
Abstract
Movement of a limb substantially decreases the intensity and sensitivity with which tactile stimuli on that limb are perceived. This movement-related tactile suppression likely interferes with performance in motor tasks that require the precise evaluation of tactile feedback, such as the adjustment of grip forces during grasping. Therefore, we hypothesise that suppression might be stronger for stimuli that are irrelevant to successful performance in a given motor task. To test this hypothesis, we measured participants’ perception of tactile intensity while performing different motor tasks. We investigated perception of both supra-threshold stimuli (Exp. 1: intensity discrimination) and of stimuli close to the detection threshold (Exp. 2: detection). We compared tactile perception between two grasping conditions (active, tactile inputs relevant), a condition where participants pointed in the air (active, tactile inputs irrelevant) and a static condition (baseline). In both experiments, we observed tactile suppression in all three movement conditions but not the predicted attenuation of tactile suppression in the grasp conditions. Contrary to our hypothesis, there was even an amplification of tactile suppression in the grasping conditions of Exp. 1, which might be related to the movement velocity. In conclusion, we did not find evidence that motor tasks modulate the strength of tactile suppression. Our results further suggest that it is important to control for possibly confounding variables, such as movement velocity and laterality, in this line of research.
Access provided by Autonomous University of Puebla. Download conference paper PDF
Similar content being viewed by others


Keywords
1 Introduction
Haptic perception is inherently active and task-dependent. For instance, the selection of motor strategies for haptic exploration depends on the estimated object property [13]. This is likely because exploratory movements modulate the precision with which properties such as the height of a Braille-like dot [7] or an object’s moment of inertia [5] can be haptically estimated. Indeed, exploratory strategies have been shown to optimise the precision of haptic estimates (e.g., [6, 15]).
Research on tactile suppression during movement, also commonly referred to as movement-related tactile gating, has generated a body of results that are difficult to reconcile with the above-mentioned literature. Comparing the tactile sensation between an active (movement) and a passive (no movement) condition, the active condition is found to reduce the perceived intensity and the tactile sensitivity (e.g., [8, 10, 14, 17]). This effect occurs robustly both for electrical stimuli applied directly in the afferent nerves, for electrical stimuli applied to the skin, and for vibrotactile stimuli applied to the skin; it was found to occur for near-threshold as well as for supra-threshold stimuli (e.g., [2, 17]); and its magnitude was found to increase with movement speed (e.g., [16]). It is puzzling that our brains, on the one hand, fine-tune the exploratory movements to maximise tactile precision while, on the other hand, suppressing the tactile sensation during movement.
One key question is: Is tactile suppression under some form of top-down neural control that enables the brain to selectively suppress only the task-irrelevant tactile stimuli? Recent research by Brozzoli et al. [1] has shown such a task-related modulation of tactile processing for visuo-tactile integration. Moreover, a recent study by Colino et al. [3] suggests that similar modulatory effects might also occur in tactile suppression: The authors studied tactile suppression during grasping, and found that stimuli were only suppressed when applied to the forearm, where they are irrelevant for grasping. Stimuli were not suppressed when they were applied to the index finger, where they are important to adjust grip forces and avoid slip of the object. However, this study might be confounded by comparably high stimulus intensities (i.e., ’readily detectable a rest’ [3]) that caused ceiling effects for the measure of tactile sensitivity in the detection task (\(d^\prime \) of approximately four). This makes it difficult to assess the strength of this attenuation. Moreover, the effect was observed when comparing stimulation in different (relevant and irrelavant) skin locations. To quantify the strength of the possible modulation of tactile suppression, one would ideally have a baseline for both suppressed and unsuppressed tactile sensation for a given stimulus location.
