
Abstract
Pure Mysore and Nistari are known oldest races of India. Both are multivoltine races which produce colored cocoons. The origin of these races is obscure. Pure Mysore was supposed to have originated from the race that Mysore King Tippu Sultan brought from China in 1875 and established in Karnataka State. Nistari is the well-known race also believed to have brought from China and established in West Bengal area. However, the wild sericigenous species of Bombyx, Theophila, and Ocinara are naturally distributed in the Himalayan ranges of Indo-China range and also in the Andaman Islands in India, besides Jawa, Sumatra, Borneo, and Malay Peninsula (Barlow, An introduction to the moths of South East Asia, 1982). Apart from these, there are wild relatives of silkworm, B. mandarina, which have been collected from Kedarnath. It is believed that the silkworm, Bombyx mori L., has evolved from B. mandarina in China and spread across the globe. There are many theories about the pattern of silkworm evolution and spread. In this article, the theories are discussed on the molecular basis.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


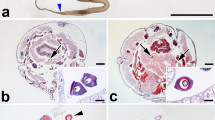
1 Introduction
Pure Mysore and Nistari are known oldest races of India. Both are Multivoltine races which produce colored cocoons. The origin of these races is obscure. Pure Mysore was supposed to have originated from the race that Mysore King Tippu Sultan brought from China in 1875 and established in Karnataka State. Nistari is the well-known race also believed to have brought from China and established in West Bengal area. However, the wild sericigenous species of Bombyx, Theophila, and Ocinara are naturally distributed in the Himalayan ranges of Indo-China range and also in Andaman Islands in India, besides Jawa, Sumatra, Borneo, and Malaya Peninsular (Barlow 1982). In addition to these species, wild relatives of silkworm, B. mandarina, has been collected from Kedarnath (Fig. 4.1). It is believed that the Silkworm, Bombyx mori L. has evolved from B. mandarina in China and spread across the globe. There are many theories about the pattern of silkworm evolution and spread. In this article, the theories are discussed on the molecular basis.

Geographical location of Kedarnath where Ancestor of Silkworm, Bombyx mandarina is also available in the wild
2 Voltinism
Voltinism is one of the important factors for discussing about the evolution and adaptation of the silkworm. The silkworm, Bombyx mori is classified based on the geographical regions, as Temperate and Tropical races. They are classified on the basis of voltinism as Univoltines, Bivoltines, and Multivoltines (or Polyvoltines); depending upon the number of generations, they undergo in a year. Generally, tropical silkworms are polyvoltines and the polyvoltine eggs do not undergo hibernation, which means that embryos develop continuously and hatch in about 10–11 days after egg laying. In univoltines and bivoltines, the embryos “pause” development and enter hibernation at blastoderm stage which is about 20–22 h after oviposition. Univoltines are temperate silkworm races, show facultative type of diapause wherein the eggs exhibit diapause phenomenon irrespective of environmental factors. Bivoltines are also temperate races which show obligatory type of diapause wherein diapause phenomenon is modified by environmental factors (Fig. 4.2).

Map showing region-wise voltinism in silkworm
Dominance of voltinism is recognized in the order of Univoltines > Bivoltine > Multivoltine. The multivoltine races always lay nonhibernating eggs. The breeds between multi- and bivoltine could lay some non-hibernating eggs under high incubation temperatures. The bivoltine strains lay non-hibernating eggs under low and hibernating eggs under high temperature of egg incubation. The breeds between bi- and univoltine could lay both non- and hibernating eggs, even under low incubation temperatures. The typical univoltine races always lay hibernating eggs (Morohoshi 2000).
Andrewartha (1952) opines that diapause is a convergent trait that has evolved independently several times in the course of evolution. Toyama (1906a, 1913) described diapause as a maternally inherited biological event under the control of sex-linked genes. Sonobe and Odake (1986) have proposed two theories related to the embryonic diapause in silkworm: (1) diapause is the phenomenon predetermined by the diapause factor during embryogenesis, and (2) diapause is the process determined by the genetic factor during embryogenesis. Mundkur et al. (2010a, b) have proposed a new theory related to diapause specially taking paternal influence into account. “diapause is the phenomenon determined by environment, maternal genes and paternal genes synergistically or exclusively” (Figs. 4.3, 4.4, and 4.5).

