
Abstract
Spiders can be a particularly important model for the study of cognition. Their close interaction with niche-constructed environmental features, such as webs, cocoons, draglines or retreats, allows for the experimental manipulation of these silken structures, and thus for a controlled study of the cognitive machinery that underlie the use and construction of these structures. There are contrasting theories about cognition, and we explore particularly the opposition between the traditional approach, the one that requires information to be processed solely within the central nervous system (CNS), and the extended cognition approach, which is less restrictive. Here we review the literature on spider cognition with an eye to the experimental data that allows the contrast between these theories of cognition, and conclude that spiders evolved to process information prior to reaching the nervous system: they use their webs to decide whether to attack or not a prey item, and we can experimentally alter their decision by manipulating web properties, such as radii tension. The experimental manipulation of web threads also alters the attentional state of the web building spider so that she predictably ignores important cues for decisions taken during the building process. Together, the experimental evidence shows that spiders extend their cognitive machinery outside the bounds of their CNS, making use of the external silken structures to offload cognitive processing. This insight may help to explain graded changes in brain/body allometry, because smaller animals could rely more on extended cognition so as not to be behaviourally limited by a relatively small brain. Extended cognition could also help explain the emergence of new levels of organisation, particularly the transition from solitary to social life. In general, extended cognition emerges as a natural bridge between two traditionally separate research agendas: the area of cognitive development (learning mechanisms) and that of evolution through natural selection.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Cognition, Plasticity, and Evolution
Although cognition clearly leads to plasticity, the reverse is not necessarily the case. Early ethologists provide not only examples of so-called instinctive behavior, most of them rigid performances elicited only in the presence of specific innate releasing stimuli, but also plenty of cases of alternative instinctive responses to alternative stimuli. Together, these early examples demonstrate that classic ethology framed instinct and plasticity in a continuum from rigid or stereotyped to ever more labile responses (Japyassú and Malange 2014).
The more a behavioral system allows for alternative responses, the more there is room for what we call cognitive performances, that is, performances that require flexible informationFootnote 1 processing abilities, with concomitant behavioral adjustments to environmental changes (Japyassú 2008). These adjustments could be adaptive, or alternatively could be unforeseen responses to new ecological challenges (Penna-Gonçalves et al. 2008), particularly when animals are exposed to environments outside of the normal range for the species (Ghalambor et al. 2007). Finally, these new responses could be stable, either through self-organization and/or learning processes; and as stable new behavioral characteristics, these responses could drive evolution to new local optima (Japyassú 2010) through evolutionary processes such as phenotypic accommodation or assimilation (West-Eberhard 2003).
Models of Cognition
We do not discuss any general definition of cognition; instead we embrace a broad and prevalent definition of cognition as the acquisition, processing, storage, and use of information (Shettleworth 2010). Because we stick to a semantic conception of informationFootnote 2, the acquisition of information is the act of making sense about some aspect of the world. Within this general framework, there is plenty of room for disagreement, and many distinct models of cognition are available in the literature.
One well-known controversy revolves around information processing. Some consider that cognition operates through a general-purpose learning mechanism, one that can solve very different kinds of problems, such as finding a rewarding food patch or finding the appropriate sexual partner. Others conceive cognition as a specialized learning device, one that is evolved to solve a single problem or a single class of problems. In this last conception, the mind comprises a myriad of neural modules, each processing adaptively one single kind of information. The first conception is prevalent among behaviorists, whereas the second is common among evolutionary biologists and psychologists, as well as among behavioral biologists (Laland and Brown 2011; Sanderson 2014).
These conceptions have opposing opinions about the quantity of previous information a cognitive mechanism should have to learn to solve problems. General-purpose learning mechanisms have no a priori information about any particular problem; instead, these mechanisms should extract information from the problem at hand, should find regularities and patterns while experiencing successive instances of that same kind of problem, thus learning the problem structure, and creating memories that help to solve similar tasks in the future. Specialized learning mechanisms, on the other hand, have a priori information that help the system to find the solution to the task. For example, although wandering jumping spiders from the genus Portia do learn how to enter safely into the orb-web of a prey-spider, they optimize the trial and error learning procedure by trying, from scratch, reasonable frequencies and intensities of the aggressive mimicry signals (Jackson and Nelson 2011). In this way, these spiders should have previous information on the most effective range of signal frequencies before starting to learn.
