
Abstract
Uruguay has several marine deposits of undoubtedly Late Pleistocene age, but there is conflicting evidence when comparing ages obtained by different methods. While 14C datings suggest younger ages (related to Marine Isotope Stage 3—MIS 3), OSL, where available, indicate older times (related to Marine Isotope Stage 5—MIS 5). The analysis of the abundant molluscan fauna and the presence of extralimital warm water taxa points to a higher than present water temperature for the Uruguayan coast. The referred discrepancies are discussed and a MIS 5 age is preferred according to all available evidence.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Marine Isotope Stage 3 (MIS 3)
- Marine Isotope Stage 5 (MIS 5)
- Late Pleistocene
- Molluscs
- Bivalves
- Gastropods
- Paleoecology
- Paleobiogeography
- Extralimital species
- Uruguay
1 Introduction
The Pleistocene Epoch was a time of global climatic and sea level changes that had a profound impact in the marine and continental realms, influencing the configuration of our present-day biota. The information for the reconstruction of these oscillations comes from different sources that include ice cores and a wide spectrum of continental and marine records. It is widely recognized the existence throughout the Pleistocene of numerous glacial–interglacial cycles characterized by cold (stadials) and warm (interestadials) lapses (e.g., Emiliani 1955; Shackleton 1969; Crowley and North 1991; Winograd et al. 1997). In recent years and especially for the Late Pleistocene, a great improvement in the knowledge of the timing and magnitude of these climatic changes in both hemispheres comes from ice cores from Greenland and Antarctica (e.g., Petit et al. 1999; North Greenland Ice Core Project Members 2004; EPICA Community Members 2004, 2006; Jouzel et al. 2007; Orombelli et al. 2010). These high resolution sources of information allowed the recognition of the pronounced and abrupt Dansgaard–Oeschger (D–O) warming events in Greenland and the more gradual Antarctic Isotope Maxima (AIM) in Antarctica (EPICA Community Members 2006; Huber et al. 2006) superimposed to the traditional isotopic stages. These millenial-scale changes were also recognized in other records (e.g., Barker et al. 2009).
The Marine Isotope Stage 5 (MIS 5) had an average duration between 130 and 71 ka B.P. and was a lapse of warm conditions with the warmest peak considered to be the Last Interglacial or MIS 5e (Zubakov and Borzenkova 1990; Winograd et al. 1997; Petit et al. 1999; Shackleton et al. 2003; Jouzel et al. 2007). The Last Interstadial or Marine Isotopic Stage 3 (MIS 3) had an average duration between 60 and 27 ka B.P. and was characterized by overall lower temperatures than present and than MIS 5 and by the record of numerous D–O events (Huber et al. 2006; Van Meerbeeck et al. 2009, 2011; Buiron et al. 2012; Long and Stoy 2013).
One of the sources of information that have contributed to the reconstruction of the environmental changes occurred in our near past come from littoral fossiliferous assemblages around the world. Pleistocene marine assemblages have been studied from the Pacific coast of North America (e.g., Valentine and Jablonski 1993; Roy et al. 1995; Powell et al. 2004), Mexico (De Diego-Forbis et al. 2004), Atlantic islands at different latitudes (Muhs et al. 2002; Ávila et al. 2009), Pacific Islands (Muhs et al. 2002), Japan (Amano 1994; Kitamura and Ubukata 2003), Australasia (Murray-Wallace and Belperio 1991; Murray-Wallace et al. 2000; Murray-Wallace 2002), Antarctica (Berkman et al. 1998), and Europe (Zazo et al. 2003; Nielsen et al. 2006; Garilli 2011). In South America, Pleistocene marine deposits are known from the Pacific coast (e.g., Ortlieb et al. 1990; Rivadeneira and Carmona 2008), Beagle Channel (Rabassa et al. 2009) and Atlantic coast, mostly from Argentina (see Aguirre and Whatley 1995; Isla et al. 2000; Aguirre 2003; Aguirre et al. 2011; Gordillo and Isla 2011; Charó et al. 2013, 2014) and in a lesser extent from Brazil (e.g., Lopes and Simone 2012; Lopes et al. 2013).
In Uruguay, undoubtedly Pleistocene fossil assemblages were first recognized by Martínez et al. (2001) in the Puerto de Nueva Palmira (western Uruguay, Colonia County) and La Coronilla (eastern Uruguay, Rocha County). Subsequently, Rojas (2007) provided a new analysis of the molluscan content of these two assemblages and included another one located at Zagarzazú (Colonia County). More recently, Martínez et al. (2013) provided a biogeographic analysis of Pleistocene and Holocene molluscan faunas of the Southwestern Atlantic, including Brazilian, Argentinean, and Uruguayan data.
After the first characterization of the Pleistocene molluscan assemblages from Uruguay provided by Martínez et al. (2001), new data on the faunal composition, biogeographic inferences, and geochronological context have become available. Thus, the aims of this contribution are (a) to update the paleontological content of the Late Pleistocene fossil assemblages from Uruguay , (b) to provide a paleoecologic and biogeographic analysis of the faunal elements that are useful for paleoenvironmental reconstruction, and (c) to discuss these results in the light of the MIS 3 versus MIS 5 temporal and climatic scenario.
2 Geographical Setting
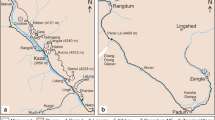
The Late Pleistocene fossiliferous deposits found so far in Uruguay are three, two of them located in the western coast, and the remaining in the eastern coast (Fig. 1). The Puerto de Nueva Palmira outcrop can nowadays be found in the area occupied by the port of the city of Nueva Palmira. About 12 km to the south the Zagarzazú site crops out in the abrasion platform of the beach. Finally, the La Coronilla fossil assemblage crops out in the abrasion platform of the homonymous beach in the Rocha County (eastern Uruguay). The Zagarzazú and La Coronilla deposits are frequently covered by sand.

Geographic location of the Late Pleistocene fossil assemblages of Puerto de Nueva Palmira, Zagarzazú, and La Coronilla
The study area comprises the Uruguayan coastal waters currently dominated by the fluviomarine gradient of the Río de la Plata and Atlantic Ocean. The Río de la Plata estuary between Argentina and Uruguay receives from the west, freshwater and sediment discharge coming from the Paraná and Uruguay rivers and from the east, the marine waters of the Atlantic Ocean (Urien 1972). It is divided by a submerged shoal (Barra del Indio) into an inner fluvial system and an outer mixohaline brackish system (Mianzan et al. 2001). The biogeography of the living biota responds to this gradient with a west to east dominance of freshwater, estuarine, and marine taxa, respectively (Masello and Menafra 1998; Giberto et al. 2004; Giménez et al. 2005; Brazeiro et al. 2006). The Río de la Plata estuary is a highly variable environment at different timescales and the boundaries and position of the salinity zones vary according to wind balance, river discharge, season, and phenomena such as El Niño Southern Oscillation (ENSO) (e.g., Guerrero et al. 1997; Acha et al. 2008; Möller et al. 2008; Nagy et al. 2008).
The large-scale dynamics of the Southwestern Atlantic Ocean are dominated by the northward flowing Malvinas (Falkland) Current and the southward flowing Brazilian Current. The former transports cold subantarctic waters along the Argentinean shelf whereas the latter carries tropical and subtropical waters along the continental margin of South America (Olson et al. 1988; Piola et al. 2000; Odebrecht and Castello 2001). Both currents meet approximately between 25° S and 45° S at the Brazil–Malvinas Confluence Zone, producing a complex oceanographic area with highly variable physicochemical and biological attributes on the shelf and slope. The temperature gradient developed by these water masses outline the current malacological provinces of the Southwestern Atlantic: the warm water Brazilian Province, the confluence Argentinean Province, and the cold Magallanic Province (e.g., Scarabino 1977; Briggs 1995; Martínez and del Río 2002; Martínez et al. 2013). Studies in Brazil , Argentina, and Uruguay showed that the faunal composition of the Argentinean Province includes a combination of species of warm-temperate and cool-temperate affinities, besides endemic ones. In this scenario, the Río de la Plata and its freshwater discharge acts as an ecological barrier and represents a broad ecotone between the southern and northern areas (Masello and Menafra 1998; Scarabino et al. 2006a, b).
3 Geological Setting
The Quaternary marine deposits of Uruguay have their origin in the transgressive–regressive events characteristic of this period of global-scale climatic oscillations. The fossiliferous deposits have been correlated with adjacent Atlantic units from Argentina and Brazil by various authors (Goñi and Hoffstetter 1964; Forti-Esteves 1974; Martínez 1990; Aguirre and Whatley 1995).
From the lithostratigraphic viewpoint they are included in the Chuy and/or Villa Soriano formations (Goñi and Hoffstetter 1964; Goso 1972). The Villa Soriano Formation represents the fossiliferous deposits that Caorsi and Goñi (1958) named “Arcillas grises del Vizcaíno” which were later formalized as a lithostratigraphic unit by Goso (1972). These deposits crop out along a narrow stripe parallel to the present coastline of Uruguay , from the Río Negro to the Merín Lagoon margins. Lithologically, this unit has wide grain-size variability, from clay to medium sands and occasionally gravel and pebbles (Preciozzi et al. 1988). Most authors consider as a characteristic of this unit the abundant fossiliferous content (e.g., Serra 1943; Caorsi and Goñi 1958); thus, in a sort of circular reasoning, most fossils have been attributed to this unit. The age of the Villa Soriano Formation has been a matter of discussion. Bossi (1966) and Preciozzi et al. (1988) considered it as of Late Pleistocene–Holocene age, meanwhile Figueiras (1962), Bossi et al. (1975), and Sprechmann (1978) considered it only of Holocene age. Martínez et al. (2006) assigned to the Villa Soriano Formation the fossiliferous deposits radiocarbon dated as Holocene. This unit has been correlated with the “Querandinense” deposits from Argentina (Goñi and Hoffstetter 1964; Aguirre and Whatley 1995) and with the Patos Group in Rio Grande do Sul State, Brazil (Martínez 1988).