The current study set out to test the hypothesis that tactile suppression during movement occurs predominantly in motor task-irrelevant skin locations, using a different experimental paradigm. We studied the perceived sensation of tactile stimuli delivered to the ring finger in two different psychophysical tasks (Experiment 1: intensity discrimination, supra-threshold stimuli; Experiment 2: detection, stimuli at threshold). We compared four different movement conditions: A baseline condition for unsuppressed sensation (Condition Static; right arm resting), a baseline condition for suppressed sensation (Condition Point: right arm moving without an additional motor-task), and two different object grasping conditions (Condition Power: power grip; Condition Pinch: pinch grip). We hypothesised that tactile suppression during grasping (Conditions Pinch and Power) would be attenuated compared to baseline (Condition Point), as tactile stimuli to the fingers are relevant for grasping. The additional contrast between the two grasp conditions (Pinch and Power) might reveal how localised such effects of task might be, as tactile inputs to the ring finger are more relevant to performing a power grip than a pinch grip.
2 Methods
2.1 Participants
Ten participants (3 male; age range 20–28 years) volunteered to participate in Exp. 1, and ten participants (1 male; age range 23–26 years) volunteered to participate in Exp. 2. All participants were right-handed and naïve with respect to the purpose and theoretical background of the experiments. They were compensated with a payment of 6 Euro per hour. The Bielefeld University ethics committee approved the study, and participants gave written informed consent prior to their participation.
2.2 Apparatus
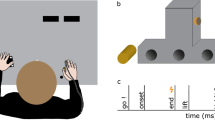
Participants were seated on a height-adjustable chair at a desk with a board (task surface) mounted 15 cm above the desk surface (see Fig. 1). They faced a computer monitor (approximately 60 cm viewing distance) that displayed trial instructions. At the beginning of a trial, participants rested both hands on the surface. Movement onsets were detected using a self-made contact sensor with a sampling frequency of 60 Hz. The task surface was transparent on the right side to allow tracking of the right hand’s movement by a LEAP infrared motion sensor that was located under the task surface. A cylindrical object for grasping stood centrally 30 cm in front of participants’ wrists (see Fig. 1 and Sect. 2.3).

The experimental apparatus as illustrated from a side view (A) and in top view (B).
The vibratory stimuli were applied using the Haptuator Original (Tactile labs). These stimulators were attached to the proximal phalanx of the left and right ring fingers (Exp. 1), or to the right ring finger only (Exp. 2). The timing of stimulus delivery in response to movement onset (see Sect. 2.4) was controlled using the Psychophysics Toolbox for Matlab [12]. Participants’ responses were submitted after a trial using the right hand by pressing a key on a keyboard that was located next to the right hand’s resting position. Participants wore over-the-ear earmuffs to avoid that they might hear activation of the vibrotactile stimulators.
2.3 Motor Tasks
At the beginning of each trial, the computer monitor displayed the instruction to put both hands in the start position (i.e., flat on the task surface). When the contact sensor detected the right hand in the correct position, the task instruction for the motor task was displayed on the monitor. In both experiments, we compared four movement conditions:
-
1.
In the Static condition, both hands were lying flat on the task surface throughout a trial to record participants’ unsuppressed tactile sensitivity (instruction on screen: “Rest”).
-
2.
In the Point condition, the participants made an out-and-back movement to the location of the cylinder with their right hand. There is no relevant tactile input during this pointing movement, so this condition gives a baseline for the participant’s suppressed tactile sensation (instruction on screen: “Point”).
-
3.
In the Pinch grip condition, participants reached and grasped the cylinder with the thumb and index finger of their right hand. In this condition, the stimulated ring finger was not relevant for successful grasping (instruction on screen: “Grasp thumb & index”).
-
4.
In the Power grip condition, participants grasped the cylinder with all fingers of the right hand. Here, the stimulated ring finger was relevant for execution of the grasping task (instruction on screen: “Grasp whole hand”).
From the recorded hand movements we computed for each trial the ring finger velocity in the horizontal plane. We centred these velocity profiles at the time of movement onset, averaged across trials, and defined the velocity during stimulation as the average velocity over the 6 time samples (i.e., 100 ms) at which the stimulation was given. Unfortunately, we had unnoticed technical issues with the tracking system after approximately 250 trials were recorded. We excluded 4 participants (Exp. 1) and 1 participant (Exp. 2) from the velocity analysis because they had fewer than 25 useable movement traces. For the remaining 6 and 9 participants we discarded on average 54.8 % (Exp. 1) and 22.9 % (Exp. 2) of the trials.