Bivoltine to multivoltine theory of evolution (Gamo and Ohtsuka 1980)

Multivoltine to Bivoltine theory of evolution (Yoshitake 1968)

KSSRDI model of silkworm diversification
Understanding these theories of silkworm diversification is important to silkworm breeders. Just like dominance of voltinism, the productivity and quality is also in the order of Univoltines > Bivoltine > Multivoltine. Tropical multivoltines are inferior in productivity. Indian breeders look towards sub-tropical bivoltines to improve the productive and qualitative traits. If the bivoltines of subtropics are introduced to Indian tropical conditions, they fail to behave as in their original environment. Over the period of time they lose their productive qualities and turn themselves into polyvoltines (Table 4.1).
3 Correlation Between Characters
In silkworms, it is observed that one character is influenced by another character generally contrasting one. We can measure their association by correlation studies. Correlations may be positive (i.e., when the quality and quantity of one character go up, those of other character also go up) or negative (i.e., when the quality and quantity of one character go up, those of other character come down). Two major causes of correlation between characters are genetic and environmental factors which result in correlation between the genotypic values of the two characters and the correlation between the environmental deviations (HoZoo 1997). The primary cause of genetic correlation is pleiotropic gene action. Pleiotropy refers to the gene’s attribute which affects more than one character, so that if one gene is segregating, it causes simultaneous variation in the character it affects. However, linkage is also a cause of genetic correlation, especially in populations derived from crosses between varieties with divergent genetic backgrounds. Such actions of a gene or interactions gene to gene as well as genotype to environment result in phenotypic, genetic, or environmental correlations between the characters of economic interest. The extent of correlation usually varies among silkworm breeds and between male and female due to differences in crossover rates which are higher in the male than in the female. For all polygenic traits, the guiding principle for any silkworm breeding scheme is information on correlation and linkage in the female, along with prior knowledge about heritability. Before beginning actual breeding experiments, it is important to know how improvement of one character will cause simultaneous change, either in a positive or negative direction, in other characters. In that aspect the ignoring of mutual genetic link between leading productive traits led to decrease in the genetic progress. Correlation between cocoon weight and pupal weight was high and positive (+0.994), between cocoon weight and shell weight was also positive but lower (+0.614), as well between pupal weight and cocoon shell weight (+0.527). Considerable negative correlation was found between pupal weight and shell ratio (−0.827) (Singh et al. 1992a, b). Fecundity correlates positively with pupal weight but negative with productivity, shell ratio, and strength of fiber. The shell ratio is a better trait for assessing the quality of cocoons for reeling because higher shell ratio led to higher silk yield, due to the proved positive correlation between shell ratio and quality of cocoons.
The phenotypic correlation not always corresponded to the genetic nature and accurately reflected the degree and character of inherited link between productive traits in Bombyx mori L. For example, this is the indicated negative phenotypic correlation between traits shell ratio and total egg weight in a batch. In that case negative phenotypic correlation informed that carrying the selection on shell ratio in positive direction will affect negatively on weight of eggs in batches. In the same time however positive genetic correlations show possibilities for differentiation of lines that combine as high shell ratio, as relatively high reproductive ability of silkworm (Grekov and Petkov 1990). The fulcrum relationship of resistance and productivity is of great concern to the breeders. Available data indirectly indicate that they are linked characters, though we don’t have the direct genetic proof of it. Various investigators have correlated silkworm characters. Table 4.2 summarizes the inference. Nagaraju (1998) has tabulated (Table 4.3) the correlation of various characters in silkworm based on 15 years of breeding data. It is generalized that, when the productivity goes up, the survivability comes down (Fig. 4.6), but neither Table 4.2 nor Table 4.3 substantiates that concept.

Seesaw balance of quality versus resistance. Yield attributes are linked to the biochemical parameters (Chatterjee et al. 1993)
4 Survival of the Fit
Fitness involves the ability of organisms (or populations or species) to survive and reproduce in the environment in which they find themselves (Allen Orr 2009). Natural selection is the process by which the best adaptations survive long enough to reproduce. Natural selection is the process by which the traits that are useful for the survival will continue on to the next generation and what does not work will be eliminated. The most important factor influencing the fitness is the environment. Let us take the example of Pure Mysore, with only two parameters, survival and cocoon weight. About 25 years ago, during the 1990s, the average cocoon weight of Pure Mysore cocoon was 1.00 g, and the survival (ERR) was 95%. Today, with improved inputs, the selection could be made up to 1.4 g cocoon weight with the same ERR. However, there is a limit. We cannot match the productivity of temperate bivoltines in temperate conditions (2.0 g cocoon weight with 95% ERR) in Pure Mysore. Let us relocate these two breeds, Pure Mysore to temperate congenial environment and temperate bivoltine to the tropical unfavorable environment. Pure Mysore in temperate conditions will gradually turn into diapausing type and show little higher cocoon weight (1.6 g) than in tropical conditions. It will never match the temperate bivoltine in productivity even if it is placed in the same environment as of temperate bivoltine breed. It looks like it simply does not have the genetic machinery for producing higher cocoon weight. The relocated bivoltine in tropical conditions lose its productivity to retain its survivability (as in case of C-Nichi). The net result is that there is no improvement in survivability, but the productivity suffered. Therefore, the environment plays a major role in deciding the quality and productivity parameters.
