
Abstract
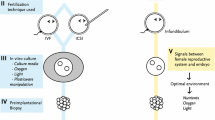
The periconception period comprises the final maturation of sperm and the processes of fertilization and early embryonic development, which take place in the oviduct. The final goal of these important events is to lead to establishment of pregnancy leading to the birth of healthy offspring. Studies in rodents and domestic animals have demonstrated that environmental conditions experienced during early development affect critical aspects of future growth, metabolism, gene expression, and physiology. Similarly, in vitro culture of embryos can be associated with changes in fetal growth, gene expression and regulation, and postnatal behavior.
In the oviduct, the cross talk between the mother and gametes/embryo begins after ovulation, between the oocyte and the female reproductive tract, and continues with the sperm and the early embryo after successful fertilization. These signals are mainly the result of direct interaction of gametes and embryos with oviductal and endometrial cells, influencing the microenvironment at the specific location. Identifying and understanding the mechanisms involved in this cross talk during the critical period of early reproductive events leading to pregnancy establishment could potentially lead to improvements in current in vitro embryo production systems in domestic mammals and humans. In this review, we discuss current knowledge of the short- and long-term consequences of in vitro embryo production on embryo development.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
1 Periconception Environment In Vivo
In vivo , mammalian oocytes and embryos develop in a complex and dynamic environment. First, in the ovarian follicle, the oocyte grows and matures, achieving full developmental competence (Fair et al. 1995). Prior to ovulation, the luteinizing hormone (LH) surge leads to the resumption of meiosis and final oocyte maturation. Oocyte maturation involves (i) nuclear maturation, i.e., progression from prophase I to metaphase II with extrusion of the first polar body, (ii) cytoplasmic maturation which involves organelle redistribution, and (iii) molecular maturation that involves the accumulation of specific mRNAs (Sirard 2001). It has been hypothesized that the quality of an oocyte is based on the presence of the appropriate set of mRNA and proteins stored during folliculogenesis (Wrenzycki et al. 2007). A defined oocyte-specific gene expression pattern arising during folliculogenesis is crucial for the acquisition of oocyte developmental competence; conversely, deficiencies in gene expression or dynamics that occur during follicle development may be linked to impaired oocyte competence (Eichenlaub-Ritter and Peschke 2002; Sirard et al. 2006).
After ovulation, in the oviduct, the oocyte undergoes fertilization and the first mitotic or cleavage divisions. Finally in the uterus, the blastocyst forms, hatches from the zona pellucida, and, depending on the species, either implants immediately [day 4.5 in rodents (Wang and Dey 2006) and days 6–10 in humans (Cha et al. 2012)], forms a large free-floating spherical structure which initiates implantation around day 40 (horses), or elongates and progressively attaches to the uterine wall (initiation between days 18–22 in cows, 15–18 in ewes, and 14–18 in pigs) (Senger 2003) (Fig. 1). These events must be properly orchestrated for successful pregnancy establishment and the delivery of a healthy offspring.

Schematic representation of the process of early embryo development in vivo in cattle. After ovulation, the matured oocyte is fertilized (D0) at the ampullary-isthmic junction, while the first cleavage division takes place around 24–30 h later in the isthmus (D1) followed by subsequent mitotic divisions up to the 16-cell stage (D4). At this stage, the embryo passes into the uterus through the uterotubal junction and forms a morula (D5–6) and then a blastocyst (D7). After hatching from the zona pellucida (D8), the morphology of the embryo changes to ovoid (D12–13), then tubular and filamentous (D16–17) before implantation begins on D19
Following copulation, semen is deposited in the female reproductive tract which, depending on the species, will be in the cranial vagina (cow, sheep, primates, and cat) or into the uterine lumen (pig, horse, and dog) (Senger 2003). Typically, billions of sperm are deposited; however, during transit through the female reproductive tract, this number is dramatically reduced as sperm are selected by different barriers [cervix and uterotubal junction (UTJ)]. Once in the oviduct, the sperm are held in a storage reservoir in the isthmus which preserves sperm fertility, reduces the incidence of polyspermy by releasing sperm gradually (Suarez 2008), and constitutes the immediate source of viable sperm at the time of ovulation (Hunter and Wilmut 1984). It has been suggested that the sperm storage site recognizes and selects a fertile sperm population in mammalian species (Teijeiro and Marini 2012). After ovulation, the fimbria of the infundibulum that surrounds the ovary allows the passage of the ovulated oocyte into the oviduct. At this point, both muscle layers and ciliated cells mechanically guide the oocyte into the lumen of the ampulla to the site of fertilization (Hunter 1988). In addition, the oocyte loses the cumulus cells, and the zona pellucida (ZP) becomes exposed directly to the oviductal fluid (OF) which prepares it for fertilization and minimizes polyspermy (Coy et al. 2008, 2012).
Once fertilization occurs, the embryo spends the first 3–4 days in the oviduct, depending on the species. In cattle, the embryo remains in the oviduct until approximately day 4 of pregnancy, by which time it is at about the 16-cell stage. Then, the embryo enters the uterus and by day 7 forms a blastocyst consisting of an inner cell mass, which gives rise to the fetus, and the trophectoderm (TE) , which forms the placenta. On days 9–10, the blastocyst hatches from the ZP and soon begins the process of elongation, which involves transitions from a spherical blastocyst on day 7 of gestation, through ovoid (days 12–13), tubular (days 14–15), and finally filamentous forms around days 16–17 (Fig. 1) (Degrelle et al. 2005). During elongation, the conceptus increases in size, more than 1000-fold, mainly through expansion of the trophectoderm (Betteridge et al. 1980) associated with an increase in protein content (Morris et al. 2000). After day 19, the elongated conceptus begins implantation with firm apposition and attachment of the trophectoderm to the endometrial luminal epithelium.
Proper communication between the conceptus-endometrium-corpus luteum (CL) is vital for pregnancy establishment. Progesterone (P4) synthesized by the CL acts indirectly via the endometrium to stimulate embryonic growth (Forde et al. 2009; Spencer et al. 2016). Conceptus elongation initiates interferon-τ (IFNT) production by TE cells (Roberts et al. 1999; Spencer and Bazer 2004; Robinson et al. 2008), which in cattle is the key signal for maternal recognition of pregnancy (Spencer and Bazer 2004). During maternal recognition of pregnancy, the mononuclear cells of the conceptus trophectoderm synthesize and secrete IFNT between days 10 and 21–25 with maximal production on days 14–16 (Bazer 1992; Roberts et al. 1999).
2 Periconception Environment In Vitro
In 1978, the first baby conceived by in vitro fertilization (IVF), Louise Brown, was born. Three years later, in 1981, the first calf obtained from IVF was born (Brackett et al. 1982). Although nearly 40 years have passed since then and many improvements have been made in in vitro embryo production (Paramio and Izquierdo 2016), even today in vitro systems are not as efficient as in vivo embryo production.
The goal of in vitro embryo production is to simulate as closely as possible the conditions that occur in vivo, to obtain high-quality embryos capable of continued development and implantation, and to result in viable births. Nowadays, in cattle, approximately 90% of oocytes cultured in vitro undergo nuclear and cytoplasmic maturation from which 80% are fertilized and cleave at least once (Lonergan et al. 2003a). Nevertheless, only between 30% and 40% reach the blastocyst stage (Rizos et al. 2008). In vitro, embryos are typically cultured until day 7 or 8 after fertilization, which corresponds to the blastocyst stage, when they are usually transferred into recipients. In heifers, the pregnancy rate following transfer of in vitro produced blastocysts is approximately 40–50% compared to about 70% when it comes to in vivo derived embryos (Hasler et al. 1995; Hoshi 2003). Thus, the challenge today is to improve current in vitro procedures providing high-quality embryos capable of continuing development and implantation after transfer to recipient and resulting in viable births.