Notwithstanding the differences between these opposing conceptions of cognition, both the general and the specialized conceptions share a common assumption. Both agree that cognition is something that happens within the brain, or the central nervous system (CNS). This is the traditional approach to cognition, whereby central cognitive processing is postulated in abstraction from bodily mechanisms, be it sensory processing or motor control. In contrast, there is a constellation of theories about cognition that downplay the importance of the brain or CNS, arguing instead that cognition extends in various ways to the non-CNS body, or even to the nearby environment. Thus, distinct perspectives oppose themselves to the above conceptions, postulating complementary or alternative theories, such as embodied, situated, embedded, extended, or enacted cognition (Wilson and Foglia 2017).
Embodied cognition theory postulates that the physical structure of the body is part of the solution to ecological problems that animals face, and thus that bodily information helps to reduce the requirements for CNS information processing (Shapiro 2010). As an example, robot bipedal locomotion is much less computationally intensive if the robot physical body has a design that narrows the range of possible movements (Matsushita et al. 2005; Pfeifer et al. 2006). Thus, one can trade information processing for adaptive morphology, and reduce the necessity for central cognition by building upon the natural properties of the materials that constitute a robot, or an animal, as researchers on morphological computation and soft robotics have been keen to demonstrate (Pfeifer et al. 2014). Embedded cognition is one strand of the situated cognition theories. It extends cognition to encompass the natural and social environment. The focus is on the strategies that organisms use to off-load cognitive processing onto the environment. This could be as simple as flagging food-caching sites to facilitate later food retrieval, or as complex as using cloud-computation to process big-data. This specialized use of external devices opens room for the idea of a cognitive performance that is distributed across the physical, social, or cultural environment (Hutchins 1995).
The thesis of extended cognition is the claim that cognitive systems themselves extend beyond the boundary of the individual organism (Clark and Chalmers 1998). The practical difference between embedded and extended approaches is that, in the embedded or situated approaches, you use the environment (for example, a computer) to process part of the information so that your brain and body can solve the problem with less effort; now, in the extended approach, the claim is that, in some cases, the environment not only helps, but instead becomes so intimately connected to the agent that it becomes part of the bodily information processing system (as if the computer was an integral part of your body, like a chip implanted on the brain).
Finally, in the enacted approach, what constitutes cognition is the dynamic coupling between environment and the biological system (Thompson 2007). This approach is similar to, but does not go so far as the dynamic systems approach (Thelen and Smith 1994) which almost eliminates the distinctiveness of cognition, or informational processes, from other biological processes.
This is not the place to go into the details of any of these theories about cognition. Instead, following Japyassú and Laland (2017), we will contrast the centralized and extended approaches to cognition. By centralized, we mean all the traditional cognitive science theories that consider cognition as something that happens within the CNS. This is in contrast to embodied, embedded, enacted, extended, and dynamic system theories, that posit cognitive relevance to the extra-CNS elements that help to solve problems. We shall call these theories collectively the extended cognition approach, because of their emphasis on extending cognition from the encapsulated brain to its external world.
We will discuss findings on spider cognition that allow the distinction between these contrasting approaches, so that we can have an empirical grasp of the theoretically rich debate on cognition. To distinguish the boundaries of the cognitive system, to decide if it is restricted to brain functioning or extends to the body or nearby environment, we will take advantage of the mutual manipulability criterion (MM, Kaplan 2012). In a nutshell, the MM specifies that two components are part of one same system if they reciprocally alter each other. For example, if manipulating something (external to the CNS) results in cognition changes and, in the opposite direction, altering the CNS internal cognitive processes result in changes in this external part of the system, then we shall conclude that this external part is indeed a constituent of the cognitive system and, accordingly, cognition extends.
Spider Extended Cognition
The idea that there is either innate or learned information within the CNS for the performance of a behavior is a trivial one. Phylogenetic or developmental memories are considered relevant for animal adaptive responses. Nevertheless, there is evidence that spiders solve problems building upon information that is not within their brains (Japyassú 2008). The interaction between unusual external features of the environment (such as an experimentally modified web) and the normal spider brain not only results in novel, unforeseen, and adaptive foraging behavior in the very first performance, but this new behavior is also a stable output of the behavioral system (Penna-Gonçalves et al. 2008). Stable behavioral outputs can result from the self-organization (see below) of the system’s components (brain/body/environment) in the very moment of the interaction of the animal with the environment, in this case, the altered web. The take-home lesson here is that the information for adaptive responses is not necessarily available in the brain beforehand: that information can emerge at the exact moment of the performance.
Self-organization is a process well known for producing emergent properties, that is, properties that only appear after the interaction between the components of the system. For example, social behavior in ants always results in the correct, collective choice of the shortest trail to alternative food sources, but no individual ant knows which is the best alternative; the best choice emerges from systematic interactions among the individuals of the colony (Sumpter 2010). The point we are exploring in this section is a kind of downgrading of this same logic, from the social to the individual level. Emergent properties could appear in the interaction between one individual and its surrounding environment, and these properties would be particularly important when the relevant features of this environment occur regularly in his ecological niche.
