
Abstract
Neuroimaging research in psychiatry has been increasing exponentially in recent years, yet many psychiatrists are relatively unfamiliar with this field. The neuroimaging findings summarized here include alterations related to fibromyalgia, chronic pain, and coping in somatoform pain disorders. Magnetic resonance imaging is the imaging method of choice for standard clinical sequences. Improvements in imaging technology now allow advanced sequences, once used exclusively for research, to be used clinically. Magnetic resonance spectroscopy (showing metabolism) offers invaluable information on living tissues, with a special contribution to the diagnosis and prognosis of diseases of the central nervous system. Voxel-based morphometry (structural information) is a recent technique that can simultaneously visualize group differences or statistical effects on gray and white matter throughout the brain. Perfusion (marker of vascularity) offers higher spatial resolution than radionuclide techniques such as positron emission tomography and single-photon emission computed tomography. Diffusion-weighted imaging (a marker of cellularity) detects subtle degradation of white matter microstructure in fibromyalgia. Diffusion tensor imaging shows integrity of surrounding white matter tracts. Functional magnetic resonance imaging is used to identify eloquent cortex.
These techniques, all of which have advanced our understanding of chronic pain and can be used to improve knowledge on the etiology of these illnesses, will be discussed.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
- Chronic pain
- Fibromyalgia
- Somatization
- Magnetic resonance imaging
- Magnetic resonance spectroscopy
- Structural brain imaging
- Perfusion
- Diffusion weighted imaging
- Diffusion tensor imaging
- Functional magnetic resonance imaging
- Treatment effects
Introduction
Chronic pain is a major public health problem. The prevalence of chronic pain in Western, industrialized countries is estimated to be between 15% and 20% of the adult population [1,2,3]. Therefore, there is a need to better understand the mechanisms that lead to chronic pain. From a neurobiological perspective, the mechanisms contributing to the transition from acute to subacute and chronic pain are heterogeneous, and are thought to occur both within the peripheral nervous system and at various levels of the central nervous system (CNS). The role of the brain in chronic pain states remains to be fully elucidated.
The World Health Organization classifies fibromyalgia syndrome (FMS) under the heading of diseases of the musculoskeletal system and connective tissue, as does the International Association for the Study of Pain [4].
Fibromyalgia (FM) is a chronic rheumatic disease characterized by the presence of diffuse musculoskeletal pain, painful sensitivity to touch in at least 11 of 18 defined trigger points, and a constellation of symptoms including fatigue, disturbed sleep, cognitive problems, and distress [5]. Furthermore, it is accepted as a central sensitivity syndrome. The American College of Rheumatology in 2010 described new diagnosis criteria [6]. The prevalence of this syndrome in Europe is approximately 2.9% [7], and the prevalence in rheumatology consultations in Spain was found to be 12% [8].
Pain is the most common and disabling symptom of FM. This pain is suspected to be caused by the altered function of structures in the CNS, including the primary and secondary sensory and motor cortices, insula, anterior cingulate cortex, thalamus, dorsolateral prefrontal cortex, and basal ganglia. If we look for analogies or parallels between these and other insults, we encounter the clinical characteristics of a number of neuropathic pain syndromes. There are similarities from the point of view of symptoms, diagnosis, and therapeutic approach. However, FMS has not demonstrated any injury or association with any known disease that affects the nervous system, and therefore one that could be considered the origin of a somatosensory disorder. Neuropathic pain is defined as a “pain caused by direct injury or disease affecting the somatosensory system” [9]. As in the case of FM, diagnosis of various neuropathic pain syndromes are performed based on clinical criteria.
Somatoform disorders (SFDs), according to the fourth edition of the Diagnostic and Statistical Manual of Mental Disorders (DSM-IV) [10], are defined by the presence of physical symptoms that suggest a general medical condition but that are not fully explained by a general medical condition, by the direct effects of a substance, or by another mental disorder. The most extreme form of this group is somatization disorder, a chronic and polysymptomatic disorder characterized by at least four unexplained gastrointestinal, sexual, and pseudo neurological symptoms [10].
The psychological profiles show the usual psychological characteristics of patients with an FM or SFD: high scores in anxiety and depression assessed with the Hospital Anxiety Depression Scale; high scores on the Pain Catastrophizing Scale and in pain assessed with the Pain Visual Analogue Scale, and low quality of life as measured by the EuroQol 5D. The Mini-Mental State Exam scores suggest symptoms of cognitive dysfunction in FM and SFDs, but at levels less severe than those found in patients with dementia [11].
Over the past decade, brain-imaging studies have shed light on neural correlates of pain perception and pain modulation, and they have also begun to disentangle the neural mechanisms that underlie chronic pain. There is now general agreement that the CNS plays a prominent role in many chronic pain states because of the “centralization” of pain. Some of the findings that strongly support this paradigm shift are outlined later.
The clinical diagnosis of FM does not exist today, given that there is no analytical testing, validated image, or specific pathological condition that serves to confirm the diagnosis. However, numerous publications describe neuroimaging abnormalities in this syndrome that affects the somatosensory system [12,13,14].
The quantitative study of anatomical and biochemical variables is important in the diagnosis, prognosis, and monitoring of different diseases [15]. The morphometry study of certain brain structures, complemented by the identification of variations of some metabolites by nuclear magnetic resonance spectroscopy, is a promising tool for the elucidation of the morphobiological component of different pathological conditions.
The study of low frequency signals in brain activity through blood oxygen level–dependent (BOLD) contrast at rest revealed synchronized cortical activity patterns, allowing the intrinsic functional architecture of the human brain to be described. The international scientific community has shared resources that will help with this analysis by means of functional magnetic resonance imaging (fMRI) at rest to obtain more accurate and advanced diagnoses in the field of neuroscience treatments.
This chapter seeks to review published scientific evidence and studies of structural and functional imaging in FMS that consider the similarities and dissimilarities between the findings in FMS and related findings in a number of neuropathic pain syndromes. The ultimate goal is to find evidence of valid indicators with which to objectify the diagnosis of these syndromes.
Magnetic Resonance Spectroscopy (MRS)
This technique enables us to study the chemical composition of living tissues, and is based on the chemical shift of atoms. The concentration of a number of metabolites is determined from spectra that may be acquired in several ways. Proton magnetic resonance spectroscopy (1H–MRS) is one of the techniques used to assess potential disruptions in neuronal integrity and associated neurochemical dysregulations. The most commonly used spectroscopy is that originating from a hydrogen nucleus (proton 1H–MRS). The position of the metabolite signal is identified on the horizontal axis by its chemical shift, scaled in units referred to as parts per million (ppm).
The brain spectrum shows peaks corresponding to the different metabolites: myo-inositol (mI), 3.56 and 4.06 ppm; choline compounds (Cho), 3.23 ppm; creatine (Cr), 3.03 and 3.94 ppm; N-acetylaspartate (NAA), 2.02; glutamine (Gln) and glutamate (Glu), 2.1–2.55 ppm and 3.8 ppm. NAA is considered a neuronal–axonal marker with a neuronal bioenergetic role [16,17,18] found in the brain and spinal cord. Recent studies, however, have indicated that NAA reflects functional rather than structural neuronal characteristics [19], suggesting that NAA is most informative in the investigation of functional abnormality. Cr is involved in energy metabolism through the Cr kinase reaction generating phosphocreatine, and in turn, adenosine triphosphate [20]. Cho containing compounds of glycerol 3-phosphocholine and phosphocholine, which are present at high levels in glial cells [21], are intermediaries in the synthesis of acetylcholine [22]. Glutamine and glutamate (Glx) are strongly compartmentalized (in neurons and in astrocytes respectively), and are directly connected to energy metabolism and neurotransmission [23].
Previous studies (see Table 30.1) described lower NAA levels within the dorsolateral prefrontal cortex of patients with chronic back pain when compared with healthy controls [24]. Lower NAA levels have also been reported in the thalamus of patients with neuropathic pain included [25, 26].
These decreases in NAA might possibly reflect a neuronal loss, indicating a neurodegenerative process to be associated with chronic pain. Hippocampal dysfunction in patients with FM has also been explored using 1H–MRS [27]. In this case control study, 15 patients and ten controls were examined, and levels and interhippocampal ratios of metabolites such as NAA, Cho, and Cr were assessed. Patients with FM had lower NAA levels than those found in controls, representing a neuronal or axonal metabolic dysfunction in the hippocampus. As the hippocampus plays a crucial role in the maintenance of cognitive functions, sleep regulation, and pain perception, the authors suggest that hippocampal metabolic dysfunction may be implicated in the symptomatology of this puzzling syndrome. Consequently, a decrease was found in the NAA/Cr ratio in the right hippocampus [28], decreased Cho and N-acetylaspartate + N-acetylaspartate glutamate (NAA + NAAG) in the left hippocampus, and also a decrease in both hippocampi of myo-inositol (mI), glutamate (Glu), the Cho/Cr ratio, and the mI/Cr ratio [11, 29]. On the other hand, an increase in NAA was found in both sides. Cho was increased on the right [27] and decreased on the left [29]. Our study confirms a significant reduction in Cho (both hippocampi ⁄ posterior cingulate cortex), mI (left hippocampus), NAA (left hippocampus ⁄ posterior insula), and Glu (left hippocampus) in both FMS and SFD groups compared with controls. Indeed, neurochemical changes that could be indicative of such damage have been reported previously [29,30,31].