5 Behavior of Temperate Silkworm Races in Indian Tropical Conditions
In Japan, silkworm eggs hatch after completion of hibernation during spring (about middle of May), in warmer climate with about 20°C temperature. They turn into moths around later half of June and lay eggs. The deposited eggs are nonpigmented pale yellow in color and nonhibernating. They hatch and grow during early July, with the temperature of 25–30°C. They become moths during later half of August and deposit eggs. The laid eggs are brown colored and hibernating (Tazima 1986) (Fig. 4.7).

Behavior of bivoltines in temperate regions. They show two successive crops in spring and following summer. Spring crop lays non-diapausing eggs, while summer crop lays diapausing eggs
The freshly laid diapausing eggs also do not have pigments, just like nonhibernating eggs, but after 10–20 days in Japan, eggs become pigmented and turn to purple black. The pigmented eggs stop their development at the age of blastoderm formation and become dormant. Eggs pass through summer, and once they encounter cold weather of winter for 3–4 months, they continue their development. Hence, “bivoltine” in Japan is alternation of diapausing and non-diapausing generations (Mundkur et al. 2004).
Extensive studies on diapause mechanism are undertaken in temperate conditions, but under tropical conditions, the studies are scanty. In Indian tropical conditions, bivoltines do not behave as they behave in Japanese conditions. They undergo hibernation in every generation. During summer season, sporadic incidences of bivoltines laying non-diapausing eggs are recorded. Conversely, during winter season, sporadic incidences of polyvoltine silkworm races laying diapausing eggs are also recorded. Generally, polyvoltine females lay nondiapausing eggs irrespective of whether they pair with polyvoltine or bivoltine males. Similarly, bivoltine females lay diapausing eggs irrespective of whether they pair with bivoltine or polyvoltine males. Therefore, diapause is a maternally inherited character, and males do not have any role in determining the voltinism of the offspring. However, an exception to this well-established phenomenon was recorded where bivoltine females (which are destined to lay hibernating eggs) lay nonhibernating eggs when they are mated with a special character race, KS-10 males (Mundkur et al. 2004, 2006, 2009, 2010b, c, d). This kind of paternal influence on diapause has opened up many views on the diapause phenomenon in silkworm Bombyx mori, from point of view of its expression under tropical conditions.
6 Difference Between Diapause and Non-diapause Eggs
Generally there are many differences between diapause and non-diapause type of silkworm eggs (Table 4.4).
7 Tropicalization of Temperate Races
A prominent race which supported sericulture industry in South India was C-Nichi. It was evolved from a Chinese × Japanese hybrid (C for Chinese × N for Nippon which means Japan and ichi means one). It lost its hibernating character and much of its productive traits and has become white, highly robust, low-productive multivoltine race. Likewise an exotic univoltine race got tropicalized and became Boropolu and sustained in West Bengal area. Several attempts were made to import and establish exotic bivoltine or univoltine races, especially from Italy, France, Russia, Japan, China, and Iran. Only a few of these races survived under Indian conditions, especially in Kashmir area, where the climatic conditions are suitable for their survival. In Southern India, which has typical tropical conditions, these bivoltines could not survive for long. Till today Pure Mysore rules the sericulture industry. Likewise in West Bengal, Nistari became the predominant silkworm race.