Webs and silk threads are a regular feature of the ecological niche of spiders. Spiders use silk for a variety of functions, from egg-sac protection, to courtship, defense, territorial marking, or foraging. Silk is an old spider adaptation, one that defines the whole taxonomic group, and thus a niche-constructed environmental feature that is particularly well suited to co-evolve with spider behavior and cognition.
Foraging is one of the main functions of spider webs and silk, and cognition is clearly involved in foraging. For example, web spiders memorize the characteristics of a single captured prey, such as the prey type, size, and location (Ades 1988; Rodríguez and Gamboa 2000; Rodríguez and Gloudeman 2011; Rodríguez et al. 2013). Also, web-builders benefit from specialized, rapid one-trial learning during the formation of search images for avoiding dangerous prey, such as ants (Henaut et al. 2014). They can even learn to change web properties, such as web asymmetry (Nakata 2012) or the size of a region of the web (Heiling and Herberstein 1999), so as to optimize future prey capture on the basis of previous foraging experience. Finally, web-building spiders even show a sense of numerosity connected to foraging. Spiders wrap successively captured prey in one single silk package, and store this package for eating later. Packages imperceptibly removed from the web are actively searched for by the spider, in a clear indication of memory. More to the point, packages with a larger number of prey items (irrespective of the prey size) are more intensively searched for than packages with a smaller number of prey items (Rodríguez et al. 2015). A sense of numerosity requires the ability of abstraction, which is an impressive cognitive feat for tiny spiders.
As shown above, there is clear evidence for the conjunction between cognition and silk thread manipulation. Our point is to evaluate if this cognitive machinery extends to web threads, through the application of the mutual manipulability criterion (MM). The available information in the literature allows the application of the MM in exemplary cases: prey capture and web building. As we see below, these exemplary cases provide positive evidence for extended cognition.
Prey capture
Animals always have to choose between alternative paths of action, and these decisions frequently rely on memories of the consequences of previous and similar decisions. The decision about proceeding to the capture of an ensnared prey item depends on the evaluation of its profitability and of the costs involved. Many spiders ignore small prey items, particularly when they are sated, but hungry spiders hunt these less profitable prey. This decision involves adaptive processing of information by the web threads. Web-building spiders can actively focus attention on a particular web portion. They do that by pulling more strongly on the web threads from the more profitable areas of the trap, a behavior that has been shown to lead to enhanced capture success in these web regions (Nakata 2010). Hungry spiders pull the radii more intensely than sated spiders, and thus respond more promptly to less profitable prey, such as fruit flies (Watanabe 2000). Also, enhanced attention to specific web areas can be artificially induced by experimentally augmenting radii tension, and spiders respond more quickly to stimuli coming from the tensed region of the web (Watanabe 2000, Nakata 2010). These attentional changes can have lasting results, as spiders can learn to focus on particularly profitable web areas, tensioning these areas more strongly as soon as the web has been built anew (Nakata 2013).
If we use the MM criteria (Kaplan 2012), it becomes clear that spider cognition extends to the web threads, because they change, and are changed by, CNS cognition. CNS cognition can alter thread tension (for example, hungry spiders tense web radii), but the reverse also is true; radii tension changes result in attentional changes, making the spider systematically ignore some prey. Radii tension modulates the decision to either attack or ignore a prey item, and the spider uses thread tension to process prey information adaptively.Footnote 3 Spiders are able to tune their webs to become more sensitive to distinct kinds of stimuli. In this sense, web threads cannot be understood as passive transmitters, or even passive filters of vibratory information. Thread properties are adjustable, and can process the same information in distinct ways, thus conveying different outputs to one same input. Spiders change thread properties in a way that is functionally similar to the way they change the properties of actual neural networks (for example, facilitating synapses), so the web must be considered parcel and part of their cognitive system. This intimate connection between the nervous system and web threads is further reinforced if one considers the informational liaisons between web-building algorithms and web structure.
Web building
Not only while sensing prey through the web, but also while building the web, the spider uses the structure and spatial distribution of threads as cognitive devices that reduce the difficulty of the very web-building process. Cognition is ubiquitous in the building process: spiders adjust mesh size to cope with distinct prey (Murakami 1983; Sandoval 1994; Schneider and Vollrath 1998; Heiling and Herberstein 2000), learning from experience with previous webs the structural changes to be accomplished in subsequent buildings (Heiling and Herberstein 1999; Venner et al. 2000). If forced to build its vertical web in a horizontal cage, Argiope argentata will first build a very irregular horizontal and planar structure, but subsequent buildings become progressively similar to regular orbs (Nogueira and Ades 2012), in a compelling example of long-term web-building learning.
