
Abstract
A significant number of HIV-infected patients develop neurological symptoms ranging from minor cognitive impairment to severe dementia (known HIV-associated dementia, HAD). Without combination antiretroviral therapy (cART) HAD occurs in 20–40 % of HIV positive subjects, with the advent of cART the incident has decreased to 5–10 %, although milder forms of cognitive deficits may occur in 30–50 % of those infected with HIV. It is believed that HIV enters the CNS during the early stages of infection by infected immune cells which initiate an inflammatory cascade which results in neuronal injury and loss. Imaging has been widely used to evaluate the effect of HIV to the brain. Specifically, proton magnetic resonance spectroscopy (1H MRS) is one of the most informative methods employed in patients suffering from HIV-associated neurocognitive disorders (HAND). MRS is able to noninvasively measure metabolic changes pertaining to neuronal injury and inflammation, thus, it can assist in the diagnosis of the disease and measure the severity of injury. In animal models of neuroAIDS, MRS has been proven extremely powerful to assess disease progression and response to treatment. Here we review the literature of preclinical models as well as MRS studies of HIV+ adults and children before and after the advent of cART regiments. In addition, we discuss technical considerations related to the disease and finally talk about future direction in HAND using MRS.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others



Keywords
- AIDS (acquired immune deficiency syndrome)
- HIV (Human immunodeficiency Virus)
- NeuroAIDS
- HAND (HIV-associated neurocognitive disorders)
- SIV (Simian immunodeficiency virus)
- Rhesus Macaque
- Brain
- MRS (magnetic resonance spectroscopy)
- MRSI (magnetic resonance spectroscopic imaging)
- cART (combination antiretroviral therapy)
- Minocycline
- Neuroinflammation
Background on Disease Pathology
Human immunodeficiency virus (HIV) affects more than 1 million people in the United States of America and ~34 million worldwide. Approximately 1.7 million people have died from AIDS and 2.5 million people were newly infected with the virus in 2011 [1]. Figure 11.1 showed the natural disease progression of HIV/AIDS: During the early phase of infection, the virus replicates extensively, reaching levels of 106–108 copies of HIV RNA/mL in blood plasma due to the absence of discernible immune responses [2]. Once seroconversion occurs (as determined by ELISA/Western Blot), it is thought that the host’s immune response begins to control viral replication, causing the high levels of virus to be quickly reduced within weeks [2]. During acute HIV-1 infection, there is frequently a marked decrease in the CD4+ T cell count [3, 4], followed by an increase toward the end of the acute stage, which typically does not return to preinfection levels in the absence of antiretroviral therapy (ART) . Throughout the latent period, there is usually a steady decline in the numbers of CD4+ lymphocytes . A diagnosis of AIDS is made if the CD4+ T-cell count falls below 200 per μL. At these later stages of the disease, viral loads are starting to increase once more, leading to symptoms, opportunistic infections, and death.

A generalized graph of the relationship between HIV copies (viral load) and CD4+ T cell counts over the average course of untreated HIV infection. This is a file from the Wikimedia Commons. Commons is a freely licensed media file repository. Adapted from Fig. 1 in Pantaleo, G et al. (February 1993). “New concepts in the immunopathogenesis of human immunodeficiency virus infection”. New England Journal of Medicine 328 (5): 327–335. PMID 8093551
Early in the AIDS epidemic, HIV-associated neurocognitive disorders (HAND) were recognized as an important clinical manifestation of the disease [5–9]. Prior to the advent of combination antiretroviral therapy (cART) , 20–40 % of HIV-infected patients developed neurological symptoms ranging from minor cognitive impairment to severe dementia (known as HIV-associated dementia, HAD or AIDS dementia complex, ADC). ADC is characterized clinically by severe cognitive, motor, and behavioral abnormalities in the absence of opportunistic infections. Physical symptoms include headaches, generalized seizures, and ataxia. Cognitive symptoms comprise forgetfulness, slowing of thought processes, global dementia, confusion, and disorientation. Abnormalities in motor functions are characterized by unsteady gait, clumsiness, tremor, limb weakness, loss of coordination, and fine motor control. Finally, behavioral changes such as social withdrawal, lethargy, personality change, and hallucinations have been perceived in patients with ADC [7, 10].
Although cART has reduced AIDS-related mortality, HAND continues to be a major problem in patients with HIV. Antiretroviral drugs suppress virus burden in the plasma, CSF, and lymphoid tissue of HIV+ individuals; however, due to their restricted CNS penetration, these drugs exhibit only limited efficacy in the treatment of HAND. While the incidence of more severe neurological symptoms of ADC has been seen to decrease with cART, less severe versions of the disease persist among the infected population [11, 12]. The overall prevalence of HAND and associated morbidity remains high at approximately ~50 % [13–15]. It is projected that the prevalence of HAND is again increasing as life expectancy among patients increases. As the virus gains resistance to ART, the incidence of severe neurodegeneration is predicted to increase [13, 16, 17] and remains a significant independent risk factor for death [18, 19].
There is a consensus that HIV enters the central nervous system (CNS) during the early stages of infection primarily through virally infected/activated monocytes from the blood across the blood–brain barrier (BBB) by a so-called Trojan horse mechanism [20, 21]. Once in the brain, infected monocytes/macrophages and microglia release viral envelope proteins and cytokines, which in turn activate uninfected macrophages and microglia [22–29]. These activated macrophages and microglia release several neurotoxic substances, such as free radicals and glutamate, which lead to neuronal injury and apoptosis [30–34]. In summary, the virus does not directly infect neurons, yet neurons suffer injury due to indirect mechanisms mediated by host proinflammatory and viral proteins [26, 31, 35–37].
Post mortem neuropathological studies have demonstrated that the virus variably affects different regions of the brain. Pre-ART, abnormalities were predominantly found in the white matter and in subcortical structures, with relative sparing of the cortex [5]. Furthermore, higher levels of virus were found in the basal ganglia and hippocampus compared to cerebellar cortex and midfrontal cortical gray matter [38]. However, it appears that the anatomic distribution and temporal progression of neuropathologic changes differ across individuals, thus, it is important to consider both cortical and subcortical brain regions in studies of neuropathogenesis and treatment of HIV-related brain disease [39].
Potential Clinical Utility of MRS in HAND
Imaging has been used widely to evaluate the effect of HIV infection on the brain. Structural neuroimaging methods such as computerized tomography (CT) and magnetic resonance imaging (MRI) are important for the diagnosis of cerebral opportunistic infections such as Progressive Multifocal Leukoencephalopathy (PML) , Toxoplasmosis , Neurosyphilis , and Cytomegalovirus (CMV) Encephalitis. Morphological alterations detected by MRI include cortical atrophy at later stages of HIV-associated dementia (HAD). Furthermore, hyperintensities in the white matter (WM) and basal ganglia (BG) were observed on T2-weighted images of HIV+ patients. Diffusion tensor imaging (DTI) has been used to study the white matter structural integrity as it is typically more sensitive compared to T2-weighted imaging [40]. More recently, also quantitative morphological MRI has been used for assessing the severity of HAND [41–43]. However, structural imaging techniques were found to be limited in the evaluation of early cognitive changes [44–47]. Thus, neuroimaging techniques that provide functional and biochemical information have proven to be more useful in the evaluation of HAND [48].
SPECT and PET are sensitive to changes related to cerebral perfusion and metabolism. Specifically, PET has been used in patients with HAND and in animal models of neuroAIDS to study glucose metabolism, neuroinflammation , microglial activation, fibrillar amyloid, and dopamine transporters [49–53]. However, nuclear medicine techniques expose the subject to radiation and have therefore limited use for monitoring diseases progression or treatment effects when repeat measurements are needed [54].
Magnetic resonance spectroscopy (MRS) offers the unique ability to measure a wide range of cerebral metabolite levels in a noninvasive manner. Specifically, proton MRS (1H MRS) is one of the most informative methods employed in neuroAIDS research. MRS has been used in HIV-infected patients to understand the cognitive deficits and to establish biomarkers in HAND. To date, more than 80 original studies have been employed to evaluate the effects of HIV infection using MRS. The resonances seen in the brain by 1H MRS are typically low weight molecules related to neuronal injury/loss and inflammation. Specifically, N-Acetylaspartate (NAA) , an established marker for neuronal density and viability is decreased in patients with advanced neurocognitive symptoms and is of particular value for the in vivo assessment of neuronal integrity [55–58]. The earliest MRS studies of neuroAIDS, published in the early 1990s, reported decreases in NAA/Creatine (NAA/Cr) in patients with advanced neurocognitive symptoms and the decrease in NAA/Cr was found to be associated with disease severity [59–61].
Choline-containing compounds are related to cell/lipid membrane metabolism, thus an increase in Cho or Cho/Cr in HIV-infected patients is possibly associated with an immune response that includes cerebral inflammation or gliosis [61–63]. Myo-inositol (mI) is an organic osmolyte which is primarily located in glial cells, thus an increase in mI is associated with gliosis or inflammation [64]. Since HAND is considered a neuroinflammatory disease mI is especially important in the evaluation of glial response. In fact, MRS studies have found an elevation in mI/Cr as a result of HIV-associated brain injury [65–67].
The total creatine (tCr) resonance consists of the sum of creatine (Cr) and phosphocreatine (PCr), a high energy reservoir for the generation of ATP. Since Cr and PCr are in equilibrium the tCr peak is commonly assumed to remain stable in size despite bioenergetic abnormalities that occur with many pathologies or with age [68]. Consequently, the tCr resonance is often used as an internal standard and is commonly referred to as simply creatine (Cr). However, elevated Cr levels have been reported in the white matter in ART naïve HIV+ ADC stage 3 patients [69] but also in the WM in simian immunodeficiency virus (SIV) infected rhesus macaques during acute infection [70] and in an accelerated macaque model of neuroAIDS a few weeks after infection [71]. In the context of HAND, an elevation in Cr may reflect enhanced high-energy phosphate turnover in activated astrocytes and microglia [71].
Other metabolites of interest include glutamate (Glu), glutamine (Gln), and Glx, which is the sum of Gln and Glu. Glu is an amino acid acting as excitatory neurotransmitter and Gln is its precursor. Even though activated microglia release neurotoxic substances, such as free radicals and glutamate, which result in excitotoxicity and finally to apoptosis [30–34], decreases in Glx levels have been reported during early and chronic HIV infection [72–76]. The observed decrease in Glx may indicate neuronal dysfunction. Furthermore, levels of lactate (Lac), a marker of anaerobic glycolysis and inflammation and lipids, indicators of cell membrane turnover, have been assessed in HIV+ patients. The Lac/Cr ratio in the lenticular nuclei was significantly greater in HIV+ patients with moderate to severe impairment compared to seronegative controls. Moreover, the (Lipids + Lac)/Cr ratio was significantly elevated in both mild and moderate to severe HIV+ patients compared to seronegative controls [77].
Consequently, quantification of these abovementioned metabolite levels can assist in the diagnosis of the disease and measuring the severity of injury. Especially, in animal models where longitudinal scans are common, MRS has been proven extremely powerful to assess disease progression and response to treatment.