8 Heritability
Most of economic characters are of quantitative type, and each is expressed by cumulative action and interaction of many genes in either an additive or nonadditive mode (HoZoo 1997). Resemblance between relatives and inbreeding depression are two basic genetic phenomena associated with quantitative characters. Resemblance through genetic causes only is defined as heritability. The reverse effect of inbreeding depression is called heterosis or hybrid vigor. In general, the closer the relationship, the closer the resemblance. Selective breeding is based on the resemblance between offspring and parents. The average level of a certain character in the next generation will be improved when the more desirable individuals are used as parents. The extent of resemblance varies with the character. Some characters which show more resemblance are more responsive to selection than others. One characteristic of a population is the degree of resemblance between relatives. A quantitative genetic approach such as heritability estimation will show how the degree of resemblance between different sorts of relatives can be used to predict the outcome of selective breeding. It can also lead a breeder to the best method of selection. Heritability expressed in quantitative terms is the proportion of total variance due to the average effects of genes and also determines the degree of resemblance between relatives. Its predictive role is of important use to breeders, because heritability expresses the reliability of the revealed phenotypic values as a guide to the concealed heritable breeding value of additive gene action or the degree of correspondence between phenotypic value and breeding value. If a breeder chooses parental silkworms according to their phenotypic values, improvement of characters in the subsequent generation can be predicted only by knowing the extent of correspondence between phenotypic values and breeding values. Therefore, the magnitude of heritability is very important as breeders select breeding methods and determine breeding procedures. A distinction is usually made between heritability in “broad” and “narrow” sense, reflecting the components of variation (variance in statistical terms) included in their estimation. Broad sense heritability can be described as the ratio between the genetic variance and phenotypic variance. Narrow-sense heritability is a more meaningful term and thus used almost exclusively by silkworm breeders. It is defined as the ratio of additive genetic variance to total phenotypic variance.
Heritability is a property of a certain character as well as of the population and of the environment to which the individual silkworms are subjected. The value of heritability varies depending on the magnitude of all the components of variance. A change in any one of these components will therefore influence heritability estimates. All the genetic components are influenced by gene frequencies, which may be different from one population to another, according to their history. Large populations will show higher heritability than small populations, in which a significant amount of fixation has occurred for some time. More variable environmental rearing conditions are expected to reduce heritability, while more uniform conditions will increase it. Whenever a value is stated for the heritability of a given character, it refers to a particular population under particular conditions. A considerable range of variation is thus commonly shown among different estimates of heritability for the same character, although it may be partly due to statistical sampling. The greater the heritability value, in general, the smaller the variation of the character because of environmental differences.
The relatively high values of h that were found out for some leading selection traits such as cocoon shell weight, filament length, etc., are informative for the big genetic variation in populations of Bombyx mori L. and for the perspectiveness of phenotype selection. In the same time, however, the relatively low values of h for the trait number of eggs in one batch determine the considerably trait magnitude by the environmental conditions (Singh et al. 1992b). Traits with low h can be improved by the inclusion of other closely correlated traits with relatively high values of h in the breeding programs. Relatively higher values of heritability were found for cocoon weight (73.60%), pupal weight (78.50%), shell weight (80.20%), growth rate (79.30%), raw silk yield (79.00%), and shell ratio (72.40%) (Chatterjee and Datta 1992; Singh and Singh 1993; Singh 1994; Singh et al. 1994; Yan 1983; Rajanna and Reddy 1990a, b). Middle values for larval period duration were found, while low heritability was detected for the traits pupation rate (28.00%) and reelability (19.00%) (Gamo and Hirabayashi 1983) (Table 4.5).
9 Hybrid Vigor and Environment
Hybrid vigor refers to positive heterosis. It is the extra vigor or improvement in performance shown by the F1 progeny over either of its parents or mid-parent value. Hybrid vigor is the manifestation of interaction between the genes or alleles of two pure lines. However the phenotypic expression is always dependent on the interaction between genotype and environment. Even if the organism possesses a very good genetic makeup, the phenotype is the result of its interaction with the environment. Thus, for optimum expression of the genotype, congenial environment is necessary.
The phenotypic expression is also dependent upon the genetic plasticity or buffering capacity of the organism. The extra vigor shown by the outcross between two inbred lines of silkworm could be due to the extra buffering capacity of the resultant organism.
10 Theories on Breeding
There are many theories of breeding methodology to make the breed best “fit” to the given environment.
10.1 Ideal Environment Breeding
Falconer (1952) advocates that “the character required is best selected for under environmental conditions which favour its fullest expression. Once developed, other characters specially required for new environment also will be present in such animals.” Many silkworm breeders subscribe to this theory (Toyama 1906a, b, 1913).
10.2 Native Environment Breeding
The high-yielding silkworm breeds developed through inbreeding of hybrids by selection for various attributes under laboratory conditions (Krishnaswami 1978). The silkworm races thus developed are utilized to obtain polyvoltine × bivoltine crossbreeds or bivoltine × bivoltine hybrids for commercial usage. The farmers often fail to produce the optimum conditions required by these combinations. As a result, only 40% of the yield attributes are expressed in the field (Nagaraju 1998). Falconer (1952, 1960, 1981) advocates “performance is best improved by selection under the conditions in which the performance is subsequently measures.” He explains that quantitative traits or metric traits show continuous variation because of P = G × E effect. Therefore, the segregation of genes cannot be followed individually (Falconer 1981). Many tropical breeders subscribe to this theory (Rajanna and Reddy 1990a, b, 1998).
