
Abstract
Metals are the oldest toxins known to humans. Metals differ from other toxic substances in that they are neither created nor destroyed by humans (Casarett and Doull’s, Toxicology: the basic science of poisons, 8th edn. McGraw-Hill, London, 2013). Metals are of great importance in our daily life and their frequent use makes their omnipresence and a constant source of human exposure. Metals such as arsenic [As], lead [Pb], mercury [Hg], aluminum [Al] and cadmium [Cd] do not have any specific role in an organism and can be toxic even at low levels. The Substance Priority List of Agency for Toxic Substances and Disease Registry (ATSDR) ranked substances based on a combination of their frequency, toxicity, and potential for human exposure. In this list, As, Pb, Hg, and Cd occupy the first, second, third, and seventh positions, respectively (ATSDR, Priority list of hazardous substances. U.S. Department of Health and Human Services, Public Health Service, Atlanta, 2016). Besides existing individually, these metals are also (or mainly) found as mixtures in various parts of the ecosystem (Cobbina SJ, Chen Y, Zhou Z, Wub X, Feng W, Wang W, Mao G, Xu H, Zhang Z, Wua X, Yang L, Chemosphere 132:79–86, 2015). Interactions among components of a mixture may change toxicokinetics and toxicodynamics (Spurgeon DJ, Jones OAH, Dorne J-L, Svendsen C, Swain S, Stürzenbaum SR, Sci Total Environ 408:3725–3734, 2010) and may result in greater (synergistic) toxicity (Lister LJ, Svendsen C, Wright J, Hooper HL, Spurgeon DJ, Environ Int 37:663–670, 2011). This is particularly worrisome when the components of the mixture individually attack the same organs. On the other hand, metals such as manganese [Mn], iron [Fe], copper [Cu], and zinc [Zn] are essential metals, and their presence in the body below or above homeostatic levels can also lead to disease states (Annangi B, Bonassi S, Marcos R, Hernández A, Mutat Res 770(Pt A):140–161, 2016). Pb, As, Cd, and Hg can induce Fe, Cu, and Zn dyshomeostasis, potentially triggering neurodegenerative disorders, such as Alzheimer’s disease (AD) and Parkinson’s disease (PD). Additionally, changes in heme synthesis have been associated with neurodegeneration, supported by evidence that a decline in heme levels might explain the age-associated loss of Fe homeostasis (Atamna H, Killile DK, Killile NB, Ames BN, Proc Natl Acad Sci U S A 99(23):14807–14812, 2002).
The sources, disposition, transport to the brain, mechanisms of toxicity, and effects in the central nervous system (CNS) and in the hematopoietic system of each one of these metals will be described. More detailed information on Pb, Mn, Al, Hg, Cu, and Zn is available in other chapters. A major focus of the chapter will be on Pb toxicity and its interaction with other metals.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others
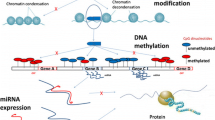

Keywords
Metals in the Environment
Heavy metal exposure can occur through contaminated air, food, and water or in hazardous occupations. While in the developed world the levels of heavy metal contamination in the environment have decreased in recent decades, developing countries in Asia and Africa continue to experience high levels of metal pollution, in particular in urban environments.
This contamination largely derives from anthropogenic sources, such as the combustion of leaded gasoline or unregulated industrial emissions. There is also a significant problem with metal contamination from mining, which results in elevated metal levels in water and air. Another major and relatively new source of metal contamination in developing countries is the practice of electronic waste recycling. Unfortunately, primitive and unsafe methods are used for the extraction of the precious metals, resulting in contamination of the local environment of highly toxic metals such as Hg and Pb (Neal and Guilarte 2013).
Environmental Exposure to Metal Mixtures
Along with actual apprehensions pertaining to human exposure to metals, it is well recognized that environmental exposures are not observed to single chemicals. The truth is that exposure to mixtures is the environmental reality in the present chemically sophisticated world (Simmons 1995). Mixtures of metals naturally occur, but metals are also often introduced into the environment as mixtures (Fairbrother et al. 2007). These mixtures are ubiquitous in air, water, and soil (Simmons 1995), and thus, people are exposed to them either concurrently or sequentially, by various routes of exposure and from a variety of sources to a large numbers of toxicants at low doses that may result in similar or dissimilar effects over exposure periods that can range from short term to a lifetime (ATSDR 2000; Pohl et al. 1997). Accordingly, the US Environmental Protection Agency (EPA) (ATSDR 2000) recommends greater efforts on understanding the combined toxic effects of metals (Fairbrother et al. 2007). Thus far the studies have largely focused on exposures to single metals (Kortenkamp and Faust 2009; Pohl et al. 1997), with few addressing chronic exposures to low levels of metal mixtures (Feron et al. 1995).
Fortunately, studies on mixtures have accelerated, incorporating more knowledge of specific modes of toxicological action and greater use of statistical methods and mathematical models (ATSDR 2000). Even so, predicting the health consequences of multiple chemical exposures is still a challenge (Pohl et al. 1997), because their study incorporates the understanding of interactions at several levels. These interactions may change toxicokinetics and toxicodynamics (Spurgeon et al. 2010). Therefore mixtures can influence adverse health effects sometimes, resulting in greater (synergistic) toxicity (Lister et al. 2011); this is particularly worrisome when the components of the mixture individually attack the same organs or, combined, overwhelm a particular mechanism that the organism uses to defend itself against toxic substances. Low doses that might not individually cause health effects, in concert, may become a public health issue (Calderon et al. 2003). It is reported that exposure to metal mixtures at concentrations below environmental quality guideline levels for individual components resulted in adverse effects that were attributed to interactions among the constituents (Yen Le et al. 2013). This issue was recently recognized by the EPA as a key gap in metal risk assessment (Abboud and Wilkinson 2013); thus a need exists for the research into the toxicity of metals, very especially with regard to metal mixtures in trace levels (Kim et al. 2009).
Criteria to Select Metal Mixtures
Given the almost infinite number of chemical mixtures, regulators are faced with a problem as to which chemicals should be chosen for assessment and regulation (Kortenkamp and Faust 2009), being important to prioritize them for research efforts (ATSRD 2004). Components produced and emitted together from industrial processes or present together in the same environmental or human body compartment are certainly to be considered. Chemicals thought to exhibit their effects through common mechanisms have been often grouped together based on similarities in chemical structure or derived from mechanistic considerations. Recently it has been argued that grouping criteria should focus on common adverse outcomes, with less emphasis on similarity of mechanisms (Kortenkamp and Faust 2009) and on chemicals having great potential impact on human health (ATSDR 2004). Reproductive, carcinogenic, and neurotoxic effects are considered potentially important health endpoints in epidemiological studies of complex mixtures, particularly when such mixtures contain trace metals (Shy 1993). Exposure to neurotoxic agents represents indeed a concern of high priority in modern society, given the ever increasing reported frequency of neurological diseases (Lucchini and Zimmerman 2009), with special concern to the induced long-term effects (Emerit et al. 2004).
Metals present unique environmental and public health issues, since these elements possess several particularities that should be taken into account when accessing the risks of their exposure; this includes the transformation into species with different valence states and the conversion between inorganic and organic forms. All these forms may possess different behaviors in the organism such as absorption, distribution, transformation, and excretion and/or different toxicities. Some metals are nutritionally essential elements at low levels but are toxic at higher levels (e.g., manganese [Mn], iron [Fe], copper [Cu], and zinc [Zn]), while others have no known biological functions (e.g., Pb, arsenic [As], cadmium [Cd], and Hg). Because metals naturally occur in the environment, many organisms developed specific mechanisms for its uptake and deposition, as well as mechanisms to regulate their accumulation, especially the accumulation of essential metals; additionally, the bioaccumulation of metals is tissue specific. All these characteristics can impact the use and interpretation of bioaccumulation data and the toxicity of metals (Fairbrother et al. 2007).
Neurotoxicity of Metals
Neurotoxicity may be defined as any adverse effect, permanent or reversible, on the structure or function of the central and/or peripheral nervous system originated by a biological, chemical, or physical agent that diminishes the ability of an organism to survive, reproduce, or adapt to its environment (Costa 1998; Costa and Manzo 1995). The nervous system can compensate for the toxic effects caused by low doses of neurotoxicants, but a prolonged and lifetime exposure even to the very low levels may lead to delayed neurodegenerative effects (Lucchini and Zimmerman 2009), with a progressive loss of neural tissues (Rachakonda et al. 2004). Thus, neurotoxic effects can be seen in later stages of life, yet the cause of these effects may be related to events occurring decades earlier. The properties that clinically identify them may bear no more than a superficial resemblance to those manifestations marking their prior stages, and this is why the earliest stages of such diseases may be confused with some other sources, such as aging (Weiss 2006). Neurotoxicity is a sensitive endpoint due to the unique and critical role of the nervous system in the control of body function, including other organs and systems, such as the endocrine and the immune system. The limited ability of neurons to regenerate after injury explains neurodegenerative disease-related loss of function, as neurons die, and the regenerative capacity is limited (Emerit et al. 2004; Mutti 1999). These disorders do not have cures (Rachakonda et al. 2004) rather they are gradually progressive, and the ability of its victims to function effectively and efficiently will be impaired at stages of the disease far earlier than its eventual detection (Weiss 2006). Thus concern exists that in the near future, low-dose long-term metal exposure may give rise to a society with lifelong loss of intelligence and motor capacities and permanent psychological disturbances (Kakkar and Jaffery 2005). These effects can produce reduction of economic productivity, and when this reduction occurs widely across a society, the resulting economic impacts may be even greater than the costs of metal pollution control itself (Landrigan et al. 2006).
Chronic exposure to low levels of metals is a contributor to neurological disease in multiple populations around the world (Christensen 1995; Witholt et al. 2000; Wright and Baccarelli 2007). Several studies demonstrate increased levels of metals in critical brain areas of neurodegenerative disease patients (Migliore and Coppedè 2009). The brain may at times compensate for the effects of an individual chemical itself acting on a particular target system; inversely, when multiple targets or functional sites within one system are impacted by different mechanisms (such as in multi-metal exposures), homeostatic capabilities may be impaired, thereby leading to cumulative damage (Lucchini and Zimmerman 2009). The actual public health concern on the potential for exacerbated cognitive and behavioral deficits resulting from children’s exposure to multiple toxic metals provides an example; investigations on the effects on cognition of at least two metals together suggesting that combinations of metals may result in increased toxicity at this level (Kordas et al. 2010). Even so, the effect of mixture interactions on neurotoxicity remains largely unknown (Tiffany-Castiglioni et al. 2006).
Sources, Routes, Disposition, Toxicity, and Mechanisms of Individual Metals
Lead
A recent assessment on the global health impacts of contaminants identified Pb among the six most toxic pollutants threatening human health (Csavina et al. 2012). Pb has many industrial uses including battery manufacture, solders, pigments, and radiation shielding. Its use as an additive in household paint has ceased, but Pb-containing paint is still found in properties built before the 1960s. Occupational exposure usually occurs by inhalation of Pb dust or fumes in Pb industries. Nonoccupational Pb exposure usually involves ingestion, such as “traditional” remedies (adults). Cooking with Pb-glazed earthenware and contaminated soil or water are other potential sources (Bradberry 2016). Concerning Pb disposition once absorbed independently of the route of exposure, 99 percent of circulating Pb is bound to erythrocytes and is dispersed into the soft tissues – brain, liver, renal cortex, aorta, lungs, spleen, teeth, and bones (Patrick 2006). The major route of excretion of absorbed Pb is the kidney.
The mechanisms of Pb neurotoxicity are no doubt complex and numerous. Pb traverses the blood–brain barrier (BBB), accumulates in the brain, and preferentially damages the prefrontal cerebral cortex, hippocampus, and cerebellum (Kwong et al. 2004).
At the biochemical level, one of the most important mechanisms of Pb toxicity is the mimicking of calcium [Ca] action and/or disruption of Ca homeostasis. Pb may also substitute for Zn in some enzymes and in Zn-finger proteins. The fetus and infant may have increased vulnerability to Pb’s neurotoxicity due in part to the immaturity of the BBB and to the lack of the high-affinity Pb-binding protein in astroglia, which sequester Pb. In addition, Pb affects virtually every neurotransmitter system in the brain, including the glutamatergic, dopaminergic, and cholinergic systems (Pohl et al. 2011).
At present Pb entrance pathways in the brain remain elusive, but the most studied candidate to date is the divalent metal ion transporter 1 (DMT1), and studies recently demonstrated that DMT1 is present in endothelial cells of the BBB (Huang et al. 2011).
As the structural basis of blood–cerebrospinal fluid barrier (BCB), epithelial cells in the choroid plexus (CP) are targets for Pb. Pb is known to accumulate in the CP; however, the mechanism of Pb uptake in the choroidal epithelial cells remains unknown. The CP, a major component of the BCB, has been shown to be involved in Pb-induced neurotoxicity (Shi and Zheng 2007). Animal studies showed accumulation of Pb in the CP at concentrations 57- and 70-fold greater than the brain cortex and cerebrospinal fluid, respectively (Zheng et al. 1991). Even though Pb serves no nutritional requirements, pathways for Pb transport, such as DMT1, Ca2+ channels, endocytosis, and anion changers, have been identified in other tissues or cells (Song et al. 2016). Pb can also exert changes on the hematopoietic system, and actually, among the most important enzymes disrupted by Pb are those involved in heme synthesis. Inhibition of delta-aminolevulinate dehydratase (ALAD) leads to accumulation of delta-aminolevulinic acid (ALA) (Bradberry 2016). Higher blood levels of Pb disturb hemoglobin (Hb) synthesis and, therefore, decrease its concentration.
Neurological effects are one of the most sensitive endpoints of Pb exposure, and children are particularly vulnerable. Exposure to high Pb levels produces encephalopathy with signs such as hyperirritability, ataxia, convulsions, stupor, and coma. In children, exposure to low Pb levels has been associated with neurobehavioral effects including impaired cognitive ability and IQ deficits. In Pb workers, reported neurobehavioral effects include malaise, forgetfulness, irritability, lethargy, headache, fatigue, impotence, decreased libido, dizziness, weakness, and paresthesia. Pb exposure in workers has also been associated with neuropsychological effects, increased prevalence and severity of white matter lesions, changes in nerve conduction velocity and postural balance, and alterations of somatosensory evoked potentials (Pohl et al. 2011).
Manganese
Mn is an essential metal ion for life since it is a cofactor for a wide variety of enzymes (Casarett and Doull’s’s 2013). There are inorganic and organic Mn compounds, with the inorganic forms being the most common in the environment. Uses of Mn include Fe and steel production, manufacture of dry cell batteries, manufacture of glass, textile bleaching, and oxidizing agent for electrode coating in welding rods. Organic compounds of Mn are present in the fuel additive, in methylcyclopentadienyl manganese tricarbonyl (MMT), and in fungicides (e.g., maneb and mancozeb). Mn is naturally present in food, with the highest concentrations typically found in soya, nuts, cereals, legumes, fruits, grains, and tea; it is also present at low levels in drinking water. Higher inhalation exposures may be experienced in occupational settings such as Mn mines, foundries, smelters, and battery manufacturing facilities (Santamaria 2008).
The route of exposure can influence the distribution, metabolism, and potential for neurotoxicity of Mn-containing compounds (Anderson 1999; Roels et al. 2012). The oral route is considered to be less important for risk assessment purposes, because Mn is poorly absorbed from the gastrointestinal (GI) tract (5%).
Mn is transported in plasma bound to a gamma 1-globulin, transferrin (TRF), and is widely distributed in the body concentrating in mitochondria, so that tissues rich in these organelles, including the brain, pancreas, liver, kidneys, and intestines, have the highest Mn concentrations. Mn readily crosses the BBB and its half-life in the brain is longer than in the whole body (Casarett and Doull’s 2013). The inhalation route is more efficient than ingestion at delivering Mn to the brain (Gianutsos et al. 1985) due to greater Mn absorption from the lungs and slower clearance of absorbed Mn from the circulation. Another efficient inhalation route is olfactory transport, an often overlooked route of direct delivery of chemicals from the nose to the brain (Brenneman et al. 2000). The principal route of excretion of Mn is in the feces (Casarett and Doull’s’s 2013).
A mechanism for the neurotoxicity of Mn has not been clearly established. A suggested mechanism of Mn neurotoxicity is the increase in auto-oxidation or turnover of intracellular catecholamines including dopamine (DA), norepinephrine, and epinephrine. This leads to the increased production of free radicals, reactive oxygen species (ROS), and other cytotoxic metabolites, along with a depletion of cellular antioxidant defense mechanisms. Other potential mechanisms include the potential substitution for Ca by Mn, the possibility that a transport mechanism for Mn is linked to the DA reuptake carrier, the inhibition of brain mitochondrial oxidative phosphorylation by Mn, and the involvement of Mn in complex interactions with other minerals (Pohl et al. 2011).