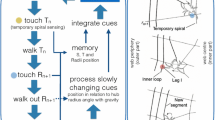
Attention is necessary while building webs, because the spider has to evaluate multiple cues to decide the position of the next threads in the emerging trap. External cues, such as prey-induced vibratory stimuli and prey nutrients (Pasquet et al. 1994; Blamires et al. 2011), wind intensity (Wu et al. 2013), gravity (Witt et al. 1976; Eberhard 1987; Vollrath 1988a, b), and humidity (Baba et al. 2014), all inform distinct aspects of the final web. Spiders also use internal cues to guide web building, such as the amount of silk supply, spider size, weight (Eberhard 1988a), and leg length (Witt et al. 1968; Vollrath 1987). Finally, the spider relies on cues put in place in the building process (position, angle, distances along threads), and on memory of these cues in previous assessments, to decide the next building steps (Eberhard 1972, 1982, 1988b, 2012a, 2012b). As a minor example of the complexities involved in web-building, we will detail the decision about the distance between successive sticky spiral segments on one specific radius of the orb. This decision involves the assessment of many distinct cues, such as reference points (the position of the inner loop of sticky spiral; the position of the outer loop of temporary spiral), the distance from the hub, the angle of the radius with gravity, the distance between radii, the measurement of distances (such as the actual temporary spiral to inner loop distance, Eberhard and Wcislo 2011). The spider also has to compare actual distances with either short-term memories of similar distances in the previous sticky spiral segment attachment, or less recent memories concerning the attachment of the previous sticky spiral loop, on the same radius (see review at Eberhard and Wcislo 2011; Eberhard, in prep.). Sometimes the spider ignores some cues in favor of others, for example when faced with conflict between distinct cues, either in natural or in experimental webs (Eberhard 2011; Eberhard and Hesselberg 2012; see review at Japyassú and Laland 2017). For example, when fixing one sticky spiral segment, the spider senses some cues on the radii (the position of the inner sticky spiral loop, and the position of the outer temporary spiral loop), to calculate the distance between the actual sticky segment and the previous, inner sticky spiral loop, thus producing a regular spacing between successive sticky spiral loops (Fig. 14.1). Nevertheless, this strategy would fail to produce regular spacing in experimental (or natural) webs with a partially destroyed spiral loop. In this case, the distances walked through the previous radius would be much smaller than the distances walked in the next radius (the one with the destroyed spiral segment), introducing a conflict between the actual and the previous cues. Facing this conflict, the spider ignores some cues (inner loop sensing) and favors others (temporary spiral sensing), avoiding the production of an irregular mesh of sticky spiral loops. Thus, due to the complexity of the task, involving the assessment of multiple cues, the experimental manipulation of the actual configuration of threads during the web-building process can actively change the spider’s attention, leading the spider more prone to ignore some cues.

Cycle of actions necessary to build the current segment of the adhesive spiral. Steps and processes within the cycle (a), with the illustration of some of the behaviors involved (b, adapted from Eberhard and Wcislo 2011). The cycle (blue arrows in a) begins and ends with the fixation of the current adhesive spiral segment (blue box). The spider fixes (the current segment) over the current radius (R n, a; spider behavior displayed at b, top figure), and then in the next radius (R n+1, a; spider behavior displayed at b, bottom figure). The spider performs successive actions (large blue arrows, a), while assessing the position of some rapidly changing cues (coloured balls). Slowly and rapidly changing cues are stored, compared to each other (to obtain distances and rates of change) and then integrated (continuous thin blue lines, a) to determine the position of the next adhesive segment fixation (in Rn+1). When confronted with conflicting cues, the spider may ignore some cues (inner loop sensing, dotted blue line, a). These cycles are repeated until the completion of the capture area (Modified from Fig.1 in Japyassú and Laland 2017)
The application of the MM criteria (Kaplan 2012) shows that web structure is part and parcel of the web-building cognitive machinery. Changing the spider’s cognitive state results in changes in the web. For example, well-fed spiders build orbs less frequently (Vollrath and Samu 1997), with smaller capture areas (Mayntz et al. 2009; Baba and Miyashita 2006; but see Vollrath and Samu 1997), and also webs with an added structure, the barrier web (Baba and Miyashita 2006). Also, the injection of drugs or the ingestion of natural substances causes changes in the web geometry (Hesselberg and Vollrath 2004; Albín et al. 2014), and it is thus clear that CNS cognitive machinery is causally connected to web structure. The reverse is also true, as experiments showing that the removal of threads (experimental webs) during the web-building process alters the spider attentional state, reviewed above, clearly demonstrate. As a result, web-building cognition extends to the web itself, as the very structure of the web can change the internal cognitive workings in the spider central nervous system.