Technical Considerations for HAND
Echo Time
As with MRI, the choice of echo time (TE) can have an enormous effect on the appearance of the information obtained in a 1H MRS study. MR spectra obtained with shorter echo times (~30 ms) allow the detection of more metabolites including glutamate, glutamine, and myo-inositol. However, the baseline is typically more distorted due to increased lipids and macromolecular background signals. Spectra obtained with longer echo times (135–144 ms) show reduced number of metabolites. However, spectra are easier to process and analyze due to the relatively flat baseline. In addition, the lactate (1.3 ppm) doublet is inverted, therefore allowing better differentiation between Lac and lipids. On the other hand, longer TE attenuates the signal-to-noise ratio (SNR) and some metabolites, e.g., mI, Glu, and Gln, which can only be detected at short echo times, would be missed, thus short TE is typically preferred (Fig. 11.2).

MRS of white matter in a normal brain. (a) Long TE spectra have less baseline distortion and are easy to process and analyze but show fewer metabolites than short TE spectra. (b) Short TE demonstrates peaks attributable to more metabolites, including lipids and macromolecules, glutamine and glutamate, and myo-inositol
Single Voxel vs. MRSI
To measure MR spectra in vivo one has to be able to define the spatial origin of the detected signal. Basically, two methods exist to obtain the spatially localized metabolic information in vivo:
-
1.
Single voxel spectroscopy (SVS) uses selective excitation pulses to localize a voxel of typically 3–8 cm3. SVS has the advantage of higher SNR and typically shorter acquisition times. However, the accumulative scan time permits acquisition of only a few locations (Fig. 11.3).
Fig. 11.3 
Single-voxel proton magnetic resonance spectroscopy (SV-MRS). (a) Voxel locations the left frontal white matter region selected for quantitative spectroscopic analysis using SV MRS. (b) Sample spectra obtained in the frontal white matter in an HIV seronegative control and an HIV seropositive individual with mild dementia using SV-MRS. The NAA peak is decreased in the HIV dementia patient. Reprinted with permission from [80]
-
2.
Magnetic resonance spectroscopic imaging (MRSI) can be obtained in two or three dimensions. MRSI allows one to collect the spectral information from a volume consisting of many voxels with individual voxel sizes of typically 0.5–3 cm3 and most importantly makes it possible to cover large brain areas although spectra are typically characterized by lower SNR compared to SVS (Fig. 11.4).
Fig. 11.4 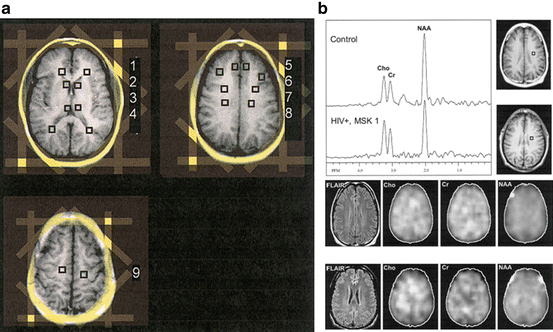
Magnetic resonance spectroscopic imaging (MRSI). (a) The MRSI locations include bilateral regions examining the (1) forceps minor, (2) caudate nucleus, (3) thalamus, (4) forceps major, (5) mesial frontal gray matter, (6) dorso-lateral gray matter, (7) frontal white matter, (8) centrum semiovale, and (9) subcortical white matter. The location of octagonal outer-volume saturation bands for lipid suppression is indicated on localizer images in yellow. (b) Sample spectra, T1-weighted and FLAIR MR images, and spectroscopic images for Cho, Cr, and NAA obtained in the frontal white matter in an HIV seronegative control and an HIV seropositive individual with mild dementia using MRSI. The NAA peak is decreased in the HIV dementia patient. FLAIR MRI scans show no abnormalities; no obvious spatial variations in the metabolic images are apparent between the HIV seronegative control and an HIV seropositive individual. Focal hyperintensity at the edges of the brain in the NAA images is due to lipid contamination from the peri-cranial tissues. Reprinted with permission from [80]


MRSI is preferred for clinical studies where it is indicated to obtain metabolic information of a large and heterogeneous lesions (e.g., in tumors), and when additional simultaneous spectral information from control regions needs to be obtained. However, HIV is considered a predominantly “global disease” and therefore, most studies have acquired SVS from 1 to 3 brain regions covering frontal white matter (FWM), basal ganglia (BG), and parietal cortex (PC) consistent with recommendations by the HIV MRS Consortium [78, 79]. In a study using both techniques, Sacktor et al. demonstrated that short TE SVS and long TE MRSI offer complementary roles in evaluating individuals with HIV dementia [80] (Fig. 11.4).
Field Strengths
MR spectroscopy inherently suffers from low SNR resulting in voxels with low spatial resolution or long acquisition times. The SNR can be improved by using higher magnetic field strengths. Physics predicts a linear increase in signal with field strength if T1 and T2 relaxation times, coil and system losses, and radiofrequency (RF) penetration effects do not change significantly. In addition, the increased chemical shift range at 3 and 7 T relative to 1.5 T results in greater separation of the resonance peaks, and consequently, allows for better quantification of those metabolites that generally overlap with others such as Glu, Gln, and mI. Thus, MRS is preferred at 3 T over 1.5 T. Figure 11.5 shows the spectra from the parietal cortex of a healthy rhesus macaque obtained at three different field strengths, 1.5, 3, and 7 T. The SNR increases at higher fields. In addition, improvements in spectral resolution are evident.

Comparison between MR spectra of rhesus macaques obtained at three different field strengths. (a) 1.5 T, (b) 3 T, (c) 7 T
Higher field strength may also result in better reproducibility. For example, in a study at Massachusetts General Hospital, four rhesus macaques underwent repeated (>4 times) SVS scans before infection to access reproducibility of the technique. Data at 1.5 T revealed coefficients of variance (CVs) within the same animals of 7–11 % for NAA/Cr, 13–16 % for Cho/Cr, and 8–12 % for mI/Cr in various brain regions including the FC, WM, and BG [81]. Four additional macaques were scanned a few years later at 3 T and revealed CVs of only 4 % for NAA/Cr, 2–8 % for Cho/Cr, and 3–7 % for mI/Cr in the FC, WM, and BG, respectively.
Ratios vs. Absolute Concentration
There are long-standing discussions on the best way to quantify MRS signals, specifically relative versus absolute quantification. The easiest method is to report metabolite ratios such as NAA/Cr. However, using Cr as an internal standard is often based on the assumption that the Cr concentration does not change during the disease process, which is sometimes, but not always true. Ratios are relatively easy to determine and are more reproducible than absolute concentrations. However, when a change is detected, it may not be possible to determine whether it is due to a change in the numerator, denominator, or both.
Changes in creatine have been reported in patients and an animal model of HAND [69–71]; increasing creatine levels may mask increases in Cho and mI. Absolute quantification of brain metabolites by MRS is more difficult to obtain. They are generally expressed in units of mmol/kg. Methods used for absolute quantification include: (a) phantom replacement techniques [82–84], (b) using the unsuppressed water signal as a reference [85–87], or (c) the use of an external reference [88, 89]. Most studies on HIV-infected patients that reported metabolite concentrations have used the water signal as reference [85].
Overview of MRS in Animal Models of HAND
In the previous chapter, we have established that in vivo MRS is a powerful tool in studying the effect of HIV infection on the brain. Due to its noninvasive nature, in vivo MRS is suitable for both cross-sectional and longitudinal studies. However, studies of HIV-infected individuals present difficulties in interpretation: the time of infection is often unknown; a number of individuals are infected with more than one strain of HIV; alcohol and substance abuse result in comorbidities. Animal models allow for greater flexibility to explore and address the questions concerning HIV neuropathogenesis in a controlled manner. Furthermore, animal models allow access to subsequent post mortem neuropathology studies helping define relationships between in vivo biomarkers of neuronal function and inflammation by MRS and postmortem histopathology.
Crucial insights into neuroAIDS have emerged from several excellent animal models, including a transgenic HIV mouse model that expresses viral surface protein gp120 in the brain [90]; injection of HIV-1-infected macrophages into the brains of severe combined immunodeficient (SCID) mice [91], which results in the development of pathologic hallmarks for HIV encephalitis (HIVE) and neuroinflammation [92–94]; a feline immunodeficiency virus (FIV) infection of cats [95–102]; and the SIV macaque models [103–107].
MRS Studies of Rodent Models
Rodent models have been applied in elucidating neuropathogenic pathways associated with HIV infection, such as the migration and distribution of monocytes in brain tissue and the neurotoxic effects of specific viral proteins [108–110]. In a SCID mouse model , HIVE was induced focally into the striatum by the unilateral injection of HIV-1ADA-infected human monocyte-derived macrophages. Histology revealed focal giant cell encephalitis, with reactive astrocytes, microgliosis, and neuronal dropout. Metabolite concentrations using in vivo MRSI at 7 T were measured 7 days after injection in infected mice compared to sham-operated and unmanipulated mice. These studies showed significant decreases in NAA in both the ipsilateral and contralateral site of injection in HIVE mice compared to sham-operated mice indicating that a highly focal encephalitis can produce global deficits for neuronal function and metabolism [111].
MRS Studies of FIV Models
Like HIV, FIV is known to infect CD4+ T lymphocytes causing immunosuppression, encephalopathy, and neurobehavioral deficits [112, 113]. The neurological abnormalities are typically more profound when FIV is induced during the developmental period of brain maturation, making this an ideal model for understanding neurologic complications observed in children infected with HIV [102, 114]. Cats infected with Maryland isolate (FIV-MD) have demonstrated reductions in NAA and NAA/Cho between 8 and 14 months of infection. FIV-infected cats also exhibited a higher proportion of quantitative electroencephalographic relative slow wave activity that correlated to lower NAA in the frontal cortex [102]. Furthermore, cats infected with high viral titers of V1CSF (a neurovirulent FIV strain which was derived from the CSF of a cat with encephalopathy) had increased Cho/Cr and reduced NAA/Cr in the frontal cortex compared to those that received low titers indicating that infectious titers in the brain during the early stages of infection determine the severity of neurovirulence [115]. Like many HIV studies, FIV infection also resulted in reduction in Glu and Glu/Cr in the cortex and white matter [115].
MRS Studies of SIV Macaque Models
The SIV macaque models are arguably the most comparable and informative of CNS disease progression [107]. The SIV-infected rhesus macaque shares very similar pathology with HIV-infected human patients, including the development of AIDS, disease of the CNS, and cognitive or behavioral deficits [103–107]. SIV is the closest known phylogenetic relative of HIV and also infects CD4+ macrophages, lymphocytes, and microglia. Its acute infection parallels that of HIV, with a very rapid change in the blood viral load observed during the first month of infection, usually around 11–12 days after infection [116–118]. Neuroinvasion occurs early in infection for both HIV and SIV [119], and their neuropathology includes gliosis, perivascular cuffing, and neuronal injury [104, 106, 120, 121]. Furthermore, antiretroviral therapy has been successfully applied to SIV-infected macaques [122, 123].
Traditional SIV Model
Acute/Early SIV Infection
Acute HIV infection is the time period from viral infection through seroconversion. It is characterized by high peak in viremia and impairment/destruction of CD4+ T lymphocytes [2, 124, 125] similar to events that are manifest in chronic infection when HIV-related dementia is more likely to occur [126]. In the SIV macaque model peak viremia is typically observed within 2 weeks after inoculation with SIVmac251 (Fig. 11.6a). We found that plasma SIV RNA peaked at 11–12 days postinfection (dpi) at ~108 copies/mL. Within days after the peak, the viral load reverted to lower levels ~106 copies/mL due to the immunologic response by the macaques. In these studies nearly all macaques had transient reductions in NAA/Cr by single voxel MRS within the first 2 weeks of infection in the frontal cortex (Fig. 11.6b) but not in the basal ganglia or white matter. Pathology confirmed neuronal dysfunction detected by significant decreases in synaptophysin, a marker for synaptic integrity, while the numbers of neurons were not affected by the virus suggesting that during early infection reversible neuronal injury occurs, but not neuronal loss [127].