These studies report a decrease in NAA in patients with chronic pain in the DLPFC and the thalamus respectively, two areas also involved in pain processing and perception. They attribute this loss to a neurodegenerative process present in chronic pain. Lower hippocampal and insular NAA levels suggest neuronal or axonal metabolic dysfunction, or a combination of these processes. We suggest that hippocampal dysfunction may be partly responsible for some of the phenomena associated with FM and somatization disorder. Blocking N-methyl-D-aspartate receptors (NMDAR) in the hippocampal formation reduces nociceptive behaviors; this reduction, in turn, supports the hypothesis that the hippocampal formation is involved in the pain-related neural processing and the expression of pain-related behaviors [32]. The default mode network (DMN) comprises a set of brain regions that are coactivated during passive task states, show an intrinsic functional relationship, and are connected via direct and indirect anatomic projections. In a previous study, we found elevated levels of Glu in the ventral posterior cingulate cortex, a key zone in the default mode network hypothesis [29]. We propose that high levels of Glu in certain regions of the brain [29] cause cellular damage and disruptions in circuits involved in the pain perception. This may be underlying the cognitive and behavioral impairments accompanying chronic pain. The chronic pain condition could cause a sustained lesion in the brain through Glu toxicity, and could explain the structural damage and significant atrophy seen in chronic pain patients.
Another remarkable fact is the correlation of all metabolites in the left hippocampus with pain and the correlation of choline in the posterior cingulate cortex with all psychological tests. In this sense, higher Cho levels and lower NAA/Cho ratios in both hippocampi have been reported in patients with FM [27]. The finding of metabolic brain differences between patients with FM and healthy controls in neural structures such as the hippocampus and amygdala (both of which pertain to the limbic system and are involved in fear, avoidance, and emotional responses experienced during pain) is compatible with a possible augmented emotional processing in patients with FM, in line with the augmented pain processing proposed by some authors [33].
Previous fMRI studies of FM have observed that augmented neural activity is due to an elevation in Glu levels, which leads to neuronal hyperexcitability. There is an increase in Glu in the amygdala [34], the posterior cingulate cortex [29], and the right posterior insula [35]. The degree of Glu elevation was associated with evoked pain sensitivity, suggesting that glutamatergic activity in this region of the brain might be partly responsible for the “gain” setting on central neural pain processing [35].
There is a study analyzing the neurotransmitter gamma aminobutyric acid (GABA) which describes a decrease in right anterior insula and an increase by the same amount in the anterior cingulate cortex [14]. Other studies report metabolic abnormalities in areas not studied by morphometry as an increased Cho/Cr ratio in the left and right caudate nuclei and right ventrolateral prefrontal cortex [36], and an increase in the Glu/Cr and Gln + Glu/Cr ratios [37].
A recent study has investigated the relationship between Glx- and GABA-derived spectroscopy values within the posterior cingulate, and the connectivity of this structure to the rest of the default mode network (DMN) [38]. The authors found that individuals with greater concentrations of Glu + Gln (Glx) and lower concentrations of GABA within the posterior cingulate have stronger connectivity values with other default mode network (DMN) regions.
One approach that might be particularly informative in FM would be to explore the association between insula connectivity and Glx/GABA levels in the same patient cohort. Alternatively, Glx and GABA concentrations in the insula may also influence functional connectivity between other brain regions and networks, as the insula has widely distributed excitatory and inhibitory connections throughout the brain [39].
Lower GABA levels within the posterior insula were associated with greater sensitivity to experimental pain. These findings suggested that lower insular GABA may also play a role in pain, namely neuronal disinhibition. In neuropathic pain syndromes, studies on metabolism spectroscopy reflect a decrease in NAA in the thalamus [26], a decrease in GABA and GABA/Cr in right reticular nucleus, and NAA and NAA/Cr in the lateral ventral posterior nucleus [40] and in the posteromedial and posterolateral nucleus, and NAA/Cr as well as the Cho/Cr combination in the intralaminar nuclei [41]. In chronic pain syndromes, metabolic changes occur in various brain regions. For example, diabetic neuropathy shows decreased NAA in the thalamus, normal levels in the anterior cingulate, and a decrease in Cr in the dorsolateral prefrontal cortex (DLPFC); while temporomandibular joint dysfunction (TMJ) shows increased NAA and Cho in the posterior insula, increased Gln in the right insula, and decreased Gln in the left insula [42]. Our group observed a decrease in NAA and increased Glu/Cr in the posterior cingulate of chronic pain patients compared to patients without pain [11, 29].
The quantification of both brain metabolites and neurotransmitters is of great interest, as it can provide indirect evidence of local neural activity and/or excitability, and may even be a predictor for therapy response. Furthermore, whether altered neurotransmitter concentrations in patients with chronic pain reflect a global (whole brain) or region-specific phenomenon (e.g., confined to regions in the pain system) needs to be investigated in greater depth.
Recent studies show a significantly higher Glu/Gln ratio in the occipital cortex of migraine patients compared with healthy control subjects, and higher Glu levels and Glu/Cr + phosphocreatine ratios in the anterior paracingulate cortex. This situation could arise from a neuronal–glial coupling of glutamatergic metabolism differences or an increased neuron/astrocyte ratio [43]. Although these data may reflect a state of neuronal hyperexcitability, they may also be associated with a nonspecific pain process. Moreover, altered glutamatergic neurotransmission seems to mediate the relationship between abnormal cortical information processing and excitability in migraine patients [44, 45].
In a study of patients with tinnitus, the patients showed higher concentrations of Glu and NAA in the auditory cortical areas, most notably in Heschl’s gyrus [46]. Another study that used MRS before, during, and after experimentally induced dental pain showed a significant absolute increase in Glu, Gln, and the Glu/Gln ratio in the insular cortex [47].
Our group has observed an increase in Glx in the posterior cingulate cortex in FM and, to a lesser extent, in somatization disorder compared with controls and levels of Glx correlates with pain-catastrophizing [11]. Our data suggest that Glx plays a role in this augmented pain processing in those individuals who have elevated Glx levels. Because higher Glx levels have been associated with an elevation in the pain catastrophizing syndrome (PCS), it is likely that Glx in the posterior cingulate is related to pain processing. We have hypothesized that increases in brain excitatory neurotransmitters could result in neuronal hyperexcitability. As part of its neurotransmitter role, Glx is an excitatory amino acid, and excessive Glx neurotransmission has been implicated in excitotoxic neuronal damage [48].
Structural Brain Imaging
Pain is defined as “an unpleasant sensory and emotional experience associated with actual or potential tissue damage, or described in terms of such damage” [4]. Pain is therefore a multidimensional phenomenon that is heavily influenced by biopsychosocial factors.
In the field of structural imaging, new approaches such as T1 and T2* mapping, as well as magnetization transfer ratios, which are often acquired during the same scanning session (multiparametric approach), will be of clinical importance by unraveling new aspects of altered microstructure in chronic pain conditions, and of methodological importance by providing a more detailed understanding of voxel-based morphometry findings [49].
Several brain structures are involved in the perception and experience of pain, such as the somatosensory cortex (primary and secondary), the cingulate gyrus (anterior), insula, thalamus, posterior parietal cortex, and prefrontal cortex. Studies show changes in the volume of brain areas involved in a “network of pain” in both FMS and neuropathic pain syndromes, and even in other forms of chronic pain. It has been suggested that prolonged nociceptive input to the brain might induce functional and morphologic maladaptive processes, which in turn further exacerbate the experience of chronic pain. Alternatively, morphologic changes might predispose toward vulnerability to develop a chronic pain state [50] (See Table 30.2). Interpretation of findings from morphometric studies must also take into account genetic and experiential factors that have recently been demonstrated to influence brain morphometry and the risk of developing chronic pain [51].
Reductions have been observed in the volume of gray matter in areas associated with pain: a level of emotional processing (anterior cingulate cortex and insula), cognitive (prefrontal cortex) and sensory (primary and secondary somatosensory cortex) processing, and the processing of stress (parahippocampal gyrus) [52].
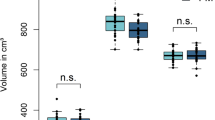
Significant gray matter reduction has been found in the prefrontal cortex, anterior cingulate cortex, and insular cortex of FM patients [53]. These regions are known to be critically involved in the modulation of subjective pain experiences. The duration of pain or functional pain disability did not correlate with gray matter volumes. A trend of inverse correlation of gray matter volume reduction in the anterior cingulate cortex was detected with the duration of pain medication intake [54]. Furthermore, reductions in gray matter volume were seen in the postcentral gyri, amygdala, hippocampi, superior frontal gyri, and anterior cingulate gyri [55]. Other authors [56] found those patients with FM had significantly less total gray matter volume and an age-associated decrease in gray matter 3.3 times greater than that of healthy controls. Greater gray matter loss was seen in patients with a longer duration of the disorder, with each year of FM being equivalent to 9.5 times the loss in normal aging. In addition, patients with FM demonstrated significantly lower gray matter density than healthy controls in brain regions such as the cingulate, the insular and medial frontal cortices, and the parahippocampal gyri. In summary, FM appears to be associated with an acceleration of age-related changes in the brain in regions that are functionally linked to core features of the disorder, including affective disturbances and chronic widespread pain. However, these results require replication because the sample was somewhat small (N = 10), and matching between controls and patients with FM was not optimal.
Changes in volume have been found in the hippocampus and insula in posttraumatic stress disorder [57, 58], in the thalamus and the prefrontal cortex in back pain [59], and also bilaterally in the prefrontal cortex in chronic fatigue syndrome [60], with global changes in this volume [61]. One possible explanation for these changes may be atrophy secondary to chronic inflammation mediated by cytokines [59].