Implications for the Evolution of Cognition
If cognition extends to external features of the environment, then the cognitive capacity of any animal should be measured not only by the volume or number of its neurons and/or synapses, but should instead also include the environmental information processing capacity. This could help explain findings that have been difficult to accommodate within the traditional view of cognition as CNS information processing. For example, it has long been shown that there are graded changes in brain–body allometry: larger animals such as mammals or reptiles cannot possibly be as small as ants or spiders, because they would have prohibitively large brains (Eberhard and Wcislo 2011). Extended cognition fits smoothly as a reasonable explanation for these otherwise incomprehensible taxa-specific brain–body allometry rules.
If smaller animals have proportionally smaller brains, they should show a relatively impaired behavioral performance, when compared to larger-brained animals, according to the traditional view of cognition. Nevertheless, this is not what the experimental evidence dictates. Tiny youngsters build as regular and functional webs as adult spiders (Hesselberg 2010; Eberhard and Wcislo 2011), small bees are capable of cognitive feats such as the formation of concepts (Giurfa et al. 2001), and the same is true about spiders that show a sense of numerosity (Rodríguez et al. 2015). From the perspective of the traditional, central cognition approach, such cognitive feats are not expected from such small-brained animals. Nevertheless, from the extended cognition approach, tiny animals are not expected to be behaviorally limited, as long as they are able to export cognitive processing to the environment, as spiders do.
For cognition to extend, the environment around the animal has to be predictable, so that the animal can be confident that some processes will regularly occur outside of its body. In this situation, CNS cognition can evolve so as to complement environmental processing, or even better, it can evolve to control the environment so as to modulate this environmental processing. This is most probably the case when the animal actively changes the environment through ecological engineering, and thus niche construction emerges as a major evolutionary process facilitating brain/environment cognitive coupling, and driving an evolutionary path that could in the end result in the incorporation of the external environmental features to the animal itself, so that the environmental feature becomes almost indissociable from the animal. This is actually the case in many social animals, like ants, termites, bees, and wasps, which are almost defined by the regular structures (nests, mounds, hives, etc.) they create.
The first step in this body/environment coupling process is the self-organization of brain neural networks and environmental features. The environmental effect of the action of one brain network can eventually feed back into the workings of another, previously unrelated brain network, creating new brain–environment feedback loops. These loops could stabilize the newly emerged (brain/environment) system in a new state, that is, could stabilize a new behavioral performance.Footnote 4 If the new state is adaptive, the brain/environment coupling is selected for, and the emergent performance becomes predictable in the long run. The important point here is that the coupling enables a reduction in CNS information processing through environmental processing, and thus small animals can evolve in the direction of complex performance even when anatomically restricted to having a small brain.
This process that begins with the self-organization of a new behavior, and continues through niche-construction to co-adapt the distributed, environmental information processing to CNS information processing, can proceed even further in social animals to the formatting of social interactions. Thus, the evolution of cognition through self-organization and niche construction helps the evolution of adaptive and distributed social information processing, and thus prompts the emergence of new levels of biological organization.
This cognitive route to the emergence of the social level mimics the emergence of the organismic level (with the onset of bodily integration through neural systems) from the simpler perceptual mechanisms of single-celled animals (Pezzulo & Levin 2015; Baluška and Levin 2016). Also, it is the basis for a nascent theoretical integration between traditionally separate research agendas: the area of cognitive development (learning mechanisms) and that of evolution through natural selection (Power et al. 2015; Watson and Szathmáry 2016). Extended cognition may after all be the proximate process instructing the evolution of new levels of biological organization.
Notes
- 1.
Information has at least three broad meanings: the statistical, the semantic, and the physical (Harms 2006). We use the semantic sense to characterize “knowledge of” or “meaning” in both the referential properties of symbols and instructional aspects of knowledge in natural biological systems.
- 2.
The actual meaning of a piece of information depends not only on the referent (the external object), but also on the internal state of the system. In the first case, meaning involves a denotative relation between a sign and its counterpart in the external world (the referent). In the second case, meaning involves a connotative relation between the sign and the internal elements of the system, a relationship that ensures an interpretation, that leads to a procedure or a path of action within the system (Harms 2004, 2006). This second, interpretive side of information requires a characterization of the connectivity between the internal elements of the system, and is thus by definition a relational conception of information. This idea of a system of mutual relations is also relevant to naturalize important properties of any cognitive system, such as agency and normativity (Moreno and Mossio 2015).
- 3.
The radial threads modulate the resonance and the attenuation of prey vibrations, as well as the velocity of their propagation, and thereby promote signal transformation through the web (Landolfa and Barth 1996). Tense threads increase the amplitude of some, and reduce the amplitude of other prey vibration frequencies (Mortimer et al. 2015).
