dinosaurs. Am J Sci 314:981–1008
Ranovharimanana L, Kammerer CF, Flynn JJ, Wyss AR (2011) New material of Dadadon isaloi (Cynodontia, Traversodontidae) from the Triassic of Madagascar. J Vert Paleont 31:1292–1302
Raugust T, Lacerda M, Schultz CL (2013) The first occurrence of Chanaresuchus bonapartei Romer 1971 (Archosauriformes, Proterochampsia) of the Middle Triassic of Brazil from the Santacruzodon Assemblage Zone, Santa Maria Formation (Paraná basin). In: Nesbitt SJ, Desojo JB, Irmis RB (eds) Anatomy, phylogeny and paleobiology of early archosaurs and their kin. Geol Soc Lond Spec Publ 379: 303–318
Reichel M, Schultz CL, Soares MB (2009) A new traversodontid cynodont (Therapsida, Eucynodontia) from the Middle Triassic Santa Maria Formation of Rio Grande do Sul, Brazil. Palaeontology 52:229–250
Reig OA (1959) Primeros datos descríptivos sobre nuevos reptiles arcosaurios del Triásico de Ischigualasto (San Juan, Argentina). Rev Assoc Geol Argent 13:257–270
Reig OA (1961) Acerca de la posición sistemática de la familia Rauisuchidae y del género Saurosuchus (Reptilia, Thecodontia). Public Mus Cienc Natur Mar del Plata 1:73–114
Reig OA (1963) La presencia de dinosaurios saurisquios en los “Estratos de Ischigualasto” (Mesotriásico superior) de las provincias de San Juan y La Rioja (República Argentina). Ameghin 3:3–20
Renesto S (2006) A reappraisal of the diversity and biogeographic significance of the Norian (Late Triassic) reptiles from the Calcare di Zorzino. New Mex Mus Nat Hist Sci Bull 37:445–456
Renesto S, Spielmann JA, Lucas SG (2009) The oldest record of drepanosaurids (Reptilia, Diapsida) from the Late Triassic (Adamanian Placerias quarry, Arizona, USA) and the stratigraphic range of the Drepanosauridae. Neues Jahrb Geol Paläont Abhand 252:315–325
Renesto S, Spielmann JA, Lucas SG, Spagnoli G (2010) The taxonomy and paleobiology of the late Triassic (Carnian-Norian: Adamanian-Apachean) drepanosaurs (Diapsida: Archosauromorpha: Drepanosauromorpha). New Mex Mus Nat Hist Sci Bull 46:1–81
Rigo M, Bertinelli A, Concheri G, Gattolin G, Godfrey L, Katz ME, Maron M, Mietto P, Muttoni G, Sprovieri M, Stellin F, Zaffani M (2016) The Pignola-Abriola section (southern Apennines, Italy): a new GSSP candidate for the base of the Rhaetian stage. Lethaia 49:287–306
Rinehart LF, Lucas SG, Heckert AB, Spielmann JA, Celeskey MD (2009) The paleobiology of Coelophysis bauri (Cope) from the Upper Triassic (Apachean) Whitaker quarry, New Mexico, with detailed analysis of a single quarry block. New Mex Mus Nat Hist Sci Bull 45:1–260
Rogers RR, Swisher CC III, Sereno PC, Monett AM, Forster CA, Martinez RC (1993) The Ischigualasto tetrapod assemblage (Late Triassic, Argentina) and 40Ar/39Ar dating of dinosaur origins. Science 260:794–797
Romer AS (1966) Vertebrate paleontology, 3rd edn. Univ Chicago Press, Chicago
Romer AS (1973) The Chanares (Argentina) Triassic reptile fauna. XX. Summary. Breviora 413:1–20
Rougier GW, de la Fuente MS, Arcucci AB (1995) Late Triassic turtles from South America. Science 268:855–858
Rowe T (1988) Definition, diagnosis and origin of Mammalia. J Vert Paleont 8:241–264
Rowe T, Gauthier J (1992) Ancestry, paleontology, and definition of the name Mammalia. Syst Biol 41:372–378
Roychowdhury T (1965) A new metoposaurid amphibian from the Triassic Maleri Formation, central India. Phil Trans R Soc Lond B250:1–52
Rubert RR, Schultz CL (2004) Um novo horizonte de correlação para o Triássico superior do Rio Grande do Sul. Pesquisas 31:71–88
Ruffell A, Simms MJ, Wignall PB (2016) The Carnian humid episode of the Late Triassic: a review. Geol Mag 153(2):271–284
Rühle v. Lilienstern H (1939) Fährten und Spüren im Chirotheriem−Sandstein von Südthüringen. Fortschr Geol Paläont 12:293–387