Acute changes in the SIV macaque model of NeuroAIDS. Based on [118] and [70]. (a) Plasma viral loads: peak viremia can be detected at 11 days postinfection (dpi). (b) Metabolic changes in the frontal cortex: acutely SIV-infected macaques showed a transient decrease in NAA/Cr at 13 dpi; elevated mI/Cr at 11, 13, and 25 dpi; and a dynamic response in Cho/Cr with increases at 11 dpi and decreases below baseline at 27 dpi
Myo-inositol levels were significantly elevated in both the frontal cortex and white matter regions, suggesting stimulation of glial metabolism within the first month of infection [70]. In addition, dynamic changes in choline were observed in the same regions. At the time of peak viremia, both choline and Cho/Cr were significantly elevated; however, between 2 and 4 weeks postinfection (wpi), Cho/Cr ratios underwent a large reduction, resulting in levels below preinfection [118] (Fig. 11.6b). An increase in Cho reflects altered membrane metabolism [128] and in the context of neuroAIDS may be reflective of cerebral inflammation, cell proliferation, recruitment, microgliosis, and/or astrocytosis [61, 63]. Evidence of the association with astrocytosis is given by our observation that during the acute phase changes in Cho/Cr mirror the changes in the astrocyte response by glial fibrillary acidic protein (GFAP) [129]. Decreases in Cho below baseline have rarely been reported in diseases not involving necrosis and still warrant further investigations.
Furthermore, the SIV Model is characterized by elevations in creatine concentrations in the white matter during the acute phase of infection. During that stage, the virus enters the brain through infected monocytes [31], resulting in glial-cell activation and proliferation . These processes may manifest themselves in high tissue metabolism, which would explain an increase in Cr. Energy change as a function of enhanced glial activation is supported by the finding that the greatest increases in mI and Cho are in the same regions as the highest increase in Cr [70].
Chronic SIV Infection
Eight rhesus macaques were infected with SIVmac251 and serially imaged with MRI and 1H MRS to terminal AIDS or the endpoint of the study at 2 years. Due to the low incidence of SIV encephalitis (SIVE) , possible diverse viral mutation and the differences in host factors the metabolic response over the time course of infection was variable. However, comparing the same animals we found a positive correlation between frontal cortex Cho/Cr and plasma viral load and a negative correlation between basal ganglia NAA/Cr and plasma viral load [130].
Rapid SIV Model
Untreated SIV+/CD8− Animals
Because of its parallels with HIV pathogenesis, the traditional SIV macaque model is hindered by the low rate of development of SIVE and the long time period for its evolution. Only approximately 25 % of infected macaques develop encephalitis and progression to terminal AIDS may take several years [107, 131]. These factors make it difficult for use in testing potential drug therapies. Therefore, attention has focused on two rapidly progressing SIV macaque models. One model employs pig-tailed macaques that are coinoculated with two SIV strains (SIV/17E-Fr and SIV/DeltaB670), which accelerates SIV CNS disease, producing SIVE in over 90 % of these animals within 3 months [132–135]. The second model retains the use of the SIV-infected rhesus macaques , but uses a monoclonal antibody to deplete the animal of CD8+ lymphocytes [136, 137]. In this model, 80 % of persistently CD8-depleted animals develop SIVE, with a course of progression to terminal AIDS within 12 weeks [33, 123, 138]. Modifications in applying the antibody have resulted in >90 % of macaques becoming persistently CD8 depleted with nearly all of them developing SIVE. This model also produces profound neuronal injury detectable within weeks by in vivo 1H MRS [123]. Thus, this accelerated model of neuroAIDS in combination with MRS provides an exceptional opportunity to efficiently explore potentially useful drug therapies that can control or reverse neuronal injury and damage.
In the SIV+/CD8− model plasma viral RNA was detectable 6 days after infection with ~107 copies Eq./mL and approached a plateau by 4 weeks after infection at ~5 × 108 copies Eq./mL (Fig. 11.7a). CSF viral loads were found to be approximately 3 orders of magnitude lower than plasma. SIV+/CD8− animals were euthanized at 4, 6, and 8 weeks after infection and the amount of viral RNA in the frontal cortex was found to be ~106 copies Eq./g at all three time points (Fig. 11.7b).

(a and b) Viral RNA and neuronal injury following SIV infection and CD8 depletion. Based on [148] and [71]. (a) Viral RNA was detectable in plasma 6 days after infection (~107 copies Eq./mL) and the viral load approaches a plateau by 4 weeks after infection. The mean plasma viral load in infected animals was 4.9 × 108 copies Eq./mL 8 weeks after infection. CSF viral loads were found to be approximately 3 orders of magnitude lower than plasma in all animals. (b) Animals were euthanized at 4, 6, and 8 weeks after infection. The amount of viral RNA in the frontal cortex of untreated animals was 7.5 × 105, 9.2 × 105, and 3.0 × 105 copies Eq./g at 4, 6, and 8 weeks after infection, respectively. (c) The neuronal marker NAA/Cr steadily declined following SIV infection, reaching decreases as low as 20 % below baseline by 8 weeks postinfection (wpi) in the parietal cortex (PC). (d) Neuronal injury is confirmed by decreases in Microtubule-associated protein 2 (MAP2), a marker of dendritic integrity
The neuronal marker NAA/Cr steadily declined following SIV infection and CD8 depletion, reaching levels as low as 20 % below baseline by 8 wpi in untreated animals (Fig. 11.7c). The decrease in NAA/Cr following infection is due to both decreases in neuronal NAA and increases in creatine, which most likely reflects the cumulative effects of altered metabolic states of neurons and glial cells, respectively (for further discussion, see Ratai et al. [71]). Neuronal injury was confirmed by decreases in microtubule-associated protein 2 (MAP2) , a marker of dendritic integrity (Fig. 11.7d).
CART and Minocycline -Treated SIV+/CD8− Animals
To gain further insights into the neuropathogenesis of AIDS we modulated disease progression with treatments. In total, 11 SIV+/CD8− rhesus macaques were treated. Seven received minocycline (MN), an antibiotic, which easily crosses the BBB, and has been found to have anti-inflammatory, neuroprotective, possibly antiviral effects [139–146]. Four of the seven MN-treated animals had persistent depletion of CD8+ lymphocytes. However, three of the MN-treated animals had partial recovery of CD8+ lymphocytes. This latter cohort was distinct from the long-term depleted cohorts in respect to its 1 order of magnitude lower viral loads and was thus grouped into a separate cohort of MN-treated short-term depleted animals. All seven animals were treated at 4 wpi for 4 weeks.
Four additional animals were treated with combination antiretroviral therapy (cART) consisting of 9-R-2-Phosphonomethoxypropyl adenine (PMPA), 5-Fluoro-1-[(2R,5S )-2-(hydroxymethyl)-[1,3]oxathiolan-5-yl]cytosine (FTC), and 2′-3′-didehydro-2′-3′-dideoxythymidine (Stavudine, Zerit®) starting at 6 wpi for 6 weeks. Both treatments, MN and cART, resulted in a moderate decline of plasma, CSF, and brain viral levels [147, 148]. During minocycline as well as cART treatment, a significant reduction of circulating activated CD14+CD16+ monocytes was observed. Without treatment, SIV progression is characterized by the expansion of these monocytes in the periphery, which play a major role in trafficking virus across the BBB into the brain [149]. More importantly, the decline in NAA/Cr was arrested with MN and cART (Fig. 11.8, here shown for the parietal cortex, NAA/Cr) resulting in higher NAA/Cr levels when compared to untreated animals sacrificed at 8 wpi. Animals that were MN treated and had partial immune reconstitution of the CD8+ T cell population had the most complete recovery. Thus, we hypothesize that possibly the best strategy to treat neuroAIDS is by the use of combination therapies targeting pathogenic factors in both the periphery and CNS [148].

Amelioration of neuronal injury with combination antiretroviral therapy (cART) and minocycline. Based on [147, 148]. NAA/Cr steadily declined following SIV infection in all animals, reaching levels as low as 20 % below baseline by 8 wpi in untreated animals (black squares). The decline in NAA/Cr was arrested with the cART (green diamonds) and minocycline treatment (red triangles) resulting in higher NAA/Cr levels when compared to untreated animals sacrificed at 8 wpi. Animals that were MN treated and had partial immune reconstitution of the CD8+ T cell population had the most complete recovery (orange triangles)
Overview of Clinical MRS in HAND
Early HIV Infection
Until recently the effect of HIV to the brain during the early stages of HIV infection (less than 1 year of seroconversion) remained relatively unknown. However, a few studies show that soon after infection brain metabolism is affected by the virus as detected by in vivo MRS. In a study by Lentz et al., eight subjects were examined by SVS MRS within 3 months of their evolving Western blot and subsequently at 2 and 6 months after their initial scan. Initially, a decrease in NAA and Glx were observed in the frontal cortex but not in the white matter semiovale compared to controls suggesting that frontal cortical gray matter to be more susceptible to early neuronal injury [76] (Fig. 11.9). Further longitudinal evaluation of this cohort showed increases in choline metabolism in the FC and WM indicative of lipid membrane turnover and glial metabolism [150]. These findings were confirmed by Sailasuta and Valcour who found increases in Cho/Cr in the WM and BG, respectively, in HIV+ patients being scanned during the first year of infection (median 2 weeks after infection) compared with HIV− controls [151, 152]. Recently, MRS markers have been correlated with markers of inflammation and neuronal injury suggesting early neuronal injury in a subset of participants with primary HIV infection through mechanisms involving central nervous system inflammation [153, 154].

Metabolic changes in NAA and Glx in the frontal cortex and white matter centrum semiovale during early and chronic infection. In the frontal cortical gray matter N-acetylaspartate (NAA) (a) and glutamate + glutamine (Glx) (b) levels are reduced in subjects during the first few months of infection, similar to subjects with chronic infection who are neurologically asymptomatic. Within the white matter of the centrum semiovale, NAA (c) was found to be reduced in chronically infected patients with HIV, but not in those during early infection. No changes from control levels for Glx in the white matter were observed (d). Reprinted with permission from [76]
Chronic HIV Infection
Observed MRS changes in chronically HIV-infected patients typically show reduction in NAA or NAA/Cr and elevations in Cho and mI or their respective ratios over Cr. The first case report of MRS studies in HIV-infected patients appeared in 1990 and discussed the strength of MRS in providing a marker of neuronal loss in patients with normal MRIs [155]. The first larger study of MRS in more than 100 HIV+ patients showed decreased NAA/Cr and elevated Cho/Cr [61]. Subsequently, it was found that Cho/Cr was elevated even in asymptomatic HIV+ patients and in patients with AIDS dementia complex while NAA/Cr was only decreased in symptomatic ADC patients [156] confirming that brain choline-containing compounds are elevated in HIV+ patients before the onset of ADC [157]. The decrease in NAA seems to correlate with the degree of neuronal dysfunction. Decreases in NAA/Cr of ~10 % have been observed in HIV+ patients with minor cognitive or motor dysfunction [67], while reductions of 15–28 % been reported in HIV+ patients with severe cognitive impairment or AIDS dementia complex compared to HIV− controls [63, 156, 158, 159].