2.1 In Vitro Maturation and Effects on Oocyte Developmental Competence
Cumulus-oocyte complexes (COCs) for research use are usually aspirated from ovaries recovered in the slaughterhouse, while those for commercial embryo transfer are recovered by transvaginal follicle puncture from live animals. In both cases, COCs are selected for in vitro maturation (IVM) based on morphological criteria as the compactness and thickness of cumulus and ooplasm homogeneity (Blondin and Sirard 1995). The cumulus cells play a critical role in the development of the oocyte by providing metabolites and nutrients, like pyruvate, oxaloacetic acid, and amino acids, thus stimulating them to resume meiosis and progress to metaphase II. Furthermore, cumulus cell expansion is an important marker for oocyte maturation, which is induced by gonadotrophin stimulation in vivo and in vitro leading to massive production of mucoid extracellular matrix protein (Chen et al. 1990). Recent evidence has shown that hyaluronic acid (HA) , an important component of the extracellular matrix, plays an important role not only in cumulus expansion but also in oocyte maturation and further embryo development (Marei et al. 2012). Furthermore, transcriptome studies have identified a large number of genes in both oocytes and their associated cumulus cells that are involved in oocyte maturation (Regassa et al. 2011). Thus, the gene expression patterns in the cumulus cells have been used as an indicator of oocyte quality (Tesfaye et al. 2009; Bunel et al. 2015). Also, the detection of glucose-6-phosphate dehydrogenase activity by brilliant cresyl blue (BCB) staining can be used as a predictor of oocyte quality, where BCB-positive oocytes are more competent to form blastocyst than BCB-negative oocytes (34.1% vs. 3.9%, respectively, P < 0.05) (Alm et al. 2005).
Oocyte developmental competence, often defined as the ability of the oocyte to mature, be fertilized, and develop to the blastocyst stage, has been associated with (i) the size of the antral follicle from which it is recovered, (ii) the stage of the follicular wave, and (iii) the site of maturation – in vivo or in vitro [for review, see Lonergan and Fair 2016]. Oocytes matured in vivo are of better quality than those matured in vitro, and this is reflected in the number of embryos obtained subsequently. Indeed, it has been shown that irrespective of whether in vitro culture (IVC) occurred in vivo or in vitro, when oocytes were matured in vivo, the resultant blastocyst rate was almost 80%, while when oocytes were matured in vitro, it was limited to about 35% (Rizos et al. 2002). In relation to in vitro maturation, oocytes originating from follicles bigger than 6 mm resulted in significantly more blastocysts than those from 2 to 6 mm follicles and those recovered prior to the LH surge (Rizos et al. 2002).
The environment to which the oocyte is exposed during maturation can influence the abundance of transcripts in the matured oocyte (Watson 2000; Lonergan et al. 2003a) and in the resulting blastocyst (Knijn et al. 2002; Russell et al. 2006). Recent evidence has shown that suboptimal conditions during oocyte IVM have an effect at the epigenetic level and on genomic imprinting (Anckaert and Fair 2015) in bovine (Heinzmann et al. 2011) and ovine (Colosimo et al. 2009) oocytes. In vitro models that mimic the in vivo situation of high-yielding dairy cows after calving, i.e., in which nonesterified fatty acid (NEFA) concentrations are elevated, indicate that not only the oocyte developmental capacity is affected but also that the phenotype of the resulting embryos is altered (Van Hoeck et al. 2011, 2013). Maturing oocytes for 24 h under high saturated NEFA conditions significantly altered metabolic footprints of day 7 embryos at the level of both gene transcription and gene function (Van Hoeck et al. 2011). Nevertheless, additional studies are required to investigate whether the expression and DNA methylation of imprinted genes in blastocysts, fetuses, and placental tissue derived from IVM oocytes are unaltered [for review see (Lonergan and Fair 2016)].
Supplementation of putative growth-promoting substances to the maturation media (e.g., gonadotrophins, steroids, and growth factors) typically results in a modest improvement in the proportion of oocytes reaching the blastocyst stage (Thompson 2000). Normally, oocytes submitted to in vitro maturation are recovered from small- to medium-sized follicles (2–8 mm) which are capable of nuclear maturation but did not have sufficient time to undergo normal cytoplasmic maturation. A range of cellular and chemical methods have been successfully employed to artificially inhibit the meiotic resumption of oocytes following removal from the follicle, thereby allowing cytoplasmic maturation in vitro in the absence of nuclear maturation [for review, see Sirard 2001]. However, none of these in vitro approaches substantially improved oocyte developmental competence.
2.2 In Vitro Fertilization and Effects on Oocyte Developmental Competence
The fate of an embryo is determined at fertilization . Delays in fertilization or fertilization by a damaged spermatozoon could conceivably lead to oocyte aging or the formation of a defective embryo, respectively (Tarin et al. 2000). Any damage to the sperm after ejaculation can lead not only to a reduced fertilization rate but also to the formation of embryos with reduced ability to develop to the blastocyst stage. This phenomenon has been demonstrated for embryos formed from sperm exposed to gossypol (Brocas et al. 1997), oxidative stress (Silva et al. 2007), and sorting for gender by flow cytometry (Wheeler et al. 2006; Wilson et al. 2006; Bermejo-Alvarez et al. 2008; Bermejo-Alvarez et al. 2010). In addition, the nature of the sire itself can affect cleavage and the ability of the resulting embryos to develop to the blastocyst stage and to establish pregnancy after transfer [for review see (Hansen et al. 2010)]. Studies in trout have shown that the oocyte is able to partially repair sperm with damaged DNA during the first cleavage; however, when DNA repair is inhibited, damaged sperm is able to fertilize the oocyte but leads to embryo loss (Perez-Cerezales et al. 2010; Perez-Cerezales et al. 2011).
Cryopreserved semen is the main source of sperm for in vitro fertilization, although the proportion of fully functional sperm in a frozen-thawed sample is quite low (Holt 1997). Therefore, before in vitro fertilization, it is necessary to separate a motile fraction of sperm using one of a variety of methods such as centrifugation on a density gradient. During this process, seminal plasma is washed away and thereby much of the antioxidant protection is lost (Marques et al. 2010). In sperm, moderate levels of reactive oxygen species (ROS) are necessary for sperm maturation, capacitation, hyperactivation, acrosome reaction, and sperm-egg fusion (Kothari et al. 2010). Nevertheless, in vitro the sperm are often exposed to supraphysiological levels of ROS (Du Plessis et al. 2008) which can affect cell membranes, DNA, and mitochondria (Agarwal et al. 2006), leading to low fertilization rates and poor embryo quality (Silva et al. 2007; Jang et al. 2010). Antioxidants have been used to decrease the impact of oxidative stress, thereby improving sperm quality and their ability to fertilize an egg and consequently increasing the number of embryos obtained (Roca et al. 2004; Roca et al. 2005; Sapanidou et al. 2015) and their quality (Jang et al. 2010; Pang et al. 2016). However, other studies have failed to demonstrate any beneficial effects following the use of antioxidants during IVF and have even reported impaired fertilization and blastocyst rates (Ali et al. 2003; Goncalves et al. 2010; Marques et al. 2010). Therefore, more studies are needed to elucidate which antioxidants are best and when they should be added to improve the efficiency of IVF.
