
Abstract
Electron paramagnetic resonance (EPR) spin-label oximetry allows the oxygen permeability coefficient to be evaluated across homogeneous lipid bilayer membranes and, in some cases, across coexisting membrane domains without their physical separation. The most pronounced effect on oxygen permeability is observed for cholesterol, which additionally induces the formation of membrane domains. In intact biological membranes, integral proteins induce the formation of boundary and trapped lipid domains with a low oxygen permeability. The effective oxygen permeability coefficient across the intact biological membrane is affected not only by the oxygen permeability coefficients evaluated for each lipid domain but also by the surface area occupied by these domains in the membrane. All these factors observed in fiber cell plasma membranes of clear human eye lenses are reviewed here.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others
Keywords
1 Introduction
The most fundamental function of biological membranes is the creation of barriers between intracellular and extracellular environments as well as between different cellular compartments. This is largely due to the hydrophobicity of the membrane interior. Water soluble components such as ions and sugars can cross biological membranes through channels or transporters. Nonelectrolytes , like molecular oxygen, can cross the lipid bilayer membrane component by passive diffusion, permeating directly through the bilayer. The oxygen flux across the lipid bilayer (J) is proportional to the difference in oxygen concentration in water on either side of the membrane (C ″ − C ′) and the membrane permeability coefficient (P M):
Here, P M describes bulk membrane properties without detailed analysis of mechanisms involved in membrane permeation. This equation implies that P M can be easily measured using a stop-flow rapid-mixing approach to create an oxygen gradient. These studies have been criticized, however, because the presence of a thick (~2 μm) unmixed water layer prevents immediate contact of oxygenated water with the membrane. It was determined that these methods can be used only for solutes with P M < 10−2 cm/s [1]. Fortunately, Diamond and Katz [2] derived an expression for the permeability coefficient of nonelectrolyte solutes across the lipid bilayer:
The oxygen partition coefficient K(x) and the oxygen diffusion coefficient D(x) are allowed to vary across the bilayer. The integration is taken across the hydrophobic portion of the membrane. Resistances r’ and r” are, conceptually, connected to transport across the head-group region [3]. Thus, the knowledge of profiles of the oxygen diffusion-concentration product across the hydrophobic region of the lipid bilayer allows evaluation the oxygen permeability coefficient across that region.
2 Methods
Electron paramagnetic resonance (EPR) spin-label oximetry approaches allowed the oxygen diffusion-concentration product in any site which can be spin labeled or spin probed by nitroxide spin labels to be obtained. Based on measurements of spin-lattice relaxation time T 1 using the saturation-recovery EPR technique, Kusumi et al. [4] defined an oxygen transport parameter as:
Since W(x) is proportional to the collision rate of oxygen with the nitroxide spin label group, it is a function of both the concentration C(x) and the translational diffusion coefficient D(x) of oxygen at depth x in the air-equilibrated membrane,
Using different lipid spin labels, with nitroxide groups located at different depths in membranes, profiles of W(x), and, thus, profiles of D(x)C(x) across membranes, can be easily obtained (see [5] for an evaluation of A). Combining the theory developed by Diamond and Katz [3] and spin-label oximetry methods , Subczynski et al. [6] developed a procedure to calculate P M based on the profiles of W(x) across the membrane for samples equilibrated with air. Since the nitroxide moiety of the lipid spin labels can be placed in the headgroup region as well as in the hydrophobic region, r’ and r” in Eq. (5.2) can also be estimated within the integral. For samples equilibrated with air, the local oxygen concentration C(x) and the oxygen concentration in the water phase C w (air) are related by the equation:
and Eq. (5.2) becomes:
Thus, P M can be evaluated in terms of experimental observable W(x) and C w (air) can be taken from published tables. Profiles of W(x) are obtained for samples equilibrated with air without the creation of oxygen gradients across the membrane.
3 Results
The EPR spin-labeling method turned out to be very powerful for oximetry measurements, allowing measurements of P M across homogeneous lipid bilayers and across coexisting membrane domains in model membranes made of commercially available phospholipids and total lipids isolated from biological membranes. It was also successfully applied to evaluate P Ms across lipid domains induced by membrane integral proteins in intact membranes. Our recent research is focused on the permeability properties of the lipid bilayer portion of fiber cell plasma membranes of eye lenses. These membranes, with their unique lipid composition (extremely high cholesterol (Chol) content) and high load with integral membrane proteins can serve as a good illustration, showing how different factors can affect the oxygen permeability of biological membranes.