2.4 Perceptual Tasks
In both experiments, vibrotactile stimuli that consisted of a 100 ms long 80 Hz pure sine wave vibration were applied to participants’ ring fingers. The stimulus intensity was modulated by varying the sine wave amplitude. In the active conditions (Point, Pinch, Power), the stimuli were applied right after the contact sensor registered a movement onset. In the rest condition (Static), the stimuli were delivered one second after the instruction to rest was displayed.
In Exp. 1 we used an intensity discrimination task. Supra-threshold stimuli were applied simultaneously to the ring fingers of both hands. One hand received a standard stimulus with a fixed intensity, the other hand received a comparison stimulus with one of seven predefined levels of stimulus intensity (Method of Constant Stimuli). When prompted, participants had to judge (forced choice) which of these two stimuli felt more intense by responding with a key press of the right hand. The seven levels of comparison stimulus intensity were spaced equidistant on a logarithmic scale which was centred at the intensity of the standard stimulus. In total there were 7 (intensity pairs) \(\times \) 20 (repetitions) = 140 trials for each of the four movement conditions (see Sect. 2.3), i.e., 560 trials in total. In half the trials, the standard stimulus was applied to the left hand; in the other half, it was applied to the right hand. We fitted a cumulative Gaussian psychometric function with two free parameters (\(\mu \), \(\sigma \)) to the responses using the Matlab function glmfit.m. The 50 % cut gave our estimate of the point of subjective equality (PSE) of left and right hand stimulus intensity.
In Exp. 2 we used a detection task with a single near-threshold stimulus intensity, whose intensity we had determined in pilot experiments. Stimuli were applied only to the right ring finger. When prompted, participants had to judge whether there had been a stimulus or not by pressing a key with the right hand. For each condition, we presented 40 trials in which the stimulus was present and 40 trials in which the stimulus was absent. Thus there were 2 (stimulus present vs. absent) \(\times \) 40 (repetitions) = 80 trials per movement condition, so 320 trials in total. As some participants had either full coverage (no false rejections) or full specificity (no false alarms), we analysed the results using the loglinear approach [9] to estimate \(d^\prime \) from the confusion matrix.
In both experiments, the order of stimulus presentation and movement condition was randomised within a repetition block. Participants had the option to label a trial as invalid if they were inattentive or thought there might have been a technical mistake (e.g., stimulus too early). These trials (Exp. 1: 2.2 %; Exp. 2: 0.75 %) were discarded and repeated at the end of the experiment. At the beginning of an experimental session, participants performed between 5 and 20 practice trials to get familiar with the task. There were no scheduled breaks, but participants were encouraged to take breaks autonomously after every 100 trials. Exp. 1 took approximately 90 min, Exp. 2 approximately 45 min to complete.
3 Results
3.1 Experiment 1
Figure 2(A) depicts the result of a representative participant. Compared to the Static baseline condition (black), the psychometric functions from the three movement conditions (Point, Power, Pinch) are shifted to the right, i.e., the PSE is larger and tactile suppression occurred. In addition, there is a leftward shift in the Static baselines condition psychometric function relative to the standard intensity (vertical striped line), suggesting that stimuli applied to the right hand were perceived as more intense than stimuli applied to the left hand.

Results experiment 1. A: Psychometric curves for an example participant. B: Population results PSE (mean and standard error). C: Population results movement velocity (mean and standard error). D: Tactile suppression (difference in PSE from Static) against mean velocity for individual participants (with least squares regression lines). (Color figure online)
The same result is observed also on the population level (Fig. 2(B)). Paired-sample t-tests confirm that tactile suppression occurred in all movement conditions, i.e., there are significant differences between PSEs in the Static condition and all of the movement conditions (Point: \(t(9)=7.3, p < 0.001\); Pinch: \(t(9)=10.5, p < 0.001\); Power: \(t(9)=16.1, p < 0.001\)). Furthermore there are significant differences between the Point condition and the two grip conditions (Pinch: \(t(9)=3.3, p = 0.002\); Power: \(t(9)=5.4, p < 0.001\)) but not between the two grip conditions (\(t(9)=0.8, p = 0.434\)). The differences between the Point condition and the two grip conditions (Pinch, Power) were in the opposite direction as we had predicted. That is, in the two grip conditions (Power, Pinch) where tactile inputs are deemed important, tactile suppression was stronger compared to the Point condition (Fig. 2(B)). Additionally, the PSE results of the Static condition are significantly smaller than 0 (two-tailed t-test: \(t(9)=23.9, p<0.001\)), which indicates that stimuli applied to the left hand were perceived as less intense than stimuli applied to the right hand. This is an unexpected result that we return to in our Discussion (Sect. 4).