The Chuy Formation was defined by Delaney (1963), although the first published mention corresponds to Goñi and Hoffstetter (1964). This unit was originally described as composed by yellow reddish medium sands. Later on, other lithological types, not included in its definition, were added to its characterization. According to Navarro (1990), the Chuy Formation extends from the Río Santa Lucía basin to the east of Uruguay. Its scarce fossil remains were reported from wells by Sprechmann (1978). The stratigraphic relationships of this unit indicate that it belongs to the Pleistocene (Goñi and Hoffstetter 1964; Goso 1972; Sprechmann 1978).
The poor definition of these lithostratigraphic units, their wide and overlapping lithological characterization, and the use of nonlithological criteria to recognize them, hinders their identification in the field and consequently the placement of fossil remains (see Martínez and Ubilla 2004; Martínez et al. 2013, for further details). Thus, the assignment of the Late Pleistocene fossiliferous deposits studied here to the Villa Soriano Formation or the Chuy Formation is problematic and not useful for practical purposes at the moment.
4 Description of the Outcrops and Previous Work
4.1 Puerto de Nueva Palmira
The fossiliferous deposit is located at around 12 m of altitude and is approximately 2 m thick (Figs. 2 and 3a–c). The lithology is medium to coarse sand with embedded clasts which maximum diameter can reach approximately 4 cm. Shells are mostly randomly distributed and densely packed. Bivalves are frequently disarticulated, although specimens with articulated valves rarely occur. Abrasion and fragmentation are very common, revealing some degree of local transport. The assemblage likely represents the accumulation of shells in multiple high energy events in a proximal environment influenced by waves (Martínez et al. 2001; Rojas 2007).

Stratigraphic sections of the studied deposits

Photographs of the fossiliferous Pleistocene outcrops. a–c Puerto de Nueva Palmira; d–f Zagarzazú; g–i La Coronilla. Images (f) and (i) show Tagelus plebeius in life position
The Puerto de Nueva Palmira fossil assemblage is one of the most mentioned in the literature concerning the Quaternary marine deposits from Uruguay. Since the first reports about this deposit, some physical changes in the area have occurred. For example, the port was built with the consequent modification of the original landscape. Teisseire (1927, 1928) reported the molluscan fauna of a deposit which he refers as located on the cliff of the cemetery of Nueva Palmira (formerly nearby the port) with an altitude of 7–10 m. This author mentions the presence of Anomalocardia brasiliana, Bullia cochlidium, Cardium muricatum, Mactra isabelleana, Pitar rostratum, Thais haemastoma, Bullia deformis, and Fissuridea patagonica. The present authors believe that this deposit studied by Teisseire and subsequent authors may be equivalent to the deposit referred here, because Teisseire (1927), Kraglievich (1928), and Fontana (1930) mentioned that in the cliff of the cemetery a new port (a Free Trade Zone) was being constructed at that time. The last author corrected the altitude assigned by Teisseire (1928) to 15 m and reported the presence, at approximately 6 m a.s.l., of varied fossil shells with gastropods that needed to be studied. He also commented (as Kraglievich 1928: Fig. 6) about the abundant representation of Corbula mactroide (sic). Both authors based on this bivalve, correlated this deposit with the “Querandinense” units. After that, Frenguelli (1930) referred to the deposit of the Puerto de Nueva Palmira, which he assigned to the “Belgranense” stage (Pleistocene) based upon its nature, position, aspect, and paleontological content. This author reported the presence of Littoridina charruana, Bulla striata, Bittium varium, Purpura haemastoma, Bullia globulosa, Ostrea spreta, Ostrea puelchana, A. brasiliana, and M. isabelleana. Regarding the presence of Corbula mactroides in this deposit, Frenguelli (1930) suggested that Kraglievich (1928) had misidentified this species with M. isabelleana which is very abundant there. But Kraglievich (1932) reaffirmed that the species found was Corbula mactroides. Later on, Roselli (1939) mentioned this deposit and commented on the diverse opinions regarding its assignment to the “Belgranense” or “Querandinense” episodes. Serra (1943) presented a stratigraphic section of the outcrop and made comments about the most abundant fossils, M. isabelleana and Purpura haemastoma. Calcaterra (1971) expressed doubts on the proposed age for this deposit based upon its elevation (more than 12 m a.s.l.) and the thickness of the overlying sediments. He also listed the presence of M. isabelleana, T. haemastoma, Arca sp., Crassostrea sp., and Buccinanops sp. Francis (1975) referred to the coastal cliff of Nueva Palmira and, besides listing some molluscan species, he made reference to the historical controversy between Kraglievich and Frenguelli on the presence or absence of Erodona mactroides in this outcrop, which he was not able to find. He was surprised about the elevation of the deposit in relation to others typically attributed to the “Querandinense”. Roselli (1976) thoroughly described the deposit and stuck to the interpretation proposed by Frenguelli (1930) concerning the Late Pleistocene age of the deposit. This author provided a list of taxa that includes Mactra (Mactratoma) isabelleana, A. brasiliana, Thais (Stramonita) haemastoma, Buccinanops globulosus, Buccinanops cochlidium, Buccinanops deformis, Plicatula spondyloidea, Ostrea spreta, Arca bisulcata, Ostrea puelchana, Tagelus (Tagelus) gibbus, Diodora patagonica, Urosalpinx rushi, Littoridina australis, Nassarium sp., Brachidontes dominguensis, Trachycardium muricatum, Balanus sp., Pitar (Pitar) rostrata, Clamys sp., Glycymeris longior, Siphonaria (Pachysiphonaria) lessoni, Crepidula dilatada ¿var. patagonica?, Calliostoma sp., Erodona mactroides, Mactra sp., ¿Olivancillaria?, ¿Chione?, Drupa (Drupa) necocheana, and two terrestrial gastropods . Afterward, the paleontology , geochronology , and paleoenvironmental conditions of the locality were studied by Martínez et al. (2001). Supplementary information may be found in Rojas (2007).
4.2 Zagarzazú
This fossiliferous deposit is located in the abrasion platform of the Zagarzazú resort approximately 0.5–1 m above mean sea level (a.m.s.l.; Figs. 2 and 3d–f). Its exposure depends on the littoral dynamics, and sometimes it may be covered by sand or water. It is composed of fine sands at the base and green clays at the top. Fossil shells are present in both levels. The fossil preservation is variable, but mostly good. Bivalve shells in life position are frequent (Fig. 3f) and complete, whole specimens with delicate shells can be found, although disarticulated and broken remains do occur as well. Few shells showing abrasion were found. The depositional environment is inferred as proximal, under low energy conditions (Rojas 2007).
The scientific knowledge of the Zagarzazú assemblage is very recent. Verde (2003) and Rojas (2003) presented preliminary reports about the ichnofossils and molluscan content. Rojas (2007) did a more extensive analysis of the molluscan fauna, paleoenvironmental conditions, and available radiometric dating.
4.3 La Coronilla
This deposit is located at the abrasion platform of the La Coronilla beach, 0.5–1 m a.s.l. (Figs. 2 and 3g–i). Similarly to the Zagarzazú locality, its exposure depends on the littoral dynamics as it is frequently covered by sand or water. It is lithologically composed of a greenish-grayish sandy claystone in which fossils are well preserved. Taxa with delicate shells occur. It is frequent the preservation of shells in life position (Fig. 3i). Fragmentation of the shells is common but not abrasion. These features and the lack of shell size sorting suggest a predominantly autochthonous–parautochthonous assemblage without significant transport (Rojas 2007; Rojas et al. 2014). Shell fragmentation can be related to the activity of biological agents. The depositional environment is inferred as proximal and mostly protected from wave action.
Previous works regarding the La Coronilla fossil fauna can be found in Martínez et al. (2001), who studied the paleontology , geochronology, and paleoenvironments. Additional information can be found in Rojas (2007). Lorenzo and Verde (2004) and Rojas et al. (2014) presented ichnological information, and Rojas and Urteaga (2011) dealt with the record of chitons in this assemblage.
5 Absolute Ages
Bracco and Ures (1998), Martínez et al. (2001), Rojas (2007), and Rojas (2010) published absolute dates for the Uruguayan Late Pleistocene assemblages, based upon conventional 14C and OSL dating. These data are compiled in Table 1.
5.1 14C Conventional
Bracco and Ures (1998) provided 14C conventional ages for the Nueva Palmira site without locating the samples in a stratigraphic column or indicating the beds from which they were collected; albeit they mentioned an elevation of 8–10 m a.s.l. They mentioned two radiocarbon ages of 31,900 ± 700 years B.P. (URU 0081) obtained from nonidentified shells and another age of 35,300 ± 1,150 years B.P. (URU 0087) from carbonate clasts included in the deposit. Martínez et al. (2001) provided 14C conventional ages obtained from infaunal bivalves of 31,000 ± 1,200 years B.P. (LP-738) on M. isabelleana and 34,600 ± 2,000 years B.P. (LP-730) on A. brasiliana. The X-ray diffraction analyses applied to evaluate the consistency of the samples gave reliable results for these ages.