Ruta M, Benton MJ (2008) Calibrated diversity, tree topology and the mother of mass extinctions: the lesson of temnospondyls. Palaeontology 51:1261–1288
Ruta M, Botha-Brink J, Mitchell SA, Benton MJ (2013) The radiation of cynodonts and the ground plan of mammalian morphological diversity. Proc R Soc B280:20131865
Schoch RR (1997) A new capitosaur amphibian from the Upper Lettenkeuper (Triassic: Ladinian) of Kupferzell (southern Germany). Neues Jahrb Geol Paläont Abhand 203:239–272
Schoch RR (2000) Biogeography and dispersal of stereospondyl amphibians. Neues Jahrb Geol Paläont Abhand 215:201–231
Schoch RR (2007) Osteology of the small archosaur Aetosaurus from the Upper Triassic of Germany. Neues Jahrb Geologie Paläont Abhand 246:1–35
Schoch RR (2008) The Capitosauria (Amphibia): characters, phylogeny, and stratigraphy. Palaeodiversity 1:189–226
Schoch RR (2014) Amphibian evolution: the life of early land vertebrates. Wiley Blackwell, Chichester
Schoch RR, Milner AR (2000) Stereospondyli. Encycl Paleoherp 3B:1–203
Schoch RR, Sues H-D (2015) A Middle Triassic stem-turtle and the evolution of the turtle body plan. Nature 523(7562):584–587. https://doi.org/10.1038/nature14472
Schoch RR, Werneburg R (1999) The Triassic labyrinthodonts from Germany. Zentralbl Geol Paläont Teil I 1998:629–650
Schoch R, Voigt S, Buchwitz M (2010) A chroniosuchid from the Triassic of Kyrgyzstan and analysis of chroniosuchian relationships. Zool J Linnean Soc 160:515–530
Schultz CL, Langer MC, Montefeltro FC (2016) A new rhynchosaur from south Brazil (Santa Maria Formation) and rhynchosaur diversity patterns across the Middle-Late Triassic boundary. Paläont Zeitschr 90(3):593–609. https://doi.org/10.1007/s12542-016-0307-7
Sengupta S (1970) Gondwana sedimentation around Bheemaram (Bhimaram), Pranhita-Godavari Valley, India. J Sediment Petrol 40:140–170
Sengupta DP (1995) Chigutisaurid temnospondyls from the Late Triassic of India and a review of the Family Chigutisauridae. Palaeontology 38:313–339
Sereno PC (1991) Basal archosaurs: phylogenetic relationships and functional implications. J Vert Paleont Mem 2:1–53
Sereno PC, Arcucci AB (1993) Dinosaurian precursors from the Middle Triassic of Argentina: Lagerpeton chanarensis. J Vert Paleont 13:385–399
Sereno PC, Arcucci AB (1994) Dinosaurian precursors from the Middle Triassic of Argentina: Marasuchus lilloensis, gen. nov. J Vert Paleont 14:53–73
Sereno PC, Wild R (1992) Procompsognathus: Theropod, “thecodont” or both ? J Vert Paleont 12:435–458
Sereno PC, Forster CA, Rogers RR, Monette AM (1993) Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria. Nature 361:64–66
Sharov AG (1970) A peculiar reptile from the Lower Triassic of Fergana. Paleont J 1970(1):127–130. [in Russian]
Sharov AG (1971) New flying reptiles from the Mesozoic of Kazakhstan and Kyrgyzstan. Dokl Akad Nauk SSSR 130:104–113. [in Russian]
Shishkin MA, Ochev VG, Tverdokhlebov VP (eds) (1995) Biostratigrafiya kontinentalnovo Triasa yuzhovo Priuralya [Biostratigraphy of the continental Triassic of the southern pre-Urals]. Nauka, Moscow. [in Russian]
Shishkin MA, Novikov IV, Gubin YM (2000a) Permian and Triassic temnospondyls from Russia. In: Benton MJ, Shishkin MA, Unwin DM, Kurochkin EN (eds) The age of dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, pp 35–59
Shishkin MA, Ochev VG, Lozovskii VR, Novikov IV (2000b) Tetrapod biostratigraphy of the Triassic of eastern Europe. In: Benton MJ, Shishkin MA, Unwin DM, Kurochkin EN (eds) The age of dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, pp 120–139
Shubin N, Sues H-D (1991) Biogeography of early Mesozoic continental tetrapods: patterns and implications. Paleobiology 17:214–230
