
Abstract
Lake Cadagno (26 ha) is a crenogenic meromictic lake located in the Swiss Alps at 1921 m asl with a maximum depth of 21 m. The presence of crystalline rocks and a dolomite vein rich in gypsum in the catchment area makes the lake a typical “sulphuretum ” dominated by coupled carbon and sulphur cycles. The chemocline lies at about 12 m depth, stabilized by density differences of salt-rich water supplied by sub-aquatic springs to the monimolimnion and of electrolyte-poor surface water feeding the mixolimnion. Steep sulphide and light gradients in the chemocline support the growth of a large bacterial plume (up to 107 cells ml−1) dominated by green sulphur bacteria (GSB) of the genus Chlorobium and purple sulphur bacteria (PSB) of the Chromatiaceae family. Since the early Holocene (10.5–8 cal kyr BP), PSB and GSB are showing long-term alternation in abundance and relative dominance. Key species are Chlorobium clathratiforme , Thiocystis chemoclinalis , Thiocystis cadagnonensis , Candidatus “ Thiodictyon syntrophicum ” and Chromatium okenii ; the latter represents only 0.3 % of the total cell number but due to its big size and high activity can contribute up to 70 % of the total carbon uptake in the chemocline. Small-celled PSB together with the sulfate-reducing bacterium Desulfocapsa thiozymogenes sp. form stable aggregates in the lake, which represent small microenvironments with an internal sulphur cycle. Eukaryotic primary producers in the anoxic zones are dominated by Cryptomonas phaseolus , whereas eukaryotic heterotrophs are represented by ciliates and choanoflagellates, but a clade of heteroloboseans and two novel clades distantly related to opisthokonts and Cercozoa are also present. Zooplankton and fish abundance in the mixolimnion of this model ecosystem are linked via food web to the chemocline microbial plume, consequently anaerobic primary production supports relative high fish productivity in the lake.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Due to their permanent stratification, meromictic lakes are interesting model ecosystems. In particular, the chemocline offers many advantages for studying the planktonic life and processes mediated by microorganisms. The physico-chemical gradients across the chemocline support the development of very intense microbial blooms (Overmann et al. 1996; Parkin and Brock 1981; Sorokin 1970; van Gemerden and Mas 1995). Along vertical gradients of light and different electron donors and acceptors, diverse physiological groups of organisms substitute each other (Guerrero et al. 1985; Jørgensen et al. 1979; Overmann et al. 1991) in distinct layers at depth intervals ranging from cm to m. These distinct layers facilitate accurate sampling and measurement of environmental characteristics. Lake Cadagno is a crenogenic meromictic lake located in the southern Swiss Alps (46°33′N, 8°43′E) at 1921 m asl, in the catchment area of a dolomite vein rich in gypsum (Piora-syncline). A permanent chemocline lies between 10 and 14 m depth, stabilized by density differences of salt-rich, high-density water constantly supplied by sub-aquatic springs to the monimolimnion and of electrolyte-poor, low-density surface water that feeds the mixolimnion. Steep sulphide gradients in the chemocline support the growth of large numbers of phototrophic sulphur bacteria (up to 107 cells ml−1). This community includes purple sulphur bacteria (PSB, family Chromatiaceae ) of the genera Chromatium, Lamprocystis, Thiocystis and Thiodictyon and green sulphur bacteria (GSB. family Chlorobiaceae) of the genus Chlorobium . Since the beginning of the last century, Lake Cadagno and the nearby Lake Ritom have been used as models to study water stratification, sulphidogenic bottom waters and blooms of PSB. Since the early 1980s, studies on ecological issues in Lake Cadagno significantly increased, supported by two major developments. The first was the establishment of research facilities at the shore of the lake, the Alpine Biology Center, and the second was the introduction of novel molecular techniques that allowed researchers to analyse microbial populations in Lake Cadagno unaffected by the difficulties in culturing them.
This contribution presents an overview of more than 30 years of studies on Lake Cadagno , including biogeochemical analyses, diversity surveys and ecological studies on the very diverse microbial community developing in it.
The present chapter is structured in the following major parts:
-
1.
Hydrology, and morphology of the lake in brief, including the lake’s origin and age
-
2.
Historical perspective and description of early investigations conducted on Lake Cadagno and description of the lake’s meromixis and the main physico-chemical features of the lake and its related stratification
-
3.
The organisms inhabiting the lake, with its diversity dominated by microorganisms, particularly in relation to the sulphur cycle
The vast majority of the studies on Lake Cadagno concern the bacterial community developing in the chemocline . The chapter on the biology of the lake reviews comprehensively studies published since 1985 and results of recent studies on prokaryotic diversity (see Sect. 5.1) and on microbial eukaryotic diversity (see Sect. 5.3).
2 Origin, Morphology and Hydrology of Lake Cadagno
Lake Cadagno (Fig. 7.1) originated due to glacial erosion during the last glacial period, about 10,000 years ago, and the formation of a moraine acting as a small dam (Boucherle and Züllig 1988; Krige 1918; Stapfer 1991; Wirth et al. 2013). The lake is located in the southern Swiss Alps at 1921 m asl (latitude, 46°33′N; longitude, 08°43′E), in Val Piora, a region characterized by its richness in mountain lakes, waterways, wetlands and ponds (Garwood 1906; Peduzzi 2011). Lake Cadagno is small (maximum length, 830 m; maximum width, 420 m; surface area, 0.26 km2; maximum depth, 21 m; average depth, 9.27 m) (Del Don et al. 2001), situated among pre-Triassic gneiss and mica-schists of central alpine crystalline rocks just south of the Gotthard Pass in the Canton Ticino (Switzerland). It has a catchment area of 2.71 km2. The southern lake shore rests on late Triassic and early Jurassic rocks folded into the so-called Piora-syncline, characterized by the presence of tectonized dolomitic limestones (magnesian limestone) and gypsum deposits with a karstic hydrology. The Piora-syncline forms a vein that separates the crystalline rocks of the Gotthard massif to the north and the metamorphic Pennine Lucomagno Nappe, about 10 km to the south, along the whole Piora valley (Otz et al. 2003).

Lake Cadagno and Piora valley. Photo taken from the west side of the lake
Lake’s sediment analysis reveals that permanent stratification, i.e. meromixis, and the presence of phototrophic sulphur bacteria must have been established very early in the history of the lake. This suggestion was first based on the analyses of organic components in the upper sediments (Putschew et al. 1995; Hebting et al. 2006) and of Cladocera (Crustacea) remains (Boucherle and Züllig 1988). Trace pigments, e.g. okenone and isorenieratene , characteristic for purple and green phototrophic bacteria, were detected in the upper 170 cm of the lake’s sediment (Züllig 1985). However, the detailed longer-term history of development of meromixis and differential redox conditions in Lake Cadagno , and the timing of the transition from oxic to euxinic (i.e. sulphidic and anoxic) conditions in the water column, remained unknown until 2009, when 10.5-m-long sediment cores covering the complete lacustrine record of Lake Cadagno were investigated using an interdisciplinary approach (Niemann et al. 2012; Ravasi et al. 2012b; Dahl et al. 2013; Wirth et al. 2013). The lake history was characterized by a first oxic-intermediate-sulphidic redox-transition period shortly after glacial retreat, and lake formation was estimated at 12.5 kyr BP (before present). During the 2300-year-long intermediate period from 12.1 to 9.8 kyr BP, strongly fluctuating Mn concentrations in the sediments associated with episodic water column mixing events were recorded (Wirth et al. 2013). At the same time, efficient Mo burial (Dahl et al. 2010) and the presence of remains of phototrophic sulphur bacteria (Ravasi et al. 2012b) provided an evidence for a sulphidic environment. In fact, at the end of this period with intermediate redox conditions, ca. 9800 ± 130 years ago, a pronounced drop in Mn and a simultaneous rise in Mo concentrations in the sediments indicated efficient Mo burial in sulphide-rich water column and thus the onset of euxinic conditions in Lake Cadagno. This redox transition was causally linked to the concurrent climate warming, which led to permafrost melting and the activation of the sub-aquatic springs. The inflow of salt-rich waters to the lake bottom likely initiated the formation of a chemocline and a stable crenogenic meromixis (créno, spring; from the Greek “Krini”) in the early Holocene. Recent biogeochemical data provide evidence that Lake Cadagno has remained sulphidic from the start of Holocene to date. Flood and mass movement events, however, generated underflows of turbidity-rich water that sporadically introduced oxygen-rich water to the hypolimnion, thereby only temporarily weakening the sulphidic conditions (Wirth et al. 2013). Such events may have influenced water turbidity and nutrient supply and may thus have caused shifts in the microbial community structure (see Sect. 5.1.1.2.) (Tonolla et al. 2005b; Decristophoris et al. 2009; Gregersen et al. 2009).
Water penetrating and circulating through the karstic dolomite and reappearing as underwater springs in the southern part of the lake has a rather high ionic strength (9–10 mM), and it flows downwards feeding the anoxic lower part of the water column (monimolimnion). Dye tracing experiments (Otz et al. 2003) have revealed that this groundwater originates from a relative small underground catchment area which extends for about 2 km to the southeast up to the so-called Calderoni Sinkhole. The sinkhole causes, uphill, a flow of the underground water towards the Lucomagno, i.e. in the opposite direction to the surface water flow. On the other hand, the upper part of the lake’s aerobic water column (mixolimnion) is fed by surface run-off from a small drainage area of about 2 km2, north of the lake, which encompasses the crystalline rocks of the Gotthard massive. The major source of surface input to Lake Cadagno is a small stream that drains water from Lake Stabbio, a small lake at 2351 m asl. The crystalline rocks of the watershed are rather resistant to chemical weathering, such that the water of the mixolimnion has a relatively low ionic strength and is oligo- to mesotrophic. The density difference between the two water masses in Lake Cadagno causes the development of a very stable stratification with a chemocline at about 10–14 m depth characterized by the rapid change of the concentrations of chemical components (Del Don et al. 2001; Hanselmann and Hutter 1998; Peduzzi et al. 1998).