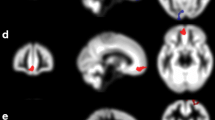
The thalamus plays a crucial role in the sensory–discriminative pain component. Both morphometry and diffusion tensor showed a volume decrease in different thalamic nuclei [55, 62]. In neuropathic pain syndromes (See Table 30.3), a decrease was also observed in the volume of the regions involved in the perception and processing of pain, and the thalamus [63, 64], the cortex of the region anterior cingulate gyrus [63, 65], and the insula [63].
The cingulate gyrus is part of the limbic system, and numerous neuroimaging studies demonstrate the involvement of the anterior cingulate region. Activation of this region is part of the emotional and cognitive component of pain [66, 67]. Both this region and the prefrontal cortex are involved in pain modulation (inhibition and facilitation).
Structural changes in these systems could contribute to the maintenance of pain and chronicity of symptoms, both in FMS and some neuropathic pain syndromes. A number of authors [68] have proposed a model in which the transition from acute to chronic pain leads to a reorganization of cortical sensory and affective pain pathways, which would mean a contribution of independent CNS peripheral nociceptive input. The morphological variations relating to these chronic pain syndromes evolve over time and respond to concepts of neuroplasticity, not to irreversible structural loss mechanisms. NAA appears to decrease in hippocampus [11, 27] as a neuronal marker, providing the molecular correlate to the volume loss observed in studies of morphometry. It can therefore be deduced that there is chronic neuronal damage in this structure in FMS. This point only could be verified by postmortem neuropathological study.
The decrease in volume of the insula [55, 56, 69] correlates with increased Glu [35] and decreased GABA [14]. These alterations suggest a metabolic dysregulation due to CNS hyperexcitability, and this is probably responsible for the overall increase in sensitivity to noxious stimuli observed in these patients. The insula could encode or extract the magnitude of the painful stimulus regardless of sensory modality [70].
Perfusion Magnetic Resonance Imaging
Pioneering single-photon emission computed tomography (SPECT) studies on somatization disorder first described hypoperfusion, primarily in the nondominant hemisphere, in the frontal, prefrontal, temporoparietal, and cerebellar areas [71].
Other researchers [72] conducted an observational study using technetium- 99 m ethyl cysteinate dimer (Tc-99 m ECD) brain SPECT to detect abnormal regional cerebral blood flow (rCBF) in 92 patients with FM. They found rCBF heterogeneity in patients with both primary and concomitant FM compared with the homogeneous rCBF observed in control patients. This difference was observed primarily in the left temporoparietal area, but was also described in the thalamus, as well as in the right temporoparietal, frontal, and basal ganglia areas. Differences in rCBF hypoperfusion in these areas between primary and concomitant FM groups were not significant. In conclusion, SPECT was not useful for differentiating primary and concomitant FM when the underlying disease activity was quiescent.
There are three main methods for studying brain perfusion by means of MRI:
Dynamic Susceptibility Contrast Imaging (DSCI)
This is the most widespread method of perfusion imaging, and is likely to be a standard sequence on most MR machines. It relies on the T2* signal drop caused by the passage of a gadolinium-containing contrast agent through the tissues. fMRI is based on measuring and analyzing the so-called BOLD effect. An increase in neural activity leads to a hemodynamic response, associated with an increase in regional blood flow and volume resulting in an increase of the oxyhemoglobin–deoxyhemoglobin ratio, which in turn leads to a reduction of local magnetic in homogeneity.
Dynamic Contrast Enhancement (DCE)
This method uses a rapid T1 sequence to measure changes in signal intensity as a bolus of gadolinium diffuses across the damaged blood-brain barrier into the extracellular, extravascular space.
Arterial Spin Labeling (ASL)
This is a newer MRI technique that uses water in arterial blood as a freely diffusible tracer to measure perfusion noninvasively, whereby the blood flowing into the brain is magnetically labeled (arterial spin labeling). This technique is still largely research-based, and provides truly quantitative values of cerebral blood flow. In future studies, the combination of arterial spin labeling and BOLD imaging might provide new insight into the interaction of neural activity with vascular responses, which is of particular importance, as neural activity cannot be measured directly using MRI techniques.
In a multimodal imaging study using H-MRS and ASL [40], it was possible to demonstrate that patients with neuropathic pain following spinal cord injury displayed lower levels of thalamic NAA, GABA, and regional blood flow (in the thalamus) compared to healthy controls and patients with spinal cord injury but no pain. This and other studies support the notion that neuropathic pain is associated with CNS reorganization, specifically within the thalamus, comprising functional as well as neurochemical mechanisms.
Diffusion Tensor Imaging (DTI)
Neuroimaging reveals changes in the white matter structure in the human brain. White matter comprises half of the human brain, and consists of bundles of myelinated axons connecting neurons in different brain regions [73]. Gray matter is composed of neuronal cell bodies and dendrites concentrated in the outer layers of the cortex.
Microstructural changes in white matter can be revealed by specialized MRI brain imaging techniques such as DTI. This method analyzes the diffusion of protons in tissue, which is more restricted in white matter than in gray matter.
Water molecules in the brain are in constant Brownian motion, and although the movement of these protons affects conventional structural imaging, diffusion-weighted imaging (DWI) and DTI allow quantification of this microscopic movement within each voxel. The main advantage of using DTI, rather than DWI, is that DTI reflects the underlying diffusion properties of the sample, independently of the orientation of the tissue with respect to the direction of measurements. DTI is thus a robust quantitative technique that is independent of how the subject has been oriented inside the scanner magnet and gradient coils.
The appropriate mathematical combination of the directional diffusion-weighted images provides quantitative measures of water diffusion for each voxel via the apparent diffusion coefficient (ADC), as well as the degree of diffusion directionality, or anisotropy. Myelin is a major diffusion barrier for water, and gives white matter its high anisotropy [74]. Demyelinating diseases are characterized by partial or total loss of myelin, with consequent loss of neuronal function. Anisotropy increases with increased myelination, diameter, and axon compaction.
Previous studies have identified and confirmed the existence of an anatomic circuitry for the functionally characterized, top-down influences on pain processing via brainstem structures in humans [75]. Fractional anisotropy (FA) is a measure of the portion of the diffusion tensor from anisotropy. Previous studies with DTI in FM patients showed alterations in the right thalamus and significantly lower fractionated anisotropy in comparison with controls. A negative correlation was observed between the FA values in the right thalamus and clinical pain in the FM group [76]. Other authors have confirmed that DTI in the brain of patients with FM appeared to be more sensitive than volumetric imaging of voxel-based morphometry (VBM), and that increased pain intensity scores were correlated with changes in DTI measurements in the right superior frontal gyrus. Increased fatigue was correlated with changes in the left superior frontal and left anterior cingulate gyrus, and self-perceived physical impairment was correlated with changes in the left postcentral gyrus. Higher intensity scores for stress symptoms were correlated negatively with diffusivity in the thalamus and FA in the left insular cortex [55].
Functional Magnetic Resonance Imaging (FMRI)
Clinical studies with functional imaging show that pain is not a static condition with a pathophysiology that is only localized in the peripheral system of muscles or tendons, but that it is a highly plastic clinical pathology that affects multiple central neural systems and defines the so-called “neural matrix” of pain or network of cortico-subcortical areas involved in pain processing.
fMRI is a noninvasive technique that detects and locates focal brain activation taking place and involved in performing a task by means of a cognitive, emotional, or sensory-motor neuron circuit. Generally, fMRI studies are based on the acquisition of images during one sequence while the patient is at rest and another while performing a task. The subsequent statistical comparison between the two phases (rest and activation) represents, in an ideal case, the focal metabolic and vascular changes in the cerebral cortex that are in operation during performance of the task being studied, or by block-design paradigms that are well connected with the episode (event-related). However, fMRI can evaluate the metabolic and vascular condition that occurs at rest (resting-state) and the actual time duration of the pattern of brain activation when performing a functional certain task under study.
The DMN comprises a set of brain regions that are coactivated during passive task states. These show an intrinsic functional relationship, and are connected via direct and indirect anatomic projections. The medial temporal lobe subsystem provides information from previous experiences in the form of memories and associations, which are the building blocks of mental simulation. The medial prefrontal subsystem facilitates the flexible use of this information during the construction of self-relevant mental simulations. These two subsystems converge on important nodes of integration, including the ventral posterior cingulate cortex (vPCC) [77].
Multiple techniques have been devised to evaluate functional brain connectivity. For correlational analyses, the main techniques are seed correlation and independent component analysis (ICA). For seed correlation, the fMRI signal is extracted from a seed region of interest, and is then correlated with the fMRI time series taken from all other brain voxels [78]. Alternatively, ICA is a data-driven technique that considers all voxels in the brain and clusters them into spatiotemporally distinct networks, which are spatially independent of one another [79].
Recent functional neuroimaging studies have enabled the neuroanatomical differentiation of the classic dimensions of pain processing. Sensory and cognitive dimensions are at the top and brain dorsal portion (contralateral primary somatosensory cortex, bilateral secondary somatosensory cortex, insular cortex in its rear portion, opercular area, thalamus, frontoparietal neocortex, and supplementary motor area). The emotional dimension involves the insular cortex, anterior cingulate cortex, basal ganglia, and prefrontal cortex. Consequently, the functional activation of the neural circuitry of pain can be modulated by either a sensory or emotional component, not to mention the involvement of the cognitive component.