For the La Coronilla deposit, Martínez et al. (2001) obtained ages of 29,500 ± 600 years B.P. (LP-884) on Ostrea equestris and 35,500 ± 1,900 years B.P. on M. isabelleana.
For Zagarzazú, Rojas (2007) obtained a 14C conventional age of 35,500 ± 1,900 years B.P. (LP-1466) on the infaunal bivalve Tagelus plebeius. The specimens used for dating were collected in life position.
The datings reported by Martínez et al. (2001) and Rojas (2007) were performed at the Laboratorio de Tritio y Carbono of the Universidad de La Plata (LATYR). All these ages were considered as minimum ages; thus, fossil assemblages are thought to be older than their measured radiocarbon age (Martínez et al. 2001; Rojas 2007).
5.2 OSL Ages
In order to extend the temporal range of numerical dating and to obtain an independent source of ages for the Pleistocene deposits, the present authors have applied the Optically Stimulated Luminescense (OSL) dating technique to sandy fossiliferous deposits. As suitable deposits for this method, the Puerto de Nueva Palmira and Zargarzazú beds were selected because of their sandy lithology. Although OSL dating has been better developed in continental environments such as aeolian sediments, this dating technique has also been used for littoral marine deposits (e.g., Mallinson et al. 2008; Simms et al. 2009; Suguio et al. 2011). From a methodological point of view, sampling was made using PVC tubes wrapped with thick black tape in order to prevent light reaching the sample. Tubes were dug horizontally into the profile and samples were then extracted. The extremes of each tube were discarded in the laboratory to use only the central part of the sample. For each sample, ages were obtained on quartz grains of 150–250 μm in the Luminescence Dating Research Laboratory, Department of Earth and Environmental Sciences of the University of Illinois at Chicago. Results were provided by Rojas (2010). Ages of 80,680 ± 5,500 years B.P. and 88,355 ± 7,070 years B.P. were obtained for the Puerto de Nueva Palmira and Zagarzazú localities, respectively. Additional data of the OSL ages can be found in Appendix.
6 Fossil Composition of the Late Pleistocene Assemblages
The Late Pleistocene fossil assemblages from Uruguay preserve a diverse and abundant bivalve and gastropod fauna (Martínez et al. 2001; Rojas 2007). In Tables 2 and 3 the present authors forwarded an update of these molluscan taxa based upon the works of Martínez et al. (2001) and Rojas (2007), due to their explicit description of sampling method employed and the availability of the specimens collected.
Other molluscs , such as polyplacophorans, have been recorded only for the La Coronilla site (Rojas and Urteaga 2011). The chiton species found in this deposit are Chaetopleura angulata, C. isabellei, C. asperrima, and Ischnochiton striolatus.
All specimens referenced in Martínez et al. (2001), Rojas (2007), and Rojas and Urteaga (2011) are housed in the Colección Paleontológica at the Facultad de Ciencias, Universidad de la República (FCDPI), Montevideo. Some of the specimens listed by Martínez et al. (2001) and Rojas (2007) were revised in order to improve their taxonomic assignment. For instance, specimens originally assigned to Chrysallida gemmulosa are reclassified herein as Fargoa bushiana. Specimens referenced as Turbonilla americana by Martínez et al. (2001) are reclassified as Turbonilla multicostata and those classified as Heleobia australis are considered as Heleobia sp., due to the difficult identification of the different species of this genus using only their shells. Specimens assigned by Martínez et al. (2001) to Clausinella gayi are now thought to be another yet unidentified venerid. Although the species Ennucula uruguayensis has been synonymized with E. puelcha, the present authors maintained it as a valid species, based on the observations reported by Scarabino et al. (2006b). Crepidula sp., listed by Martínez et al. (2001) from Puerto de Nueva Palmira is herein identified as Bostrycapulus odites. Parvanachis isabellei from Martínez et al. (2001) and Parvanachis paessleri from Rojas (2007) are presented here as Parvanachis spp. because the assignment of these specimens to a particular species of Parvanachis deserves caution and further comparisons.
In addition, occasional remains of other non-molluscan taxa such as balanids, decapods, ostracods, bryozoans, serpulids, corals, echinoid, and fishes have been found (Rojas 2007). Bioerosion traces on shells reported by Verde (2003) for the Zagarzazú locality include Entobia isp., Gastrochaenolites isp., and Meandropolydora isp. Lorenzo and Verde (2004) found Entobia isp., Caulostrepsis taenicola, and Oichnus paraboloides on molluscan shells from Puerto de Nueva Palmira and La Coronilla localities. Recently, Rojas et al. (2014) reported the first gastropod drill hole on a fossil chiton plate. This corresponds to the ichnospecies Oichnus simplex placed on an intermediate valve of C. angulata found in the La Coronilla site. Verde (2003) recorded ichnofossils in soft sediments from the Zagarzazú locality. These structures correspond to Ophiomorpha nodosa and Thalassinoides isp., along with other yet unidentified bivalve ichnofossils.
7 Analysis of the Molluscan Fauna
A total of 85 molluscan taxa (36 gastropods , 45 bivalves, and 4 polyplacophorans) have been recorded so far in the Late Pleistocene fossil assemblages of Uruguay . Ongoing research suggests that the molluscan diversity is even higher. La Coronilla locality has the richest assemblage, recording 70 molluscan taxa. It is followed by the Puerto de Nueva Palmira and Zagarzazú deposits that record 32 taxa and 27 taxa, respectively.
According to the available information (Clavijo et al. 2005; Martínez et al. 2006; Rojas 2007; Rojas and Urteaga 2011) the gastropods Bittiolum varium, Melanella sp., F. bushiana, Iselica anomala, Littoraria flava, Lucapinella henseli, T. multicostata and the Vitrinellidae specimens, the bivalves Anatina anatina, Atrina seminuda, Cardiomya sp., Cyclinella tenuis, E. uruguayensis, Ervilia concentrica, Gastrochaena sp., Gouldia cerina, Laevicardium sp., Limaria sp., Macoma constricta, Musculus sp., Pitar palmeri, and Merisca martinicensis have been exclusively recorded in Pleistocene deposits and are absent from the Holocene molluscan assemblages.
All recorded molluscs are extant and live along the western coast of the Atlantic Ocean. Only Siphonaria lessoni is a cold water species influenced by the Malvinas Current (Scarabino 1977) which represents approximately 1 % of the taxa recorded in the Pleistocene assemblages. Meanwhile, in the recent Uruguayan coast approximately 19 % of the molluscan fauna are considered to be cold water species (Sicardi 1967; Sprechmann 1978).
The molluscs found in the Late Pleistocene assemblages also record 17 taxa that show a retraction in their recent biogeographic range (Figs. 4, 5, and 6). The gastropods B. varium, Bulla occidentalis, F. bushiana, and L. flava, the bivalves A. anatina, A. brasiliana, C. tenuis, E. concentrica, G. cerina, Laevicardium sp., Limaria sp., M. constricta, Chione subrostrata, P. palmeri, Scapharca brasiliana, and M. martinicensis, and the chiton I. striolatus have their southernmost boundary of distribution in Brazilian waters. Most of these taxa were recorded exclusively at the La Coronilla assemblage. From the precedent group, only A. brasiliana has been recorded in all Pleistocene deposits studied. G. cerina, N. subrostrata, and B. varium were recorded both at La Coronilla and Puerto de Nueva Palmira localities. For the moment, B. occidentalis has been only recorded at Puerto de Nueva Palmira, L. flava only in Zagarzazú and M. constricta is shared by Zagarzazú and Puerto de Nueva Palmira localities.

Extralimital warm water gastropods from the Pleistocene fossil assemblages from Uruguay. a Littoraria flava (FCDPI 3948); b Bittiolum varium (FCDPI 6661); c Fargoa bushiana (FCDPI 4274); d Bulla occidentalis (FCDPI 3060). Scale bar a–c 1 mm; d 5 mm

Extralimital warm water bivalves from the Pleistocene fossil assemblages from Uruguay. a Scapharca brasiliana (FCDPI 4202); b Ervilia concentrica (FCDPI 4217); c Macoma constricta (FCDPI 4145); d Limaria sp. (FCDPI 2881); e Laevicardium sp. (FCDPI 4204); f Pitar palmeri (FCDPI 4222); Scale bar: a, c, e, f 5 mm; b, d 1 mm

Extralimital warm water bivalves from the Pleistocene assemblages from Uruguay. a Anatina anatina (FCDPI 4192); b Gouldia cerina (FCDPI 4220); c Chione subrostrata (FCDPI 4206); d Anomalocardia brasiliana (FCDPI 4205); e Cyclinella tenuis (FCDPI 4221); f Merisca martinicensis (FCDPI 4717);. Scale bar: a 10 mm; b 1 mm; c–f 5 mm

Current southernmost limit of geographic distribution of the extralimital warm water molluscs found in the Late Pleistocene assemblages from Uruguay. RG Rio Grande do Sul; SC Santa Catarina; P Parana; SP São Paulo. Biogeographic data were taken from Ríos (1994, 2009), Scarabino and Zaffaroni (2004), Amaral et al. (2006), Pimenta et al. (2009), Rosenberg (2009)
8 Discussion
Several lines of evidence must be considered in order to assign the Uruguayan Late Pleistocene fossil assemblages either to MIS 3 or MIS 5. These lines are: (a) the numerical ages obtained for the deposits, (b) sea level and deposit elevation above mean sea level, (c) the environmental conditions inferred from the identified fossils.