3 Historical Overview: Scientific Studies and Hydroelectric Exploitation
Lake Cadagno has attracted scientists since the end of the nineteenth century. In fact, since the foundation of limnology in 1904, the lakes of the Piora region have created considerable scientific interest (Bourcart 1906; Garwood 1906; Burckhardt 1910); for a review on the historic data sets on lakes Ritom and Cadagno, see Peduzzi and Bianconi (2012). Stratification phenomena, as well as sulphur-rich deep waters and mass developments of phototrophic sulphur bacteria , are among the early observations on the lake. A monograph on the nearby Lake Ritom (Collet et al. 1918) reported results of studies carried out during the construction of the Ritom hydroelectric dam (1915–1920). Already then, the importance of the sulphur cycle and the bacteria involved therein were mentioned (Düggeli 1924). Studies on fish populations (Surbeck 1917) and earlier researches on phytoplankton and sulphur bacteria were carried out on Lake Ritom and Lake Cadagno (Bachmann 1924, 1928; Borner 1928a, b; Burckhardt 1910; Düggeli 1924). These early published works are important documents reporting the loss of meromixis in Lake Ritom following the construction of the Ritom dam. Similar results were reported for a French alpine lake, Lac de la Girotte in Savoie, in 1926 (Delebeque 1898; Barades 1926). Lake Cadagno is probably the most unique among the lakes in the entire alpine range that exhibit crenogenic meromixis. In 1948, a small dam was built raising the lake’s surface water level from 1918 to 1921 m asl, and up to the year 2007, the lake was exploited for production of electricity resulting in a 3-m decrease in the lake’s water level during the winter. During almost 60 years of exploitation of the lake for hydroelectric generation, it retained its permanent stratification. The lowering of the water level led to a loss of about 720,000 m3 water, i.e. about 30 % of the lake’s total water volume (2.42 million m3). This comprised 50 % of the water volume with O2 concentration >4 mg l−1 (around the minimum survival limit for Salmonidae). Aerobic organisms in the mixolimnion were subjected to a significant reduction in vital living space. In addition to that, 45,000 m2 of littoral dried up causing, on the one hand, the death of numerous populations of sediment dwellers (e.g. Ephemeroptera, Plecoptera and Trichoptera), which were a potential source of food for fish species and, on the other hand, the loss of spawning areas for the fish reproduction (Tonolla et al. 1988). At present, the lake is no more actively being exploited for the production of electricity, and therefore its water level remains fairly constant at 1921 m asl.
Studies on Lake Cadagno intensified during the twentieth century, more so since the 1980s. The opening of the Alpine Biology Center (ABC) in Piora in 1994 in cooperation with the Universities of Geneva, Zurich, Lugano and recently the newly opened University of Applied Sciences and Arts of Southern Switzerland (SUPSI), as well as participating cantons, the municipality and the patriciate of Quinto, have led to a substantial increase in scientific studies in the region.
The presence of scientific database is an invaluable source of information for researchers, as it provides an important historical perspective and solid data sets as a basis for future research. For a complete bibliography on scientific studies on the region and on Lake Cadagno before 1998, refer to Chronological list of publications on Lake Cadagno and Piora Valley in Peduzzi et al. (1998), and for the period after 1998, check on the website of the Alpine Biology Center, Piora ( http://www.piora.org/ ).
4 Lake Water Chemistry and Meromixis
The dolomite vein present in the Piora valley is in direct contact with the southern part of the Cadagno basin. The water which percolates through these easily weathered rocks becomes enriched in calcium, magnesium, carbonate and sulphate ions and enters the waterbody through sublacustrine springs. The analyses of seven underwater springs located in the southern part of the lake at depths between 8.0 and 12.5 m provided the following averages for: conductivity, 1359 (±145) μS cm−1; calcium, 171.1 (±22.5) mg l−1; magnesium, 74.7 (±10.1) mg l−1; sulphate, 622.9 (±81.5) mg l−1; and carbonate, 120.7 (±35.6) mg l−1 (Del Don et al. 2001). The inflow of this water contributes to the establishment of the crenogenic meromixis in the lake. The chemocline is permanent and stabilized by the difference in density between the two water strata: the upper mixolimnion and the lower monimolimnion (Del Don et al. 2001; Lehmann and Bachofen 1999; Tonolla et al. 1998b).
The concentrations of nutrients in the oxic mixolimnion (between 0 and 10–12 m depth) are low, with phosphate (PO4−P) close to the detection limit (<0.3 μg l−1), nitrate (NO3−N) below 11.5 μg l−1 and dissolved inorganic carbon (DIC) ca. 10 mg l−1 (Tonolla et al. 1998b, 1999). The monimolimnion is anoxic and rich in reduced compounds due to the action of anaerobic sulphur bacteria. Moreover, it is a steady food source indispensable for the development of the dense community of phototrophic sulphur bacteria in the chemocline . High concentrations of sulphur compounds (HS− up to 30 mg l−1; \( {\mathrm{SO}}_4^{2-} \) up to 200 mg l−1) ammonium (NH4−N up to 3.9 mg l−1), phosphate (PO4−P up to 0.132 mg l−1), carbonates (\( {\mathrm{HCO}}_3^{-} \) and \( {\mathrm{CO}}_3^{2-} \) up to 50 mg l−1) (Del Don 2001; Tonolla et al. 1998b) and dissolved organic carbon (DOC, up to 6 mg C l−1) (Bertoni et al. 1998) are the basis for the massive development of populations of these phototrophic sulphur bacteria, which reach maximum densities of 107 cells per ml−1 during summers.
With sulphate concentrations ranging between 100 and 200 mg l−1 (1–2 mM), Lake Cadagno contains up to ten times more sulphate than most freshwater lakes (Hanselmann and Hutter 1998). In addition, sulphate, bicarbonate, calcium and manganese are the dominant ions in Lake Cadagno (Tonolla et al. 1998a).
For the water stability, the massive development of large-celled, flagellated phototrophic purple sulphur bacteria in the chemocline may also influence the profiles of temperature and salinity by their active movements, contributing to maintain constant values over a depth interval of about 0.5 m. The water mixing caused by the organisms at this level destabilizes the density gradient and the physico-chemical stratification (Wüest 1994).
5 Biology of Lake Cadagno
5.1 Prokaryotic Diversity
The prokaryotic diversity of Lake Cadagno has been studied by several workers in the last three decades (Züllig 1985; Peduzzi et al. 1998, 2000, 2012; Tonolla et al. 1999, 2000, 2005a). Because of the difficulty in culturing environmental microorganisms, the study of the bacterial populations was focused, as early as in the 1990s, on direct detection methods such as nucleic acid staining with DAPI or acridine orange (Bensandoun et al. 1998; Tonolla et al. 1998a). Such approaches revealed the presence of several bacterial morphotypes (e.g. Chromatium okenii , Lamprocystis sp., Desulfocapsa and morphotype R) and allowed an evaluation of the bacterial diversity in the lake’s water column. The phylogenetic analysis of bacterial communities continued with the amplification and generation of gene clone libraries for 16S ribosomal RNA genes (Bosshard et al. 2000a; Demarta et al. 1998; Schramm et al. 2003). The microbiological analyses did not only focus on the chemocline (Tonolla et al. 1999, 2000, 2005a; Bosshard et al. 2000a, b; Decristophoris et al. 2009; Halm et al. 2009), but also on the monimolimnion, the mixolimnion (Tonolla et al. 2005b; Gregersen et al. 2009; Milucka et al. 2015) and the anoxic sediment (Schubert et al. 2011). They allowed the identification of novel bacterial species, later isolated from the chemocline of the lake (e.g. Thiocystis chemoclinalis sp. nov., Thiocystis cadagnonensis sp. nov. and Candidatus “Thiodictyon synthrophicum” sp. nov.) (Peduzzi et al. 2011; Peduzzi et al. 2012).
Recently a next-generation sequencing technology, 454 pyrosequencing, was also applied to explore the prokaryotic diversity in the layers of the water column (mixolimnion, chemocline , monimolimnion) and the superficial anoxic sediment of Lake Cadagno (unpublished data). This generated 22,700 sequences of the hypervariable V1–V3 regions (average length, 400 bp) of the 16S rRNA gene. Through PICRUSt algorithm (Langille et al. 2013), 3244 sequences (1227 sequences for mixolimnion, 621 for chemocline, 1326 for monimolimnion and 70 for anoxic sediment) could be assigned to 18 known bacterial phyla, 11 of which were dominant (>1 % relative abundance). This number is higher than the 12 phyla identified in the previous studies (Ravasi et al. 2012a). The monimolimnion noticeably differed, with 13 phyla identified by 454 pyrosequencing, compared with eight phyla previously identified. Also, microbial communities differed among the different layers in the water column and the anoxic sediment. The mixolimnion was dominated by the phylum Verrucomicrobia (51 %), which was less abundant in the chemocline (8 %) and monimolimnion (4 %) and absent in the anoxic sediment. Other important phyla of mixolimnion were Actinobacteria (14%) and Bacteroidetes (3 %). Proteobacteria were the most represented phylum in the chemocline communities (76 %); they were also well represented in the mixolimnion (31 %), monimolimnion (19 %) and anoxic sediment (18 %). Other important communities in the chemocline were Chlorobi (8 %), Actinobacteria (5 %) and Bacteroidetes (2 %). Other than the presence of Proteobacteria, the monimolimnion communities were mainly comprised of Chlorobi (66 %), Actinobacteria (6 %), Bacteroidetes (3 %) and Chloroflexi (1 %). The anoxic sediment was the most diverse habitat, with various phyla almost equally represented, in addition to the previously mentioned Proteobacteria (Chloroflexi 19 %, Chlorobi 15 %, Actinobacteria 14 %, Cyanobacteria 11 %, OP8 8 %, Bacteroidetes 7 %, Firmicutes 2 % and Nitrospirae and Acidobacteria 1 %). At a finer taxonomical resolution, Alphaproteobacteria were the dominant class of Proteobacteria in the mixolimnion, with the family of Pelagibacteraceae (aerobic oxygenic bacteria). Alphaproteobacteria also represented a good part of the Proteobacteria found in the anoxic sediment ( Hyphomicrobiaceae , Pelagibacteraceae and Rhodobacteraceae ). The class Betaproteobacteria was represented in all samples, mainly by Comamonadaceae . Deltaproteobacteria were also present in all habitats analysed, except in the mixolimnion; they were best notable in the monimolimnion, with the sulphate reducers Desulfobulbaceae, and in the sediment (clade OM27). Epsilonproteobacteria were present in chemocline and monimolimnion ( Helicobacteraceae ). Gammaproteobacteria were the most dominant Proteobacteria class in the chemocline, with the sulphur purple photosynthetic Chromatiaceae , and were also prominent in the monimolimnion (Chromatiaceae) and the sediment (Moraxellaceae, Pseudoalteromonadaceae, Sinobacteraceae, Vibrionaceae).
The diversity of archaea in Lake Cadagno has received less attention than the bacteria. Nonetheless, the few studies that were conducted have highlighted interesting patterns of distribution. Bottinelli (2008) explored the coexistence of sulphate-reducing bacteria and methanogenic archaea in the sediments of the lake. The analysis of 16S rRNA gene sequences allowed the detection of few phylotypes, with more diversity in the deeper part of the sediments. Schubert et al. (2011) studied the archaeal community involved in methane production or degradation in upper sediments of Lake Cadagno, constructing 16S rRNA gene clone libraries from 2 to 4 cm sediment (42 clones) and 9- to 15-cm (43 clones) sediment depth. These depths represented the peak (2–4-cm sediment) and bottom (9–15-cm sediment) of the AOM (anaerobic oxidation of methane) zone. The archaeal diversity revealed in this study was also low, with only nine different phylotypes detected, based on a 97 % 16S rRNA gene sequence similarity criterion. The majority of 16S rRNA gene sequences belonged to the euryarchaeotal marine benthic group D (MBGD), found in a variety of limnic and marine habitats. In total, six distinct MBGD phylotypes were retrieved, three of which were only found in 2–4-cm sediment depth. The second archaeal lineage detected was the recently described AOM-associated archaea (AAA) clade of Euryarchaeota . As third and fourth archaeal lineages, the crenarchaeotal marine benthic groups B and C (MBGB and MBGC) were identified. Catalysed reporter deposition-fluorescence in situ hybridization (CARD–FISH) analysis revealed a high abundance of Deltaproteobacteria , especially of free-living sulphate-reducing bacteria of the Desulfosarcina/Desulfococcus branch of Deltaproteobacteria in the AOM zone. Here, loose aggregations of AAA cells were found that might be responsible for oxidation of methane in the lake sediments (Schubert et al. 2011). A more recent study by Milucka et al. (2015) on methane oxidation coupled to oxygenic photosynthesis in the chemocline of Lake Cadagno revealed that, despite the geochemical evidence for high methane-oxidizing activity, no anaerobic methane-oxidizing archaea (ANME)-1 or (ANME)-2 were detected in samples from the anoxic zone (i.e. from 10 and 13 m) by CARD–FISH with any of the used probes.
