
Abstract
Genotypes may differ considerably in the magnitude of performance in different environments. When genotypes perform differently in variable environments, genotype x environment interaction (G x E or GEI) has occurred. GEI poses a big challenge to maize breeders when developing varieties for release to farmers in a large geographical area, which is the case in West Africa (WA). Compulsorily, multi-locational environment trials (METs), involving the evaluation of a set of genotypes or varieties in many different environments within agroclimatic zones of WA, are used to identify stable, high-yielding genotypes for registration, release, and commercialization. The test locations used for the different types of Regional Uniform Variety Trials (RUVTs) since the time of WECAMAN (RUVT-early and RUVT extra-early) and now the DTMA/STMA trials for drought-tolerant varieties are not very representative and precisely discriminating. Therefore, there is a need to sample more testing sites in WA, using modern cultivars with high variation in mean performance; varieties developed during the last decade to identify sites which possess high representativeness, discriminating ability, and repeatability. In addition, because WA is large with several distinct agroclimatic zones that cut across the countries of the sub-region, there is a need to identify mega-environments for cultivar evaluation, release, and commercialization across WA. Such sites would be more appropriate for use in evaluating and selecting superior drought-tolerant genotypes as well as for serving as core testing sites within each agroclimatic zone, thereby minimizing human labor and research expenses. Several statistical tools are available for the analysis of data from METs. Among the most powerful tools are the additive main effects and multiplicative interaction (AMMI) and the GGE biplot. Many sets of data obtained during the last three decades were subjected to the statistical tools and the procedures used, results obtained, interpretation of the results, and possible application of the outcome to boost maize production in WA are presented in this Chapter.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


1 Introduction
Failure of genotypes to perform consistently in variable environments is termed genotype x environment interaction (G x E or GEI). GEI is advantageous when the breeder is developing varieties for specific ecological niches—narrow adaptation—but it is a disadvantage when developing varieties for broad adaptation. In this case, the genetic worth of the individual genotypes cannot be predicted accurately from their phenotypic performance. Breeders have used several statistical approaches to handle GEI in crops, including:
-
Obtaining optimum combinations of number of locations, years, and replications for yield trials to maximize heritability and progress from selection
-
Identifying the representative or “ideal” evaluation sites in which the performance of the test genotypes correlates highly with their performance in the range of environments for which they are being developed
-
Stratification of the evaluation environments and/or the genotypes being evaluated
-
Using regression analysis to characterize the environmental response of individual genotypes
The advent of the personal computer, along with the development of many user-friendly software packages, has greatly facilitated research in this area, and several sophisticated statistical programs are now available for handling GEI data with relative ease.
In crop species, economic traits such as grain yield are inherited quantitatively and are, therefore, greatly subject to environmental variation. Breeders carry out genotype evaluation in multiple environments in what is termed multi-environment trials (METs). Data from METs are subjected to the ANOVA (Table 17.1), and significant GEI is further analyzed to decompose the components. The ANOVA model for METs is
In this model:
Y ijlk = the observation on the ith genotype in the jth year, the lth location, and the kth replication, μ = the overall mean of the trait in the experiment, Y j = the effect of the jth year, L l = the effect of the jth location, YL jl = the effect of the interaction of the jth year with the lth location, Rk (jl) = the effect of the kth replication within the lth location in the jth year, G i = the effect of the ith genotype, GY ij = the effect of the interaction of the ith genotype with the jth year, GL il = the effect of the interaction of the ith genotype with the lth location, GYL ijl = the effect of the ith genotype with the ith year and the lth location, and ε ijlk = the random error or residual.
2 Multilocational Testing and Genotype by Environment Interactions in West and Central Africa
The performances of breeding materials or crop varieties differ from one environment to another. While the effect of environment may, in general, be additive in some instances, in others it may not be. Additive environmental effect means that the relative ranking of varieties is maintained. In effect, all or most varieties improve or decrease in their performance by a similar factor from one environment to another. However, varieties may differ considerably in the magnitude of performance they show from a good to a better environment. When the latter is present for a set of genotypes or varieties, genotype–environment interaction is implied. Multilocational trials or multi-environment trials (METs), involving the evaluation of a set of genotypes or varieties in several or many environments that are dissimilar, are required to determine the magnitude of genotype–environment interactions and, by implication, stability of performance of the genotypes or varieties. Multilocational testing can provide information on genotypes with high average performance and in effect stability over a range of different environments. It is also possible, using the results from multilocational trials, to identify genotypes that are best adapted to particular environments. Families that show superiority in such environments are recombined in successive cycles of improvement. Analysis of variance of data obtained from multilocational trials for a set of genotypes can also yield estimates of heritability of important traits considered.
