
Abstract
The concept of reserve has been developed to account for the discontinuity between the extent of brain damage at its clinical manifestation in the form of cognitive decline or dementia. In this chapter, we discuss contributors to cognitive reserve from various stages of the life-course, including childhood, early adulthood, middle age, and late life. Evidence from observational, as well as intervention trials is presented and assessed. We conclude by arguing that reserve formation in dementia risk is a life-course process whereby baseline cognitive abilities are subjected to modulation by subsequent experiences at diverse stages over the entire life-course. Variations among individuals in their ability to withstand age-related brain changes is ultimately dependent on their life-time accumulation of mental, physical, and lifestyle inputs into cognitive reserve.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
- Reserve
- Cognitive reserve
- Brain reserve
- Dementia
- Life-course
- Education
- Occupational attainment
- Childhood cognitive ability
- Social networks
- Leisure activities
5.1 Introduction
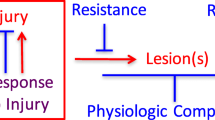
It has been reported that about a quarter to two-thirds of people characterized as cognitively intact throughout longitudinal assessments in fact fulfil pathological criteria for dementia at autopsy (Neuropathology Group 2001; Steffener and Stern 2012; Crystal et al. 1988; Morris et al. 1996; Price and Morris 1999; Mortimer et al. 2003). These findings provided an empirical underpinning for the well-established regularity in clinical practice that the same degree of head injury, or stroke, often leads to drastically different levels of cognitive impairment across individuals. As a result, the concept of reserve was developed to account for the discontinuity between the extent of brain damage and its clinical manifestation (Stern 2002, 2009). Reserve hypothesis postulates that functioning may be maintained in the presence of brain changes or insults due to the aspects of brain structure and function that can buffer the effects of neuropathology. Therefore, the greater the reserve, the more severe is the pathology required to manifest as functional impairment (Richards et al. 2004). The distinction between brain structure and function in the definition of reserve has led to the subdivision of the concept into two components: brain reserve and cognitive reserve, each emphasizing distinctive aspects of protection against the mounting pathology.
Brain reserve encompasses protection from cognitive impairment or dementia due to anatomical features of the brain, such as size, neural count, or synaptic connectivity (Katzman 1993). The brain reserve hypothesis argues for a critical point of pathology, beyond which any further deterioration will invariably lead to functional impairment (Satz 1993). Individuals with larger brains, more neurons, or better-connected synapses should be able to sustain more insults prior to the clinical manifestation. The brain reserve hypothesis is also known as the passive model because it assumes that the effects of a given insult will lead to identical effects across individuals and that the eventual differences are only due to the brain reserve capacity—the damage is either sufficient or insufficient to deplete this capacity beyond a fixed critical level. This model, therefore, does not account for individual differences in brain network efficiency or flexibility, which might exist even at identical levels of brain size or synaptic count.
Cognitive reserve is a concept related to brain reserve, in a way that it is also meant to help cope with underlying brain damage. The difference is that cognitive reserve emphasizes efficiency and resilience of the brain networks rather than the biological characteristics of these networks (Stern 2009). Efficiency—brain network’s ability to operate effectively despite sustained disruption, and flexibility—recruitment of alternate networks when faced with disruption to the standard approaches, are the centrepieces of the cognitive reserve model. In contrast to the brain reserve model, a fixed threshold for functional impairment is not assumed in the framework of cognitive reserve—instead, the critical threshold varies across individuals as a function of how efficient or resilient the brain networks are at utilizing the remaining neuron connections. Therefore, the effects of brain damage might vary across individuals depending on their ability to cope with the damage, even if the underlying brain reserve capacity is held constant.
The concept of reserve has been called upon to account for a well-established finding in epidemiological literature that high educational (Katzman 1993; Meng and D’Arcy 2012) and occupational attainment (Qiu et al. 2003; Stern et al. 1994) appear to protect against cognitive decline and dementia. Explaining this finding using the predictions of the brain reserve hypothesis would imply a conclusion that the brains of highly educated individuals are considerably different in terms of anatomical features than the brains of individuals with less advanced education. The cognitive reserve explanation provides a more plausible explanation: people with higher educational or occupational attainment are able to process tasks in a more efficient manner because their brains actively attempt to cope with brain damage by using pre-existing cognitive processing approaches, or by activating compensatory mechanisms (Fratiglioni and Wang 2007)—and high education or occupational attainment might indicate the extent to which these strategies will be successfully used across individuals.