The Anoxygenic Phototrophic Sulphur Bacteria in the Chemocline
One of the main features of sulphur-rich Lake Cadagno is the diverse community of microorganisms which develops in the chemocline especially in the spring and summer months. The community, which is dominated by anoxygenic phototrophic sulphur bacteria , also contains microorganisms that acquire energy via anaerobic respiration, aerobic respiration and fermentation (Bosshard et al. 2000a). Both green sulphur bacteria (GSB) and purple sulphur bacteria (PSB) have been observed in Lake Cadagno, and a number of their strains have been successfully isolated, cultivated in the laboratory and described as new bacterial species (i.e. Eichler and Pfennig 1988; Tonolla et al. 1999; Peduzzi et al. 2011, 2012). Thiocystis chemoclinalis , T. cadagnonensis and Candidatus “ Thiodictyon syntrophicum ” were recently described (Peduzzi et al. 2011, 2012).
Anoxygenic phototrophic sulphur bacteria are phylogenetically diverse, but all harvest light energy using antenna complexes containing bacteriochlorophylls, and accessory pigments, such as carotenoids. The antenna complexes adsorb light radiation and transfer the captured energy to a reaction centre where it is subsequently used to generate ATP and reductant (NAD(P)H). Anoxygenic phototrophic sulphur bacteria utilize either the pheophytin–quinone-type reaction centres (PSB and Chloroflexus sp.) or the Fe–S reaction centres (GSB and Heliobacteria sp.). In addition to sulphide, some of these bacteria utilize elemental sulphur and thiosulphate as electron donors (Frigaard and Dahl 2009).They can also utilize hydrogen (H2(g)) as a reductant. Many anoxygenic phototrophs can also oxidize ferrous iron to reduce CO2 (Widdel et al. 1993; Ehrenreich and Widdel 1994; Overmann and Garcia-Pichel 2006; Hegler et al. 2008). The oxidation of reduced inorganic sulphur by sulphide-oxidizing anoxygenic phototrophic bacteria is usually coupled to CO2 reduction to form cell biomass following a number of possible pathways (Eqs. 7.1–7.5):
Taxonomy of Anoxygenic Phototrophs from Lake Cadagno
In Lake Cadagno , the cell concentration maxima of anoxygenic phototrophs ( Chromatiaceae and Chlorobiaceae) in the summer are located between 10.5 and 13.5 m depth in the chemocline (Schanz et al. 1998; Bosshard et al. 2000a). The community of phototrophic bacteria in the chemocline is diverse but dominated by a few species of purple and green sulphur bacteria (Tonolla et al. 1999; Bosshard et al. 2000a). The main anoxygenic phototrophic bacteria identified in Lake Cadagno (Table 7.1) are three flagellated PSB, one large-celled, Chromatium okenii , and two small-celled, Thiocystis chemoclinalis and T. cadagnonenensis; four small-celled PSB Lamprocystis purpurea , Candidatus “ Thiodictyon syntrophicum ”, Lamprocystis roseopersicina and Lamprocystis sp. strain D; and two GSB Chlorobium clathratiforme and Chlorobium phaeobacteroides (Bosshard et al. 2000a, b; Tonolla et al. 1999, 2003, 2005b; Peduzzi et al. 2011, 2012). The main sulphide-oxidizing anoxygenic phototrophs are grouped according to their physiology, morphology, pigment composition and membrane structure (Table 7.1).
The PSB belong to the gamma division of the Proteobacteria . They have BChl a as the main bacteriochlorophyll and contain carotenoids, such as okenone . The reaction centre is of Type II containing pheophytin and quinone. They fix CO2 using the reductive pentose phosphate cycle. Metabolically, they can be photoautotrophs or photoheterotrophs and in some cases also chemolithotrophs growing in the absence of light (Table 7.1). Two examples of strains that can grow chemolithotrophically are Lamprocystis purpurea and Candidatus “ Thiodictyon syntrophicum ”. PSB are mostly obligate anaerobes, although some may tolerate low-oxygen conditions. The four main small-celled PSB found in Lake Cadagno are between 1.4 and 4 μm in diameter and spherical to oval in shape. The best described of the group, L. purpurea and Candidatus “T. syntrophicum”, utilize BChl a and the carotenoid okenone to harvest light for photosynthesis. The small-celled PSB in Lake Cadagno are immotile and contain gas vacuoles and intracellular sulphur globules. They also form aggregates , notably together with sulphate-reducing bacteria (SRB) belonging to the Desulfovibrionaceae (Tonolla et al. 2003; Peduzzi et al. 2003b; review by Tonolla et al. 2004). This association with the SRB is considered to provide them with a environmental advantage by providing substrate (Descristophoris et al. 2009). The 16S rRNA gene sequence of Lamprocystis roseopersicina is very similar to that of Lamprocystis purpurea, a finding that supports the reclassification of the latter species from Amoebobacter to Lamprocystis (Imhoff 2001). The large-celled PSB in the lake, Chromatium okenii , also belongs to the gamma division of the Proteobacteria (Table 7.1). This bacterium is rod shaped and has large cells (4.5–6 × 8–15 μm). C. okenii are motile using flagella and they harbour intracellular sulphur globules (Fig 7.2).

Light microscopy images of (a) green sulphur bacterium Chlorobium clathratiforme and (b) purple sulphur bacterium Chromatium okenii showing intercellular sulphur globules
The GSB belong to a separate phylum ( Chlorobi ). They have a Type I reaction centre (Fe–S) and are generally considered strict photoautotrophs fixing CO2 using the reverse citric acid cycle. The occurrence of the concentrated pigments in chlorosomes allows the GSB to use light at very low intensities (Frigaard and Bryant 2006). The abundance of the brown-coloured GSB, Chlorobium clathratiforme, increased in 2000 in Lake Cadagno and thereafter formed a significant population in the chemocline (Tonolla et al. 2005b; Gregersen et al. 2009; Habicht et al. 2011). Chlorobium clathratiforme has irregular, rod-shaped cells with typical dimensions of ~1 by 4–7 μm (Fig 7.2). They contain BChl e and the carotenoid isorenieratene . The cells are net-forming and nonmotile and display cultivation-dependent gas vacuoles.
Temporal and Spatial Distribution of the Anoxygenic Phototrophic Sulphur Bacteria
The population density of anoxygenic phototrophic sulphur bacteria in the chemocline varies with depth in the anaerobic water column (upper monimolimnion), seasonally and over longer timescales. As expected from the geochemical characteristics of the lake, there is a strong change in community composition between the oxic and anoxic layers (Bosshard et al. 2000b). The seasonal distribution of the anoxygenic phototroph population in the chemocline suggests that the population is adapted to changing geochemical niches (Tonolla et al. 1999). In March and June 1999, bacterial populations were evenly distributed in depth, but a microstratification of the population was observed as the season progressed and the geochemical gradient developed (Tonolla et al. 2003). A shift in dominance among the Chromatiaceae from Chromatium okenii in the spring and early summer to an increase in Lamprocystis purpurea in the late summer and autumn was observed by Bosshard et al. (2000a, b). However, this pattern has not been consistent in recent decades. In another seasonal study, Tonolla et al. (2003) observed high densities of C. okenii in the late autumn, but small-celled forms of the PSB were most abundant during spring and summer. Thus, even among the PSB populations, a number of ecophysiological factors contribute to the population composition. These results are further confounded by the motility of C. okenii, likely a response to light and nutrient availability.
The changes in the anoxygenic phototroph community structure in Lake Cadagno during the last decades are intriguing. Also, the relative abundance of PSB and GSB has changed dramatically over the past 15 years. A study on in situ hybridization during 1994–2003 shows that up to 2001, PSB were the most prominent phototrophic sulphur bacteria forming 70–95 % of the total bacterial population (review by Tonolla et al. 2004). In 2000, the dominance shifted from PSB to GSB, due to a dramatic increase of Chlorobium clathratiforme (Tonolla et al. 2005b). The bacterial biomass increased threefold, due entirely to the increase in abundance of GSB, while PSB densities remained stable possibly because of associations with SRB in the aggregates (Decristophoris et al. 2009). After this change, up to 95 % of the phototrophic sulphur bacteria community was comprised of Chl. clathratiforme (Tonolla et al. 2005b). Indeed, subsequent studies revealed that Chl. clathratiforme in Lake Cadagno is a clonal population forming up to 70 % of the bacterial 16S rRNA gene sequences obtained in the chemocline at 11–12-m depth, where the cell concentration was the highest (Gregersen et al. 2009). The population shift since 2000 could be due to a change in environmental conditions, i.e. the introduction of a new strain from outside by natural or anthropogenic means, or because of a favourable mutation in the existing population, e.g. one that allows a more effective metabolism in the dark. A study of Chl. clathratiforme in the water column of Lake Cadagno suggests that it may carry out the fermentation of polyglucose in the dark, effectively giving this bacterium a competitive advantage (Habicht et al. 2011).
As expected, the PSB are located higher in the anoxic water column than are GSB. Due to the differences in the light-harvesting mechanism in PSB and GSB, it seems logical that these groups of anoxygenic phototrophic bacteria would easily coexist, each inhabiting its niche determined by light intensity and possibly wavelength of light. The concentrations of chlorophyll in oxygenic phototrophs in the oxic upper layers of Lake Cadagno are low. At water depths corresponding to the depth of chemocline , the wavelength distribution of the available light is largely determined by the light absorbance by water itself. The wavelengths reaching a depth of 10 m mainly correspond to that absorbed by carotenoids and the Qx band of BChl e. The intensity of near-infrared radiation at wavelengths above 700 nm is much less so the characteristic long-wavelength absorbance of aggregated BChl e in the chlorosomes of Chl. clathratiforme or protein-associated BChl a in the PSB is unlikely to be important in the lake. Nonetheless, long-term studies of the phototrophic population reveal that the changes in light-related limnological properties of the lake, such as turbidity, light availability and light intensity over long timescales (decades), profoundly influence the population composition (Tonolla et al. 2005b).