Several methods are available for the analysis and interpretation of multilocational trial (MET) data (Yates and Cochran 1938; Williams 1952; Finlay and Wilkinson 1963; Eberhart and Russell 1966; Gollob 1968; Mandel 1969, 1971; Zobel et al. 1988; Gauch 1988; Gauch and Zobel 1997; Cornelius et al. 1993; Crossa and Cornelius 1997; Yan et al. 2000, 2007). Of these statistical tools, the two most powerful for the analysis of MET data are the additive main effects and multiplicative interaction (AMMI) model proposed by Zobel et al. (1988), Gauch (1988), and Gauch and Zobel (1997) and GGE biplot methodology proposed by Yan et al. (2000). Several reviews have compared and contrasted AMMI and GGE biplot with respect to their suitability for GEI analysis (Gauch 2006; Yan and Tinker 2006; Yan et al. 2007; Gauch et al. 2008; Yang et al. 2009). A major disadvantage of the AMMI model is that it is insensitive to the most important part of the crossover GE. Furthermore, since there is no clear biological separation between the two terms G and GE, the AMMI model does not offer any advantage to the breeder for genotypic and site evaluation when analyzing MET data (Yan et al. 2007). On the other hand, the GGE biplot is a powerful statistical model that takes care of some of the disadvantages of AMMI. The method is an effective statistical tool for identifying the most outstanding cultivar in a given environment and the most suitable environment for each cultivar, the comparison of any pair of cultivars in individual environments, the best cultivars for each environment and mega-environment differentiation, the average yield and stability of the genotypes, and the discriminating ability and representativeness of the environments (Yan et al. 2007; Yan and Kang 2003; Yan and Tinker 2006). The decision as to whether location groups could be considered as mega-environments is based on the consistency of location groupings and of the winning genotypes in the individual location groups across years (Yan et al. 2000, 2007, 2010). There has been a limited use of the GGE biplot method for analysis of METs on maize yield data in WCA by Badu-Apraku et al. (2008a, 2009, 2010). However, there is, presently, an increasing use of the GGE biplot for MET data analysis by researchers of the sub-region (Badu-Apraku et al. 2011a, b, c).
3 Repeatability of Traits and Locations in the West African Biotic and Abiotic Stress Screening Sites of IITA
Repeatability is computed as the proportion of the genetic variance over the total phenotypic variance (Fehr 1987). It represents the upper limit for broad-sense heritabilities. Repeatability and broad-sense heritability mean the same thing if the ratio of genotypic to phenotypic variance is considered. However, the term heritability can be misleading, as it can mean that the trait is inherited from one generation to another. However, sometimes the expression is used for one generation with no genetically constant selection units (hybrids, crosses) which are evaluated in different environments with varying repetitions per environment. Under such circumstances, the word repeatability is more appropriate. In this case, reference is made to how repeatable a trait is when assessed in two environments compared to four environments. The repeatability of a trait increases as more replications are used across environments or in the same environment across years.
Repeatability in a series of randomized complete block designs may be represented as follows:
where l is the number of locations, y is the number of years, and r is the number of replications.
Heritability, which is the ratio of practical importance, may be much less than the repeatability, but it cannot be greater. The repeatability differs very much according to the nature of the character and also according to the genetic properties of the population and the environmental conditions under which the individuals are kept. Falconer (1981) indicated that two assumptions are implicit in the idea of repeatability. The first is that the variances of the different measurements are equal and have their components in the same proportions. The second is that the different measurements reflect what is genetically the same character. Unless these assumptions are valid, repeatability becomes a vague concept.
The regional drought-tolerant early maturing variety trials of the Drought-Tolerant Maize for Africa (DTMA) Project serve as the vehicle for testing, identifying, and exchanging drought-tolerant extra-early (80–85 days) and early (90–95 days) varieties and hybrids of maize with broad adaptation to a particular agroecological zone among the four partner countries, Mali, Nigeria, Ghana, and Benin in West Africa (WA). Promising varieties identified based on trial results are used in farmer participatory on-farm trials and demonstrations which serve as important vehicles to showcase the effectiveness of new technology to farmers and thus are instrumental in the identification, release, and commercialization of drought-tolerant maize varieties and hybrids in the participating countries. The trials also give the NARS partners the opportunity to identify promising cultivars for the introgression of favorable alleles into the breeding populations of national maize programs to diversify and broaden the genetic base.
Results of multilocational trials in WA have established the existence of GEIs (Fakorede and Adeyemo 1986; Badu-Apraku et al. 1995, 2003, 2007, 2008a). This implies the need for extensive testing of cultivars in multiple environments over years before taking decisions on cultivar recommendations. However, due to the scarce resources of the national maize research programs of WA, there is a need to conduct cultivar evaluation in a limited number of environments. As pointed out by Yan et al. (2007), it is important to reexamine target environments for their uniqueness as some environments may never provide unique information, because they are always similar to some other environments in separating and ranking genotypes. This facilitates the identification of core testing sites where evaluation of cultivars can be done without losing valuable information about genotypes. Furthermore, stratification of maize evaluation environments can help increase heritability of measured traits, accelerate the rate of gain from selection, strengthen the potential competitiveness for seed production, and maximize grain yields for farmers (Gauch and Zobel 1997). It is therefore very important to develop a better understanding of the target agroecosystems used for the evaluation of drought-tolerant cultivars in WA and to determine if it could be subdivided into different mega-environments to facilitate a more meaningful cultivar evaluation and recommendation. It is anticipated that locations selected for METs should constitute a sample of environments that adequately represents the range of environmental conditions of the target geographical region. Furthermore, a protocol on seeds was ratified by the heads of states of ECOWAS member countries in 2009, and the West African Catalogue of Plant Species and Varieties (COAFEV) is presently available in the sub-region (FAO 2008). The seed catalogue contains the list of varieties whose seeds can be produced and commercialized within the territories of the 17 member countries of ECOWAS and consists of the varieties registered in the national catalogues of the member states. The catalogue offers a unique opportunity for the deployment of good-quality seeds of improved maize varieties and hybrids across the borders of the ECOWAS countries for production and marketing. As a result of these new developments and the implications of global warming, desertification, and recurrent drought in the sub-region, there is a need for reexamination of the current mega-environments in WA and the identification of core testing sites in each of the mega-environments used for the evaluation of the three different regional trials in WA. Several studies have been conducted to examine the effects of genotype and GEI in early and extra-early cultivar evaluation with particular emphasis on identifying core testing sites in the mega-environments of the lowlands of WCA for the early- and extra-early maturity groups. In one study, Badu-Apraku et al. (2011c) analyzed grain-yield data of the Regional Uniform Variety Trials-early (RUVT-early) containing 18 early cultivars evaluated for 3 years in 15 sites representing the dry savanna, moist savanna, and forest–savanna transition zones of WCA (Table 17.2). One of the objectives of the study was to classify the sites of the experiment into mega-environments based on the method proposed by Yan et al. (2007). According to this classification, test locations may be grouped into three types: (1) locations with low genotype discrimination that should not be selected as test locations; (2) locations with high genotype discrimination, representative of the mega-environment as well as close to the ideal mega-environment, which should, therefore, be chosen for superior genotype selection, when few test locations can be managed due to budget constraints; and (3) locations with high genotype discrimination that do not represent the mega-environment, which could be used for unstable genotype evaluation.