Figure 2(C) shows the velocity of the ring finger at the time of stimulation. Previous studies have shown that the strength of tactile suppression depends on movement velocity [4, 16], so differences in velocity might explain the differences between conditions that we observe. Qualitatively, differences in velocity between conditions follow the same pattern as differences in PSEs, i.e., on average movement velocity is a bit lower in the Point condition than in the Pinch and Power conditions. However, these differences are not significant (Point vs. Pinch \(t(5)=-1.0, p = 0.247\); Point vs. Power \(t(5)=-1.1, p = 0.321\); Power vs. Pinch \(t(5)=-0.5, p = 0.607\)). We assessed whether there was a positive relationship between velocity and tactile suppression effects (i.e., the increase in PSE relative to the static condition), but a correlation analysis did not reveal any significant effects (Absolute value of all Pearson correlation coefficients \(|r|\le 0.6\); Fig. 2 (D)). This might be due to noise in the velocity measurements, as many trials had to be discarded from motion analysis (cf. Sect. 2.3), or to the relatively small range of velocities.
In summary, the results show that perceived intensity was lower in the movement conditions than in the Static condition, which conforms with previous studies on this matter [14, 17]. There is no evidence that task-relevancy attenuates this movement-related tactile suppression, in disagreement with our hypothesis and the results by Colino et al. [3]. On the contrary, tactile suppression was even amplified in the grasping conditions (Pinch, Power) compared to the Point condition. This unexpected effect might be explained by differences in motor planning, muscle activation, or movement velocity between the movement conditions, even if an effect of velocity is not evident in our results.
A possible explanation of why our results differ from those reported by Colino et al. [3] is that we used a different perceptual task. Wrongly assessing the intensity of a supra-threshold stimulus is not the same as detecting whether a stimulus occurred in the first place. We therefore performed a second experiment, where we measured perceptual sensitivity (yes/no detection task) for weaker vibrotactile stimuli.

Results experiment 2. A: Population results \(d^\prime \) (normalised for visualisation by subtracting average individual \(d^\prime \) values; mean and standard error). B: Population results movement velocity (mean and standard error). C: Tactile suppression (negative difference in \(d^\prime \) from Static) against mean velocity for individual participants (with least squares regression lines). (Color figure online)
3.2 Experiment 2
Figure 3(A) depicts the population results of Experiment 2 on tactile sensitivity (\(d^\prime \)). Here, suppression corresponds to a lower \(d^\prime \) in the movement conditions (Point, Power, Pinch) relative to the Static condition. Tactile suppression relative to the Static condition was observed in all movement conditions: the Point condition (\(t(9)=4.8, p = 0.001\)), Pinch condition (\(t(9)=3.7, p = 0.005\)), and the Power condition (\(t(9)=3.4, p = 0.008\)). We found no effect of motor task on tactile suppression, as indicated by t-tests comparing movement conditions: Point vs. Pinch: \(t(9)=0.3, p = 0.751\); Point vs. Power: \(t(9)=-0.1, p = 0.898\); Pinch vs. Power: \(t(9)=-0.5, p = 0.646\)).
In Exp. 2, there was a significantly lower movement velocity in the Point condition than in the Power condition (\(t(8)=-2.4, p = 0.041\)), but no differences between the other conditions (Point vs. Pinch \(t(8)=-1.3, p = 0.234\); Power vs. Pinch \(t(8)=-1.4, p = 0.213\)). We again performed a regression analysis to investigate whether the strength of tactile suppression between participants might be explained by differences in velocity, in which case we would expect a positive correlation between velocity and the decrease in \(d^\prime \) relative to Static condition (Fig. 3(C)). However, we found no significant relationship in any of the three movement conditions (all \(|r|\le 0.52\)).