Smith RMH, Swartt R (2002) Changing fluvial environments and vertebrate taphonomy in response to climatic drying in a mid-Triassic rift valley fill: the Omingonde Formation (Karoo Supergroup) of central Namibia. PALAIOS 17:249–267
Smith RMH, Rubidge B, van der Walt M (2012) Therapsid biodiversity patterns and paleoenvironments of the Karoo basin, South Africa. In: Chinsamy-Turan A (ed) Forerunners of mammals. Radiation. Histology. Biology. Indiana Univ Press, Bloomington, pp 31–62
Soares MB, Schultz CL, Horn BLD (2011) New information on Riograndia guaibensis Bonaparte, Ferigolo & Ribeiro, 2011 (Eucynodontia, Trithelodontidae) from the Late Triassic of southern Brazil: anatomical and biostratigraphic implications. Anais Acada Brasil Ciênc 83:329–354
Spielmann JA, Lucas SG (2012) Tetrapod fauna of the Upper Triassic Redonda Formation, east-central New Mexico: the characteristic assemblage of the Apachean land-vertebrate faunachron. New Mex Mus Nat Hist Sci Bull 55:1–119
Spielmann JA, Hunt AP, Lucas SG, Heckert AB (2006a) Revision of Redondasuchus (Archosauria: Aetosauria) from the Upper Triassic Redonda Formation, New Mexico, with description of a new species. New Mex Mus Nat Hist Sci Bull 37:583–587
Spielmann JA, Lucas SG, Hunt AP (2006b) The vertebrate macrofauna of the Upper Triassic (Apachean) Redonda Formation, east-central New Mexico. New Mex Mus Nat Hist Sci Bull 37:502–509
Spielmann JA, Lucas SG, Hunt AP, Heckert AB (2006c) Reinterpretation of the holotype of Malerisaurus langstoni, a diapsid reptile from the Upper Triassic Chinle Group of West Texas. New Mex Mus Nat Hist Sci Bull 37:543–547
Spielmann JA, Lucas SG, Heckert AB (2007) Tetrapod fauna of the Upper Triassic (Revueltian) Owl Rock Formation, Chinle Group, Arizona. New Mex Mus Nat Hist Sci Bull 41:371–383
Spielmann JA, Lucas SG, Rinehart LF, Heckert AB (2008) The Late Triassic archosauromorph Trilophosaurus. New Mex Mus Nat Hist Sci Bull 43:1–177
Spielmann JA, Lucas SG, Hunt AP (2013) The first Norian (Revueltian) rhynchosaur: Bull Canyon Formation, New Mexico, U.S.A. New Mex Mus Nat Hist Sci Bull 61:562–565
Sprinkle DA, Kowallis BJ, Jensen PH (2011) Correlation and age of the Nugget Sandstone and Glen Canyon Group, Utah. Utah Geol Assoc Publ 40:131–149
Stocker MR (2012) A new taxonomic arrangement for Paleorhinus scurriensis. Earth Environ Trans R Soc Edinb 103:251–263
Stocker MR, Butler RJ (2013) Phytosauria. In: Nesbitt SJ, Desojo JB, Irmis RB (eds) Anatomy, phylogeny and palaeobiology of early archosaurs and their kin. Geol Soc Lond Spec Publ 379: 91–117
Stocker MR, Zhao L, Nesbitt SJ, Wu X, Li C (2017) A short-snouted, Middle Triassic phytosaur and its implications for the morphological evolution and biogeography of Phytosauria. Nat Sci Rep 7:46028. https://doi.org/10.1038/srep46028
Sues H-D (1992) A remarkable new armored archosaur from the Upper Triassic of Virginia. J Vert Paleont 12:142–149
Sues H-D, Baird D (1993) A skull of a sphenodontian lepidosaur from the New Haven Arkose (Upper Triassic: Norian) of Connecticut. J Vert Paleont 13:370–372
Sues H-D, Fraser NC (2010) Triassic life on land: the great transition. Columbia University Press, New York
Sues H-D, Schoch RR (2013) Anatomy and phylogenetic relationships of Calamops paludosus (Temnospondyli, Stereospondyli) from the Triassic of the Newark basin, Pennsylvania. J Vert Paleont 35:1061–1070
Sues H-D, Olsen PE, Kroehler PA (1994) Small tetrapods from the Upper Triassic of the Richmond basin (Newark Supergroup), Virginia. In: Fraser NC, Sues H-D (eds) In the shadow of the dinosaurs. Cambridge University Press, New York, pp 161–170