- 4.
Although it is notoriously difficult to detect novelty in a lifelong, complete repertoire of actions (because some performances could be simply rare in place of nonexistent), sometimes novelty is the only possibility, for example when the behaviour is impossible without a particular experimental manipulation. This is the case of the reeling attack tactic, whereby the spider reels a dry thread so that an entangled prey comes close enough to be wrapped. Reeling attack is the default foraging strategy for a whole family of cobweavers, but orbweavers cannot possibly attack through reeling under natural conditions, because their orbweb’s radii are firmly attached to the frame (and thus cannot be reeled). Surprisingly, orbweavers on experimental orbwebs (with a radii artificially cut free from the frame) do promptly reel-attack their prey in the very first trial; this new behaviour is stable, occurring predictably in the experimental orbwebs, and in all the species studied (Penna-Gonçalves et al. 2008). Since orbweavers never attack naturally through reeling, and considering this behaviour is impossible in normal orbwebs, this experimental result requires explanation, because these spiders cannot possibly have an adapted neural network for controlling a reeling attack. The explanation is rather simple: orbweavers do reel threads in natural circumstances, but only when building their webs, and never in a foraging context (prey attack). Thus, the cut-free radius of the experimentally modified orbweb provides the opportunity for the spider to perform a known behavior within a novel, prey-attack context. This is precisely the case of self-organization discussed above. A novelty (predatory reeling in orbweavers) emerges and stabilizes through an environmental (cut-free radius) modification that allows a feedback between two existing neural networks. The cut-free radius allows the co-occurrence of a (natural) web-building behaviour (reeling) with an attack behaviour (prey-wrapping), with the consequent emergence of a new foraging tactic: the reel-attack. This exemplifies how self-organization can produce new and stable behaviors; in this case, the evolutionary appearance of the reel-attack requires only the evolution of a specific environmental feature (a detachable radius), and this is precisely what occurred in the transition from ancestral orbwebs to derived cobwebs.
References
Ades C (1988) Memória e aprendizagem em aranhas. Biotemas 1:2–27
Albin A, Lacava M, Viera C (2014) Effects of gonyleptidine on orb web spider Araneus Lathyrinus (Holmberg 1875). Arachnol 16:154–156
Baba Y, Miyashita T (2006) Does individual internal state affect the presence of a barrier web in Argiope bruennichii (Araneae: Araneidae)? J Ethol 24(1):75–78
Baba YG, Kusahara M, Maezono Y, Miyashita T (2014) Adjustment of web-building initiation to high humidity: a constraint by humidity-dependent thread stickiness in the spider Cyrtarachne. Naturwissenschaften 101(7):587–593
Baluška F, Levin M (2016) On having no head: cognition throughout biological systems. Front Psychol 7:902
Blamires SJ, Chao YC, Liao CP, Tso IM (2011) Multiple prey cues induce foraging flexibility in a trapbuilding predator. Anim Behav 81(5):955–961
Clark A, Chalmers D (1998) The extended mind. Analysis 58:10–23
Eberhard WG (1972) The web of Uloborus diversus (Araneae: Uloboridae). J Zool 166(4):417–465
Eberhard WG (1982) Behavioral characters for the higher classification of orb-weaving spiders. Evolution 36:1067–1095
Eberhard WG (1987) Effects of gravity on temporary spiral construction by Leucauge mariana (Araneae: Araneidae). J Ethol 5(1):29–36
Eberhard WG (1988a) Behavioral flexibility in orb-web construction: effects of supplies in different silk glands and spider size and weight. J Arachnol 295–302
Eberhard WG (1988b) Memory of distances and directions moved as cues during temporary spiral construction in the spider Leucauge mariana (Araneae: Araneidae). J Insect Behav 1(1):51–66
Eberhard WG (2011) Are smaller animals behaviorally limited? Lack of clear constraints in miniature spiders. Anim Behav 81:813–823
Eberhard WG, Wcislo WT (2011) Grade changes in brain-body allometry: morphological and behavioural correlates of brain size in miniature spiders, insects and other invertebrates. Adv Insect Physiol 40:155