The discriminating power of an environment refers to the ability of the environment to identify an ideal genotype, while the representativeness refers to the ability of a test location to typify the mega-environment. Using the discriminating power versus representativeness view of GGE biplot analysis of the results of the test locations, the test environments in this study, four mega-environments, were identified as follows:
-
Group 1 – Katibougou, Sotouboua, Ejura, and Bagou
-
Group 2 – Manga (MAN), Nyankpala (NYP), Bagauda (BG), Yendi (YD), Angaredebou (ANG), Mokwa (MK), Katibougou (KX), and Zaria (ZA)
-
Group 3 – Ativeme and Ikenne (IKN)
-
Group 4 – Ina
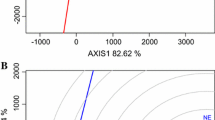
There was high correlation between the test locations Ejura, Sotouboua, Bagou, and Katibougou (Fig. 17.1) in their ranking of the genotypes, an indication that a promising early-maturing cultivar selected in any one of these locations will also be suitable for production in the other locations within the same mega-environments in the same or different countries. Similarly, MAN, NYP, BG, YD, ANG, MOK, KX, and ZA showed high correlation in their ranking of the genotypes in the second group indicating that a promising cultivar identified in one location will be most likely adapted to the other locations in this group. Selection of a cultivar out of these two groups of locations will most likely result in cultivars adapted to IKN and other locations within the same mega-environment. Ina stands alone in mega-environment 4 in its ranking of the genotypes and was unique in the ranking of the genotypes. Kita was identified as the ideal location, while Zaria was close to the ideal location.

The “discriminating power and representativeness” view of GGE biplot based on genotype x environment yield data of 18 early-maturing maize cultivars evaluated in 15 locations across WA between 2006 and 2008. The data were not transformed (“Transform=0”), were not standardized (“Scaling=0”), and were environment-centered (“Centering=2”). The biplot was based on genotype-focused singular value partitioning (“SVP = 2”) and is therefore appropriate for visualizing the relationships among environments. Principal component (PC) 1 and PC 2 explained 51.1% of yield variation
The four mega-environments identified in the study were different from the maize agroecological zones identified by earlier researchers (Fajemisin et al. 1985; Menkir et al. 2003; Setimela et al. 2007). This result was not surprising since the study by Menkir (2003) was based on climatic data obtained from the geographic information system (GIS) for a large number of locations in SSA while those by Fajemisin et al. (1985) involved intermediate-to-late maturing cultivars. Besides, the earlier studies employed methods different from those used by Badu-Apraku et al. (2010, 2011a). Moreover, fewer locations were sampled with no representative location from the mid-altitude agroecology. These reasons might have accounted for the differences in the results of the studies. Validation of the discriminating power of the mega-environments was done using repeatability as the indicator. The repeatability values computed for grain yield and nine other traits, using the data of Badu-Apraku et al. (2011, unpublished data), are summarized in Table 17.3. The output was subjected to factor analysis for the purpose of grouping the 15 locations into factors, which were considered as mega-environments. Repeatability for individual traits varied widely among locations. For example, repeatability for grain yield ranged from −0.91 at Ina to 0.64 at IKN. Across sites, most of the traits had low repeatability values, mostly below 0.4 with only days to anthesis and silk having values above 0.4 (Table 17.3). Similarly, across traits, repeatability values were low for most sites with only Ikenne and Bagauda having values of about 0.4, while other sites had values much lower. As noted in earlier reports on RUVTs (Fakorede et al. 2007), grain yield had a negative relationship with coefficient of variation (CV) also in this study, but grain yield and CV did not influence repeatability.
Five factors (mega-environments) were identified, and together they accounted for 87% of the variation among the locations for repeatability (Table 17.4). Although the proportions of the variation attributable to the factors were not too far apart (about 15–22%), the number of locations per factor varied from two to five. The five mega-environments identified for evaluating early-maturing maize germplasm in this study are:
-
1.
Kati, Angaradebou, Mokwa, Ejura, and Nyankpala with factor loadings of 0.62–0.82. This group accounted for about 22% of the variability among sites for the repeatability values.
-
2.
Manga and Kita with about equal factor loadings of approximately 0.94 each, accounting for about 19% of the variability among sites.
-
3.
Sotouboua, Bagou, and Ativeme with loadings of 0.94, 0.84, and −0.62, respectively. This group accounted for about 16% of the variability among sites. Ativeme had a negative relationship with this group.
-
4.
Bagauda and Ikenne with loadings of about 0.88 and 0.85, respectively, explaining about 15% of the variation among sites.
-
5.
Yendi, Zaria, and Ina with loadings of 0.69–0.91, also accounted for 15% of the variation among sites.
The mega-environments identified in this study were not identical with those from earlier studies, but it was probably the most reliable because it took into consideration several traits of the maize plant. Although both GGE biplot analysis employed by Badu-Apraku et al. (2011c) and factor analysis used the principal component analysis approach, factor analysis has the added advantage that the factor loadings may be subjected to rotation to maximize the correlation among locations loaded on the same factor while minimizing the relationship between factors. Essentially, the mega-environments delineated by the factor analysis are orthogonal to each other (Fakorede 1979).
