
Abstract
The acrosome, a single exocytotic vesicle on the head of sperm, has an essential role in fertilization, but the exact mechanisms by which it facilitates sperm–egg interactions remain unresolved. The acrosome contains dozens of secretory proteins that are packaged into the forming structure during spermatogenesis; many of these proteins are localized into specific topographical areas of the acrosome, while others are more diffusely distributed. Acrosomal proteins can also be biochemically classified as components of the acrosomal matrix, a large, relatively insoluble complex, or as soluble proteins. This review focuses on recent findings using genetically modified mice (gene knockouts and transgenic “green acrosome” mice) to study the effects of eliminating acrosomal matrix-associated proteins on sperm structure and function. Some gene knockouts produce infertile phenotypes with obviously missing, specific activities that affect acrosome biogenesis during spermatogenesis or interfere with acrosome function in mature sperm. Mutations that delete some components produce fertile phenotypes with subtler effects that provide useful insights into acrosomal matrix function in fertilization. In general, these studies enable the reassessment of paradigms to explain acrosome formation and function and provide novel, objective insights into the roles of acrosomal matrix proteins in fertilization. The use of genetically engineered mouse models has yielded new mechanistic information that complements recent, important in vivo imaging studies.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
2.1 Introduction
The acrosome is a single exocytotic vesicle overlying the nucleus of most animal sperm. While the sperm of some taxa (e.g., teleost fish) do not possess an acrosome, it is generally thought that mammalian sperm acrosomes may perform two principal exocytosis-dependent functions: (1) to serve as a site for the sequestration and release of proteins required for binding to and penetration of the zona pellucida and (2) to enable the sperm to fuse with the oolemma. For the past several decades, research has been largely guided by the concept that mammalian sperm acrosomes must be intact to bind to the zona pellucida prior to undergoing exocytosis, enabling the sperm to penetrate this vestment and fertilize the egg (Wassarman et al. 2004). Although this widely accepted dogma has been challenged on many levels, recent experiments utilizing modern, highly sophisticated imaging techniques have enabled investigators to track the acrosomal status of individual fertilizing spermatozoa. Using genetically engineered mouse strains that express green fluorescent protein in their acrosomes, these experiments have shown that almost all fertilizing mouse sperm have undergone acrosomal exocytosis prior to the time they penetrate the zona pellucida (Jin et al. 2011).
These contemporary experiments raise new questions of how the acrosome forms during spermatogenesis and functions in the fertilization process. Specific issues that need to be addressed include (1) the contribution of acrosomal biogenesis to the fertility of individual sperm, (2) the timing of acrosomal exocytosis in vivo, (3) the role of acrosomal compartmentalization (including structurally distinct domains within the vesicle and biochemical segregation of overlapping soluble materials and insoluble acrosomal matrix compartments), and (4) the functions of specific proteins in fertilization. While recent results challenge or even contradict experiments that were based upon previous reductionist paradigms, many of the older studies were carefully designed and yielded a solid body of evidence that should be reevaluated from a more holistic standpoint to see whether a reinterpretation sheds new light on the question of the physiological role of the acrosome in fertilization.
In a previous review, we focused on the structure and function of the sperm acrosomal matrix in fertilization, where we espoused three principal points: (1) the acrosome is not a “bag of enzymes,” simply comprised of soluble substances that are rapidly released when the outer acrosomal membrane fuses with the plasma membrane, (2) acrosomal exocytosis is not an “all-or-none” event, where the acrosome is either “intact” or “reacted” but instead incorporates intermediate steps involving the progressive loss of acrosomal matrix material, and (3) the acrosomal matrix is a molecular scaffold assembly that is dismantled by a self-regulated mechanism driven, in part, by proteolysis (Buffone et al. 2008). We will not recapitulate these points in this review, but we will discuss them in light of the many new findings from studies using genetically modified mice.
It is abundantly clear that fertilization is a complicated process that requires models to extend from a cluster of hundreds or thousands of sperm to the single-cell level. Given the heterogeneous nature in the structure and physiology in a population of sperm, it is not surprising that aspects of acrosomal structure, including the matrix, could depend on events during spermatogenesis and could impact function at several steps in the female reproductive tract or the cumulus–oocyte complex. Furthermore, the heterogeneity in the timing and location of acrosomal exocytosis in individual cells present within a population of spermatozoa is not yet fully understood. Thus, it is a challenge to sort out real population cell behaviors (e.g., disruption of cumulus cells) from irrelevant or deleterious cell behaviors (spontaneous exocytosis, cell demise). Perhaps the most important question is, what is the location and timing of these events in the one sperm that successfully fertilizes the egg? We must also recognize that there might be multiple means (pathways) to an end (fertilization). In light of the contemporary imaging studies indicating that most mouse sperm that fertilize mouse oocytes enclosed by cumulus masses have lost the green fluorescent protein marker of intact acrosomes (Jin et al. 2011; Hirohashi et al. 2011), we recognize that work in this area is still very active so it would be premature to attempt to develop a consensus model. Nevertheless, it is useful at this time to consider results utilizing another modern approach: gene knockouts in mice. Toward this goal, recently developed, genetically modified mouse models provide valuable new insights into the molecular mechanisms of mammalian fertilization involving the sperm acrosome.
2.2 Gene Knockout Studies in Mice: What They Do and Do Not Tell Us About Acrosomal Matrix Function
The knockout of a specific reproductive gene in the mouse is usually engineered with an explicit outcome (hypothesis) in mind: to demonstrate the importance of a particular protein by interfering with fertility. However, the resulting phenotypes are often different and/or are more complicated than expected. In many cases, knockouts of individual genes encoding acrosomal matrix proteins have not caused sterility; this outcome can be explained away with the understanding that sperm function and fertilization are critical for survival of a species, so it makes sense to have built-in redundancy. Nevertheless, closer inspection of some of these “fertile” knockouts has revealed important insights into the functions of acrosomal matrix and other fertilization proteins. Also, knockouts of multiple proteins can address questions of complementary proteins in a complex system. As the body of literature involving genetically engineered mice has grown, there have been many surprising new observations that force us to think more broadly and to ask different questions than had previously been considered. For example, we now know not to assume that acrosomal exocytosis will only occur on the ZP, and we also understand proteins thought to be involved in sperm–egg interactions can have other functions such as acrosome formation, transiting the uterus and oviduct, etc. Thus, we feel the literature is rich with data from gene knockouts, and this review is an attempt to bring both widely known and understated results to the reader’s attention with hope that new avenues for research will come to light (Table 2.1).