Sues H-D, Baird D, Olsen PE (1997) Procolophonidae (Amniota: Parareptilia) from the Upper Triassic of Nova Scotia, Canada. J Vert Paleont 17(3):79A
Sues H-D, Olsen PE, Carter JG (1999) A Late Triassic traversodont cynodont from the Newark Supergroup of North Carolina. J Vert Paleont 19:351–354
Sues HD, Olsen PE, Carter JG, Scott DM (2003) A new crocodylomorph archosaur from the Upper Triassic of North Carolina. J Vert Paleont 23:329–343
Sues HD, Nesbitt SJ, Berman DS, Henrici AC (2011) A late-surviving basal theropod dinosaur from the latest Triassic of North America. Proc Biol Sci 278(1723):3459–3464
Sulej T (2002) Species discrimination of the Late Triassic temnospondyl amphibian Metoposaurus diagnosticus. Acta Palaeont Polon 47:535–546
Sulej T (2005) A new rauisuchian reptile (Diapsida: Archosauria) from the Late Triassic of Poland. J Vert Paleont 225:78–86
Sulej T (2007) Osteology, variability and evolution of Metoposaurus, a temnospondyl from the Late Triassic of Poland. Palaeont Polon 64:29–139
Sulej T (2010) The skull of an Early Late Triassic aetosaur and the evolution of the stagonolepidid archosaurian reptiles. Zool J Linnean Soc 158:860–881
Sulej T, Majer D (2005) The temnospondyl amphibian Cyclotosaurus from the Upper Triassic of Poland. Palaeontology 48:157–170
Sulej T, Bronowicz R, Tałanda M, Niedźwiedzki G (2011) A new dicynodont-archosaur assemblage from the Late Triassic (Carnian) of Poland. Earth Environ Sci Trans R Soc Edinb 101:261–269
Sulej T, Niedźwiedzki G, Bronowicz R (2012) A new Late Triassic vertebrate fauna from Poland with turtles, aetosaurs, and coelophysoid dinosaurs. J Vert Paleont 32:1033–1041
Sullivan RM, Lucas SG (1999) Eucoelophysis baldwini, a new theropod dinosaur from the Upper Triassic of New Mexico, and the status of the original types of Coelophysis. J Vert Paleont 19:81–90
Sullivan RM, Lucas SG, Randall KA (1995) Late Triassic vertebrate fauna from the Zions View locality (Little Conewago Creek), York County, Pennsylvania. J Vert Paleont 15(3):55A
Świło M, Niedźwiedzki G, Sulej T (2014) Mammal-like tooth from the Upper Triassic of Poland. Acta Palaeont Polon 59:815–820
Szajna MJ, Silvestri SM (1996) A new occurrence of the ichnogenus Brachychirotherium: implications for the Triassic-Jurassic mass extinction event. In: Morales M (ed) The continental Jurassic. Mus N Arizona Bull 60: 275–283
Szulc J, Racki G, Jewuła K, Środoń J (2015) How many Upper Triassic bone-bearing levels in the Upper Silesia (southern Poland)? Ann Soc Geol Polon 85:587–626
Tatarinov LP (2005) A new cynodont (Reptilia, Theriodonta) from the Madygen Formation (Triassic) of Fergana, Kyrgyztan. Paleont J 39:192–198
Thomson TJ, Lovelace DM (2014) Swim track morphotypes and new track localities from the Moenkopi and Red Peak formations (Lower-Middle Triassic) with preliminary interpretations of aquatic behaviors. New Mex Mus Nat Hist Sci Bull 62:103–128
Thulborn T, Turner S (2003) The last dicynodont: an Australian Cretaceous relict. Proc R Soc Lond B 270:985–993
Tixeront M (1971) Lithostratigraphie et minéralisations cuprifères et uranifères stratiformes syngenetiques et familières des formations detritiques Permo-Triasiques du couloir d’Argana (Haut-Atlas occidental, Maroc) et possibilitiés de recherches. Maroc Dir Mines Géol Div Géol Rep Serv Études Gîtes Min 921:1–37
Unwin DM, Alifanov VR, Benton MJ (2000) Enigmatic small reptiles from the Midde-Late Triassic of Kyrgyzstan. In: Benton MJ, Shishkin DM, Kurochkin EN (eds) The age of dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, pp 177–186
Valencio DA, Mendía JE, Vilas JF (1975) Palaeomagnetism and K-Ar ages of Triassic igneous rocks from the Ischigualasto-Ischichuca basin and Puesto Viejo Formation, Argentina. Earth Planet Sci Lett 26:319–330