10.3 Shuttle Breeding
High-yielding breeds bred in laboratory have the limitations of limited population rearing. Chatterjee (1993) supported the breeding plan of taking breeding materials during F10 to the different Regional Research Stations so that they are exposed to various climates. Then they are brought back to the main breeding laboratory for applying appropriate selection pressure.
10.4 Voltinism Breeding
Discovery of pnd and npnd genes and their behavior in tropical environment lead to this special branch of breeding (Chatterjee 1993; Murakami 1986, 1988, 1989; Murakami and Ohtsuki 1989; Subramanya and Murakami 1994). Murakami (1986) showed that npnd (nonpigmented non-diapausing) is a sex-linked gene and responsible for multivoltinism exhibiting maternal inheritance. He also revealed that npnd is epistatic to pnd (pigmented non-diapausing) which was described by Katsumata (1968).
10.5 Avoltinism Breeding
The tropical bivoltine and multivoltine race exhibit distinct economic advantages. Survival is generally attributed to multivoltines and productivity to bivoltines. Voltinism remains a barrier and bottleneck in balancing and harvesting these two critical attributes. Discovery of new dominant voltinism gene “Id” has become a new tool in silkworm breeding. The presence of Id gene overcomes the effect of voltinism. It can be transduced to the desired race thereby making it a “voltine-less” race. A new terminology is coined for such character, “avoltinism,” which means voltinism-less or “without voltinism.” This tool holds the promise for breaking the inverse relationship and balancing the polyvoltine survival traits with bivoltine quantitative and qualitative traits (Mundkur et al. 2004, 2010a, b, c, d, 2011, 2012).
10.6 Biotechnology, Genomics, and Marker-Assisted Selection Breeding
Review of progress in applying molecular genetic and genomic technologies to studies in the domesticated silkworm, Bombyx mori, highlighting its use as a model for Lepidoptera, and in sericulture and biotechnology has been done by many scientists (Nagaraju 1999; Nagaraju et al. 2001; Goldsmith et al. 2005). Dense molecular linkage maps are being integrated with classical linkage maps for positional cloning and marker-assisted selection. Classical mutations have been identified by a candidate gene approach. Cytogenetic and sequence analyses show that the W chromosome is composed largely of nested full-length long terminal repeat retrotransposons. Z-chromosome-linked sequences show a lack of dosage compensation. The downstream sex differentiation mechanism has been studied via the silkworm homologue of double-sex. Expressed sequence tagged databases have been used to discover Lepidoptera-specific genes, provide evidence for horizontal gene transfer, and construct microarrays. Physical maps using large-fragment bacterial artificial chromosome libraries have been constructed, and whole-genome shotgun sequencing is underway. Germline transformation and transient expression systems are well established and available for functional studies, high-level protein expression, and gene silencing via RNA interference.
11 Understanding Tolerance and Resistance in Silkworm Breeds
Resistance is an absolute term where the individual completely immunizes itself to a particular stress. This kind of situation is observed in case of biotrophic pathogen (pathogen needing a live host to draw nutrient) infection when the host contains a resistance gene and the infecting pathogen contains its corresponding a virulence gene. Resistance can be summarized as the ability of the organism to limit pathogen growth and infection.
Tolerance is rather a relative term, and it is also man made to some extent. Tolerance does not limit infection but instead reduces or offsets its negative fitness consequences. Generally biotic stress challenges the resistance of the individual, and abiotic stress challenges the tolerance and resistance, and tolerance may provide equivalent short-term benefits but have fundamentally different epidemiological consequences and thus exhibit different evolutionary behaviors (Felix Horns and Hood Michael 2012).
The defense reaction in silkworm against bacteria is the production of antibacterial protein in the hemolymph. The defense reaction in silkworm against nuclear polyhedrosis virus (NPV) includes (1) antiviral substance in the gut juice of the silkworm (Aizawa 1962, 1991) and (2) viral inhibitory factors (VIF) produced in the silkworm hemolymph (Hayashita et al. 1968). Red fluorescent protein (RFP) also shows profound antiviral activity.