2.3 Acrosome Formation/Globozoospermia
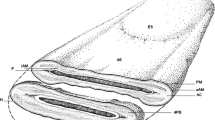
The function of the sperm acrosome is dependent upon its structure. The sperm of each species has a characteristic configuration with defined acrosomal shapes that are determined, in part, by the underlying nuclear morphologies. However, each acrosome shares the common features of an inner acrosomal membrane (IAM) that is laminated to the nuclear foundation, a fusogenic outer acrosomal membrane (OAM) that is closely apposed to the plasma membrane over the acrosome, and an equatorial segment (ES), which forms the posterior acrosomal margin where the IAM and OAM meet. Acrosomal formation begins during late meiosis when small, proacrosomal vesicles form in pachytene spermatocytes (Bloom et al. 1975; Holstein and Roosen-Runge 1981; Anakwe and Gerton 1990). After the meiotic divisions, the resulting haploid spermatids inherit the proacrosomal vesicles and continue acrosome biogenesis in earnest. Proacrosomal vesicles eventually coalesce into a single acrosomal vesicle that then attaches to and begins to spread over the future anterior segment of the nucleus. The points of contact between the developing acrosome and spermatid nucleus define the prospective IAM. The acrosome and the nucleus continue developing throughout spermiogenesis until they coordinately achieve structures similar to those found in ejaculated sperm. Final steps of metamorphosis occur during epididymal transit.
In recent years, the molecular mechanisms supporting the transmogrification of a round spermatid acrosome to the form seen in mature testicular spermatozoa have been elucidated by many studies utilizing mutations or gene knockouts in the mouse. One of the earliest examples of the effect of acrosomal dysplasia is the blind-sterile mouse, whose round spermatids exhibit disorganized proacrosomal granules with no further development beyond step 3 of spermiogenesis (Sotomayor and Handel 1986). This phenotype is caused by mutations in the Tbc1d20 gene, which encodes a GTPase-activating protein specific for Rab1 and Rab2 small GTPase families (Liegel et al. 2013). Another GTPase-activating protein, AGFG1 (also known as Hrb), specific in this case for ARF proteins, shows a similar phenotype (Kang-Decker et al. 2001; Kierszenbaum et al. 2004). Vesicle trafficking is key to formation of the acrosome since mutations affecting PICK1, the Golgi-associated PDZ- and coiled-coil motif-containing protein (GOPC), the casein kinase II α′ catalytic subunit (CSNK2A2), and islet cell autoantigen 1-like protein (ICA1L) cause sterility or subfertility in males (Xu et al. 1999; Yao et al. 2002; Xiao et al. 2009; He et al. 2015). Mutations affecting SEC23IP, a phospholipase A1-like protein that interacts with Sec23, also cause male subfertility by interfering with acrosome biogenesis and leading to round-headed spermatozoa (Arimitsu et al. 2011). Spermatids of infertile male mice lacking GalNAc transferase 3 (GALNT3) produce proacrosomal vesicles of various sizes, which attach to the nuclear envelope but do not coalesce to form a single acrosomal vesicle, apparently causing oligoasthenoteratozoospermia through a severe reduction of mucin-type O-glycans, impaired acrosome formation, and increased apoptosis (Miyazaki et al. 2012). Acrosomal dysplasia does occur in cases where certain acrosomal matrix components such as zona pellucida-binding protein 1 (ZPBP) or 2 (ZPBP2) are missing (Lin et al. 2007).
Abnormalities in nuclear envelope structure can also interfere with the ability of the acrosome to form properly. Disruption of the mouse homologue (Gmcl1) of the Drosophila melanogaster germ cell-less gene causes insufficient chromatin condensation and abnormal acrosome structures in mutant sperm (Kimura et al. 2003). When the inner nuclear membrane protein DPY19L2 is absent, the nuclear dense lamina as well as the junction between the acroplaxome and the nuclear envelope are destabilized. As a result, the linking of the acrosome and the manchette to the nucleus is blocked; this disrupts vesicular trafficking, causes sperm nuclear shaping to fail, and leads to the elimination of the unbound acrosomal vesicle (Pierre et al. 2012). In elongating spermatids, abnormalities of anterior head shaping involving CSNK2A2 or the sperm acrosome-associated 1 protein (SPACA1) also lead to globozoospermia (Xu et al. 1999; Fujihara et al. 2012). Spermatids with mutant PAFAH1B1 protein (the regulatory subunit of platelet-activating factor acetylhydrolase 1b) are sterile because the spermatids fail to form acrosomes properly and exhibit nuclei distorted in size and shape (Nayernia et al. 2003).
Final maturation of the sperm acrosome occurs within the epididymis. The classic case is the morphological alteration in the acrosome of guinea pig sperm where the head changes drastically from a planar, tennis racket-shaped structure to a form reminiscent of a cupped hand or xistera, the handheld basket used in jai alai, as the sperm transit from the testis to the cauda epididymis (Fawcett and Hollenberg 1963). Another example is seen in the post-testicular development of spermatozoa of the tammar wallaby where the immature, uncondensed acrosome has a distinctive “thumbtack” appearance that is remodeled into a streamlined, condensed structure on the sperm head (Setiadi et al. 1997). Phospholipase A2, group III (PLA2G3), which is secreted in the proximal (caput) region of the mouse epididymis, is reported to have a role in epididymal maturation of sperm (Sato et al. 2010); however, the Pla2g3 gene is also expressed in the testis, so it is difficult to attribute the subfertility of Pla2g3 −/−males to the lack of the Pla2g3 protein in the epididymis rather than the testis. The molecular mechanisms for these structural alterations are not fully understood but could be due, in part, to changes in protein disulfide cross-links, oligosaccharide modifications, and/or repackaging of the acrosomal components (Anakwe et al. 1991; Setiadi et al. 1997; Fléchon 2015).