Elevated mI or mI/Cr has been reported at various stages of HIV dementia and has been shown to increase with dementia severity [54, 65–67, 160]. Elevated Cr levels have been reported in the frontal white matter of chronic HIV patients [69]. Further changes in chronically infected patients include decreases in glutamate or Glx in the frontal white matter [72, 73]. Lower brain glutamate levels are associated with poorer cognitive deficits in HIV patients [74, 75].
Post Post-cART
As previously stated with the advent of ART, the incidence of severe neurological symptoms of HAD has been seen to decrease; however, less severe versions of the disease persist among the infected population [11, 12]. The characteristic involvement of the basal ganglia in pre-ART specimens is less commonly seen in post-ART specimens, which display neuroinflammation in the hippocampus and in adjacent parts of the entorhinal and temporal cortices [15, 161, 162].
Longitudinal studies in HIV+ patients with mild dementia showed improvement of CD4 counts and HIV Dementia Scale score when treated with cART. The initially increased Cho/Cr levels reversed in the midfrontal cortex and in the basal ganglia and the initially elevated mI/Cr and mI levels in the basal ganglia also decreased [163]. Another study that followed HIV+ patients before and after cART showed no significant improvement in Cho or mI after 3 months of treatment, but only after 6–9 months [54].
Clinical Trials Using MRS
A few clinical trials have included MRS as one of the outcome measures for neuroprotective drugs in HIV+ patients. In a Phase II randomized, double-blind, placebo-controlled, multicenter trial within the Adult AIDS Clinical Trials Group, 140 HIV-infected patients with mild to severe ADC receiving stable antiretroviral therapy were enrolled to assess the safety and efficacy of memantine, an N-methyl-d-aspartate receptor antagonist as treatment of HIV-associated cognitive impairment. While no significant differences in cognitive performance (neuropsychological z score (NPZ)-8) were shown 16 weeks later, the MRS data suggested that memantine may ameliorate neuronal metabolism, an important step to stabilizing or preventing neuronal injury. These results underscore the need for longer studies to assess the full potential of neuroprotective agents [164]. On the other hand, a study that assessed the effectiveness of selegiline transdermal system (STS) in reversing HIV-induced neurologic injury revealed no effect on either MRS metabolites, oxidative stress, or NPZ-8 scores [165].
Comorbidities
HAND is exacerbated by several comorbidities . Numerous studies have shown that neuronal injury is enhanced by both licit (alcohol) and illicit drugs (methamphetamine) [43, 166, 167]. As the HIV-infected population ages, other age-related neurological disorders will likely affect HIV+ patients. Several studies started to investigate the relationship between HIV and age [79].
Of note, increasing evidence suggests that certain antiretrovirals may be neurotoxic [168]. Nucleoside reverse transcriptase inhibitors (NRTIs) suppress HIV replication but are often associated with mitochondrial toxicity. Chronically infected HIV+ patients on long-term cART regimens that included NRTIs such as didanosine and/or stavudine revealed significant reductions in NAA in the FWM compared with HIV− controls while HIV+ individuals receiving alternative cART regimens that did not include NRTIs exhibited only intermediate decreases in NAA [169].
HIV-Infected Children
Overall, fewer MRS studies have been performed on the brains of HIV-infected children. Most HIV+ children under the age of 13 are infected during pregnancy, childbirth, or breastfeeding. A study by Keller et al. showed that while healthy children showed an increase in NAA with age in the frontal WM and hippocampus, HIV+ children did not show the age-related increase. Furthermore, HIV+ children with high plasma viral loads had lower Cho, mI, and Cr in the basal ganglia but higher Cho in the midfrontal gray matter compared to children with lower viral loads [170].
A study by Mbugua et al. imaged a cohort of 5-year-old HIV+ children in South Africa and found lower CD4/CD8 ratio in early infancy (mean age = 8 ± 2 weeks old) to be significantly associated with lower NAA and Cho levels in the basal ganglia [171]. Furthermore, a longitudinal study by Holmes et al. compared HIV uninfected, exposed (HEU) and HIV unexposed, uninfected (HUU) young children; they found a significant increase in NAA levels with age in HUU only as well as significantly higher mean Cho levels in HEU children at age 7 years [172].
Summary and Future Perspectives
In summary, MRS offers a valuable imaging technique to assess disease status and response to therapeutics in patients with HAND. Since MRS changes are sensitive to early subtle neuropathogenic changes MRS studies certainly add information to conventional MRI exams. In animal models MRS has successfully been utilized to understand the neuropathogenesis of the disease. Specifically, the combination of MRS with immunology, virology, and pathology in animal models has led to a greater understanding of the inflammatory and neuronal events caused by the virus.
To date most human studies have been cross sectional; however, MRS is especially powerful when applied longitudinally to monitor individual patients. Thus, the next step in neuroimaging studies should include standardized MRS protocols in multicenter clinical trials to evaluate potential treatments for patients suffering from HAND. Significant effort has been made by the AIDS Clinical Trial Group (ACTG) using the same imaging paradigm at multiple sites [154].
Furthermore, most studies performed single voxel spectroscopy on either 1.5 or 3 T. However, more novel MRS techniques have been developed to more accurately measure metabolites that show overlapping peaks in the regular one-dimensional (1D) MRS spectra. For instance, using 2D localized chemical shift correlated spectroscopy (L-COSY) enables converting a crowded, overlapping 1D MR spectrum to a better resolved 2D spectrum through the addition of a spectral dimension [173] and allows for better detection of metabolites at lower concentrations and delineation of the components of otherwise overlapping peaks such as glutamate/glutamine, myo-inositol, aspartate (Asp), gamma-aminobutyrate (GABA), taurine, glutathione (GSH), threonine, and macromolecules [174].
In addition, GABA and GSH can specifically be detected with so-called editing techniques either using double-quantum coherence (DQC) [175, 176] or difference editing techniques, e.g., MEGA PRESS [177, 178]. Specifically, the quantification of GSH levels may be of pertinence in HIV-infected patients. HIV is associated with substantial oxidative stress [179]. Glutathione is thought to be an extremely important antioxidant for HIV-infected patients, because it appears to interfere with HIV’s entry into its target cells [180].
Greater signal-to-noise ratio, spatial and spectral resolution can also be gained from higher field strengths. 7 T MRS in combination with LCModel [181] allows for the quantification of 17 metabolites [182]. 7 T MRS has successfully been employed in the CD8-depleted macaque model of neuroAIDS [183]. In addition, Gonen et al., have successfully implemented a 3D MRSI sequence with 0.05 cm3 isotropic spatial resolution at 7 T [184].
Other technical advances include motion- corrected MR spectroscopy exams. For children between 2 months and 7 years of age, motion artifact and patient cooperation often limit the feasibility of MRI and MRS [185]. Using image-based navigators, it is possible to correct motion in structural imaging, single-voxel and multivoxel spectroscopy prospectively [186–192].
References
Worldwide HIV AIDS Statistics (2014) http://www.avert.org/worldwide-hiv-aids-statistics.htm
Daar ES, Moudgil T, Meyer RD, Ho DD (1991) Transient high levels of viremia in patients with primary human immunodeficiency virus type 1 infection. N Engl J Med 324(14):961–964
Gupta KK (1993) Acute immunosuppression with HIV seroconversion. N Engl J Med 328(4):288–289
Vento S, Di Perri G, Cruciani M, Garofano T, Concia E, Bassetti D (1993) Rapid decline of CD4+ cells after IFN alpha treatment in HIV-1 infection. Lancet 341(8850):958–959
Navia BA, Cho ES, Petito CK, Price RW (1986) The AIDS dementia complex: II. Neuropathology. Ann Neurol 19(6):525–535
Navia BA, Jordan BD, Price RW (1986) The AIDS dementia complex: I. Clinical features. Ann Neurol 19(6):517–524
Price RW, Brew B, Sidtis J, Rosenblum M, Scheck AC, Cleary P (1988) The brain in AIDS: central nervous system HIV-1 infection and AIDS dementia complex. Science 239(4840):586–592
Gray F, Hurtrel M, Hurtrel B (1993) Early central nervous system changes in human immunodeficiency virus (HIV)-infection. Neuropathol Appl Neurobiol 19(1):3–9
Vaughan JT, Hetherington HP, Otu JO, Pan JW, Pohost GM (1994) High frequency volume coils for clinical NMR imaging and spectroscopy. Magn Reson Med 32(2):206–218
Price RW, Brew BJ (1988) The AIDS dementia complex. J Infect Dis 158(5):1079–1083
Brodt HR, Kamps BS, Gute P, Knupp B, Staszewski S, Helm EB (1997) Changing incidence of AIDS-defining illnesses in the era of antiretroviral combination therapy. AIDS 11(14):1731–1738
McArthur JC (2004) HIV dementia: an evolving disease. J Neuroimmunol 157(1–2):3–10
Nath A, Schiess N, Venkatesan A, Rumbaugh J, Sacktor N, McArthur J (2008) Evolution of HIV dementia with HIV infection. Int Rev Psychiatry 20(1):25–31
Heaton RK, Clifford DB, Franklin DR Jr, Woods SP, Ake C, Vaida F, Ellis RJ, Letendre SL, Marcotte TD, Atkinson JH, Rivera-Mindt M, Vigil OR, Taylor MJ, Collier AC, Marra CM, Gelman BB, McArthur JC, Morgello S, Simpson DM, McCutchan JA, Abramson I, Gamst A, Fennema-Notestine C, Jernigan TL, Wong J, Grant I (2010) HIV-associated neurocognitive disorders persist in the era of potent antiretroviral therapy: CHARTER Study. Neurology 75(23):2087–2096