Role of the Anoxygenic Phototrophic Sulphur Bacteria in the Primary Production of Lake Cadagno
Although the chemocline represents only a small part of the total lake volume, here about half of the total daily photoassimilation of carbon in the lake occurs (Camacho et al. 2001). Photosynthetic 14C-assimilation, measured with radioisotope (14C) technique, generally peaks at 1–3 m (phytoplankton) and at 10–12 m, the location of the chemocline with the anoxygenic phototrophic bacteria community. Very high values of phototrophic and chemotrophic C-assimilation are generally observed below 10 m (Bossard et al. 2001). Indeed, in a study on Lake Cadagno in September 1999, Camacho and coworkers found high rates of dark carbon fixation by chemolithotrophs in the chemocline, suggesting a high contribution of these organisms to the overall carbon cycle of the lake (Camacho et al. 2001). Recently, the rates of CO2 assimilation of the most abundant phototrophic sulphur bacteria of the chemocline were measured using both nanoscale secondary ion mass spectrometry (nanoSIMS ) (Musat et al. 2008; Zimmermann et al. 2015) and 14CO2 quantitative assimilation in dialysis bags (Storelli et al. 2013). Thus, the strongest assimilator in the presence of light was the large-celled PSB C. okenii, while in the dark the major assimilator was the small-celled PSB Candidatus “T. syntrophicum”, which was also a strong CO2 assimilator in the light. While C. okenii represents only ca. 0.3 % of the total cell number, it contributes 70 % of the total uptake of carbon in the system (Musat et al. 2008), an assimilation effectiveness theorized by the authors to be based in C. okenii’s mobility towards substrate and optimal light conditions. Likewise, although it only contributes just about 2 % of the total bacterial populations in the chemocline, Candidatus “T. syntrophicum” appears to also play a greater role in CO2 fixation in Lake Cadagno, irrespective of if there is light (Storelli et al. 2013). The most abundant population of the chemocline, GSB Chl. clathratiforme, showed only low CO2 fixation rates. These results were supported by a micro-autoradiography analysis which revealed that about 10 % of the cells of Chl. clathratiforme from below the photic zone in Lake Cadagno fixed CO2 in the dark, and 20–55 % of the cells from the photic zone fixed CO2 in the light (Habicht et al. 2011). Although GSB are considered strictly phototrophic, their low rates of CO2 photoassimilation are surprising. Another metabolic pathway probably contributes to biomass production. At present, only acetate and pyruvate are known to be used by GSB as organic carbon source in the presence of sulphide and CO2 (Van Gemerden and Mas 1995).
Non-photosynthetic carbon assimilation is often observed in phototrophic sulphur bacteria but the pathway is not yet fully understood. In order to gain insight into the process of dark CO2 fixation, two-dimensional differential gel electrophoresis (2D-DIGE) was used to monitor the global changes in the proteome of Candidatus “T. syntrophicum” strain Cad16T in anoxic autotrophic cultures, irrespective of availability of light. Interestingly, three enzymes more abundant in the dark are part of a hypothetical anaerobic dicarboxylate/4-hydroxybutyrate (DC/HB) cycle, an autotrophic CO2 fixation pathway found essentially in archaea (Berg 2011). The substrates needed for this process [NAD(P)H and acetyl-CoA] should be provided by the degradation of the storage globules of poly(3-hydroxybutyrate) (PHB), whose synthesis was shown to be higher in the presence of light (Storelli et al. 2014).
The Sulphate-Reducing Bacteria (SRB)
Within the chemocline , non-phototrophic bacteria are also present: these include SRB that can account for a significant fraction of the total community (Tonolla et al. 1998a). Their seasonal depth distribution was consistent with the variations of the phototrophic PSB. Over an annual cycle, however, differences in abundance of bacteria detected with probes SRB385 (detecting most Desulfovibrionaceae ) and SRB385Db (detecting most Desulfobacteriaceae) were observed (Peduzzi et al. 2003a). In situ determination of sulphide turnover rates (Luthy et al. 2000) and high-resolution images of sulphide concentrations in the water column (Lehmann and Bachofen 1999) confirmed the presence of active SRB in the bacterial layer dominated by the phototrophic sulphur bacteria and introduced the possibility of a significant sulphur cycling within the chemocline coupled to a rapid turnover of sulphide (Fritz and Bachofen 2000). The vertical distribution profiles of SRB detected with probes SRB385 and SRB385Db were similar to those of the phototrophic sulphur bacteria and amounted on average 24 % of the DAPI-stained bacteria in the chemocline before the rise of the GSB Chl. clathratiforme population at the beginning of the twenty-first century. This percentage shows that SRB made up a significant part of the bacterial population in the chemocline of Lake Cadagno (Tonolla et al. 1998a, 1999, 2000). The in situ observations also reveal majority of the SRB consisted of one particular morphotype, which was observed in cell aggregates formed by small-celled phototrophs (Fig 7.3).

Aggregates of small-celled purple sulphur bacteria (in purple-violet, coccoid cells) and their association with sulphate-reducing bacteria of the genus Desulfocapsa (pink-red rod shaped cells) in samples from the chemocline of Lake Cadagno. Confocal laser scanning microscopy image
Desulfomonile tidjei in the monimolimnion of Lake Cadagno is easily recognizable as it emits a green fluorescence after acridine orange staining (Bensadoun et al. 1998) and is detectable with the two specific probes DsmA445 and DsmB455 (Tonolla et al. 2005a). Its vertical distribution was unusual, compared with other SRB, as it did not produce a maximum in abundance in the chemocline , but its distribution pattern followed that of hydrogen sulphide profiles. It accounted from 17 to 44 % of the total bacterial numbers, especially at 15–17 m depth (Bensadoun et al. 1998; Tonolla et al. 1998a, 2005a). Seasonal maximum was most frequently around 30 %, the calculated biovolumes reaching values between 10 and 17 % of the total bacterial volume (Tonolla et al. 1998a, 2005a).
5.2 Interactions and Functions of the Microbial Populations
Long-term Alternation Between Green and Purple Bacteria
The long-term changes of GSB and PSB in Lake Cadagno were followed by analysing pigments of photosynthetic sulphur bacteria and 16S rRNA gene sequences in long-core sediment samples. The carotenoid pigments okenone and isorenieratene were extracted and identified, and their concentrations measured semi-quantitatively by chromatography (Wirth et al. 2013). Total DNA was extracted, and, for each of the sediment samples, the number of copies of 16S rRNA genes of nine targeted phototrophic sulphur bacterial populations (seven Chromatiaceae and two Chlorobiaceae) was determined by quantitative PCR (Ravasi et al. 2012b).
Out of the seven Chromatiaceae populations analysed using DNA sequences, six contained okenone and only one, Lamprocystis roseopersicina , contained rhodopinal (Peduzzi et al. 2011, 2012). During 1990–2012, L. roseopersicina had very low numbers in the water column and therefore constituted a minor fraction of the total Chromatiaceae population (Tonolla et al. 2005b). Both the investigated Chlorobiaceae produce isorenieratene . Therefore, as expected, records from lipid biomarkers (carotenoid pigments) and DNA analysis showed a consistent pattern (Wirth et al. 2013).
The GSB/PSB ratio was found to vary during the evolution of the lake. In the early Holocene (10.5–8 cal kyr BP), GSB weakly dominated over PSB. On the contrary, the middle Holocene (8–5 cal kyr BP) was characterized by a minor prevalence of PSB. In the period <5 cal kyr BP, flood frequencies in the lake area increased. Floods and mass movement events (i.e. mobilization, mixing and redeposition of sediments from shallower parts of the lake to the deepest lake area) generated turbiditic underflows that sporadically introduced O2-rich water to the hypolimnion, thus briefly weakening the sulphidic conditions. Concomitantly with the rise in flood frequency, the relative abundance of GSB increased again. Here, the two isorenieratene peaks at 3.7 and 3.4 cal kyr BP seemed to correspond to flood-frequency highs with a time lag of roughly 50–100 years (e.g. at 4.3, 3.8, 3.4 and weakly at 2.5 thousand years ago). Enhanced light absorption due to higher suspended particle concentrations and algal/cyanobacterial growth in the surface and subsurface waters during floods likely reduced light availability within the chemocline . Since GSB are known to be more low-light-tolerant than PSB (Biebl and Pfennig 1978; Brocks and Schaeffer 2008), this could have triggered the observed shift in the bacterial community from a PSB- to a GSB-dominated population (Tonolla et al. 2005a; Decristophoris et al. 2009; Gregersen et al. 2009; Wirth et al. 2013). In any case, there appears to be a time lag of 50–100 years between the deposition of the most prominent flood layers and the subsequent restructurings of the phototrophic community. It is thus uncertain what exactly controlled past shifts in the microbial community structure in the water column of Lake Cadagno . Between 1.5 and 0.5 cal kyr BP, flood frequency decreased, while the Mo burial rate again increased and PSB became more abundant. However, with increasing flood activity towards the present time, the bacterial community structure has again shifted to dominance of GSB.
Microbial Cell–Cell Interactions: The Case of Lake Cadagno aggregates
We already showed that in the chemocline, up to 15 % of the cells of the total microbial community were associated in three-dimensional cell–cell aggregates (Peduzzi et al. 2003b; Decristophoris et al. 2009). The isolate Cad16 was recently proposed as Candidatus “ Thiodictyon syntrophicum ” sp. nov. strain Cad16T, a provisionally novel species within the genus Thiodictyon (Peduzzi et al. 2012). This PSB strain Cad16T together with the SRB Desulfocapsa thiozymogenes (isolate Cad626) forms stable aggregates in the lake. We will further refer to these aggregates as Lake Cadagno aggregates. Aggregation of cells of Candidatus “T. syntrophicum” and Desulfocapsa thiozymogenes is not mandatory as it is for other consortia (Overmann and Schubert 2002), since in their natural environment both partners can be encountered as free-living cells although numerically inconspicuous (Peduzzi et al. 2003b; Decristophoris et al. 2009).
Major community shifts and changes have recently been observed not only in the phototrophic sulphur bacteria but also in size and composition of aggregates in Lake Cadagno (see in Sect. 5.1.1.2: Temporal and spatial distribution of the anoxygenic phototrophic sulphur bacteria). Aggregates’ particle size decreased, whereas in 2004, compared with 1998, the number of SRB cells in the aggregates increased about three times (Fig 7.3) (Decristophoris et al. 2009). A reduction in size of Lake Cadagno aggregates, due to reduced abundance of small-celled PSB, was previously described for winter and spring periods when ice and snow cover had reduced light transmission (Peduzzi et al. 2003b). Although the driving forces that are regulating aggregate formation and composition are still not well understood, overall, these findings suggest that the aggregate formation is a dynamic association responding to shifts in environmental conditions (e.g. changes in sulphide and oxygen concentration as suggested also by Overmann 1997) and to changes in global microbial community structure.
Ecological consequences of aggregate formation may be important for other members of the microbial community not directly involved in aggregation since aggregation might significantly alter competition dynamics among the phototrophic sulphur bacteria community. During periods of intense sulphide photo-oxidation, anoxygenic photosynthesis at the upper boundary of the bacterial layer is limited by the availability of reduced sulphur compounds, a situation encountered in Lake Cadagno (Luthy et al. 2000) and also Lake Mahoney (Overmann 1997) and other lakes (Shira, Shunet, Cisò, etc.) treated in this book (see Chaps. 7 and 9 of part II). Since sulphide and light occur in vertical opposing gradients with depth in the lake, phototrophic sulphur bacteria can find suitable conditions for growth only in a narrow zone of overlap between the two opposing gradients. Under these conditions, Candidatus “T. syntrophicum” in aggregates could have a growth advantage over non-associated phototrophic sulphur bacteria since the sulphate-reducing partner could serve as a permanent source of sulphide. Thus, an internal sulphur cycling has the potential to influence competition dynamics among the anaerobic photosynthetic community.