The five factors identified in this latter study represent five mega-environments. It is anticipated that cultivars with superior performance in any of the locations with positive loadings on a particular factor will perform well in other locations loaded on the factor. Cultivars that perform well in locations with positive loadings on the factor are expected to perform poorly in the location with negative loadings.
The information on the repeatability of traits of the 18 early-maturing cultivars presented in Table 17.5 are useful for identification of locations with high repeatability when averaged across traits, traits with high repeatability when averaged across locations, and location x trait interaction for repeatability. Based on this interpretation, only days to anthesis and silk were moderately repeatable across locations in the study; repeatability values for all other traits were low. Similarly, apart from Ikenne, Bagauda, Ikenne, Kita, Mokwa, and Katibouguo with repeatability estimates of 0.5 or more for grain yield, the aggregate phenotypes of the maize cultivars in the study were poorly repeatable in terms of grain yield at the different testing sites. However, repeatability for some other traits was high in some locations and low or even zero in some others. It is particularly striking that some of the locations with high grain yield and relatively low CV such as Zaria had low repeatability for yield and the aggregate phenotype.
Badu-Apraku et al. (2011b) identified Zaria, Ilorin, Ikenne, Ejura, Kita, Babile, Ina, and Angaredebou in WA as the core testing sites of the three mega-environments for testing the Regional Uniform Variety Trials-Extra-early. In another study, involving the testing sites for the Regional Early Trials, Badu-Apraku et al. (2011a) classified the test environments into four mega-environments. Four test locations were highly correlated in their ranking of the genotypes in group 1, suggesting that a promising early-maturing cultivar selected in one of these locations in one country will also be suitable for production in the other locations within the same mega-environments in different countries (Badu-Apraku et al. 2011a). Similarly, eight test locations were highly correlated in their ranking of the genotypes in group 2. The implication of this is that a promising cultivar identified in one of these locations will likely be adapted to the other locations. The identification of the core testing sites is expected to facilitate the selection of high-yielding and stable cultivars in the four different regional trials of WA [Regional Uniform Variety Trial (RUVT)-early, RUVT-extra-early, drought-tolerant (DT) regional early, and the DT regional extra-early variety trials] and seed production and marketing across the countries of WA.
The selection of suitable breeding and testing sites is crucial to the success of a maize breeding program. A test location must be discriminating so that genetic differences among genotypes can be easily detected. It is important therefore that the test locations are representative of the target environments so that selected genotypes have the desired adaptation and are representative of the target environment as well as repeatable so that genotypes selected from year to year will have superior performance. According to Yan et al. (2007), only test locations with high discriminating ability are useful, and only those that are also representative can be used in selecting superior genotypes. The repeatability of genotype ranking across years within test locations is also an essential aspect in test location evaluation. The GEI of the testing sites of the RUVT-early and extra-early varieties in WCA has been studied, and the test locations were characterized and stratified into mega-environments and core testing sites to facilitate efficient and less costly testing of varieties (Badu-Apraku et al. 2011a, b). On the contrary, the testing locations of the regional drought-tolerant trials confined to the drought-prone locations in the four partner countries of the Drought Tolerant Maize for Africa (DTMA) Project , Nigeria, Ghana, Benin, and Mali (Table 17.3), have not been extensively studied. Therefore, there was a need for information on the representativeness, discriminating ability, and repeatability of the testing sites of the DT Regional Variety Trials in WA to facilitate the understanding of the responses of drought-tolerant maize genotypes in target drought environments. This was important in designing an efficient and economic selection strategy for the IITA Maize Breeding Program. Badu-Apraku et al. (2013) evaluated 12 early-maturing maize cultivars for 3 years at 16 locations in WA to determine the representativeness, the discriminating ability, and the repeatability of the test locations used for the evaluation of the DT Regional Early Variety Trials and to identify core testing sites to facilitate testing, seed production, and commercialization of drought-tolerant cultivars in WA. The GGE biplot analysis showed that Zaria (Nigeria), Nyankpala (Ghana), and Ejura (Ghana) possessed the highest discriminating ability. Two mega-environments were identified. Bagou, Nyankpala, Bagauda, Ikenne, and Mokwa represented the first mega-environment (ME1); Ejura, Ina, and Sotuba constituted the second (ME2). The ME1 would be more useful for evaluating early maize genotypes for tolerance to drought than ME2 because locations in ME1 were more strongly correlated to Ikenne (managed drought stress site). Among the testing sites , Bagou and Mokwa were found to be closely related to Ikenne in their ranking of the cultivars for drought tolerance; Zaria was the exact opposite, indicating that this was the least suitable location for evaluating genotypes for drought tolerance. Nyankpala and Ikenne were identified as the core testing site for ME1 and Ejura for ME2. TZE Comp 3 C2F2 was identified as the highest-yielding cultivar for ME1 and Syn DTE STR-Y for ME2, indicating that they could be used as check cultivars. Ikenne, Nyankpala, and Ejura had moderately high repeatability. They were closer to the average environment axis of each mega-environment suggesting that they will be useful for culling unstable genotypes during multilocational testing. Other sites were less representative and not repeatable and will not be useful for evaluating early maize cultivars for drought tolerance.