(a) Numerical ages
The standard radiocarbon ages obtained for the three Pleistocene fossil assemblages fall within the scope of MIS 3. These ages were considered by Martínez et al. (2001) and Rojas (2007) as minimum ages. Due to the warmer water affinities of the molluscan fauna, these authors considered that the real age of the assemblages would fall within MIS 5e. In relation to the ages reported by Martínez et al. (2001), Tonni et al. (2010) argued that the ages reported for the Puerto de Nueva Palmira and the La Coronilla deposits were finite ages and statistically distinguishable from the dating limit of the radiocarbon dating method. Thus, Tonni et al. (2010) based their opinion only on the numerical ages, but not taking into account the paleoecological information, and considered that the Uruguayan Pleistocene deposits are representative of the final period of MIS 3.
However, the OSL ages recently obtained for the Puerto de Nueva Palmira and Zagarzazú localities are significantly older compared to those obtained by 14C, falling within MIS 5a. The OSL ages, obtained from coastal deposits elsewhere, have shown to be an accurate dating method (e.g., Mallinson et al. 2008; Simms et al. 2009; Suguio et al. 2011) that indicates the time lapse occurred since the last exposure of quartz sand to sunlight. In fact, for instance, Simms et al. (2009) assigned on the basis of OSL dating to MIS 5a deposits from the northwestern Gulf of Mexico which had previously radiocarbon dated as of MIS 3.
(b) Sea level and deposits elevation
During the Late Pleistocene , sea level above or at its present position has been widely recognized for MIS 5e up to approximately 9 m a.s.l. (e.g., Stirling et al. 1998; McCulloch and Esat 2000; Shackleton 2000; Murray-Wallace 2002; Waelbroeck et al. 2002; Hearty et al. 2007; Schellmann and Radtke 2004; Siddall et al. 2007; Alley et al. 2010; Muhs et al. 2011; Dutton and Lambeck 2012). Coastal deposits of this substage usually crop out several meters above present sea level.
Regarding the MIS 5a substage, considerable controversy exists about the position of sea level in that moment. The estimates for eustatic sea level altitude range approximately from +3 (or even +6 and +10) to −30 m, relative to modern sea level (Ludwig et al. 1996; Muhs et al. 2002; Potter and Lambeck 2003; Potter et al. 2004; Wehmiller et al. 2004; Dorale et al. 2010; Abad et al. 2013; Medina-Elizalde 2013). The elevation of the MIS 5a deposits is very variable depending on the studied sites and their tectonic setting (see Coyne et al. 2007, and references therein). Although they often represent submerged outcrops, for example, Muhs et al. (2002) and Wehmiller et al. (2004) report MIS 5e and 5a deposits at similar low elevations.
There is a certain consensus that MIS 3 interestadial sea level was well below its present position, with estimates from approximately −80 to −20 m (Yokoyama et al. 2001; Murray-Wallace 2002; Siddall et al. 2003, 2008). The elevation of MIS 3 deposits around the world is variable, but they are mostly found below present sea level although they have been also reported up to 8 m a.s.l. (see Hodgson et al. 2009).
Considering the sea level scenarios throughout the Late Pleistocene and taking into account that the studied deposits are considered to be in a mostly tectonically stable area, the Pleistocene assemblages from Uruguay are likely to have been deposited during a sea level stand higher than the present one, such as the inferred for MIS 5. Regarding the height of the Uruguayan deposits, the La Coronilla and Zagarzazú deposits are located at about 0.5–1 m a.s.l., whereas the Puerto de Nueva Palmira site is placed about 12–13 m a.s.l. (Martínez et al. 2001; Rojas 2007). These elevation differences are notorious and deserve an explanation, especially considering that the Puerto de Nueva Palmira and Zagarzazú deposits are separated by only about 12 km.
Taphonomy and biological indicators suggest that the Zagarzazú assemblage was deposited in shallow waters protected from waves and probably close to the coastline, due to the presence of the Thalassinoides–Ophiomorpha association (see Verde 2003). The Puerto de Nueva Palmira assemblage reflects the deposition during high energy events that could represent accumulation in the more proximal foreshore. Thus, the water table could have been thicker in Zagarzazú and thinner in Puerto de Nueva Palmira deposit, explaining at least in part the height difference between both deposits. Like the Zagarzazú assemblage, the La Coronilla one is also inferred to have been deposited in a shallower water environment protected from waves. Thus the coastline at that time would have been situated inland, possibly reducing the elevation difference of the deposits. However, even considering the depth of deposition, the Puerto de Nueva Palmira assemblage is placed at an important altitude. Although there is still no published reference to neotectonics in the Uruguayan coast, Martínez et al. (2001) considered likely that neotectonics had resulted in a local uplift of the Puerto de Nueva Palmira deposit, which remains to be proved. Another possibility to take into account is that the Uruguayan Pleistocene deposits have different ages, as suggested by the available datings. Consequently, they may have been deposited during different sea-level position scenarios. For instance, the Puerto de Nueva Palmira assemblage could have been deposited during MIS 5e and the Zagarzazú deposit during MIS 5a.
(c) Paleoenvironmental inferences from the molluscan assemblages
Another line of evidence that should be considered in order to infer the most probable age of the Late Pleistocene invertebrate assemblages from Uruguay is the paleoenvironmental information provided by the fauna. As it has been mentioned above, only one species (S. lessoni) recorded in the Puerto de Nueva Palmira assemblage is considered to be a cold water taxon, which currently lives in rocky shores of the Uruguayan coast (e.g., Scarabino et al. 2006a; Brazeiro et al. 2006). However, the Late Pleistocene assemblages include 17 extralimital warm water molluscan taxa that currently do not reach the Uruguayan coast (Fig. 7). These molluscs represent 20 % of the taxa recorded providing a strong environmental signal. The former presence of these taxa in the Uruguayan coast points to the existence of higher temperatures than present (Martínez et al. 2001; Rojas 2007; Rojas and Urteaga 2011). The majority of the warm water molluscs recorded has their southernmost distribution boundary in Santa Catarina, Brazil . Thus, it can be inferred that the temperature regime of the Uruguayan coast when these assemblages lived could have been similar to those found today in that region of the Brazilian coast. Considering the Late Pleistocene period, the only time interval in which similar or higher temperatures than present are inferred is the substage MIS 5e. For example, taking into account Southern Hemisphere data, temperatures about 3 °C (Petit et al. 1999) or +4 to +5 °C higher than present were reconstructed from Antarctic ice cores (EPICA 2006; Jouzel et al. 2007). Bianchi and Gersonde (2002) and Cortese and Abelmann (2002) inferred +2 to +3 °C higher than present temperatures based on diatoms and radiolarians from marine sediment records in the Atlantic sector of the Southern Ocean. Also, a global dataset of ice, marine, and terrestrial sequences suggests that global temperatures were on average 1.5 °C higher than today, also showing a strong latitudinal temperature gradient with greater warming at higher latitudes (Turney and Jones 2010).
Additionally, MIS 5e fossil assemblages worldwide commonly record extralimital warm water species. This is the case for Pacific and western Atlantic islands like Hawaii and Bermuda but also around North America and Greenland (see Muhs et al. 2002), for Mediterranean and eastern Atlantic islands assemblages (e.g., Zazo et al. 2003, 2013; Garilli 2011; Montesinos et al. 2014). Most studies considering shifting geographic ranges of species invoke weaker cold currents and stronger warm currents to explain biogeographic changes shown by calid water taxa during the warmer MIS 5e (e.g., Muhs et al. 2002, among others). Similarly, a stronger and southward reaching warm Brazilian Current has been invoked to explain the presence of warm water molluscs in the Late Pleistocene deposits from Uruguay (Martínez et al. 2001; Rojas 2007). Also, the same mechanism has been proposed for the finding of northward displaced species in Holocene Argentinean assemblages, especially those from the province of Buenos Aires (Aguirre 1991, 1993; Aguirre and Farinati 2000; Aguirre et al. 2005).
The environmental and paleobiogeographical information derived from the Late Pleistocene fossil assemblages from Uruguay fits in the environmental scenario reconstructed for MIS 5e from multiple lines of evidence. However, some issues remain to be considered. For example, the La Coronilla assemblage contains 12 extralimital warm water molluscan taxa, whereas the Puerto de Nueva Palmira records six taxa and the Zagarzazú assemblage records three taxa. As it was explained before, different depositional conditions are inferred for these three assemblages. Thus, taphonomic processes may have played a role in the preservation potential of the fauna. Another variable that must be considered is salinity. The La Coronilla assemblage is located in the Atlantic sector of the Uruguayan coast; thus, fully marine conditions are to be expected. However, the Puerto de Nueva Palmira and Zagarzazú deposits are in the western coast of Uruguay, where freshwater environments develop today. Although the salinity requirements of the taxa recorded in these assemblages suggest higher salinities in that area, a western freshwater discharge was supposed to be already working then (e.g., Potter 1998; Iriondo 2004; Veroslavsky and Ubilla 2007), lowering salinity to some extent. Thus, this difference with the La Coronilla assemblage may be playing a role in the lower species richness of the western deposits. There is still another possibility to consider. Maybe the three assemblages are not coetaneous, as suggested by the available OSL ages, implying deposition under rather different environmental conditions. Meanwhile the warmer water character of the La Coronilla fossil assemblage probably represents a MIS 5e deposit, the Puerto de Nueva Palmira, and Zagarzazú assemblages may be likely representing the occurrence of MIS 5a deposits.