2.4 Zona Interacting Proteins
For many years, the prevailing notion was that sperm bound to the zona pellucida of unfertilized eggs through cell surface receptors or binding proteins present on the exterior of acrosome-intact sperm. Biochemical studies in the mouse led to the identification of ZP3R (originally called “sp56”) as the putative ZP3 receptor protein (Bleil and Wassarman 1990; Cheng et al. 1994; Bookbinder et al. 1995). Subsequent experiments established that ZP3R and its guinea pig homologue are actually components of the acrosomal matrix (Foster et al. 1997; Kim et al. 2001). ZP3R monomers are assembled into a multimeric complex in the particulate acrosomal matrix. During the course of spontaneous acrosomal exocytosis and coincident with release of ZP3R from the acrosome under capacitating conditions, the monomer is proteolytically processed from 67,000 M r to 43,000 M r (Buffone et al. 2009). The processing of ZP3R is dramatically reduced when sperm are incubated under non-capacitating conditions. The cleavage probably takes place in complement control protein domain (CCP) 6 or the bridge region between CCP6 and CCP7, which is not present in the guinea pig orthologue AM67. This cleavage has functional consequences since the cleaved form of ZP3R does not bind to unfertilized eggs. However, although ZP3R binds the zona pellucida, the elimination of ZP3R in the mouse by homologous recombination did not affect fertility in the nullizygous mice (Muro et al. 2012).
Other candidates proposed as proteins responsible for zona pellucida binding also turned out to be components of the acrosomal matrix. Zona-binding proteins include the homologues known as sp38 [boar (Mori et al. 1993)], IAM38 [bull (Yu et al. 2006)], ZPBP and ZPBP2 [mouse (Lin et al. 2007)]. In the pig, this protein competes with proacrosin for binding to the zona pellucida (Mori et al. 1993). As discussed previously, elimination of the Zpbp and/or Zpbp2 genes by homologous recombination results in infertility or subfertility in male mice due to multiple effects on acrosome biogenesis and other spermatogenic defects leading to dysfunctional spermatozoa (Lin et al. 2007). The serine zymogen proacrosin is another zona-binding protein (Howes et al. 2001; Furlong et al. 2005), but, as described below, elimination of the Acr gene by homologous recombination does not cause infertility in mice (Baba et al. 1994; Adham et al. 1997). Zonadhesin (ZAN) was originally identified biochemically as a binding protein of the porcine zona pellucida (Hardy and Garbers 1995) and then demonstrated to be associated with the acrosomal matrix and membranes (Bi et al. 2003). Homologues in other species were identified and a gene knockout of Zan was performed in the mouse, which does not affect fertility but, instead, enables the mouse sperm to bind more effectively to pig, cow, and rabbit ZP without enhancing attachment to the mouse ZP, essentially eliminating the filter for the species-specific interactions between the mutant mouse sperm and oocytes of other animals (Tardif et al. 2010). From the studies performed to date, it is clear that there are several zona pellucida-binding proteins present within the acrosomal matrix of sperm. Some of these are involved in essential steps of acrosome biogenesis such as ZPBP and ZPBP2. However, the elimination of other zona-binding proteins by homologous recombination does not affect fertility per se but may yield a subtler phenotype such as a loss of species-specific sperm–zona binding or the slower dissipation of the acrosomal matrix when exocytosis is initiated, leading to a delayed rate of fertilization. These traits appear to be redundant, yet they likely impart a competitive advantage to sperm from wild-type mice when one or more males mate with the same female.
2.5 Oocyte Cumulus Complex and Zona Pellucida Transit: Proteases, Regulatory Proteins, and Binding Proteins
Mammalian oocytes are surrounded by two formidable barriers to spermatozoa: the oocyte cumulus complex (OCC) that consists of multiple layers of cumulus cells with an extracellular matrix rich in hyaluronic acid and the ZP that is comprised of three or four glycoproteins (Litscher and Wassarman 2014). The sperm surface and acrosome contain several proteases that are purported to have a role in at least some limited digestion of the OCC extracellular matrix and ZP, but it remains unclear whether these proteases are essential for sperm to transit these barriers or whether mechanical forces produced by the sperm tail are sufficient (Bedford 1998; Litscher and Wassarman 2014). In addition, the acrosomal matrix contains numerous proteins that can bind to the ZP and presumably enable a spermatozoon to burrow into the ZP by the combined mechanical forces of the tail and limited enzymatic digestion by proteases. Gene knockouts of individual and multiple proteases have provided some important and unforeseen insights into this process. Indeed, the pattern of surprising knockout results was set by perhaps the first knockout in this field—the proacrosin (ACR) gene. For many years, proacrosin was purported to be the main sperm protease for ZP penetration, but, as discussed previously, gene knockout experiments showed that proacrosin was not essential for zona penetration as mice were fertile and proacrosin-null sperm were able to transit the ZP (Baba et al. 1994). Further study showed that proacrosin-null sperm dispersed acrosomal contents more slowly than wild-type sperm, indicating that the role of acrosin was to disperse the acrosomal matrix (Yamagata et al. 1998). This also inspired research to identify novel acrosomal proteases involved in OCC and ZP transit.
PRSS21 (TESP5, testisin) is a candidate serine protease to complement acrosin; it is a GPI-linked protein on the surface of the sperm head, cytoplasmic droplet, and midpiece of the tail (Honda et al. 2002). PRSS21 knockout mice were fertile in long-term mating trials over several weeks, but in vitro analysis showed that the PRSS21-null sperm had decreased motility, abnormal morphology, and poor performance in fertilization assays (Netzel-Arnett et al. 2009).