Both brain reserve and cognitive reserve models are closely related to the general concept of brain plasticity (Mahncke et al. 2006)—the ability of the brain to re-arrange structure as well as neural connections, allowing it to change with learning, to repair, and to compensate. Plasticity is central to memory foundation and learning processes because it provides the brain with a life-long possibility to change and adjust when facing environmental demands and stimuli. The focus on life-long development is important for the cognitive reserve construct in particular, which, as an active process, is likely formed as a combination of baseline capacity, which is then subjected to modulation by multiple experiences and exposures throughout the entire life-course. Through this life-long process an individual develops a level of cognitive reserve which in turn can mitigate the effects of pathology on the clinical diagnosis later in life.
In this chapter, we will explore the life-course perspective to cognitive reserve in the framework of cognitive decline and dementia in greater detail. We start out by laying out some well-established epidemiological evidence in favour of the cognitive reserve hypothesis, with particular emphasis on reserve contributors from various stages of the life-course, including childhood. For each reserve contributor, we also discuss potential neural and cellular mechanisms that could account for the reported findings. We proceed by presenting a tentative model of cognitive reserve over the life-course, which emphasizes various reserve components and contributors from different points throughout the life-span. Conclusions are offered at the end of the chapter.
5.2 Early Adulthood: Educational Attainment
Evidence of an elevated risk of dementia in people with low educational attainment is cited most often in support of a cognitive reserve hypothesis for late-life dementia and cognitive decline. The evidence is so abundant and consistent that the recently discovered decline in dementia prevalence in the West has been in part attributed to expanding educational opportunities in rich industrialized societies (Langa 2015). A recent meta-analysis (Meng and D’Arcy 2012) has found more than 100 studies reporting either protective effects of high education or detrimental effects of low attainment on the risk of dementia from a wide variety of contexts. Most often, an explanation of these effects involves a reference to the general cognitive reserve hypothesis, emphasizing brain networks efficiencies and flexibilities, whereby individuals with high attainment are suspected to have more efficient brain networks which remain operational even under the burden of pathology, or are able to re-distribute tasks away from the damaged areas in favour of the networks unaffected by the pathology (Herlitz and Dekhtyar 2013).
An alternative to the cognitive reserve explanation behind the association between education and late-life cognitive decline or dementia is the so-called brain-battering hypothesis (Del Ser et al. 1999). It builds on the notion that well-educated individuals are more prone to engage in healthier lifestyles associated with a lower burden of vascular and behavioural risks (Gottfredson 2004), which are then implicated in late-life dementia risk (Norton et al. 2014). Therefore, any eventual association between education and cognitive decline or dementia is not due to education mitigating the impact of pathology on the clinical expression of the disease, but rather due to education protecting individuals from developing vascular neuropathology which, ultimately, is the cause of dementia. The brain-battering hypothesis was first used to account for a peculiar finding that individuals with low education were both diagnosed as demented and eventually died earlier than the highly educated ones, although they did not differ in the rate of cognitive decline or the neurodegenerative burden; the only difference between the two groups was in terms of cerebrovascular pathology (Del Ser et al. 1999). Further, lifestyle factors, such as smoking, were found to alter the association between education and non-Alzheimer disease dementia (Cobb et al. 1995), suggesting mediation by vascular factors. Finally, mixed findings have been reported between education and measures of brain pathology or structure, casting doubt on the model of cognitive reserve (Roe et al. 2008; Christensen et al. 2007; Koepsell et al. 2008), although there have been issues with representativeness, sample size, and selection of pathological measures in these studies. Another possibility for the association between education and dementia is the so-called detection bias, suggesting that subjects with dementia who are less-educated would be diagnosed at an earlier pathological stage of the disease as compared to those with higher education. This is especially relevant since some studies have found that low education has been associated with an elevated incidence of dementia, whereas at the same time no association has been established between low education and the rate of memory decline or mortality of dementia patients (Qiu et al. 2001; Tuokko et al. 2003).
Although detection bias and the earlier-discussed brain-battering hypothesis are both legitimate alternatives to the general theory of cognitive reserve in explaining the link between education and cognitive decline, increasing evidence is pointing to the existence of a neural basis for cognitive reserve in dementia risk. The risk of Alzheimer’s disease was more than halved in the group with higher educational level, even after adjustment for demographic, socioeconomic, lifestyle, and vascular characteristics in a large Finnish study (Ngandu et al. 2007). Similarly, a large study based on harmonization of longitudinal clinical data and neuropathology from three long-standing population-based studies that included post-mortem donation of 872 brains (Brayne et al. 2010) has shown that education did not protect against the accumulation of neurodegenerative or vascular pathologies in the brain at death. It did, however, mitigate the association between pathology burden and cognitive decline, so that for a specific pathological burden, those with more education were at a lower risk of dementia in late life. Considering the fact that evidence has been derived from relatively large sample size, population-based nature of the data, and longitudinal assessments, it is becoming increasingly likely that that the association between low education and increased incidence of dementia is not due to an increased burden of neuropathology, but rather due to increased vulnerability of less-educated individuals to cognitive deterioration due to neuropathological insults (Brayne 2010).