4 Discussion
In this study we examined whether tactile suppression might be modulated by a ‘clever’, centrally controlled mechanism that knows whether tactile sensory inputs at a specific location are relevant to behavioural success in a given task. This presumed mechanism would process relevant stimuli at the regular, unsuppressed intensity but would attenuate the perceived intensity of task-irrelevant stimuli. Such task-dependent processing of tactile stimuli has been reported in other contexts [1] and preliminary results in the literature [3] suggested that also tactile suppression might be modulated in such a way. However, the results here do not indicate the existence of such mechanism. We tested participants’ intensity discrimination performance (Exp. 1) and detection performance (Exp. 2) for vibrotactile stimuli comparing two grasping conditions (Pinch, Power), where tactile inputs to the finger were presumed relevant to avoid slip, with a pointing-in-the-air condition (Point), where tactile inputs are irrelevant. The results show no effect of task on detection (Sect. 3.2) and even the opposite effect of task than hypothesised on intensity discrimination (Sect. 3.1), i.e., more suppression for the grasping conditions. We therefore conclude that movement related tactile suppression is likely not modulated in a top-down manner by task-relevancy.
Of course, it is possible that such a modulation does indeed exist and that our experimental design was unsuitable to detect it. For instance, we investigated suppression early on during the outward hand movement and in one skin location. Instead, the task-relevancy in tactile suppression might be specific in both space and time. As for the spatial aspect, it might be that only a highly specific part of the moving limb is considered task-relevant and that in our grasping task the proximal part of the finger was not included. Similarly, the task-relevancy effect might occur later during the movement, close to the time of grasping contact. It is also possible that motor-task relevancy alone is not sufficient to cause attenuation of movement-related tactile suppression. A study by Juravle et al. [11] suggests that there might be an interaction between motor tasks and the ecological validity of the corresponding perceptual task.
However, given the influence of movement velocity on tactile suppression [4, 16], one has to also bear a different explanation in mind for why differences between tasks were observed in some studies but not in ours. Different motor tasks can lead to differences in muscle activation and movement velocity, which then might modulate the strength of tactile suppression effects recorded. In our design, we picked actions that were very similar (i.e., grasping and pointing) and we chose to compare very early stages of movement (i.e., movement onset) where no larger velocity differences were expected. Nevertheless, even in our experiments there is a tendency for slower movements in the Point condition than in the Grasp conditions (Figs. 2C and 3B). This finding illustrates how hard it is to obtain equivalent experimental conditions when comparing tactile suppression between movement conditions. It is therefore essential to control for such low-level confounds when assessing the effect of task-relevancy (or other task-related factors) on tactile suppression.
An additional unexpected result is the difference in perceived tactile intensity between the right and left hands (Static condition in Exp. 1, Sect. 3.1 and in Fig. 2B). We included only right-handed participants, so this unexpected laterality effect might have been caused by hand dominance. Alternatively, there might have been attentional effects on perceived tactile intensity, as the right hand was active in 75 % of the trials, whereas the left hands was always passive. This might have shifted attention to the right hand. Irrespective of the nature of this laterality effect, it is also a possible confound in experiments that compare perceived intensity between an active and a passive hand (e.g., [3]).
In summary, while we cannot rule out that the motor task modulates tactile suppression, our results suggest that such an influence is minor. Instead, we found that tactile suppression is modulated more by low-level factors, such as laterality and possibly also movement velocity, which have to be controlled when performing this kind of experiment.