Double knockout ACR/PRSS21 male mice were able to produce offspring in vivo but with significantly reduced litter size (Kawano et al. 2010). In vitro, ACR/PRSS21-null sperm failed to fertilize both cumulus-free and cumulus-intact oocytes, which suggests a role for these proteases in both cumulus penetration and ZP transit. Sperm penetration through the OCC was slower for ACR/PRSS21 sperm than wild-type sperm, and OCCs incubated with extracts of these sperm showed slower dispersal times than wild-type sperm extracts. Analysis of in vitro fertilization experiments showed numerous effects: the acrosomal contents of ACR/PRSS21 knockout sperm failed to disperse, the number of sperm bound to the ZP and the percentage of oocytes with fused sperm were very low, and the ratio of sperm bound to the ZP, percentage of ZP penetrated by sperm, number of sperm in the perivitelline space, and percent fertilized oocytes were zero or negligible. Scanning electron microscopy showed that oocytes incubated with ACR/PRSS21-null sperm did not have any sperm tracks, the putative paths of sperm through the ZP caused by limited enzymatic digestion. It is notable that the ACR/PRSS21 sperm appear to remain acrosome intact in these images, or perhaps they are acrosome reacted but retain a significant portion of undispersed acrosomal matrix material. The authors hypothesize that this may be due to the lack of PRSS21 activity rather than dispersal of the acrosomal matrix since ACR is known to activate the PRSS21 protease. They propose that ACR-null mice may have compensatory PRSS21 enzymatic activity for penetrating the ZP while PRSS21-null mice may have compensatory acrosomal matrix dispersal activity. ACR/PRSS21 double knockout mice lack both mechanisms and are subfertile due to the failure to fertilize all ovulated oocytes (Kawano et al. 2010). The authors propose that sperm lacking these two proteases are able to fertilize oocytes due to a compensatory mechanism in the uterine fluid; it is also tempting to speculate that at least some sperm lacking these proteases are able to fertilize oocytes due to simple mechanical forces from sperm motility. The authors suggest a role in cumulus dispersal and penetration, which is interesting since sperm lacking known hyaluronidases SPAM1 (Baba et al. 2002) and HYAL5 (Kimura et al. 2009) were able to fertilize oocytes. Together with double knockout experiments of SPAM1 and ACR or SPAM1 and PRSS21 (Zhou et al. 2012), these results suggest that ACR and PRSS21 may work in concert with hyaluronidases for cumulus penetration. These results are also consistent with the notion that acrosomal exocytosis occurs before or during interaction with the cumulus (Jin et al. 2011).
A likely substrate of acrosin in the guinea pig acrosomal matrix is AM50, the orthologue of NPTX2 (neuronal pentraxin 2 or Narp) in mice. AM50, like other pentraxins, forms large disulfide-bonded pentameric or decameric complexes and is a major structural component of the acrosomal matrix in guinea pigs (Noland et al. 1994; Reid and Blobel 1994). When acrosin is activated during acrosomal exocytosis in guinea pig sperm or in isolated acrosomal matrices, AM50 is processed at acrosin-specific sites coincident with acrosomal matrix disassembly (Westbrook-Case et al. 1994; Kim et al. 2011). NPTX2 is present in the mouse and knockout of this protein yields fertile males (Bjartmar et al. 2006), although analysis of sperm performance has not been done. Curiously, pentraxin 3 (PTX3) is produced by mouse cumulus cells during cumulus expansion and localizes in the matrix; in vivo, Ptx3 −/− female mice have been reported to be infertile or subfertile due to severe abnormalities of the cumulus oophorus, although oocytes can be successfully fertilized in vitro (Varani et al. 2002; Salustri et al. 2004).
Protein C Inhibitor (PCI), a widely expressed serine protease inhibitor in seminal plasma, is present in the acrosome, has been shown to inhibit human sperm acrosin activity, and is detected on the surface of damaged or abnormal human sperm where it is assumed to prevent inappropriate acrosin activity (Moore et al. 1993; Hermans et al. 1994; Zheng et al. 1994; Uhrin et al. 2007). PCI has also been linked to fertility in humans as infertile patients had inactive PCI (He et al. 1999). PCI-null female mice are fertile but males are infertile and the primary defect appears to be with the Sertoli cell–spermatogenic cell junctions in the testis since spermatogenic cells were prematurely released into the seminiferous tubule lumen and immature and abnormal germ cells were found in the epididymis (Uhrin et al. 2000; Uhrin et al. 2007). Further analysis of PCI-null epididymal sperm showed 95 % abnormal sperm morphology (many with separated heads and tails and abnormally shaped heads), significantly reduced motility, and failure to bind either cumulus-intact or cumulus-free eggs (Uhrin et al. 2000; Uhrin et al. 2007). Whether the observed lack of cumulus and ZP binding and penetration in absence of PCI is due to problems of unregulated acrosin activity is a question to be investigated, but it is interesting to note that ACR/PRSS21-null mice also had this phenotype. Perhaps unregulated acrosin produces premature spontaneous acrosomal exocytosis and proteolysis of the acrosomal matrix such that sperm–ZP binding sites of the acrosomal matrix and/or ZP are prematurely processed and thus nonfunctional.
The pro-protein convertase PCSK4 is reported to be localized in the developing acrosome during spermatogenesis and, in mature sperm, it is present on the plasma membrane over the acrosome (Gyamera-Acheampong et al. 2006); whether there are two PCSK4 pools (acrosomal and cell surface) or it migrates from the acrosome to the sperm surface is unclear. PCSK4-null mice are infertile (Mbikay et al. 1997; Gyamera-Acheampong et al. 2006) and have abnormal acrosomal shapes (Tardif et al. 2012) that are likely caused by reduced ACRBP processing during spermiogenesis that leads to improper packaging of proacrosin and perhaps other acrosomal matrix proteins into the acrosome (Kanemori et al. 2013). PCSK4-null mature sperm had accelerated tyrosine phosphorylation during capacitation, improper processing of the egg-binding protein ADAM2, increased spontaneous acrosomal exocytosis, and reduced ZP binding. Thus, PCSK4 has multiple functions in developing and mature sperm and identification of substrate proteins will help elucidate the mechanisms through which this protein functions.