Using the GGE biplot, Akaogu et al. (2012) studied the interrelationship among the test environments for evaluating extra-early hybrids in Nigeria by the IITA Maize Program. In the biplot view presented in Fig. 17.2, the straight line from the origin to the coordinates where an environment falls is denoted as the research environment vector, while the straight line with a single arrow which passes the origin and the average environment represents the average environment axis (AEA). The length of the vector describes its discriminating power, while the angle between an environment and AEA measures its representativeness. According to Yan et al. (2010), the shorter environmental vectors indicate that the specific environments were not strongly correlated with environments with longer vectors and that they were probably not strongly correlated with one another either. Thus, MO11-N and MO11-I had relatively long vectors and were considered as more powerful in discriminating among the hybrids, while IK11-N and IK10-N environments had small angles with AEA and were considered as the most representative of the test environments. The high repeatability of IKDT is not surprising since the induced drought stress treatment was effectively managed so that the irrigation system provided the same amount of water to all plots. Furthermore, the Ikenne site has deep uniform soils with high water-holding capacity. The high repeatability of IKDT has confirmed the effectiveness of the screening methodology adopted by IITA’s Maize Improvement Program for selecting drought-tolerant genotypes. In contrast IKDT had high representativeness, discriminating ability, and repeatability, suggesting that it is the ideal test location. Based on the discriminativeness and representativeness of the research environments used in the present study, it was concluded that Mokwa has the ability to discriminate well among the extra-early maturing hybrids.

The discriminating power and representativeness view of GGE biplot based on genotype x environment yield of 21 extra-early hybrids evaluated at three locations between 2010 and 2011
According to Yan et al. (2007), representativeness of a test site is a key factor for determining how it should be used in cultivar evaluation , assuming adequate discriminating ability. On the other hand, the usefulness of repeatability in MET data analysis lies in the fact that it is an essential step for assessing the representativeness of the test locations. Not until repeatability analysis based on multiyear and multilocational data is performed, a test location cannot be declared as of high, low, null, or negative representation of a mega-environment (Yan et al. 2011). For a test location to be described as highly representative, it must be repeatable across years in ranking genotypes. Based on repeatability analysis, a test location may be classified into one of four categories (Yan et al. 2011). The type I environments are highly representative test locations, which are also highly repeatable by definition. This type of test locations is considered ideal for use as core test locations. Genotypic differences observed at such locations are both repeatable across years and representative of the mega-environment. It is crucial for a breeding program to have a core test location of this type, particularly for early generation selection when it is not feasible because of availability of limited amount of seed to conduct multilocational tests. In our studies, none of the test locations could be classified as a core test location because of their relatively low repeatability and representativeness. Type II environments are low or moderately representative test locations, which may be highly repeatable or less repeatable. Type III environments are test locations that have zero representativeness but are highly repeatable. Such locations may be used to cull unstable genotypes, and, when employed, it is important that the selection intensity is low, to prevent useful genotypes from being mistakenly discarded. Type IV consists of test locations with negative representativeness. Such locations must not be used as test locations for the mega-environment of interest as the selection would be counterproductive. In the present study, Mokwa, Samaru, and Ejura were classified into type II for each of ME1, ME2, and ME3, respectively, owing to their relatively moderate repeatability and proximity to the average environment axis. These locations are expected to be especially useful in the multilocational test stage for selecting against unstable genotypes. Other locations used in this study are less representative and not repeatable, suggesting that the locations are not very useful for evaluating early-maturing maize genotypes for drought tolerance. Furthermore, the identification of Ejura and Ikenne as the most closely related test locations suggests that the ranking of the genotypes at Ikenne under induced moisture stress was quite similar to the ranking at Ejura, under natural drought. In addition, the result also implied that the mega-environment Ejura (ME3) could be considered the best location for testing early maize genotypes for drought tolerance under naturally occurring drought. According to Yan et al. (2011), the repeatability for a given location may vary with the set of genotypes involved. If highly unadapted germplasm or landraces or cultivars released long ago which may perform poorly every year as well as everywhere are included, the genotype main effect may be artificially inflated, leading to a higher estimation of repeatability. On the other hand, if the variation in mean performance among tested genotypes is very small, a low repeatability will result. In the present study, the sum of squares of the genotypic main effects for all measured traits accounted for only a small proportion of the total sum of squares thus suggesting that there was little variation in mean performance among the genotypes and thus accounting for the low repeatability obtained.
In conclusion, the results of the present study showed that the test locations used for the evaluation of the early-maturing maize genotypes for drought tolerance in the DTMA regional drought trials are not very representative and discriminating. Also, no core testing sites could be identified among the locations used in this study. Therefore, there is a need to sample more testing sites in WA using the modern cultivars with high variation in mean performance developed during the last decade to identify sites such as Ejura which possess high representativeness, discriminating ability, and repeatability. Such sites would be more appropriate for use in evaluating and selecting superior drought-tolerant genotypes as well as for serving as core testing sites within the identified mega-environments for cultivar evaluation, release, and commercialization across WA.
The development and commercialization of early and extra-early maize varieties and hybrids that are very responsive to added inputs have extended the boundaries of suitability of maize to the drier areas of the northern Guinea savanna and Sudan savanna of WCA. Despite the immense potential of maize in the savannas of WCA, production is greatly constrained by Striga hermonthica parasitism, recurrent drought, and low soil fertility, especially low levels of soil nitrogen (N). These constraints are more pronounced in the drier savannas that are characterized by reduced annual rainfall. To promote rapid adoption and commercialization of maize in the moist and dry savannas of WCA, there is a need for extra-early and early cultivars with combined resistance or tolerance to Striga infestation, drought, and low soil N.