Peduzzi et al. (2003a, b) discussed the potential physiological interactions between the microbial partners in Lake Cadagno aggregates . However, from the results presented, it is evident that the role of both sulphur and carbon compounds in the interactions of the bacteria in this association needs to be elucidated in detail. Exploiting nanoSIMS provided more evidence that the metabolic interaction and exchange are not limited to sulphur compounds but most probably also involve organic metabolites (Musat et al. 2008, 2012).
5.3 Novel Eukaryotic Microbes in the Anoxic Water Column
Eukaryotic microbial species diversity in freshwater ecosystems is not well understood and especially so in permanently anoxic systems (Saccà 2012). Currently, only a few molecular sequencing studies exist that address this issue in meromictic freshwater lakes (Lefèvre et al. 2007; Triadó-Margarit and Casamayor 2012; Gies et al. 2014). From a species biodiversity perspective, meromictic lakes are the only freshwater lakes suitable for obligate anoxic organisms and should, at least in theory, hold a higher biodiversity and possibly also higher endemism, the surrounding oxic environment simply acting as a barrier to dispersal.
The anaerobic eukaryotic biodiversity of Lake Cadagno was examined by constructing SSU rRNA clone libraries (Frigaard, unpublished data). Samples for the SSU rRNA clone libraries were collected from three depths: the lower oxic mixolimnion (10 m), the chemocline (11.5 m) and the anoxic monimolimnion (15 m), during 24–28 September 2006. The cell size fraction of the microorganisms analysed was in the range between 0.2 and 149 μm. Between 49 and 98 SSU rRNA sequences per depth were retrieved from the libraries made. Further details about the physico-chemical measurements, sampling and sample preparation are described elsewhere (Gregersen et al. 2009). At the time of sampling, the Chl a concentration in the samples from the mixolimnion, chemocline and monimolimnion was approx. 7, 45 and <1 μg L−1, respectively. The ratio of Chl c to Chl a was about 0.25 in both the mixolimnion and chemocline, indicating that Chl c-containing algae were present. Chl c was undetectable in the monimolimnion.
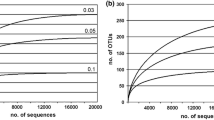
The non-parametric richness estimator SChao1 values were about twofold higher in the monimolimnion than in the mixolimnion and about fourfold higher than in the oxycline, which indicates a more complex eukaryotic community at the time of sampling in the monimolimnion than in the oxycline or mixolimnion. Cryptophytes and ciliates dominated the libraries from Lake Cadagno (Fig. 7.4). The dominance of cryptophytes in the chemocline corresponds well to the high concentration of Chl c found here (11 μg L−1). The other common groups encountered in the samples included choanoflagellates (related to Monosiga and Desmarella) and metazoan rotifers (Monogononta), retrieved from all three libraries, and fungi and ichthyosporean sequences retrieved from the mixo- and monimolimnic samples. The absence of Chl b in the examined samples corresponds well to the absence of sequences from chlorarachniophytes, green algae and euglenophytes.

Distribution of the main eukaryotic groups present in Lake Cadagno based on the number of clones as compared with total numbers of clones retrieved (see text for details). “Others” include sequences and OTUs (operational taxonomic units) of uncertain taxonomic origin as well as OTUs of heterolobosean and katablepharidean origin
One cryptophyte operational taxonomic unit (OTU) dominated both the oxycline and monimolimnion clone libraries based on the number of clones retrieved, but no reference sequence in GenBank matched the retrieved SSU rRNA sequences. However, the cloning generated nine nucleomorph SSU rRNA sequences identical to the nucleomorph sequence of Cryptomonas phaseolus (Gregersen et al. 2009). This species has been found to be sulphide tolerant and to be the dominant single cryptophyte species in the deep chlorophyll maximum in Lake Cisó, a holomictic lake in Spain that periodically experiences total anoxia with high levels of sulphide in the hypolimnion (Pedrós-Alió et al. 1987). C. phaseolus was also dominant in the holomictic Lake Großer Vätersee in Germany (Gervais et al. 2003), which also has sulphide-rich hypolimnic water. It forms stable populations, probably due to the lack of competition from other eukaryotes in the oxycline, one of the normal mechanisms believed to account for succession (Gasol et al. 1992).
Within the alveolates, the dominant group was the Ciliophora (ciliates ). As expected, different sequences were retrieved from the oxic and anoxic samples, as ciliates in general are adapted to varying oxygen concentrations. The ciliate sequences found in the monimolimnic sample were most closely related to sequences from species known to be obligate anaerobes: Caenomorpha uniserialis , Cyclidium porcatum , Metopus contortus and Trimyema compressum . All four species have been found to harbour endosymbiotic methanogenic bacteria (Wagener et al. 1990) and also other bacteria with unknown function (Clarke et al. 1993; Bruggen et al. 1983).
In the genetic library for the monimolimnion, four sequences retrieved were related to the heterolobosean amoeba Sawyeria marylandensis (Fig. 7.5). The latter was isolated from an anoxic salt marsh sediment sample and was found to be micro-aerophilic (O’Kelly et al. 2003). Together with Monopylocystis visvesvarai and Psalteriomonas lanterna, it forms a highly supported clade within the heteroloboseans . As the Lake Cadagno OTU was found to be unique (<91 % sequence identity), a water sample from the lake was screened for possible amoeboid cells, but none were encountered. However, in both the oxycline and monimolimnic samples, a round organism was present in large numbers, feeding ferociously on the photosynthetic sulphur bacteria present in large numbers (Fig. 7.6a). Single-cell PCR of this organism revealed that it had an identical SSU rRNA with the heterolobosean OTU. Phase-contrast microscopy revealed two flagella in this organism, one long and thin and the other one short and thick (Fig. 7.6b). The general outline and presence of flagella distinguish this organism from the outline of Sawyeria marylandensis, and it is likely that it represents both a new genus and a new species. However, we consider that for a formal description, an ultrastructural examination needs to be conducted.

Phylogenetic tree of a subset of monimolimnic SSU rRNA sequences (including the heterolobosean sequences) based on Bayesian analysis. Support values are Bayesian posterior probability values and neighbour-joining bootstrap support values

Light micrographs of the novel heterolobosean organism observed in Lake Cadagno. (a) Bright-field micrograph showing the general outline of the cell. The cell interior is filled with ingested phototrophic sulphur bacteria. (b) Phase-contrast micrograph showing the ingestion of phototrophic sulphur bacteria. Arrow points to the longest flagellum; the smaller flagellum is not visible. The bacterium slides along the flagellum and ingestion occurs along close to the flagellar base. Scale bars = 10 μm
While the abundance of alveolate ciliates , choanoflagellates and fungi sequences in Lake Cadagno is similar to that found in molecular studies on protists in Lake Pavin in France (Lefèvre et al. 2007), the absence of cercozoan sequences in Lake Cadagno is surprising, unlike in the mixolimnion and in the oxycline of Lake Pavin. Lefranc et al. (2005) retrieved cercozoan sequences only from the oligomesotrophic Lake Pavin and the eutrophic Lake Aydat, in France. Thus, the absence of cercozoan organisms might be a consequence of nutrient limitation. Lake Cadagno can be characterized as an oligotrophic to oligomesotrophic lake, and this might partly explain the absence of cercozoans. However, further studies are needed to clarify whether chemical differences or population dynamics could explain the observed difference.
In conclusion, eukaryotic primary producers in the anoxic zones were dominated by Cryptomonas phaseolus . Eukaryotic heterotrophs were dominated by ciliates and choanoflagellates , but also included a novel clade of heteroloboseans and two novel clades of uncertain position distantly related to opisthokonts and Cercozoa. Using single-cell PCR, the novel heterolobosean clade was linked to an abundant heterotrophic flagellate in the lake that feeds on anaerobic phototrophic sulphur bacteria .
5.4 Phyto- and Zooplankton
Güttinger and Straub (1998) addressed the question whether the special conditions (meromicticity) of the lake had any influence on the composition of the diatom flora. As expected, live diatoms in Lake Cadagno have been found in the oxic layer only and to a depth of about 10 m. A list of diatom species observed in the water column in earlier (e.g. Schanz et al. 1988) and recent studies is given in Güttinger and Straub (1998). However, in their qualitative study, no species were found, which are not common in other alkaline alpine lakes of non-meromictic character (Güttinger and Straub 1998).
Other authors followed phytoplankton summer dynamics in Lake Cadagno (Schanz and Stalder 1998). At the beginning of summer, phytoplanktonic communities in the upper 10 m are characterized by a uniform vertical distribution of pennate diatoms (above all species of the genus Fragilaria , i.e. F. crotonensis and F. capucina) and centric diatoms (species of the genera Stephanodiscus , with St. hantzschii, and Cyclotella) as well as green algae (mostly Scenedesmus costato-granulatus and Dictyosphaerium subsolitatum ). Pennate and centric diatoms become dominant later in the summer. In autumn, green algae ( Echinocoleum elegans , Sphaerocystis schroeterii and Oocystis lacustris ) dominate (Camacho et al. 2000; Schanz and Stalder 1998).
Chlorophyll a concentrations in the water column vary considerably, e.g. in summer 1988 between 26 mg m−2 in late July and 4 mg m−2 in early September or in summer 1986 between 18 and 65 mg m−2, thus depending on the period of sampling, while phytoplankton primary production varies between 2 and 17 mg C m−3 h−1, from 0 to 9.5 m depth (Schanz and Friedl 1993; Bossard et al. 2001). The comparison of 1987 and 1997 production rates did not show any statistically significant variation (Bossard et al. 2001; Pasini and Schanz 1998).
Dense developments of Fragilaria capucina and Cyclotella comensis are sometimes noted in the oxic–anoxic transition zone of the upper chemocline . A few centimetres deeper, the massive presence of cryptomonads Cryptomonas erosa and Cryptomonas phaseolus sometimes gives the water an intense green colour. At the same depth, often by nearly anoxic conditions, an important zooplankton community thrives all year round (Camacho et al. 2001; Tonolla et al. 1988). The dominant species are Cladocera Daphnia longispina complex and Bosmina longirostris , which often represent up to 40 % of the zooplankton density. Rotifers constitute, however, the largest share of zooplankton diversity (15 species, according to Riccardi et al. 2012), with Conochilus sp. (35 % of total zooplankton density) and Asplanchna priodonta (10 %) (Camacho et al. 2001). Other zooplankton species encountered in significant quantities are the calanoid copepod Acanthodiaptomus denticornis and the cyclopoid Cyclops abyssorum . The complete list of zooplankton species of Lake Cadagno is available in Riccardi et al. (2012).