The transport of Mn into the brain ultimately depends on its ability to cross the BBB. Similarly to the case of Fe, the transport of Mn across the BBB and its cellular uptake can happen through TRF-dependent and TRF-independent mechanisms (Quintanar 2008). Mn(II) from the bloodstream can also be directly transported by DMT1, or it can cross the cellular membrane using glutamate-activated ionic channels that would normally transport Ca into the cell. Increased plasma Cu and Mn concentrations may lead to brain deposits and CNS damage. It was recently suggested that Mn enters the CNS predominantly through the BCB and that high Mn concentration impairs the integrity of this barrier (Dusek et al. 2015).
Even less information is available pertaining to eventual Mn effects in the heme biosynthetic pathway; one study suggested that Mn can actually interfere at this level (Qato and Maines 1985). More specifically, Mn(II) seems to inhibit aminolevulinic acid synthase (ALAS) activity in the brain and liver. In Maines’ study (1980) the inhibition of liver and erythrocyte ALAD by Mn was observed, while in another work a competitive inhibition of ferrochelatase (FECH) by Mn was exhibited (Hift et al. 2011).
Inhalation of high levels of Mn (as seen in occupational studies) can lead to a syndrome of disabling neurological effects in humans called manganism with symptoms like tremors, difficulty in walking, and facial muscle spasms. Initial symptoms of Mn toxicity that can progress into manganism include irritability, aggressiveness, and hallucinations. Effects similar to the preclinical neurological effects and mood effects seen in occupational studies have also been associated with environmental exposures to Mn in air. In addition, there is evidence that oral exposure to Mn may produce similar neurological effects as reported for inhalation exposure. Exposure to excess levels of Mn in drinking water has been associated with subtle learning and behavioral deficits in children (Pohl et al. 2011).
Aluminum
Al is the most abundant metal and the third most abundant element in the Earth’s crust. Due to its high reactivity, Al is not found in the free state in nature (Casarett and Doull’s’s 2013).
As per World Health Organization (WHO) reports, humans get inevitably exposed to Al through food, cooking utensils, deodorants, and antacids (Kaur et al. 2006) and for purifying water and vaccine adjuvants apart from occupational exposure in gun, automobile, aerospace, and defense-related factories (Sinczuk-Walczaki et al. 2003; Singh and Goel 2015), where inhalation absorption dominates (Buchta et al. 2005). Al overload in dialysis patients has also been reported (Abreo et al. 1990).
It has been reported that only 10% of the ingested Al is absorbed in the GI tract (Gorsky et al. 1979). Al has been shown to accumulate in various mammalian tissues such as the brain, bone, liver, and kidney (Wills et al. 1993; Sahin et al. 1994). Al uptake in the brain is much slower as compared to other organs, but once gained access into the brain, Al distributes into the various regions, namely, the medial striatum, corpus callosum, and cingulate bundle (Kumar and Gill 2014).
After inhalation, Al is distributed into the whole organism. It is excreted only by renal elimination (Buchta et al. 2005).
Although the mechanism of Al-induced neurotoxicity remains elusive, recent reports suggest elevated oxidative and inflammatory stress markers (Kumar et al. 2009) to be majorly responsible for disruption of intraneuronal metal homeostasis (Julka and Gill 1995) as well as axonal transport and long-term potentiation (Wenting et al. 2014). Thus, involvement of multiple mechanisms in Al-induced neurotoxicity warrants multi-targeted approach for effective treatment (Singh and Goel 2015).
It causes oxidative damage by binding to prooxidant metals like Fe and Cu and modulates their ability to promote metal-based oxidative events. Also, Al can directly compete with and even substitute several other essential metals in vivo. Strong evidence suggests that Al forms Al-superoxide anion complex, which is a more potent oxidant than superoxide anion (Oteiza et al. 1993; Nehru and Anand 2005).
Al promotes accumulation of insoluble amyloid β-protein and aggregation of hyper-phosphorylated tau protein, which comprises neurofibrillary tangles (NFTs) (Kawahara and Kato-Negishi 2011) and causes detrimental changes to cholinergic neurotransmission (Sehti et al., Sethi et al. 2009).
Animal studies indicate that Al exposure can affect the permeability of the BBB, cholinergic activity, signal transduction pathways, and lipid peroxidation (LPO), impair neuronal glutamate–nitric oxide–cyclic GMP pathway, and interfere with the metabolism of essential trace elements (Pohl et al. 2011).
Al gains access to the brain through TRF-mediated transport, which subsequently leads to neurotoxicity (Yokel 2006; Singh and Goel 2015). Al is capable of crossing the BBB (Banks and Kastin 1985; Exley 2001), which leads to an increase of Al concentration in the hippocampus (Struys-Ponsar et al. 1997), cortex, singulated bundles, and corpus callosum (Sethi et al. 2009).
Al induces changes in hemato-biochemical parameters in vivo (Ghorbel et al. 2015). It has been demonstrated that Al overload affects two enzymes involved in heme formation, ALAS (the rate-limiting enzyme) and ALAD, as well as the major enzyme of heme degradation, HO (heme oxygenase). Despite, Al increases ALAS activity rather than ALAD, suggesting that Al might promote heme formation. The catabolism of heme prevails over its synthesis (Lin et al. 2013). It has also been proved that Al(III) overdose leads to microcytic anemia, due to its capacity to interfere in heme synthesis, whether by affecting the protoporphyrin biosynthesis or by interfering with Fe metabolism (Bazzoni et al. 2005).
Experimental evidence of Al-induced neurotoxicity subsists since 1965, whereby administration of Al has been reported to induce formation of NFTs in rabbits similar to that found in AD (Klatzo et al. 1965), increasing the risk of neurodegenerative diseases such as AD, Parkinsonism, ALS, etc. (Becaria et al. 2003). Various studies have indicated neuropathological, neurobehavioral, neurophysical, and neurochemical changes following Al exposure (Miu et al., Miu et al. 2003; Colomina et al. 2002; Kaur et al. 2006; Walton 2012). Al concentrations are also elevated in DA-related brain regions of PD patients (Yasui et al. 1992), showing a correlation between PD and Al exposure.
Also, in patients with reduced renal function, prolonged dialysis with Al-containing dialysates has produced a neurotoxicity syndrome (dialysis dementia) characterized by the gradual loss of motor, speech, and cognitive functions (Pohl et al. 2011).
Mercury
Hg is a heavy metal that exists in three chemical forms, metallic or elemental (Hg0), inorganic (Hg1+ and Hg2+), and organic, mostly as methylmercury (MeHg) (Park and Zheng 2012; Hsu-Kim et al. 2013). The general population is primarily exposed to MeHg through their diet (particularly seafood) and Hg0 from dental amalgams (Clarkson et al. 2007). A few studies have reported substantial human exposure to inorganic Hg, which may come from the use of personal products, such as skin-lightening cosmetics (Al-Saleh et al. 2016). Because of its antibacterial/antifungal properties, the organic Hg compound thiomersal is used as a preservative in medical preparations (Lohren et al. 2015). In recent years, man-made MeHg contamination has decreased considerably, due to improvements in industrial manufacturing and efforts to minimize the release of Hg in the environment. However, Hg remains a global pollutant, and there are regions in the world, primarily in developing countries, where the levels of environmental contamination remain high (Ceccatelli et al. 2010).
Hg vapor emitted from amalgam dental fillings is the major source of Hg vapor affecting the general public. Elemental Hg vapor is primarily distributed in the kidneys and oxidizes into inorganic Hg that is predominately excreted in the urine (Al-Saleh et al. 2016). The elemental Hg can then be converted into inorganic Hg in the body which can accumulate in the brain. Ingested MeHg is nearly completely absorbed in the GI tract. Organic Hg deposits in various organs, including the blood, brain, and kidney. More than 90% of blood MeHg is in the red blood cells (RBCs) where MeHg appears to be bound to cysteine residues in Hb. Following MeHg exposure, Hg compounds are excreted mainly via the kidney and the GI tract. Demethylation of MeHg, occurring mostly in the liver, is a key step in the excretion process. Both MeHg and the inorganic Hg formed in the liver are excreted in the bile conjugated with glutathione (GSH) and related compounds. However, MeHg undergoes enterohepatic recirculation. The halftime of excretion varies in different species (70 days in humans) (Ceccatelli et al. 2010).
MeHg distributes to all the areas of the brain by crossing the BBB through mechanisms that are not fully characterized. It is possible that neutral amino acid carrier systems are used for the transport of MeHg–cysteine complexes. Demethylation of MeHg seems to take place in the brain. The formed inorganic Hg has a very long half-life in the brain, especially in the thalamus and pituitary. Inorganic Hg produced by demethylation may be sequestered by metallothioneins (MTs), a family of cysteine-rich proteins that binds with high affinity to metals (Cd, Zn, and Hg) (Ceccatelli et al. 2010).
The cytotoxicity of MeHg has been attributed to three major mechanisms: (1) perturbation of intracellular Ca2+ levels; (2) induction of oxidative stress (OS) either by overproduction of ROS or by reduced oxidative defense capacity; and (3) interactions with sulfhydryl groups, thus forming complexes with thiol-containing compounds [2] targeting proteins and peptides containing cysteine and methionine. Uncontrolled release of Ca2+ from the mitochondria has been reported to occur during OS. The level of ROS increases after exposure to MeHg in brain tissue and in various in vitro neuronal models. MeHg is accumulated in the mitochondria, where it decreases the rate of oxygen consumption, alters the electron transport chain by impairing complex III, and induces loss of the mitochondrial membrane potential. MeHg is also known to interfere with the uptake of cystine, the key precursor of GSH synthesis, via XAG transporters in astrocytes (Ceccatelli et al. 2010).
Previous studies indicate that the BBB is significantly more sensitive to organic Hg species as compared to inorganic compounds (Lohren et al. 2015). However, iHg compounds (e.g., HgCl2) can act as a direct BBB toxicant, increasing thus its permeability in rodents (Zheng et al. 2003). In the human body, Hg ions including MeHg (CH3Hg+) are preferably conjugated to reduced SH groups including cysteine and GSH. The disposition of Hg is regulated by the availability of ligands as well as the ability of the resulting complexes to serve as substrates for a variety of transporters (Ballatori 2002). MeHg-L-cysteine has some structural similarity to the amino acid methionine (Hoffmeyer et al. 2006). Thus, the amino acid transporters, which carry methionine into cells, actually transport MeHg-L-cysteine across membranes (Kerper et al. 1992; Simmons-Willis et al. 2002). Once MeHg has entered the cell, it binds to GSH. The conjugate is a substrate for ATP-binding cassette (ABC) transporters that mediate cellular efflux of glutathione S-conjugates (Strak et al. 2016).
An increase in the urinary concentration of specific porphyrins has been described as a biomarker of prolonged exposure to all forms of Hg (Bowers et al. 1992; Woods et al. 1991; Woods 1995) based upon selective interference with the fifth (uroporphyrinogen decarboxylase) (Woods et al. 1984) and sixth (coproporphyrinogen oxidase) (Woods and Southern 1989) enzymes of the heme biosynthetic pathway in kidney cells, a principal target of Hg. Hg induces a specific change in the urinary porphyrin excretion pattern characterized by increased concentrations of pentacarboxyporphyrin and coproporphyrin, along with the appearance of an atypical porphyrin identified empirically as keto-isocoproporphyrin (Heyer et al. 2006).
The neurotoxicity of MeHg was first recognized in adults during the Minamata outbreaks in 1953 (Ekino et al. 2007), but many subsequent studies reported its toxicity in fetal neurodevelopment (Grandjean and Herz 2011). Data for the neurodevelopmental risk of MeHg at low levels are however still limited due to the different interpretations or study designs (Al-Saleh et al. 2016). All signs and symptoms of toxicity in adults are confined mostly to the nervous system where it affects primarily the granule layer of the cerebellum and the visual cortex of the cerebrum (Kaur et al. 2006).
Exposure to Hg produces neurological and behavioral effects in humans. Adverse neurological effects following acute inhalation of high concentrations of Hg vapor include a number of cognitive, personality, sensory, and motor disturbances. In addition, chronic inhalation exposure has produced signs of neurotoxicity including tremors, unsteady walking, irritability, poor concentration, short-term memory deficits, tremulous speech, blurred vision, performance decrements in psychomotor skills, paresthesias, and decreased nerve conduction (Pohl et al. 2011).
Developmental exposure to MeHg can have long-term consequences, supporting the hypothesis of an increased risk for neurodegenerative disorders later in life (Ceccatelli et al. 2010).
Copper
Cu is an essential metal for all living organisms and is a component of many metalloproteins such as the antioxidant enzyme Cu–Zn superoxide dismutase (SOD) and cytochrome oxidase. Cu salts are used in fungicides, algicides, fertilizers, electroplating, dyes, inks, disinfectants, and wood preservatives (Bradberry 2016). It is mainly used in electric and electronic industry. Mining also contributes to environmental contamination, in soil and water (Angelovicová and Fazekasová 2014).
The brain concentrates heavy metals including Cu for metabolic use. As a cofactor of several enzymes and/or as structural component, Cu is involved in many physiological pathways in the brain (Scheiber et al. 2014). In general, Cu contents are higher in the gray matter (1.6–6.5 mg/g wet weight) than in the white matter (0.9–2.5 mg/g wet weight); Cu is enriched in the locus coeruleus and the substantia nigra, which both are pigmented tissues and contain catecholaminergic cells. Both brain Cu content and distribution change during development, with age and in neurodegenerative diseases (Scheiber et al. 2014).
Although Cu is an essential element, it also plays a role in the pathogenesis of neurodegenerative disease such as AD (Lu et al. 2006). Free reduced Cu(I) can bind to SH groups and inactivates enzymes such as glucose-6-phosphate dehydrogenase and glutathione reductase. In addition, Cu can interact with oxygen species (e.g., superoxide anions, hydrogen peroxide) and catalyze the production of reactive toxic hydroxyl radicals (Bradberry 2016). Cumulative evidence has implied that an imbalanced Cu homeostasis in the brain contributes to the pathogenesis of neurodegenerative disorders such as idiopathic Parkinson’s disease (IPD), AD, and familial amyotrophic lateral sclerosis (ALS). Increased concentration of redox available Cu has been reported in PD CSF, and its concentration was correlated with motor impairment (Dusek et al. 2015).
Brain Cu is derived from peripheral Cu that is transported across the BBB and/or the BCB, which separate the brain interstitial space from blood and CSF, respectively (Zheng and Monnot 2012). At both barriers Cu is transported primarily as free ion. Although the Cu uptake into cerebral capillaries is much slower than into the CP, the Cu acquired by cerebral capillaries appears to be more readily transported into the brain parenchyma than Cu from the CP to the CSF. In fact, recent evidence indicates that the role of the BCB in brain Cu homeostasis is rather to export Cu from the CSF to the blood than to import Cu. The BBB represents the major route for the transport of Cu from the blood circulation into the brain parenchyma, where Cu is utilized and subsequently released into the CSF via the brain interstitial fluid. The Cu in the CSF can be taken up by choroid epithelial cells, from where it may be stored or exported to the blood. Thus, while the BBB determines the influx of Cu into the brain, the BCB contributes to the maintenance of the Cu homeostasis in the brain extracellular fluids (Gunshin et al. 1997).
The Cu transporter Ctr1 is likely to be the major pathway for Cu entry into brain cells. Experimental evidence was provided that DMT1, which is also expressed in brain cells, is involved in apical Cu uptake by intestinal cells. However, while some authors defend that DMT1 clearly can transport Cu (Garrick et al. 2006), others consider that DMT1 is a Fe-preferring transporter that does not transport Cu (Illing et al. 2012). The alternative Cu transport could be mediated by members of the ZIP (ZRT-/IRT-like protein) family of metal transporters (Scheiber et al. 2014).
The synthesis of hemoproteins may also be affected by Cu deficiency. It is well known that Cu deficiency can lead to anemias which might be explained by an intracellular defect in heme biosynthesis at FECH (Wagner and Tephly 1975).
Alterations of Cu homeostasis have also been associated with neurodegenerative diseases such as prion, AD, PD, and Huntington’s disease (Scheiber et al. 2014). In AD Ab peptides have been shown to bind with high affinity (Atwood et al. 2000), and senile plaques are strongly enriched in Cu (Lovell et al. 1998). In addition, Cu ions induce the precipitation of Ab peptides in vitro. These observations suggest that Cu triggers the formation of plaques in the brain (Atwood et al. 2000; Tougu et al. 2011).