Conclusion
Silkworm breeders of Indian tropical conditions are trying very best for the substantial improvement in the cocoon yield, silk recovery, and quality in the silkworm breeds they develop, through the integration of all the available approaches which include conventional as well as molecular biology tools. Foregoing discussions clearly indicate that the environment is the critical limiting factor, which include biotic and abiotic factors, which determine the final performance of the developed breeds. Southern zone of India is characterized by semiarid areas by tropical conditions, often experiencing deficit rainfall and high temperature, with higher biotic as well as abiotic stress. Natural selection for this area is the polyvoltine breed with moderate production with higher survival rates. Balanced increase in productivity, quality, and survivability is possible by considering all the relevant factors in breeding.
References
Aizawa K (1962) Antiviral substance in the gut juice of the silkworm, Bombyx mori (L). J Insect Pathol 4:72–76
Aizawa K (1991) Defense reactions of the silkworm, Bombyx mori (L), against the nuclear polyhedrosis. Sericologia 31:201–203
Allen Orr A (2009) Fitness and its role in evolutionary genetics. Nat Rev Genet 10(8):531–539
Andrewartha MG (1952) Diapause in relation to the ecology of insects. Bio Rev 27:50–107
Barlow HS (1982) An introduction to the moths of South East Asia. Malayan Nature Society, Kaulalumpur, pp 1–305
Chatterjee SN (1993) Silkworm breeding in India. Sericologia 33(3):427–447
Chatterjee SN, Datta RK (1992) Hierarchical clustering of 54 races and strains of the mulberry silkworm, Bombyx mori L: significance of biochemical parameters. Theor Appl Genet 85(4):394–402
Chatterjee SN, Rao CGP, Chatterjee GK, Ashwath SK, Patnaik AK (1993) Correlation between yield and biochemical parameters in the mulberry silkworm, Bombyx mori L. Theor Appl Genet 87(3):385–391
Falconer DS (1952) The problem of environment and selection. Am Nat 86:293–298
Falconer DS (1960) Introduction to quantitative genetics. Oliver & Boyd, Edinburgh
Falconer DS (1981) Introduction to quantitative genetics, 2nd edn. Longman, London
Felix Horns F, Hood Michael E (2012) The evolution of disease resistance and tolerance in spatially structured populations. Ecol Evol 2(7):1705–1711
Gamo T, Hirabayashi T (1983) Genetic analysis of growth rate, pupation rate and some quantitative characters by diallel cross in the silkworm, Bombyx mori. Jpn J Breed 33:178–190
Gamo T, Ohtsuka Y (1980) Phylogenetic studies on the racial differences of the silkworm, Bombyx mori, on the basis of polymorphic genes in haemolymph proteins. Bull Sericult Exp Sta 28:15–50
Goldsmith MR, Shimada T, Abe H (2005) The genetics and genomics of the silkworm, Bombyx mori. Annu Rev Entomol 50:71–100
Gowda BLV, Narayanaswamy TK, Munirajappa R (1988) Impact of pupal weight on growth and development of the following generation in the silkworm Indian race NB7 (Bombyx mori). Sericologia 29:481–489
Gowda BLV, Sannaveerappanayar VT, Shivayogeshwar B (1989) Fecundity and hatchability in mulberry silkworm, Bombyx mori (L.) as influenced by pupal weight. International congress on tropical sericultural practices, part VI, Bangalore, 18–23 February 1989, pp 21–24
Grekov D, Petkov N (1990) Correlation between some traits and possibilities for prognostication heterosis of F1 crosses on hybridization of Bombyx mori L. Zhivotnov’s Nauki 27:80
Hayashita K, Nishida J, Matsubara F (1968) Inactivation of nuclear polyhedrosis virus in the digestive juice of silkworm, Bombyx mori (L). I. Comparison of antiviral activities in the digestive juice of larvae reared on natural and artificial diets. Jpn J Appl Entomol Zool 12:189–193
HoZoo L (1997) Principles and techniques of silkworm breeding. ESCAP, United Nations, New York
Jayaswal KP, Singh T, Sen SK (1990) Correlation between some economic parameters and their application in silkworm breeding. Indian Silk 29:25–27
Jayaswal KP, Singh T, Subba RG (1991) Effect of female pupal weight on fecundity of mulberry silkworm Bombyx mori. Indian J Seric 30:141–143