Yield losses due to Striga hermonthica may range from 10% to 100% depending on the variety and the environmental conditions (Kroschel 1999). Striga infestation is extremely difficult to control and constitutes a major threat to the rapid spread of maize into the WCA savanna. Available Striga control measures include host plant resistance and cultural, chemical, and manual methods (Odhiambo and Ransom 1994; Kim et al. 1998). However, the use of host plant resistance or tolerance is considered the most economical, sustainable, and environmentally friendly for resource-poor farmers. Therefore, a breeding program for Striga resistance was initiated in Côte d’Ivoire in 1994 by the IITA to develop maize populations, cultivars, and inbred lines with combined earliness or extra-earliness and resistance/tolerance to S. hermonthica, drought, and low soil N. High-yielding early and extra-early drought and Striga-resistant/Striga-tolerant populations, inbred lines, varieties, and hybrids have been developed using drought-tolerant and Striga-resistant germplasm from diverse sources identified through several years of extensive testing in WCA (Badu-Apraku et al. 2001).
Genotype x environment interaction (GEI) poses a major challenge to breeders during multilocational evaluation of cultivars under S. hermonthica infestation because it may result in lack of consistency in the expression of resistance across environments. As a result, our strategy to ensure that the Striga resistance of the genotypes from our program does not break down in other environments has been to use the locations Ferkessédougou and Sinématiali in Côte d’Ivoire, Mokwa and Abuja in Nigeria and Angaredebougou, and Ina in Benin Republic as the principal screening and evaluation sites for Striga resistance for the maize populations, derived inbred lines, and cultivars in the program. We have observed in our program that Striga-resistant genotypes developed in a particular environment show the best performance in that particular environment. However, there is no reported situation where a Striga-resistant genotype in one environment has been found susceptible in another environment in WCA, suggesting that the resistance genes in our program could be durable (Kling et al. 2000). Despite the efforts to reduce GEI and hence improve the stability of performance of the products from our breeding program , recent studies by Badu-Apraku et al. (2006, 2008b) and Badu-Apraku and Lum (2010) have revealed significant GEI for most traits under Striga infestation, suggesting that the cultivars responded differently to the environments and that the differential response among cultivars was due to varying climatic conditions, soil type, and crop management practices at diverse test locations, which might have significantly influenced infestation and subsequent growth and development of Striga. Even though the test locations used in our screening for Striga resistance have provided a broad range of growing conditions, the significant GEI for grain yield and the Striga traits suggested that changes in the relative rankings of cultivars could be substantial across the diverse growing environments. This calls for the need to examine the GEI patterns and the consistency of the ranking of the Striga-resistant cultivars across the test environments in WCA. Therefore, 16 early-maturing cultivars were evaluated at two locations in Nigeria and three locations in the Republic of Benin from 2007 to 2009 to examine the grain yield, the stability, and the consistency of the rankings of the early-maturing cultivars under Striga-infested and Striga-free environments, assess the consistency of the rankings of the cultivars based on grain yield and other Striga traits under Striga-infested and Striga-free conditions, and assess the repeatability of measured traits and the test locations in Nigeria and Benin.
The combined analysis of variance showed significant cultivar and cultivar × environment interactions for grain yield and other traits under Striga-infested and Striga-free environments. The test of concordance was highly significant for grain yield (W = 0.68), number of emerged Striga plants (W = 0.74), and Striga damage (W = 0.56) under Striga infestation, indicating stability of resistance in the cultivars developed from diverse sources under artificial S. hermonthica infestation across environments. There was high consistency of the rankings of the cultivars for grain yield and other Striga-resistant traits under Striga-infested and Striga-free environments in Benin and Nigeria. Furthermore, the results indicated that S. hermonthica-resistant cultivars developed in Nigeria were also resistant in Benin. The AMMI biplot analysis for grain yield revealed POOL15SR/ACR94TZECOMP5-W/ACR94TZECOMP5-W and 2004 TZE-Y Pop DT STR C4 as the most stable cultivars with above average mean grain yield in Striga-infested environments and can be combined with other crop management options to control the parasite in the Striga endemic environments. Cultivars TZE Comp 5-W C7F2 and TZE Comp5-Y C6 S6 (Set B) had less Striga damage and number of emerged Striga plants across test environments. These cultivars could therefore serve as unique sources of favorable alleles for improving Striga resistance in maize in different production environments and farming systems.
4 Conclusion
Several international trials are packaged annually by IITA and sent out to NARS partners for evaluation. The test entries are usually composed of newly developed varieties by IITA and NARS scientists, and proprietary varieties developed by seed companies. A standard check variety is included along with field book containing the field design and sheets for data collection. The collaborator supplies a second check variety and carries out the trial in his/her location. At the end of the season, the field books are returned to IITA for analysis. The analysis has consistently showed statistically significant GEI, implying the need to subject the data to further analysis to decompose the GEI. Location effects carry the largest portion of GEI in trials conducted within and among countries in WCA; therefore, management of field trials needs refinement so that specific location factors, such as soil type, timing, rate and composition of fertilizer, weed control, and pest control, are done as precisely as possible. Using estimates of repeatability values, multivariate statistical methods, such as factor analysis, we grouped sites with similar characteristics for maize production. Five mega-environments were identified for evaluating maize germplasm. Sites within each mega-environment cut across several countries in WA, for example, mega-environment 1 had Katibougou (Mali), Angaradebou (Benin), Mokwa (Nigeria), Ejura, and Nyankpala (Ghana) as similar locations for maize growth and productivity. This mega-environment accounted for 22% of the total variation for grain yield in the study; others explained 19, 16, 15, and 15%, respectively. AMMI and GGE biplot were quite effective for identifying stable, high-yielding genotypes for specific locations. This is a definite advantage of GEI analysis.
References
Akaogu, I.C., B. Badu-Apraku, V.O. Adetimirin, I. Vroh, M. Oyekunle, and R.O. Akinwale. 2012. Genetic diversity assessment of extra-early maturing yellow maize inbreds and hybrid performance in Striga-infested and Striga-free environments. The Journal of Agricultural Science 151: 519–537.