5.3 Mid-Life: Occupational Attainment and Mental Activity
While education is viewed as a contributor to cognitive reserve from early adulthood, there has also been considerable interest in epidemiological literature in reserve contributors from mid-life, of which occupational attainment has been one of the most prominent (Karp et al. 2009; Andel et al. 2005). It is believed that demanding occupational roles enhance cognitive reserve by maintaining intellectual flexibility even into the ages when cognitive function is otherwise expected to decline (Schooler 2004). Furthermore, prolonged mental stimulation at work prevents disuse and subsequent atrophy of cognitive processes and skills (Fratiglioni et al. 2004), as predicted by the “use it or lose it” hypothesis (Salthouse 1991).
A number of epidemiological studies have found that individuals in manual occupations are at an elevated risk of poor cognitive performance (Dartigues et al. 1992) and dementia (Qiu et al. 2003; Smyth et al. 2004). One of the explanations offered to account for these findings concerns differences between occupations in the extent of complexity of tasks an individual performs daily, with substantively complex occupational roles expected to provide the most demanding practice and exercise (Schooler et al. 1999). The work complexity aspect has been subsequently assessed in a number of epidemiological studies, with occupational tasks graded along the broad dimensions of complexity with data, people, and things (Karp et al. 2009; Andel et al. 2006; Kröger et al. 2008). Results from these studies have indicated that individuals in occupational roles requiring expertise in tasks dealing with people and data are at a reduced risk of poor cognitive functioning (Smart et al. 2014) and dementia in late life (Dekhtyar et al. 2015a). These effects are not explained by shared genetic- or early-life environmental confounding as demonstrated in a co-twin study (Andel et al. 2005).
Similarly to the effects of education, several imaging and autopsy-verified studies have generally corroborated the findings reported in epidemiological literature, suggesting the existence of a neural basis for the relationship between occupational attainment and cognitive decline and dementia. For instance, it has been shown that higher occupational attainment was associated with a greater pathologic burden (lower regional cerebral blood flow in medial frontal cortex and left dorsolateral frontal cortex) in patients with frontotemporal dementia (FTD), suggesting a reserve pathway because increased pathologic load must have been initially present for clinical symptoms to appear (Borroni et al. 2009). Furthermore, FTD patients with occupations requiring advanced verbal abilities also had reduced regional glucose utilization in the left inferior frontal gyrus, whereas those with more physically demanding roles at work displayed reduced metabolic rate of glucose utilization in the supplementary motor area (Spreng et al. 2011), further indicating that a neural basis might underlie the effects. A recent study has also shown that survival was prolonged in autopsy-confirmed FTD patients with higher occupational attainment (Massimo et al. 2015). Finally, in the context of AD, controlling for clinical dementia severity, less relative perfusion has been found in the parietal region in individuals whose occupations were characterized by higher interpersonal skills (Stern et al. 1995).
Whereas the association between occupational complexity and dementia has mostly been attributed to cognitive reserve pathways emphasizing neural efficiency or neural compensation, it is also possible that other mechanisms are also implicated. Some studies have suggested the detrimental effects of work stress could also be at play. A study from Sweden found that low job control (in both demanding and non-demanding jobs) was associated with an elevated risk of dementia, even after accounting for vascular factors (Wang et al. 2012b).
5.4 Late Life: Social Networks and Leisure Activities
Although the effects of mid-life cognitive stimulation through complex occupational demands is well-established, a number of studies have suggested that late-life stimulating activities can serve as an additional line of defence against cognitive decline and dementia. Of these stimulating activities, the effects of social networks and leisure activities have been particularly emphasized. Living alone and having limited social contacts have been found to be associated with more than a 50% increase in the risk of dementia in later life in a community-based cohort of 1203 non-demented individuals aged 75 or more from Stockholm, Sweden (Fratiglioni et al. 2004). Interestingly, infrequent social contacts with network resources have not been found to elevate the risk of dementia, provided those contacts were deemed as satisfying. A later study using a more detailed assessment of social networks based on the validated Lubben Social Network Scale reported largely comparative findings, albeit in an exclusively female study population (Crooks et al. 2008). Several pathways have been suggested to account for the findings, including the possible precipitating effects of limited social contacts in situations when pathological alterations are already in place, akin to the reserve model predictions (Fratiglioni et al. 2000). A biological mechanism could also be at work, with previous literature suggesting a link between social integration and immune system (Seeman 1996), whereas both degenerative and vascular dementia also exhibit considerable inflammatory components.