Eberhard WG (2012a) Cues guiding placement of the first loop of the sticky spiral in orbs of Micrathena duodecimspinosa (Araneidae) and Leucauge mariana (Tetragnathidae). Arachnology 16(7):224–227
Eberhard WG (2012b) Correlations between Leg positions and spaces between sticky lines in the orbs of Micrathena duodecimspinosa (Araneae: Araneidae). Arachnology 15(7):235–240
Eberhard WG, Hesselberg T (2012) Cues that spiders (Araneae: Araneidae, Tetragnathidae) use to build orbs: lapses in attention to one set of cues because of dissonance with others? Ethology 118(7):610–620
Ghalambor CK, McKay JK, Carroll SP, Reznick DN (2007) Adaptive versus non-adaptive phenotypic plasticity and the potential for contemporary adaptation in new environments. Funct Ecol 21:394–407
Giurfa M, Zhang S, Jenett A, Menzel R, Srinivasan MV (2001) The concepts of ‘sameness’ and ‘difference’ in an insect. Nature 410(6831):930–933
Harms WF (2004) Information and meaning in evolutionary processes. Cambridge University Press, Cambridge
Harms WF (2006) What is information? Three concepts. Biol Theory 1:230–242
Heiling AM, Herberstein ME (1999) The role of experience in web-building spiders (Araneidae). Anim Cogn 2:171–177
Heiling AM, Herberstein ME (2000) Interpretations of orb-web variability: a review of past and current ideas. Ekologia(Bratislava)/Ecology(Bratislava) 19:97–106
Hesselberg T, Vollrath F (2004) The effects of neurotoxins on web-geometry and web-building behaviour in Araneus diadematus Cl. Physiol Behav 82(2):519–529
Hesselberg T (2010) Ontogenetic Changes in Web Design in Two Orb‐Web Spiders. Ethology 116(6):535–545
Hénaut Y, Machkour-M’Rabet S, Lachaud JP (2014) The role of learning in risk-avoidance strategies during spider–ant interactions. Anim Cogn 17:185–195
Hutchins E (1995) Cognition in the wild. MIT Press, Cambridge
Jackson RR, Nelson XJ (2011) Reliance on trial and error signal derivation by Portia Africana, an araneophagic jumping spider from East Africa. J Ethol 29:301–307
Japyassú HF (2008) Cognições mínimas. In: Vianna B (ed) Biologia da libertação. Mazza Edições, Belo Horizonte, pp 97–113
Japyassú HF (2010) Fenótipos amplificáveis em pequenas cognições. Rev Etol 9:63–71
Japyassú HF, Malange J (2014) Plasticity, stereotypy, intra-individual variability and personality: handle with care. Behav Processes 109:40–47
Japyassú, H. F., & Laland, K. N. (2017). Extended spider cognition. Animal Cognition, 1-21
Kaplan DM (2012) How to demarcate the boundaries of cognition. Biol Philos 27:545–570
Laland KN, Brown GR (2011) Sense and nonsense: evolutionary perspectives on human behavior. Oxford University Press, Oxford
Landolfa MA, Barth FG (1996) Vibrations in the orb web of the spider Nephila clavipes: cues for discrimination and orientation. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 179(4):493–508
Matsushita K, Lungarella M, Paul C, Yokoi H (2005) Locomoting with less computation but more morphology. In Robotics and Automation, 2005. ICRA 2005. Proceedings of the 2005 IEEE International Conference on (pp. 2008–2013). IEEE
Mayntz, D., Toft, S., & Vollrath, F. (2009). Nutrient balance affects foraging behaviour of a trap-building predator. Biology letters, rsbl20090431
Moreno A, Mossio M (2015) Biological autonomy: a philosophical and theoretical enquiry, vol 12. Springer, Berlin
Mortimer B, Holland C, Windmill JF, Vollrath F (2015) Unpicking the signal thread of the sector web spider Zygiella x-notata. J R Soc Interface 12(113), 20150633
Murakami Y (1983) Factors determining the prey size of the orb-web spider, Argiope amoena (L. Koch) (Argiopidae). Oecologia 57(1–2):72–77
Nakata K (2010) Attention focusing in a sit-and-wait forager: a spider controls its prey-detection ability in different web sectors by adjusting thread tension. Proc R Soc B 277:29–33
Nakata K (2012) Plasticity in an extended phenotype and reversed up–down asymmetry of spider orb-webs. Anim Behav 83:821–826
Nakata K (2013) Spatial learning affects thread tension control in orb-web spiders. Biol Lett 9(4):20130052
Nogueira SDC, Ades C (2012) Evidence of learning in the web construction of the spider Argiope argentata (Araneae: Araneidae). Rev Etol 11(1):23–36
Pasquet A, Ridwan A, Leborgne R (1994) Presence of potential prey affects web-building in an orb-weaving spider Zygiella x-notata. Anim Behav 47(2):477–480