Recently, possible responses of the biological parameters of the Lake to climate changes were investigated through the analysis of the populations of different diatom genera found in the sediments over the last 100 years (Tiffay et al. 2015). A significant shift in relative genus abundance was observed, with a dominance of small-size planktonic diatoms in recent years (1985–2012) compared with periphytic/benthic diatoms dominated in earlier years (1900–1950). Interestingly, taxa previously not observed in the Lake (Güttinger and Straub 1998, Riccardi et al. 2012) were also identified in the sediments: Aneumastus sp., Craticula sp., Gyrosigma sp., Meridion sp. and Reimeria sp. (Tiffay et al. 2015).
In contrast to Mahoney Lake in Canada (Overmann 1997), Lake Cadagno planktonic community in oxic layer takes advantage of the bacterial development in the chemocline for growth, and zooplankton is a link between bacterial primary production and the higher levels of the food chain (e.g. fish). In situ hybridization has showed the presence of PSB in the intestine of these predators (e.g. Cyclops abyssorum ). This was further confirmed by the analysis of stable carbon isotopes in bacteria and zooplankton by which the contribution of the bacterial layer in providing carbon to the zooplankton through the latter’s predation on bacteria was estimated to be around 50 % (Camacho et al. 2001).
5.5 Fish
Meromictic conditions in Lake Cadagno support high fish productivity. The massive development of anaerobic photosynthetic bacteria in the chemocline provides a supplementary form of primary production along with that of the algae. On the whole, bacterial and phytoplankton primary productions are the beginning of a food chain that ensures the development of a fairly large fish population (Tonolla et al. 1988). Fish productivity, estimated at 25–30 kg ha−1 (Marrer 1975; Tonolla et al. 1988), represents relatively high values for an alpine oligomesotrophic lake situated at an altitude close to 2000 m asl. In fact, the lakes of the Piora region, Lakes Ritom and Cadagno in particular, have a long history of fish management, documented as far back as the fifteenth century (Surbeck 1917). For the inhabitants of this mountain region, fish represented an important supplementary source of proteins (Peduzzi 2000). Moreover, the fish of both lakes Ritom and Cadagno were renowned for their natural salmon-like colouration supposedly due to the carotenoids of the PSB inhabiting the chemocline. The presence of fish is due to man-made introductions for exploitation of fishery. The species present are rainbow trout (Oncorhynchus mykiss), brown trout (Salmo trutta), lake charr (Salvelinus umbla), brook trout (Salvelinus fontinalis), lake trout (Salvelinus namaycush), bullhead (Cottus gobio), Phoxinus lumaireul and Italian chub (Squalius squalus), though the last two species are not caught by fishermen. Brook trout and lake trout were introduced to the lake in the 1950s and fry of the commercial fish species are introduced annually (Polli 2012).
6 Concluding Remarks
Lake Cadagno represents an ideal model lake for studying the role of microbes in aquatic environments. The scientific projects performed over many decades offer an important source of information supporting ongoing and future scientific projects. Despite its constant meromictic state, Lake Cadagno is unique in that it is responding to the ongoing climatic changes, and man-made changes are influencing the lake itself as well as the catchment area. In addition, continuing studies of the major bacteria involved in the sulphur cycle, as well as studies of microbial interactions and the investigation of the microbes from the mixolimnion and from the sulphidogenic sediments, will contribute to advance our knowledge of this interesting ecosystem.
Meromictic lakes such as Lake Cadagno can be used as a model for studying biogeochemical processes mediated by microorganisms and characterized by permanently anoxic water columns, as earlier reported for Lake Cadagno (Hanselmann and Hutter 1998; Putschew et al. 1995). In this perspective, of great interest are the investigations promoted by the Nordic Center of Earth Evolution focusing on the long history and evolution of life on Earth.
Since anoxic and sulphidic marine conditions may have prevailed during the Precambrian (Canfield et al. 1998; Meyer et al. 2011), Lake Cadagno has the potential to provide insights into the biogeochemical cycling of elements in the Proterozoic oceans and thus can be considered as the present-day analogue to the ancient prokaryote-dominated oceans (Canfield et al. 2010; Honeycutt et al. 2008), considering particularly the bacteria involved in the sulphur cycling (Johnston 2011). Phototrophic sulphur bacteria, especially the GSB Chlorobium spp., have been suggested to proliferate in ancient anoxic ocean basins (Crowe et al. 2008; Summons and Powell 1987; Halm et al. 2009). They are, indeed, believed to have been important for the carbon fixation and chemical development of the anoxic Precambrian ocean, where they may have contributed to the genesis of banded iron formations and the stabilization of the sulphide-rich ocean during the mid-Proterozoic period, Proterozoic aeon 2500–542 million years ago (Brocks et al. 2005; Crowe et al. 2008; Habicht et al. 2011; Johnston et al. 2009).
Further, we applied in Lake Cadagno very innovative investigation techniques, which are among the most promising and powerful in the field of molecular microbial ecology to study of the lake’s ecosystem. Thus, Lake Cadagno and its sulphur phototrophic bacteria and associated sulphate-reducing bacteria have proven to represent an excellent model system to test and develop novel investigation techniques in both environmental microbiology and microbial ecology. Particularly interesting are the methods described in Musat et al. (2008), Halm et al. (2009), Habicht et al. (2011) and Storelli et al. (2013) to infer or measure in situ activities of single cells of diverse groups of organisms (e.g. Chromatium okenii ). These techniques have now the potential to be applied to more difficult environments such as some other aquatic environments, running waters, sediments and microbial mats or soils.
In the studies reviewed here, much of the attention was devoted to population dynamics of phototrophic sulphur bacteria comprising both an apparently monophyletic GSB community and a multiple and diverse PSB community. One question raised but yet unanswered is about the coexistence in the apparently same ecological niche of diverse, but closely related, populations of phototrophic sulphur bacteria. This coexistence and alternation has been present on a long timescale, since the formation of euxinic conditions in the lake, soon after its formation, about 10,000 years BP.
As already pointed out above, other functional groups in the bacterial community such as SRB or, as highlighted in a recent work (Berg et al. 2016), bacteria involved in the iron cycle have a great impact in the functioning of this ecosystem. In particular, sulphate-reducing bacteria in Lake Cadagno aggregates may well play a major role in competition dynamics among the phototrophic populations. Thus, the mechanisms underlying PSB–SRB aggregate formation certainly deserve more attention. The study of aggregates, with the tools and techniques described above, potentially integrates all levels of biological analysis from molecular to ecological and holds the promise to give new insights into the metabolic coupling between organisms in aquatic systems.
There are striking parallels and analogies between Lake Cadagno and other stratified meromictic ecosystems worldwide, from microbial mats to ancient oceans. The most interesting analogy is with a low sulphidic and prokaryote-dominated Proterozoic ocean (2500–542 million years ago). Research on this particular ecosystem thus holds the promise to give insights into many fascinating aspects of lake biology and biogeology like life and Earth history and into some of the growing interdisciplinary fields in environmental microbiology: the microbial community dynamics and interactions, as well as the single-cell trait-based biodiversity in microbial communities and its link to ecosystem functioning.
References
Bachmann H (1924) Der Ritomsee. Schweiz Z Hydrol 2:7–28
Bachmann H (1928) Das Phytoplankton der Pioraseen nebst einigen Beiträgen zur Kenntnis des Phytoplanktons schweizerischer Alpenseen. Schweiz Z Hydrol 5:50–103
Barades J (1926) Le percement du Lac de la Girotte. L’Illus 4338:400–401
Bensadoun J-C, Tonolla M, Demarta A et al (1998) Vertical distribution and microscopic characterization of a non-cultivable microorganism (morphotype R) of Lake Cadagno. Doc Ist Ital Idrobiol 63:45–51
Berg IA (2011) Ecological aspects of the distribution of different autotrophic CO2 fixation pathways. Appl Environ Microbiol 77:1925–1936
Berg JS, Michellod D, Pjevac P, Martinez-Perez C, Buckner CRT, Hach PF, Schubert CJ, Milucka J, Kuypers MMM (2016) Intensive cryptic microbial iron cycling in the low iron water column of the meromictic Lake Cadagno. Environ Microbiol. doi:10.1111/1462-2920.13587
Bertoni R, Callieri C, Pugnetti A (1998) Dinamica del carbonio organico nel Lago di Cadagno e attività microbiche nel mixolimnio. Doc Ist Ital Idrobiol 63:11–17
Biebl H, Pfennig N (1978) Growth yields of green sulfur bacteria in mixed cultures with sulfur and sulfate reducing bacteria. Arch Microbiol 117:9–16
Borner L (1928a) Die Bodenfauna des Lago Ritom und seines Deltagebiets vor der Absenkung (1916) I. Schweiz Z Hydrol 4:121–162
Borner L (1928b) Die Bodenfauna des Lago Ritom und seines Deltagebiets vor der Absenkung (1916) II. Schweiz Z Hydrol 4:1–30
Bossard P, Gammeter S, Lehman C et al (2001) Limnological description of the lakes Zürich, Lucerne, and Cadagno. Aquat Sci 63:225–249
Bosshard PP, Santini Y, Grüter D et al (2000a) Bacterial diversity and community composition in the chemocline of in the meromictic Lake Cadagno as revealed by 16S rDNA analysis. FEMS Microbiol Ecol 31:173–182
Bosshard PP, Stettler R, Bachofen R (2000b) Seasonal and spatial community dynamics in the meromictic Lake Cadagno. Arch Microbiol 174:168–174