In PD, strong and growing evidence suggests abnormalities in Cu homeostasis. Parkinsonism is a frequent symptom in neurological Wilson’s disease, which is an inherited disorder of Cu metabolism that is characterized by excessive deposition of Cu in the liver, brain, and other tissues (Lorincz 2010). Cu has been demonstrated to bind to both soluble and membrane-bound α-synuclein with high affinity (Dudzik et al. 2012) and to accelerate aggregation of soluble a-synuclein (Davies et al. 2013).
Zinc
Zn is a nutritionally essential metal, and a deficiency results in severe health consequences. At the other extreme, excess of Zn is relatively uncommon and occurs only at very high levels. Zn is ubiquitous in the environment, so that it is present in most foodstuffs, water, and air. The principal industrial uses of Zn include its applications as a corrosion protector for Fe and steel, application in batteries, and production of metal alloys, brass, and bronze. Zn oxide is the most widely used compound in industry, in the production of paints, plastics, cosmetics, pharmaceuticals, textiles, and electrical and electronic equipment (Peakall and Burger 2003).
The distribution of endogenous Zn is high in the bone, testis, and liver (Yasuno et al. 2011). High concentrations of Zn are also found in the brain and in pancreatic cells. In the brain, Zn highly occurs in the hippocampus, amygdala, and cortex (Kozlowski et al. 2009).
The molecular mechanisms by which Zn2+ triggers neuronal injury have not been elucidated clearly. There are several possibilities in which Zn exerts their adverse effects, including impairment of mitochondrial superoxide production (Sensi et al. 2000), disruption of metabolic enzyme activity (Sensi and Jeng 2004), and activation of p38 and voltage-dependent potassium channels (Zhu et al. 2013). Deregulation of neuronal Zn(II) homeostasis is believed to be strictly connected to mitochondrial dysfunction and OS, making the cation a possible contributor to the activation of pathophysiological pathways involved in brain aging and/or neurodegeneration. Zn, in its ionic form, can also exert important modulatory effects on neurotransmission and synaptic function, as well as regulate many signaling pathways (Kozlowski et al. 2009).
Zn permeability for TRPM7 channels is fourfold higher than that of Ca. Recently, TRPM7 channel is reported to play an important role for Zn2þ-mediated neuronal injury and may represent a novel target for neurological disorders where Zn2þ toxicity plays an important role (Kim et al. 2016a, b).
MTs exert a critical role in buffering cytosolic Zn(II). MT-3 seems to be particularly relevant to neuronal Zn(II) homeostasis in critical brain regions such as the hippocampus where it is abundantly present in the same hippocampal glutamatergic terminals that are also strongly enriched in vesicular Zn(II) (Kozlowski et al. 2009). Zn is selectively stored and released from presynaptic vessels of neurons found primarily in the mammalian cerebral cortex (Nriagu 2007).
The most common effects associated with long-term excessive Zn intakes (ranging from 150 mg/day to 1–2 g/day) have included sideroblastic anemia, hypochromic microcytic anemia, leukopenia, lymphadenopathy, neutropenia, hypocupremia, and hypoferremia. Changes in serum lipid profile, serum ferritin, and erythrocyte SOD activity have been reported in a number of patients who have ingested high doses of Zn. Zn is required for the activity of ALAD which plays a protective role in heme biosynthesis (Nriagu 2007).
Concerning neurotoxic effects, an interesting body of scientific literature suggests that Zn is a neurotoxin. There is evidence indicating that the readily available Zn2+ could, in certain pathological states, induce neuronal injury. Exposure of mature cortical neuronal cultures to several hundred μM concentrations of Zn2+ induced neuronal death. These findings indicate that Zn dyshomeostasis is likely a key modulator of neuronal injury. A previous study has reported that Zn2+ induces neurotoxicity in a concentration- and time-dependent manner (Zhu et al. 2012). Because the Zn-releasing neurons also release glutamate, they are sometimes referred to as “gluzinergic” neurons. Zn can modulate the overall excitability of the brain possibly through its effects on glutamate, gamma-aminobutyric acid (GABA) receptors of this network (Nriagu 2007).
Arsenic
Arsenic is particularly difficult to characterize as a single element because its chemistry is so complex and there are many different As compounds. It may be trivalent or pentavalent and is widely distributed in nature. The most common inorganic trivalent As compounds are As trioxide, sodium arsenite, and As trichloride (Casarett and Doull’s 2013).
Arsenic is one of the oldest poisons known to men and its applications throughout history are wide and varied. The catastrophe of As toxicity, caused by As-contaminated water, has already been reported in many countries. Yet, an estimated 100 million people worldwide are exposed to excessive amounts of As via drinking water (in the range of ppm) (Watanabe and Hirano 2013; Krüger et al. 2009). The atmospheric deposition of As through the burning of charcoal and activities of metal foundry are examples of human activities that contributes to As environmental contamination (O’Neil 1995), being the excessive use of pesticides and fertilizers and mining the factors that most contribute to As soil contamination (Adriano 2001).
Both arsenate (pentavalent inorganic As) and arsenite are well absorbed by oral and inhalation routes. Absorption by the dermal route has not been well characterized but is low compared to other routes (Casarett and Doull’s).
Once absorbed, arsenates are partially reduced to arsenites, yielding a mixture of As(III) and As(V) in the blood. As(III) undergoes methylation primarily in the liver to form monomethylarsonic acid (MMA) and dimethylarsinic acid (DMA). The rate and relative proportion of methylation production varies among species. Most inorganic As is promptly excreted in the urine as a mixture of As(III), As(V), MMA, and DMA. Smaller amounts are excreted in feces. In most species, including humans, ingested organic arsenical compounds such as MMA and DMA undergo limited metabolism, do not readily enter the cell, and are primarily excreted unchanged in the urine (ATSDR 2007). Some of these metabolites are more potent and toxic than the originally ingested inorganic form of As (iAs), including mono- and dimethylated arsenicals (Watanabe and Hirano 2013; Krüger et al. 2009).
All forms of As, including inorganic and methylated arsenicals, accumulate in many parts of the brain, with the highest accumulation in the pituitary (Sanchez-Pena et al. Sánchez-Peña et al. 2010).
The metabolism of iAs consumes GSH, which is the main antioxidant in the CNS (Dringen 2000). Arsenic may induce OS by cycling between oxidation states of metals such as Fe or by interacting with antioxidants and increasing inflammation, resulting in the accumulation of free radicals in cells. Inadequate GSH availability may modulate iAs biotransformation and determine disease (Ramos-Chávez et al. 2015).
The role of OS as the leading mechanism in As-induced neurological defects has been widely supported by in vitro and in vivo studies. OS may be the initiating mechanism for As-induced neurotoxicity. Arsenic-induced DNA damage and apoptosis in neuronal cells may follow an intrinsic mitochondrial apoptotic pathway, mediating through increased intracellular Ca that triggers mitochondrial stress and generation of ROS (Flora 2011).
In occupationally As-exposed subjects, a positive correlation between compromised subjective neurological symptoms, visual evoke potential, electroneurographic and electroencephalographic results, and As concentration in air and urine was established (Flora 2011).
Studies in murine models have demonstrated that iAs crosses the BBB and is methylated in different brain regions that express the As 3 methyltransferase (AS3MT) enzyme (Ramos-Chávez et al. 2015). It was identified in two uptake pathways: aquaglyceroporin (AQP) channels, in particular the liver isoform AQP9, and the glucose permease (GLUT1) conduct trivalent As(OH)3 and CH3As(OH)2 which both have oxidative status of +3. It was proposed that GLUT1 is the major pathway for the movement of trivalent inorganic and methylated As into the brain and heart, where AQPs are not abundantly expressed, and this uptake may contribute to cardiovascular disease and neurotoxicity (Jiang et al. 2010)
Considerable evidence supports the observation that As can influence many aspects of the heme system. Previous research has shown that As decreases heme metabolism and can bind to Hb, resulting in lower Hb concentrations. Arsenic has been shown to alter erythrocyte morphology and induce erythrocyte death. Arsenic also depresses bone marrow, which can lead to anemia (Hb < 120 g/L in nonpregnant adults), leukopenia, and thrombocytopenia. Several studies have shown that As alters heme metabolism and contributes to lower Hb concentrations (Kile et al. 2016).
Epidemiological studies show that As can cause serious neurological effects after inhalation or oral exposure. Common effects seen in humans orally exposed to As are peripheral and/or central neuropathy (Pohl et al. 2011). A recently published meta-analysis focused on the negative impact of As exposure on intelligence measured by IQ tests (Rodriguez-Barranco et al. 2013). A 2007 study found a significant association between urinary As concentrations greater than 50 μg/L and poor scores on tests measuring visual-spatial reasoning, language and vocabulary, memory, intelligence, and math skills in 6–8-year-old children from Mexico (Rosado et al. 2007).
Exposure to high levels of As produces mainly CNS effects, and exposure to low levels produces mainly peripheral nervous system effects (Pohl et al. 2011). According to Naujokas et al. (2013), exposure to low As concentrations has been shown to increase susceptibility to cognitive dysfunction. The mechanism of As-induced neurological changes has not been determined. However, some of the neurological effects of high-level oral exposure are thought to be the result of direct cytotoxicity. In addition, animal studies have shown altered neurotransmitter concentrations in some areas of the brain after oral exposure to As (Pohl et al. 2011).
Iron
The major scientific and medical interest in Fe is as an essential metal, but toxicologic considerations are important in terms of Fe deficiency, accidental acute exposures, and chronic Fe overload. Environmental exposure to Fe does occur, for example, from drinking water, Fe pipes, and cookware (Rush et al. 2009). But further evidence indicates that overload of Fe, which is released from Hb, also contributes to brain injury after intracerebral hemorrhage. The toxicity resulting from Fe deposition in neurons is primarily mediated by the increasing of Fe that can generate radical species via the Fenton’s reaction since radical species are frequently associated with cytotoxicity, which is the initiating stimuli for cell death (Dai et al. 2013).
The total amount of Fe in the body is mainly present in the form of Hb (60–70%), myoglobin, cytochromes, and other Fe-containing enzymes (10%) as well as in ferritin and hemosiderin (Appel et al. 2001).
Fe absorption is accomplished by enterocytes in the proximal small intestine, near the gastroduodenal junction. Its access to the circulation is modulated by transport via both the apical and basolateral membranes, which is operated by specific transporter proteins and accessory enzymes. Nonheme Fe is first reduced to Fe(II), which is transported by DMT1 (Skjørringe et al. 2015). Some of the absorbed Fe is stored in enterocytes’ ferritin, and some is exported to the circulation by ferroportin (FP). Absorbed Fe is rapidly delivered to TRF, which under normal conditions accounts for nearly all serum Fe. In normal human subjects, plasma TRF is only approximately 30% saturated. The absence of a regulated mechanism for Fe excretion determines the necessity of a tight balance between the sites of Fe absorption, uptake, transport, storage, and utilization for maintenance of Fe homeostasis (Kozlowski et al. 2009). After the liver, the brain contains the highest quantity of Fe, ca. 60 mg of nonheme Fe distributed uniquely according to brain structures. Fe is the most abundant trace element in the brain where it is essential for normal brain development and function (Ward et al. 2014). It plays a crucial role for many processes including oxygen transport, the synthesis of DNA and RNA, and the formation of myelin and development of the neuronal dendritic tree (Lieu et al. 2001). The substantia nigra and globus pallidus in normal adult human brain can contain Fe levels, which exceed hepatic levels, in the range of 3.3–3.8 mM Fe. These high brain Fe concentrations can be attributed primarily to the rapid rate of oxidative metabolism necessary to maintain ionic membrane gradients, axonal transport, and synaptic transmission (Kozlowski et al. 2009). Postmortem and in vivo magnetic resonance imaging studies have shown that Fe accumulation follows an exponential saturation function with only little changes after the fourth to fifth decades of life (Pirpamer et al. 2016).
Ferrous Fe showed higher influx into cells than ferric Fe and induced more ROS production which resulted in higher susceptibility of neuron death. The types of neuronal cell death which were induced by Fe overload were testified as necrosis, apoptosis, and autophagic cell death, relying on the level of Fe dosage (Dai et al. 2013).
Two main events linked to increased ROS generation have been identified in the degenerating substantia nigra: (a) increased Fe levels and (b) reduced antioxidant defenses. Fe deposition seems to be a specific hallmark of PD (Rubio-Osornio et al. 2013).
The toxicity of Fe deposition in neurons is primarily mediated by the increasing of Fe that can generate radical species via the Fenton’s reaction, since radical species are frequently associated with cytotoxicity, which is the initiating stimulus for cell death (Dai et al. 2013).
Brain Fe unpaired regulation may also result from the disrupted expression of brain Fe metabolism proteins induced by nongenetic factors. These currently undetermined factors may disrupt normal control mechanisms of protein expression and lead to Fe imbalance in the brain, inducing then OS and neuronal death in some neurodegenerative disorders. Fe accumulation in the brain occurs gradually over time with concurrent increases in ferritin. Fe overload results in a large increase in the chelatable free Fe pool, which is too large to be sequestered by ferritin within cells. Fe toxicity, largely based on Fenton chemistry, mainly affects the mitochondrial inner membrane respiratory complexes (Kozlowski et al. 2009).
The transport of Fe across the BBB must be regulated, but the permeation mechanism has not been completely clarified so far. The uptake of TRF-bound Fe by TFR-mediated endocytosis from the blood into cerebral endothelial cells is no different in nature from the uptake into other cell types. After permeation across the BBB or blood–CSF barrier, Fe is likely to bind quickly to the TRF secreted by the oligodendrocytes and CP epithelial cells, which, diversely from what happens in other tissues, becomes fully saturated with Fe. The excess Fe will bind to other transporters, including small molecules like citrate or ascorbate. The widespread distribution of TFR in neurons clearly indicates that neurons can acquire Fe by means of TFR-mediated uptake of TF–Fe (Kozlowski et al. 2009).
Heme is an Fe-containing porphyrin that functions as a cofactor in a wide array of cellular processes. The terminal step of heme biosynthesis, which occurs in the mitochondrial matrix, is the insertion of Fe into protoporphyrin IX (Korolnek and Hamza 2014). Further evidence indicates that overload of Fe, which is released from Hb, also contributes to brain injury after intracerebral hemorrhage (Dai et al. 2013). Fe can be released from the breakdown of Hb following aneurysm or blood disease (Rush et al. 2009).
Fe deposition has also been associated with inflammatory, neurodegenerative, and cerebral small vessel disease. Even in normal elderly persons, elevated levels of Fe relate to worse cognitive performance (Pirpamer et al. 2016). Syndromes with neurodegeneration with brain Fe accumulation (NBIA) are a group of neurodegenerative disorders characterized by abnormalities in brain Fe metabolism with excess Fe accumulation in the globus pallidus and to a lesser degree in the substantia nigra and adjacent areas (Schneider 2016).
The brain of Alzheimer’s diseased humans is characterized by the accumulation of Fe within senile plaques (ca. 1 mM) and NFTs and also by lowered expression of TRF receptor. As a consequence, these brains are subject to high levels of OS. Fe may also promote a deposition and may affect the enzymatic processing of the amyloid precursor protein. As for PD, DA cell loss and disease progression are accompanied by the accumulation of high Fe concentrations that are particularly associated with aggregation of alpha-synuclein (especially the mutated form found in familial PD) within Lewy bodies. An increased Fe content can be detected in the substantia nigra of most PD patients, and up to a 255% increase in intracellular Fe concentration has been observed in postmortem PD brains. Together with Fe accumulation, the lowered expression of ferritin within the substantia nigra of PD patients results into OS and decreased GSH levels, thus directly contributing to DA neuronal toxicity (Kozlowski et al. 2009).
Cadmium
Cd is a toxic nonessential transition metal classified as a human carcinogenic (ATSDR 2004). It is characterized by a long half-life in humans (Jin et al. 1998) with a low rate of excretion from the body. The Cd content of the body increases with age in industrialized societies, from less than 1 μg in the newborn to 15–20 mg in adults (Notarachille et al. 2014).
There are several sources of human exposure to Cd, including employment in primary metal industries, production of certain batteries, some electroplating processes, and consumption of tobacco (ATSDR 2012).
Cd is poorly absorbed after oral ingestion. The estimated absorption of Cd is less than 5% from the GI tract in humans and about 1% in animals. Once absorbed, the movement of Cd from blood to tissue is rapid. Over 60% of the body burden of Cd is localized in the liver and kidney. However, the factors that influence absorption and tissue distribution of Cd are not well understood (Liu and Klaassen 1996).