More recent studies into the relationship between social networks and dementia have explored specific activity dimensions, with leisure activities receiving considerable interest. A systematic review has found that 52 original articles linking leisure activities (mental, physical, and social components) and dementia have been published between 1991 and 2011 (Wang et al. 2012a). Observational studies are mostly consistent in indicating beneficial effects of physical (Tolppanen et al. 2015), mental (Karp et al. 2006), and social leisure activities (Paillard-Borg et al. 2009; Verghese et al. 2003) on dementia. Importantly, various leisure activities could have a different effect on the pattern of cognitive decline in old age. Thus, a recent study using detailed data from China has reported that in almost 1500 men and women aged 65 and older, higher levels of mental ability were associated with slower decline in global cognition, language, and executive function. In contrast, higher levels of physical leisure activities were related with less decline in episodic memory and language, with a clear dose-response pattern observable across both mental and physical leisure types (Wang et al. 2013). While observational studies are mostly consistent at indicating protective effects of physical and mental abilities on dementia, randomized control trials have so far lacked similar consistency (Wang et al. 2012a), although some promising findings been reported in a trial of a multi-domain intervention consisting of diet, exercise, cognitive training, and vascular monitoring (Ngandu et al. 2015). Similarly, transitioning from a sedentary lifestyle to moderate physical activity has been shown to have a beneficial effect on cognitive functioning and dementia (Lovden et al. 2013). In general, more research is required to unequivocally conclude that stimulating (socially, physically, and mentally) lifestyles in old age provide an additional line of defence, over and above of that already developed through educational or occupational stimulation.
5.5 Early Life: Childhood Cognitive Ability
Although education, a contributor to cognitive reserve from early adulthood, occupational attainment, a contributor from mid-life, and social networks/leisure activities from late life have all been implicated, to various degrees, in late-life cognitive decline and dementia, a number of questions have remained. One of the most pertinent issues concerns the directionality of the effects of education and occupational attainment: are mentally stimulating lifestyles true risk factors for cognitive decline and dementia, or are their effects simply a consequence of differences in prior cognitive ability? The former possibility, known in the literature as the differential preservation hypothesis, suggests that intellectual stimulation and development of mental abilities can in fact delay age-related decline—therefore the extent to which either factor (higher education or occupation) is present, differentially affects the trajectory of cognitive change and decline (Salthouse 2006; Gow et al. 2014). In contrast, the preserved differentiation hypothesis argues that educational and occupational effects on cognitive functioning in late life are ultimately confounded by differences in baseline cognitive abilities (Smart et al. 2014).
The possibility that early-life cognitive abilities might be implicated in late-life cognitive decline and dementia has been suggested in the literature previously, although testing the hypothesis proved difficult due to considerable data limitations. For instance, an investigation of autobiographies written by 22-year old nuns has revealed that those nuns who would later develop also had lower linguistic ability, which served as a marker of intelligence, as opposed to the nuns who would not develop the disease (Snowdon et al. 1996). A later study based on the same data indicated that low early-life linguistic ability was also related to lower brain weight, higher degree of cerebral atrophy, more advanced neurofibrillary pathology, and neuropathologic criteria for AD (Riley et al. 2005). Although a seminal study that provided tentative support for the hypothesis that early-life cognitive ability might underlie the risk of late-life decline and dementia, the Nun study had limitations: its population was not representative and the sample size was small (especially in the neuropathological study), which means that the issue of preserved differentiation versus differential preservation of cognitive abilities throughout the life-course remained unaddressed.
Some of the more comprehensive investigations of the effects of early cognitive ability on late-life cognitive ageing have come from the data on Scottish children (known as the Lothian cohorts), born in the 1920–1930s who took an identical cognitive test at the age of 11, and were subsequently enrolled in a study of cognitive ageing after the age of 70 (Deary et al. 1921). It has been reported that childhood intelligence was not associated with early-onset dementia, whereas differences by early-life intelligence did exist in the case of late-onset dementia (Whalley et al. 2000). A subsequent study on the same material revealed that lower childhood IQ was a risk factor for vascular dementia, but not Alzheimer disease, suggesting potential mechanisms via the vascular pathology, akin to the brain-battering hypothesis (McGurn et al. 2008). A recent study has explicitly examined the relative importance of the preserved differentiation vs. the differential preservation hypotheses using the Lothian cohorts’ data, although in the context of cognitive ageing and not dementia. It was shown that complex occupational roles in mid-life were associated with improved cognitive performance in later life, even after controlling for IQ at age 11 (Smart et al. 2014). On the one hand, these results confirmed the differential preservation hypothesis, suggesting that engagement in complex environments in later life may help preserve cognitive function in advanced ages. On the other hand, since the effect size of occupational complexity was reduced by more than half once childhood intelligence was accounted for, it appears that engagement in complex activities is also partly a consequence of earlier cognitive abilities, as would be predicted by the preserved differentiation hypothesis.