Bottinelli M (2008) Approche moléculaire à l’étude des bactéries sulfato-réductrices et des Archaea méthanogènes dans les sédiments des lacs Cadagno et Rotsee. Dissertation, Université de Genève, Terre et Environnement 71
Boucherle MM, Züllig H (1988) Lago Cadagno: an environmental history. In: Lang G, Schlüchter C (eds) Lake, mire and river environments. Balkema Publishers, Rotterdam, pp 234–295
Bourcart FE (1906) Les lacs alpins suisses, étude chimique et physique. Dissertation, Université de Genève, Georg & Co., Genève
Brocks JJ, Love GD, Summons RE et al (2005) Biomarker evidence for green and purple sulphur bacteria in a stratified Palaeoproterozoic sea. Nature 437:866–870
Brocks JJ, Schaeffer P (2008) Okenane, a biomarker for purple sulfur bacteria (Chromatiaceae), and other new carotenoid derivatives from the 1640 Ma Barney Creek Formation. Geochim Cosmochim Acta 72:1396–1414
Bruggen JJA, Stumm CK, Vogels GD (1983) Symbiosis of methanogenic bacteria and sapropelic protozoa. Arch Microbiol 136:89–95
Burckhardt G (1910) Das Plankton des Lago Ritom unter dem Einfluss der Schwefelquellen. Verh Schweiz Naturforsch Ges 93:302–303
Camacho A, Vicente E, Miracle MM (2000) Spatio-temporal distribution and growth dynamics of phototrophic sulfur bacteria populations in the sulfide-rich lake Arcas. Aquat Sci 62:334–349
Camacho A, Erez J, Chicote A et al (2001) Microbial microstratification inorganic carbon photoassimilation and dark carbon fixation at the chemocline of the meromictic Lake Cadagno (Switzerland) and its relevance to the food web. Aquat Sci 63:91–106
Canfield DE, Thamdrup B, Fleischer S (1998) Isotope fractionation and sulfur metabolism by pure and enrichment cultures of elemental sulfur disproportionating bacteria. Limnol Oceanogr 43:253–264
Canfield DE, Farquhar J, Aubrey L et al (2010) High isotope fractionations during sulfate reduction in a low-sulfate euxinic ocean analog. Geology 38:415–418
Clarke KJ, Finlay BJ, Esteban G et al (1993) Cyclidium porcatum n. sp.: a free-living anaerobic scuticociliate containing a stable complex of hydrogenosomes, eubacteria and archaeobacteria. Eur J Protistol 29:262–270
Collet LW, Mellet R, Ghezzi C (1918) Le lac Ritom. Département suisse de l’intérieur. Communication du service des eaux (Dr C Mutzner) Berne Nr. 13
Crowe SA, Jones C, Katsev S et al (2008) Photoferrotrophs thrive in an Archean ocean analogue. Proc Natl Acad Sci USA 105:15938–15943
Dahl TW, Anbar AD, Gordon GW et al (2010) The behavior of molybdenum and its isotopes across the chemocline and in the sediments of sulfidic Lake Cadagno, Switzerland. Geochim Cosmochim Acta 74:144–163
Dahl TW, Chappaz A, Fitts JP et al (2013) Molybdenum reduction in a sulfidic lake: evidence from X-ray absorption fine-structure spectroscopy and implications for the Mo paleoproxy. Geochim Cosmochim Acta 103:213–231
Decristophoris PMA, Peduzzi S, Ruggeri-Bernardi N et al (2009) Fine scale analysis of shifts in bacterial community structure in the chemocline of meromictic Lake Cadagno, Switzerland. J Limnol 68:16–24
Del Don C, Hanselmann KW, Peduzzi R et al (2001) The meromictic alpine Lake Cadagno: orographycal and biogeochemical description. Aquat Sci 63:70–90
Delebeque A (1898) Les lacs français. Chamerot & Renouard, Paris
Demarta A, Tonolla M, Caminada AP et al (1998) Phylogenetic diversity of the bacterial community from the anoxic layer of the mero-mictic Lake Cadagno. Doc Ist Ital Idrobiol 63:19–30
Düggeli M (1924) Hydrobiologische Untersuchungen im Pioragebiet. Bakteriologische Untersuchungen am Ritomsee. Schweiz Z Hydrol 2:62–205
Eichler B, Pfennig N (1988) A new purple sulfur bacterium from stratified freshwater lakes, Amoebobacter purpureus sp. nov. Arch Microbiol 149:395–400
Ehrenreich A, Widdel F (1994) Anaerobic oxidation of ferrous iron by purple bacteria, a new tyoe of phototrophic metabolism. Appl Environ Microbiol 60:4517–4526
Fischer C, Wiggli M, Schanz F et al (1996) Light environment and synthesis of bacteriochlorophyll by populations of Chromatium okenii under natural environmental conditions. FEMS Microbiol Ecol 21:1–9
Frigaard NU, Bryant D (2006) Chlorosomes: antenna organelles in photosynthetic green bacteria. Microbiol Monogr 2:79–114
Frigaard NU, Dahl C (2009) Sulfur metabolism in phototrophic sulfur bacteria. Adv Microb Physiol 54:103–200
Fritz M, Bachofen R (2000) Volatile organic sulfur compounds in a meromictic Alpine lake. Acta Hydrochim Hydrob 28:185–192
Garwood EJ (1906) The tarns of Canton Ticino. Q J Geol Soc 62:165–195
Gasol JM, Guerrero R, Pedrós-Alió C (1992) Spatial and temporal dynamics of a metalimnetic Cryptomonas peak. J Plankton Res 14:1565–1579
Gervais F, Siedel U, Heilmann B et al (2003) Small-scale vertical distribution of phytoplankton, nutrients and sulphide below the oxycline of a mesotrophic lake. J Plankton Res 25:273–278
Gies EA, Konwar KM, Beatty JT et al (2014) Illuminating microbial dark matter in meromictic Sakinaw Lake. Appl Environ Microbiol 80:6807–6818
Gregersen LH, Habicht KS, Peduzzi S et al (2009) Dominance of a clonal green sulfur bacterial population in a stratified lake. FEMS Microbiol Ecol 70:30–41
Guerrero R, Montesinos E, Pedrós-Alió C, Esteve I, Mas J, van Gemerden H (1985) Phototrophic sulfur bacteria in two Spanish lakes: vertical distribution and limiting factors. Limnol Oceanogr 30:919–931
Güttinger W, Straub F (1998) Diatoms of Lake Cadagno. Doc Ist Ital Idrobiol 63:57–64
Habicht KS, Miller M, Cox RP et al (2011) Comparative proteomics and activity of a green sulfur bacterium through the water column of Lake Cadagno, Switzerland. Environ Microbiol 13:203–215
Halm H, Musat N, Lam P et al (2009) Co-occurrence of denitrification and nitrogen fixation in a meromictic lake, Lake Cadagno (Switzerland). Environ Microbiol 11:1945–1958
Hanselmann K, Hutter R (1998) Geomicrobiological coupling of sulfur and iron cycling in anoxic sediments of a meromictic lake: sulfate reduction and sulfide sources and sinks in Lake Cadagno. Doc Ist Ital Idrobiol 63:85–98
Hebting Y, Schaeffer P, Behrens A et al (2006) Biomarker evidence for a major preservation pathway of sedimentary organic carbon. Science 312:1627–1631
Hegler F, Posth NR, Jiang J et al (2008) Physiology of phototrophic iron(II)-oxidizing bacteria: implications for modern and ancient environments. FEMS Microbiol Ecol 66:250–260
Honeycutt C, Canfield D, Bjerrum C (2008) One dimensional physico-chemical ecological modelling of sulfidic oceans: applications to proterozoic ocean chemistry and euxinic photoic zones. Geophysical Research Abstracts, vol 10, EGU: 2008-A-08712, 2008 EGU General Assembly 2008
Imhoff JF (2001) Transfer of Pfennigia purpurea Tindall 1999 (Amoebobacter purpureus Eichler and Pfennig 1988) to the genus Lamprocystis as Lamprocystis purpurea comb. nov. Int J Syst Evol Microbiol 51:1699–1701
Johnston DT, Wolfe-Simon F, Pearson A et al (2009) Anoxygenic photosynthesis modulate proterozoic oxygen. Proc Natl Acad Sci USA 106:16925–16929
Johnston DT (2011) Multiple sulfur isotopes and the evolution of earth’s surface sulfur cycle. Earth-Sci Rev 106:161–183
Jørgensen BB, Kuenen JG, Cohen Y (1979) Microbial transformations of sulfur compounds in a stratified lake (Solar Lake, Sinai). Limnol Oceanogr 24:799–822
Krige LJ (1918) Petrographische Untersuchungen im Val Piora und Umgebung. Ecol Geol Helv 14:549–654
Langille MGI, Zaneveld J, Caporaso JG et al (2013) Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat Biotechnol 31:814–823
Lefèvre E, Bardot C, Noël C et al (2007) Unveiling fungal zooflagellates as members of freshwater picoeukaryotes: evidence from a molecular diversity study in a deep meromictic lake. Environ Microbiol 9:61–71
Lefranc M, Thénot A, Lepère C et al (2005) Genetic diversity of small eukaryotes in lakes differing by their trophic status. Appl Environ Microbiol 71:5935–5942
Lehmann C, Bachofen R (1999) Images of concentrations of dissolved sulphide in the sediment of a lake and implications for internal sulphur cycling. Sedimentology 46:537–544
Luthy L, Fritz M, Bachofen R (2000) In situ determination of sulfide turnover rates in a meromictic alpine lake. Appl Environ Microbiol 66:712–717
Marrer H (1975) Zur fischereilichen Bewirtschaftung meromiktischer Alpenseen (Lago di Cadagno, Tessin). Schweiz Z Hydrol 37:213–219
Meyer KM, Macalady JL, Fulton JM et al (2011) Carotenoid biomarkers as an imperfect reflection of the anoxygenic phototrophic community in meromictic Fayetteville Green Lake. Geobiology 9:321–329
Milucka J, Kirf M, Lu L et al (2015) Methane oxidation coupled to oxygenic photosynthesis in anoxic waters. ISME J. doi:10.1038/ismej.2015.12
Musat N, Halm H, Winterholler B et al (2008) A single-cell view on the ecophysiology of anaerobic phototrophic bacteria. Proc Natl Acad Sci 105:17861–17866
Musat N, Foster R, Vagner T et al (2012) Detecting metabolic activities in single cells, with emphasis on nanoSIMS. FEMS Microbiol Rev 36:486–511
Niemann H, Stadnitskaia A, Wirth SB et al (2012) Bacterial GDGTs in holocene sediments and catchment soils of a high Alpine lake: application of the MBT/CBT-paleothermometer. Clim Past 8:889–906
O’Kelly CJ, Silberman JD, Amaral Zettler LA et al (2003) Monopylocystis visvesvarai n. gen., n. sp. and Sawyeria marylandensis n. gen., n. sp.: two new amitochondrial Heterolobosean amoebae from anoxic environments. Protist 154:281–290
Otz MH, Otz HK, Otz I et al (2003) Surface water/groundwater interaction in the Piora aquifer, Switzerland: evidence from dye tracing tests. Hydrgeol J 11:228–239
Overmann J, Beatty T, Hall KJ, Pfennig N, Northcote TG (1991) Characterization of a dense, purple sulfur bacterial layer in a meromictic lake. Limnol Oceanogr 36:846–859
Overmann J, Beatty T, Krouse HR, Hall KJ (1996) The sulfur cycle in the chemocline of a meromictic salt lake. Limnol Oceanogr 41:147–156
Overmann J (1997) Mahoney lake: A case study of the ecological significance of phototrophic sulfur bacteria. In: Jones JG (ed) Advances in microbial ecology, vol 15. Plenum Press, New York, p 251
Overmann J, Schubert K (2002) Phototrophic consortia: model systems for symbiotic interrelations between prokaryotes. Arch Microbiol 177:201–208