2.3 In Vitro Embryo Culture: Short- and Long-Term Effects
Embryo culture is the longest step during the process of in vitro embryo production and the step during which the greatest reduction in development occurs, achieving only 30–40% of blastocyst rate. The presumptive zygotes are recovered around 18 h after fertilization and usually are cultured until days 7–9 (Fig. 2). It has been clearly shown that the intrinsic quality of the oocyte is the main factor affecting blastocyst yield, while the postfertilization culture environment affects the quality of the produced blastocysts (Rizos et al. 2002).

Schematic representation of in vitro embryo production in cattle. In vitro maturation (IVM) of cumulus-oocyte complexes (COCs) is carried out for 24 h under an atmosphere of 38.5 °C, 5% CO2, and maximum humidity. In vitro fertilization (IVF) is accomplished by co-incubation of matured COCs with selected motile sperm at a final concentration of 1 × 106 for 18 h, under the same conditions as IVM. In vitro culture (IVC) of the presumptive zygotes is maintained until D7–D9 at 38.5 °C, 5% CO2, 5% O2, and maximum humidity. The embryos can be cultured in different systems including coculture with BOECs in monolayers, suspension or polarized, and with products derived from BOECs such as microvesicles and conditioned media. Oviductal fluid recovered from the oviduct may also be used as well as its microvesicles
Culture conditions in vitro produce embryos of inferior quality compared to those derived from in vivo in terms of morphology, cryotolerance, transcript expression profiles, and pregnancy rate after transfer [for review, see Lonergan 2007]. To highlight the importance of developing in vitro systems that are as close as possible to the in vivo situation, different experiments have been carried out alternating in vitro with in vivo embryo culture. Thus, the culture of in vitro produced bovine zygotes in vivo in the sheep (Rizos et al. 2002), cow (Tesfaye et al. 2007), or mouse (Rizos et al. 2007) oviduct increases the quality of the resulting blastocysts so that it resembles that of totally in vivo produced embryos. Conversely, the in vitro culture of in vivo produced bovine zygotes results in blastocysts of low quality (Rizos et al. 2002). In addition, we have demonstrated that in vitro produced bovine zygotes cultured either in vitro or in vivo in the sheep oviduct at different stages of development up to the blastocyst stage exhibit a clear temporal sensitivity to their culture environment on their transcriptome and cryotolerance (Lonergan et al. 2003b). Thus, blastocysts cultured for the first 2 days in vivo and the last 4 days in vitro had the lowest survival rates after cryopreservation; those cultured only for the last 2 days in vitro had intermediate rates of survival, and those which spent the last 4 days of culture in vivo had high rates of survival, compared with those cultured entirely in vivo. Based on these studies, it was concluded that the period around the time of embryonic genome activation (EGA) is critical to the quality of the embryo. Similarly, Gad et al. (2012) investigated the consequences of culture conditions before and during the EGA on bovine embryonic developmental rates and global gene expression patterns using the homologous cow oviduct. Embryo development was similar irrespective of where culture took place; however, the transcriptome of the blastocysts was clearly influenced by culture conditions, confirming once more the significant influence of culture conditions during EGA.
In vitro embryo culture has also been associated with epigenetic alterations in the embryo. In vivo, after fertilization the embryo’s genome is epigenetically reprogrammed. This process involves the creation of the methylation patterns needed for normal development by activating and silencing specific genes (Reik et al. 2001). During this period, the embryo is especially vulnerable to in vitro induced epigenetic defects (El Hajj and Haaf 2013). Indeed, animal studies have revealed a link between different assisted reproductive technologies (ARTs) and imprinting disorders, via altered DNA-methylation patterns and histone codes (Urrego et al. 2014). In addition, imprinting disorders are more prevalent in gametes and embryos after ARTs than in their counterparts derived from in vivo production (Urrego et al. 2014). In vitro embryo culture has been associated with abnormal reprogramming of imprinted genes such as SNPRN, IGF2, or H19 in cattle (Nowak-Imialek et al. 2008; Curchoe et al. 2009; Suzuki et al. 2009) and mice (Khosla et al. 2001). These alterations to the epigenetic profile may have a direct effect on the subsequent embryo and fetal development.
From the above, it can be concluded that the actual in vitro culture conditions provide a suboptimal environment for early embryonic development and have short- and long-term consequences. The short-term consequences include alterations in morphology, cryotolerance, and gene expression patterns, while the long-term consequences are reflected in abnormal offspring development and behavior (Rizos et al. 2008).
Embryos can be cultured in defined or semi-defined media, cocultured with oviduct epithelial cells or their conditioned media and with extracellular vesicles (Maillo et al. 2016). Nowadays, one of the most commonly used media for the culture of bovine embryos is synthetic oviductal fluid (SOF) which is frequently supplemented with fetal calf serum (FCS) and/or bovine serum albumin (BSA) (Tervit et al. 1972; Holm et al. 1999). The presence of serum in the IVC media has a stimulatory effect on the speed of development, with more blastocysts on day 6 of culture than either in its absence or with BSA (Gutierrez-Adan et al. 2001; Rizos et al. 2003). However, serum can have a negative effect on embryo quality as manifested by reduced cryotolerance and altered gene expression (Lazzari et al. 2002; Rizos et al. 2003; Wrenzycki et al. 2005) and pregnancy rate after transfer. Lazzari et al. (2002) evidenced that IVC of bovine embryos in the presence of serum or BSA significantly increased the number of cells in day 7 blastocysts and the relative abundance of transcripts for several genes including HSP70.1, Cu ⁄ Zn-SOD, GLUT-3, GLUT-4, bFGF, and IGFI-R compared with embryos cultured in vivo (either in the sheep oviduct or produced entirely in vivo). Moreover, these deviations were linked to gestation length and birth weight of the derived calves. Both in vitro systems, FCS and BSA , were associated with a significantly elevated incidence of deviations in embryonic development and a higher proportion of calves with increased birth weight (Lazzari et al. 2002). In a recent study, it was shown that BSA removal over a 24 h period (from D6 to D7) in an individual embryo culture system decreased embryo development and cell counts in the inner cell mass, although the embryos tended to improve their survival after vitrification and also to result in a lower incidence of miscarriage (Murillo-Ríos et al. 2016).
Culture of embryos with FCS has been associated with alterations in the phenotype of newborn offspring in cattle (Farin et al. 2006), sheep (Walker et al. 1996), and mice (Fernandez-Gonzalez et al. 2004) named “large offspring syndrome (LOS) .” LOS is characterized by aberrant placental development, extended gestation length, sudden perinatal death, breathing difficulties, more male calves, and large size at birth (Farin et al. 2006). Insulin-like growth factor 2 (IGF2) is an imprinted gene that regulates fetal and placental development in cattle and other species (Constancia et al. 2002). In sheep, the altered expression of IGF2R has been correlated with the incidence of LOS (Young et al. 2001) suggesting that IGF2 could be sensitive to epigenetic disorders during in vitro conditions. In addition, fetuses and placental tissue derived from IVP embryos presented an aberrant expression of imprinted and non-imprinted genes (Perecin et al. 2009; Farin et al. 2010). These phenotypic alterations may be the result of failure to properly establish or maintain DNA methylation and histone modifications during in vitro conditions (Farin et al. 2006). This also reflects the likelihood that epigenetic alterations during early embryo development are maintained in subsequent embryo and fetal development.