Cd can also be absorbed by the inhalation and dermal routes regardless its chemical form (chloride, carbonate, oxide, sulfide, sulfate, or other forms), although dermal route of exposure is relatively insignificant and of low concern (Wester et al. 1992). Cd can reach the CNS being uptaken from the nasal mucosa or olfactory pathways (Lafuente and Esquifino 1999). Cd is not known to undergo direct metabolic conversions. It has a high affinity for the SH groups of albumin and MT. The interaction between Cd and MT plays a critical role in the toxicokinetics and toxicity; Cd is retained in both organs, liver and kidney, bound mainly to MT.It has a retention halftime of 73 days in the liver and a lifetime in the kidneys. Since a small fraction of the Cd presented to the GI tract is absorbed, most of the oral dose is excreted via the feces. The amount of Cd excreted in urine represents only a small fraction of the total body burden unless renal damage is present. Absorption of Cd is also influenced by metal ions such as Zn, Fe, Ca, and chromium (Cr) (ATSDR 2012).
Cd-induced injury in the cerebral microvessels is thought to be associated with OS. Following in vivo Cd exposure, there was an early increase followed by a later decrease in microvessel enzymes involved in cellular redox reactions, such as SOD, glutathione peroxidase, and catalase. Thus, a depletion of microvessel antioxidant defense systems and a resultant increase in LPO may provoke microvessel damage (Shukla et al. 1996). Cd significantly increases the levels of LPO in parietal cortex, striatum, and cerebellum as compared to a control group in developing rats exposed to Cd (Méndez-Armenta et al. 2003).
Under normal conditions and by oral route, Cd barely reaches the brain in adults due to the presence of the BBB (Yang et al. 2016). Differently, concerning inhalation pathway, Cd is transported along the primary olfactory neurons to their terminal in the olfactory bulbs, thereby bypassing the intact BBB, the olfactory route could therefore be a likely way to reach the brain and should be taken into account for occupational risk assessments for this metal (Tjälve and Henriksson 1999; Bondier et al. 2008).
In the brain, Cd tends to accumulate in the CP at concentrations much greater than those found in the CSF and elsewhere in brain tissues. A postmortem human study revealed that Cd concentration in the CP was about two to three times higher than that found in the brain cortex (Zheng 2001). Due to differences in the BBB integrity (Antonio et al. 2002), Cd is thus more toxic to newborn and young rats than to adult rats. Cd can increase the permeability of the BBB in rats (Shukla et al. 1987) and penetrate and accumulate in the brain of developing and adult rats, leading to brain intracellular accumulation, cellular dysfunction, and cerebral edema (Mendez-Armenta and Ríos Méndez-Armenta and Ríos 2007).
Cd causes anemia through three known mechanisms, i.e., hemolysis due to a deformity of peripheral RBCs, Fe deficiency through the competition with duodenal Fe absorption, and hypoproduction of erythropoietin (Horiguchi et al. 1994, 2010), an erythroid-specific glycoprotein hormone produced mainly from the kidney that promotes RBC formation (Ebert and Bunn 1999). However, there has been rare investigation on the direct toxicity of Cd to erythroid precursors (Wang et al. 2013).
In addition to liver and kidney target organs, the CNS is also subjected to Cd toxicity (Lafuente and Esquifino 1999). Cd can enter the brain parenchyma and neurons, causing neurological alterations in humans and animal models by inhibiting thiol-containing enzymes and decreasing serotonin and acetylcholine levels (Jomova and Valko 2011).
Exposure to Cd also severely affects the function of the nervous system (López Alonso et al. 2003) with symptoms including headache and vertigo, olfactory dysfunction, parkinsonian-like symptoms, slowing of vasomotor functioning, peripheral neuropathy, decreased equilibrium, decreased ability to concentrate, and learning disabilities (Kim et al. 2005; Monroe and Halvorsen 2006).
Concerning biochemical changes of CNS in response to Cd, it can inhibit the release of acetylcholine, probably by interfering with Ca metabolism (Desi 1998). Cd can affect the degree and balance of excitation/inhibition in synaptic neurotransmission as well as the antioxidant levels in animal brain (Mendez-Armenta and Ríos Méndez-Armenta and Ríos 2007).
Moreover, there are studies showing the neurotoxicity of Cd at μM range on cell culture models such as neurons and glial cells (Lopez et al. 2006).
Metal Mixture Neurotoxicity
In all environmental media, mixtures of neurotoxic metals naturally occur, and metals are often introduced into the environment as mixtures (Fairbrother et al. 2007). In this context, Pb, As, Hg, and Cd are metals/metalloids included in a WHO’s list of ten chemicals of major public concern (Prüss-Ustün et al. 2016), with Pb and As being among the leading toxic agents detected in the environment (Järup 2003). The four metals are thought to exhibit their neurotoxic effects (Pohl et al. 2011; Kaur et al. 2006) through common mechanisms, such as the generation of ROS (Patrick 2006; Flora 2011; Kaur et al. 2006; Méndez-Armenta et al. 2003) and interaction with essential metals (Pohl et al. 2011; Lin et al. 2013; Freitas Fonseca et al. 2014; Nriagu 2007). Hence, interactions among Pb, As, Hg, and Cd will be addressed, and emphasis will be also given to the interactions of these metals with Fe, Cu, and Zn. Indeed, a mechanistic relationship is established between the dyshomeostasis of these essential metals and OS, as well as associations between both conditions and neurodegeneration (Kozlowski et al. 2009).
Mechanisms Associated with Interactions of Lead with other Metals
Some populations at high risk for Pb toxicity are also overexposed to other metals through environmental pollution (Shukla et al. 1987), most of them nearby areas of industrial activity. Namely, chronic exposure to low levels of Pb and Cd through food, water, and air is described as common in industrial areas (Markiewicz-Górka et al. 2015). Other authors mention that sources of co-exposure to Pb and As are primarily through soil and dust deriving from pyrometallurgical nonferrous metal production or mining activity (Csavina 2012). It is also described that Pb, Al, and Zn are constituents, in major or trace amounts, of raw materials and wastes associated with the production of alumina (Phol et al. 2011). These examples illustrate that some populations are exposed to potential worrisome levels of metal mixtures. Accordingly, children residing near ore smelters have been shown to be exposed to Pb, As, Cd, Zn, and Cu (Shukla et al. 1987), while the levels of Pb, as well as As, Hg, Cd, Mn, Zn, and Cu, in whole blood of residents from mining communities were found to exceed the permissible WHO guideline values (Obiri et al. 2016).
Other co-exposure scenarios to the general population are not rare, since Pb can occur simultaneously with As, Hg, and Cd in various parts of the ecosystem (Cobbina et al. 2015). Mixtures of metals can be present in the air, being an example the determination of urban metal levels in Pakistan which led to find that Pb and also Cd, Zn, Fe, and Cu are among the dominant contributors in indoor particulates, while Pb, Zn, and Fe constituted the major components in outdoor particulates. The excessive concentrations of Pb, Cd, and Zn were associated with automobile emissions. Even so, the Cd and Cu levels estimated in this study were considerably lower than those for Mexico City, in Mexico (Nazir et al. 2011). The contamination of mixtures of heavy metals in the aquatic environment has also attracted global attention owing to its abundance, persistence, and environmental toxicity. A study performed in a river in Bangladesh showed that the levels of Pb, As, and Cd indicated that water was not safe for drinking and/or cooking (Ali et al. 2016). Metal emissions can also contaminate the soil, with mixtures of Pb, Mn, Zn, and Cu occurring in this environmental compartment (Roneya and Colman 2004). Furthermore, heavy metals prone to bioaccumulate in the food chain might be dangerous to human health. In corroboration, the metal contents in plants and soil show significant correlations for Pb, Cd, Cu, and Zn, while the contents of the metals in vegetables often exceed those allowable for normal human and animal consumption. It has been estimated that if an adult consumed 2 kg potatoes, 2 kg tomatoes, and 1 kg carrots in a week, his/her food would exceed by 12% the maximum allowable level (MAL) for Cd; the daily maximum allowable rate of ingested Pb could be reached by consuming 880 g of vegetables (equal parts of potatoes, tomatoes, carrots, and cucumbers) (Islam et al. 2007). Another study in Korean pregnant women showed that co-exposure to Pb and Hg could come from frequent fish and cereal or vegetable consumption; while fish consumption was positively associated with Hg levels in cord blood, cereal and vegetable consumptions were positively associated with Pb levels (Kim et al. 2016a, b). Pb can also migrate from artisanal Al cookware and enter food at unacceptable levels that can significantly contribute to a child or adult’s body burden of this metal (Weidenhamer et al. 2014).
-
Disposition in the Brain
There is a general recognition that each mixture component may affect the disposition of other mixture components in the CNS. Being the brain a common target organ for Pb and other metals, potential additive or synergic effects induced by metal mixtures are expected (Mendez-Armenta and Ríos Méndez-Armenta and Ríos 2007).
Concerning Pb interactions with As, the intraperitoneal administration of both metals resulted in increased brain regional Pb levels in rats, accompanied by a significant decrease of As levels in some other regions (Mejía et al. 1997). In the same way as As, Cd can increase the level of Pb in certain brain regions with a magnitude greater than that observed after Cd exposure alone; the accumulation of Cd in several brain regions also increased. It is plausible the possibility that the co-exposure of Pb and Cd can damage the BBB. In this study, the levels of blood Pb decreased suggesting that blood Pb level as a diagnostic tool for Pb toxicity in co-exposed conditions is of insignificant value (Shukla et al. 1987). Again, when Pb is administrated together with Hg or with As/Hg, Pb brain accumulation can increase by 83.6% and 76.1%, respectively (Cobbina et al. 2015).
Pb can also interact with essential metals, such as Mn, resulting in changes in Pb disposition. Even very low doses of Mn when administrated together with Pb can raise striatal Mn, and also Pb (Suchla and Chandra 1987), as well as augments of Pb concentration in the whole brain of adult rodents and in pups after their co-exposure during gestation and/or lactation (Mejía et al. 1997). Overall, these informations indicate that Mn in the presence of Pb increases its distribution and/or retention in the brain (Pohl et al. 2011). The presence of excess Mn in the brain might increase the affinity of brain tissue to bind Pb, as it was suggested by in vitro results (Kalia et al. 1984; Shukla et al. Shukla et al. 1987; Wright and Bacarelli 2007). Analogous outcomes arise when Pb is administrated in vivo through a ternary metal mixture of Pb/As/Mn. Increased levels of Pb in the brain as compared with single exposures to each one of these metals, including the exposure to Pb alone, were already observed. Again, blood Pb levels fail to reflect the increased Pb deposition in the brain, raising concern that blood Pb levels may underestimate risk associated with co-exposure to Pb and other metals (Andrade et al. 2014b).
Fe, Zn, and Cu are also essential metals, however, considered protective of the toxic effects of Pb (Pohl et al. 2011; Klauder and Petering 1975). Interactions of Pb with Fe are broadly referred, and at the brain level, while Pb exposure damages the integrity of the BBB in young animals, Fe supplement may prevent against Pb-induced BBB disruption, with significantly reduced Pb levels in this organ (Wang et al. 2007). Other studies demonstrated that Pb exposure significantly increased Pb concentrations in cerebral cortex and low Fe dose significantly reduced the cortex Pb levels. Remarkably, Fe high dose increased the cortex Pb levels (Zhi et al. 2015). Cu has been shown to impede the GI absorption of Pb, with in vivo studies showing that at higher supplemental Cu doses and higher Cu/Pb dose ratios, supplemental Cu can decrease blood, liver, and kidney concentrations of Pb; however, levels of Pb in the brain were not affected (Pohl et al. 2011). Concomitantly, other studies with ternary and quaternary mixtures led to observe that the exposure to the mixtures Pb/Hg/Cd and Pb/Hg/As/Cd increased brain Cu by 31.9% and 40.8%, respectively (Cobbina et al. 2015). By its turn, Zn can also reduce Pb toxicity due to its antagonistic effect on Pb absorption from the GI tract (Basha et al. 2003). Furthermore, hippocampal Pb levels decreased, as compared to exposure to Pb alone, when rodents were co-exposed to Pb and Zn (Basha et al. 2003; Piao et al. 2007). The co-exposure to Pb, As, Cd, and Hg can result in brain Zn reduction by 65.1%, which was attributable to mimicry of Zn by complexes of toxic metals like As in the mixture (Cobbina et al. 2015).
The mechanisms responsible for the elevation of Pb in the brain, when the metal is in the presence of As, Cd, and/or Hg, are not elucidated. Nevertheless respecting to essential metals, their transporters can be “hijacked” by nonessential metals possessing physicochemical similarities (Martinez-Finley et al. 2012), modifying their uptake and accumulation (Spurgeon et al. 2010). Namely, DMT1 is present in several tissues including the BBB endothelium [Wang et al. 2011) and in both glia and neurons. This transporter is also most likely the major pathway by which Pb is transported into the brain (Wang et al. 2011); it is also involved in the uptake of Hg and Cd at least in intestinal cells (Vazquez et al. 2015; Tallkvist et al. 2001). Gu et al. (2009) also observed an effect greater than additive on DMT1 protein synthesis, enhancing transport of ions in the developing rat brain resulting from exposure to both Pb and Cd orally (von Stackelberg et al. 2013). Plausibly competition for DMT1 may have a relevant role in the increased deposition of Pb in the brain, when in the presence of Cd, Hg, and/or Mn. Differently GLUT1 may be a major pathway uptake of As in the epithelial cells of the BBB (Liu et al. 2006), and hence, the mechanisms leading to increased Pb brain levels after co-exposure to Pb and As remain to be elucidated (Andrade et al. 2014b).
DMT1 is also involved in active transport of Mn, Fe, Cu, and Zn, although Zn shows a different relative capacity (Espinoza et al. 2012; Garrick et al. 2006). These essential metals are harmful in oversupply, and thus, DMT1’s role in their homeostasis is very relevant (Garrick et al. 2006).
Deficiencies in Fe can contribute to increased brain Pb levels; during periods of low Fe stores, expression of DMT1 in the duodenum is greatly increased, allowing not only increased Fe absorption but also Pb absorption (Cobbina et al. 2015). DMT1 regulation in the duodenum is sensitive to levels of Fe uptake, and the transporter has a much higher affinity for Fe over Pb (Wang et al. 2011). Nevertheless, the possibility that the expression of other Fe influx transport protein might contribute to increased brain Pb accumulation is not excluded (Zhu et al. 2013). On the other hand, Pb may limit Fe absorption, however through a different mechanism, one similar to Cd which downregulates the expression of DMT1 to 70% of controls when added to human intestinal cells (Kwong et al. 2004). Concomitantly, Gu et al. (2009) have reported that exposures to Pb and Cd synergistically increase DMT1 protein expression in the CNS of developing rats. Largely studies are still needed to clarify the affinities of different metals to DMT1, in order to provide a better understanding regarding interactions among metals during its transport in different tissues. According to a report of an in vitro study, the following order for DMT1 transport affinities is Mn >?Cd >?Fe > Pb > Zn with, as mentioned by the authors, doubts pertaining to Cd and Fe and uncertainty on where to place Cu (Garrick et al. 2006).
Another study proposes a different order: Cd2+ > Fe2+ > Mn2+ >> Zn2+ (Illing et al. 2012). Besides, while some authors defend that DMT1 clearly can transport Cu (Garrick et al. 2006), others consider that DMT1 is a Fe-preferring transporter that does not transport Cu (Illing et al. 2012). It is also very likely that additional mechanisms contribute to changes in the disposition of Pb, when the metal is present with other metals. Actually once within the cells, a particular metal may occupy abundant binding sites (Kalia et al. 1984) on metalloproteins or target molecules, modifying the compartmentalization of other metals, thus leading to aberrant binding and toxicity (Spurgeon et al. 2010). Furthermore a given metal may induce overexpression of transporters and/or binding proteins that alter the uptake of other metals (Kalia et al. 1984; Molina et al. 2011). In this perspective emphasis will be given to MTs, which are Zn-rich metal-binding proteins (Nordberg and Nordberg 2009). Zn induces the expression of a MT that has been shown to sequester Pb in vitro, protecting cells against its cytotoxicity (Roneya and Colman 2004). Many investigators proposed that Pb and Zn compete for similar binding sites on a MT-like transport protein and prevent the absorption of Pb from the GI tract (Basha et al. 2003). Studies on the ability of metals to displace Zn from MTs indicated that Cd had the highest capacity to displace Zn from hepatic MT, followed by Pb, Cu, and Hg. Differently, As had a limited ability to displace Zn from MT, while Al, Fe, and Mn had no effect on Zn binding (Walkess et al. 1984).
Overall, the simultaneous exposure of Pb to As, Mn, and/or Hg induces increased accumulation of Pb in the brain, while in a different way when Pb is in the presence of Zn and/or Cu, its accumulation decreases.