The emotional component can mediate the intensity threshold at which a stimulus is perceived as painful. In fact, this threshold appears to be much lower in patients diagnosed with chronic functional pain (such as FM) than in healthy controls, and can generate an abnormal activation of the neuronal pain circuit. This has given rise to the collective denomination of these syndromes as central susceptibility syndromes. Patients with FM have been found to have greater connectivity between the DMN and the insular cortex, which is a brain region known to process evoked pain, and the executive attention network [80]. Resting-state functional magnetic resonance imaging (rfMRI) data from 18 patients with FM and 18 age-matched healthy control subjects were analyzed using dual-regression ICA, which is a data-driven approach for the identification of independent brain networks. Intrinsic, or resting-state, connectivity was evaluated in multiple brain networks: the DMN, the executive attention network (EAN), and the medial visual network (MVN), with the medial visual network serving as a negative control. Spontaneous pain levels were also analyzed for covariance with intrinsic connectivity. These findings indicate that resting brain activity within multiple networks is associated with spontaneous clinical pain in patients with FM. These findings may also have broader implications for how subjective experiences such as pain arise from a complex interplay among multiple brain networks (See Table 30.4).
While acute experimental pain induces default mode network deactivation in healthy subjects [81], chronic back pain is associated with mitigated DMN deactivation to visual attention tasks [82]. The DMN showed greater connectivity to the insula cortex and secondary somatosensory cortex (S2) (brain regions known to process evoked experimental pain and somatosensation), while the EAN showed greater intra-network connectivity in FM patients. Both the DMN and EAN were more connected to the insula in patients reporting greater spontaneous clinical pain at the time of the scan, which suggested a close link between DMN–insula connectivity and clinical pain.
Reduced resting connectivity within the somatosensory system and increased connectivity between the default mode network and somatosensory processing regions, such as the secondary somatosensory cortex, were recently reported [83]. Such independent, confirmatory data are important for any neuroimaging-based markers of disease in FM, and further research is needed. Interestingly, this study also found altered connectivity with brain regions supporting visual and auditory processing, which may relate to the multisensory dysfunction sometimes reported in these patients.
A study used a seed voxel region of interest approach, and showed a pattern of both increased and decreased brain connectivity in FM patients [84]. Increased connectivity was found between DMN areas such as medial prefrontal cortex (mPFC) and posterior cingulate cortex (PCC), and also between anterior cingulate cortex (ACC) and the insula. These results support the fact that DMN and insula resting connectivity is disrupted in FM.
A recent study explored structural and fMRI changes in FM patients, and found an interesting association with age [85]. Younger, but not older, FM patients showed decoupling between the insula and anterior mid-cingulate cortex, two brain regions that are normally strongly connected in healthy adults, as part of a salience network. Another study reported increased frequency power (for a broad 0.01–0.25 Hz band) in somatosensory (primary somatosensory cortex, S1), cognitive (DLPFC), and affective (amygdala) brain regions in FM patients [86].
During mild pain events, a relationship has been demonstrated between catastrophizing and activity in cortical regions associated with affective, attention, and motor aspects of pain. This includes the dorsolateral prefrontal, insular, rostral anterior cingulate, premotor, and parietal cortices. During more intense pain, prefrontal cortical regions involved in top-down pain modulation are negatively correlated with catastrophizing. An explanation for this may be that a cortical vigilance network is engaged during mild pain. However, diminished prefrontal cortical modulation impedes disengagement and suppression during more intense pain [87]. These findings may also involve catastrophizing in the progression toward or persistence of chronic pain.
Patients with somatoform pain disorders are thought to have an early-acquired defect in stress regulation. The fMRI [88] was used to search for common alterations in the pain-responsive and stress-responsive cortical areas. They studied a group of 17 patients and an age-matched control group by inducing pin-prick pain, cognitive stress, and emotional stress. The patients demonstrated increased activation of pain-processing areas (thalamus, basal ganglia, and operculo-insular cortex) during first pain exposure, and increased activation of some prefrontal, temporal, and parietal regions was also observed. Temporal and parietal areas were also activated during cognitive stress, and activation was reduced during emotional stress. However, hippocampal volume was not significantly reduced in the patient group. This study supports the current concept that central processing of pain and cognitive stress is increased in these patients, possibly owing to exaggerated memory or anticipation of pain exposure, or both, and to a disturbance in stress regulating systems. Though surprising, the finding of a reduced responsiveness to emotional stress is not contradictory to this hypothesis. Some sort of neglect or coping mechanisms may have developed over time as a response to earlier adverse events.
Another fMRI study [89] researched the cerebral processing of noxious heat stimuli as objective markers for pain sensation in 12 right-handed women diagnosed with somatoform pain disorder and 13 age-matched, healthy volunteers. Compared with controls, patients with pain disorder responded to induced pain with hypoactivation of the ventromedial prefrontal/orbitofrontal cortex, and hyperactivation of the amygdala, parahippocampal gyrus, and anterior insula. The finding of altered cerebral processing of experimentally induced pain in patients with somatoform pain disorder supports the hypothesis of dysfunctional pain processing, particularly in affect-regulating regions.
An fMRI analysis [13] revealed no differences in activity in brain regions related with attention and affection, or regions with sensory projections from the stimulated body area. However, when there was a primary lesion in the descending pain regulating system (the rostral anterior cingulate cortex), the patients failed to respond to pain provocation. The attenuated response to pain in these cases is the first demonstration of a specific brain region where the impairment of pain inhibition in FMS patients is expressed. These results validate previous reports of dysfunctional endogenous pain inhibition in FM, and advance the understanding of the central pathophysiologic mechanisms, providing a new direction for the development of successful treatments in FM.
Although the etiology of this disorder remains largely unknown, emerging data suggest that FM arises through augmentation of central pain processing pathways. This hypothesis is largely based upon findings of previous functional neuroimaging studies, showing that FM patients display augmented neuronal responses to both innocuous and painful stimuli [12, 33], confirming the allodynia and hyperalgesia seen in this condition [90].
Studies with functional neuroimaging support the hypothesis of central pain augmentation in FM. Differences of activation in the fronto-cingulate cortex, the supplemental motor areas, and the thalamus were found between both groups with distinct differences in BOLD signal changes over the duration of pain stimulation, and even during anticipation of pain. These results support the hypothesis that central mechanisms of pain processing in the medial pain system and favorable cognitive/affective factors even during the anticipation of pain may play an important role for pain processing in patients with FMS [54].
The default network is disrupted during painful stimuli [91, 92] in FM [93] and depression [94], thereby further encouraging researchers to consider how the functions of the DMN might be important in understanding diseases of the mind. The functional connectivity pattern within the DMN is altered during pain, selectively in the prefrontal cortex and posterior cingulate cortex–precuneus (increased connectivity), and in the lateral parietal cortex (decreased connectivity). A limited number of functional neuroimaging pain studies have shown that the activity in a network, including the posterior cingulate cortex–precuneus, the inferior parietal lobule, and the medial prefrontal cortex, was consistently reduced in response to a range of painful stimuli [91, 92].
New acquisition techniques and new analysis strategies have emerged that enable new conceptual approaches to the acquisition of data, such as network and multivariate pattern analyses, and in particular, support vector machines (SVM) [95]. The emergence of connectivity analyses, both functional connectivity (resting state) and structural connectivity, as enabled by DTI and tractography, have allowed not only for the analysis of the connectedness of two remote brain areas, but also for the construction and analysis of large networks consisting of multiple brain sites. One such approach is the graph theory, where graphs are mathematical structures to model relations between objects.
The use of the graph theory in the analysis of chronic pain states has been limited. When investigating patients with migraine (without aura) and healthy controls, and applying the graph theory based on resting-state functional connectivity analyses [96], a disruption was found in whole-brain networks with an increase in disease duration, in which areas implicated in sensory discrimination constituted an abnormal network configuration.
Treatment Effects
Mindfulness meditation has beneficial effects on a number of psychiatric, functional somatic, and stress-related symptoms, and therefore has been increasingly incorporated into psychotherapeutic programs [97, 98], with subjects reporting better pain-related quality of life and greater life satisfaction [99]. Altogether, 25 systematic reviews were found; they investigated the evidence of complementary and alternative medicine (CAM) for the FMS [100]. In general, they were exercised-based CAM therapies, manipulative therapies, mind–body therapies, acupuncture, hydrotherapy, phytotherapy, and homeopathy. Consistently positive results were found for tai chi, yoga, meditation and mindfulness-based interventions, hypnosis or guided imagery, electromyogram (EMG) biofeedback, and balneotherapy/hydrotherapy. Inconsistent results were found for qigong, acupuncture, chiropractic interventions, electroencephalogram (EEG) biofeedback, and nutritional supplements. Inconclusive results were found for homeopathy and phytotherapy. Despite a growing body of scientific evidence of CAM therapies for the management of fibromyalgia syndrome (FMS), systematic reviews still show methodological flaws that limit definite conclusions about their efficacy and safety.
Neuroimaging studies were conducted to analyze the brains of people with and without meditation experience. During the first 20 min inside the MRI scanner, they had spontaneous thoughts, and for the next 20 min they developed a simple exercise task, which consisted of focusing only on their breathing. As they began to practice this exercise, meditation with the usual respiratory concentration, medial prefrontal cortex activity decreased in all patients. This part of the DMN is considered relevant to self-centered mental processes. Moreover, although the blood flow in the medial prefrontal region of the inexperienced meditators decreased a few minutes later than that of the experienced meditators, the blood supply of the area was reduced for the duration of the exercise, suggesting the calming effects of meditation [101].