Oxidative stress is also important for the developing embryo during the embryo culture step. Substances such as ammonia, oxygen radicals, or growth factors can produce lipid peroxidation, membrane injury, and structural damage, leading to decreased cryotolerance and apoptosis (Somfai et al. 2007). Thus, antioxidant supplementation of the culture media has been shown to protect the embryo against oxidative stress. It also helps to maintain intra- and extracellular redox balance, which is necessary to reduce the toxicity of ROS, improving embryo development, increasing cryotolerance and cell differentiation, and inhibiting apoptosis during culture (Guerin and Menezo 2011; Takahashi 2012). Vitamins such as tocopherol, ascorbic acid, folic acid, or cyanocobalamin play a key role in reducing oxidative damage and improving blastocyst development rate in mouse embryos (Wang et al. 2002). Recently, it has been demonstrated that the supplementation of culture media with other antioxidants, such as cocretin which has a high antioxidant capacity, leads to an improvement of cryotolerance of bovine embryos (Zullo et al. 2016). Another antioxidant, retinol, seems to have a positive effect on early embryo development and quality (Livingston et al. 2004). Similar results were reported by Lawrence et al. (2004) suggesting that retinol protects early bovine embryos against damaging effect of heat stress during in vitro culture.
Despite the many efforts to improve in vitro culture media, conditions within the oviduct have not been fully recapitulated in vitro. As knowledge concerning embryo requirements during early embryonic development increases, in vitro culture systems have evolved to mimic more precisely what occurs in the oviduct during this period. Therefore, at this moment, it is known that the embryo requires an evolving array of energy substrates (Gardner et al. 1996; Quesenberry et al. 2015); consumption of pyruvate and glucose is low until the 16-cell stage and increases significantly with morula compaction and blastocyst formation (de Souza et al. 2015).
As previously mentioned, in an attempt to mimic in vivo conditions and improve the quality of the embryos produced, different systems of embryo culture have been developed. They offer unique advantages such as the gradual change of culture media to suit the specific requirements of the developing embryo, thus overcoming limitations of conventional culture systems. Coculture in vitro with bovine oviductal epithelial cells (BOECs) has been considered to help with the production of good quality embryos (Ulbrich et al. 2010). These cells can be grown as monolayers, cell suspensions, or as polarized cultures (Fig. 2). A study by Cordova et al. (2013) showed that the use of BOEC in in vitro embryo culture at the early stages of embryo development, up to day 4, improves embryo development and embryo quality in terms of expression of specific gene transcripts. This period of culture coincides with the in vivo period when the embryo is still in the oviduct. However, the drawback of monolayers is that they dedifferentiate, losing important morphological characteristics (Rottmayer et al. 2006) including reduction of cell height, loss of cilia, and loss of secretory granules and bulbous protrusions (Thibodeaux et al. 1992; Walter 1995). Recently, we demonstrated that an established BOEC monolayer can be used successfully for coculture with no differences in embryo development when compared either with coculture with fresh recovered cells or normal culture in SOF (Lopera-Vásquez et al. 2016). Furthermore, the quality of the produced blastocysts in terms of cryotolerance and number of TE and ICM cells was higher than those produced using media supplemented with FCS. This method gives an advantage over the classical coculture systems as it provides homogeneous results. An alternative to monolayers is the short-term (24 h) epithelial cell suspension culture, which maintains morphological characteristics as well as the gene markers present in the cell in vivo such as OVGP1, estrogen, and P4 receptors (Rottmayer et al. 2006). Preliminary results from our group showed that BOECs in suspension cultures are closer to in vivo controls than monolayers in terms of morphology and oviductal epithelial cell markers OVGP1, GPX4, and FOXJ1 (Hamdi et al. 2015). Finally, polarized cell cultures maintain the polarized asymmetrical structure of the oviductal epithelial cells, and it seems that this system preserves detailed morphological features of the porcine oviduct as well as oviduct-specific markers (Miessen et al. 2011).
Oviductal fluid (OF) has been used as a supplement during in vitro embryo production. Its composition is very complex, containing simple and complex carbohydrates, ions, lipids, phospholipids, and proteins (Avilés et al. 2010). Porcine oocytes treated with OF before fertilization showed significantly increased cleavage rates and blastocyst yield, suggesting that OF protects the embryo against adverse effects on mitochondrial DNA transcription or replication and apoptosis (Lloyd et al. 2009). In contrast, Cebrian-Serrano et al. (2013) evidenced that the exposure of cattle oocytes to OF before fertilization had no effect on embryo development and morphology of the resulting blastocysts; however, blastocysts produced from oocytes treated with OF showed differences in specific transcripts (Cebrian-Serrano et al. 2013). Recently, we showed that low concentrations of OF (<5%) in bovine embryo culture media as a substitute for serum had a positive effect on development and quality in terms of cryotolerance, cell number, and expression of qualitatively related genes (Lopera-Vasquez et al. 2015).
Extracellular vesicles (EVs) is a general term encompassing several different vesicle types, released by somatic cells, that are present in body fluids and contain bioactive molecules (i.e., proteins, RNAs, mRNAs, and miRNAs) (Simons and Raposo 2009; Silveira et al. 2012) and lipids (Raposo and Stoorvogel 2013). The EV denomination is commonly size and origin associated, being exosomes (30–200 nm) from endosomal origin and microvesicles (MV) (100–1000 nm) from the plasma membrane. EVs are important in intercellular communication and play a key role in the regulation of physiological and pathological processes. It has been demonstrated that EV can horizontally transfer mRNAs to other cells, whereupon the mRNA can then be translated into functional proteins in the new location (Hergenreider et al. 2012). EVs have been identified in vivo in all body fluids including amniotic fluid, urine, and blood (Simpson et al. 2008). Currently, in reproduction, knowledge of the role of these secreted vesicles is limited to those from follicular fluid (Silveira et al. 2012), the endometrial environment (Ng et al. 2013), seminal plasma (Piehl et al. 2013), and uterine fluid (Burns et al. 2014). Recently, Burns et al. (2016) demonstrated that EVs emanate from both the conceptus trophectoderm and uterine epithelia and are involved in intercellular communication between those tissues during the establishment of pregnancy in sheep. Therefore, EV can be used as a supplement during in vitro embryo culture. Thus, Saadeldin et al. (2014) showed that the addition of exosomes isolated from the conditioned medium of parthenogenetic embryos increased the developmental competence of cloned embryos. In vitro derived embryos are also known to secrete EVs into culture media, where they may play a role in promoting development (Saadeldin et al. 2015). Recently, we provided evidence that EVs derived from BOEC conditioned media improve blastocyst quality and induce cryoprotection in in vitro cultures to the same extent as classical coculture with fresh BOEC monolayers (Lopera-Vásquez et al. 2016). Thus, the presence of EV in OF and their effect on early embryonic development may be of great importance and may provide information and new insights on early embryo-maternal communication and improve embryo quality in our current IVP systems.
3 Conclusion
During the last few decades, many advances have been achieved in our understanding of early reproductive events based on in vivo and in vitro studies, and assisted reproductive technologies are commonly used in humans and many animal species with success. Nevertheless, the quality of in vitro embryos is still inferior compared to their in vivo counterparts, and in vitro conditions have short- and long-term effects on the resulting embryo, fetus, and offspring. Therefore, a better understanding of how the embryo develops physiologically in the reproductive tract (oviduct and uterus) will provide the knowledge to develop new strategies to decipher the mechanisms involved in oocyte developmental competence that help and improve the current systems of in vitro embryo production. Together, such improvements will lead to the production of better quality embryos. Moreover, proper in vitro models that mimic the physiological situation as closely as possible will be developed.