Gannon P, Khan MZ, Kolson DL (2013) Current understanding of HIV-associated neurocognitive disorders pathogenesis. Curr Opin Neurol 24(3):275–283
Sacktor N (2002) The epidemiology of human immunodeficiency virus-associated neurological disease in the era of highly active antiretroviral therapy. J Neurovirol 8(Suppl 2):115–121
Sacktor N, McDermott MP, Marder K, Schifitto G, Selnes OA, McArthur JC, Stern Y, Albert S, Palumbo D, Kieburtz K, De Marcaida JA, Cohen B, Epstein L (2002) HIV-associated cognitive impairment before and after the advent of combination therapy. J Neurovirol 8(2):136–142
Ellis RJ, Deutsch R, Heaton RK, Marcotte TD, McCutchan JA, Nelson JA, Abramson I, Thal LJ, Atkinson JH, Wallace MR, Grant I (1997) Neurocognitive impairment is an independent risk factor for death in HIV infection. San Diego HIV Neurobehavioral Research Center Group. Arch Neurol 54(4):416–424
Liner KJ II, Hall CD, Robertson KR (2008) Effects of antiretroviral therapy on cognitive impairment. Curr HIV/AIDS Rep 5(2):64–71
Peluso R, Haase A, Stowring L, Edwards M, Ventura P (1985) A Trojan Horse mechanism for the spread of visna virus in monocytes. Virology 147(1):231–236
Hickey WF, Hsu BL, Kimura H (1991) T-lymphocyte entry into the central nervous system. J Neurosci Res 28(2):254–260
Bonwetsch R, Croul S, Richardson MW, Lorenzana C, Valle LD, Sverstiuk AE, Amini S, Morgello S, Khalili K, Rappaport J (1999) Role of HIV-1 Tat and CC chemokine MIP-1alpha in the pathogenesis of HIV associated central nervous system disorders. J Neurovirol 5(6):685–694
Catani MV, Corasaniti MT, Navarra M, Nistico G, Finazzi-Agro A, Melino G (2000) gp120 induces cell death in human neuroblastoma cells through the CXCR4 and CCR5 chemokine receptors. J Neurochem 74(6):2373–2379
Cheng J, Nath A, Knudsen B, Hochman S, Geiger JD, Ma M, Magnuson DS (1998) Neuronal excitatory properties of human immunodeficiency virus type 1 Tat protein. Neuroscience 82(1):97–106
Meucci O, Fatatis A, Simen AA, Bushell TJ, Gray PW, Miller RJ (1998) Chemokines regulate hippocampal neuronal signaling and gp120 neurotoxicity. Proc Natl Acad Sci U S A 95(24):14500–14505
Nath A (2002) Human immunodeficiency virus (HIV) proteins in neuropathogenesis of HIV dementia. J Infect Dis 186(Suppl 2):S193–S198
Nath A, Psooy K, Martin C, Knudsen B, Magnuson DS, Haughey N, Geiger JD (1996) Identification of a human immunodeficiency virus type 1 Tat epitope that is neuroexcitatory and neurotoxic. J Virol 70(3):1475–1480
Ohagen A, Ghosh S, He J, Huang K, Chen Y, Yuan M, Osathanondh R, Gartner S, Shi B, Shaw G, Gabuzda D (1999) Apoptosis induced by infection of primary brain cultures with diverse human immunodeficiency virus type 1 isolates: evidence for a role of the envelope. J Virol 73(2):897–906
D’Aversa TG, Eugenin EA, Berman JW (2005) NeuroAIDS: contributions of the human immunodeficiency virus-1 proteins Tat and gp120 as well as CD40 to microglial activation. J Neurosci Res 81(3):436–446
Crowe S, Zhu T, Muller WA (2003) The contribution of monocyte infection and trafficking to viral persistence, and maintenance of the viral reservoir in HIV infection. J Leukoc Biol 74(5):635–641
Gartner S (2000) HIV infection and dementia. Science 287(5453):602–604
Persidsky Y, Gendelman HE (2003) Mononuclear phagocyte immunity and the neuropathogenesis of HIV-1 infection. J Leukoc Biol 74(5):691–701
Williams KC, Hickey WF (2002) Central nervous system damage, monocytes and macrophages, and neurological disorders in AIDS. Annu Rev Neurosci 25:537–562
Roy S, Wainberg MA (1988) Role of the mononuclear phagocyte system in the development of acquired immunodeficiency syndrome (AIDS). J Leukoc Biol 43(1):91–97
Kaul M, Garden GA, Lipton SA (2001) Pathways to neuronal injury and apoptosis in HIV-associated dementia. Nature 410(6831):988–994
Kaul M, Zheng J, Okamoto S, Gendelman HE, Lipton SA (2005) HIV-1 infection and AIDS: consequences for the central nervous system. Cell Death Differ 12(Suppl 1):878–892
Kraft-Terry SD, Buch SJ, Fox HS, Gendelman HE (2009) A coat of many colors: neuroimmune crosstalk in human immunodeficiency virus infection. Neuron 64(1):133–145
Wiley CA, Soontornniyomkij V, Radhakrishnan L, Masliah E, Mellors J, Hermann SA, Dailey P, Achim CL (1998) Distribution of brain HIV load in AIDS. Brain Pathol 8(2):277–284
Moore DJ, Masliah E, Rippeth JD, Gonzalez R, Carey CL, Cherner M, Ellis RJ, Achim CL, Marcotte TD, Heaton RK, Grant I (2006) Cortical and subcortical neurodegeneration is associated with HIV neurocognitive impairment. AIDS 20(6):879–887
Filippi CG, Ulug AM, Ryan E, Ferrando SJ, van Gorp W (2001) Diffusion tensor imaging of patients with HIV and normal-appearing white matter on MR images of the brain. AJNR Am J Neuroradiol 22(2):277–283
Bonnet F, Amieva H, Marquant F, Bernard C, Bruyand M, Dauchy FA, Mercie P, Greib C, Richert L, Neau D, Catheline G, Dehail P, Dabis F, Morlat P, Dartigues JF, Chene G (2013) Cognitive disorders in HIV-infected patients: are they HIV-related? AIDS 27(3):391–400
Ances BM, Ortega M, Vaida F, Heaps J, Paul R (2012) Independent effects of HIV, aging, and HAART on brain volumetric measures. J Acquir Immune Defic Syndr 59(5):469–477
Pfefferbaum A, Rosenbloom MJ, Sassoon SA, Kemper CA, Deresinski S, Rohlfing T, Sullivan EV (2012) Regional brain structural dysmorphology in human immunodeficiency virus infection: effects of acquired immune deficiency syndrome, alcoholism, and age. Biol Psychiatry 72(5):361–370
Aylward EH, Henderer JD, McArthur JC, Brettschneider PD, Harris GJ, Barta PE, Pearlson GD (1993) Reduced basal ganglia volume in HIV-1-associated dementia: results from quantitative neuroimaging. Neurology 43(10):2099–2104
Broderick DF, Wippold FJ II, Clifford DB, Kido D, Wilson BS (1993) White matter lesions and cerebral atrophy on MR images in patients with and without AIDS dementia complex. AJR Am J Roentgenol 161(1):177–181
Dooneief G, Bello J, Todak G, Mun IK, Marder K, Malouf R, Gorman J, Hilal S, Stern Y, Mayeux R (1992) A prospective controlled study of magnetic resonance imaging of the brain in gay men and parenteral drug users with human immunodeficiency virus infection. Arch Neurol 49(1):38–43
McArthur JC, Kumar AJ, Johnson DW, Selnes OA, Becker JT, Herman C, Cohen BA, Saah A (1990) Incidental white matter hyperintensities on magnetic resonance imaging in HIV-1 infection. Multicenter AIDS Cohort Study. J Acquir Immune Defic Syndr 3(3):252–259
Boska MD, Mosley RL, Nawab M, Nelson JA, Zelivyanskaya M, Poluektova L, Uberti M, Dou H, Lewis TB, Gendelman HE (2004) Advances in neuroimaging for HIV-1 associated neurological dysfunction: clues to the diagnosis, pathogenesis and therapeutic monitoring. Curr HIV Res 2(1):61–78
Hammoud DA, Endres CJ, Chander AR, Guilarte TR, Wong DF, Sacktor NC, McArthur JC, Pomper MG (2005) Imaging glial cell activation with [11C]-R-PK11195 in patients with AIDS. J Neurovirol 11(4):346–355
Wiley CA, Lopresti BJ, Becker JT, Boada F, Lopez OL, Mellors J, Meltzer CC, Wisniewski SR, Mathis CA (2006) Positron emission tomography imaging of peripheral benzodiazepine receptor binding in human immunodeficiency virus-infected subjects with and without cognitive impairment. J Neurovirol 12(4):262–271
Andersen AB, Law I, Krabbe KS, Bruunsgaard H, Ostrowski SR, Ullum H, Hojgaard L, Lebech A, Gerstoft J, Kjaer A (2010) Cerebral FDG-PET scanning abnormalities in optimally treated HIV patients. J Neuroinflammation 7:13
Chang L, Wang GJ, Volkow ND, Ernst T, Telang F, Logan J, Fowler JS (2008) Decreased brain dopamine transporters are related to cognitive deficits in HIV patients with or without cocaine abuse. Neuroimage 42(2):869–878
Ances BM, Christensen JJ, Teshome M, Taylor J, Xiong C, Aldea P, Fagan AM, Holtzman DM, Morris JC, Mintun MA, Clifford DB (2010) Cognitively unimpaired HIV-positive subjects do not have increased 11C-PiB: a case-control study. Neurology 75(2):111–115
Chang L, Feger U, Ernst TM (2011) Bioimaging. In: The neurology of AIDS, 3rd edn. Oxford University Press, Oxford
Birken DL, Oldendorf WH (1989) N-Acetyl-L-aspartic acid: a literature review of a compound prominent in 1H-NMR spectroscopic studies of brain. Neurosci Biobehav Rev 13(1):23–31
Simmons ML, Frondoza CG, Coyle JT (1991) Immunocytochemical localization of N-acetyl-aspartate with monoclonal antibodies. Neuroscience 45(1):37–45
Moffett JR, Namboodiri MA, Cangro CB, Neale JH (1991) Immunohistochemical localization of N-acetylaspartate in rat brain. Neuroreport 2(3):131–134
Urenjak J, Williams SR, Gadian DG, Noble M (1993) Proton nuclear magnetic resonance spectroscopy unambiguously identifies different neural cell types. J Neurosci 13(3):981–989
Menon DK, Ainsworth JG, Cox IJ, Coker RC, Sargentoni J, Coutts GA, Baudouin CJ, Kocsis AE, Harris JR (1992) Proton MR spectroscopy of the brain in AIDS dementia complex. J Comput Assist Tomogr 16(4):538–542
Meyerhoff DJ, MacKay S, Bachman L, Poole N, Dillon WP, Weiner MW, Fein G (1993) Reduced brain N-acetylaspartate suggests neuronal loss in cognitively impaired human immunodeficiency virus-seropositive individuals: in vivo 1H magnetic resonance spectroscopic imaging. Neurology 43(3 Pt 1):509–515
Chong WK, Sweeney B, Wilkinson ID, Paley M, Hall-Craggs MA, Kendall BE, Shepard JK, Beecham M, Miller RF, Weller IV et al (1993) Proton spectroscopy of the brain in HIV infection: correlation with clinical, immunologic, and MR imaging findings. Radiology 188(1):119–124
Barker PB, Lee RR, McArthur JC (1995) AIDS dementia complex: evaluation with proton MR spectroscopic imaging. Radiology 195(1):58–64