Jonaka N (1986) Some parameters of basic quantitative breeding characters in silkworm, Bombyx mori. II. Correlations between the basic breeding characters. Genet Sel 19:144–150
Katsumata F (1968) Non maternal inheritance in Voltinism observed in the crossing experiments between Indonesian polyvoltine and Japanese bivoltine races of silkworm, Bombyx mori. J Seric Sci Jpn 37:453–461
Krishnaswami S (1978) New technology of silkworm rearing. Bulletin No. 2. Central Sericultural Research and Training Institute, Mysore, Central Silk Board, India, pp 1–23
Liu Q-X, He S-M (1991) Studies on the efficiency of selection for improving technological characters of cocoon filament of Bombyx mori L. Canye Kaxue 17:75–79
Long NV, Petkov N (1987) Breeding—genetic studies in some silkworm (Bombyx mori) breeds. I. Variability and correlations of quantitative characters. Genet Sel 20:58–62
Miyahara T (1978) Selection of long filament length basic variety. I. Effect of selection in the later generations. Acta Sericologia Sinica 106:73–78
Morohoshi S (2000) Development physiology of silkworms. In: Translation of second Japanese edition of Kaiko no Hatsuiku Seiri by Morohoshi S, (1979). Oxford and IBH Pub.Co.Pvt.Ltd, New Delhi
Mundkur R, Murthy M, Raghuraman MR, Bongale UD (2004) Prevention of diapause in bivoltine eggs of the silkworm, Bombyx mori L., through a cross with the race KS-10 as male parent. Int J Ind Entomol 9(1):107–109
Mundkur R, Murthy M, Rao SK, Raghuraman R, Bongale UD (2006) Paternal inheritance of voltinism: a new tool to understand the gene regulation in the silkworm, Bombyx mori. In: Proceedings of international symposium on insect genetics and genome, CDFD, Hyderabad, 9–11 Jan 2006
Mundkur R, Murthy M, Latha R, Krishna Rao S, Sekharappa BM (2009) Indication of expression of inhibitor of diapauses gene in silkworm, Bombyx mori. In: National conference on recent trends in animal physiology, Manasa Gangotri, Mysore, 29–30 Oct 2009
Mundkur R, Katti SR, Chandrakala MV, Sekharappa BM (2010a) Diapause in silkworm, Bombyx mori L with special reference to the paternal influence: a review. Sericologia 50(2):145–170
Mundkur R, Murthy M, Latha R, Krishna Rao S, Sekharappa BM (2010b) Avoltinism breeding: a new concept in silkworm breeding. In: Silkworm breeder’s meet, RSRS Coonoor, 2–3 Dec 2010
Mundkur R, Murthy M, Latha R, Krishna R, Sekharappa BM (2010c) “Id” inhibitor of diapauses, a new gene as diapauses switch in silkworm, Bombyx mori. In: Nattional seminar on emerging trends in animal biotchnology, Kakatiya University, Warangal, 26–27 Mar 2010
Mundkur R, Murthy M, Latha R, Rajanna GS, Krishna Rao S (2010d) Reporting of a new gene inhibitor of diapause(Id) in the silkworm, Bombyx mori L. Sericologia 50(3):359–364
Mundkur R, Murthy M, Krishna Rao S (2011) Modification of diapause induced by inhibitor of diapause gene (Id), after fertilization in silkworm, Bombyx mori. In: International conference on life science research for rural and agricultural development, CPRS, Patna, Bihar, 27–29 Dec 2011
Mundkur R, Murthy M, Krishna Rao S (2012) Termination of diapause by Inhibitor of diapause gene (Id) after fertilization in silkworm, Bombyx mori. In: National seminar on advances in zoology and life processes, Goa University, 9–11 Feb 2012
Murakami A (1986) Changes in embryonic diapause in the tropical multivoltine silkworm stock Cambodge. In: Abstract: 34th Annual meetings of the Tokyo district Sericul. Sci Soc 27
Murakami A (1988) Ecogenetical studies on tropical mulberry Bombyx mori. In: Proceedings of the international congress on tropical sericulture practices, Bangalore, India, pp 11–23
Murakami A (1989) Genetic studies on tropical races of silkworm (Bombyx mori) with special reference to cross breeding strategy between tropical and temperate races. 2. Multivoltine silkworm strains in Japan and their origin. JARQ 23:123–127
Murakami A, Ohtsuki Y (1989) Genetic studies on tropical races of silkworm, Bombyx mori, with special reference to cross breeding strategy between tropical and temperate races. 1. Genetic nature of the tropical multivoltine strain, Cambodge. JARQ 23:37–45
Nagaraju J (1998) Silk yield attributes and complexities. In: Reddy GS (ed) Silkworm breeding. Oxford Press, New Delhi, pp 168–185