Penna-Gonçalves V, Garcia CRM, Japyassú HF (2008) Homology in a context dependent predatory behavior in spiders (Araneae). J Arachnol 36:352–359
Pezzulo G, Levin M (2015) Remembering the body: applications of computational neuroscience to the top-down control of regeneration of limbs and other complex organs. Integr Biol 7:1487–1517
Pfeifer R, Iida F, Gómez G (2006) Morphological computation for adaptive behavior and cognition. In: International congress series, vol 1291. Elsevier, Kitakyushu (Japan), pp 22–29
Pfeifer R, Iida F, Lungarella M (2014) Cognition from the bottom up: on biological inspiration, body morphology, and soft materials. Trends Cogn Sci 18:404–413
Power DA, Watson RA, Szathmáry E, Mills R, Powers ST, Doncaster CP, Czapp B (2015) What can ecosystems learn? Expanding evolutionary ecology with learning theory. Biol Direct 10:69
Rodríguez RL, Gamboa E (2000) Memory of captured prey in three web spiders (Araneae, Araneidae, Linyphiidae, Tetragnathidae). Anim Cogn 3:91–97
Rodríguez RL, Gloudeman MD (2011) Estimating the repeatability of memories of captured prey formed by Frontinella Communis spiders (Araneae: Linyphiidae). Anim Cogn 14:675–682
Rodríguez RL, Kolodziej RC, Höbel G (2013) Memory of prey larders in golden orb-web spiders, Nephila clavipes (Araneae: Nephilidae). Behaviour 150:1345–1356
Rodríguez RL, Briceño RD, Briceño-Aguilar E, Höbel G (2015) Nephila Clavipes spiders (Araneae: Nephilidae) keep track of captured prey counts: testing for a sense of numerosity in an orb-weaver. Anim Cogn 18:307–314
Sanderson SK (2014) Human nature and the evolution of society. Westview Press, Boulder
Sandoval, C. P. (1994). Plasticity in web design in the spider Parawixia bistriata: a response to variable prey type. Functional Ecology, 701-707
Schneider JM, Vollrath F (1998) The effect of prey type on the geometry of the capture web of Araneus diadematus. Naturwissenschaften 85(8):391–394
Shapiro LA (2010) Embodied cognition. Routledge, London
Shettleworth SJ (2010) Cognition, evolution, and behavior. Oxford University Press
Sumpter DJ (2010) Collective animal behavior. Princeton University Press, Princeton
Thelen E, Smith LB (1994) A dynamic systems approach to the development of cognition and action. MIT Press, Cambridge
Thompson E (2007) Mind in life: biology, phenomenology, and the sciences of mind. Harvard University Press, Cambridge
Venner S, Pasquet A, Leborgne R (2000) Web-building behaviour in the orb-weaving spider Zygiella xnotata: influence of experience. Anim Behav 59(3):603–611
Vollrath F (1987) Altered geometry of webs in spiders with regenerated leg. Nature 328:247–248
Vollrath F (1988a) Untangling the spider’s web. Trends Ecol Evol 3(12):331–335
Vollrath F (1988b) Spiral orientation of Araneus diadematus orb-webs built during vertical rotation. J Comp Physiol A 162(3):413–419
Vollrath F, Samu F (1997) The effect of starvation on web geometry in an orb-weaving spider. Bull Br Arachnol Soc 10:295–297
Watanabe T (2000) Web tuning of an orb-web spider, Octonoba Sybotides, regulates prey-catching behavior. Proc R Soc B 267:565–569
Watson RA, Szathmáry E (2016) How can evolution learn? Trends Ecol Evol 31:147–157
West-Eberhard MJ (2003) Developmental plasticity and evolution. Oxford University Press, New York
Wilson RA, Foglia L (2017) Embodied Cognition, in The Stanford Encyclopedia of Philosophy (Spring 2017 Edition), Edward N. Zalta (ed.), forthcoming URL = https://plato.stanford.edu/archives/spr2017/entries/embodied-cognition/
Witt PN, Reed C, Peakall DB (1968) A spider’s web: problems in regulatory biology. Springer, New York
Witt, P. N., Scarboro, M. B., Daniels, R., Peakall, D. B., & Gause, R. L. (1976). Spider web-building in outer space: evaluation of records from the Skylab spider experiment. Journal of Arachnology, 115-124
Wu CC, Blamires SJ, Wu CL, Tso IM (2013) Wind induces variations in spider web geometry and sticky spiral droplet volume. J Exp Biol 216(17):3342–3349
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter

Cite this chapter
Japyassú, H.F. (2017). Plasticity and Cognition in Spiders. In: Viera, C., Gonzaga, M. (eds) Behaviour and Ecology of Spiders. Springer, Cham. https://doi.org/10.1007/978-3-319-65717-2_14
Download citation
DOI: https://doi.org/10.1007/978-3-319-65717-2_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-65716-5
Online ISBN: 978-3-319-65717-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)