-
Mechanisms of Toxicity
In the same way as other metals, Pb may exert its toxic action by multi-mechanisms. Emphasis should be given to OS, since it is an important convergent point on the mechanisms of metal toxicity (Whittaker et al. 2010) representing a pathway that leads to the destruction of cells, including neurons and vascular cells in the CNS (Chong et al. 2005). While OS is a major mechanism of Pb-induced toxicity (Flora et al. 2012), exposures to As (Jomova and Valko 2011), Al (Kumar and Gill 2014), Cd (Mendez-Armenta et al., Méndez-Armenta et al. 2003), and Hg (Kaur et al. 2006) are also associated with excessive ROS production. Additionally, Pb exposure can lead to dyshomeostasis of essential metals in the brain, such as Mn (Pohl et al. 2011), Fe (Dai et al. 2013), Cu (Bradberry 2016), and Zn (McCord and Aizenman 2014). The dyshomeostasis of these metals is also known to induce OS (Pohl et al. 2011; Dai et al. 2013; Bradberry 2016; (McCord and Aizenman 2014) and has been associated with the induction of neurotoxicity.
It is expected that co-exposure to mixtures of Pb with other nonessential metals results in increased generation of ROS and/or decreases in the levels of antioxidants in the brain. Indeed, previous in vivo assays concerning exposure to metals’ mixtures representative of groundwater contamination in different parts of India involved the administration of a mixture of metals that included Pb, As, Cd, Mn, and Fe; their concentrations were equal to their WHO maximum permissible limit (MPL). In this work, increased LPO and decreased GSH level and activities of antioxidants in the brain (Jadhav et al. 2007) were observed. Similarly, the in vivo exposure to Pb, Hg, and Cd at the MPLs for each metal stipulated in the National Standard of The Republic of China for Municipal Water Standards resulted in a significant reduction in the activities of the brain antioxidant enzymes SOD and catalase and increases in malondialdehyde which is a marker of LPO (Cobbina et al. 2015). Elevated levels of brain nitric oxide synthase, an indicator of nitric oxide (NO), also suggested the induction of nitrosative stress (NS); NS is a parallel process, similar to OS but with involvement of ROS, known to create major toxicities to the brain and already associated with several neurodegenerative conditions (Klandorf and Van Dyke 2012, Cobbina et al. 2015). Still considering OS, in vitro works showed that astrocytes treated with Pb, As, and Cd triggered ROS generation, resulting in apoptosis of the mixture-treated astrocytes greater than would have been predicted by the individual metal treatment. Other toxicological evidences exist indicating that Mn interacts with Pb and Cd greater than additively, increasing the formation of ROS (von Stackelberg et al. 2013).
Another mechanism through Pb exerts CNS toxicity is the disruption of the normal physiological balance of trace metals in the brain, and essential metals may become compartmentally toxic by accumulation at levels that exceed the normal metal-buffering capacity within the cell (Zhou et al. 2014).
Cumulative evidences have implied that an imbalanced Fe, Cu, and Zn homeostasis in the CNS correlates with the pathogenesis of neurodegenerative disorders (Zheng et al. 2014; Molina et al. 2011; Szewczyk 2013). Actually, Pb has the ability to substitute other bivalent cations like Fe (Flora et al. 2012) and provoke Fe elevation in the brain tissue of Pb-exposed rodents. It is proposed that Pb influences cellular Fe influx or efflux, but changes in brain Fe levels might not be the result of an effect on DMT1 (Zhou et al. 2014) and rather on ferroportin 1 (FP1). This transporter might directly contribute to Fe efflux from neurons when overexpressed, thus preventing cellular Fe accumulation; Pb exposure might downregulate FP1 expression inducing cellular Fe accumulation in cells (Zhou et al. 2014). In agreement, Pb-induced increased Fe content in the old-aged rats’ brain might be the result of the decrease of the expression of FP1. Furthermore, the effect of Pb on FP1 expression is regulated at transcriptional and posttranscriptional levels. Overall the perturbation in Fe homeostasis may contribute to the neurotoxic consequences induced by Pb exposure, and FP1 may play a role in Pb-induced Fe accumulation in the brain (Zhu et al. 2013).
Concerning Cu, Pb can induce Cu accumulation in brain tissue as it is reported in in vitro and in vivo studies. Excess Cu concentration is detrimental to cells due to free Cu capability to form toxic free radicals, resulting in OS in neuronal cells. Metals such as Pb (and also Cd and Hg) can affect SH integrity within the MT metal-binding sites. It was hypothesized that exposure of MT to transition metals such as Cu, in combination with certain heavy metals with higher affinity for MT (Pb, Cd, and Hg), could lead to a release of transition metals and, hence, potentiate metal-dependent OS. Interestingly, there might be a bifunctional role of MT in protecting against and enhancing Cu- dependent OS depending upon exposure to Cu/heavy metal (Fabisiak et al. 1999). Still respecting to Cu, it is proposed that Pb upregulates the expression of the Cu transporter CTR1, which plays a major role in cellular Cu uptake and is abundant in the BBB, BCB, and brain parenchyma. Concurrently, Pb might downregulate the expression of P-type Cu-transporting ATPase (ATP7A), which is a major component of the intracellular Cu homeostasis apparatus. A consequent reduction of Cu efflux from the cells may thereby result in disturbed Cu homeostasis in the brain (Zheng et al. 2014). Less information is available on Pb-induced Zn dyshomeostasis. Even so, a large percentage of behavior-disordered persons exhibit an imbalance in levels of Cu and Zn in urine, blood, and other tissues, being suggested that Pb-induced imbalance in Zn (as well as Cu) may mediate insidious neurotoxic effect of Pb (Ademuyiwa et al. 2010).
-
Effects in the Hematopoietic System
Pb directly affects the hematopoietic system through reduction of the life span of circulating erythrocytes, by increasing the fragility of cell membranes. This metal can also restrain the synthesis of Hb, by inhibiting various key enzymes involved in the heme synthesis pathway. The combined aftermath of these two processes leads to anemia (Flora et al. 2012). Eight enzymes catalyze the reactions leading to heme synthesis (Hift et al., 2011; Kauppinen 2005), and most importantly, those enzymes have been shown to be specifically susceptible to impairment by a variety of toxic agents (Bleiberg et al. 1967; Quintanilla-Vega et al. 1996), including metals other than Pb. Blood ALAD and Zn protoporphyrin have been demonstrated to be sensitive for metal interactions at low doses (Wang and Fowler 2008). When interferences occur with the enzymes of the heme biosynthesis, there is characteristically an excessive accumulation and excretion of ALA and/or porphyrins (Adhikari et al. 2006; Guolo et al. 1996). Because individual porphyrins differ by the side-chain substituents, different metals may induce specific and different changes in porphyrin excretion patterns (Woods et al. 2009). Therefore, co-exposure of Pb with other metals with the capability to interfere with this metabolic pathway may certainly result in other characteristic urinary porphyrin profiles.
Indeed, Pb/As co-exposure can lead to synergistic inhibition of blood ALAD as compared to a single exposure either to Pb or As (Wang and Fowler 2008), while an additive effect on coproporphyrin excretion, but without alteration on ALA or uroporphyrin excretion, was already noticed (Fowler and Mahaffey 1978; Mahaffey et al. 1981). By its turn (in sequence), it is reported that while Cd itself do not markedly alter urinary excretion of porphyrins, simultaneous treatment with Pb and Cd increases in the urinary excretion of these heme precursors. In addition, the decrease in urinary ALA excretion observed after concomitant administration of Pb and Cd may result from Cd inhibition of the formation of active metabolites of vitamin D, which appear to play a role in Pb absorption (Fowler and Mahaffey 1978).
When the metals Pb, As, and Cd are administrated as a ternary mixture, interactions also occur, with additive increases in the urinary excretion of porphyrins, along with greater blood Zn protoporphyrin levels, than those observed with single exposures (Fowler and Mahaffey 1978). Another study established that exposure to the same mixture increases ALA, Fe, and Cu levels in vivo. The authors also observed that increases in ALA were followed by statistically significant increases in kidney Cu (Cobbina et al. 2015). Increased RBCs were also noted after administration of Pb or Cd (or As), and more cells were observed when the three metals were concomitantly administered. Despite increased numbers of circulating RBCs, Hb and hematocrit were reduced, especially with the Pb–Cd combination (Mahaffey and Fowler 1977). It was speculated that both Cd and Pb could affect Hb through interference with Fe metabolism, which is an essential element for Hb production. Cd can induce anemia by competing with Fe absorption, and in the same way, Pb is taken up by the Fe absorption machinery secondarily blocking Fe through competitive inhibition (Kwong et al. 2004).
Similarly to Pb, the mechanism by which excess Al induces anemia seems to be a reversible block in heme synthesis, due either to a defect in porphyrin synthesis or to impaired Fe utilization. The exposure to Al can induce an increase in the percentage of uroporphyrin and a decrease in coproporphyrin in urine (Nasiadek et al. 2001). However, Al can have an inhibitory effect in vitro, whereas in in vivo studies it activates the enzyme; Pb and Al together depress the enzyme activity in an additive way that can be reactivated by the addition of Zn (Abdulla et al. 1979). In line, Hg body burden was already shown to correlate with changes in urinary porphyrin profiles and with specific neurobehavioral deficits (Woods 1996). However these outcomes and the lack of studies pertaining to eventual Pb interactions with Hg on the heme synthesis pathway reveal the pertinence of studies on this matter. It is also known that Mn interferes with Fe homeostasis (Molina et al. 2011) and can in vitro inhibit FECH (Hift et al., 2011). This enzyme catalyzes the terminal step in the biosynthesis of heme, converting protoporphyrin IX into heme B through the insertion of Fe (Korolnek and Hamza 2014). More recently, it was demonstrated that Mn induces modifications in the excretion of porphyrins by increasing total porphyrins and modifying their profile. Results obtained after co-exposure to Pb, As, and Mn were suggestive that combined effects of the three metals resulted in higher heme synthesis disorders than those induced by exposure to each one of the metals alone (Andrade et al. 2014a). However, there is a lack of works clarifying the eventual interactions between Pb and Mn at this level.
Both Pb poisoning and Fe deficiency are capable of independently producing anemia. In the erythrocyte, both conditions affect the same cellular processes resulting in similar outcomes, with evidence for a synergistic relationship. During anemia, Pb becomes even more concentrated in RBC, seeming reasonable that the increased toxicity to erythrocytes during Fe deficiency anemia and Pb poisoning is partly due to an increased concentration of Pb in these cells (Kwong et al. 2004). Combined effects of Pb and Fe on heme synthesis are also described; when the pathway is inhibited at the final step, Zn instead of Fe is incorporated into protoporphyrin resulting in elevated levels of Zn protoporphyrin, which is a measure of heme synthesis inhibition. Indeed Fe deficiency and Pb poisoning are each one capable of inhibiting heme synthesis at this final step, and accordingly the concentrations of Zn protoporphyrin are dramatically higher in the presence of both conditions than in either Fe deficiency or Pb poisoning alone (Kwong et al. 2004). Furthermore, FECH is especially sensitive to low Fe levels in the presence of Pb poisoning. Actually Pb accumulates at mitochondria where apparently interferes with transport of Fe, the site of FECH. Concomitantly, while the levels of urinary ALA are usually not affected by Fe deficiency, they are elevated in Pb poisoning due to Pb’s inhibition of ALAD. The simultaneous occurrence of Fe deficiency and elevated Pb levels result in an almost twofold increase in urinary ALA, than with the presence of elevated Pb levels alone (Kwong et al. 2004). Pertaining to Pb interactions with Zn, orally exposed children benefitted of a protective effect of Zn on the hematopoietic effects of Pb, in the same way as demonstrated by in vivo works, even with Pb at higher doses. There are mechanistic evidences that Zn excess protects and reactivates Pb-inhibited ALAD, which is a Zn-containing enzyme (Pohl et al. 2011). Actually Zn may induce proteins that sequester Pb and donate Zn to ALAD (Roneya and Colman 2004). Indeed, supplemental Zn can protect against the Pb-induced increases in Zn protoporphyrin and urinary ALA excretion. However, these protective effects were seen at higher but not lower Pb doses and when basal levels of Zn in the diet were adequate (Roneya and Colman 2004). Additionally, effects of the mixed exposure to Pb, Zn, and Cu on workers’ erythrocytes include inhibition of ALAD and increased free erythrocyte protoporphyrin in comparison with values in unexposed population; these effects are characteristic of Pb. Thus, characteristic hematological effects of Pb were seen in workers co-exposed to low levels of Zn and Cu. However, whether or not Zn and Cu afforded some protection against these effects cannot be determined from this study (ATSDR 2004).
Interestingly, associations exist among Pb poisoning, hematological disarrays, and neurotoxicity. Pb’s inhibition of heme synthesis and disruption of protoporphyrin utilization appear to have a role in the demyelination of neuronal axons and the development of peripheral neuropathy, which is not unusual in adult’s Pb intoxication. Pb-intoxicated individuals have a high concentration of blood protoporphyrins and excrete an excess of protoporphyrins that are thought to be precursors of a substance essential for myelin maintenance (Kwong et al. 2004). Moreover, ALA which accumulates due to Pb exposure resembles GABA, and stimulation of GABA receptors is thought to be an important mechanism of Pb-induced neurotoxicity (Bradberry 2016).
-
Neurotoxic Effects
Neurotoxic effects are considered among the most potentially important health endpoints in epidemiological studies of complex mixtures, most particularly when such mixtures contain trace metals (Shy 1993). Not many reports are found respecting specifically to the neurotoxic effects induced by mixtures of metals. Even so, the mixture of As and Pb already showed to induce neuropsychological effects in children living in Morales (Mexico) despite that no conclusive results were obtained regarding interactions between these elements in this work (Carrizales et al. 2006). In turn, studies with rodents showed that while Pb alone increased movement and vertical activity and Cd alone decreased movement and increased rest time, Cd antagonized Pb-induced effects in rats co-exposed to both metals (Pohl et al. 2011). A synergistic effect from a mixture of Pb, As, and Cd on the developmental neurotoxicity was also observed with respect to glial and neuronal functions (Zhu et al. 2014). Other studies with the ternary mixture of the same metals revealed reductions in myelin thickness and axon density of the optic nerve and a decrease in thickness of nerve fiber, plexiform layer, and retinal ganglion cell counts in the retina (Cobbina et al. 2015). Concerning Hg, exposure to low Pb and Hg doses for 60 days can affect the learning and memory abilities of mice, evidenced by marginally high escape latency and low swimming speeds in the Morris water maze test. Similar trends were observed after exposure to the mixture Pb/Hg/Cd (Cobbina et al. 2015). In addition, animal studies provide compelling evidence that exposure to both Pb and Mn leads to synergistic neurological effects (Wright and Baccarelli 2007), with references to adversely affect cognitive function in an additive manner (Pohl et al. 2011). Decreased learning of conditioned avoidance responses to either Pb or Mn alone is described, and also gestational exposure to the same mixture reduces brain weight to a greater extent than either each metal alone (Wright and Baccarelli 2007). Despite an unaware of clinical data investigating joint exposures to Pb and Mn, Mexican children co-exposed to Pb and Mn had greater negative effects on cognitive performance than children exposed only to Pb (Neal and Guilarte 2013). Cognitive deficits were shown to be associated not only with Fe deficiency but also with blood Pb concentration in children, and it was proposed that Fe deficiency may affect verbal IQ directly and/or via increased blood Pb concentration due to Fe deficiency (Jeong et al. 2015). With respect to Zn, it is reported a protective effect of this metal against inhibition of smooth muscle contractility by Pb. Nevertheless, some authors consider that these studies do not provide evidence of potentiation, but rather a protective effect, or no effect, of Zn on Pb neurotoxicity (Pohl et al. 2011). Based on animal studies and mechanistic understanding, the predicted direction of joint toxic action for neurological effects is less than additive for the effects of Zn on Pb, and a similar direction is reported for Cu (Roneya and Colman 2004). A series of studies on Japanese workers exposed to Pb, Zn, and Cu at a gun metal foundry revealed that Zn and Cu may antagonize some of the neurological effects of Pb in occupationally exposed adults. Indirect evidence of potential antagonism by Zn and Cu or by Zn in Pb inhibition of peripheral nerve conduction velocity was found in some of these studies, but not in others. Other information from this series of studies include indirect evidence of potential antagonism by Zn and Cu of Pb inhibition of central nerve conduction and potential antagonism by Zn of Pb inhibition of autonomic nervous function (ATSDR 2004).
References
Abboud P, Wilkinson KJ. Role of metal mixtures (Ca, Cu and Pb) on Cd bioaccumulation and phytochelatin production by Chlamydomonas reinhardtii. Environ Pollut. 2013;179:33–8.