Globally, there are fewer coastal records comprising MIS 5a and as mentioned above, there is conflicting evidence about the sea-level position and temperature regimes of this substage relative to the present conditions. Some studies of tectonically stable areas from the Northern Hemisphere suggest that sea level during MIS 5a was above the present one, with temperature regimes similar to present conditions explained by the increased melting of ice sheets (e.g., Ludwig et al. 1996; Muhs et al. 2002; Dorale et al. 2010). Some studies of pollen and diatoms from Baffin Island, Canada, show that climate was warmer than the Holocene around 80 ka B.P., implying the almost complete deglaciation of the Laurentide ice sheet (Miller et al. 1999). Regarding the Southern Hemisphere data, controversies are recorded by pollen and beetles (e.g., Burge and Shulmeister 2007; Fletcher and Thomas 2010) from Tasmania and New Zealand.
Considering the OSL ages obtained for Puerto de Nueva Palmira and Zagarzazú deposits and the fact that they record less extralimital warm water molluscs than the fossil assemblage of La Coronilla, these western assemblages could represent deposits of the less understood MIS 5a substage.
Ongoing research on the Late Pleistocene deposits from Uruguay suggests that the knowledge of the molluscan composition of the assemblages can be improved and richness seems to be higher than that reported by previous studies (Martínez et al. 2001; Rojas 2007; Martínez et al. 2013). Along with the paleoecological interpretations, new geochronological data including AMS and other dating methods such as U/Th are needed to achieve a higher resolution in the timing of the environmental and faunal changes occurred in the Uruguayan coast during the last thousand years.
9 Conclusions
Uruguay has three undoubtedly Late Pleistocene localities with marine fossiliferous deposits; the western Puerto de Nueva Palmira, Zagarzazú assemblages and the eastern La Coronilla assemblage. Conflicting ages were obtained from different dating methods. Whereas standard radiocarbon ages are related to MIS 3, OSL datings from the western deposits are older and related to MIS 5a. The paleoecological and biogeographical analyses of the abundant molluscan fauna point to a higher than present water temperature for the Uruguayan coast, due to the presence of a significant number of extralimital warm water taxa. These taxa that currently live in Brazilian waters are well represented in the La Coronilla assemblage and have a lesser representation in the Puerto de Nueva Palmira and Zagarzazú assemblages. Higher temperatures than present and widespread biogeographic changes of marine fauna have been widely recognized for MIS 5e (Last Interglacial) assemblages. Thus, all available evidence suggest that the Late Pleistocene assemblages from Uruguay were deposited during MIS 5, thus the radiocarbon dating indicating MIS 3 times should be considered as minimum ages. The La Coronilla assemblage most likely represents a MIS 5e assemblage, meanwhile Puerto de Nueva Palmira and Zagarzazú deposits could have been originated during the MIS 5a substage, as suggested by the available OSL ages and the lower representation of extralimital northern molluscs .
References
Abad M, Rodríguez-Vidal J, Aboumaria K, Zaghloul MN, Cáceres LM, Ruiz F, Martínez-Aguirre A, Izquierdo T, Chamorro S (2013) Evidence of MIS 5 sea-level highstands in Gebel Mousa coast (Strait of Gibraltar, North of Africa). Geomorph 182:133–146
Acha EM, Mianzan H, Guerrero R, Carreto J, Giberto D, Montoya N, Carignan M (2008) An overview of physical and ecological processes in the Rio de la Plata. Est Cont Shelf Res 28:1579–1588
Aguirre ML (1991) Holocene macrobenthic molluscan associations from northeastern Buenos Aires Province, Argentina. Quat South Am Antarct Peninsula 7:161–195
Aguirre ML (1993) Palaeobiogeography of the Holocene molluscan fauna from northeastern Buenos Aires Province, Argentina: its relation to coastal evolution and sea level changes. Palaeogeogr Palaeoclimatol Palaeoecol 102:1–26
Aguirre ML (2003) Late Pleistocene and Holocene palaeoenvironments in Golfo San Jorge, Patagonia: molluscan evidence. Mar Geol 194:3–30
Aguirre ML, Farinati EA (2000) Moluscos del Cuaternario marino de la Argentina. Bol Acad Nac Cienc Córdoba 64:235–333
Aguirre M, Whatley RC (1995) Late quaternary marginal marine deposits and palaeoenvironments from northeastern Buenos Aires Province, Argentina: a review. Quat Sci Rev 14:223–254
Aguirre ML, Donato M, Richiano S, Farinati EA (2011) Pleistocene and Holocene interglacial molluscan assemblages from Patagonia and Bonaerensian littoral (Argentina, SW Atlantic): palaeobiodiversity and palaeobiogeography. Palaeogeogr Palaeoclimatol Palaeoecol 308:277–292
Aguirre ML, Negro Sirch Y, Richiano S (2005) Late Quaternary molluscan assemblages from the coastal area of Bahía Bustamante (Patagonia, Argentina): paleoecology and paleoenvironments. J South Am Earth Sci 20:13–32
Alley RB, Andrews JT, Brigham-Grette J, Clarke GKC, Cuffey KM, Fitzpatrick JJ, Funder S, Marshall SJ, Miller GH, Mitrovica JX, Muhs DR, Otto-Bliesner BL, Polyak L, White JWC (2010) History of the Greenland Ice Sheet: paleoclimatic insights. Quat Sci Rev 29:1728–1756
Amano K (1994) An attempt to estimate the surface temperature of the Japan Sea in the Early Pleistocene by using a molluscan assemblage. Palaeogeogr Palaeoclimatol Palaeoecol 108:369–378
Amaral ACZ, Rizzo AE, Arruda EP (2006) Manual de identificação dos invertebrados marinhos da Região Sudeste-Sul do Brasil, vol 1. Edusp, Sao Paulo
Ávila SP, Madeira P, Zazo C, Kroh A, Kirby M, Marques da Silva C, Cachão M, de Frias Martins AM (2009) Palaeoecology of the Pleistocene (MIS 5.5) outcrops of Santa Maria Island (Azores) in a complex oceanic tectonic setting. Palaeogeogr Palaeoclimatol Palaeoecol 274:18–31
Barker S, Diz P, Vautravers MJ, Pike J, Knorr G, Hall IR, Broecker WS (2009) Interhemispheric Atlantic seesaw response during the last deglaciation. Nature 457:1097–1102
Berkman PA, Andrews JT, Bjorck S, Colhoun EA, Emslie SD, Goodwin ID, Hall BL, Hart CP, Hirakawa K, Igarashi A, Ingolfsson O, Lopez-Martinez J, Lyons WB, Mabin MCG, Quilty PG, Taviani M, Yoshida Y (1998) Circum-Antarctic coastal environmental shifts during the Late Quaternary reflected by emerged marine deposits. Ant Sci 10(3):345–362
Bianchi C, Gersonde R (2002) The Southern Ocean surface between Marine Isotope Stages 6 and 5d: shape and timing of climate changes. Palaeogeogr Palaeoclimatol Palaeoecol 187:151–177
Bossi J (1966) Geología del Uruguay. Departamento de Publicaciones de la Universidad de la República, Montevideo, Uruguay
Bossi J, Ferrando LA, Fernández N, Elizalde G, Morales H, Ledesma JJ, Carballo E, Medina E, Ford I, Montaña JR (1975) Carta Geológica del Uruguay. Escala 1:1.000.000. Dirección de Suelos y Fertilizantes, Ministerio de Agricultura y Pesca, Montevideo
Bracco R, Ures C (1998) Las variaciones del nivel del mar y el desarrollo de las culturas prehistóricas en el Uruguay. Actas Congr Uruguayo Geol 2:16–20
Brazeiro A, Borthagaray AI, Giménez L (2006) Patrones geográficos de diversidad bentónica en el litoral rocoso de Uruguay. In: Menafra R, Rodríguez L, Scarabino F, Conde D (eds) Bases para la conservación y el manejo de la costa uruguaya. Vida Silvestre, Montevideo
Briggs JC (1995) Global biogeography. Developments in palaeontology and stratigraphy, vol 14. Elsevier, Amsterdam
Buiron D, Stenni B, Chappellaz J, Landais A, Baumgartner M, Bonazza M, Capron E, Frezzotti M, Kageyama M, Lemieux-Dudon B, Masson-Delmotte V, Parrenin F, Schilt A, Selmo E, Severi M, Swingedouw D, Udisti R (2012) Regional imprints of millennial variability during the MIS 3 period around Antarctica. Quat Sci Rev 48:99–112
Burge PI, Shulmeister J (2007) An MIS 5a to MIS 4 (or early MIS 3) environmental and climatic reconstruction from the northwest South Island, New Zealand, using beetle fossils. J Quat Sci 22:501–516
Calcaterra A (1971) Algunas observaciones sobre la fauna de moluscos de la Formación Querandina (Deptos de Colonia y Soriano, Uruguay). Com Soc Malac Uruguay 3(20):79–82
Caorsi J, Goñi J (1958) Geología Uruguaya, vol 37. Boletín Instituto Geológico Uruguayo