References
Agarwal A, Said TM, Bedaiwy MA, Banerjee J, Alvarez JG (2006) Oxidative stress in an assisted reproductive techniques setting. Fertil Steril 86:503–512
Ali AA, Bilodeau JF, Sirard MA (2003) Antioxidant requirements for bovine oocytes varies during in vitro maturation, fertilization and development. Theriogenology 59:939–949
Alm H, Torner H, Lohrke B, Viergutz T, Ghoneim IM, Kanitz W (2005) Bovine blastocyst development rate in vitro is influenced by selection of oocytes by brilliant cresyl blue staining before IVM as indicator for glucose-6-phosphate dehydrogenase activity. Theriogenology 63:2194–2205
Anckaert E, Fair T (2015) DNA methylation reprogramming during oogenesis and interference by reproductive technologies: studies in mouse and bovine models. Reprod Fertil Dev 27:739–754
Avilés M, Gutiérrez-Adán A, Coy P (2010) Oviductal secretions: will they be key factors for the future ARTs? Mol Hum Reprod 16:896–906
Bazer FW (1992) Mediators of maternal recognition of pregnancy in mammals. Proc Soc Exp Biol Med 199:373–384
Bermejo-Alvarez P, Lonergan P, Rath D, Gutierrez-Adan A, Rizos D (2010) Developmental kinetics and gene expression in male and female bovine embryos produced in vitro with sex-sorted spermatozoa. Reprod Fertil Dev 22:426–436
Bermejo-Alvarez P, Rizos D, Rath D, Lonergan P, Gutierrez-Adan A (2008) Can bovine in vitro-matured oocytes selectively process X- or Y-sorted sperm differentially? Biol Reprod 79:594–597
Betteridge KJ, Eaglesome MD, Randall GC, Mitchell D (1980) Collection, description and transfer of embryos from cattle 10–16 days after oestrus. J Reprod Fertil 59:205–216
Blondin P, Sirard MA (1995) Oocyte and follicular morphology as determining characteristics for developmental competence in bovine oocytes. Mol Reprod Dev 41:54–62
Brackett BG, Bousquet D, Boice ML, Donawick WJ, Evans JF, Dressel MA (1982) Normal development following in vitro fertilization in the cow. Biol Reprod 27:147–158
Brocas C, Rivera RM, Paula-Lopes FF, McDowell LR, Calhoun MC, Staples CR, Wilkinson NS, Boning AJ, Chenoweth PJ, Hansen PJ (1997) Deleterious actions of gossypol on bovine spermatozoa, oocytes, and embryos. Biol Reprod 57:901–907
Bunel A, Jorssen EP, Merckx E, Leroy JL, Bols PE, Sirard MA (2015) Individual bovine in vitro embryo production and cumulus cell transcriptomic analysis to distinguish cumulus-oocyte complexes with high or low developmental potential. Theriogenology 83:228–237
Burns G, Brooks K, Wildung M, Navakanitworakul R, Christenson LK, Spencer TE (2014) Extracellular vesicles in luminal fluid of the ovine uterus. PLoS One 9:e90913
Burns GW, Brooks KE, Spencer TE (2016) Extracellular vesicles originate from the conceptus and uterus during early pregnancy in sheep. Biol Reprod 94(3):56
Cebrian-Serrano A, Salvador I, Garcia-Rosello E, Pericuesta E, Perez-Cerezales S, Gutierrez-Adan A, Coy P, Silvestre MA (2013) Effect of the bovine oviductal fluid on in vitro fertilization, development and gene expression of in vitro-produced bovine blastocysts. Reprod Domest Anim 48:331–338
Cha J, Sun X, Dey SK (2012) Mechanisms of implantation: strategies for successful pregnancy. Nat Med 18:1754–1767
Chen L, Wert SE, Hendrix EM, Russell PT, Cannon M, Larsen WJ (1990) Hyaluronic acid synthesis and gap junction endocytosis are necessary for normal expansion of the cumulus mass. Mol Reprod Dev 26:236–247
Colosimo A, Di Rocco G, Curini V, Russo V, Capacchietti G, Berardinelli P, Mattioli M, Barboni B (2009) Characterization of the methylation status of five imprinted genes in sheep gametes. Anim Genet 40:900–908
Constancia M, Hemberger M, Hughes J, Dean W, Ferguson-Smith A, Fundele R, Stewart F, Kelsey G, Fowden A, Sibley C, Reik W (2002) Placental-specific IGF-II is a major modulator of placental and fetal growth. Nature 417:945–948
Cordova A, Perreau C, Schmaltz-Panneau B, Locatelli Y, Ponsart C, Mermillod P (2013) Use of an in vitro model in bovine to evidence a functional and molecular dialogue between preimplantation embryo and oviduct epithelial cells. Gynecol Obstet Fertil 41:537–539
Coy P, Cánovas S, Mondéjar I, Saavedra MD, Romar R, Grullón L, Matás C, Avilés M (2008) Oviduct-specific glycoprotein and heparin modulate sperm-zona pellucida interaction during fertilization and contribute to the control of polyspermy. Proc Natl Acad Sci U S A 105:15809–15814
Coy P, Jimenez-Movilla M, Garcia-Vazquez FA, Mondejar I, Grullon L, Romar R (2012) Oocytes use the plasminogen-plasmin system to remove supernumerary spermatozoa. Hum Reprod 27:1985–1993
Curchoe CL, Zhang S, Yang L, Page R, Tian XC (2009) Hypomethylation trends in the intergenic region of the imprinted IGF2 and H19 genes in cloned cattle. Anim Reprod Sci 116:213–225
de Souza D, Salles L, Rosa e Silva A (2015) Aspects of energetic substrate metabolism of in vitro and in vivo bovine embryos. Braz J Med Biol Res 48:191–197
Degrelle SA, Campion E, Cabau C, Piumi F, Reinaud P, Richard C, Renard JP, Hue I (2005) Molecular evidence for a critical period in mural trophoblast development in bovine blastocysts. Dev Biol 288:448–460
Du Plessis S, Makker K, Desai N, Agarwal A (2008) Impact of oxidative stress on IVF. Expert Rev Obstet Gynecol 3:539–554
Eichenlaub-Ritter U, Peschke M (2002) Expression in in-vivo and in-vitro growing and maturing oocytes: focus on regulation of expression at the translational level. Hum Reprod Update 8:21–41
El Hajj N, Haaf T (2013) Epigenetic disturbances in in vitro cultured gametes and embryos: implications for human assisted reproduction. Fertil Steril 99:632–641
Fair T, Hyttel P, Greve T (1995) Bovine oocyte diameter in relation to maturational competence and transcriptional activity. Mol Reprod Dev 42:437–442
Farin CE, Farmer WT, Farin PW (2010) Pregnancy recognition and abnormal offspring syndrome in cattle. Reprod Fertil Dev 22:75–87
Farin PW, Piedrahita JA, Farin CE (2006) Errors in development of fetuses and placentas from in vitro-produced bovine embryos. Theriogenology 65:178–191
Fernandez-Gonzalez R, Moreira P, Bilbao A, Jimenez A, Perez-Crespo M, Ramirez MA, Rodriguez De Fonseca F, Pintado B, Gutierrez-Adan A (2004) Long-term effect of in vitro culture of mouse embryos with serum on mRNA expression of imprinting genes, development, and behavior. Proc Natl Acad Sci U S A 101:5880–5885