2.6 Inner Acrosomal Membrane Proteins
As acrosomal exocytosis proceeds and acrosomal matrix proteins are released, some components of the IAM remain associated with the sperm and are in position to interact with egg vestments. The term “secondary binding” has been used to describe the interactions of these proteins with the ZP, particularly ZP2, but we envision these interactions to be less well defined and to involve multiple binding and release events. In this model, sperm would adhere to the ZP via multiple proteins that are then released as a result of limited proteolysis to produce repeated binding and release at the molecular level; the continual force of the tail would drive the head progressively further into the ZP. Numerous proteins have the ZP-binding and/or enzymatic properties that are consistent with this model.
The cellular differentiation marker CD46 is ubiquitously expressed on the surface of most mammalian cells and is involved in complement protection in many species, including humans, but not in rats, mice, and guinea pigs. In mice, it appears to be expressed solely in the testis, and, in sperm, CD46 is not on the PM as would be expected, but is located inside the acrosome on the IAM. It was thought that this might be to compensate for loss of sperm surface CD proteins that protect the cell from attack by complement, but sperm from CD46 knockout mice are not more susceptible to complement than control sperm, so the function of CD46 is not likely to be complement protection. Interestingly, CD46 knockout mouse sperm have enhanced fertility; the sperm show accelerated spontaneous acrosomal exocytosis and have an elevated litter size in one mouse strain (DBA/2N), and CD46-null sperm fertilized superovulated oocytes at a significantly higher rate than controls (Inoue et al. 2003). Thus, CD46 appears to be important in stabilizing the acrosomal membranes. How it does this from the inner acrosomal membrane is a mystery, but it is possible that, during acrosome formation in spermatogenesis, CD46 may interact with other proteins destined for the periacrosomal membranes. Consequently, CD46 might direct or influence sorting of proteins that stabilize these membranes so that, in the absence of CD46, these proteins are misdirected or do not function properly to stabilize the acrosomal membranes.
Several IAM proteins are known to have a role in sperm–ZP binding but do not have knockout models yet. Yu et al. (2006) also have extracted proteasomal activity from the IAM, but, again, there is no gene knockout data available with regard to its role in fertilization. SPACA4 (SAMP14) is present in the IAM, OAM, and ES of human sperm; it is GPI-linked and is exposed after acrosomal exocytosis (Shetty et al. 2003). Antibodies to recombinant SPACA4 inhibit binding/fusion of human sperm to zona-free hamster eggs, but a knockout mouse line has not been generated.
2.7 Equatorial Segment Proteins
Equatorin (EQTN) localization is limited to the equatorial segment and the protein is resistant to fairly strong extraction conditions (high salt, 0.1 % Triton X-100, sonication), persists following acrosomal exocytosis on sperm in the perivitelline space, and is even present on sperm inside the egg (Manandhar and Toshimori 2001). Following acrosomal exocytosis, some EQTN is redistributed onto the PM over the ES (and some on the IAM), but the significance of this is not clear. Male EQTN-null mice are subfertile (~50 % of controls) and have reduced litter sizes (5.8 vs. 12.3 in control). EQTN-null sperm undergo normal acrosomal biogenesis (acrosomes are normal by immunofluorescence and by transmission electron microscopy), but do not undergo acrosomal exocytosis above control levels. Co-immunoprecipitation with SNAP25 and syntaxin 1a suggests a role in vesicle fusion during acrosomal exocytosis, but since the protein does not have a canonical transmembrane domain, it must do this via a membrane protein that interacts with the vesicle fusion proteins (Wolkowicz et al. 2003; Hao et al. 2014). Also, since the outer acrosomal and plasma membranes don’t fuse in the ES, EQTN is not likely to be involved in fusion in that region. The reason why EQTN affects fusion of the OAM and PM in the more anterior zones is an area for more investigation.
Sperm equatorial segment protein 1 (SPESP1) is another ES-limited protein that is important for fertility in both expected and surprising ways. SPESP1-null and heterozygous males had delays in fertility and smaller litter sizes than wild-type males, and heterozygous and null sperm numbers in the oviduct were significantly reduced (Fujihara et al. 2010). Interestingly, in vitro assays showed that the number of sperm, percent progressively motile sperm, and motility patterns were the same for SPESP1-null, heterozygous, and wild-type mice, but the percent of fertilized oocytes and the percent fusibility with zona-free oocytes were significantly less in SPESP1-null and heterozygotes than wild type. Furthermore, in SPESP1-null sperm following acrosomal exocytosis, the plasma and outer acrosomal membranes were found to undergo fusion throughout the entire equatorial segment; since these membranes are typically nonfusogenic, these results show that SPESP1 is important in maintaining the stability of the ES which has an impact on fusibility with the egg plasma membrane (Fujihara et al. 2010).