Meyerhoff DJ, Bloomer C, Cardenas V, Norman D, Weiner MW, Fein G (1999) Elevated subcortical choline metabolites in cognitively and clinically asymptomatic HIV+ patients. Neurology 52(5):995–1003
Brand A, Richter-Landsberg C, Leibfritz D (1993) Multinuclear NMR studies on the energy metabolism of glial and neuronal cells. Dev Neurosci 15(3–5):289–298
Laubenberger J, Haussinger D, Bayer S, Thielemann S, Schneider B, Mundinger A, Hennig J, Langer M (1996) HIV-related metabolic abnormalities in the brain: depiction with proton MR spectroscopy with short echo times. Radiology 199(3):805–810
Lopez-Villegas D, Lenkinski RE, Frank I (1997) Biochemical changes in the frontal lobe of HIV-infected individuals detected by magnetic resonance spectroscopy. Proc Natl Acad Sci U S A 94(18):9854–9859
Chang L, Ernst T, Leonido-Yee M, Walot I, Singer E (1999) Cerebral metabolite abnormalities correlate with clinical severity of HIV-1 cognitive motor complex. Neurology 52(1):100–108
Saunders DE, Howe FA, van den Boogaart A, Griffiths JR, Brown MM (1999) Aging of the adult human brain: in vivo quantitation of metabolite content with proton magnetic resonance spectroscopy. J Magn Reson Imaging 9(5):711–716
Chang L, Ernst T, Witt MD, Ames N, Gaiefsky M, Miller E (2002) Relationships among brain metabolites, cognitive function, and viral loads in antiretroviral-naive HIV patients. Neuroimage 17(3):1638–1648
Ratai EM, Pilkenton SJ, Greco JB, Lentz MR, Bombardier JP, Turk KW, He J, Joo CG, Lee V, Westmoreland S, Halpern E, Lackner AA, Gonzalez RG (2009) In vivo proton magnetic resonance spectroscopy reveals region specific metabolic responses to SIV infection in the macaque brain. BMC Neurosci 10:63
Ratai EM, Annamalai L, Burdo T, Joo CG, Bombardier JP, Fell R, Hakimelahi R, He J, Lentz MR, Campbell J, Curran E, Halpern EF, Masliah E, Westmoreland SV, Williams KC, Gonzalez RG (2011) Brain creatine elevation and N-acetylaspartate reduction indicates neuronal dysfunction in the setting of enhanced glial energy metabolism in a macaque model of NeuroAIDS. Magn Reson Med 66(3):625–634
Sailasuta N, Shriner K, Ross B (2009) Evidence of reduced glutamate in the frontal lobe of HIV-seropositive patients. NMR Biomed 22(3):326–331
Harezlak J, Buchthal S, Taylor M, Schifitto G, Zhong J, Daar E, Alger J, Singer E, Campbell T, Yiannoutsos C, Cohen R, Navia B (2011) Persistence of HIV-associated cognitive impairment, inflammation, and neuronal injury in era of highly active antiretroviral treatment. AIDS 25(5):625–633
Ernst T, Jiang CS, Nakama H, Buchthal S, Chang L (2010) Lower brain glutamate is associated with cognitive deficits in HIV patients: a new mechanism for HIV-associated neurocognitive disorder. J Magn Reson Imaging 32(5):1045–1053
Mohamed MA, Barker PB, Skolasky RL, Selnes OA, Moxley RT, Pomper MG, Sacktor NC (2010) Brain metabolism and cognitive impairment in HIV infection: a 3-T magnetic resonance spectroscopy study. Magn Reson Imaging 28(9):1251–1257
Lentz MR, Kim WK, Lee V, Bazner S, Halpern EF, Venna N, Williams K, Rosenberg ES, Gonzalez RG (2009) Changes in MRS neuronal markers and T cell phenotypes observed during early HIV infection. Neurology 72(17):1465–1472
Roc AC, Ances BM, Chawla S, Korczykowski M, Wolf RL, Kolson DL, Detre JA, Poptani H (2007) Detection of human immunodeficiency virus induced inflammation and oxidative stress in lenticular nuclei with magnetic resonance spectroscopy despite antiretroviral therapy. Arch Neurol 64(9):1249–1257
Lee PL, Yiannoutsos CT, Ernst T, Chang L, Marra CM, Jarvik JG, Richards TL, Kwok EW, Kolson DL, Simpson D, Tang CY, Schifitto G, Ketonen LM, Meyerhoff DJ, Lenkinski RE, Gonzalez RG, Navia BA (2003) A multi-center 1H MRS study of the AIDS dementia complex: validation and preliminary analysis. J Magn Reson Imaging 17(6):625–633
Chang L, Lee PL, Yiannoutsos CT, Ernst T, Marra CM, Richards T, Kolson D, Schifitto G, Jarvik JG, Miller EN, Lenkinski R, Gonzalez G, Navia BA (2004) A multicenter in vivo proton-MRS study of HIV-associated dementia and its relationship to age. Neuroimage 23(4):1336–1347
Sacktor N, Skolasky RL, Ernst T, Mao X, Selnes O, Pomper MG, Chang L, Zhong K, Shungu DC, Marder K, Shibata D, Schifitto G, Bobo L, Barker PB (2005) A multicenter study of two magnetic resonance spectroscopy techniques in individuals with HIV dementia. J Magn Reson Imaging 21(4):325–333
Greco JB, Sakaie KE, Aminipour S, Lee PL, Chang LL, He J, Westmoreland S, Lackner AA, Gonzalez RG (2002) Magnetic resonance spectroscopy: an in vivo tool for monitoring cerebral injury in SIV-infected macaques. J Med Primatol 31(4–5):228–236
Jacobs MA, Horska A, van Zijl PC, Barker PB (2001) Quantitative proton MR spectroscopic imaging of normal human cerebellum and brain stem. Magn Reson Med 46(4):699–705
Soher BJ, van Zijl PC, Duyn JH, Barker PB (1996) Quantitative proton MR spectroscopic imaging of the human brain. Magn Reson Med 35(3):356–363
Michaelis T, Merboldt KD, Bruhn H, Hanicke W, Frahm J (1993) Absolute concentrations of metabolites in the adult human brain in vivo: quantification of localized proton MR spectra. Radiology 187(1):219–227
Barker PB, Soher BJ, Blackband SJ, Chatham JC, Mathews VP, Bryan RN (1993) Quantitation of proton NMR spectra of the human brain using tissue water as an internal concentration reference. NMR Biomed 6(1):89–94
Christiansen P, Henriksen O, Stubgaard M, Gideon P, Larsson HB (1993) In vivo quantification of brain metabolites by 1H-MRS using water as an internal standard. Magn Reson Imaging 11(1):107–118
Ernst T, Kreis R, Ross BD (1993) Absolute quantitation of water and metabolites in the human brain; Part I: Compartments and water. J Magn Res B102:1–8
Tofts PS, Wray S (1988) A critical assessment of methods of measuring metabolite concentrations by NMR spectroscopy. NMR Biomed 1(1):1–10
Hennig J, Pfister H, Ernst T, Ott D (1992) Direct absolute quantification of metabolites in the human brain with in vivo localized proton spectroscopy. NMR Biomed 5(4):193–199
Toggas SM, Masliah E, Rockenstein EM, Rall GF, Abraham CR, Mucke L (1994) Central nervous system damage produced by expression of the HIV-1 coat protein gp120 in transgenic mice. Nature 367(6459):188–193
Persidsky Y, Limoges J, McComb R, Bock P, Baldwin T, Tyor W, Patil A, Nottet HS, Epstein L, Gelbard H, Flanagan E, Reinhard J, Pirruccello SJ, Gendelman HE (1996) Human immunodeficiency virus encephalitis in SCID mice. Am J Pathol 149(3):1027–1053
Persidsky Y, Buttini M, Limoges J, Bock P, Gendelman HE (1997) An analysis of HIV-1-associated inflammatory products in brain tissue of humans and SCID mice with HIV-1 encephalitis. J Neurovirol 3(6):401–416
Persidsky Y, Gendelman HE (2002) Murine models for human immunodeficiency virus type 1-associated dementia: the development of new treatment testing paradigms. J Neurovirol 8(Suppl 2):49–52
Zink WE, Anderson E, Boyle J, Hock L, Rodriguez-Sierra J, Xiong H, Gendelman HE, Persidsky Y (2002) Impaired spatial cognition and synaptic potentiation in a murine model of human immunodeficiency virus type 1 encephalitis. J Neurosci 22(6):2096–2105
Jacobson S, Henriksen SJ, Prospero-Garcia O, Phillips TR, Elder JH, Young WG, Bloom FE, Fox HS (1997) Cortical neuronal cytoskeletal changes associated with FIV infection. J Neurovirol 3(4):283–289
Podell M, Oglesbee M, Mathes L, Krakowka S, Olmstead R, Lafrado L (1993) AIDS-associated encephalopathy with experimental feline immunodeficiency virus infection. J Acquir Immune Defic Syndr 6(7):758–771
Phillips TR, Prospero-Garcia O, Wheeler DW, Wagaman PC, Lerner DL, Fox HS, Whalen LR, Bloom FE, Elder JH, Henriksen SJ (1996) Neurologic dysfunctions caused by a molecular clone of feline immunodeficiency virus, FIV-PPR. J Neurovirol 2(6):388–396
Silvotti L, Corradi A, Brandi G, Cabassi A, Bendinelli M, Magnan M, Piedimonte G (1997) FIV induced encephalopathy: early brain lesions in the absence of viral replication in monocyte/macrophages. A pathogenetic model. Vet Immunol Immunopathol 55(4):263–271
Prospero-Garcia O, Herold N, Waters AK, Phillips TR, Elder JH, Henriksen SJ (1994) Intraventricular administration of a FIV-envelope protein induces sleep architecture changes in rats. Brain Res 659(1–2):254–258
Poli A, Abramo F, Di Iorio C, Cantile C, Carli MA, Pollera C, Vago L, Tosoni A, Costanzi G (1997) Neuropathology in cats experimentally infected with feline immunodeficiency virus: a morphological, immunocytochemical and morphometric study. J Neurovirol 3(5):361–368
Podell M, March PA, Buck WR, Mathes LE (2000) The feline model of neuroAIDS: understanding the progression towards AIDS dementia. J Psychopharmacol 14(3):205–213
Podell M, Maruyama K, Smith M, Hayes KA, Buck WR, Ruehlmann DS, Mathes LE (1999) Frontal lobe neuronal injury correlates to altered function in FIV-infected cats. J Acquir Immune Defic Syndr 22(1):10–18
Desrosiers RC (1990) The simian immunodeficiency viruses. Annu Rev Immunol 8:557–578
Zink MC, Amedee AM, Mankowski JL, Craig L, Didier P, Carter DL, Munoz A, Murphey-Corb M, Clements JE (1997) Pathogenesis of SIV encephalitis. Selection and replication of neurovirulent SIV. Am J Pathol 151(3):793–803
Zink MC, Spelman JP, Robinson RB, Clements JE (1998) SIV infection of macaques—modeling the progression to AIDS dementia. J Neurovirol 4(3):249–259
Murray EA, Rausch DM, Lendvay J, Sharer LR, Eiden LE (1992) Cognitive and motor impairments associated with SIV infection in rhesus monkeys. Science 255(5049):1246–1249
Burudi EME, Fox HS (2001) Simian immunodeficiency virus model of HIV-induced central nervous system dysfunction. Adv Virus Res 56:435–468
Bruce-Keller AJ, Chauhan A, Dimayuga FO, Gee J, Keller JN, Nath A (2003) Synaptic transport of human immunodeficiency virus-Tat protein causes neurotoxicity and gliosis in rat brain. J Neurosci 23(23):8417–8422