Forde N, Carter F, Fair T, Crowe MA, Evans AC, Spencer TE, Bazer FW, McBride R, Boland MP, O'Gaora P, Lonergan P, Roche JF (2009) Progesterone-regulated changes in endometrial gene expression contribute to advanced conceptus development in cattle. Biol Reprod 81:784–794
Gad A, Hoelker M, Besenfelder U, Havlicek V, Cinar U, Rings F, Held E, Dufort I, Sirard MA, Schellander K, Tesfaye D (2012) Molecular mechanisms and pathways involved in bovine embryonic genome activation and their regulation by alternative in vivo and in vitro culture conditions. Biol Reprod 87:100
Gardner DK, Lane M, Calderon I, Leeton J (1996) Environment of the preimplantation human embryo in vivo: metabolite analysis of oviduct and uterine fluids and metabolism of cumulus cells. Fertil Steril 65:349–353
Goncalves FS, Barretto LS, Arruda RP, Perri SH, Mingoti GZ (2010) Effect of antioxidants during bovine in vitro fertilization procedures on spermatozoa and embryo development. Reprod Domest Anim 45:129–135
Guerin P, Menezo Y (2011) Review: role of tubal environment in preimplantation embryogenesis: application to co-culture assays. Zygote 19:47–54
Gutierrez-Adan A, Lonergan P, Rizos D, Ward FA, Boland MP, Pintado B, de la Fuente J (2001) Effect of the in vitro culture system on the kinetics of blastocyst development and sex ratio of bovine embryos. Theriogenology 55:1117–1126
Hamdi M, Lopera R, Maillo V, Núñez C, Gutierrez-Adan A, Lonergan P, Bermejo-Alvarez P, Rizos D (2015) Bovine oviduct epithelial cells: an in vitro model to study early embryo-maternal communication. Anim Reprod 12:798
Hansen PJ, Block J, Loureiro B, Bonilla L, Hendricks KE (2010) Effects of gamete source and culture conditions on the competence of in vitro-produced embryos for post-transfer survival in cattle. Reprod Fertil Dev 22:59–66
Hasler JF, Henderson WB, Hurtgen PJ, Jin ZQ, McCauley AD, Mower SA, Neely B, Shuey LS, Stokes JE, Trimmer SA (1995) Production, freezing and transfer of bovine IVF embryos and subsequent calving results. Theriogenology 43:141–152
Heinzmann J, Hansmann T, Herrmann D, Wrenzycki C, Zechner U, Haaf T, Niemann H (2011) Epigenetic profile of developmentally important genes in bovine oocytes. Mol Reprod Dev 78:188–201
Hergenreider E, Heydt S, Tréguer K, Boettger T, Horrevoets AJG, Zeiher AM, Scheffer MP, Frangakis AS, Yin X, Mayr M, Braun T, Urbich C, Boon RA, Dimmeler S (2012) Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs. Nat Cell Biol 14:249–256
Holm P, Booth PJ, Schmidt MH, Greve T, Callesen H (1999) High bovine blastocyst development in a static in vitro production system using SOFaa medium supplemented with sodium citrate and myo-inositol with or without serum-proteins. Theriogenology 52:683–700
Holt WV (1997) Alternative strategies for the long-term preservation of spermatozoa. Reprod Fertil Dev 9:309–319
Hoshi H (2003) In vitro production of bovine embryos and their application for embryo transfer. Theriogenology 59:675–685
Hunter RHF (1988) The fallopian tubes: their role in fertility and infertility. Springer-Verlag, Berlin, 191 pp.
Hunter RH, Wilmut I (1984) Sperm transport in the cow: peri-ovulatory redistribution of viable cells within the oviduct. Reprod Nutr Dev 24:597–608
Jang HY, Kim YH, Kim BW, Park IC, Cheong HT, Kim JT, Park CK, Kong HS, Lee HK, Yang BK (2010) Ameliorative effects of melatonin against hydrogen peroxide-induced oxidative stress on boar sperm characteristics and subsequent in vitro embryo development. Reprod Domest Anim 45:943–950
Khosla S, Dean W, Brown D, Reik W, Feil R (2001) Culture of preimplantation mouse embryos affects fetal development and the expression of imprinted genes. Biol Reprod 64:918–926
Knijn HM, Wrenzycki C, Hendriksen PJ, Vos PL, Herrmann D, van der Weijden GC, Niemann H, Dieleman SJ (2002) Effects of oocyte maturation regimen on the relative abundance of gene transcripts in bovine blastocysts derived in vitro or in vivo. Reproduction 124:365–375
Kothari S, Thompson A, Agarwal A, du Plessis SS (2010) Free radicals: their beneficial and detrimental effects on sperm function. Indian J Exp Biol 48:425–435
Lawrence JL, Payton RR, Godkin JD, Saxton AM, Schrick FN, Edwards JL (2004) Retinol improves development of bovine oocytes compromised by heat stress during maturation. J Dairy Sci 87:2449–2454
Lazzari G, Wrenzycki C, Herrmann D, Duchi R, Kruip T, Niemann H, Galli C (2002) Cellular and molecular deviations in bovine in vitro-produced embryos are related to the large offspring syndrome. Biol Reprod 67:767–775
Livingston T, Eberhardt D, Edwards JL, Godkin J (2004) Retinol improves bovine embryonic development in vitro. Reprod Biol Endocrinol 2:83
Lloyd A, Pratt K, Siebrasse E, Moran MD, Duina AA (2009) Uncoupling of the patterns of chromatin association of different transcription elongation factors by a histone H3 mutant in Saccharomyces cerevisiae. Eukaryot Cell 8:257–260
Lonergan P (2007) State-of-the-art embryo technologies in cattle. Soc Reprod Fertil Suppl 64:315–325
Lonergan P, Fair T (2016) Maturation of oocytes in vitro. Annu Rev Anim Biosci 4:255–268
Lonergan P, Rizos D, Gutierrez-Adan A, Fair T, Boland MP (2003a) Effect of culture environment on embryo quality and gene expression – experience from animal studies. Reprod Biomed Online 7:657–663
Lonergan P, Rizos D, Kanka J, Nemcova L, Mbaye AM, Kingston M, Wade M, Duffy P, Boland MP (2003b) Temporal sensitivity of bovine embryos to culture environment after fertilization and the implications for blastocyst quality. Reproduction 126:337–346
Lopera-Vasquez R, Hamdi M, Maillo V, Lloreda V, Coy P, Gutierrez-Adan A, Bermejo-Alvarez P, Rizos D (2015) Effect of bovine oviductal fluid on development and quality of bovine embryos produced in vitro. Reprod Fertil Dev. doi:10.1071/RD15238
Lopera-Vásquez R, Hamdi M, Fernandez-Fuertes B, Maillo V, Beltrán-Breña P, Calle A, Redruello A, López-Martín S, Gutierrez-Adán A, Yañez-Mó M, Ramirez MÁ, Rizos D (2016) Extracellular vesicles from BOEC in in vitro embryo development and quality. PLoS One 11:e0148083
Maillo V, Sánchez-Calabuig MJ, Lopera-Vasquez R, Hamdi M, Gutierrez-Adan A, Lonergan P, Rizos D (2016) Oviductal response to gametes and early embryos in mammals. Reproduction 152:R127–R141
Marei WF, Ghafari F, Fouladi-Nashta AA (2012) Role of hyaluronic acid in maturation and further early embryo development of bovine oocytes. Theriogenology 78:670–677
Marques A, Santos P, Antunes G, Chaveiro A, Moreira da Silva F (2010) Effect of alpha-tocopherol on bovine in vitro fertilization. Reprod Domest Anim 45:81–85