It is, however, important to note that early-life cognitive ability may be associated with cognitive ageing not only by virtue of life-long stability of cognitive abilities, i.e. by affecting the level of functioning in old age, but also by influencing the rate of decline. Longitudinal assessments of cognitive ageing are not uncommon, although only a handful of studies also managed to collect early-life ability measures, and contradictory findings have been reported thus far. For instance, the latent growth curve analysis of the Lothian cohort data has shown that intelligence at age 11 was associated with the baseline level of cognitive ability at age 79, but not with the rate of decline 8 years later (Gow et al. 2011). On the other hand, childhood cognitive ability was associated with both the level as well as the change in cognitive abilities between ages 43–53 in the National Survey of Health and Development data (Gow et al. 2012). Although more studies are needed to clarify the issue, it appears that early-life ability can affect late-life cognitive declines, and not just the levels of ability, although this may not happen uniformly across all stages of the ageing process.
One way of contributing to the debate about the relative importance of preserved differentiation versus differential preservation in the context of cognitive ageing over the entire life-course would be by examining whether the late-life risk of dementia is reduced in individuals with high education or in complex occupation, after childhood cognitive ability is taken into account. Only two studies so far has been able to combine information on childhood cognitive ability, educational attainment from early adulthood, and life-time occupational complexity in a life-course model of cognitive reserve in dementia (Dekhtyar et al. 2015a).
A large population-based study from Uppsala, Sweden has followed more than 7500 individuals aged 65 for 29 years to detect their incident dementia. To capture childhood cognitive ability, school grades from the third year of elementary school (age 10) were extracted from the school archives, while information was also collected on formal education, and life-time occupational attainment. It was reported that participants with the lowest 20% of school grades from age 10 were at an elevated risk of dementia, relative to the rest of the population (HR: 1.21, p < 0.05). Effects of education on dementia disappeared once early-life school performance was accounted for, whereas higher occupational complexity with data preserved its association with a reduced risk of dementia (Dekhtyar et al. 2015a). These findings were later confirmed in a study of about 450 men and women aged 75 or more who underwent detailed neuropsychological assessment, as opposed to the inpatient diagnosis which was used in the study from Uppsala. An even greater risk of dementia (HR: 1.5; p < 0.05) was found in individuals with the lowest childhood school grades. These findings indicate that baseline cognitive abilities have a long-term effect on the risk of dementia; this risk can be somewhat modulated by subsequent stimulating work environments, although initial abilities appear more decisive for the late-life risk of dementia.
5.6 Life-Course Model of Reserve
An earlier discussion has indicated that factors influencing the development of reserve are located at various stages of individual’s life-course. In the case of dementia risk in old age, pre-morbid cognitive ability is of outmost importance, since it modifies the clinical expression of pathology (Richards and Deary 2005). The fundamentals of pre-morbid cognitive abilities are the current elements of brain structure (brain reserve) and function (cognitive reserve), with the former emphasizing inputs based on structural neural network complexity, while the latter targeting the functional processing efficiency and flexibility. Ultimately, both structural and functional inputs are in turn influenced by a variety of factors originating as early as childhood, if not before, considering a wealth of literature of genetic (Deary et al. 2004) and early-environmental influences on adult functioning (Dekhtyar et al. 2015b). We have identified childhood cognitive ability, education, occupational complexity, and late-life leisure activity here as some of the most pertinent inputs into the model of reserve over the life-course, but other influencing factors clearly contribute, including health behaviours, socioeconomic environment, and lifestyles (Richards and Deary 2005). Therefore, although most of neuropathology most likely applies to the older ages, the factors allowing the toleration of this pathology occur throughout the entire life-course.
In conclusion, based on the evidence presented in this chapter, it is becoming increasingly clear that the development of reserve that can mitigate the impact of pathology on the clinical expression of disease occurs at various stages throughout the life-course. Baseline cognitive abilities lay the foundation of reserve formation, which is subsequently enhanced by intellectual stimulation provided by educational attainment. By exerting continued demands on the brain, occupational tasks preserve this acquired buffer, much the same way as late-life social engagement and rewarding leisure activities. Ultimately, cognitive reserve can be conceived as a sum of its mental, physical, and lifestyle inputs over the entire life course, and the brain’s ability to withstand the changes associated with ageing will to a large extent reflect the gradual accumulation of these inputs.
References
Andel R, Crowe M, Pedersen NL, Mortimer J, Crimmins E, Johansson B, et al. Complexity of work and risk of Alzheimer’s disease: a population-based study of Swedish twins. J Gerontol B Psychol Sci Soc Sci. 2005;60(5):P251–8.
Andel R, Vigen C, Mack WJ, Clark LJ, Gatz M. The effect of education and occupational complexity on rate of cognitive decline in Alzheimer’s patients. J Int Neuropsychol Soc. 2006;12(01):147–52.