3.1 Model Lipid Bilayer Membranes
3.1.1 Effect of Acyl Chains Unsaturation
Incorporation of either a cis or trans double bond at the C9–C10 position of the acyl chain decreased the oxygen diffusion-concentration product at all locations (depths) in the phospholipid bilayers [5]. We think that the key feature of oxygen transport in the lipid bilayer is the small size of the molecule. Since molecular oxygen is small, its diffusion in the membrane must be quite different from that of lipids and other molecules of larger sizes. Oxygen transport in the membrane is closely related to the creation and movements of many small, vacant pockets due to rapid gauche-trans isomerization of acyl chains and conformational mismatch between lipid molecules. The presence of a rigid double bond would reduce the dynamics of the chain around the double bond. These data indicate that the dynamic gauche-trans isomerization of acyl chains plays an important role in oxygen diffusion in the membrane and are in agreement with the arguments advanced by Pace and Chan [7] on the relationship between the diffusion of small molecules in the membrane and kink migration and/or kink formation due to the acyl chain isomerization.
3.1.2 Effect of Cholesterol Intercalation
Based on our observations, we can conclude that the intercalation of Chol in saturated membranes decreases the oxygen diffusion-concentration product in the polar headgroup and hydrocarbon regions (to the depth to which the rigid ring structure of Chol is immersed), but little affects this product in the membrane center [6]. In unsaturated membranes, intercalation of Chol also decreases the oxygen diffusion-concentration product in and near the polar headgroup region but increases this product in the membrane center [5]. Since the major barrier for oxygen permeation is located in and near the polar headgroup region, the presence of Chol decreases total membrane permeability for oxygen despite the increase in the oxygen diffusion-concentration product in the bilayer center. Interestingly, a high cholesterol content is responsible for creating hydrophobic channels for oxygen transport parallel to the membrane surface, concurrently, is responsible for creating the rigidity barrier to oxygen transport across the membrane [8].
3.1.3 Cholesterol-Induced Phases and Domains
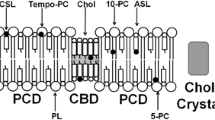
When investigating effects of Chol on oxygen transport across lipid bilayer membranes, one must to take into account the membrane phases and domains presented in membranes at different Chol contents. Membranes made of dimyristoylphosphatidylcholine (DMPC ) and Chol form one of the simplest paradigms for the study of formation, coexistance, and separation of phases and domains. As shown in the phase diagram presented by Almeida et al. [9] and extended by Mainali et al. [10], at a physiological temperature, the liquid-disordered phase can adopt up to ~10 mol% Chol. This is a homogenous phase with properties (that is, the profile of the oxygen diffusion-concentration product) that change smoothly up to 10 mol% Chol. Above this concentration, a new liquid-ordered phase is formed with a Chol concentration of ~30 mol%. Thus, for Chol contents ranging between 10–30 mol%, these two phases coexist with very different oxygen diffusion-concentration product profiles [11]. The liquid-ordered phase can adopt ~30–50 mol% Chol. Increased Chol concentration drastically changed the profile of the oxygen diffusion-concentration product, which is bell-shaped at 30 mol% and rectangular-shaped (with abrupt increase of three–four times between C9 and C10) at 50 mol% Chol. Because the liquid-ordered phase of DMPC cannot adopt more than 50 mol% Chol, at higher contents, the excess of Chol forms the pure Chol bilayer domains (CBDs) surrounded by the phospholipid bilayer saturated with Chol [10]. At a Chol content above 66 mol% Chol (which is the Chol solubility threshold for DMPC), these pure CBDs collapse, forming Chol crystals, presumably outside the lipid bilayer. The EPR spin-labeling approach allows estimation of the permeability coefficient for oxygen across the CBD with the use of only two Chol analogue spin labels; thus, based only on approximate profiles of the oxygen diffusion-concentration product [12]. These data indicate that CBDs most strongly affect the total oxygen permeability coefficient. We conclude that oxygen permeation through all these phases and domains should be considered when the total oxygen permeability across lipid bilayer membranes is evaluated.