2.8 Ten Questions
In light of recent developments in the field of fertilization using contemporary imaging techniques and molecular biology to create targeted mutations in genes considered to be important for fertilization, several issues remain that need to be addressed. These include, but are not limited to, the following ten questions: (1) Does acrosomal exocytosis need to occur in a specific region of the female reproductive tract or peri-oocyte environment for fertilization to take place? Current evidence suggests that most sperm have initiated acrosomal exocytosis prior to penetrating the zona pellucida of cumulus-enclosed oocytes. (2) Is binding of the zona pellucida by acrosomal matrix components exposed on the sperm surface required for fertilization or is the affinity of these proteins assistive but not essential for fertilization to take place? For example, ZP3R binds to the oocyte zona pellucida but is not required for fertilization. On the other hand, in the absence of ZAN, the species-specific filter that blocks heterospecific sperm from other phylogenetic classes from binding to mouse oocytes is lost; it would be interesting to determine if ZAN operates to filter the binding of sperm related at the genus or strain level. (3) Can we develop better tools to assess the integrity of the membranes overlying the acrosomal contents during various stages of capacitation and acrosomal exocytosis? (4) Do the components of the acrosome influence the interactions of sperm with the oviductal epithelium, either independently on a single-cell level or perhaps more generally through influencing the reciprocal connectivity between neighboring sperm and the female reproductive tract? (5) Do “acrosome-reacted” sperm that are experimentally recovered from the perivitelline space and are used successfully to fertilize other eggs still contain functional acrosomal matrix material? These experiments raise questions concerning the functions of the acrosomal contents in fertilization (they may be assistive but not required). (6) Are the different terms used to describe the acrosomal matrix important or are they describing the same properties from varied perspectives? Some reports refer to the acrosomal matrix as a particulate compartment of the acrosomal contents (Buffone et al. 2008). Others make reference to a crystalloid nature observed by transmission electron microscopy (Fléchon 2015). Still others classify the acrosomal matrix as an amyloid (Guyonnet et al. 2014). (7) What role does pH play in the intra-acrosomal dynamics and post-exocytotic dispersion of the contents? For example, the pH of the acrosome is acidic and gradually begins to alkalinize during the course of epididymal maturation; once exocytosis is initiated, the contents are rapidly exposed to the more alkaline pH of the milieu surrounding the sperm. (8) Does the change in pH occurring during these periods activate or inactivate acrosomal enzymes? This is true for some proteases such as the proacrosin zymogen activation, but there may be other enzymes exhibiting similar properties. (9) What are the substrates for acrosomal hydrolases? Initially, it was thought that acrosin acted upon the zona pellucida. However, it is clear that this serine protease is involved in the dispersion of the acrosomal matrix, but the actual substrates have yet to be elucidated. (10) Why are acrosomal proteins differentially compartmentalized? The component segregation could be important for differential binding and/or release during exocytosis. Alternatively, the different domains might simply influence the shape of the acrosome. Clearly, there is still much to learn concerning the involvement of acrosomal matrix components in directing the structure of the acrosome during spermatogenesis and epididymal maturation as well as the function of the acrosome in the fertilization process.
References
Adham IM, Nayernia K, Engel W (1997) Spermatozoa lacking acrosin protein show delayed fertilization. Mol Reprod Dev 46:370–376. doi:10.1002/(SICI)1098-2795(199703)46:<370::AID-MRD16>3.0.CO;2-2
Anakwe OO, Gerton GL (1990) Acrosome biogenesis begins during meiosis: Evidence from the synthesis and distribution of an acrosomal glycoprotein, acrogranin, during guinea pig spermatogenesis. Biol Reprod 42:317–328
Anakwe OO, Sharma S, Hoff HB et al (1991) Maturation of guinea pig sperm in the epididymis involves the modification of proacrosin oligosaccharide side chains. Mol Reprod Dev 29:294–301. doi:10.1002/mrd.1080290313
Arimitsu N, Kogure T, Baba T et al (2011) p125/Sec23-interacting protein (Sec23ip) is required for spermiogenesis. FEBS Lett 585:2171–2176. doi:10.1016/j.febslet.2011.05.050
Baba T, Azuma S, Kashiwabara S, Toyoda Y (1994) Sperm from mice carrying a targeted mutation of the acrosin gene can penetrate the oocyte zona pellucida and effect fertilization. J Biol Chem 269:31845–31849
Baba D, Kashiwabara S, Honda A et al (2002) Mouse sperm lacking cell surface hyaluronidase PH-20 can pass through the layer of cumulus cells and fertilize the egg. J Biol Chem 277:30310–30314. doi:10.1074/jbc.M204596200
Bedford JM (1998) Mammalian fertilization misread? Sperm penetration of the eutherian zona pellucida is unlikely to be a lytic event. Biol Reprod 59:1275–1287. doi:10.1095/biolreprod59.6.1275
Bi M, Hickox JR, Winfrey VP et al (2003) Processing, localization and binding activity of zonadhesin suggest a function in sperm adhesion to the zona pellucida during exocytosis of the acrosome. Biochem J 375:477–488. doi:10.1042/bj20030753
Bjartmar L, Huberman AD, Ullian EM et al (2006) Neuronal pentraxins mediate synaptic refinement in the developing visual system. J Neurosci 26:6269–6281. doi:10.1523/JNEUROSCI.4212-05.2006