Borroni B, Premi E, Agosti C, Alberici A, Garibotto V, Bellelli G, et al. Revisiting brain reserve hypothesis in frontotemporal dementia: evidence from a brain perfusion study. Dement Geriatr Cogn Disord. 2009;28(2):130–5.
Brayne C, Ince PG, Keage HAD, McKeith IG, Matthews FE, Polvikoski T, et al. Education, the brain and dementia: neuroprotection or compensation? Brain. 2010;2210–6.
Christensen H, Anstey KJ, Parslow RA, Maller J, Mackinnon A, Sachdev P. The brain reserve hypothesis, brain atrophy and aging. Gerontology. 2007;53(2):82–95.
Cobb J, Wolf PA, Au R, White R. D’agostino R. The effect of education on the incidence of dementia and Alzheimer’s disease in the Framingham study. Neurology. 1995;45(9):1707–12.
Crooks VC, Lubben J, Petitti DB, Little D, Chiu V. Social network, cognitive function, and dementia incidence among elderly women. Am J Public Health. 2008;98(7):1221–7.
Crystal H, Dickson D, Fuld P, Masur D, Scott R, Mehler M, et al. Clinico‐pathologic studies in dementia nondemented subjects with pathologically confirmed Alzheimer’s disease. Neurology. 1988;38(11):1682.
Dartigues J-F, Gagnon M, Letenneur L, Barberger-Gateau P, Commenges D, Evaldre M, et al. Principal lifetime occupation and cognitive impairment in a French elderly cohort (Paquid). Am J Epidemiol. 1992;135(9):981–8.
Deary IJ, Whiteman MC, Pattie A, Starr JM, Hayward C, Wright AF, et al. Apolipoprotein e gene variability and cognitive functions at age 79: a follow-up of the Scottish mental survey of 1932. Psychol Aging. 2004;19(2):367.
Deary IJ, Gow AJ, Pattie A, Starr JM. Cohort profile: the Lothian birth cohorts of 1921 and 1936. Int J Epidemiol. 2012;41(6):1576–84.
Dekhtyar S, Wang HX, Scott K, Goodman A, Koupil I, Herlitz A. A life-course study of cognitive reserve in dementia-from childhood to old age. Am J Geriatr Psychiatry. 2015a;23(9):885–96.
Dekhtyar S, Wang H-X, Scott K, Goodman A, Koupil I, Herlitz A. Associations of head circumference at birth with early life school performance and later-life occupational prestige. Longitudinal Life Course Stud. 2015b;6(1):26–42.
Del Ser T, Hachinski V, Merskey H, Munoz DG. An autopsy-verified study of the effect of education on degenerative dementia. Brain. 1999;122:2309–19.
Fratiglioni L, Wang HX. Brain reserve hypothesis in dementia. J Alzheimer’s Dis. 2007;12(1):11–22.
Fratiglioni L, Wang H-X, Ericsson K, Maytan M, Winblad B. Influence of social network on occurrence of dementia: a community-based longitudinal study. Lancet. 2000;355(9212):1315–9.
Fratiglioni L, Paillard-Borg S, Winblad B. An active and socially integrated lifestyle in late life might protect against dementia. Lancet Neurol. 2004;3(6):343–53.
Gottfredson LS. Intelligence: is it the epidemiologists’ elusive “fundamental cause” of social class inequalities in health? J Pers Soc Psychol. 2004;86(1):174.
Gow AJ, Johnson W, Pattie A, Brett CE, Roberts B, Starr JM, et al. Stability and change in intelligence from age 11 to ages 70, 79, and 87: the lothian birth cohorts of 1921 and 1936. Psychol Aging. 2011;26(1):232–40.
Gow AJ, Johnson W, Mishra G, Richards M, Kuh D, Deary IJ. Is age kinder to the initially more able?: yes, and no. Intelligence. 2012;40(1):49–59.
Gow AJ, Avlund K, Mortensen EL. Occupational characteristics and cognitive aging in the glostrup 1914 cohort. J Gerontol Ser B Psychol Sci Soc Sci. 2014;69(2):228–36.
Herlitz A, Dekhtyar S. A life-span approach to dementia. In: Nilsson L-G, Ohta N, editors. Dementia and memory. Hove and New York: Psychology Press; 2013. p. 110–23.
Karp A, Paillard-Borg S, Wang H-X, Silverstein M, Winblad B, Fratiglioni L. Mental, physical and social components in leisure activities equally contribute to decrease dementia risk. Dement Geriatr Cogn Disord. 2006;21(2):65–73.
Karp A, Andel R, Parker MG, Wang H-X, Winblad B, Fratiglioni L. Mentally stimulating activities at work during midlife and dementia risk after age 75: follow-up study from the Kungsholmen project. Am J Geriatr Psychiatry. 2009;17(3):227–36.