Okamoto S, Kang YJ, Brechtel CW, Siviglia E, Russo R, Clemente A, Harrop A, McKercher S, Kaul M, Lipton SA (2007) HIV/gp120 decreases adult neural progenitor cell proliferation via checkpoint kinase-mediated cell-cycle withdrawal and G1 arrest. Cell Stem Cell 1(2):230–236
Zelivyanskaya ML, Nelson JA, Poluektova L, Uberti M, Mellon M, Gendelman HE, Boska MD (2003) Tracking superparamagnetic iron oxide labeled monocytes in brain by high-field magnetic resonance imaging. J Neurosci Res 73(3):284–295
Nelson JA, Dou H, Ellison B, Uberti M, Xiong H, Anderson E, Mellon M, Gelbard HA, Boska M, Gendelman HE (2005) Coregistration of quantitative proton magnetic resonance spectroscopic imaging with neuropathological and neurophysiological analyses defines the extent of neuronal impairments in murine human immunodeficiency virus type-1 encephalitis. J Neurosci Res 80(4):562–575
Phipps AJ, Hayes KA, Buck WR, Podell M, Mathes LE (2000) Neurophysiologic and immunologic abnormalities associated with feline immunodeficiency virus molecular clone FIV-PPR DNA inoculation. J Acquir Immune Defic Syndr 23(1):8–16
Podell M, Hayes K, Oglesbee M, Mathes L (1997) Progressive encephalopathy associated with CD4/CD8 inversion in adult FIV-infected cats. J Acquir Immune Defic Syndr Hum Retrovirol 15(5):332–340
Belman AL (1997) Pediatric neuro-AIDS. Update. Neuroimaging Clin N Am 7(3):593–613
Johnston JB, Silva C, Hiebert T, Buist R, Dawood MR, Peeling J, Power C (2002) Neurovirulence depends on virus input titer in brain in feline immunodeficiency virus infection: evidence for activation of innate immunity and neuronal injury. J Neurovirol 8(5):420–431
Reimann KA, Tenner-Racz K, Racz P, Montefiori DC, Yasutomi Y, Lin W, Ransil BJ, Letvin NL (1994) Immunopathogenic events in acute infection of rhesus monkeys with simian immunodeficiency virus of macaques. J Virol 68(4):2362–2370
Staprans SI, Dailey PJ, Rosenthal A, Horton C, Grant RM, Lerche N, Feinberg MB (1999) Simian immunodeficiency virus disease course is predicted by the extent of virus replication during primary infection. J Virol 73(6):4829–4839
Greco JB, Westmoreland SV, Ratai EM, Lentz MR, Sakaie K, He J, Sehgal PK, Masliah E, Lackner AA, Gonzalez RG (2004) In vivo 1H MRS of brain injury and repair during acute SIV infection in the macaque model of neuroAIDS. Magn Reson Med 51(6):1108–1114
Chakrabarti L, Hurtrel M, Maire MA, Vazeux R, Dormont D, Montagnier L, Hurtrel B (1991) Early viral replication in the brain of SIV-infected rhesus monkeys. Am J Pathol 139(6):1273–1280
Sharer LR, Baskin GB, Cho ES, Murphey-Corb M, Blumberg BM, Epstein LG (1988) Comparison of simian immunodeficiency virus and human immunodeficiency virus encephalitides in the immature host. Ann Neurol 23(Suppl):S108–S112
Simon MA, Chalifoux LV, Ringler DJ (1992) Pathologic features of SIV-induced disease and the association of macrophage infection with disease evolution. AIDS Res Hum Retroviruses 8(3):327–337
Fox HS, Weed MR, Huitron-Resendiz S, Baig J, Horn TFW, Dailey PJ, Bischofberger N, Henriksen SJ (2000) Antiretroviral treatment normalizes neurophysiological but not movement abnormalities in simian immunodeficiency virus-infected monkeys. J Clin Invest 106(1):37–45
Williams K, Westmoreland S, Greco J, Ratai E, Lentz M, Kim WK, Fuller RA, Kim JP, Autissier P, Sehgal PK, Schinazi RF, Bischofberger N, Piatak M, Lifson JD, Masliah E, Gonzalez RG (2005) Magnetic resonance spectroscopy reveals that activated monocytes contribute to neuronal injury in SIV neuroAIDS. J Clin Invest 115(9):2534–2545
Altfeld M, Rosenberg ES, Shankarappa R, Mukherjee JS, Hecht FM, Eldridge RL, Addo MM, Poon SH, Phillips MN, Robbins GK, Sax PE, Boswell S, Kahn JO, Brander C, Goulder PJ, Levy JA, Mullins JI, Walker BD (2001) Cellular immune responses and viral diversity in individuals treated during acute and early HIV-1 infection. J Exp Med 193(2):169–180
Rosenberg E, Cotton D (1997) Primary HIV infection and the acute retroviral syndrome. AIDS Clin Care 9(3):19, 23–25
McArthur JC, Hoover DR, Bacellar H, Miller EN, Cohen BA, Becker JT, Graham NM, McArthur JH, Selnes OA, Jacobson LP et al (1993) Dementia in AIDS patients: incidence and risk factors. Multicenter AIDS Cohort Study. Neurology 43(11):2245–2252
Lentz MR, Kim JP, Westmoreland SV, Greco JB, Fuller RA, Ratai EM, He J, Sehgal PK, Halpern EF, Lackner AA, Masliah E, Gonzalez RG (2005) Quantitative neuropathologic correlates of changes in ratio of N-acetylaspartate to creatine in macaque brain. Radiology 235(2):461–468
Blusztajn JK, Wurtman RJ (1983) Choline and cholinergic neurons. Science 221(4611):614–620
Kim JP, Lentz MR, Westmoreland SV, Greco JB, Ratai EM, Halpern E, Lackner AA, Masliah E, Gonzalez RG (2005) Relationships between astrogliosis and 1H MR spectroscopic measures of brain choline/creatine and myo-inositol/creatine in a primate model. AJNR Am J Neuroradiol 26(4):752–759
Fuller RA, Westmoreland SV, Ratai E, Greco JB, Kim JP, Lentz MR, He J, Sehgal PK, Masliah E, Halpern E, Lackner AA, Gonzalez RG (2004) A prospective longitudinal in vivo 1H MR spectroscopy study of the SIV/macaque model of neuroAIDS. BMC Neurosci 5(1):10
Westmoreland SV, Halpern E, Lackner AA (1998) Simian immunodeficiency virus encephalitis in rhesus macaques is associated with rapid disease progression. J Neurovirol 4(3):260–268
Zink MC, Suryanarayana K, Mankowski JL, Shen A, Piatak M Jr, Spelman JP, Carter DL, Adams RJ, Lifson JD, Clements JE (1999) High viral load in the cerebrospinal fluid and brain correlates with severity of simian immunodeficiency virus encephalitis. J Virol 73(12):10480–10488
Zink M, Clements J (2000) A rapid, reproducible model of AIDS and encephelitis in SIV infected macaques demonstrates the role of viral load in CNS disease. NeuroAIDS 3(5):online—October
Zink MC, Clements JE (2002) A novel simian immunodeficiency virus model that provides insight into mechanisms of human immunodeficiency virus central nervous system disease. J Neurovirol 8(Suppl 2):42–48
Weed MR, Hienz RD, Brady JV, Adams RJ, Mankowski JL, Clements JE, Zink MC (2003) Central nervous system correlates of behavioral deficits following simian immunodeficiency virus infection. J Neurovirol 9(4):452–464
Schmitz JE, Kuroda MJ, Santra S, Sasseville VG, Simon MA, Lifton MA, Racz P, Tenner-Racz K, Dalesandro M, Scallon BJ, Ghrayeb J, Forman MA, Montefiori DC, Rieber EP, Letvin NL, Reimann KA (1999) Control of viremia in simian immunodeficiency virus infection by CD8+ lymphocytes. Science 283(5403):857–860
Schmitz JE, Simon MA, Kuroda MJ, Lifton MA, Ollert MW, Vogel CW, Racz P, Tenner-Racz K, Scallon BJ, Dalesandro M, Ghrayeb J, Rieber EP, Sasseville VG, Reimann KA (1999) A nonhuman primate model for the selective elimination of CD8+ lymphocytes using a mouse-human chimeric monoclonal antibody. Am J Pathol 154(6):1923–1932
Williams K, Alvarez X, Lackner AA (2001) Central nervous system perivascular cells are immunoregulatory cells that connect the CNS with the peripheral immune system. Glia 36(2):156–164
Domercq M, Matute C (2004) Neuroprotection by tetracyclines. Trends Pharmacol Sci 25(12):609–612
Baptiste DC, Fehlings MG (2006) Pharmacological approaches to repair the injured spinal cord. J Neurotrauma 23(3–4):318–334
Stirling DP, Koochesfahani KM, Steeves JD, Tetzlaff W (2005) Minocycline as a neuroprotective agent. Neuroscientist 11(4):308–322
Tikka T, Fiebich BL, Goldsteins G, Keinanen R, Koistinaho J (2001) Minocycline, a tetracycline derivative, is neuroprotective against excitotoxicity by inhibiting activation and proliferation of microglia. J Neurosci 21(8):2580–2588
Lin S, Zhang Y, Dodel R, Farlow MR, Paul SM, Du Y (2001) Minocycline blocks nitric oxide-induced neurotoxicity by inhibition p38 MAP kinase in rat cerebellar granule neurons. Neurosci Lett 315(1–2):61–64
Si Q, Cosenza M, Kim MO, Zhao ML, Brownlee M, Goldstein H, Lee S (2004) A novel action of minocycline: inhibition of human immunodeficiency virus type 1 infection in microglia. J Neurovirol 10(5):284–292
Dutta K, Mishra MK, Nazmi A, Kumawat KL, Basu A (2010) Minocycline differentially modulates macrophage mediated peripheral immune response following Japanese encephalitis virus infection. Immunobiology 215(11):884–893
Zink MC, Uhrlaub J, DeWitt J, Voelker T, Bullock B, Mankowski J, Tarwater P, Clements J, Barber S (2005) Neuroprotective and anti-human immunodeficiency virus activity of minocycline. JAMA 293(16):2003–2011
Ratai EM, Bombardier JP, Joo CG, Annamalai L, Burdo TH, Campbell J, Fell R, Hakimelahi R, He J, Autissier P, Lentz MR, Halpern EF, Masliah E, Williams KC, Westmoreland SV, Gonzalez RG (2010) Proton magnetic resonance spectroscopy reveals neuroprotection by oral minocycline in a nonhuman primate model of accelerated NeuroAIDS. PLoS One 5(5):e10523
Ratai EM, Fell R, He J, Piatak M, Lifson JD, Burdo TH, Campbell J, Autissier P, Annamalai L, Masliah E, Westmoreland SV, Williams KC, Gilberto Gonzalez R (2012) Reduction of brain virus by minocycline and combination anti-retroviral therapy produces neuronal protection in a primate model of AIDS. In: ISMRM 20th Annual Meeting & Exhibition, Melbourne
Campbell JH, Burdo TH, Autissier P, Bombardier JP, Westmoreland SV, Soulas C, Gonzalez RG, Ratai EM, Williams KC (2011) Minocycline inhibition of monocyte activation correlates with neuronal protection in SIV NeuroAIDS. PLoS One 6(4):e18688
Lentz MR, Kim WK, Kim H, Soulas C, Lee V, Venna N, Halpern EF, Rosenberg ES, Williams K, Gonzalez RG (2011) Alterations in brain metabolism during the first year of HIV infection. J Neurovirol 17(3):220–229
Sailasuta N, Ross W, Ananworanich J, Chalermchai T, DeGruttola V, Lerdlum S, Pothisri M, Busovaca E, Ratto-Kim S, Jagodzinski L, Spudich S, Michael N, Kim JH, Valcour V (2012) Change in brain magnetic resonance spectroscopy after treatment during acute HIV infection. PLoS One 7(11):e49272