Anatomical likelihood estimation (ALE) meta-analysis found eight brain regions of GM that were consistently enhanced in meditators [102]. Three studies [103,104,105] showed an apparent pattern of structural increase in WM in meditators versus controls. Our research found that meditators showed a lower apparent diffusion coefficient (ADC) in the left posterior parietal white matter than did controls, and that the ADC was negatively correlated with years of meditation.
Similar research [106] addressed the functional connectivity of the DMN in subjects who commonly practiced mindfulness versus subjects who did not. Their results indicated both reduced activation of two main nodes of the DMN (posterior cingulated cortex and medial prefrontal cortex), and that experienced meditators showed activation of the medial prefrontal cortex, insula, and temporal lobes during meditation, a differential pattern of functional connectivity both during resting and during mindfulness exercises. Other authors [107] show that the activity in a subregion of the DMN, the ventromedial prefrontal cortex, is inversely correlated with years of meditation experience, suggesting that the experience of meditation can enable more efficient cognitive processes subserved by this region. Another study [108], also reported a higher functional connectivity in the DMN in meditator subjects (medial prefrontal cortex), suggesting that meditation practice is associated with functional changes in areas of the DMN even when not practicing. In summary, existing studies suggest differential patterns in meditators’ functional connectivity, consistent with reduced mind-wandering, a greater awareness of the present moment, and self-referential processing than those found in non-meditators [106, 109].
Meditation may be able to reinforce positive feelings, especially compassion and benevolence. To test this hypothesis [110], subjects performed compassion exercises while lying down in a brain scanner. Half of the 30 volunteers had several years of experience in Buddhist meditation techniques. The control group comprised age-matched participants with no experience in this type of group meditation. Emotional reactions were provoked with either the laughter of a baby or a deeply distressed groan. Such acoustic signals primarily stimulated those areas that had been shown in other studies to process emotional stimuli (the insula, the anterior cingulate cortex, and secondary somatosensory area). The major differences between experienced meditators and novices were observed in the insula. Many of these phenomena are explained through mechanisms of neuronal plasticity: An intense effort results in alterations in the structure and mode of operation of certain areas of the brain.
One review [111] described up to 17 research studies in which the therapeutic potential of mindfulness in pain was analyzed. Another study [112] showed that Zen meditators have pain sensitivity thresholds higher than non-meditator subjects. This is where regulation comes into play as a basic feature of meditation [113]. It appears that one effect of reduced activation of certain areas is a reduction in the connectivity between them. Connectivity has been associated with complex functions that are performed by multiple brain structures in combination. The study showed increased activation of areas typically associated with pain, such as the insula, thalamus, anterior cingulate cortex, and prefrontal cortex [33, 114]. If only this increased activity is observed, it might seem that meditators are feeling more pain than nonmeditators, which contrasts with the poor results obtained when they were asked to rate their pain. Connectivity studies, however, show that meditation reduces the connectivity between these areas related to pain regulation.
Similarly, several authors have studied the role of the prefrontal cortex using emotion-regulation tasks. For example, in the first study to address this [115], meditators were asked to perform a task of emotion recognition, with results showing lower connectivity between the prefrontal cortex and the right amygdala than in participants who did not practice meditation. The authors hypothesized that meditators tend to treat emotional states as “objects” of care. By treating these conditions as transient mental products, this allows the meditator to maintain greater distance from emotional experiences. This contrasts with the usual way of thinking and feeling emotions and thoughts, in which they are considered “facts” or “reality”. Results in this field have been obtained by other authors [116, 117], although some of these [118] pointed to the amygdala as a major participant in the regulation of emotions. These authors studied the regulation of anxiety through meditation techniques, which showed reduced amygdala activity after performing a series of exercises.
However, other researchers [119] showed that DMN–insula connectivity, which was increased in FM patients, was reduced following 4 weeks of nonpharmacological acupuncture and sham acupuncture therapy, resulting in reduced pain in these patients. The authors suggested that connectivity between the DMN and insula may serve as a possible surrogate biomarker for pain reduction in FM.
Such studies could indeed play a role in clinical practice, as they could, in addition to behavioral measures such as anxiety and catastrophizing scores, help to identify patients at a high risk of developing chronic pain, implying the necessity of early therapeutic intervention. Likewise, it is desirable to determine whether or not patients are likely to respond to a certain therapy. In two recently performed studies, functional connectivity was demonstrated to predict clinical improvement in response to pregabalin or milnacipran intake in FM patients [120, 121].
Interestingly, functional brain imaging has also been used to predict response to placebo treatment [121,122,123]. In the context of personalized medicine, placebo treatment might indeed be a therapeutic option in several patients with chronic pain, and brain imaging could help to identify patients who are likely to benefit from a placebo treatment.
Other authors investigating analgesic mechanisms of pregabalin in an FM group have seen that reductions in clinical pain were associated with reductions in functional connectivity between the DMN and the posterior insular cortex, corroborating the theory that the interaction of these two regions might play a specific role in chronic pain [120].
A recent study by our group, with 63 patients, showed preliminary evidence of the utility of memantine for the treatment of FM. Compared with a placebo group, memantine significantly decreased ratings on a pain visual analog scale (Cohen’s d = 1.43 at 6 months) and pain measured with a sphygmomanometer (d = 1.05). All other secondary outcomes except anxiety also improved, with moderate-to-large effect sizes at 6 months. Compared with placebo, the absolute risk reduction obtained with memantine was 16.13% (95% confidence interval = 2.0–32.6%), and the number needed to treat was 6.2 (95% confidence interval = 3–47) [124].
Interestingly, in another study conducted by our group using MRS at baseline and 6 months, in 13 patients with FM treated with memantine and 12 with placebo, the patients treated with memantine exhibited a significant increase in Glu, the Glu/Cr ratio, Glx, and total NAA + NAAG in the posterior cingulate cortex compared with those on placebo. Furthermore, the memantine group exhibited increases in Cr and Cho in the right posterior insula, while a correlation between Cho and the Fibromyalgia Impact Questionnaire (FIQ) in the posterior insula was also observed, demonstrating that memantine treatment resulted in an increase in cerebral metabolism in FM patients, and suggesting its utility for the treatment of the illness [125].
The combination of ASL and BOLD imaging might provide new insight into the interaction of neural activity with vascular responses, which is of particular importance given that neural activity cannot be measured directly using MRI techniques. Some authors have applied ASL and a Gaussian process binary classifier to distinguish intraindividually between a nonpain condition (prior to molar extraction) and postsurgical pain (after third molar extraction from the lower jaw), reaching a classification accuracy of 95 [126]. Others have reported that the functional connectivity between the nucleus accumbens (NAc) and the prefrontal cortex in patients with subacute back pain was predictive of whether the pain persisted [127]. A similar association was described for the structural connectivity of the prefrontal cortex as assessed by DTI [128].
Conclusions
This chapter discussbes several techniques used for the diagnosis of FM. At present, there are no other noninvasive techniques that can provide equivalent information and, as a consequence, MRS, DTI tractography, and fMRI are expected to be a powerful combined technique for researching brain anatomy and disease in situ in human beings.
The main findings among patients with chronic pain are an increased functional connectivity between the pain system and the DMS, decrease in gray matter volume in the insular cortex and anterior cingulate cortex, and also decreased GABA concentrations in the insular cortex or thalamus.
New acquisition techniques and new analysis strategies have emerged that enable new conceptual approaches to the acquisition of data, such as network and multivariate pattern analyses, and in particular, support vector machines (SVM).
Continued improvements in the design of imaging equipment and analysis algorithms are progressively improving the specificity of the biological parameters that can be calculated, allowing detailed quantitative characterization of microvascular structure in a wide range of pathological tissues, including FM.
References
Sjøgren P, Ekholm O, Peuckmann V, Grønbaek M. Epidemiology of chronic pain in Denmark: an update. Eur J Pain. 2009;13:287–92.
Blyth FM, March LM, Brnabic AJ, Jorm LR, Williamson M, Cousins MJ. Chronic pain in Australia: a prevalence study. Pain. 2001;89:127–34.
Manchikanti L, Boswell MV, Singh V, Derby R, Fellows B, Falco FJ, Datta S, Smith HS, Hirsch JA. Comprehensive review of neurophysiologic basis and diagnostic interventions in managing chronic spinal pain. Pain Physician. 2009;12:E71–120.
International Association for the Study of Pain (IASP). Classification of chronic pain. 2nd ed. IASP Task Force on Taxonomy, Merskey H, Bogduk N, editors. Seattle: IASP Press; 1994.
Wolfe F, Smythe HA, Yunus MB, Bennet RM, Bombardier C, Goldenberg ADL. American College of Rheumatology 1990. Criteria for the classification of fibromyalgia. Report of the Multicenter Criteria Committee. Arthritis Rheum. 1990;33:160–72.
Wolfe F, Clauw DJ, Fitzcharles MA, Goldenberg DL, Katz RS, Mease P, Russell AS, Russell IJ, Winfield JB, Yunus MB. The American College of Rheumatology preliminary diagnostic criteria for fibromyalgia and measurement of symptom severity. Arthritis Care Res. 2010;62:600–10.
Branco JC, Bannwarth B, Failde I, Abello Carbonell J, Blotman F, Spaeth M, Saraiva F, Nacci F, Thomas E, Caubère JP, Le Lay K, Taieb C, Matucci-Cerinic M. Prevalence of fibromyalgia: a survey in five European countries. Semin Arthritis Rheum. 2010;39:448–53.
Gamero Ruiz F, Gabriel Sánchez R, Carbonell Abelló J, Tornero Molina J, Sanchez-Magro I. Pain in Spanish rheumatology outpatient offices: EPIDOR epidemiological study. Rev Clin Esp. 2005;205:157–63.