Katzman R. Education and the prevalence of dementia and Alzheimer’s disease. Neurology. 1993;43(1):13–20.
Koepsell TD, Kurland BF, Harel O, Johnson EA, Zhou XH, Kukull WA. Education, cognitive function, and severity of neuropathology in Alzheimer disease. Neurology. 2008;70(19 Pt 2):1732–9.
Kröger E, Andel R, Lindsay J, Benounissa Z, Verreault R, Laurin D. Is complexity of work associated with risk of dementia? the Canadian study of health and aging. Am J Epidemiol. 2008;167(7):820–30.
Langa KM. Is the risk of Alzheimer’s disease and dementia declining? Alzheimers Res Ther. 2015;7(1):34.
Lovden M, Xu W, Wang HX. Lifestyle change and the prevention of cognitive decline and dementia: what is the evidence? Curr opin psychiatry. 2013;26(3):239–43.
Mahncke HW, Bronstone A, Merzenich MM. Brain plasticity and functional losses in the aged: scientific bases for a novel intervention. Prog Brain Res. 2006;157:81–109.
Massimo L, Zee J, Xie SX, McMillan CT, Rascovsky K, Irwin DJ, et al. Occupational attainment influences survival in autopsy-confirmed frontotemporal degeneration. Neurology. 2015;84(20):2070–5.
McGurn B, Deary IJ, Starr JM. Childhood cognitive ability and risk of late-onset Alzheimer and vascular dementia. Neurology. 2008;71(14):1051–6.
Meng X, D’Arcy C. Education and dementia in the context of the cognitive reserve hypothesis: a systematic review with meta-analyses and qualitative analyses. PLoS ONE. 2012;7(6):e38268.
Morris JC, Storandt M, McKeel DW Jr, Rubin EH, Price JL, Grant EA, et al. Cerebral amyloid deposition and diffuse plaques in “normal” aging: Evidence for presymptomatic and very mild Alzheimer’s disease. Neurology. 1996;46(3):707–19.
Mortimer JA, Snowdon DA, Markesbery WR. Head circumference, education and risk of dementia: findings from the nun study. J Clin Exp Neuropsychol. 2003;25(5):671–9.
Neuropathology Group. Pathological correlates of late-onset dementia in a multicentre, community-based population in England and Wales. Neuropathology Group of the Medical Research Council Cognitive Function and Ageing Study (MRC CFAS). Lancet. 2001;357(9251):169–75.
Ngandu T, von Strauss E, Helkala EL, Winblad B, Nissinen A, Tuomilehto J, et al. Education and dementia: what lies behind the association? Neurology. 2007;69(14):1442–50.
Ngandu T, Lehtisalo J, Solomon A, Levälahti E, Ahtiluoto S, Antikainen R, et al. A 2 year multidomain intervention of diet, exercise, cognitive training, and vascular risk monitoring versus control to prevent cognitive decline in at-risk elderly people (FINGER): a randomised controlled trial. Lancet. 2015;385(9984):2255–63.
Norton S, Matthews FE, Barnes DE, Yaffe K, Brayne C. Potential for primary prevention of Alzheimer’s disease: an analysis of population-based data. Lancet Neurol. 2014;13(8):788–94.
Paillard-Borg S, Fratiglioni L, Winblad B, Wang HX. Leisure activities in late life in relation to dementia risk: principal component analysis. Dement Geriatr Cogn Disord. 2009;28(2):136–44.
Price JL, Morris JC. Tangles and plaques in nondemented aging and “preclinical” Alzheimer’s disease. Ann Neurol. 1999;45(3):358–68.
Qiu C, Backman L, Winblad B, Aguero-Torres H, Fratiglioni L. The influence of education on clinically diagnosed dementia incidence and mortality data from the Kungsholmen project. Arch Neurol. 2001;58(12):2034–9.
Qiu C, Karp A, von Strauss E, Winblad B, Fratiglioni L, Bellander T. Lifetime principal occupation and risk of Alzheimer’s disease in the Kungsholmen project. Am J Ind Med. 2003;43(2):204–11.
Richards M, Deary IJ. A life course approach to cognitive reserve: a model for cognitive aging and development? Ann Neurol. 2005;58(4):617–22.
Richards M, Shipley B, Fuhrer R, Wadsworth M. Cognitive ability in childhood and cognitive decline in mid-life: longitudinal birth cohort study. BMJ. 2004;328(7439):552.
Riley KP, Snowdon DA, Desrosiers MF, Markesbery WR. Early life linguistic ability, late life cognitive function, and neuropathology: findings from the nun study. Neurobiol Aging. 2005;26(3):341–7.