Badu-Apraku, B., and A.F. Lum. 2010. The pattern of grain yield response of normal and quality protein maize cultivars in stress and nonstress environments. Agronomy Journal 102: 381–394.
Badu-Apraku, B., J. M. Fajemisin and A.O. Diallo. 1995. The performance of early and extra-early varieties across environments in West and Central Africa. In Contributing to food self-sufficiency: Maize Research and Development in West and Central Africa, ed. B. Badu-Apraku, M.O. Akoroda, M. Ouedraogo, and F.M. Quin, 149–159. Proceedings of a Regional Maize Workshop, 29 May–2 June 1995, IITA, Cotonou.
Badu-Apraku, B., M. A. B. Fakorede, M. Ouedraogo and R. J. Carsky. 2001. Impact, challenges, and prospects of maize research and development in West and Central Africa. Proc. Regional Maize Workshop, 4–7 May 1999, IITA-Cotonou, Benin Republic. WECAMAN/IITA, Ibadan, Nigeria.
Badu-Apraku, B., F.J. Abamu, A. Menkir, M.A.B. Fakorede, K. Obeng-Antwi, and C. The. 2003. Genotype by environment interactions in the regional early maize variety trials in West and Central Africa. Maydica 48: 93–104.
Badu-Apraku, B., M.A.B. Fakorede, and A.F. Lum. 2006. Evaluation of experimental varieties from recurrent selection for Striga resistance in two extra-early maize populations in the savannas of West and Central Africa. Experimental Agriculture 43: 183–200.
Badu-Apraku, B., A. Menkir, and A.F. Lum. 2007. Genetic variability for grain yield and components in an early tropical yellow maize population under Striga hermonthica infestation. Crop Improvement 20: 107–122.
Badu-Apraku, B., M.A.B. Fakorede, and A.F. Lum. 2008a. S1 family selection in early maturing maize population in Striga-infested and Striga-free environments. Crop Science 48: 1984–1994.
Badu-Apraku, B., A.F. Lum, M.A.B. Fakorede, A. Menkir, Y. Chabi, C. The, M. Abdulai, S. Jacob, and S. Agbaje. 2008b. Performance of cultivars derived from recurrent selection for grain yield and Striga resistance in early maize. Crop Science 48: 99–112.
Badu-Apraku, B., M.A.B. Fakorede, A.F. Lum, and R. Akinwale. 2009. Improvement of yield and other traits of extra-early maize under stress and nonstress environments. Agronomy Journal 101: 381–389.
Badu-Apraku, B., A. Menkir, S.O. Ajala, R.O. Akinwale, M. Oyekunle, and K. Obeng-Antwi. 2010. Performance of tropical early-maturing maize cultivars in multiple stress environments. Canadian Journal of Plant Science 90: 1–22.
Badu-Apraku, B., A.F. Lum, R.O. Akinwale, and M. Oyekunle. 2011a. Biplot analysis of diallel crosses of early maturing tropical yellow maize inbreds in stress and nonstress environments. Crop Science 51: 173–188.
Badu-Apraku, B., M. Oyekunle, and R.O. Akinwale. 2011b. Combining ability of early-maturing white maize inbreds under stress and nonstress environments. Agronomy Journal 103: 544–557.
Badu-Apraku, B., R.O. Akinwale, S.O. Ajala, A. Menkir, M.A.B. Fakorede, and M. Oyekunle. 2011c. Relationships among traits of tropical early maize cultivars in contrasting environments. Agronomy Journal 103: 717–729.
Badu-Apraku, B., R.O. Akinwale, K. Obeng-antwi, A. Haruna, R. Kanton, I. Usuman, S.G. Ado, N. Coulibaly, G.C. Yallou, and M. Oyekunle. 2013. Assessing the representativeness and repeatability of testing sites for drought-tolerant maize in West Africa. Canadian Journal of Plant Science 93: 699–714.
Cornelius, P.L., D.A. Van Sanford, and M. Seyedsadr. 1993. Clustering cultivars into groups without rank change interaction. Crop Science 33: 1193–1200.
Crossa, J., and P.L. Cornelius. 1997. Sites regression and shifted multiplicative model clustering of cultivar trials sites under heterogeneity of variances. Crop Science 37: 406–415.
Eberhart, S.A., and W.A. Russell. 1966. Stability parameters for comparing varieties. Crop Science 6: 36–40.
Fajemisin, J., Y. Efron, S.K. Kim, F.H. Khadr, Z.T. Dabrowski, J.H. Mareck, M. Bjarnason, V. Parkinson, L.A. Everett, and A. Diallo. 1985. Population and varietal development in maize for tropical Africa through resistance breeding approach. In Breeding strategies for maize production improvement in the tropics, ed. A. Brandolini and F. Salamini, 385–407. Florence: FAO and Istituto Agronomico per L’ Oltremare.
Fakorede, M.A.B. 1979. Interrelationships among grain yield and agronomic traits in a synthetic population of maize. Maydica 24: 181–192.
Fakorede, M.A.B., and M.O. Adeyemo. 1986. Genotype x environment components of variance for three types of maize varieties in the rainforest zone of south-west of Nigeria. Nigerian Journal of Agronomy 2: 43–46.
Fakorede, M.A.B., B. Badu-Apraku, A.F. Lum, A. Menkir and M. Ouedraogo. 2007. A review of NARES-IARC-Donor collaboration to develop demand-driven technologies for improved maize production in West and Central Africa. In Demand-driven technologies for sustainable maize production in West and Central Africa, ed. B. Badu-Apraku, M.A.B. Fakorede, A.F. Lum, A. Menkir, and M. Ouédraogo, 3–24. Proceedings of the Fifth Biennial Regional Maize Workshop, 3–6 May 2005, IITA, Cotonou, WECAMAN/IITA.