Overmann J, Garcia-Pichel F (2006) The phototrophic way of life. In: Dworkin M, Falkow S, Rosenberg E et al (eds) The prokaryotes, Ecophysiology and biochemistry, vol 2. Springer, New York, pp 32–85
Parkin TB, Brock TD (1981) The role of phototrophic bacteria in the sulfur cycle of a meromictic lake. Limnol Oceanogr 26:880–890
Pasini P, Schanz F (1998) Influence of UV-radiation on the primary production of two high mountain lakes in the Piora region. Doc Ist Ital Idrobiol 63:65–70
Pedrós-Alió C, Gasol JM, Guerrero R (1987) On the ecology of a Cryptomonas phaseolus population forming a metalimnetic bloom in Lake Cisó, Spain: annual distribution and loss factors. Limnol Oceanogr 32:285–298
Peduzzi R, Bachofen R, Tonolla M (1998) Lake Cadagno: a meromictic alpine lake. Doc Ist Ital Idrobiol 63
Peduzzi R (2000) Piora e San Gottardo, due secoli di ricerca scientifica. In: Peduzzi R (ed) La ricerca alpina e le trasversali. Atti delle 178a assemblea annuale dell’ASSN 1998, vol 1. Documenta Centro di Biologia Alpina, Piora, pp 17–25
Peduzzi R, Bianconi F (2012) Storia della ricerca biologica e limnologica. In: Rampazzi F, Tonolla M, Peduzzi R (eds) Biodiversità della Val Piora—Risultati e prospettive delle “Giornate della biodiversità”. Mem Soc Ticinese Sci Nat 11:13–19
Peduzzi S, Tonolla M, Hahn D (2003a) Vertical distribution of sulfate-reducing bacteria in the chemocline of Lake Cadagno, Switzerland, over an annual cycle. Aquat Microb Ecol 30:295–302
Peduzzi S, Tonolla M, Hahn D (2003b) Isolation and characterization of aggregate-forming sulfate-reducing and purple sulfur bacteria from the chemocline of meromictic Lake Cadagno, Switzerland. FEMS Microbiol Ecol 45:29–37
Peduzzi S (2011) Wasserkraft Val Piora-Piotta. Wege durch die Wasserwelt—5.1. Region Tessin, Hydrologischer Atlas der Schweiz-Geographisches Institut der Universität Bern, Bern: p 42
Peduzzi S, Welsh A, Demarta A et al (2011) Thiocystis chemoclinalis sp. nov. and Thiocystis cadagnonensis sp. nov., motile purple sulfur bacteria isolated from the chemocline of a meromictic lake. Int J Syst Evol Microbiol 61:1682–1687
Peduzzi S, Storelli N, Welsh A et al (2012) Candidatus “Thiodictyon syntrophicum”, sp. nov., a new purple sulfur bacterium isolated from the chemocline of Lake Cadagno forming aggregates and specific associations with Desulfocapsa sp. Syst Appl Microbiol 35:139–144
Pfennig N, Cohen-Bazire G (1967) Some properties of the green bacterium Pelodictyon clathratiforme. Arch Mikrobiol 59:226–236
Polli B (2012) Pesci della Val Piora (Cantone Ticino, Svizzera). In: Rampazzi F, Tonolla M, Peduzzi R (eds) Biodiversità della Val Piora—Risultati e prospettive delle “Giornate della biodiversità”. Memorie della Società Ticinese di Scienze Naturali 11:237–240
Putschew A, Scholz-Bättcher BM, Rullkätter J (1995) Organic geochemistry of sulfur-rich surface sediments of meromictic lake Cadagno, swiss Alps. In: Vairavamurthy MA, Schoonen MAA (eds) Geochemical transformations of sedimentary sulfur. ACS Symposium Series 612, Oxford Press, pp 59–78
Ravasi DF, Peduzzi R, Tonolla M (2012a). La diversità microbica del Lago Cadagno e degli endoliti della Val Piora (Cantone Ticino, Svizzera). In: Rampazzi F, Tonolla M, Peduzzi R (eds) Biodiversità della Val Piora—Risultati e prospettive delle “Giornate della biodiversità”. Mem Soc Ticinese Sci Nat 11:69–78
Ravasi DF, Peduzzi S, Guidi V et al (2012b) Development of a real-time PCR method for the detection of fossil 16S rDNA fragments of phototrophic sulfur bacteria in the sediments of Lake Cadagno. Geobiology 10:196–204
Riccardi N, Austoni M, Kamburska L et al (2012) Phytoplankton and zooplankton species distribution in the high altitude lakes of the Piora valley (Canton Ticino, Switzerland). In: Rampazzi F, Tonolla M, Peduzzi R (eds) Biodiversità della Val Piora—Risultati e prospettive delle “Giornate della biodiversità”. Mem Soc Ticinese di Sci Nat 11:77–91
Saccà A (2012) The role of eukaryotes in the anaerobic food web of stratified lakes. In: Altenbach A, Bernhard JM., Seckbach J (eds) anoxia: evidence for eukaryote survival and paleontological strategies. Cell Orig Life Extrem Habitat Astrobiol 21:403–419
Schanz F, Elber F, Huerlimann J (1988) Ricerche algali nella regione di Piora: Confronto dei risultati del periodo 1915–1928 e del 1987. Boll Soc Ticinese Sci Nat LXXVI:35–46
Schanz F, Friedl C (1993) Environmental effects on light utilization by phyto-plankton in a meromictic alpine lake. Verh Int Ver Limnol 25:621–624
Schanz F, Fischer-Romero C, Bachofen R (1998) Photosynthetic production; photoadaptatin of phototrophic sulfur bacteria in Lake Cadagno (Switzerland). Limnol Oceanogr 43:1262–1269
Schanz F, Stalder S (1998) Phytoplankton summer dynamics: sedimentation in the themally stratified Lake Cadagno. Doc Ist Ital Idrobiol 63:71–76
Schramm A, Fuchs BM, Nielsen JL et al (2003) Fluorescence in situ hybridization of 16S rRNA gene clones (Clone-FISH) for probe validation; screening of clone libraries. Environ Microbiol 4:713–720
Schubert CJ, Vazquez F, Lösekann-Behrens T et al (2011) Evidence for anaerobic oxidation of methane in sediments of a freshwater system (Lago di Cadagno). FEMS Microbiol Ecol 76:26–38
Sorokin YI (1970) Interrelations between sulphur and carbon turnover in meromictic lakes. Arch Hydrobiol 66:391–446
Stapfer A (1991) Pollenanalytische Untersuchungen im Val Piora (Tessin). Ein Beitrag zur Klima und Vegetationsgeschichte der Nacheiszeit. Geogr Helv 46:156–164
Storelli N, Peduzzi S, Saad M et al (2013) CO2 assimilation in the chemocline of Lake Cadagno is dominated by a few types of phototrophic purple sulfur bacteria. FEMS Microbiol Ecol 84:421–432
Storelli N, Saad M, Frigaard N-U et al (2014) Proteomic analysis of the purple sulfur bacterium Candidatus “Thiodictyon syntrophicum” strain Cad16T isolated from Lake Cadagno. EuPA Open Proteom 2:17–30
Summons RE, Powell TG (1987) Identification of aryl isoprenoids in source rocks and crude oils: biological markers for the green sulphur bacteria. Geochim Cosmochim Acta 51:557–566
Surbeck G (1917) Ueber die Fische des Ritom-, Cadagno- und Tomsees in Val Piora. Verh Schweiz Nat Ges 99:264–265
Tiffay M-C, Stoll S, Peduzzi S (2015) Ecosystem responses to climate variations of the high Alpine Lake Cadagno over the last 100 years. RS02. SEFS 9 Symposium for European Freshwater Sciences
Tonolla M, Del Don C, Boscolo P et al (1988) The problem of fish management in an artificially regulated meromictic lake: Lake Cadagno (Canton Tessin, Switzerland). Riv Ital Acquacolt 23:57–68
Tonolla M, Demarta A, Hahn D et al (1998a) Microscopic and molecular in situ characterization of bacterial populations in the meromictic Lake Cadagno. Doc Ist Ital Idrobiol 63:31–44
Tonolla M, Demarta A, Peduzzi R (1998b) The chemistry of Lake Cadagno. Doc Ist Ital Idrobiol 63:11–17
Tonolla M, Demarta A, Peduzzi R et al (1999) In situ analysis of phototrophic sulfur bacteria in the chemocline of meromictic Lake Cadagno (Switzerland). Appl Environ Microbiol 65:1325–1330
Tonolla M, Demarta A, Peduzzi S et al (2000) In situ analysis of sulfate-reducing bacteria related to Desulfocapsa thiozymogenes in the chemocline of meromictic Lake Cadagno (Switzerland). Appl Environ Microbiol 66:820–824
Tonolla M, Peduzzi S, Hahn D et al (2003) Spatio-temporal distribution of phototrophic sulfur bacteria in the chemocline of meromictic Lake Cadagno (Switzerland). FEMS Microbiol Ecol 43:89–98
Tonolla M, Peduzzi S, Demarta A et al (2004) Phototrophic sulfur and sulfate-reducing bacteria in the chemocline of meromictic Lake Cadagno, Switzerland. J Limnol 63:161–170
Tonolla M, Bottinelli M, Demarta A et al (2005a) Molecular identification of an uncultured bacterium (“morphotype R”) in meromictic Lake Cadagno, Switzerland. FEMS Microbiol Ecol 53:235–244
Tonolla M, Peduzzi R, Hahn D (2005b) Long-term population dynamics of phototrophic sulfur bacteria in the chemocline of Lake Cadagno, Switzerland. Appl Environ Microbiol 71:3544–3550
Triadó-Margarit X, Casamayor EO (2012) Genetic diversity of planktonic eukaryotes in high mountain lakes (Central Pyrenees, Spain). Environ Microbiol 14:2445–2456
Van Gemerden H, Mas J (1995) Ecology of phototrophic sulfur bacteria. In: Blankenship RE, Madigan MT, Bauer CE (eds) Anoxygenic photosynthetic bacteria. Kluwer Academic, The Netherlands, pp 49–85
Wagener S, Schulz S, Hanselmann K (1990) Abundance and distribution of anaerobic protozoa and their contribution to methane production in Lake Cadagno (Switzerland). FEMS Microbiol Ecol 74:39–48
Wirth SB, Gilli A, Niemann H et al (2013) Combining sedimentological, trace metal (Mn, Mo) and molecular evidence for reconstructing past water-column redox conditions: the example of meromictic Lake Cadagno (Swiss Alps). Geochim Cosmochim Acta 120:220–238
Widdel F, Schnell S, Heising S et al (1993) Ferrous iron oxidation by anoxygenic phototrophic bacteria. Nature 362:834–836
Wüest A (1994) Interaktionen in Seen: Die Biologie als Quelle dominanter physikalischer Kräfte. Limnologica 24:93–104
Zimmermann M, Escrig S, Hübschmann T et al (2015) Phenotypic heterogeneity in metabolic traits among single cells of a rare bacterial species in its natural environment quantified with a combination of flow cell sorting and NanoSIMS. Front Microbiol 6:243
Züllig H (1985) Pigmente phototropher Bakterien in Seesedimenten und ihre Bedeutung für die Seeforschung. Schweiz Z Hydrol 47:87–126
Acknowledgements
We are grateful to Prof. R. Peduzzi and the Alpine Biology Foundation, Piora, for the cooperation and support. We also would like to thank K. Habicht, D. Canfield, C. Schubert, X. Perret, J. Zopfi, M. Bueche, J. A. Wuest, A. Bruder, T. Sommer, M. Kuypers and M. Saad for their collaboration and contributions to specific projects.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Tonolla, M. et al. (2017). Lake Cadagno: Microbial Life in Crenogenic Meromixis. In: Gulati, R., Zadereev, E., Degermendzhi, A. (eds) Ecology of Meromictic Lakes. Ecological Studies, vol 228. Springer, Cham. https://doi.org/10.1007/978-3-319-49143-1_7
Download citation
DOI: https://doi.org/10.1007/978-3-319-49143-1_7
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-49141-7
Online ISBN: 978-3-319-49143-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)