Treede RD, Jensen TS, Campbell JN, Cruccu G, Dostrovsky JO, Griffin JW, Hansson P, Hughes R, Nurmikko T, Serra J. Neuropathic pain: redefinition and a grading system for clinical and research purposes. Neurology. 2008;70:1630–5.
American Psychiatric Association. Diagnostic and statistical manual of mental disorders DSM-IV-TR. 4th (text revision) ed. Washington, DC: American Psychiatric Association; 2000.
Fayed N, Andres E, Rojas G, Moreno S, Serrano-Blanco A, Roca M, Garcia-Campayo J. Brain dysfunction in fibromyalgia and somatization disorder using proton magnetic resonance spectroscopy: a controlled study. Acta Psychiatr Scand. 2012;126:115–25.
Cook DB, Lange G, Ciccone DS, Liu WC, Steffener J, Natelson BH. Functional imaging of pain in patients with primary Fibromyalgia. J Rheumatol. 2004;31:364–78.
Jensen KB, Kosek E, Petzke F, Carville S, Fransson P, Marcus H, Williams SC, Choy E, Giesecke T, Mainguy Y, Gracely R, Ingvar M. Evidence of dysfunctional pain inhibition in Fibromyalgia reflected in rACC during provoked pain. Pain. 2009;144:95–100.
Foerster BR, Petrou M, Edden RAE, Clauw DJ, Sundgren PC, Schmidt-Wilcke T, Lowe SE, Harte SE, Clauw DJ, Harris RE. Reduced insular gamma-aminobutyric acid in fibromyalgia. Arthritis Rheum. 2012;64:579–83.
Ashburner J. Computational Neuroanatomy. PhD thesis, London: University College; 2000.
Barker PB. N-acetyl aspartate – a neuronal marker? Ann Neurol. 2001;49:423–4.
Moffett JR, Ross B, Arun P, Madhavarao CN, Namboodiri AM. N-Acetylaspartate in the CNS: from neurodiagnostics to neurobiology. Prog Neurobiol. 2007;81:89–131.
Urenjak J, Williams SR, Gadian DG. Specific expression of N-acetylaspartate in neurons, oligodendrocyte-type-2 astrocyte progenitors, and immature oligodendrocytes in vitro. J Neurochem. 1992;59:55–61.
Baslow MH. The vertebrate brain, evidence of its modular organization and operating system: insights in to the brain's basic units of structure, function, and operation and how they influence neuronal signaling and behavior. Front Behav Neurosci. 2011;5:5.
Sartorius A, Lugenbiel P, Mahlstedt MM. Proton magnetic resonance spectroscopic creatine correlates with creatine transporter protein density in rat brain. J Neurosci Methods. 2008;172:215–9.
Gill SS, Small RK, Thomas DG, Patel P, Porteous R, Van Bruggen N, Gadian DG, Kauppinen RA, Williams SR. Brain metabolites as 1H NMR markers of neuronal and glial disorders. NMR Biomed. 1989;2:196–200.
Katz-Brull R, Koudinov AR, Degani H. Choline in the aging brain. Brain Res. 2002;951:158–65.
Escartin C, Valette J, Lebon V, Bonvento G. Neuron–astrocyte interactions in the regulation of brain energy metabolism: a focus on NMR spectroscopy. J Neurochem. 2006;99:393–401.
Grachev ID, Fredrickson BE, Apkarian AV. Abnormal brain chemistry in chronic back pain: an in vivo proton magnetic resonance spectroscopy study. Pain. 2000;89:7e18.
Gustin SM, Peck CC, Wilcox SL, Nash PG, Murray GM, Henderson LA. Different pain, different brain: thalamic anatomy in neuropathic and non-neuropathic chronic pain syndromes. J Neurosci. 2011;31:5956–64.
Fukui S, Matsuno M, Inubushi T, Nosaka S. N-Acetylaspartate concentrations in the thalami of neuropathic pain patients and healthy comparison subjects measured with (1) H-MRS. Magn Reson Imaging. 2006;24:75–9.
Emad Y, Ragab Y, Zeinhom F, El-Khouly G, Abou-Zeid A, Rasker JJ. Hippocampus dysfunction may explain symptoms of fibromyalgia syndrome. A study with single-voxel magnetic resonance spectroscopy. J Rheumatol. 2008;35:1371–7.
Wood PB, Glabus MF, Simpson R, Patterson JC. Changes in gray matter density in fibromyalgia: correlation with dopamine metabolism. J Pain. 2009;10:609–18.
Fayed N, Garcia-Campayo J, Magallón R, Andrés-Bergareche H, Luciano JV, Andres E, Beltrán J. Localized 1H-NMR spectroscopy in patients with fibromyalgia: a controlled study of changes in cerebral glutamate/glutamine, inositol, choline, and N-acetylaspartate. Arthritis Res Ther. 2010;12:R134.
Grachev ID, Thomas PS, Ramachandran TS. Decreased levels of N-acetylaspartate in dorsolateral prefrontal cortex in a case of intractable severe sympathetically mediated chronic pain (complex regional pain syndrome, type I). Brain Cogn. 2002;49:102–13.
Pattany PM, Yezierski RP, Widerström-Noga EG, Bowen BC, Martinez-Arizala A, Garcia BR, Quencer RM. Proton magnetic resonance spectroscopy of the thalamus in patients with chronic neuropathic pain after spinal cord injury. AJNR Am J Neuroradiol. 2002;23:901–5.
Mckenna JE, Melzack R. Blocking NMDA receptors in the hippocampal dentate gyrus with AP5 produces analgesia in the formalin pain test. Exp Neurol. 2001;172:92–9.
Gracely RH, Petzke F, Wolf JM, Clauw DJ. Functional magnetic resonance imaging evidence of augmented pain processing in fibromyalgia. Arthritis Rheum. 2002;46:1333–43.
Valdés M, Collado A, Bargalló N, Vázquez M, Rami L, Gómez E, Salamero M. Increased glutamate⁄glutamine compounds in the brains of patients with fibromyalgia: a magnetic resonance spectroscopy study. Arthritis Rheum. 2010;62:1829–36.
Harris RE, Sundgren PC, Craig AD, Kirshenbaum E, Sen A, Napadow V, Clauw DJ. Elevated insular glutamate in fibromyalgia is associated with experimental pain. Arthritis Rheum. 2009;60:3146–52.
Petrou M, Harris RE, Foerster BR, McLean SA, Sen A, Clauw DJ, Sundgren PC. Proton MR spectroscopy in the evaluation of cerebral metabolism in patients with fibromyalgia: comparison with healthy controls and correlation with symptom severity. Am J Neuroradiol. 2008;29:913–8.
Feraco P, Bacci A, Pedrabissi F, Passamonti L, Zampogna G, Malavolta N, Leonardi M. Metabolic abnormalities in pain-processing regions of patients with fibromyalgia: a 3 T MR spectroscopy study. Am J Neuroradiol. 2011;32:1585–90.
Kapogiannis D, Reiter DA, Willette AA, Mattson MP. Posteromedial cortex glutamate and GABA predict intrinsic functional connectivity of the default mode network. Neuroimage. 2013;64:112–9.
Napadow V, Harris RE. What has functional connectivity and chemical neuroimaging in fibromyalgia taught us about the mechanisms and management of ‘centralized’ pain? Arthritis Res Ther. 2014;16:425.
Gustin SM, Wrigley PJ, Youssef AM, McIndoe L, Wilcox SL, Rae CD, Edden RA, Siddall PJ, Henderson LA. Thalamic activity and biochemical changes in individuals with neuropathic pain after spinal cord injury. Pain. 2014;155:1027–36.
Wang Y, Li D, Bao F, Ma S, Guo C, Jin C, Zhang M. Thalamic metabolic alterations with cognitive dysfunction in idiopathic trigeminal neuralgia: a multivoxel spectroscopy study. Neuroradiology. 2014;56:685–93.
Chang L, Munsaka SM, Kraft-Terry S, Ernst T. Magnetic resonance spectroscopy to assess neuroinflammation and neuropathic pain. J Neuroimmune Pharmacol. 2013;8:576–93.
González de la Aleja J, Ramos A, Mato-Abad V, Martínez-Salio A, Hernández-Tamames JA, Molina JA, Hernández-Gallego J, Alvarez-Linera J. Higher glutamate to glutamine ratios in occipital regions in women with migraine during the interictal state. Headache. 2013;53:365–75.
Siniatchkin M, Sendacki M, Moeller F, Wolff S, Jansen O, Siebner H, Stephani U. Abnormal changes of synaptic excitability in migraine with aura. Cereb Cortex. 2012;22:2207–16.
Reyngoudt H, Achten E, Paemeleire K. Magnetic resonance spectroscopy in migraine: what have we learned so far? Cephalalgia. 2012;32:845–59.
Cacace AT, Silver SM. Applications of magnetic resonance spectroscopy to tinnitus research: initial data, current issues, and future perspectives. Prog Brain Res. 2007;166:71–81.
Gutzeit A, Meier D, Froehlich JM, Hergan K, Kos S, V Weymarn C, Lutz K, Ettlin D, Binkert CA, Mutschler J, Sartoretti-Schefer S, Brügger M. Differential NMR spectroscopy reactions of anterior/posterior and right/left insular subdivisions due to acute dental pain. Eur Radiol. 2013;23:450–60.
Bleich S, Römer K, Wiltfang J, Kornhuber J. Glutamate and the glutamate receptor system: a target for drug action. Int J Geriatr Psychiatry. 2003;18:S33–40.