Valcour V, Chalermchai T, Sailasuta N, Marovich M, Lerdlum S, Suttichom D, Suwanwela NC, Jagodzinski L, Michael N, Spudich S, van Griensven F, de Souza M, Kim J, Ananworanich J (2012) Central nervous system viral invasion and inflammation during acute HIV infection. J Infect Dis 206(2):275–282
Peluso MJ, Meyerhoff DJ, Price RW, Peterson J, Lee E, Young AC, Walter R, Fuchs D, Brew BJ, Cinque P, Robertson K, Hagberg L, Zetterberg H, Gisslen M, Spudich S (2013) Cerebrospinal fluid and neuroimaging biomarker abnormalities suggest early neurological injury in a subset of individuals during primary HIV infection. J Infect Dis 207(11):1703–1712
Masters MC, Ances BM (2014) Role of neuroimaging in HIV-associated neurocognitive disorders. Semin Neurol 34(1):89–102
Menon DK, Baudouin CJ, Tomlinson D, Hoyle C (1990) Proton MR spectroscopy and imaging of the brain in AIDS: evidence of neuronal loss in regions that appear normal with imaging. J Comput Assist Tomogr 14(6):882–885
Salvan AM, Vion-Dury J, Confort-Gouny S, Nicoli F, Lamoureux S, Cozzone PJ (1997) Brain proton magnetic resonance spectroscopy in HIV-related encephalopathy: identification of evolving metabolic patterns in relation to dementia and therapy. AIDS Res Hum Retroviruses 13(12):1055–1066
Tracey I, Carr CA, Guimaraes AR, Worth JL, Navia BA, Gonzalez RG (1996) Brain choline-containing compounds are elevated in HIV-positive patients before the onset of AIDS dementia complex: a proton magnetic resonance spectroscopic study. Neurology 46(3):783–788
Marcus CD, Taylor-Robinson SD, Sargentoni J, Ainsworth JG, Frize G, Easterbrook PJ, Shaunak S, Bryant DJ (1998) 1H MR spectroscopy of the brain in HIV-1-seropositive subjects: evidence for diffuse metabolic abnormalities. Metab Brain Dis 13(2):123–136
Moller HE, Vermathen P, Lentschig MG, Schuierer G, Schwarz S, Wiedermann D, Evers S, Husstedt IW (1999) Metabolic characterization of AIDS dementia complex by spectroscopic imaging. J Magn Reson Imaging 9(1):10–18
von Giesen HJ, Wittsack HJ, Wenserski F, Koller H, Hefter H, Arendt G (2001) Basal ganglia metabolite abnormalities in minor motor disorders associated with human immunodeficiency virus type 1. Arch Neurol 58(8):1281–1286
Anthony IC, Ramage SN, Carnie FW, Simmonds P, Bell JE (2005) Influence of HAART on HIV-related CNS disease and neuroinflammation. J Neuropathol Exp Neurol 64(6):529–536
Anthony IC, Bell JE (2008) The neuropathology of HIV/AIDS. Int Rev Psychiatry 20(1):15–24
Chang L, Ernst T, Leonido-Yee M, Witt M, Speck O, Walot I, Miller EN (1999) Highly active antiretroviral therapy reverses brain metabolite abnormalities in mild HIV dementia. Neurology 53(4):782–789
Schifitto G, Navia BA, Yiannoutsos CT, Marra CM, Chang L, Ernst T, Jarvik JG, Miller EN, Singer EJ, Ellis RJ, Kolson DL, Simpson D, Nath A, Berger J, Shriver SL, Millar LL, Colquhoun D, Lenkinski R, Gonzalez RG, Lipton SA (2007) Memantine and HIV-associated cognitive impairment: a neuropsychological and proton magnetic resonance spectroscopy study. AIDS 21(14):1877–1886
Schifitto G, Yiannoutsos CT, Ernst T, Navia BA, Nath A, Sacktor N, Anderson C, Marra CM, Clifford DB (2009) Selegiline and oxidative stress in HIV-associated cognitive impairment. Neurology 73(23):1975–1981
Pfefferbaum A, Adalsteinsson E, Sullivan EV (2005) Cortical NAA deficits in HIV infection without dementia: influence of alcoholism comorbidity. Neuropsychopharmacology 30(7):1392–1399
Chang L, Ernst T, Speck O, Grob CS (2005) Additive effects of HIV and chronic methamphetamine use on brain metabolite abnormalities. Am J Psychiatry 162(2):361–369
Marra CM, Zhao Y, Clifford DB, Letendre S, Evans S, Henry K, Ellis RJ, Rodriguez B, Coombs RW, Schifitto G, McArthur JC, Robertson K (2009) Impact of combination antiretroviral therapy on cerebrospinal fluid HIV RNA and neurocognitive performance. AIDS 23(11):1359–1366
Schweinsburg BC, Taylor MJ, Alhassoon OM, Gonzalez R, Brown GG, Ellis RJ, Letendre S, Videen JS, McCutchan JA, Patterson TL, Grant I (2005) Brain mitochondrial injury in human immunodeficiency virus-seropositive (HIV+) individuals taking nucleoside reverse transcriptase inhibitors. J Neurovirol 11(4):356–364
Keller MA, Venkatraman TN, Thomas A, Deveikis A, LoPresti C, Hayes J, Berman N, Walot I, Padilla S, Johnston-Jones J, Ernst T, Chang L (2004) Altered neurometabolite development in HIV-infected children: correlation with neuropsychological tests. Neurology 62(10):1810–1817
Mbugua K, Holmes MJ, Hess AT, Little F, Cotton MF, Dobbels E, van der Kouwe AJ, Laughton B, Meintjes EM (2014) Effects of ART timing and HIV progression on neuro-metabolite levels in basal ganglia at age 5 years. In: 20th Annual Meeting of the Organization for Human Brain Mapping, Hamburg, Germany
Holmes MJ, Mbugua K, Little F, Cotton MF, van der Kouwe AJ, Laughton B, Meintjes EM (2014) A longitudinal study of the effects of HIV exposure on metabolite levels in the Midfrontal Gray Matter in children: at 5 and 7 years. In: 20th Annual AIDS conference, Melbourne, Australia
Banakar S, Thomas MA, Deveikis A, Watzl JQ, Hayes J, Keller MA (2008) Two-dimensional 1H MR spectroscopy of the brain in human immunodeficiency virus (HIV)-infected children. J Magn Reson Imaging 27(4):710–717
Thomas MA, Yue K, Binesh N, Davanzo P, Kumar A, Siegel B, Frye M, Curran J, Lufkin R, Martin P, Guze B (2001) Localized two-dimensional shift correlated MR spectroscopy of human brain. Magn Reson Med 46(1):58–67
Trabesinger AH, Boesiger P (2001) Improved selectivity of double quantum coherence filtering for the detection of glutathione in the human brain in vivo. Magn Reson Med 45(4):708–710
Trabesinger AH, Weber OM, Duc CO, Boesiger P (1999) Detection of glutathione in the human brain in vivo by means of double quantum coherence filtering. Magn Reson Med 42(2):283–289
Terpstra M, Henry PG, Gruetter R (2003) Measurement of reduced glutathione (GSH) in human brain using LCModel analysis of difference-edited spectra. Magn Reson Med 50(1):19–23
Terpstra M, Marjanska M, Henry PG, Tkac I, Gruetter R (2006) Detection of an antioxidant profile in the human brain in vivo via double editing with MEGA-PRESS. Magn Reson Med 56(6):1192–1199
Srinivas A, Dias BF (2008) Antioxidants in HIV positive children. Indian J Pediatr 75(4):347–350
Markovic I, Clouse KA (2004) Recent advances in understanding the molecular mechanisms of HIV-1 entry and fusion: revisiting current targets and considering new options for therapeutic intervention. Curr HIV Res 2(3):223–234
Provencher SW (1993) Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn Reson Med 30(6):672–679
Tkac I, Oz G, Adriany G, Ugurbil K, Gruetter R (2009) In vivo 1H NMR spectroscopy of the human brain at high magnetic fields: metabolite quantification at 4T vs. 7T. Magn Reson Med 62(4):868–879
Ratai EM, Pilkenton S, He J, Fell R, Bombardier JP, Joo CG, Lentz MR, Kim WK, Burdo TH, Autissier P, Annamalai L, Curran E, O’Neil SP, Westmoreland SV, Williams KC, Masliah E, Gilberto Gonzalez R (2011) CD8+ lymphocyte depletion without SIV infection does not produce metabolic changes or pathological abnormalities in the rhesus macaque brain. J Med Primatol 40(5):300–309
Gonen O, Liu S, Goelman G, Ratai EM, Pilkenton S, Lentz MR, Gonzalez RG (2008) Proton MR spectroscopic imaging of rhesus macaque brain in vivo at 7T. Magn Reson Med 59(4):692–699
Caruso PA, Johnson J, Thibert R, Rapalino O, Rincon S, Ratai EM (2013) The use of magnetic resonance spectroscopy in the evaluation of epilepsy. Neuroimaging Clin N Am 23(3):407–424
Hess AT, Tisdall MD, Andronesi OC, Meintjes EM, van der Kouwe AJ (2011) Real-time motion and B0 corrected single voxel spectroscopy using volumetric navigators. Magn Reson Med 66(2):314–323
Tisdall MD, Hess AT, Reuter M, Meintjes EM, Fischl B, van der Kouwe AJ (2012) Volumetric navigators for prospective motion correction and selective reacquisition in neuroanatomical MRI. Magn Reson Med 68(2):389–399
Bogner W, Hess AT, Gagoski B, Tisdall MD, van der Kouwe AJ, Trattnig S, Rosen B, Andronesi OC (2013) Real-time motion- and B-correction for LASER-localized spiral-accelerated 3D-MRSI of the brain at 3T. Neuroimage 88C:22–31
Kuperman JM, Brown TT, Ahmadi ME, Erhart MJ, White NS, Roddey JC, Shankaranarayanan A, Han ET, Rettmann D, Dale AM (2011) Prospective motion correction improves diagnostic utility of pediatric MRI scans. Pediatr Radiol 41(12):1578–1582
Brown TT, Kuperman JM, Erhart M, White NS, Roddey JC, Shankaranarayanan A, Han ET, Rettmann D, Dale AM (2010) Prospective motion correction of high-resolution magnetic resonance imaging data in children. Neuroimage 53(1):139–145
Andrews-Shigaki BC, Armstrong BS, Zaitsev M, Ernst T (2011) Prospective motion correction for magnetic resonance spectroscopy using single camera Retro-Grate reflector optical tracking. J Magn Reson Imaging 33(2):498–504
Keating B, Ernst T (2012) Real-time dynamic frequency and shim correction for single-voxel magnetic resonance spectroscopy. Magn Reson Med 68(5):1339–1345
Acknowledgements
This work was partly funded by NIH grants R21NS059331, R01NS050041, R01NS040237, R01NS37654, R01MH62962, MH59754, MH62512, RR00168, R24 RR016001, N01 AI040101, and P41RR14075. The author wishes to thank Dr. R Gilberto Gonzalez for valuable discussions.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Ratai, EM. (2016). Magnetic Resonance Spectroscopy in HIV-Associated Neurocognitive Disorders: HAND. In: Öz, G. (eds) Magnetic Resonance Spectroscopy of Degenerative Brain Diseases. Contemporary Clinical Neuroscience. Springer, Cham. https://doi.org/10.1007/978-3-319-33555-1_11
Download citation
DOI: https://doi.org/10.1007/978-3-319-33555-1_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-33553-7
Online ISBN: 978-3-319-33555-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)