Miessen K, Sharbati S, Einspanier R, Schoen J (2011) Modelling the porcine oviduct epithelium: a polarized in vitro system suitable for long-term cultivation. Theriogenology 76:900–910
Morris DG, Diskin MG, Sreenan JM (2000) Protein synthesis and phosphorylation by elongating 13–15-day-old cattle blastocysts. Reprod Fertil Dev 12:39–44
Murillo-Ríos A, Maillo V, Muñoz M, Gutiérrez-Adán A, Carrocera S, Martín-González D, Fernandez-Buznego A, Gómez E (2016) Short- and long-term outcomes of the absence of protein during bovine blastocyst formation in vitro. Reprod Fertil Dev 29(6):1064–1073
Ng YH, Rome S, Jalabert A, Forterre A, Singh H, Hincks CL, Salamonsen LA (2013) Endometrial exosomes/microvesicles in the uterine microenvironment: a new paradigm for embryo-endometrial cross talk at implantation. PLoS One 8:e58502
Nowak-Imialek M, Wrenzycki C, Herrmann D, Lucas-Hahn A, Lagutina I, Lemme E, Lazzari G, Galli C, Niemann H (2008) Messenger RNA expression patterns of histone-associated genes in bovine preimplantation embryos derived from different origins. Mol Reprod Dev 75:731–743
Pang YW, Sun YQ, Sun WJ, WH D, Hao HS, Zhao SJ, Zhu HB (2016) Melatonin inhibits paraquat-induced cell death in bovine preimplantation embryos. J Pineal Res 60:155–166
Paramio MT, Izquierdo D (2016) Recent advances in in vitro embryo production in small ruminants. Theriogenology 86(1):152–159
Perecin F, Meo SC, Yamazaki W, Ferreira CR, Merighe GK, Meirelles FV, Garcia JM (2009) Imprinted gene expression in in vivo- and in vitro-produced bovine embryos and chorio-allantoic membranes. Genet Mol Res 8:76–85
Perez-Cerezales S, Gutierrez-Adan A, Martinez-Paramo S, Beirao J, Herraez MP (2011) Altered gene transcription and telomere length in trout embryo and larvae obtained with DNA cryodamaged sperm. Theriogenology 76:1234–1245
Perez-Cerezales S, Martinez-Paramo S, Beirao J, Herraez MP (2010) Fertilization capacity with rainbow trout DNA-damaged sperm and embryo developmental success. Reproduction 139:989–997
Piehl LL, Fischman ML, Hellman U, Cisale H, Miranda PV (2013) Boar seminal plasma exosomes: effect on sperm function and protein identification by sequencing. Theriogenology 79:1071–1082
Quesenberry PJ, Aliotta J, Deregibus MC, Camussi G (2015) Role of extracellular RNA-carrying vesicles in cell differentiation and reprogramming. Stem Cell Res Ther 6:153
Raposo G, Stoorvogel W (2013) Extracellular vesicles: Exosomes, microvesicles, and friends. J Cell Biol 200:373–383
Regassa A, Rings F, Hoelker M, Cinar U, Tholen E, Looft C, Schellander K, Tesfaye D (2011) Transcriptome dynamics and molecular cross-talk between bovine oocyte and its companion cumulus cells. BMC Genomics 12:57
Reik W, Dean W, Walter J (2001) Epigenetic reprogramming in mammalian development. Science 293:1089–1093
Rizos D, Clemente M, Bermejo-Alvarez P, de La Fuente J, Lonergan P, Gutierrez-Adan A (2008) Consequences of in vitro culture conditions on embryo development and quality. Reprod Domest Anim 43(Suppl 4):44–50
Rizos D, Gutierrez-Adan A, Perez-Garnelo S, De La Fuente J, Boland MP, Lonergan P (2003) Bovine embryo culture in the presence or absence of serum: implications for blastocyst development, cryotolerance, and messenger RNA expression. Biol Reprod 68:236–243
Rizos D, Pintado B, de la Fuente J, Lonergan P, Gutiérrez-Adán A (2007) Development and pattern of mRNA relative abundance of bovine embryos cultured in the isolated mouse oviduct in organ culture. Mol Reprod Dev 74:716–723
Rizos D, Ward F, Duffy P, Boland MP, Lonergan P (2002) Consequences of bovine oocyte maturation, fertilization or early embryo development in vitro versus in vivo: implications for blastocyst yield and blastocyst quality. Mol Reprod Dev 61:234–248
Roberts RM, Ealy AD, Alexenko AP, Han CS, Ezashi T (1999) Trophoblast interferons. Placenta 20:259–264
Robinson RS, Hammond AJ, Wathes DC, Hunter MG, Mann GE (2008) Corpus luteum-endometrium-embryo interactions in the dairy cow: underlying mechanisms and clinical relevance. Reprod Domest Anim 43(Suppl 2):104–112
Roca J, Gil MA, Hernandez M, Parrilla I, Vazquez JM, Martinez EA (2004) Survival and fertility of boar spermatozoa after freeze-thawing in extender supplemented with butylated hydroxytoluene. J Androl 25:397–405
Roca J, Rodriguez MJ, Gil MA, Carvajal G, Garcia EM, Cuello C, Vazquez JM, Martinez EA (2005) Survival and in vitro fertility of boar spermatozoa frozen in the presence of superoxide dismutase and/or catalase. J Androl 26:15–24
Rottmayer R, Ulbrich SE, Kolle S, Prelle K, Neumueller C, Sinowatz F, Meyer HH, Wolf E, Hiendleder S (2006) A bovine oviduct epithelial cell suspension culture system suitable for studying embryo-maternal interactions: morphological and functional characterization. Reproduction 132:637–648
Russell DF, Baqir S, Bordignon J, Betts DH (2006) The impact of oocyte maturation media on early bovine embryonic development. Mol Reprod Dev 73:1255–1270
Saadeldin IM, Kim SJ, Choi YB, Lee BC (2014) Improvement of cloned embryos development by co-culturing with parthenotes: a possible role of exosomes/microvesicles for embryos paracrine communication. Cell Reprogram 16:223–234
Saadeldin IM, HJ O, Lee BC (2015) Embryonic-maternal cross-talk via exosomes: potential implications. Stem Cells Cloning 8:103–107
Sapanidou V, Taitzoglou I, Tsakmakidis I, Kourtzelis I, Fletouris D, Theodoridis A, Zervos I, Tsantarliotou M (2015) Antioxidant effect of crocin on bovine sperm quality and in vitro fertilization. Theriogenology 84:1273–1282
Senger P (2003) Pathways to pregnancy and parturition. Current conceptions, Inc., Pullman
Silva PF, Gadella BM, Colenbrander B, Roelen BA (2007) Exposure of bovine sperm to pro-oxidants impairs the developmental competence of the embryo after the first cleavage. Theriogenology 67:609–619
Silveira JC, Veeramachaneni DNR, Winger QA, Carnevale EM, Bouma GJ (2012) Cell-secreted vesicles in equine ovarian follicular fluid contain miRNAs and proteins: a possible new form of cell communication within the ovarian follicle. Biol Reprod 86:71
Simons M, Raposo G (2009) Exosomes – vesicular carriers for intercellular communication. Curr Opin Cell Biol 21:575–581
Simpson RJ, Jensen SS, Lim JWE (2008) Proteomic profiling of exosomes: current perspectives. Proteomics 8:4083–4099
Sirard MA (2001) Resumption of meiosis: mechanism involved in meiotic progression and its relation with developmental competence. Theriogenology 55:1241–1254