Charó MP, Fucks EE, Gordillo S (2013) Moluscos marinos bentónicos del Cuaternario de Bahía Anegada (sur de Buenos Aires, Argentina): variaciones faunísticas en el Pleistoceno tardío y Holoceno. Rev Mex Cienc Geol 30(2):404–416
Charó MP, Gordillo S, Fucks EE, Giaconi LM (2014) Late Quaternary molluscs from the northern San Matías Gulf (Northern Patagonia, Argentina), southwestern Atlantic: faunistic changes and paleoenvironmental interpretation. Quatern Int 352:26–47
Clavijo C, Scarabino F, Rojas A, Martínez S (2005) Lista sistemática de los moluscos marinos y estuarinos del Cuaternario del Uruguay. Com Soc Malac Uruguay 9(88):381–411
Cortese G, Abelmann A (2002) Radiolarian-based paleotemperatures during the last 160 kyr at ODP Site 1089 (Southern Ocean, Atlantic Sector). Palaeogeogr Palaeoclimatol Palaeoecol 182:259–286
Coyne MK, Jones B, Ford D (2007) Highstands during Marine Isotope Stage 5: evidence from the Ironshore Formation of Grand Cayman. Br West Indies Quat Sci Rev 26:536–559
Crowley TJ, North GR (1991) Paleoclimatology. Oxford University Press Inc, New York
De Diego-Forbis T, Douglas R, Gorsline D, Nava-Sanchez E, Mack L, Banner J (2004) Late Pleistocene (last interglacial) terrace deposits, Bahia Coyote, Baja California Sur, Mexico. Quatern Int 120:29–40
Delaney PJC (1963) Fisiografia e geología de superficie da Planicie Costeira do Rio Grande do Sul. Dissertation, Universidade de Sao Paulo
Dorale JA, Onac BP, Fornos JJ, Gines J, Gines A, Tuccimei P, Peate DW (2010) Sea-level highstand 81,000 years ago in Mallorca. Science 327:860–863
Dutton A, Lambeck K (2012) Ice volume and sea level during the last interglacial. Science 337:216–219
Emiliani C (1955) Pleistocene temperatures. J Geol 63:538–578
EPICA Community Members (2004) Eight glacial cycles from an Antarctic ice core. Nature 429:623–628
EPICA Community Members (2006) One-to-one coupling of glacial climate variability in Greenland and Antarctica. Nature 444:195–198
Figueiras A (1962) Sobre nuevos hallazgos de moluscos subfósiles de la Transgresión Querandina. Com Soc Malac Uruguay 1(3):53–68
Fletcher MS, Thomas I (2010) A quantitative Late Quaternary temperature reconstruction from western Tasmania, Australia. Quat Sci Rev 29:2351–2361
Fontana MA (1930) Memoria de la excursión científica a Nueva Palmira: Abril de 1927. Rev Soc Am Arq 4:119–178
Forti Esteves IR (1974) Bioestratigrafia e palaeoecologia (Mollusca) do Quaternario da Planicie costeira do Rio Grande do Sul (Brasil). Annais do Congresso Brasileiro de Geologia 28:133–149
Francis JC (1975) Esquema bioestratigráfico regional de la República Oriental de Uruguay. Actas 1er Congr Arg Paleont Biostr 2:539–568
Frenguelli J (1930) Apuntes de Geología Uruguaya. Bol Inst Geol Perf 11:1–47
Garilli V (2011) Mediterranean quaternary interglacial molluscan assemblages: palaeobiogeographical and palaeoceanographical responses to climate change. Palaeogeogr Palaeoclimatol Palaeoecol 312:98–114
Giberto DA, Bremeca CS, Acha EM, Mianzan H (2004) Large-scale spatial patterns of benthic assemblages in the SW Atlantic: the Río de la Plata estuary and adjacent shelf waters. Est Coast Shelf Sci 61:1–13
Giménez L, Borthagaray AI, Rodríguez M, Brazeiro A, Dimitriadis C (2005) Scale-dependent patterns of macrofaunal distribution in soft-sediment intertidal habitats along a large-scale estuarine gradient. Helgol Mar Res 59:224–236
Goñi JC, Hoffstetter R (1964) Uruguay. Lexique stratigraphique. international, 5 Amerique Latine, 9a. Centre National de la Recherche Scientifique, Paris
Gordillo S, Isla FI (2011) Faunistic changes between the Middle/Late Pleistocene and the Holocene on the Atlantic coast of Tierra del Fuego: molluscan evidence. Quat Int 233:101–112
Goso H (1972) Cuaternario. Technical report, Programa de Estudio y Levantamiento de Suelos, Ministerio de Ganadería y Agricultura, Montevideo
Guerrero RA, Acha EM, Framiñan MB, Lasta CA (1997) Physical oceanography of the Río de la Plata Estuary, Argentina. Cont Shelf Res 17(7):727–742
Hearty PJ, Hollin JT, Neumann AC, O’Leary MJ, McCulloch M (2007) Global sea-level fluctuations during the Last Interglaciation (MIS 5e). Quat Sci Revs 26:2090–2112
Hodgson DA, Verleyen E, Vyverman W, Sabbe K, Leng MJ, Pickering MD, Keely BJ (2009) A geological constraint on relative sea level in Marine Isotope Stage 3 in the Larsemann Hills, Lambert Glacier region, East Antarctica (31366–33228 cal yr BP). Quat Sci Rev 28:2689–2696
Huber C, Leuenberger M, Spahni R, Flückiger J, Schwander J, Stocker TF, Johnsen S, Landais A, Jouzel J (2006) Isotope calibrated Greenland temperature record over Marine Isotope Stage 3 and its relation to CH4. Earth Planet Sci Lett 243:504–519
Iriondo M (2004) The littoral complex at the Paraná River mouth. Quat Int 114:143–154
Isla FI, Rutter NW, Schnack EJ, Zarate MA (2000) La transgresión Belgranense en Buenos Aires. Una revisión a cien años de su definición. Rev Cuaternario Cienc Ambientales 4:3–14
Jain M, Bøtter-Jensen L, Singhvi AK (2003) Dose evaluation using multiple-aliquot quartz OSL: test of methods and a new protocol for improved accuracy and precision. Radiat Meas 37(1):67–80
Jouzel J, Masson-Delmotte V, Cattani O, Dreyfus G, Falourd S, Hoffmann G, Minster B, Nouet J, Barnola JM, Chappellaz J, Fischer H, Gallet JC, Johnsen S, Leuenberger M, Loulergue L, Luethi D, Oerter H, Parrenin F, Raisbeck G, Raynaud D, Schilt A, Schwander J, Selmo E, Souchez R, Spahni R, Stauffer B, Steffensen JP, Stenni B, Stocker TF, Tison JL, Werner M, Wolff EW (2007) Orbital and millennial Antarctic climate variability over the past 800,000 years. Science 317:793–796
Kitamura A, Ubukata T (2003) The sequence of local recolonization of warm-water marine molluscan species during a deglacial warming climate phase: a case study from the early Pleistocene of the Sea of Japan. Palaeogeogr Palaeoclimatol Palaeoecol 199:83–94
Kraglievich L (1928) Apuntes sobre la Geología y Paleontología de la República Oriental del Uruguay. Rev Am Arq 2:5–61
Kraglievich L (1932) Nuevos apuntes para la geología y paleontología uruguayas. Anales Mus Hist Nat Montvideo 2(3):257–320
Long J, Stoy P (2013) Quantifying the periodicity of Heinrich and Dansgaard-Oeschger events during Marine Oxygen Isotope Stage 3. Quat Res 79:413–423
Lopes RP, Simone LRL (2012) New fossil records of Pleistocene marine mollusks in southern Brazil. Rev Bras Paleontol 15(1):49–56
Lopes RP, Simone LRL, Dillenburg SR, Schultz CL, Pereira JC (2013) A Middle Pleistocene marine molluscan assemblage from the southern coastal plain of Rio Grande do Sul State, Brazil. Rev Bras Paleontol 16(3):343–360
Lorenzo N, Verde M (2004) Estructuras de bioerosión en moluscos marinos de la Formación Villa Soriano (Pleistoceno tardío—Holoceno) de Uruguay. Rev Bras Paleontol 7:319–328
Ludwig KR, Muhs DR, Simmons KR, Halley RB, Shinn EA (1996) Sea-level records at 80 ka from tectonically stable platforms: Florida and Bermuda. Geology 24(3):211–214
Mallinson D, Burdette K, Mahan S, Brook G (2008) Optically stimulated lumninescense age controls on late Pleistocene and Holocene coastal lithosomes, North Carolina, USA. Quat Res 69:97–109
Martínez S (1988) Moluscos fósiles holocenos de la margen occidental de la Laguna Merín (Formación Villa Soriano, Uruguay). Com Soc Malacol Uruguay 6:409–418
Martínez S (1990) Taphonomy and Palaeoecology of Holocene mollusks from the western margin of the Merin Lagoon (Villa Soriano Fm., Uruguay). Quat South Am Antarct Peninsula 7:121–135
Martínez S, Ubilla M (2004) El Cuaternario de Uruguay. In: Veroslavsky G, Ubilla M, Martínez S (eds) Cuencas sedimentarias de Uruguay: Geología, palaeontología y recursos naturales, Cenozoico. DIRAC, Montevideo
Martínez S, del Río CJ (2002) Late Miocene molluscs from the Southwestern Atlantic Ocean (Argentina and Uruguay): a palaeobiogeographic analysis. Palaeogeogr Palaeoclimatol Palaeoecol 188:167–187
Martínez S, del Río C, Rojas A (2013) Biogeography of the quaternary molluscs of the Southwestern Atlantic Ocean. Springer Briefs in Earth System Sciences, South America and the southern hemisphere
Martínez S, Rojas A, Ubilla M, Verde M, Perea D, Piñeiro G (2006) Molluscan assemblages from the marine Holocene of Uruguay: composition, geochronology, and paleoenvironmental signals. Amegh 43(2):385–397