Lorio S, Lutti A, Kherif F, Ruef A, Dukart J, Chowdhury R, Frackowiak RS, Ashburner J, Helms G, Weiskopf N, Draganski B. Disentangling in vivo the effects of iron content and atrophy on the ageing human brain. Neuroimage. 2014;103:280–9.
Schmidt-Wilcke T. Variations in brain volume and regional morphology associated with chronic pain. Curr Rheumatol Rep. 2008;10:467–74.
Wood PB. Variations in brain gray matter associated with chronic pain. Curr Rheumatol Rep. 2010;12:462–9.
Cagnie B, Coppieters I, Denecker S, Six J, Danneels L, Meeus M. Central sensitization in fibromyalgia? A systematic review on structural and functional brain MRI. Semin Arthritis Rheum. 2014;44:68–75.
Valet M, Gündel H, Sprenger T, Sorg C, Mühlau M, Zimmer C, Henningsen P, Tölle TR. Patients with pain disorder show gray-matter loss in pain-processing structures: a voxel-based morphometric study. Psychosom Med. 2009;71:49–56.
Burgmer M, Pogatzki-Zahn E, Gaubitz M, Wessoleck E, Heuft G, Pfleiderer B. Altered brain activity during pain processing in fibromyalgia. Neuroimage. 2009;44:502–8.
Lutz J, Jäger L, de Quervain D, Krauseneck T, Padberg F, Wichnalek M, Beyer A, Stahl R, Zirngibl B, Morhard D, Reiser M, Schelling G. White and gray matter abnormalities in the brain of patients with fibromyalgia: a diffusion-tensor and volumetric imaging study. Arthritis Rheum. 2008;58:3960–9.
Kuchinad A, Schweinhardt P, Seminowicz DA, Wood PB, Chizh BA, Bushnell MC. Accelerated brain gray matter loss in fibromyalgia patients: premature aging of the brain? J Neurosci. 2007;27:4004–7.
Villarreal G, Hamilton DA, Petropoulos H, Driscoll I, Rowland LM, Griego JA, Kodituwakku PW, Hart BL, Escalona R, Brooks WM. Reduced hippocampal volume and total white matter volume in posttraumatic stress disorder. Biol Psychiatry. 2002;52:119–25.
Chen S, Xia W, Li L, Liu J, He Z, Zhang Z, Yan L, Zhang J, Hu D. Gray matter density reduction in the insula in fire survivors with posttraumatic stress disorder: a voxel-based morphometric study. Psychiatry Res. 2006;146:65–72.
Apkarian AV, Sosa Y, Sonty S, Levy RM, Harden RN, Parrish TB, Gitelman DR. Chronic back pain is associated with decreased prefrontal and thalamic gray matter density. J Neurosci. 2004;24:10410–5.
Okada T, Tanaka M, Kuratsune H, Watanabe Y, Sadato N. Mechanisms underlying fatigue: a voxel-based morphometric study of chronic fatigue syndrome. BMC Neurol. 2004;4:14.
de Lange FP, Kalkman JS, Bleijenberg G, Hagoort P, van der Meer JW, Toni I. Gray matter volume reduction in the chronic fatigue syndrome. Neuroimage. 2005;26:777–81.
Schmidt-Wilcke T, Luerding R, Weigand T, Jürgens T, Schuierer G, Leinisch E, Bogdahn U. Striatal grey matter increase in patients suffering from fibromyalgia. A voxel-based morphometry study. J Pain. 2007;132:S109–16.
Obermann M, Rodriguez-Raecke R, Naegel S, Holle D, Mueller D, Yoon MS, Theysohn N, Blex S, Diener HC, Katsarava Z. Gray matter volume reduction reflects chronic pain in trigeminal neuralgia. Neuroimage. 2013;74:352–8.
Henderson LA, Peck CC, Petersen ET, Rae CD, Youssef AM, Reeves JM, Wilcox SL, Akhter R, Murray GM, Gustin SM. Chronic pain: lost inhibition? J Neurosci. 2013;33:7574–82.
Schmidt-Wilcke T, Hierlmeier S, Leinisch E. Altered regional brain morphology in patients with chronic facial pain. Headache. 2010;50:1278–85.
Apkarian AV, Bushnell MC, Treede RD, Zubieta JK. Human brain mechanisms of pain perception and regulation in health and disease. Eur J Pain. 2005;9:463–84.
Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S. The role of the medial frontal cortex in cognitive control. Science. 2004;306:443–7.
Apkarian AV, Baliki MN, Geha PY. Towards a theory of chronic pain. Prog Neurobiol. 2009;87:81–97.
Robinson ME, Craggs JG, Price DD, Perlstein WM, Staud R. Gray matter volumes of pain related brain areas are decreased in fibromyalgia syndrome. Gray matter volumes of pain related brain areas are decreased in fibromyalgia syndrome. J Pain. 2011;12:436–43.
Baliki MN, Geha PY, Apkarian AV. Parsing pain perception between nociceptive representation and magnitude estimation. J Neurophysiol. 2009;101:875–87.
Garcia-Campayo J, Sanz C, Baringo T, Ceballos C. SPECT scan in somatisation disorder patients: an exploratory study of eleven cases. Aust N Z J Psychiatry. 2001;35:359–63.
Chen JJ, Wang JY, Chang YM. Regional cerebral blood flow between primary and concomitant fibromyalgia patients: a possible way to differentiate concomitant fibromyalgia from the primary disease. Scand J Rheumatol. 2007;36:22.
Fields RD. White matter in learning, cognition and psychiatric disorders. Trends Neurosci. 2008;31:317–76.
Sundgren PC, Dong Q, Gómez-Hassan D, Mukherji SK, Maly P, Welsh R. Diffusion tensor imaging of the brain: review of clinical applications. Neuroradiology. 2004;46:339–50.
Hadjipavlou G, Dunckley P, Behrens TE, Tracey I. Determining anatomical connectivities between cortical and brainstem pain processing regions in humans: a diffusion tensor imaging study in healthy controls. Pain. 2006;123:169–78.
Sundgren PC, Petrou M, Harris RE. Diffusion-weighted and diffusion tensor imaging in fibromyalgia patients: a prospective study of whole brain diffusivity, apparent diffusion coefficient, and fraction anisotropy in different regions of the brain and correlation with symptom severity. Acad Radiol. 2007;14:839–46.
Buckner RL, Andrews-Hanna JR, Schacter DL. The brain’s default network: anatomy, function, and relevance to disease. Ann N Y Acad Sci. 2008;1124:1–38.
Biswal B, Yetkin FZ, Haughton VM, Hyde JS. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn Reson Med. 1995;34:537–41.
Beckmann CF, DeLuca M, Devlin JT, Smith SM. Investigations into resting-state connectivity using independent component analysis. Philos Trans R Soc Lond B Biol Sci. 2005;360:1001–13.
Napadow V, LaCount L, Park K, As-Sanie S, Clauw DJ, Harris RE. Intrinsic brain connectivity in fibromyalgia is associated with chronic pain intensity. Arthritis Rheum. 2010;62:2545–55.
Seminowicz DA, Davis KD. Interactions of pain intensity and cognitive load: the brain stays on task. Cereb Cortex. 2007;17:1412–22.
Baliki M, Geha P, Apkarian A, Chialvo D. Impaired brain de-activation in chronic pain. In: Society for Neuroscience Annual Meeting. San Diego: Society for Neuroscience. 2007;825-822/II819.
Pujol J, Macia D, Garcia-Fontanals A, Blanco-Hinojo L, Lopez-Sola M, Garcia-Blanco S, Poca-Dias V, Harrison BJ, Contreras-Rodriguez O, Monfort J, Garcia-Fructuoso F, Deus J. The contribution of sensory system functional connectivity reduction to clinical pain in fibromyalgia. Pain. 2014;155:1492–503.
Cifre I, Sitges C, Fraiman D, Munoz MA, Balenzuela P, Gonzalez-Roldan A, Martinez-Jauand M, Birbaumer N, Chialvo DR, Montoya P. Disrupted functional connectivity of the pain network in fibromyalgia. Psychosom Med. 2012;74:55–62.
Ceko M, Bushnell MC, Fitzcharles MA, Schweinhardt P. Fibromyalgia interacts with age to change the brain. Neuroimage Clin. 2013;3:249–60.
Kim JY, Kim SH, Seo J, Kim SH, Han SW, Nam EJ, Kim SK, Lee HJ, Lee SJ, Kim YT, Chang Y. Increased power spectral density in resting-state pain-related brain networks in fibromyalgia. Pain. 2013;154:1792–7.
Seminowicz DA, Davis KD. Cortical responses to pain in healthy individuals depends on pain catastrophizing. Pain. 2006;120:297–306.
Stoeter P, Bauermann T, Nickel R, Corluka L, Gawehn J, Vucurevic G, Vossel G, Egle UT. Cerebral activation in patients with somatoform pain disorder exposed to pain and stress: an fMRI study. Neuroimage. 2007;36:418–30.
Gundel H, Valet M, Sorg C, Huber D, Zimmer C, Sprenger T, Tölle TR. Altered cerebral response to noxious heat stimulation in patients with somatoform pain disorder. Pain. 2008;137:413–21.
Petzke F, Clauw DJ, Ambrose K, Khine A, Gracely RH. Increased pain sensitivity in fibromyalgia: effects of stimulus type and mode of presentation. Pain. 2003;105:403–13.