Roe CM, Xiong C, Miller JP, Cairns NJ, Morris JC. Interaction of neuritic plaques and education predicts dementia. Alzheimer Dis Assoc Disord. 2008;22(2):188–93.
Salthouse TA. Theoretical perspectives on cognitive aging. Psychology Press; 1991.
Salthouse TA. Mental exercise and mental aging: evaluating the validity of the “Use It or Lose It” hypothesis. Perspect Psychol Sci. 2006;1(1):68–87.
Satz P. Brain reserve capacity on symptom onset after brain injury: a formulation and review of evidence for threshold theory. Neuropsychology. 1993;7(3):273.
Schooler C, Mulatu MS, Oates G. The continuing effects of substantively complex work on the intellectual functioning of older workers. Psychol Aging. 1999;14(3):483.
Schooler C, Mulatu MS, Oates G. Occupational self‐direction, intellectual functioning, and self‐directed orientation in older workers: findings and implications for individuals and societies. Am J Sociol. 2004;110(1):161–97.
Seeman TE. Social ties and health: the benefits of social integration. Ann Epidemiol. 1996;6(5):442–51.
Smart EL, Gow AJ, Deary IJ. Occupational complexity and lifetime cognitive abilities. Neurology. 2014;83(24):2285–91.
Smyth KA, Fritsch T, Cook TB, McClendon MJ, Santillan CE, Friedland RP. Worker functions and traits associated with occupations and the development of AD. Neurology. 2004;63(3):498–503.
Snowdon DA, Kemper SJ, Mortimer JA, Greiner LH, Wekstein DR, Markesbery WR. Linguistic ability in early life and cognitive function and Alzheimer’s disease in late life: findings from the nun study. JAMA. 1996;275(7):528–32.
Spreng RN, Drzezga A, Diehl-Schmid J, Kurz A, Levine B, Perneczky R. Relationship between occupation attributes and brain metabolism in frontotemporal dementia. Neuropsychologia. 2011;49(13):3699–703.
Steffener J, Stern Y. Exploring the neural basis of cognitive reserve in aging. Biochim Biophys Acta (BBA)-Mol Basis Dis. 2012;1822(3):467–73.
Stern Y. What is cognitive reserve? theory and research application of the reserve concept. J Int Neuropsychol Soc. 2002;8(03):448–60.
Stern Y. Cognitive reserve. Neuropsychologia. 2009;47(10):2015–28.
Stern Y, Gurland B, Tatemichi TK, Tang MX, Wilder D, Mayeux R. Influence of education and occupation on the incidence of Alzheimer’s disease. JAMA. 1994;271(13):1004–10.
Stern Y, Alexander GE, Prohovnik I, Stricks L, Link B, Lennon MC, et al. Relationship between lifetime occupation and parietal flow: implications for a reserve against Alzheimer’s disease pathology. Neurology. 1995;45(1):55–60.
Tolppanen A-M, Solomon A, Kulmala J, Kåreholt I, Ngandu T, Rusanen M, et al. Leisure-time physical activity from mid- to late life, body mass index, and risk of dementia. Alzheimer’s Dementia. 2015;11(4):434–43.
Tuokko H, Garrett DD, McDowell I, Silverberg N, Kristjansson B. Cognitive decline in high-functioning older adults: reserve or ascertainment bias? Aging Ment Health. 2003;7(4):259–70.
Verghese J, Lipton RB, Katz MJ, Hall CB, Derby CA, Kuslansky G, et al. Leisure activities and the risk of dementia in the elderly. N Engl J Med. 2003;348(25):2508–16.
Wang H-X, Xu W, Pei J-J. Leisure activities, cognition and dementia. Biochim Biophys Acta (BBA)-Mol Basis Dis. 2012a;1822(3):482–91.
Wang H-X, Wahlberg M, Karp A, Winblad B, Fratiglioni L. Psychosocial stress at work is associated with increased dementia risk in late life. Alzheimer’s Dement. 2012b;8(2):114–20.
Wang H-X, Jin Y, Hendrie HC, Liang C, Yang L, Cheng Y, et al. Late life leisure activities and risk of cognitive decline. J Gerontol Ser A Biol Sci Med Sci. 2013;68(2):205–13.
Whalley LJ, Starr JM, Athawes R, Hunter D, Pattie A, Deary IJ. Childhood mental ability and dementia. Neurology. 2000;55(10):1455–9.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Dekhtyar, S., Wang, HX. (2017). Cognitive Reserve: A Life-Course Perspective. In: Petrosini, L. (eds) Neurobiological and Psychological Aspects of Brain Recovery. Contemporary Clinical Neuroscience. Springer, Cham. https://doi.org/10.1007/978-3-319-52067-4_5
Download citation
DOI: https://doi.org/10.1007/978-3-319-52067-4_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-52065-0
Online ISBN: 978-3-319-52067-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)