Sirard MA, Richard F, Blondin P, Robert C (2006) Contribution of the oocyte to embryo quality. Theriogenology 65:126–136
Somfai T, Ozawa M, Noguchi J, Kaneko H, Kuriani Karja NW, Farhudin M, Dinnyes A, Nagai T, Kikuchi K (2007) Developmental competence of in vitro-fertilized porcine oocytes after in vitro maturation and solid surface vitrification: effect of cryopreservation on oocyte antioxidative system and cell cycle stage. Cryobiology 55:115–126
Spencer TE, Bazer FW (2004) Conceptus signals for establishment and maintenance of pregnancy. Reprod Biol Endocrinol 2:49
Spencer TE, Forde N, Lonergan P (2016) The role of progesterone and conceptus-derived factors in uterine biology during early pregnancy in ruminants. J Dairy Sci 99:5941–5950
Suarez SS (2008) Regulation of sperm storage and movement in the mammalian oviduct. Int J Dev Biol 52:455–462
Suzuki J Jr, Therrien J, Filion F, Lefebvre R, Goff AK, Smith LC (2009) In vitro culture and somatic cell nuclear transfer affect imprinting of SNRPN gene in pre- and post-implantation stages of development in cattle. BMC Dev Biol 9:9
Takahashi M (2012) Oxidative stress and redox regulation on in vitro development of mammalian embryos. J Reprod Dev 58:1–9
Tarin JJ, Perez-Albala S, Cano A (2000) Consequences on offspring of abnormal function in ageing gametes. Hum Reprod Update 6:532–549
Teijeiro JM, Marini PE (2012) The effect of oviductal deleted in malignant brain tumor 1 over porcine sperm is mediated by a signal transduction pathway that involves pro-AKAP4 phosphorylation. Reproduction 143:773–785
Tervit HR, Whittingham DG, Rowson LE (1972) Successful culture in vitro of sheep and cattle ova. J Reprod Fertil 30:493–497
Tesfaye D, Ghanem N, Carter F, Fair T, Sirard MA, Hoelker M, Schellander K, Lonergan P (2009) Gene expression profile of cumulus cells derived from cumulus-oocyte complexes matured either in vivo or in vitro. Reprod Fertil Dev 21:451–461
Tesfaye D, Lonergan P, Hoelker M, Rings F, Nganvongpanit K, Havlicek V, Besenfelder U, Jennen D, Tholen E, Schellander K (2007) Suppression of connexin 43 and E-cadherin transcripts in in vitro derived bovine embryos following culture in vitro or in vivo in the homologous bovine oviduct. Mol Reprod Dev 74:978–988
Thibodeaux JK, Roussel JD, White KL, Broussard JR, Godke RA (1992) The use of image analysis to evaluate the development of uterine and oviduct epithelial cells during in vitro culture. A potential quality assurance procedure for in vitro laboratories. Arch Pathol Lab Med 116:444–448
Thompson JG (2000) In vitro culture and embryo metabolism of cattle and sheep embryos – a decade of achievement. Anim Reprod Sci 60-61:263–275
Ulbrich SE, Zitta K, Hiendleder S, Wolf E (2010) In vitro systems for intercepting early embryo-maternal cross-talk in the bovine oviduct. Theriogenology 73:802–816
Urrego R, Rodriguez-Osorio N, Niemann H (2014) Epigenetic disorders and altered gene expression after use of assisted reproductive technologies in domestic cattle. Epigenetics 9:803–815
Van Hoeck V, Leroy JL, Arias Alvarez M, Rizos D, Gutierrez-Adan A, Schnorbusch K, Bols PE, Leese HJ, Sturmey RG (2013) Oocyte developmental failure in response to elevated nonesterified fatty acid concentrations: mechanistic insights. Reproduction 145:33–44
Van Hoeck V, Sturmey RG, Bermejo-Alvarez P, Rizos D, Gutierrez-Adan A, Leese HJ, Bols PE, Leroy JL (2011) Elevated non-esterified fatty acid concentrations during bovine oocyte maturation compromise early embryo physiology. PLoS One 6:e23183
Walker SK, Hartwich K, Seamark R (1996) The production of unusually large offspring following embryo manipulation: concepts and challenges. Theriogenology 45:111–120
Walter I (1995) Culture of bovine oviduct epithelial cells (BOEC). Anat Rec 243:347–356
Wang H, Dey SK (2006) Roadmap to embryo implantation: clues from mouse models. Nat Rev Genet 7:185–199
Wang X, Falcone T, Attaran M, Goldberg JM, Agarwal A, Sharma RK (2002) Vitamin C and vitamin E supplementation reduce oxidative stress-induced embryo toxicity and improve the blastocyst development rate. Fertil Steril 78:1272–1277
Watson PF (2000) The causes of reduced fertility with cryopreserved semen. Anim Reprod Sci 60-61:481–492
Wheeler MB, Rutledge JJ, Fischer-Brown A, VanEtten T, Malusky S, Beebe DJ (2006) Application of sexed semen technology to in vitro embryo production in cattle. Theriogenology 65:219–227
Wilson RD, Fricke PM, Leibfried-Rutledge ML, Rutledge JJ, Penfield CM, Weigel KA (2006) In vitro production of bovine embryos using sex-sorted sperm. Theriogenology 65:1007–1015
Wrenzycki C, Herrmann D, Lucas-Hahn A, Korsawe K, Lemme E, Niemann H (2005) Messenger RNA expression patterns in bovine embryos derived from in vitro procedures and their implications for development. Reprod Fertil Dev 17:23–35
Wrenzycki C, Herrmann D, Niemann H (2007) Messenger RNA in oocytes and embryos in relation to embryo viability. Theriogenology 68(Suppl 1):S77–S83
Young LE, Fernandes K, McEvoy TG, Butterwith SC, Gutierrez CG, Carolan C, Broadbent PJ, Robinson JJ, Wilmut I, Sinclair KD (2001) Epigenetic change in IGF2R is associated with fetal overgrowth after sheep embryo culture. Nat Genet 27:153–154
Zullo G, De Canditiis C, Pero ME, Albero G, Salzano A, Neglia G, Campanile G, Gasparrini B (2016) Crocetin improves the quality of in vitro-produced bovine embryos: implications for blastocyst development, cryotolerance, and apoptosis. Theriogenology 86(8):1879–1885
Acknowledgements
Funded by the Spanish Ministry of Economy and Competitiveness AGL2015- 70140-R. P.L. was supported by funding from the European Union Seventh Framework Programme FP7/2007–2013 under grant agreement n° 312097 (“FECUND”).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Rizos, D., Maillo, V., Sánchez-Calabuig, MJ., Lonergan, P. (2017). The Consequences of Maternal-Embryonic Cross Talk During the Periconception Period on Subsequent Embryonic Development. In: Fazeli, A., Holt, W. (eds) Periconception in Physiology and Medicine. Advances in Experimental Medicine and Biology, vol 1014. Springer, Cham. https://doi.org/10.1007/978-3-319-62414-3_4
Download citation
DOI: https://doi.org/10.1007/978-3-319-62414-3_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-62412-9
Online ISBN: 978-3-319-62414-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)