Voigt S, Buchwitz M, Fischer J, Kogan I, Moisan P, Schneider JW, Spindler F, Brosig A, Preusse M, Scholze F, Linnemann U (2017) Triassic life in an inland lake basin of the warm-temperate biome—the Madygen Lagerstätte (southwest Kyrgyzstan, Central Asia). In: Fraser NC, Sues H-D (eds) Terrestrial conservation Lagerstätten: windows into the evolution of life on land. Dunedin Academic Press, Edinburgh. in press
von Huene F (1940) The tetrapod fauna of the Upper Triassic Maleri beds. Palaeont Indica New Ser 32(1):1–42
Watson DMS (1914) Eunotosaurus africanus Seeley, and the ancestry of the Chelonia. Proc Zool Soc Lond 11:1011–1020
Weems RE, Lucas SG (2015) A revision of the Norian conchostracan zonation in North America and its implications for Late Triassic North American tectonic history. New Mex Mus Nat Hist Sci Bull 67:303–317
Weems RE, Tanner LH, Lucas SG (2016) Synthesis and revision of the lithostratigraphic groups and formations in the upper Permian?-Lower Jurassic Newark Supergroup of eastern North America. Stratigraphy 13:111–153
Westphal F (1970) Phytosaurier-Hautplatten aus der Trias von Madagaskar-ein Beitrag zur Gondwana-Paläogeographie. Neues Jahrb Geol Paläont Monats 1970:632–638
Whiteside DI, Marshall JEA (2008) The age, fauna and palaeoenvironment of the Late Triassic fissure deposits of Tytherington, South Gloucestershire, UK. Geol Mag 145:105–147
Whiteside JH, Grogan DS, Olsen PE, Kent DV (2011) Climatically driven biogeographic provinces of Late Triassic tropical Pangea. Proc Natl Acad Sci U S A 108:8972–8977
Wild R (1978) Die Saurier von Kupferzell-Bauersbach. Württemb Franken Jahrb 62:1–16
Wild R (1980) The fossil deposits of Kupferzell, southwest Germany. Mes Vert Life 1:15–18
Wild R (1989) Aetosaurus (Reptilia: Thecodontia) from the Upper Triassic (Norian) of Cene near Bergamo, Italy, with a revision of the genus. Riv Mus Civ Sci Nat 14:1–24
Wu X, Chatterjee S (1993) Dibothrosuchus elaphros, a crocodylomorph from the Lower Jurassic of China and the phylogeny of the Sphenosuchia. J Vert Paleont 13:58–89
Yadagiri P, Rao BRJ (1987) Contribution to the stratigraphy and vertebrate fauna of the Lower Jurassic Kota Formation, Pranhita-Godavari Valley, India. Palaeobotanist 36:230–244
Yates AM (2003) A definite prosauropod dinosaur from the Lower Eliot Formation (Norian: Upper Triassic) of South Africa. Palaeont Afr 39:63–68
Yates AM, Kitching JW (2003) The earliest known sauropod dinosaur and the first steps toward sauropod locomotion. Proc R Soc Lond B270:1753–1758
Zeigler KE, Heckert AB, Lucas SG eds (2003) Paleontology and geology of the Upper Triassic (Revueltian) Snyder quarry, New Mexico. New Mex Mus Nat Hist Sci Bull 24: 1–132
Acknowledgments
I thank many colleagues for access to specimens in museum collections and/or for assistance in the field. I am particularly grateful to Jaime Powell for access to the Neoatosauroides fossils in Tucuman, Argentina. Andy Heckert, Adrian Hunt and Larry Tanner have collaborated extensively with me in the field studying Late Triassic rocks and fossils, and they have taught me much. Adrian Hunt and Julien Kimmig provided helpful reviews of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG
About this chapter

Cite this chapter
Lucas, S.G. (2018). Late Triassic Terrestrial Tetrapods: Biostratigraphy, Biochronology and Biotic Events. In: Tanner, L. (eds) The Late Triassic World. Topics in Geobiology, vol 46. Springer, Cham. https://doi.org/10.1007/978-3-319-68009-5_10
Download citation
DOI: https://doi.org/10.1007/978-3-319-68009-5_10
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-68008-8
Online ISBN: 978-3-319-68009-5
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)