Lui F, Duzzi D, Corradini M, Serafini M, Baraldi P, Porro CA. Touch or pain? Spatiotemporal patterns of cortical fMRI activity following brief mechanical stimuli. Pain. 2008;138:362–74.
Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A default mode of brain function. Proc Natl Acad Sci U S A. 2001;98:676–82.
Jorge LL, Amaro Jr E. Brain imaging in fibromyalgia. Curr Pain Headache Rep. 2012;16:388–98.
Sheline YI, Barch DM, Price JL, Rundle MM, Vaishnavi SN, Snyder AZ, Mintun MA, Wang S, Coalson RS, Raichle ME. The default mode network and self-referential processes in depression. Proc Natl Acad Sci U S A. 2009;106:1942–7.
O’Doneel LJ, Schultz T. Statistical and machine learning methods for neuroimaging: examples, challenges, and extensions to diffusion imaging data. In: Hotz I, Schultz T, editors. Visualization and processing of higher order descriptors for multi-valued data, mathematics and visualization. Cham: Springer; 2015.
Liu J, Zhao L, Lei F, Zhang Y, Yuan K, Gong Q, Liang F, Tian J. Disrupted resting-state functional connectivity and its changing trend in migraine suffers. Hum Brain Mapp. 2015;36:1892–907.
Baer RA. Mindfulness training as a clinical intervention: a conceptual and empirical review. Clin Psychol Sci Pract. 2003;10:125–43.
Grossman P, Niemann L, Schmidt S, Walach H. Mindfulness-based stress reduction and health benefits. A meta-analysis. J Psychosom Res. 2004;57:35–43.
Lauche R, Langhorst J, Paul A, Dobos G, Cramer H. Self-reported health and satisfaction of patients with chronic diseases who meditate: a case–control study. Qual Life Res. 2014;23:2639–44.
Lauche R, Cramer H, Häuser W. A systematic overview of reviews for complementary and alternative therapies in the treatment of the fibromyalgia syndrome. Evid Based Complement Alternat Med. 2015;1–13.
Hölzel BK, Ott U, Hempel H, Hackl A, Wolf K, Stark R, Vaitl D. Differential engagement of anterior cingulate and adjacent medial frontal cortex in adept meditators and non-meditators. Neurosci Lett. 2007;421:16–21.
Fox KC, Nijeboer S, Dixon ML, Floman JL, Ellamil M, Rumak SP, Sedlmeier P, Christoff K. Is meditation associated with altered brain structure? A systematic review and meta-analysis of morphometric neuroimaging in meditation practitioners. Neurosci Biobehav Rev. 2014;43C:48–73.
Fayed N, Lopez del Hoyo Y, Andres E, Serrano-Blanco A, Bellón J, Aguilar K, Cebolla A, Garcia-Campayo J. Brain changes in long-term Zen meditators using proton magnetic resonance spectroscopy and diffusion tensor imaging: a controlled study. PLoS One. 2013;8:e58476.
Kang D, Jo HJ, Jung WH, Kim SH, Jung YH, Choi CH, Lee US, An SC, Jang JH, Kwon JS. The effect of meditation on brain structure: cortical thickness mapping and diffusion tensor imaging. Soc Cogn Affect Neurosci. 2013;8:27–33.
Tang YY, Lu Q, Fan M, Yang Y, Posner MI. Mechanisms of white matter changes induced by meditation. Proc Natl Acad Sci. 2012;109:10570–4.
Brewer JA, Worhunsky PD, Gray JR, Tang YY, Weber J, Kober H. Meditation experience is associated with differences in default mode network activity and connectivity. Proc Natl Acad Sci. 2011;108:20254–9.
Hasenkamp W, Barsalou LW. Effects of meditation experience on functional connectivity of distributed brain networks. Front Hum Neurosci. 2012;6:38.
Jang JH, Jung WH, Kang DH, Byun MS, Kwon SJ, Choi CH, Kwon JS. Increased default mode network connectivity associated with meditation. Neurosci Lett. 2011;487:358–62.
Taylor VA, Daneault V, Grant J, Scavone G, Breton E, Roffe-Vidal S, Lavarenne AS, Marrelec G, Benali H, Beauregard M. Impact of meditation training on the default mode network during a restful state. Soc Cogn Affect Neurosci. 2013;8:4–14.
Lutz A, Brefczynski-Lewis J, Johnstone T, Davidson RJ. Regulation of the neural circuitry of emotion by compassion meditation: effects of meditative expertise. PLoS One. 2008;26:e1897.
Khoury B, Lecomte T, Fortin G, Masse M, Therien P, Bouchard V, Hofmann SG. Mindfulness-based therapy: a comprehensive meta-analysis. Clin Psychol Rev. 2013;33:763–71.
Grant JA, Courtemanche J, Rainville P. A non-elaborative mental stance and decoupling of executive and pain-related cortices predicts low pain sensitivity in Zen meditators. Pain. 2011;152:150–6.
Hölzel BK, Lazar SW, Gard T, Schuman-Olivier Z, Vago DR, Ott U. How does mindfulness meditation work? Proposing mechanisms of action from a conceptual and neural perspective. Perpsect Psychol Sci. 2011;6:537–59.
Price DD. Psychological and neural mechanisms of the affective dimension of pain. Science (New York, NY). 2000;288:1769–72.
Creswell JD, Way BM, Eisenberger NI, Lieberman MD. Neural correlates of dispositional mindfulness during affect labeling. Psychosom Med. 2007;69:560–5.
Harris RE, Clauw DJ, Scott DJ, McLean SA, Gracely RH, Zubieta JK. Decreased central mu-opioid receptor availability in fibromyalgia. J Neurosci. 2007;27:10000e6.
Arienzo D, Babiloni C, Ferretti A, Caulo M, Del Gratta C, Tartaro A, Rossini PM, Romani GL. Somatotopy of anterior cingulate cortex (ACC) and supplementary motor area (SMA) for electric stimulation of the median and tibial nerves: an fMRI study. Neuroimage. 2006;33:700–5.
Goldin PR, Gross JJ. Effects of mindfulness-based stress reduction (MBSR) on emotion regulation in social anxiety disorder. Emotion. 2010;10:83–91.
Napadow V, Kim J, Clauw DJ, Harris RE. Decreased intrinsic brain connectivity is associated with reduced clinical pain in fibromyalgia. Arthritis Rheum. 2012;64:2398–403.
Harris RE, Napadow V, Huggins JP, Pauer L, Kim J, Hampson J, Sundgren PC, Foerster B, Petrou M, Schmidt-Wilcke T, Clauw DJ. Pregabalin rectifies aberrant brain chemistry, connectivity, and functional response in chronic pain patients. Anesthesiology. 2013;119:1453–64.
Schmidt-Wilcke T, Ichesco E, Hampson JP, Kairys A, Peltier S, Harte S, Clauw DJ, Harris RE. Resting state connectivity correlates with drug and placebo response in fibromyalgia patients. Neuroimage Clin. 2014;6:252–61.
Hashmi JA, Baria AT, Baliki MN, Huang L, Schnitzer TJ, Apkarian AV. Brain networks predicting placebo analgesia in a clinical trial for chronic back pain. Pain. 2012;153:2393–402.
Sevel LS, O'Shea AM, Letzen JE, Craggs JG, Price DD, Robinson ME. Effective connectivity predicts future placebo analgesic response: a dynamic causal modeling study of pain processing in healthy controls. Neuroimage. 2015;110:87–94.
Olivan-Blázquez B, Herrera-Mercadal P, Puebla-Guedea M, Pérez-Yus MC, Andrés E, Fayed N, López-Del-Hoyo Y, Magallon R, Roca M, Garcia-Campayo J. Efficacy of memantine in the treatment of fibromyalgia: a double-blind, randomised, controlled trial with 6-month follow-up. Pain. 2014;155:2517–25.
Fayed N, Olivan-Blázquez B, Herrera-Mercadal P, Puebla-Guedea M, Pérez-Yus MC, Andrés E, López del Hoyo Y, Magallon R, Viguera L, Garcia-Campayo J. Changes in metabolites after treatment with memantine in fibromyalgia. A double-blind randomized controlled trial with magnetic resonance spectroscopy with a 6-month follow-up. CNS Neurosci Ther. 2014;20:999–1007.
O'Muircheartaigh J, Marquand A, Hodkinson DJ, Krause K, Khawaja N, Renton TF, Huggins JP, Vennart W, Williams SC, Howard MA. Multivariate decoding of cerebral blood flow measures in a clinical model of on-going postsurgical pain. Hum Brain Mapp. 2015;36:633–42.
Baliki MN, Petre B, Torbey S, Herrmann KM, Huang L, Schnitzer TJ, Fields HL, Apkarian AV. Corticostriatal functional connectivity predicts transition to chronic back pain. Nat Neurosci. 2012;15:1117–9.
Mansour AR, Baliki MN, Huang L, Torbey S, Herrmann KM, Schnitzer TJ, Apkarian AV. Brain white matter structural properties predict transition to chronic pain. Pain. 2013;154:2160–8.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Miguel, N.F., Garcia-Campayo, J., González-Toledo, E., Viguera, L. (2017). Neuroimaging in Chronic Pain, Fibromyalgia, and Somatization. In: Gargiulo, P., Mesones-Arroyo, H. (eds) Psychiatry and Neuroscience Update - Vol. II. Springer, Cham. https://doi.org/10.1007/978-3-319-53126-7_30
Download citation
DOI: https://doi.org/10.1007/978-3-319-53126-7_30
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-53125-0
Online ISBN: 978-3-319-53126-7
eBook Packages: MedicineMedicine (R0)