Abdulla M, Svensson S, Haeger-Aronsen B. Antagonistic effects of zinc and aluminum on lead inhibition of delta-aminolevulinic acid dehydratase.Arch. Environ Health. 1979;34(6):464–9.
Abreo K, Glass J, Sella M. Aluminum inhibits hemoglobin synthesis but enhances iron uptake in friend erythroleukemia cells. Kidney Int. 1990;37:677–81.
Ademuyiwa A, Agarwal R, Chandra R, Behari JR. Effects of sub-chronic low-level lead exposure on the homeostasis of copper and zinc in rat tissues. J Trace Elem Med Biol. 2010;24:207–11.
Adhikari A, Penatti CAA, Resende RR, Ulrich H, Britto LRG, Bechara EJH. 5-Aminolevulinate and 4, 5-dioxovalerate ions decrease GABAA receptor density in neuronal cells, synaptosomes and rat brain. Brain Res. 2006;1093:95–104.
Adriano DC. Trace elements in terrestrial environments. New York: Eds. Springer; 2001. 867p.
Agency for Toxic Substances and Disease Registry. Supplementary guidance for conducting health risk assessment of chemical mixtures, Risk Assessment Forum U.S. Atlanta: U.S. Department of Health and Human Services, Public Health Service; 2000.
Agency for Toxic Substances and Disease Registry. Priority list of hazardous substances. Atlanta: U.S. Department of Health and Human Services, Public Health Service. 2016. https://www.atsdr.cdc.gov/spl/, 5th September 2016, 5 pm.
Agency for Toxic Substances and Disease Registry (ATSDR). Interaction profile for: lead, manganese, zinc and copper. Atlanta: U.S. Department of Health and Human Services, Public Health Service; 2004.
Agency for Toxic Substances and Disease Registry (ATSDR). Arsenic CAS# 7440–38-2. Atlanta, GA: U.S. Department of Health and Human Services, Public Health Service; 2007.
Agency for Toxic Substances and Disease Registry (ATSDR). Toxicological profile for Cadmium. Atlanta: U.S. Department of Health and Human Services, Public Health Service; 2012.
Ali MM, Alia ML, Islam S, Rahman Z. Preliminary assessment of heavy metals in water and sediment of Karnaphuli River. Bangladesh Environ Earth Sci. 2016;73:1837–48.
Al-Saleh I, Nester M, Abduljabbar M, Al-Rouqi R, Eltabache C, Al-Rajudi T, Elkhati R. Mercury (Hg) exposure and its effects on Saudi breastfed infant’s neurodevelopment. Int J Hyg Environ Health. 2016;219:129–41.
Anderson D. Factors contributing to biomarker responses in exposed workers. Mutat Res. 1999;428:197–202.
Andrade V, Mateus ML, Batoréu MC, Aschner M, Marreilha dos Santos AP. Changes in rat urinary porphyrin profiles predict the magnitude of the neurotoxic effects induced by a mixture of lead, arsenic and manganese. Neurotoxicology. 2014a;45:168–77.
Andrade V, Mateus ML, Santos D, Aschner M, Batoréu MC, Marreilha dos Santos AP. Arsenic and manganese alter lead deposition in the rat. Biol Trace Elem Res. 2014b;158(3):384–91.
Angelovicová L, Fazekasová D. Contamination of the soil and water environment by heavy metals in the former mining area of Rudňany (Slovakia). Soil Water Res. 2014;9(1):18–24.
Annangi B, Bonassi S, Marcos R, Hernández A. Biomonitoring of humans exposed to arsenic, chromium, nickel, vanadium, and complex mixtures of metals by using the micronucleus test in lymphocytes. Mutat Res. 2016;770(Pt A):140–61.
Antonio MT, López N and Leret ML. Pb and Cd poisoning during development alters cerebellar and striatal function in rats. Toxicology. 2002; 176: 59–66.
Appel MJ, Kuper CF, Woutersen RA. Disposition, accumulation and toxicity of iron fed as iron (II) sulfate or as sodium iron EDTA in rats. Food Chem Toxicol. 2001;39:261–9.
Atamna H, Killile DK, Killile NB, Ames BN. Heme deficiency may be a factor in the mitochondrial and neuronal decay of aging. Proc Natl Acad Sci U S A. 2002;99(23):14807–12.
Atwood CS, Scarpa RC, Huang X, Moir RD, Jones WD, Fairlie DP, Tanzi R, Bush AI. Characterization of copper interactions with Alzheimer amyloid b peptides: identification of an Attomolar-affinity copper binding site on amyloid b1-42. J Neurochem. 2000;43(2):560–8.
Ballatori N. Transport of toxic metals by molecular mimicry. Environ Health Perspect. 2002;110(5):689–94.
Banks WA, Kastin AJ. The aluminum-induced increase in blood-brain barrier permeability to delta-sleep-inducing peptide occurs throughout the brain and is independent of phosphorus and acetylcholinesterase levels. Psychopharmacology. 1985;86(1–2):84–9.
Basha R, Wei W, Brydie M, Razmiafshari M, Zawia NH. Lead-induced developmental perturbations in hippocampal Sp1 DNA-binding are prevented by zinc supplementation: in vivo evidence for Pb and Zn competition. Int J Devl Neuroscience. 2003;21:1–12.
Bazzoni GB, Bollini AN, Hernandez GN, Contini MC, Chiarotto MM, Rasia ML. In vivo effect of aluminium upon the physical properties of the erythrocyte membrane. J Inorg Biochem. 2005;99:822–7.
Becaria A, Campbell A, Bondy SC. Aluminum and copper interact in the promotion of oxidative but not in? Ammatory events: implications for Alzheimer’s disease. J Alzheimers Dis. 2003;5:31–8.
Bleiberg J, Wallen M, Brodkin R, Applebaum IL. Industrially acquired porphyria. Arch Dermatol. 1967;80:793–7.
Bondier JR, Michel G, Propper A. Harmful effects of cadmium on olfactory system in mice. Inhal Toxicol. 2008;20(13):1169–77.
Bowers MA, Aicher LD, Davis HA, Woods JS. Quantitative determination of porphyrins in rat and human urine and evaluation of urinary porphyrin profiles during mercury and lead exposures. J Lab Clin Med. 1992;120:272–81.
Bradberry SM. Metals (cobalt, copper, lead, mercury). Medicine. 2016;44(3):182–4.
Brenneman KA, Wong BA, Buccellato MA, Costa ER, Gross EA, Dorman DC. Direct olfactory transport of inhaled manganese (54MnCl2) to the rat brain: toxicokinetic investigations in a unilateral nasal occlusion model. Toxicol Appl Pharmacol. 2000;169:238–48.
Buchta M, Kiesswetter E, Schaper M, Zschiesche W, Schaller KH, Kuhlmann A, Letzel S. Neurotoxicity of exposures to aluminium welding fumes in the truck trailer construction industry. Environ Toxicol Pharmacol. 2005;19:677–85.
Calderon J, Ortiz-Perez D, Yanez L, Díaz-Barriga F. Human exposure to metals. Pathways of exposure, biomarkers of effect, and host factors. Ecotoxicol Environ Saf. 2003;56:93–103.
Carrizales L, Razoa I, Tellez-Hernandez J, Torres-Nerioa R, Torres A, Batres LE, Cubillas A-C, Díaz-Barriga F. Exposure to arsenic and lead of children living near a copper-smelter in San Luis Potosi, Mexico: Importance of soil contamination for exposure. Environ Res. 2006;101:1–10.
Casarett & Doull’s. Toxicology: the basic science of poisons. 8th ed. London: McGraw-Hill; 2013.
Ceccatelli S, Daréb E, Moors M. Methylmercury-induced neurotoxicity and apoptosis. Chem Biol Interact. 2010;188:301–8.
Chong ZZ, Li F, Maiese K. Oxidative stress in the brain: novel cellular targets that govern survival during neurodegenerative disease. Prog Neurobiol. 2005;75:207–46.
Christensen JM. Human exposure to toxic metals: factors influencing interpretation of biomonitoring results. Sci Total Environ. 1995;166:89–135.
Clarkson TW, Vyas JB, Ballatori N. Mechanisms of mercury disposition in the body. Am J Ind Med. 2007;50(10):757–64.
Cobbina SJ, Chen Y, Zhou Z, Wub X, Feng W, Wang W, Mao G, Xu H, Zhang Z, Wua X, Yang L. Low concentration toxic metal mixture interactions: effects on essential and non-essential metals in brain, liver, and kidneys of mice on sub-chronic exposure. Chemosphere. 2015;132:79–86.
Colomina MT, Roig JL, Sánchez DJ, Domingo JL. Influence of age on aluminum-induced neurobehavioral effects and morphological changes in rat brain. Neurotoxicology. 2002;23(6):775–81.
Costa LG. Biochemical and molecular neurotoxicology: relevance to biomarker development, neurotoxicity testing and risk assessment. Toxicol Lett. 1998;102-103:417–21.
Costa LG, Manzo L. Biochemical markers of neurotoxicity: research epidemiological applications. Toxicol Lett. 1995;77(1–3):137–44.
Csavina J, Field J, Taylor MP, Gao S, Landázuri A, Betterton EA, Sáez AE. A review on the importance of metals and metalloids in atmospheric dust and aerosol from mining operations. Sci Total Environ. 2012;433:58–73.
Dai M-C, Zhong Z-H, Sun Y-H, Sun Q-F, Wang Y-T, Yang G-Y, Bian L-G. Curcumin protects against iron induced neurotoxicity in primary cortical neurons by attenuating necroptosis. Neurosci Lett. 2013;536:41–6.
Davies KM, Hare DJ, Cottam V, Chen N, Hilgers L, Halliday G, et al. Localization of copper and copper transporters in the human brain. Metallomics. 2013;5:43–51.
Desi I, Nagymajtenyi L and Schulz H. Behavioural and neurotoxicological changes caused by cadmium treatment of rats during development. J Appl Toxicol. 1998; 18(1), 63–70, 1998.
Dringen R. Metabolism and functions of glutathione in brain. Prog Neurobiol. 2000;62:649–71.
Dudzik CG, Walter ED, Abrams BS, Jurica MS, Millhauser GL. Coordination of copper to the membrane-bound form of alpha-synuclein. Biochemistry. 2012;52:53–60.
Dusek P, Roosc PM, Litwin T, Schneider SA, Flaten TP, Aaseth J. The neurotoxicity of iron, copper and manganese in Parkinson’s and Wilson’s diseases. J Trace Elem Med Biol. 2015;31:193–203.
Ebert BL, Bunn HF. Regulation of the erythropoietin gene. Blood. 1999;94:1864–77.
Ekino S, Susa M, Ninomiya T, Imamura K, Kitamura T. Minamata disease revisited: an update on the acute and chronic manifestations of methyl mercury poisoning. J Neur Sci. 2007;262:131–44.
Emerit J, Edeas M, Bricaire F. Neurodegenerative diseases and oxidative stress. Biomed Pharmacother. 2004;58:39–46.
Espinoza A, Le Blanc S, Olivares M, Pizarro F, Ruz M, Arredondo M. Iron, copper, and zinc transport: inhibition of divalent metal transporter 1 (DMT1) and human copper transporter 1 (hCTR1) by shRNA. Biol Trace Elem Res. 2012;146(2):281–6.
Exley C. Aluminum and Alzheimer’s disease. J Alzheimers Dis. 2001;3(6):551–2.
Fabisiak JP, Pearce LL, Borisenko GG, Tyhurina YY, Tyurin VA, Razzack J, Lazo JS, Pitt BR, Kagan VE. Bifunctional anti/prooxidant potential of metallothionein: redox signaling of copper binding and release. Antioxid Redox Signal. 1999;1:349–64.
Fairbrother A, Wenste R, Sappington K, Wood W. Framework for metals risk assessment. Ecotoxicol Environ Saf. 2007;68:145–227.
Feron VJ, Groten JP, Jonker D, Cassee FR, van Bladeren PJ. Toxicology of chemical mixtures: challenges for today and the future. Toxicology. 1995;105:415–27.
Flora SJS. Arsenic-induced oxidative stress and its reversibility. Free Radic Biol Med. 2011;51:257–81.
Flora G, Gupta D, Tiwari A. Toxicity of lead: a review with recent updates. Iternterdis Toxicol. 2012;5(2):47–58.
Fowler BA, Mahaffey KR. Interactions among lead, cadmium, and arsenic in relation to porphyrin excretion patterns. Environ Health Perspect. 1978;25:87–90.
Freitas Fonseca M, De Souza Hacon S, Grandjean P, Choi AL, Rodrigues Bastos W. Iron status as a covariate in methylmercury-associated neurotoxicity risk. Chemosphere. 2014;100:89–96.
Garrick MD, Singleton S, Vargas F, Kuo HC, Zhao L, Knopfel M, Davidson T, Costa M, Paradkar P, Roth JA, Garrick LM. DMT1: which metals does it transport? Biol Res. 2006;39:79–85.
Ghorbel I, Maktouf S, Kallel C, Chaabouni SE, Boudawara T, Zeghal N. Disruption of erythrocyte antioxidant defense system, hematological parameters, induction of pro-inflammatory cytokines and DNA damage in liver of co-exposed rats to aluminium and acrylamide. Chem Biol Interact. 2015;236:31–40.
Gianutsos G, Seltzer MD, Saymeh R, Wu M-LW, Michel RG. Brain manganese accumulation following systemic administration of different forms. Arch Toxicol. 1985;57:272–5.
Gorsky JE, Dietz AA, Spencer H, Osis D. Metabolic balance of aluminum studied in six men. Clin Chem. 1979;25(10):1739–43.
Grandjean P, Herz KT. Methylmercury and brain development: imprecision and underestimation of developmental neurotoxicity in humans. Mt Sinai J Med. 2011;78(1):107–18.
Gu C, Chen S, Xu X, Zheng L, Li Y, Wu K, Liu J, Qi Z, Han D, Chen G, Huo X. Lead and cadmium synergistically enhance the expression of divalent metal transporter 1 protein in central nervous system of developing rats. Neurochem Res. 2009;34:1150–6.
Gunshin H, Mackenzie B, Berger UV, Gunshin Y, Romero MF, Boron WF, Nussberger S, Gollan JL, Hediger MA. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature. 1997;388:482–8.
Guolo M, Stella AM, Melito V, Parera V, Batle AMC. Altered 5-aminolevulinic acid metabolism leading to pseudophorphyria in hemodialysed patients. lnt J Biochem Cell Bid. 1996;28:311–7.
Heyer NJ, Bittner AC Jr, Echeverria D, Woods JS. A cascade analysis of the interaction of mercury and coproporphyrinogen oxidase (CPOX) polymorphism on the heme biosynthetic pathway and porphyrin production. Toxicol Lett. 2006;161:159–66.
Hift RJ, Thunell S, Brun A. Drugs in porphyria: from observation to a modern algorithm-based system for the prediction of porphyrogenicity. Pharmacol Ther. 2011;132(2):158–69.
Hoffmeyer RE, Singh SP, Doonan CJ, Ross ARS, Hughes RJ, Pickering IJ, George GN. Molecular mimicry in mercury toxicology. Chem Res Toxicol. 2006;19(6):753–9.
Horiguchi H, Teranishi H, Niiya K, Aoshima K, Katoh T, Sakuragawa N, Kasuya M. Hypoproduction of erythropoietin contributes to anemia in chronic cadmium intoxication: clinical study on Itai-itai disease in Japan. Arch Toxicol. 1994;68(10):632–6.
Horiguchi H, Aoshima K, Oguma R, Sasaki S, Miyamoto K, Hosoi Y, Katoh T, Kayama F. Latest status of cadmium accumulation and its effects on kidneys, bone, and erythropoiesis in inhabitants of the formerly cadmium-polluted Jinzu River Basin in Toyama, Japan, after restoration of rice paddies. Int Arch Occup Environ Health. 2010;83:953–70.
Hsu-Kim H, Kucharzyk KH, Zhang T, Deshusses MA. Mechanisms regulating mercury bioavailability for methylating microorganisms in the aquatic environment: a critical review. Environ Sci Technol. 2013;47(6):2441–56.
Huang P, Chen C, Wang H, Li G, Jing H, Han Y, Li N, Xiao Y, Yu Q, Liu Y, Wang P, Shi Z, Sun Z. Manganese effects in the liver following subacute or subchronic manganese chloride exposure in rats. Ecotoxicol Environ Saf. 2011;74:615–22.
Illing AC, Shawki A, Cunningham CL, Mackenzie B. Substrate profile and metal-ion selectivity of human divalent metal-ion transporter-1. J Biol Chem. 2012;287(36):30485–96.