Martínez S, Ubilla M, Verde M, Perea D, Rojas A, Guérèquiz R, Piñeiro G (2001) Paleoecology and geochronology of Uruguayan coastal marine Pleistocene deposits. Quat Res 55:246–254
Masello A, Menafra R (1998) Comunidades macrobentónicas de la zona costera uruguaya y áreas adyacentes. In: Wells PG, Daborn GR (Eds) El Río de la Plata—Una revisión ambiental. Un informe del Proyecto ECOPLATA. Dalhousie University, Halifax
McCulloch MT, Esat T (2000) The coral record of last interglacial sea levels and sea surface temperatures. Chem Geol 169:107–129
Medina-Elizalde M (2013) A global compilation of coral sea-level benchmarks: implications and new challenges. Earth Planet Sci Lett 362:310–318
Mianzan H, Lasta C, Acha M, Guerrero R, Macchi G, Bremen C (2001) The Río de la Plata Estuary, Argentina-Uruguay. In: Seeliger U, Kjerfve B (eds) Coastal marine ecosystems of Latin America. Springer-Verlag, Berlin
Miller GH, Mode WN, Wolfe AP, Sauer PE, Bennike O, Forman SL, Short SK, Staford TW Jr (1999) Stratifed interglacial lacustrine sediments from Baffin Island, Arctic Canada: chronology and paleoenvironmental implications. Quat Sci Rev 18:789–810
Möller OO Jr, Piola AR, Freitas AC, Campos EJD (2008) The effects of river discharge and seasonal winds on the shelf off southeastern South America. Cont Shelf Res 28:1607–1624
Montesinos M, Ramos AJG, Lomoschitz A, Coca J, Redondo A, Betancort JF, Meco J (2014) Extralimital Senegalese species during Marine Isotope Stages 5.5 and 11in the Canary Islands (29° N): Sea surface temperature estimates. Palaeogeogr Palaeoclimatol Palaeoecol 410:153–163
Muhs DR, Simmons KR, Randall Schumann R, Halley RB (2011) Sea-level history of the past two interglacial periods: new evidence from U-series dating of reef corals from south Florida. Quat Sci Rev 30:570–590
Muhs DR, Simmons KR, Steinke B (2002) Timing and warmth of the Last Interglacial period: new U-series evidence from Hawaii and Bermuda and a new fossil compilation for North America. Quat Sci Revs 21:1355–1383
Murray-Wallace CV (2002) Pleistocene coastal stratigraphy, sea-level highstands and neotectonism of the southern Australian passive continental margin—a review. J Quat Sci 17(5–6):469–489
Murray-Wallace CV, Belperio AP (1991) The last interglacial shoreline in Australia—a review. Quat Sci Rev 10(5):441–461
Murray-Wallace CV, Beu AG, Kendrick GW, Brown LJ, Belperio AP, Sherwood JE (2000) Palaeoclimatic implications of the occurrence of the arcoid bivalve Anadara trapezia (Deshayes) in the Quaternary of Australasia. Quat Sci Rev 19(6):559–590
Nagy GJ, Severov DN, Pshennikov VA, De los Santos M, Lagomarsino JJ, Sans K, Morozov EG (2008) Rio de la Plata estuarine system: relationship between river flow and frontal variability. Adv Space Res 41:1876–1881
Navarro R (1990) A brief review on the knowledge on the Quaternary of the Southern and Southeastern Coast of Uruguay. Quat South Am Antarct Peninsula 7:107–117
Nielsen JK, Hanken N-M, Nielsen JK, Hansen KS (2006) Biostratigraphy and palaeoecology of the marine Pleistocene of Rhodes, Greece: Scleractinia, Serpulidae, Mollusca and Brachiopoda. Bull Geosci 81(3):173–196
North Greenland Ice Core Project Members (2004) High-resolution record of Northern Hemisphere climate extending into the last interglacial period. Nature 431:147–151
Odebrecht C, Castello JP (2001) The convergence ecosystem in the Southwest Atlantic. In: Seeliger U, Kjerfve B (eds) Coastal marine ecosystems of Latin America. Springer, Berlin
Olson DB, Podestá GP, Evans RH, Brown OB (1988) Temporal variations in the separation of Brazil and Malvinas Currents. Deep Sea Res 35(12):1971–1990
Orombelli G, Maggi V, Delmonte B (2010) Quaternary stratigraphy and ice cores. Quat Int 219:55–65
Ortlieb L, Devries T, Diaz A (1990) Ocurrencia de Chione broggi (Pilsbry & Olsson, 1943) (Pelecypoda) en depositos litorales cuaternarios del sur del Perú: implicaciones paleoceanográficas. Bol Soc Geol Perú 81:127–134
Petit J-R, Jouzel J, Raynaud D, Barkov NI, Barnola JM, Basile I, Bender M, Chappellaz J, Davis M, Delaygue G, Delmotte M, Kotlyakov VM, Legrand M, Lipenkov VY, Lorius C, Pepin L, Ritz C, Saltzman E, Stievenard M (1999) Climate and atmospheric history of the past 420000 years from the Vostok ice core, Antarctica. Nature 399:429–436
Pimenta AD, Absalão RS, Miyaji C (2009) A taxonomic review of the genera Boonea, Chrysallida, Parthenina, Ivara, Fargoa, Mumiola, Odostomella and Trabecula (Gastropoda, Pyramidellidae, Odostomiinae) from Brazil. Zootaxa 2049:39–66
Piola AR, Campos EJD, Möller OO Jr, Charo M, Martínez C (2000) Subtropical shelf off eastern South America. J Geophys Res 105:6565–6578
Potter E-K, Lambeck K (2003) Reconciliation of sea-level observations in the Western North Atlantic during the last glacial cycle. Earth Planet Sci Lett 217:171–181
Potter E-K, Esat TM, Schellmann G, Radtke U, Lambeck K, McCulloch MT (2004) Suborbital-period sea-level oscillations during marine isotope substages 5a and 5c. Earth Planet Sci Lett 225:191–204
Potter PE (1998) The Mesozoic and Cenozoic paleodrainage of South America: a natural history. J South Am Earth Sci 10:331–344
Powell CL II, Grant LB, Conkling SW (2004) Paleoecologic Analysis and Age of a Late Pleistocene Fossil Assemblage at a Locality in Newport Beach, Upper Newport Bay, Orange County, California. The Veliger 47(3):171–180
Preciozzi F, Spoturno J, Heinzen W, Rossi P (1988) Memoria explicativa de la Carta Geológica del Uruguay a la escala 1: 500.000. DINAMIGE, Montevideo
Prescott JR, Hutton JT (1994) Cosmic ray contributions to dose rates for luminescence and ESR dating. Radiat Meas 23:497–500
Rabassa J, Gordillo S, Ocampo C, Rivas Hurtado P (2009) The southernmost evidence for an interglacial transgression (Sangamon?) in South America. First record of upraised Pleistocene marine deposits in Isla Navarino (Beagle Channel, Southern Chile). Geol Acta 6(3):251–258
Ríos E (1994) Seashells of Brazil. Editora Fundaçao Universidade do Rio Grande, Rio Grande
Ríos E (2009) Compendium of Brazilian sea shells. Evangraf, Rio Grande
Rivadeneira MM, Carmona ER (2008) A Late Pleistocene macro-benthic assemblage in Caleta Patillos, northern Chile: paleoecological and paleobiogeographical interpretations. Rev Geol Chile 35:163–173
Rojas A (2003) Moluscos marinos cuaternarios del Balneario Zagarzazú, Departamento de Colonia, Uruguay. Pub Esp Soc Zool Uruguay 1:83
Rojas A (2007) Moluscos de aguas cálidas del Cuaternario Marino del Uruguay. Dissertation, Universidad de la República
Rojas A (2010) Primeras edades obtenidas por datación por luminiscencia (OSL) en depósitos marinos fosilíferos del Pleistoceno tardío de Uruguay. X Congr Argent Paleontol y Bioestratigr y VII Congr Lat Paleontol 1:205
Rojas A, Urteaga D (2011) Late Pleistocene and Holocene chitons (Mollusca, Polyplacophora) from Uruguay: palaeobiogeography and palaeoenvironmental reconstruction in mid latitudes of the Southwestern Atlantic. Geobios 44:377–386
Rojas A, Verde M, Urteaga D, Scarabino F, Martínez S (2014) The first predatory drillhole on a fossil chiton plate: an occasional prey item or an erroneous attack? Palaios 29:414–419
Roselli FL (1939) Apuntes de Geología y Paleontología uruguayas. Bol Soc Am Cs Nat “Kraglievich-Fontana” 1(2):29–102
Roselli FL (1976) Contribución al estudio de la Geo Paleontología Departamentos de Colonia y Soriano (República Oriental del Uruguay). Imprenta Cooperativa, Montevideo
Rosenberg G (2009) Malacolog 4.1.1: a database of Western Atlantic Marine Mollusca. [WWW database (version 4.1.1)]. http://www.malacolog.org/. Accessed 26 Nov 2014
Roy K, Jablonski D, Valentine JW (1995) Thermally anomalous assemblages revisited: patterns in the extraprovincial latitudinal range shifts of Pleistocene marine mollusks. Geology 23(12):1071–1074
Scarabino F, Zaffaroni JC (2004) Estatus faunístico de veinte especies de moluscos citadas para aguas uruguayas. Com Zool Mus Nac Hist Nat Antr 13(202):1–15
Scarabino F, Zaffaroni JC, Carranza A, Clavijo C, Nin M (2006a) Gasterópodos marinos y estuarinos de la costa uruguaya: faunística, distribución, taxonomía y conservación. In: Menafra R, Rodríguez L, Scarabino F, Conde D (eds) Bases para la conservación y el manejo de la costa uruguaya. Vida Silvestre, Montevideo