Falconer, D.S. 1981. Introduction to Quantitative Genetics, 267 pp. Essex: Longman House.
FAO. 2008. FAO Statistics, Food and Agriculture Organization of the United Nations, Rome. http://faostat.fao.org/.
Fehr, W.R. 1987. Principles of Cultivar Development. New: Macmillan Publishing.
Finlay, K.W., and G.N. Wilkinson. 1963. The analysis of adaptation in a plant breeding programme. Australian Journal of Agricultural Research 14: 742–754.
Gauch, H.G. 1988. Model selection and validation for yield trials with interaction. Biometrics 44: 705–715.
———. 2006. Statistical analysis of regional yield trial: AMMI analysis of factorial designs. Amsterdam: Elsevier.
Gauch, H.G., and R.W. Zobel. 1997. Identifying mega-environments and targeting genotypes. Crop Science 37: 311–326.
Gauch, H.G., H. Piepho, and P. Annicchiarico. 2008. Statistical analysis of yield trials by AMMI and GGE: Further considerations. Crop Science 48: 866–889.
Gollob, H.F. 1968. A statistical model which combines features of factor analytic and analysis of variance techniques. Psychometrika 33: 73–115.
Kim, S.K., J.M. Fajemisin, C. Thé, A. Adepoju, J.G. Kling, B. Badu-Apraku, M. Versteeg, R. Carsky, and S.T.O. Lagoke. 1998. Development of synthetic maize populations for resistance to Striga hermonthica. Plant Breeding 117: 203–209.
Kling, J. G., J. M. Fajemisin, B. Badu-Apraku, A. Diallo, A. Menkir, and A. Melake-Berhan. 2000. Striga resistance breeding in maize. In Breeding for Striga resistance in cereals, ed. B.I.G. Haussmann et al., 103–118. Proceedings of a Workshop held at IITA, Ibadan, Nigeria, 16–20 Aug 1999, Margraf Verlag, Weikersheim.
Kroschel, J. 1999. Analysis of the Striga problem: The first step towards future joint action. In Joint Action to Control Striga in Africa: Advances in Parasitic Weed Control at on Farm Level, ed. J. Kroschel et al., vol. 1, 3–26. Weikersheim: Margraf Verlag.
Mandel, J. 1969. The partitioning of interaction in analysis of variance. Journal of Research of the National Bureau of Standards B. Mathematical Sciences 73: 309–328.
———. 1971. A new analysis of variance model for non-additive data. Technometrics 13: 1–18.
Menkir, A., B. Badu-Apraku, C. Thé, and A. Adepoju. 2003. Evaluation of heterotic patterns of IITA lowland white maize inbred lines. Maydica 48: 161–170.
Odhiambo, G. D. and J. K. Ransom. 1994. Long term strategies for Striga control. In Maize research for stress environments, ed. D.C Jewell, et al., 263–266. Proceedings of the 4th Eastern and Southern Africa Regional Maize Conference, Harare, Zimbabwe, 28 March–1 April 1994, CIMMYT, Mexico.
Setimela, P.S., B. Vivek, M. Bänziger, J. Crossa, and F. Maideni. 2007. Evaluation of early to medium maturing open pollinated maize varieties in SADC region using GGE biplot based on the SREG model. Field Crops Research 103: 161–169.
Williams, E.J. 1952. The interpretation of interactions in factorial experiments. Biometrika 39: 65–81.
Yan, W., and M.S. Kang. 2003. GGE Biplot analysis: A graphical tool for breeders, geneticists, and agronomists. Boca Raton: CRC Press.
Yan, W., and N.A. Tinker. 2006. Biplot analysis of multi-environment trial data: Principles and applications. Canadian Journal of Plant Science 86: 623–645.
Yan, W., L. Hunt, Q. Sheng, and Z. Szlavnics. 2000. Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Science 40: 597–605.
Yan, W., M.S. Kang, S. Ma, S. Woods, and P.L. Cornelius. 2007. GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Science 47: 596–605.
Yan, W., J. Fregeau-Reid, D. Pageau, R. Martin, J. Mitchell-Fetch, M. Etienne, J. Rowsell, P. Scott, M. Price, B. de Haan, A. Cummiskey, J. Lajeunesse, J. Durand, and E. Sparry. 2010. Identifying essential test locations for oat breeding in Eastern Canada. Crop Science 50: 505–515.
Yan, W., D. Pageau, J. Fregeau-Reid, and J. Durand. 2011. Assessing the representativeness and repeatability of test locations for genotype evaluation. Crop Science 51: 1603–1610.
Yang, R., J. Crossa, P.L. Cornelius, and J. Burgueno. 2009. Biplot analysis of genotype x environment interaction: proceed with caution. Crop Science 49: 1564–1576.
Yates, F., and W.G. Cochran. 1938. The analysis of groups of experiments. Journal of Agriculture Science (Cambridge) 28: 556–580.
Zobel, R.W., M.J. Wright, and H.G. Gauch. 1988. Statistical analysis of a yield trial. Agronomy Journal 80: 388–393.
Author information
Authors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter

Cite this chapter
Badu-Apraku, B., Fakorede, M.A.B. (2017). Genotype x Environment Interaction and Repeatability of Traits. In: Advances in Genetic Enhancement of Early and Extra-Early Maize for Sub-Saharan Africa. Springer, Cham. https://doi.org/10.1007/978-3-319-64852-1_17
Download citation
DOI: https://doi.org/10.1007/978-3-319-64852-1_17
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-64851-4
Online ISBN: 978-3-319-64852-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)