Nagaraju J (1999) Biotechnology: a novel concept for silkworm improvement. In: Devaiah MC, Narayanaswamy KC, Maribashetty VG (eds) Advances in mulberry sericulture. C.V.G. Publications, Bangalore, pp 208–242
Nagaraju J, Klimenko V, Couble P (2001) The silkworm, Bombyx mori: a model genetic system. In: Reeve ECR (ed) Encyclopaedia of genetics. Fitzroy Dearborn Publishers, London, pp 219–239
Ozdzenska B, Kremky J (1987) Estimation of heritability and genotypic, phenotypic and environmental correlations in out-bred populations of mulberry silkworm, Bombyx mori L. Sericologia 27:633–638
Petkov N (1981a) The possibility of forecasting the effectiveness of selecting lines of the silkworm, Bombyx Mori based on weight and silk content of cocoons. Selskotopanska Nauka 19:92–96
Petkov N (1981b) Variability and correlations between some characteristic features of silkworm (Bombyx mori L). Zhivotnov’d Nauki 18:83–86
Petkov N (1981c) Phenotypic correlations and regressions between some silkworm (Bombyx mori) breeding characters. Genet Sel 14:386–390
Rajanna GS, Reddy GS (1990a) Studies on the variability and interrelationship between some quantitative characters in different breeds of silkworm, Bombyx mori L. Sericologia 30:67–74
Rajanna GS, Reddy GS (1990b) New dimensions in tropical silkworm race breeding. Indian Silk 28:8–12
Rajanna GS, Reddy GS (1998) New approaches to silkworm improvement aimed at narrowing yield gaps. In: Reddy GS (ed) Silkworm breeding. Oxford Press, New Delhi, pp 168–185
Satenahalli SB, Govindan R, Goud JV, Magdum SB (1990) Genetic parameters and correlation coefficient analysis in silkworm, Bombyx mori. Mysore J Agric Sci 24:491–495
Singh T (1994) Correlation between pupal weight and fecundity in Bombyx mori (L). Ann Entomol 12:5–7
Singh T, Singh K (1993) Heritability and correlations between some economic characters in Bombyx mori (L). Sci Cult 59:51–52
Singh T, Jayaswal KP, Subba RG (1992a) Correlation studies between some breeding parameters of silkworm, Bombyx mori (L). J Zool Res 5:47–50
Singh T, Singh K, Das M (1992b) Astounding effect of correlated parameters in silk industry. Indian Text J 102:26–30
Singh T, Jayaswal KP, Subba RG (1992c) Correlation studies between some breeding parameters of silkworm, Bombyx mori (L). J Zool Res 5:47–50
Singh T, Chandrashekharaiah A, Samson MV (1994) Selection strategies in relation to correlation and heritability in the silkworm, Bombyx mori L. Bull Seric Res 5:37–41
Singh T, Bhat MM, Khan MA (2011) Critical analysis of correlation and heritability phenomenon in the silkworm, Bombyx mori (Lepidoptera: bombycidae). Adv Biosci Biotechnol 2(5):347–353
Sonobe H, Odake H (1986) Studies on embryonic diapause in the pnd mutant of the silkworm, Bombyx mori; V. Identification of a pnd+ gene-specific protein. Rouxs Arch Dev Biol 195:229–235
Subramanya G, Murakami A (1994) Climatic differential phenotypic expression of voltine genes in Bombyx mori L. Indian J Seric 33(2):103–109
Tazima Y (1986) The genetics of the silkworm. Logos Press, Academic Press, London
Toyama K (1906a) Mendel’s law of heredity as applied to the silkworm crosses. Biol Zentralbl 26:321–334
Toyama K (1906b) Breeding methods of silkworm. Sangyo Shimpo 158:282–286
Toyama K (1913) Maternal inheritance and mendelism. J Genet 2:351–405
Yan LL (1983) The estimates of heritability of pupal weight, cocoon weight and number of eggs laid in the silkworm (Bombyx mori) and its genetic correlation between these characters and path coefficient analysis. J Seric Sci China 9:149–155
Yoshitake N (1968) Phylogenetic aspects on the origin of Japanese race of the ilkworm, Bombyx mori L. J Sericol Sci Jpn 37:83–87
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Muniraju, E., Mundkur, R. (2018). Tracing of Evolution in Silkworm, Bombyx mori L., on the Basis of Molecular Studies. In: Kumar, D., Gong, C. (eds) Trends in Insect Molecular Biology and Biotechnology. Springer, Cham. https://doi.org/10.1007/978-3-319-61343-7_4
Download citation
DOI: https://doi.org/10.1007/978-3-319-61343-7_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-61342-0
Online ISBN: 978-3-319-61343-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)