Scarabino F, Zaffaroni JC, Clavijo C, Carranza A, Nin M (2006b) Bivalvos marinos y estuarinos de la costa uruguaya: faunística, distribución, taxonomía y conservación. In: Menafra R, Rodríguez L, Scarabino F, Conde D (eds) Bases para la conservación y el manejo de la costa uruguaya. Vida Silvestre, Montevideo
Scarabino V (1977) Moluscos del Golfo San Matías (Provincia de Río Negro, República Argentina). Inventario y claves para su identificación. Com Soc Malacol Uruguay 4:177–285
Schellmann G, Radtke U (2004) A revised morpho- and chronostratigraphy of the Late and Middle Pleistocene coral reef terraces on Southern Barbados (West Indies). Earth Sci Rev 64:157–187
Serra N (1943) Memoria explicativa del mapa geológico del Departamento de Colonia. Bol Inst Geol Uruguay 30
Shackleton NJ (1969) The Last Interglacial in the Marine and Terrestrial Records. Proc R Soc Lond B Biol Sci 174(1034):135–154
Shackleton NJ (2000) The 100,000 year Ice Age cycle identified and found to lag temperature, carbon dioxide, and orbital eccentricity. Science 289:1897–1902
Shackleton NJ, Sánchez Goñi MF, Pailler D, Lancelot Y (2003) Marine isotope substage 5e and the Eemian Interglacial. Global Planet Change 36:151–155
Sicardi OE (1967) La influencia de las Corrientes marinas sobre la malacofauna uruguaya. Com Soc Malacol Uruguay 2(12):49–60
Siddall M, Rohling EJ, Almogi-Labin A, Hemleben Ch, Meischner D, Schmelzer I, Smeed DA (2003) Sea-level fluctuations during the last glacial cycle. Nature 423:853–858
Siddall M, Rohling EJ, Thompson WG, Waelbroeck C (2008) Marine isotope stage 3 sea level fluctuations: data synthesis and new outlook. Rev Geophys 46:RG4003
Siddall M, Chappell J, Potter E-K (2007) Eustatic sea level during past interglacials. In: Sirocko F, Claussen M, Sanchez-Goni MF, Litt T (Eds) The climate of past interglacials. Developments in Quaternary Science Series No. 7. Elsevier, Amsterdam
Simms AR, DeWitt R, Rodríguez AB, Lambeck K, Anderson JB (2009) Revisiting marine isotope stage 3 and 5a (MIS3-5a) sea levels within the northwestern Gulf of Mexico. Global Planet Change 66:100–111
Sprechmann P (1978) The paleoecology and paleogeography of the Uruguayan coastal area during the Neogene and Quaternary. Zitt 4:3–72
Stirling CH, Esat TM, Lambeck K, McCulloch MT (1998) Timing and duration of the Last Interglacial: evidence for a restricted interval of widespread coral reef growth. Earth Planet Sci Lett 160:745–762
Suguio K, Bezerra FHR, Barreto AMF (2011) Luminescence dated Late Pleistocene wave-built terraces in northeastern Brazil. Acad Bras Cienc 83(3):907–920
Teisseire A (1927) Expedición a los departamentos de Colonia y Soriano. Rev Amigos Arqueol 1:47–61
Teisseire A (1928) Contribución al estudio de la geología y la paleontología de la República Oriental del Uruguay. Región de Colonia. Anales Univ 122(series 37):319–469
Tonni EP, Carbonari JE, Huarte RA (2010) Marine Sediments Attributed to Marine Isotope Stage 3 in the Southeastern Buenos Aires Province, Argentina. Curr Res Pleist 27:154–156
Turney CSM, Jones RT (2010) Does the Agulhas Current amplify global temperatures during super-interglacials? J Quat Sci 25(6):839–843
Urien CM (1972) Río de la Plate estuary environments. In: Nelson BW (ed) Environmental framework of coastal plain estuaries, Boulder; Geol Soc Am Mem 133:213–234
Valentine JW, Jablonski D (1993) Fossil communities: compositional variation at many time scales. In: Ricklefs RE, Schluter D (eds) Species diversity in ecological communities: Historical and geographical perspectives. University of Chicago Press, Chicago
Van Meerbeeck CJ, Renssen H, Roche DM (2009) How did Marine Isotope Stage 3 and Last Glacial Maximum climates differ? Perspectives from equilibrium simulations. Clim Past 5:33–51
Van Meerbeeck CJ, Renssen H, Roche DM, Wohlfarth B, Bohncke SJP, Bos JAA, Engels S, Helmens KF, Sánchez-Goñi MF, Svensson A, Vandenberghe J (2011) The nature of MIS 3 stadial e interstadial transitions in Europe: new insights from model data comparisons. Quat Sci Rev 30:3618–3637
Verde M (2003) Marine invertebrate ichnofossils from the Quaternary of Uruguay. Pub Esp Soc Zool Uruguay 1:94
Veroslavsky G, Ubilla M (2007) A “snapshot” of the evolution of the Uruguay River (Del Plata Basin): the Salto depositional sequence (Pleistocene, Uruguay, South America). Quat Sci Rev 26:2913–2923
Waelbroeck C, Labeyrie L, Michel E, Duplessy JC, McManus JF, Lambeck K, Balbon E, Labracherie M (2002) Sea-level and deep water temperature changes derived from benthic foraminifera isotopic records. Quat Sci Rev 21:295–305
Wehmiller JF, Simmons KR, Cheng H, Edwards L, Martin-McNaughton J, York LL, Krantz DE, Shen C (2004) Uranium-series coral ages from the US Atlantic Coastal Plain—The “80 ka problem” revisited. Quat Int 120:3–14
Winograd IJ, Landwehr JM, Ludwig KR, Coplen TB, Riggs AC (1997) Duration and structure of the past four interglaciations. Quat Res 48:141–154
Yokoyama Y, Esat TM, Lambeck K (2001) Coupled climate and sea-level changes deduced from Huon Peninsula coral terraces of the last ice age. Earth Planet Sci Lett 193:579–587
Zazo C, Goy JL, Dabrio CJ, Bardají T, Hillaire-Marcel C, Ghaleb B, González-Delgado JA, Soler V (2003) Pleistocene raised marine terraces of the Spanish Mediterranean and Atlantic coasts: records of coastal, sea-level highstands and climate changes. Mar Geol 194(1–2):103–133
Zazo C, Goy JL, Dabrio CJ, Lario J, González-Delgado JA, Bardají T, Hillaire-Marcel C, Cabero A, Ghaleb B, Borja F, Silva PG, Roquero E, Soler V (2013) Retracing the Quaternary history of sea-level changes in the Spanish Mediterranean-Atlantic coasts: geomorphological and sedimentological approach. Geomorphology 196:36–49
Zubakov V, Borzenkova I (1990) Global paleoclimate of the Late Cenozoic. Elsevier, New York
Acknowledgments
We acknowledge the invitation by Eduardo P. Tonni and Jorge Rabassa to attend the “Simposio multidisciplinario, El estadio isotópico 3 en Argentina y el sur de América del Sur: 60.000 a 25.000 años atrás”, at La Plata, Argentina , from which this publication has derived. Germán M. Gasparini kindly helped with editorial issues. We thank Fabrizio Scarabino for providing samples of the La Coronilla assemblage. Juan Carlos Zaffaroni and Alexandre Dias Pimenta helped with micromollusc determinations. Mariano Verde assisted during the fieldwork and Mariana Demicheli aided in the preparation of samples and picking of specimens. Martín Ubilla helped during OSL sampling and provided useful bibliography. We also thank the personnel at Museo Lucas Rosselli and Administración Nacional de Puertos (ANP) of Nueva Palmira for allowing the access to the Puerto de Nueva Palmira deposit. This is a contribution to “Programa de Desarrollo de las Ciencias Básicas” (PEDECIBA-Biología), Comisión Sectorial de Investigación Científica (CSIC), and Agencia Nacional de Investigación e Innovación (ANII/FCE2007_034 and ANII/FCE2007_058) for providing funding for this research.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Appendix: Data of the OSL ages obtained for Puerto de Nueva Palmira and Zagarzazú
Appendix: Data of the OSL ages obtained for Puerto de Nueva Palmira and Zagarzazú
Sample code | Lab number | Equivalent dose (Grays)a | U (ppm)b | Th (ppm)b | K2O (%)b | H2O (%)c | Cosmic dose (mGrays/year)d | Total dose (mGrays/year) | OSL age (year)e |
|---|---|---|---|---|---|---|---|---|---|
PNPLO1 | UIC2632 | 73.58 ± 4.78 | 0.6 ± 0.1 | 1.5 ± 0.1 | 0.64 ± 0.01 | 10 ± 3 | 0.016 ± 0.002 | 0.91 ± 0.04 | 80,680 ± 5,500 |
ZZZLO1 | UIC2633 | 124.53 ± 8.16 | 0.7 ± 0.1 | 2.1 ± 0.1 | 1.34 ± 0.01 | 20 ± 5 | 0.016 ± 0.002 | 1.41 ± 0.06 | 88,355 ± 7,070 |
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Rojas, A., Martínez, S. (2016). Marine Isotope Stage 3 (MIS 3) Versus Marine Isotope Stage 5 (MIS 5) Fossiliferous Marine Deposits from Uruguay. In: Gasparini, G., Rabassa, J., Deschamps, C., Tonni, E. (eds) Marine Isotope Stage 3 in Southern South America, 60 KA B.P.-30 KA B.P.. Springer Earth System Sciences. Springer, Cham. https://doi.org/10.1007/978-3-319-40000-6_14
Download citation
DOI: https://doi.org/10.1007/978-3-319-40000-6_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-39998-0
Online ISBN: 978-3-319-40000-6
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)