3.1.4 Lens Lipid Membranes
Lens lipid membranes are formed from the total lipid extracts from different regions of eye lenses. The Chol content in the eye lens fiber-cell plasma membrane is extremely high. Chol saturates the bulk phospholipid bilayer and induces formation of CBDs within the membrane [13]. The saturating Chol content in fiber-cell membranes keeps the bulk physical properties of lens-lipid membranes (including profiles of the oxygen-diffusion concentration product) consistent and independent of changes in phospholipid composition [14]. The phospholipid composition of fiber-cell membranes changes significantly with age and between different regions of the lens. Surprisingly, independent of these differences, profiles of the oxygen diffusion-concentration product across phospholipid domains are very similar in all of the investigated membranes [15]. They have a rectangular shape with an abrupt increase in the oxygen diffusion-concentration product at the C9–C10 position. In the headgroups and hydrocarbon regions to the depth of C9, the product is as low as in gel-phase membranes. At locations deeper than C9, it is as high as in fluid-phase membranes. These profiles are typical for lipid bilayers saturated with Chol and are very different from the bell-shaped profile across membranes without Chol.
3.2 Intact Fiber Cell Plasma Membranes of Eye Lenses
Integral membrane proteins are practically impermeable to oxygen [16]. However, they affect (decrease) the oxygen permeability of the lipid bilayer, creating boundary lipids (around integral proteins) and trapped lipids (in protein-rich domains) for which oxygen transport is significantly lower [17]. In human lens membranes, more than 50% of lipid molecules are in contact with integral proteins.
3.2.1 Lipid Domains
Our most recent studies have focused on intact lens membranes isolated from human lenses [18]. We hypothesized that we should find the presence of the four purported lipid domains that are induced in the lipid bilayer portion because of the presence of integral membrane proteins, namely, bulk, boundary, and trapped lipids, as well as pure CBDs. We were able to confirm the existence of three of them: bulk, boundary, and trapped lipid domains [18].
3.2.2 Oxygen Permeability across Domains in Intact Fiber Cell Membranes
The EPR spin-labeling approach allowed us to evaluate P M across bulk plus boundary lipid domains, and across trapper lipids in both cortical and nuclear eye lens membranes [18]. Separation of results for bulk lipids and boundary lipids is not possible because the lipids exchange too quickly between these domains. Results obtained for clear lenses from donors from groups age 0–20, 21–40, 41–60, and 61–80 years [18] indicate that for all membrane domains in cortical and nuclear fiber cell membranes, the permeability for oxygen was significantly lower than across water layers of the same thickness as these domains. The lowering effect was mainly due to the presence of membrane proteins and the formation of boundary and trapped lipid domains. For all age groups investigated, the P M across the bulk plus boundary domain was always smaller (by 30%, on average) in nuclear than in cortical membranes. This difference was even greater for trapped lipids where the P M across trapped lipids in nuclear membranes, on average, was 45% smaller compared to that in cortical membranes. The permeability of the trapped lipid domain in cortical and nuclear membranes was ~4.7 and ~8.5 times smaller, respectively, then the permeability across water layers of the same thickness as the domain. Thus, the trapped lipid domain forms a major membrane barrier for oxygen transport into the lens center, and this barrier was significantly greater in the lens nucleus. However, for all investigated age groups, the values of the P M measured for corresponding membrane domains did not change significantly with age.
4 Conclusions
The effective P M across the lipid bilayer portion of the fiber cell plasma membrane is, in the first approximation, equal to the weighted sum of the P Ms evaluated for each lipid domain. The weight for each domain is proportional to the surface area occupied by the domain, divided by the total surface occupied by the lipid bilayer portion of the membrane. An EPR spin-labeling method has been developed that allows quantitative evaluation of the amounts of phospholipids and Chol (which determines the surface of the domain) in lipid domains of intact membranes [19]. Application of this method to fiber-cell plasma membranes isolated from cortical and nuclear regions of clear eye lenses indicates that the relative amounts of boundary and trapped phospholipids, and trapped Chol in nuclear membranes increase significantly with donor age. The amount of lipids in those domains in cortical membranes did not change significantly with age. The combination of results for oxygen permeability across lipid domains with those regarding the amount of lipids in domains allowed us to conclude that trapped lipids in the lipid bilayer portion of nuclear fiber cell membranes form a high barrier to oxygen permeation into the lens center, and that this barrier increases with age. In cortical fiber cell membranes, barriers formed by the lipid bilayer domains were significantly lower than in nuclear fiber cells and did not change with age, although these barriers were still considerably greater than across a water layer of the same thickness as domains. Proteins are nearly impermeable to oxygen [16]. Thus, the total effective P M across the intact membrane is equal to the oxygen permeability coefficient evaluated for the lipid bilayer portion of the membrane, multiplied by the factor proportional to the surface area of the lipid bilayer portion, divided by the surface area of the entire membrane.