Bleil JD, Wassarman PM (1990) Identification of a ZP3-binding protein on acrosome-intact mouse sperm by photoaffinity crosslinking. Proc Natl Acad Sci 87:5563–5567
Bloom W, Fawcett DW, Maximow AA (1975) A textbook of histology, 10th edn. Saunders, Philadelphia
Bookbinder LH, Cheng A, Bleil JD (1995) Tissue- and species-specific expression of sp56, a mouse sperm fertilization protein. Science 269:86–89
Buffone MG, Foster JA, Gerton GL (2008) The role of the acrosomal matrix in fertilization. Int J Dev Biol 52:511–522. doi:10.1387/ijdb.072532mb
Buffone MG, Kim K-S, Doak BJ et al (2009) Functional consequences of cleavage, dissociation and exocytotic release of ZP3R, a C4BP-related protein, from the mouse sperm acrosomal matrix. J Cell Sci 122:3153–3160. doi:10.1242/jcs.052977
Cheng A, Le T, Palacios M et al (1994) Sperm-egg recognition in the mouse: Characterization of sp56, a sperm protein having specific affinity for ZP3. J Cell Biol 125:867–878
Fawcett DW, Hollenberg RD (1963) Changes in the acrosome of the guinea pig spermatozoa during passage through the epididymis. Z Zellforsch Mikrosk Anat 60:276–292
Fléchon J-E (2015) The acrosome of eutherian mammals. Cell Tissue Res 363:147–157. doi: 10.1007/s00441-015-2238-0
Foster JA, Friday BB, Maulit MT et al (1997) AM67, a secretory component of the guinea pig sperm acrosomal matrix, is related to mouse sperm protein sp56 and the complement component 4-binding proteins. J Biol Chem 272:12714–12722
Fujihara Y, Murakami M, Inoue N et al (2010) Sperm equatorial segment protein 1, SPESP1, is required for fully fertile sperm in mouse. J Cell Sci 123:1531–1536. doi:10.1242/jcs.067363
Fujihara Y, Satouh Y, Inoue N et al (2012) SPACA1-deficient male mice are infertile with abnormally shaped sperm heads reminiscent of globozoospermia. Development 139:3583–3589. doi:10.1242/dev.081778
Furlong LI, Harris JD, Vazquez-Levin MH (2005) Binding of recombinant human proacrosin/acrosin to zona pellucida (ZP) glycoproteins. I. Studies with recombinant human ZPA, ZPB, and ZPC. Fertil Steril 83:1780–1790. doi:10.1016/j.fertnstert.2004.12.042
Guyonnet B, Egge N, Cornwall GA (2014) Functional amyloids in the mouse sperm acrosome. Mol Cell Biol 34:2624–2634. doi:10.1128/MCB.00073-14
Gyamera-Acheampong C, Tantibhedhyangkul J, Weerachatyanukul W et al (2006) Sperm from mice genetically deficient for the PCSK4 proteinase exhibit accelerated capacitation, precocious acrosome reaction, reduced binding to egg zona pellucida, and impaired fertilizing ability. Biol Reprod 74:666–673. doi:10.1095/biolreprod.105.046821
Hao J, Chen M, Ji S et al (2014) Equatorin is not essential for acrosome biogenesis but is required for the acrosome reaction. Biochem Biophys Res Commun 444:537–542. doi:10.1016/j.bbrc.2014.01.080
Hardy DM, Garbers DL (1995) A sperm membrane protein that binds in a species-specific manner to the egg extracellular matrix is homologous to von Willebrand factor. J Biol Chem 270:26025–26028. doi:10.1074/jbc.270.44.26025
He S, Lin Y-L, Liu Y-X (1999) Functionally inactive protein C inhibitor in seminal plasma may be associated with infertility. Mol Hum Reprod 5:513–519. doi:10.1093/molehr/5.6.513
He J, Xia M, Tsang WH et al (2015) ICA1L forms BAR-domain complexes with PICK1 and is crucial for acrosome formation in spermiogenesis. J Cell Sci 128:3822–3836. doi:10.1242/jcs.173534
Hermans JM, Jones R, Stone SR (1994) Rapid inhibition of the sperm protease acrosin by protein C inhibitor. Biochemistry (Mosc) 33:5440–5444. doi:10.1021/bi00184a012
Hirohashi N, Gerton GL, Buffone MG (2011) Video imaging of the sperm acrosome reaction during in vitro fertilization. Commun Integr Biol 4:471–476. doi:10.4161/cib.4.4.15636
Holstein AF, Roosen-Runge EC (1981) Atlas of human spermatogenesis. Grosse, Berlin
Honda A, Yamagata K, Sugiura S et al (2002) A mouse serine protease TESP5 is selectively included into lipid rafts of sperm membrane presumably as a glycosylphosphatidylinositol-anchored protein. J Biol Chem 277:16976–16984
Howes E, Pascall JC, Engel W, Jones R (2001) Interactions between mouse ZP2 glycoprotein and proacrosin; a mechanism for secondary binding of sperm to the zona pellucida during fertilization. J Cell Sci 114:4127–4136
Inoue N, Ikawa M, Nakanishi T et al (2003) Disruption of mouse CD46 causes an accelerated spontaneous acrosome reaction in sperm. Mol Cell Biol 23:2614–2622. doi:10.1128/MCB.23.7.2614-2622.2003
Jin M, Fujiwara E, Kakiuchi Y et al (2011) Most fertilizing mouse spermatozoa begin their acrosome reaction before contact with the zona pellucida during in vitro fertilization. Proc Natl Acad Sci 108:4892–4896. doi:10.1073/pnas.1018202108
Kanemori Y, Ryu J-H, Sudo M et al (2013) Two functional forms of ACRBP/sp32 are produced by pre-mRNA alternative splicing in the mouse. Biol Reprod 88:105. doi:10.1095/biolreprod.112.107425
Kang-Decker N, Mantchev GT, Juneja SC et al (2001) Lack of acrosome formation in Hrb-deficient mice. Science 294:1531–1533. doi:10.1126/science.1063665
Kawano N, Kang W, Yamashita M et al (2010) Mice lacking two sperm serine proteases, ACR and PRSS21, are subfertile, but the mutant sperm are infertile in vitro. Biol Reprod 83:359–369. doi:10.1095/biolreprod.109.083089
Kierszenbaum AL, Tres LL, Rivkin E et al (2004) The acroplaxome is the docking site of Golgi-derived myosin Va/Rab27a/b-containing proacrosomal vesicles in wild-type and Hrb mutant mouse spermatids. Biol Reprod 70:1400–1410. doi:10.1095/biolreprod.103.025346
Kim KS, Cha MC, Gerton GL (2001) Mouse sperm protein sp56 is a component of the acrosomal matrix. Biol Reprod 64:36–43
Kim K-S, Foster JA, Kvasnicka KW, Gerton GL (2011) Transitional states of acrosomal exocytosis and proteolytic processing of the acrosomal matrix in guinea pig sperm. Mol Reprod Dev 78:930–941. doi:10.1002/mrd.21387