Islam E, Yang X, He Z and Mahmood Q. Assessing potential dietary toxicity of heavy metals in selected vegetables and food crops. J Zhejiang Univ Sci B. 2007; 8(1): 1–13.
Jadhav S, Sarkar S, Patil R, Tripathi H. Effects of subchronic exposure via drinking water to a mixture of eight water-contaminating metals: a biochemical and histopathological study in male rats. Arch Environ Con Tox. 2007;53:667–77.
Järup L. Hazards of heavy metal contamination. Br Med Bull. 2003;68:167–82.
Jeong KS, Park H, Hac E, Hong Y-C, Hae M, Park H, Kimf B-N, Leeg SJ, Lee KY, Kim JH, Kim Y. Evidence that cognitive deficit in children is associated not only with iron deficiency, but also with blood lead concentration: a preliminary study. J Trace Elem Med Biol. 2015;29:336–41.
Jiang X, McDermott JR, Ajees AA, Rosen BP, Liu Z. Trivalent arsenicals and glucose use different translocation pathways in mammalian GLUT1. Metallomics. 2010;2(3):211–9.
Jin T, Lu J and Nordberg M.Toxicokinetics and biochemistry of cadmium with special emphasis on the role of metallothionein. Neurotoxicology. 1998; 19(4-5):529-35.
Jomova K, Valko M. Advances in metal-induced oxidative stress and human disease. Toxicology. 2011;283(2–3):65–87.
Julka D, Gill KD. Development of a possible peripheral marker for aluminum neurotoxicity. Med Sci Res. 1995;23:311–4.
Kakkar P, Jaffery FN. Biological markers for metal toxicity. Environ Toxicol Pharmacol. 2005;19:335–49.
Kalia K, Chandra SV, Viswanathan PN. Effect of 54Mn and lead interaction on their binding with tissue proteins: in vitro studies. Ind Health. 1984;22:207–18.
Kauppinen R. Porphyrias Lancet. 2005;365:241–52.
Kaur A, Joshi K, Minz RW, Gill KD. Neurofilament phosphorylation and disruption: a possible mechanism of chronic aluminium toxicity in Wistar rats. Toxicology. 2006;219(1–3):1–10.
Kawahara M, Kato-Negishi M. Link between aluminum and the pathogenesis of Alzheimer’s disease: the integration of the aluminum and amyloid Cascade hypotheses. Int J Alzheimers Dis. 2011;2011:276393.
Kerper LE, Ballatori N, Clarkson TW. Methylmercury transport across the blood-brain barrier by an amino acid carrier. Am J Phys. 1992;262:761–5.
Kile ML, Fara JM, Ronnenberg AG, Quamruzzaman Q, Rahman M, Mostofa G, Afroz S, Christiani DC. A cross sectional study of anemia and iron deficiency as risk factors for arsenic-induced skin lesions in Bangladeshi women. BMC Public Health. 2016;16:158.
Kim S, Moon C, Eun S, Ryu P, Jo S. Identification of ASK1, MKK4, JNK, c-Jun, and caspase-3 as a signaling cascade involved in cadmium-induced neuronal cell apoptosis. Biochem Biophys Res Commun. 2005;328:326–34.
Kim Y, Kim BN, Hong Y-C, Shin M-S, Yoo H-J, Kim J-W, Bhang S-Y, Cho S-C. Co-exposure to environmental lead and manganese affects the intelligence of school-aged children. Neurotoxicology. 2009;30:564–71.
Kim JH, Lee SJ, Kim SY, Choi G, Lee JJ, Kim HJ, Kim S, Park J, Moon HB, Choi K, Kim S, Choi SR. Association of food consumption during pregnancy with mercury and lead levels in cord blood. Sci Total Environ. 2016a;29(563–564):118–24.
Kim Y, Oh HG, Cho YY, Kwon O-H, Park MK, Chung S. Stress hormone potentiates Zn2+-induced neurotoxicity via TRPM7 channel in dopaminergic neuron. Biochem Biophys Res Commun. 2016b;470:362–7.
Klandorf H, Van Dyke K. Oxidative and nitrosative stresses: their role in health and disease in man and birds. Oxidative stress – molecular mechanisms and biological effects. (Chapter 3). Ed. Volodymyr Lushchak and Halyna M. Semchyshyn. 2012. ISBN 978-953-51-0554-1, Published: April 25, 2012 under CC BY 3.0 license.
Klatzo I, Wisniewski H, Streicher E. Experimental production of neurofibrillary degeneration: 1. Light microscopic observations. J Neuropathol Exp Neurol. 1965;24:187–99.
Klauder DS, Petering HG. Protective value of dietary copper and iron against some toxic effects of lead in rats. Environ Health Perspect. 1975;12:77–80.
Kordas K, Queirolo EI, Ettinger AS, Wright RO, Stoltzfus RJ. Prevalence and predictors of exposure to multiple metals in preschool children from Montevideo. Uruguay Sci Total Environ. 2010;408:4488–94.
Korolnek T, Hamza I. Like iron in the blood of the people: the requirement for heme trafficking in iron metabolism. Front Pharmacol. 2014;4;5,126.
Kortenkamp and Faust. State of the art report on mixture toxicity – final report. UE Comission. 2009. http://ec.europa.eu/environment/chemicals/effects/pdf/report_mixture_toxicity.pdf, 13th June 2014, 2 p.m.
Kozlowski H, Janicka-Klos A, Brasun J, Gaggelli E, Valensin D, Valensin G. Copper, iron, and zinc ions homeostasis and their role in neurodegenerative disorders (metal uptake, transport, distribution and regulation). Coord Chem Rev. 2009;253:2665–85.
Krüger K, Straub H, Hirner AV, Hippler J, Binding N, Musshoff U. Effects of monomethylarsonic and monomethylarsonous acid on evoked synaptic potentials in hippocampal slices of adult and young rats. Toxicol Appl Pharmacol. 2009;236(1):115–23.
Kumar V, Gill KD. Oxidative stress and mitochondrial dysfunction in aluminium neurotoxicity and its amelioration: a review. Neurotoxicology. 2014;41:154–66.
Kumar A, Dogra S, Prakash A. Protective effect of curcumin (Curcuma longa), against aluminium toxicity: possible behavioral and biochemical alterations in rats. Behav Brain Res. 2009;205(2):384–90.
Kwong WT, Friello P, Semba RD. Interactions between iron deficiency and lead poisoning: epidemiology and pathogenesis. Sci Total Environ. 2004;330:21–37.
Lafuente A, Esquifino A. Cadmium effects on hypothalamic activity and pituitary hormone secretion in the male. Toxicol Lett. 1999;110(3):209–18.
Landrigan P, Nordberg M, Lucchini R, Nordberg G, Grandjean P, Iregren A, Alessio L. The declaration of Brescia on prevention of the neurotoxicity of metals. Am J Ind Med. 2006;50(10):709–11.
Lieu PT, Heiskala M, Peterson PA, Yang Y. The roles of iron in health and disease. Mol Asp Med. 2001;22:1–87.
Lin C-Y, Hsiao W-C, Huang C-J, Kao C-F, Hsua G-S-W. Heme oxygenase-1 induction by the ROS-JNK pathway plays a role in aluminum-induced anemia. J Inorg Biochem. 2013;128:221–8.
Lister LJ, Svendsen C, Wright J, Hooper HL, Spurgeon DJ. Modelling the joint effects of a metal and a pesticide on reproduction and toxicokinetics in Lumbricid earthworms. Environ Int. 2011;37:663–70.
Liu J, Klaassen CD. Absorption and distribution of cadmium in Metallothionein-I transgenic mice. Fund Appl Toxicol. 1996;29:294–300.
Liu Z, Sanchez MA, Jiang X, Boles E, Landfear SM, Rosen BP. Mammalian glucose permease GLUT1 facilitates transport of arsenic trioxide and methylarsonous acid. Biochem Biophys Res Commun. 2006;351(2):424–30.
Lohren H, Pieper I, Blagojevic L, Bornhorst J, Galla H-J, Schwerdtle T. Neurotoxicity of organic and inorganic mercury species – effects on and transfer across the blood-cerebrospinal fluid barrier, cytotoxic effects in target cells. Perspect Sci. 2015;3:21–2.
López Alonso M, Prieto Montaña F, Miranda M, Castillo C, Hernández J, Benedito JL. Cadmium and lead accumulation in cattle in NW Spain. Vet Hum Toxicol. 2003;45(3):128–30.
Lopez E, Arce C, Oset-Gasque MJ, Canadas S, González MP. Cadmium induces reactive oxygen species generation and lipid peroxidation in cortical neurons in culture. Free Rad Biol Med. 2006;40:940–51.
Lorincz MT. Neurologic Wilson’s disease. Ann N Y Acad Sci. 2010;1184:173–87.
Lovell MA, Robertson JD, Teesdale WJ, Campbell JL, Markesbery WR. Copper, iron and zinc in Alzheimer’s disease senile plaques. J Neurol Sci. 1998;158(1):47–52.
Lu J, Zheng Y-L, Wu D-M, Sun D-X, Shan Q, Fan S-H. Trace amounts of copper induce neurotoxicity in the cholesterol-fed mice through apoptosis. FEBS Lett. 2006;580:6730–40.
Lucchini R, Zimmerman N. Lifetime cumulative exposure as a threat for neurodegeneration: need for prevention strategies on a global scale. Neurotoxicology. 2009;30(6):1144–8.
Mahaffey KR, Fowler BA. Effects of concurrent Administration of Lead, cadmium, and arsenic in the rat. Environ Health Perspect. 1977;19:165–71.
Mahaffey KR, Capar SG, Gladen BC, Fowler BA. Concurrent exposure to lead, cadmium, and arsenic. Effects on toxicity and tissue metal concentrations in the rat. J Lab Clin Med. 1981;98(4):463–81.
Maines MD. Regional distribution of the enzymes of haem biosynthesis and the inhibition of 5-aminolaevulinate synthase by manganese in the rat brain. Biochem J. 1980;190:315–21.
Markiewicz-Górka I, Januszewska L, Michalak A, Prokopowicz A, Januszewska E, Pawlas N, Pawlas K. Effects of chronic exposure to lead, cadmium, and manganese mixtures on oxidative stress in rat liver and heart. Arh Hig Rada Toksikol. 2015;66(1):51–62.
Martinez-Finley EJ, Chakraborty S, Fretham SJB, Aschner M. Cellular transport and homeostasis of essential and nonessential metals. Metallomics. 2012;4(7):593–605.
McCord MC, Aizenman E. The role of intracellular zinc release in aging, oxidative stress, and Alzheimer’s disease. Front Aging Neurosci. 2014;6:77.
Mejía JJ, Diáz-Barriga F, Calderón J, Ríos C, Jiménez-Capdeville ME. Effects of lead-arsenic combined exposure on central Monoaminergic systems. Neurotoxicol Teratol. 1997;19(6):489–97.
Méndez-Armenta M, Ríos C. Cadmium neurotoxicity. Environ Toxicol Pharmacol. 2007;23(3):350–8.
Méndez-Armenta M, Villeda-Hernández J, Barroso-Moguel R, Nava-Ruiz C, Jiménez-Capdeville ME, Rios C. Brain regional lipid peroxidation and metallothionein levels of developing rats exposed to cadmium and dexamethasone. Toxicol Lett. 2003;144(2):151–7.
Migliore L, Coppedè F. Environmental-induced oxidative stress in neurodegenerative disorders and aging. Mut Res. 2009;674:73–84.
Miu AC, Andreescu CE, Vasiu R, Olteanu AI. A behavioral and histological study of the effects of long-term exposure of adult rats to aluminum. Int J Neurosci. 2003;113(9):1197–211.
Molina RM, Phattanarudee S, Kim J, Thompson K, Wessling-Resnick M, Maher TJ, Brain JD. Ingestion of Mn and Pb by rats during and after pregnancy alters iron metabolism and behavior in offspring. Neurotoxicology. 2011;32(4):413–22.
Monroe RK, Halvorsen SW. Cadmium blocks receptor-mediated Jak/STAT signaling in neurons by oxidative stress. Free Radic Biol Med. 2006;41(3):493–502.
Mutti A. Biological monitoring in occupational and environmental toxicology. Toxicol Lett. 1999;108:77–89.
Nasiadek M, Chmielnicka J, Subdys J. Analysis of urinary Porphyrins in rats exposed to aluminum and iron. Ecotoxicol Environ Saf. 2001;48(1):11–7.
Naujokas MF, Anderson B, Ahsan H, Aposhian HV, Graziano JH, Thompson C, Suk WA. The broad scope of health effects from chronic arsenic exposure: update on a worldwide public health problem. Environ Health Perspect. 2013;121(3):295–302.
Nazir R, Shaheen N, Shah MH. Indoor/outdoor relationship of trace metals in the atmospheric particulate matter of an industrial area. Atmos Res. 2011;101:765–72.
Neal AP, Guilarte TR. Mechanisms of heavy metal neurotoxicity: lead and manganese. Toxicol Res (Camb). 2013;(2):99–114.
Nehru B, Anand P. Oxidative damage following chronic aluminium exposure in adult and pup rat brains. J Trace Elem Med Biol. 2005;19(2–3):203–8.
Nordberg M, Nordberg GF. Metallothioneins: historical development and overview. Met Ions Life Sci. 2009;5:1–29.
Notarachille G, Arnesano F, Calò V, Meleleo D. Heavy metals toxicity: effect of cadmium ions on amyloid beta protein 1-42. Possible implications for Alzheimer’s disease. Biometals. 2014;27(2):371–88.
Nriagu J. Zinc toxicity in humans. School of Public Health, University of Michigan, Elsevier B.V. 2007.
O’Neil P. Heavy metals in soils. In: Alloway BJ, editor. Arsenic. London: Blackie Academic and Professional Arsenic; 1995. p. 105–21.
Obiri S, Yeboah PO, Osae S, Adu-Kumi S. Levels of arsenic, mercury, cadmium, copper, lead, zinc and manganese in serum and whole blood of resident adults from mining and non-mining communities in Ghana. Environ Sci Pollut Res Int. 2016;23(16):16589–97.
Oteiza PI, Keen CL, Han B, Golub MS. Aluminum accumulation and neurotoxicity in Swiss-Webster mice after long-term dietary exposure to aluminum and citrate. Metabolism. 1993;42(10):1296–300.
Park JD, Zheng W. Human exposure and health effects of inorganic and elemental mercury. J Prev Med Public Health. 2012;45(6):344–52.
Patrick L. Lead toxicity, a review of the literature. Part I: exposure, evaluation, and treatment. Altern Med Rev. 2006;11(1):2–22.
Peakall D, Burger J. Methodologies for assessing exposure to metals: speciation, bioavailability of metals, and ecological host factors. Ecotoxicol Environ Saf. 2003;56(1):110–21.
Piao F, Cheng F, Chen H, Li G, Sun X, Liu S, Yamauchi T, Yokoyama K. Effects of Zn administration on Pb toxicities in rats. Ind Health. 2007;45:546–51.
Pirpamer L, Hofer E, Gesierich B, De Guio F, Freudenberger P, Seiler S, Duering M, Jouvent E, Duchesnay E, Dichgans M, Ropele S, Schmidt R. Determinants of iron accumulation in the normal aging brain. Neurobiol Aging. 2016;43:149–55.
Pohl HR, Hansen H, Chou C-HSJ. Public health guidance values for chemical mixtures: current practice and future directions. Regul Toxicol Pharmacol. 1997;26:322–9.
Pohl HR, Roney N, Abadin HG. Metal ions affecting the neurological system. Met Ions Life Sci. 2011;8:247–62.
Prüss-Ustün A, Wolf J, Corvalán C, Bos R, Neira M. Preventing disease through healthy environments. A global assessment of the burden of disease from environmental risks. Geneva: World Health Organization (WHO); 2016.
Qato MK, Maines MD. Regulation of heme and drug metabolism activities in the brain by manganese. Biochem Biophys Res Commun. 1985;128(1):18–24.
Quintanar L. Manganese neurotoxicity: a bioinorganic chemist’s perspective. Inorg Chim Acta. 2008;361:875–84.
Quintanilla-Vega B, Hernandez A, Mendoza-Figueroa T. Reduction in porphyrin excretion as a sensitive indicator of lead toxicity in primary cultures of adult rat hepatocytes. Toxicol In Vitro. 1996;10:675–83.
Rachakonda V, Pan TH, Le WD. Biomarkers of neurodegenerative disorders: how good are they? Cell Res. 2004;14(5):349–60.
Ramos-Chávez LA, Rendón-López CRR, Zepeda A, Silva-Adaya D, Del Razo LM, Gonsebatt ME. Neurological effects of inorganic arsenic exposure: altered cysteine/glutamate transport, NMDA expression and spatial memory impairment. Front Cell Neurosci. 2015;9:9–21.