References
Brozzoli, C., Pavani, F., Urquizar, C., Cardinali, L., Farnè, A.: Grasping actions remap peripersonal space. NeuroReport 20(3), 913–917 (2009)
Chapman, C.E., Bushnell, M.C., Miron, D., Duncan, G.H., Lund, J.P.: Sensory perception during movement in man. Exp. Brain Res. 68(3), 516–524 (1987)
Colino, F.L., Buckingham, G., Cheng, D.T., van Donkelaar, P., Binsted, G.: Tactile gating in a reaching and grasping task. Physiol. Rep. 2(3), e00267 (2014)
Cybulska-Klosowicz, A., Meftah, E.M., Raby, M., Lemieux, M.L., Chapman, C.E.: A critical speed for gating of tactile detection during voluntary movement. Exp. Brain Res. 210(2), 291–301 (2011)
Debats, N.B., Kingma, I., Beek, P.J., Smeets, J.B.J.: Moving the weber fraction: the perceptual precision for moment of inertia increases with exploration force. PLoS ONE 7(9), e42941 (2012)
Drewing, K.: After experience with the task humans actively optimize shape discrimination in touch by utilizing effects of exploratory movement direction. Acta Psychol. 141(3), 295–303 (2012)
Drewing, K., Kaim, L.: Haptic shape perception from force and position signals varies with exploratory movement direction and the exploring finger. Attention Percept. Psychophys. 71(5), 1174–1184 (2009)
Garland, H.T., Angel, R.W.: Modulation of tactile sensitivity during movement. Neurology 24(4), 361 (1974)
Hautus, M.J.: Corrections for extreme proportions and their biasing effects on estimated values of \(d^\prime \). Behav. Res. Methods Instrum. Comput. 27(1), 46–51 (1995)
Juravle, G., Deubel, H., Tan, H.Z., Spence, C.: Changes in tactile sensitivity over the time-course of a goal-directed movement. Behav. Brain Res. 208(2), 391–401 (2010)
Juravle, G., McGlone, F., Spence, C.: Context-dependent changes in tactile perception during movement execution. Front. Psychol. 4(913) (2013)
Kleiner, M., Brainard, D., Pelli, D.: What’s new in psychtoolbox-3? In: Abstract Supplement of the 30th European Conference on Visual Perception (ECVP), vol. 36, p. 14 (2007)
Lederman, S.J., Klatzky, R.L.: Hand movements: a window into haptic object recognition. Cogn. Psychol. 19(3), 342–368 (1987)
Milne, R.J., Aniss, A.M., Kay, N.E., Gandevia, S.C.: Reduction in perceived intensity of cutaneous stimuli during movement: a quantitative study. Exp. Brain Res. 70(3), 569–576 (1988)
Plaisier, M.A., van Dam, L.C.J., Glowania, C., Ernst, M.O.: Exploration mode affects visuohaptic integration of surface orientation. J. Vis. 14(13), 22 (2014)
Schmidt, R.F., Torebjörk, H.E., Schady, W.J.L.: Gating of tactile input from the hand. Exp. Brain Res. 79(1), 103–108 (1990)
Williams, S.R., Chapman, C.E.: Time course and magnitude of movement-related gating of tactile detection in humans. II. Effects of stimulus intensity. J. Neurophysiol. 84(2), 863–875 (2000)
Acknowledgments
NBD was supported by the German Research Foundation (DFG) grant HE 1187/19-1. MR, CG and MOE were supported by the Cluster of Excellence Cognitive Interaction Technology ‘CITEC’ (EXC 277), which is funded by the DFG. MOE was additionally supported by the FP7/2007-2013 project n8 601165 WEARHAP. We would like to thank Irene Senna for fruitful discussions and Miriam Henning for her work on pilot experiments.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this paper
Cite this paper
Debats, N.B., Rohde, M., Glowania, C., Oppenborn, A., Ernst, M.O. (2016). The Influence of Motor Task on Tactile Suppression During Action. In: Bello, F., Kajimoto, H., Visell, Y. (eds) Haptics: Perception, Devices, Control, and Applications. EuroHaptics 2016. Lecture Notes in Computer Science(), vol 9774. Springer, Cham. https://doi.org/10.1007/978-3-319-42321-0_15
Download citation
DOI: https://doi.org/10.1007/978-3-319-42321-0_15
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-42320-3
Online ISBN: 978-3-319-42321-0
eBook Packages: Computer ScienceComputer Science (R0)