References
Missner A, Pohl P (2009) 110 years of the meyer–Overton rule: predicting membrane permeability of gases and other small compounds. ChemPhysChem 10:1405–1414
Diamond JM, Katz Y (1974) Interpretation of nonelectrolyte partition coefficients between dimyristoyl lecithin and water. J Membr Biol 17:121–154
Dix JA, Kivelson D, Diamond JM (1978) Molecular motion of small nonelectrolyte molecules in lecithin bilayers. J Membr Biol 40:315–342
Kusumi A, Subczynski WK, Hyde JS (1982) Oxygen transport parameter in membranes as deduced by saturation recovery measurements of spin-lattice relaxation times of spin labels. Proc Natl Acad Sci U S A 79:1854–1858
Subczynski WK, Hyde JS, Kusumi A (1991) Effect of alkyl chain unsaturation and cholesterol intercalation on oxygen transport in membranes. Biochemistry 30:8578–8590
Subczynski WK, Hyde JS, Kusumi A (1989) Oxygen permeability of phosphatidylcholine-cholesterol membranes. Proc Natl Acad Sci U S A 86:4474–4478
Pace RJ, Chan SI (1982) Molecular motions in lipid bilayers. III. Lateral and transverse diffusion in bilayers. J Chem Phys 76:4241–4247
Subczynski WK, Wisniewska A (2000) Physical properties of lipid bilayer membranes: relevance to membrane biological functions. Acta Biochim Pol 47:613–625
Almeida PF, Vaz WL, Thompson TE (1992) Lateral diffusion in the liquid phases of dimyristoylphosphatidylcholine/cholesterol lipid bilayers: a free volume analysis. Biochemistry 31:6739–6747
Mainali L, Raguz M, Subczynski WK (2013) Formation of cholesterol bilayer domains precedes formation of cholesterol crystals in cholesterol/dimyristoylphosphatidylcholine membranes: EPR and DSC studies. J Phys Chem B 117:8994–9003
Subczynski WK, Wisniewska A, Hyde JS et al (2007) Three-dimensional dynamic structure of the liquid-ordered domain as examined by a EPR oxygen probing. Biophys J 92:1573–1584
Raguz M, Mainali L, Widomska J et al (2011) Using spin-label electron paramagnetic resonance (EPR) to discriminate and characterize the cholesterol bilayer domain. Chem Phys Lipids 164:819–829
Mainali L, Raguz M, O'Brien WJ et al (2013) Properties of membranes derived from the total lipids extracted from the human lens cortex and nucleus. Biochim Biophys Acta 1828:1432–1440
Subczynski WK, Raguz M, Widomska J et al (2012) Functions of cholesterol and the cholesterol bilayer domain specific to the fiber-cell plasma membrane of the eye lens. J Membr Biol 245:51–68
Mainali L, Raguz M, O’Brien WJ et al (2016) Changes in the properties and organization of human lens lipid membranes occurring with age. Curr Eye Res, DOI: 10.1080/02713683.2016.1231325
Altenbach C, Greenhalgh DA, Khorana HG et al (1994) A collision gradient method to determine the immersion depth of nitroxides in lipid bilayers: application to spin-labeled mutants of bacteriorhodopsin. Proc Natl Acad Sci U S A 91:1667–1671
Ashikawa I, Yin J-J, Subczynski WK et al (1994) Molecular organization and dynamics in bacteriorhodopsin-rich reconstituted membranes: discrimination of lipid environments by the oxygen transport parameter using a pulse ESR spin-labeling technique. Biochemistry 33:4947–4952
Raguz M, Mainali L, O'Brien WJ et al (2015) Lipid domains in intact fiber-cell plasma membranes isolated from cortical and nuclear regions of human eye lenses of donors from different age groups. Exp Eye Res 132:78–90
Raguz M, Mainali L, O'Brien WJ et al (2015) Amounts of phospholipids and cholesterol in lipid domains formed in intact lens membranes. Exp Eye Res 140:179–186
Acknowledgments
Supported by grants EY015526, EB002052, and EY001931 from the NIH.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Subczynski, W.K., Widomska, J., Mainali, L. (2017). Factors Determining the Oxygen Permeability of Biological Membranes: Oxygen Transport Across Eye Lens Fiber-Cell Plasma Membranes. In: Halpern, H., LaManna, J., Harrison, D., Epel, B. (eds) Oxygen Transport to Tissue XXXIX. Advances in Experimental Medicine and Biology, vol 977. Springer, Cham. https://doi.org/10.1007/978-3-319-55231-6_5
Download citation
DOI: https://doi.org/10.1007/978-3-319-55231-6_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-55229-3
Online ISBN: 978-3-319-55231-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)