Rodríguez-Barranco M, Lacasaña M, Aguilar-Garduño C, Alguacil J, Gil F, González-Alzaga B, Rojas-García A. Association of arsenic, cadmium and manganese exposure with neurodevelopment and behavioural disorders in children: a systematic review and meta-analysis. Sci Total Environ. 2013;1(454–455):562–77.
Roels HA, Bowler RM, Kim Y, Claus Henn B, Mergler D, Hoet P, Gocheva VV, Bellinger DC, Wright RO, Harris MG, Chang Y, Bouchard MF, Riojas-Rodriguez H, Menezes-Filho JA, Tellez-Rojo MM. Manganese exposure and cognitive deficits: a growing concern for manganese neurotoxicity. Neurotoxicology. 2012;33:872–80.
Roneya N, Colman J. Interaction profile for lead, manganese, zinc, and copper. Environ Toxicol Pharmacol. 2004;18:231–4.
Rosado JL, Ronquillo D, Kordas K, Rojas O, Alatorre J, Lopez P, Garcia-Vargas G, Caamaño MC, Cebrián ME, Stoltzfus RJ. Arsenic exposure and cognitive performance in Mexican schoolchildren. Environ Health Perspect. 2007;115(9):1371–5.
Rubio-Osornio M, Montes S, Heras-Romero Y, Guevara J, Rubio C, Aguilera P, Rivera-Mancia S, Floriano-Sánchez E, Monroy-Noyola A, Ríos C. Induction of ferroxidase enzymatic activity by copper reduces MPP+−evoked neurotoxicity in rats. Neurosci Res. 2013;75:250–5.
Rush T, Hjelmhaug J, Lobner D. Effects of chelators on mercury, iron, and lead neurotoxicity in cortical culture. Neurotoxicology. 2009;30:47–51.
Sahin G, Varol I, Temizer A, Benli K, Demirdamar R, Duru S. Determination of aluminum levels in the kidney, liver, and brain of mice treated with aluminum hydroxide. Biol Trace Elem Res. 1994;41(1–2):129–35.
Sánchez-Peña LC, Petrosyan P, Morales M, González NB, Gutiérrez-Ospina G, Del Razo LM, Gonsebatt ME. Arsenic species, AS3MT amount, and AS3MT gene expression in different brain regions of mouse exposed to arsenite. Environ Res. 2010;110(5):428–34.
Santamaria AB. Manganese exposure, essentiality & toxicity. Indian J Med Res. 2008;128:484–500.
Scheiber IF, Mercer JFB, Dringen R. Metabolism and functions of copper in brain. Prog Neurobiol. 2014;116:33–57.
Schneider SA. Neurodegenerations with brain iron accumulation. Parkinsonism Rel Disord. 2016;22:21–5.
Sensi SL, Jeng JM. Rethinking the excitotoxic ionic milieu: the emerging role of Zn(2+) in ischemic neuronal injury. Curr Mol Med. 2004;4(2):87–111.
Sensi SL, Yin HZ, Weiss JH. AMPA/kainate receptor-triggered Zn2+ entry into cortical neurons induces mitochondrial Zn2+ uptake and persistent mitochondrial dysfunction. Eur J Neurosci. 2000;12:3813–8.
Sethi P, Jyoti A, Hussain E, Sharma D. Curcumin attenuates aluminium-induced functional neurotoxicity in rats. Pharmacol Biochem Behav. 2009;93:31–9.
Shi LZ, Zheng W. Early lead exposure increases the leakage of the blood–cerebrospinal fluid barrier, in vitro. Hum Exp Toxicol. 2007;26(3):159–67.
Shukla GS, Hussain T, Chandra SV. Possible role of regional superoxide dismutase activity and lipid peroxide levels in cadmium neurotoxicity: in vivo and in vitro studies in growing rats. Life Sci. 1987;41(19):2215–21.
Shukla GS and Chandra SV. Concurrent exposure to lead, manganese, and cadmium and their distribution to various brain regions, liver, kidney, and testis of growing rats. Archives of Environ Contam Toxicol. 1987; 16(3):303–310.
Shukla A, Shukla GS, Srimal RC. Cadmium-induced alterations in blood-brain barrier permeability and its possible correlation with decreased microvessel antioxidant potential in rat. Hum Exp Toxicol. 1996;15(5):400–5.
Shy CM. Epidemiological studies of neurotoxic, reproductive, and carcinogenic effects of complex mixtures. Environ Health Perspect. 1993;101(4):183–8.
Simmons JE. Chemical mixtures: challenge for toxicology and risk assessment. Toxicology. 1995;105:11–9.
Simmons-Willis TA, Koh AS, Clarkson TW, Ballatori N. Transport of a neurotoxicant by molecular mimicry : the methylmercury-L-cysteine complex is a substrate for human L-type large neutral amino acid transporter (LAT) 1 and LAT2. Biochem J. 2002;367:239–46.
Sinczuk-Walczaki H, Szymczak M, Razniewska G, Matczak W, Szymczak W. Effects of occupational exposure to aluminium on nervous system: clinical and electroencephalographic findings. Int J Occup Med Environ Health. 2003;16(4):301–10.
Singh T, Goel RK. Neuroprotective effect of Allium cepa L. in aluminium chloride induced neurotoxicity. Neurotoxicology. 2015;49:1–7.
Skjørringe T, Burkhart A, Johnsen KB, Moos T. Divalent metal transporter 1 (DMT1) in the brain: implications for a role in iron transport at the blood-brain barrier, and neuronal and glial pathology. Front Mol Neurosci. 2015;8(19):1–13.
Song H, Zheng G, Liu Y, Shen X-F, Zhao Z-H, Aschner M, Luo W-J, Chen J-Y. Cellular uptake of lead in the blood-cerebrospinal fluid barrier: novel roles of Connexin 43 hemichannel and its down-regulations via Erk phosphorylation. Toxicol Appl Pharmacol. 2016;15(297):1–11.
Spurgeon DJ, Jones OAH, Dorne J-L, Svendsen C, Swain S, Stürzenbaum SR. Systems toxicology approaches for understanding the joint effects of environmental chemical mixtures. Sci Total Environ. 2010;408:3725–34.
Strak E, Ellinger I, Balthasar C, Scheinast M, Schatz J, Szattler T, Bleichert S, Saleh L, Knöfler M, Zeisler H, Hengstschläger M, Rosner M, Salzer H, Gundacker C. Mercury toxicokinetics of the healthy human term placenta involve amino acid transporters and ABC transporters. Toxicology. 2016;340:34–42.
Struys-Ponsar C, Kerkhofs A, Gauthier A, Soffié M, van den Bosch de Aguilar P. Effects of aluminum exposure on behavioral parameters in the rat. Pharmacol Biochem Behav. 1997;56(4):643–8.
Szewczyk B. Zinc homeostasis and neurodegenerative disorders. Front Aging Neurosci. 2013;5:33.
Tallkvist J, Bowlus CL, Lonnerdal B. DMT1 gene expression and cadmium absorption in human absorptive enterocytes. Toxicol Lett. 2001;122:171–7.
Tiffany-Castiglioni E, Hong S, Qian Y, Tang Y, Donnelly KC. In vitro models for assessing neurotoxicity of mixtures. Neurotoxicology. 2006;27:835–9.
Tjälve H, Henriksson J. Uptake of metals in the brain via olfactory pathways. Neurotoxicology. 1999;20(2–3):181–95.
Tougu V, Tiiman A, Palumaa P. Interactions of Zn(II) and cu(II) ions with Alzheimer’s amyloid-beta peptide metal ion binding, contribution to fibrillization and toxicity. Metallomics. 2011;3:250–61.
Vazquez M, Velez D, Devesa V, Puig S. Participation of divalent cation transporter DMT1 in the uptake of inorganic mercury. Toxicology. 2015;331:119–24.
von Stackelberg K. Guzy E, Chu T, Henn BC. Mixtures, metals, genes and pathways: a systematic review. Working paper prepared for: methods for research synthesis: a cross-disciplinary workshop. Harvard Center for Risk Analysis. 2013.
Wagner GS, Tephly TR. A possible role of copper in the regulation of heme biosynthesis through ferrochelatase. Adv Exp Med Biol. 1975;58:343–54.
Walton JR. Aluminum disruption of calcium homeostasis and signal transduction resembles change that occurs in aging and Alzheimer’s disease. J Alzheimers Dis. 2012;29(2):255–73.
Waalkes MP, Harvey MJ, Klaassen CD. Relative in vitro affinity of hepatic metallothionein for metals. Toxicol Lett. 1984; 20(1):33-9.
Wang G, Fowler BA. Roles of biomarkers in evaluating interactions among mixtures of lead, cadmium and arsenic. Toxicol Appl Pharmacol. 2008;233:92–9.
Wang Q, Luo W, Zheng W, Liu Y, Xu H, Zheng G, Dai Z, Zhang W, Chen Y, Chen J. Iron supplement prevents lead-induced disruption of the blood–brain barrier during rat development. Toxicol Appl Pharmacol. 2007;219(1):33–41.
Wang Q, Luo W, Zhang W, Liu M, Song H, Chen J. Involvement of DMT1 +IRE in the transport of lead in an in vitro BBB model. Toxicol In Vitro. 2011;25:991–8.
Wang L, Wang X, Zhang S, Qu G, Liu S. A protective role of heme-regulated eIF2a kinase in cadmium-induced toxicity in erythroid cells. Food Chem Toxicol. 2013;62:880–91.
Ward RJ, Zucca FA, Duyn JH, Crichton RR, Zecca L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014;13(10):1045–60.
Watanabe T, Hirano S. Metabolism of arsenic and its toxicological relevance. Arch Toxicol. 2013;87(6):969–79.
Weidenhamer JD, Lobunski PA, Kuepouo G, Corbin RW, Gottesfeld P. Lead exposure from aluminum cookware in Cameroon. Sci Total Environ. 2014;496:339–47.
Weiss B. Economic implications of manganese neurotoxicity. Neurotoxicology. 2006;27:362–8.
Wenting L, Ping L, Haitao J, Meng Q, Xiaofei R. Therapeutic effect of taurine against aluminum-induced impairment on learning, memory and brain neurotransmitters in rats. Neurol Sci. 2014;35(10):1579–84.
Wester RC, Maibach HI, Sedik L, Melendres J, DiZio S, Wade M. In vitro percutaneous absorption of cadmium from water and soil into human skin. Fund Appl Toxicol. 1992;19(1):1–5.
Whittaker MH, Wang G, Chen X-Q, Lipsky M, Smith D, Gwiazda R, Fowler BA. Exposure to Pb, Cd, and As mixtures potentiates the production of oxidative stress precursors: 30-day, 90-day, and 180-day drinking water studies in rats. Toxicol Appl Pharmacol. 2010;254(2):154–66.
Wills MR, Hewitt CD, Sturgill BC, et al. Long-term oral or intravenous aluminum administration in rabbits I. Renal and hepatic changes. Ann Clin Lab Sci. 1993;23(1):1–16.
Witholt R, Gwiazda RH, Smith DR. The neurobehavioral effects of subchronic manganese exposure in the presence and absence of pre-parkinsonism. Neurotoxicol Teratol. 2000;22:851–61.
Woods JS. Porphyrin metabolism as indicator of metal exposure and toxicity. In: Goyer RA, Cherian MG, editors. Handbook of experimental pharmacology. Vol. 115. Chap. 2. Toxicology of metals, biochemical aspects. Berlin: Springer; 1995. p. 19–52.
Woods JS. Altered porphyrin metabolism as a biomarker of mercury exposure and toxicity. Can J Physiol Pharmacol. 1996;74:210–5.
Woods JS, Southern MR. Studies on the etiology of trace metal-induced porphyria: effect of porphyrinogenic metals on coproporphyrinogen oxidase in rat liver and kidney. Toxicol Appl Pharmacol. 1989;97:183–90.
Woods JS, Eaton DL, Lukens CB. Studies on porphyrin metabolism in the kidney. Effects of trace metals and glutathione of renal uroporphyrinogen decarboxylase. Mol Pharmacol. 1984;26:336–41.
Woods JS, Bowers MA, Davis HA. Urinary porphyrin profiles as biomarkers of trace metal exposure and toxicity: studies on urinary porphyrin excretion patterns in rats during prolonged exposure to methyl mercury. Toxicol Appl Pharmacol. 1991;110:464–76.
Woods JS, Martin MD, Leroux BG, DeRouen TA, Bernardo MF, Luis HS, Leitão JG, Simmonds PL, Rue TC. Urinary porphyrin excretion in normal children and adolescents. Clin Chim Acta. 2009;405:104–9.
Wright RO, Baccarelli A. Metals and neurotoxicology. J Nutr. 2007;137(12):2809–13.
Yang XF, Han QG, Liu DY, Fan GY, Ma JY, Wang ZL. Microstructure and ultrastructure alterations in the pallium of immature mice exposed to cadmium. Biol Trace Elem Res. 2016;1–7.
Yasui M, Kihira T, Ota K. Calcium, magnesium and aluminum concentrations in Parkinson’s disease. Neurotoxicology. 1992;13:593–600.
Yasuno T, Okamoto H, Nagai M, Kimura S, Yamamoto T, Nagano K, Furubayashi T, Yoshikawa Y, Yasui H, Katsumi H, Sakane T, Yamamoto A. The disposition and intestinal absorption of zinc in rats. Eur J Pharm Sci. 2011;44:410–5.
Yen Le TT, Vijver MG, Kinraide TB, Peijnenburg SWJGM, Hendriks AJ. Modelling metal interactions and metal toxicity to lettuce Lactuca sativa following mixture exposure (Cu2+-Zn2+ and Cu2+-Ag+). Environ Pollut. 2013;176:185–92.
Yokel RA. Blood-brain barrier flux of aluminum, manganese, iron and other metals suspected to contribute to metal-induced neurodegeneration. J Alzheimers Dis. 2006;10:223–53.
Zheng W. Toxicology of choroid plexus: special reference to metal-induced neurotoxicities. Microsc Res Tech. 2001;52(1):89–103.
Zheng W, Monnot AD. Regulation of brain iron and copper homeostasis by brain barrier systems: implication in neurodegenerative diseases. Pharmacol Ther. 2012;133(2):177–88.
Zheng W, Perry DF, Nelson DL, Aposhian HV. Protection of cerebrospinal fluid against toxic metals by the choroid plexus. FASEB J. 1991;5:2188–93.
Zheng W, Aschner M, Ghersi-Egeac J-F. Brain barrier systems: a new frontier in metal neurotoxicological research. Toxicol Appl Pharmacol. 2003;192:1–11.
Zheng G, Zhang J, Xuc Y, Shen X, Song H, Jing J, Luo W, Zheng W, Chen J. Involvement of CTR1 and ATP7A in lead (Pb)-induced copper (Cu)accumulation in choroidal epithelial cells. Toxicol Lett. 2014;225:110–8.
Zhi D, Tao AIJ, Fang HJ, Sun RB, Shi Y, Wang LL, Wang Q. Influence of iron supplementation on DMT1 (IRE)-induced transport of lead by brain barrier systems in vivo. Biomed Environ Sci. 2015;28(9):651–9.
Zhou F, Chen Y, Fan G, Feng C, Dub G, Zhu G, Li Y, Jiao H, Guan L, Wang Z. Lead-induced iron overload and attenuated effects of ferroportin 1 overexpression in PC12 cells. Toxicol In Vitro. 2014;28:1339–48.
Zhu L, Ji X-J, Wang H-D, Pan H, Chen M, Lu T-J. Zinc neurotoxicity to hippocampal neurons in vitro induces ubiquitin conjugation that requires p38 activation. Brain Res. 2012;1438:1–7.
Zhu G, Fan G, Feng C, Li Y, Chen Y, Zhou F, Du G, Jiao H, Liu Z, Xiao X, Lin F, Yand J. The effect of lead exposure on brain iron homeostasis and the expression of DMT1/FP1 in the brain in developing and aged rats. Toxicol Lett. 2013;216:108–23.
Zhu H, Jia Y, Cao H, Meng F, Liu X. Biochemical and histopathological effects of subchronic oral exposure of rats to a mixture of five toxic elements. Food Chem Toxicol. 2014;71:166–75.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Andrade, V.M., Aschner, M., Marreilha dos Santos, A.P. (2017). Neurotoxicity of Metal Mixtures. In: Aschner, M., Costa, L. (eds) Neurotoxicity of Metals. Advances in Neurobiology, vol 18. Springer, Cham. https://doi.org/10.1007/978-3-319-60189-2_12
Download citation
DOI: https://doi.org/10.1007/978-3-319-60189-2_12
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-60188-5
Online ISBN: 978-3-319-60189-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)