Kimura T, Ito C, Watanabe S et al (2003) Mouse germ cell-less as an essential component for nuclear integrity. Mol Cell Biol 23:1304–1315. doi:10.1128/MCB.23.4.1304-1315.2003
Kimura M, Kim E, Kang W et al (2009) Functional roles of mouse sperm hyaluronidases, HYAL5 and SPAM1, in fertilization. Biol Reprod 81:939–947. doi:10.1095/biolreprod.109.078816
Liegel RP, Handley MT, Ronchetti A et al (2013) Loss-of-function mutations in TBC1D20 cause cataracts and male infertility in blind-sterile mice and Warburg micro syndrome in humans. Am J Hum Genet 93:1001–1014. doi:10.1016/j.ajhg.2013.10.011
Lin Y-N, Roy A, Yan W et al (2007) Loss of zona pellucida binding proteins in the acrosomal matrix disrupts acrosome biogenesis and sperm morphogenesis. Mol Cell Biol 27:6794–6805. doi:10.1128/MCB.01029-07
Litscher ES, Wassarman PM (2014) Evolution, structure, and synthesis of vertebrate egg-coat proteins. Trends Dev Biol 8:65–76
Manandhar G, Toshimori K (2001) Exposure of sperm head equatorin after acrosome reaction and its fate after fertilization in mice. Biol Reprod 65:1425–1436. doi:10.1095/biolreprod65.5.1425
Mbikay M, Tadros H, Ishida N et al (1997) Impaired fertility in mice deficient for the testicular germ-cell protease PC4. Proc Natl Acad Sci 94:6842–6846
Miyazaki T, Mori M, Yoshida CA et al (2012) Galnt3 deficiency disrupts acrosome formation and leads to oligoasthenoteratozoospermia. Histochem Cell Biol 139:339–354. doi:10.1007/s00418-012-1031-3
Moore A, Penfold LM, Johnson JL et al (1993) Human sperm–egg binding is inhibited by peptides corresponding to core region of an acrosomal serine protease inhibitor. Mol Reprod Dev 34:280–291. doi:10.1002/mrd.1080340308
Mori E, Baba T, Iwamatsu A, Mori T (1993) Purification and characterization of a 38-kDa protein, sp38, with zona pellucida-binding property from porcine epididymal sperm. Biochem Biophys Res Commun 196:196–202. doi:10.1006/bbrc.1993.2234
Muro Y, Buffone MG, Okabe M, Gerton GL (2012) Function of the acrosomal matrix: zona pellucida 3 receptor (ZP3R/sp56) is not essential for mouse fertilization. Biol Reprod 86:1–6. doi:10.1095/biolreprod.111.095877
Nayernia K, Vauti F, Meinhardt A et al (2003) Inactivation of a testis-specific Lis1 transcript in mice prevents spermatid differentiation and causes male infertility. J Biol Chem 278:48377–48385. doi:10.1074/jbc.M309583200
Netzel-Arnett S, Bugge TH, Hess RA et al (2009) The glycosylphosphatidylinositol-anchored serine protease PRSS21 (testisin) imparts murine epididymal sperm cell maturation and fertilizing ability. Biol Reprod 81:921–932. doi:10.1095/biolreprod.109.076273
Noland TD, Friday BB, Maulit MT, Gerton GL (1994) The sperm acrosomal matrix contains a novel member of the pentaxin family of calcium-dependent binding proteins. J Biol Chem 269:32607–32614
Pierre V, Martinez G, Coutton C et al (2012) Absence of Dpy19l2, a new inner nuclear membrane protein, causes globozoospermia in mice by preventing the anchoring of the acrosome to the nucleus. Development 139:2955–2965. doi:10.1242/dev.077982
Reid MS, Blobel CP (1994) Apexin, an acrosomal pentaxin. J Biol Chem 269:32615–32620
Salustri A, Garlanda C, Hirsch E et al (2004) PTX3 plays a key role in the organization of the cumulus oophorus extracellular matrix and in vivo fertilization. Development 131:1577–1586. doi:10.1242/dev.01056
Sato H, Taketomi Y, Isogai Y et al (2010) Group III secreted phospholipase A2 regulates epididymal sperm maturation and fertility in mice. J Clin Invest 120:1400–1414. doi:10.1172/JCI40493
Setiadi D, Lin M, Rodger JC (1997) Posttesticular development of spermatozoa of the tammar wallaby (Macropus eugenii). J Anat 190:275–288. doi:10.1046/j.1469-7580.1997.19020275.x
Shetty J, Wolkowicz MJ, Digilio LC et al (2003) SAMP14, a novel, acrosomal membrane-associated, glycosylphosphatidylinositol-anchored member of the Ly-6/urokinase-type plasminogen activator receptor superfamily with a role in sperm-egg Interaction. J Biol Chem 278:30506–30515. doi:10.1074/jbc.M301713200
Sotomayor RE, Handel MA (1986) Failure of acrosome assembly in a male sterile mouse mutant. Biol Reprod 34:171–182. doi:10.1095/biolreprod34.1.171
Tardif S, Wilson MD, Wagner R et al (2010) Zonadhesin is essential for species specificity of sperm adhesion to the egg’s zona pellucida. J Biol Chem 285(32):24863–24870. doi:10.1074/jbc.M110.123125
Tardif S, Guyonnet B, Cormier N, Cornwall GA (2012) Alteration in the processing of the ACRBP/sp32 protein and sperm head/acrosome malformations in proprotein convertase 4 (PCSK4) null mice. Mol Hum Reprod 18:298–307. doi:10.1093/molehr/gas009
Uhrin P, Dewerchin M, Hilpert M et al (2000) Disruption of the protein C inhibitor gene results in impaired spermatogenesis and male infertility. J Clin Invest 106:1531–1539. doi:10.1172/JCI10768
Uhrin P, Schöfer C, Zaujec J et al (2007) Male fertility and protein C inhibitor/plasminogen activator inhibitor-3 (PCI): localization of PCI in mouse testis and failure of single plasminogen activator knockout to restore spermatogenesis in PCI-deficient mice. Fertil Steril 88:1049–1057. doi:10.1016/j.fertnstert.2006.11.193
Varani S, Elvin JA, Yan C et al (2002) Knockout of pentraxin 3, a downstream target of growth differentiation factor-9, causes female subfertility. Mol Endocrinol 16:1154–1167. doi:10.1210/me.16.6.1154
Wassarman PM, Jovine L, Litscher ES et al (2004) Egg–sperm interactions at fertilization in mammals. Eur J Obstet Gynecol Reprod Biol 115(Suppl 1):S57–S60. doi:10.1016/j.ejogrb.2004.01.025
Westbrook-Case VA, Winfrey VP, Olson GE (1994) A domain-specific 50-kilodalton structural protein of the acrosomal matrix is processed and released during the acrosome reaction in the guinea pig. Biol Reprod 51:1–13. doi:10.1095/biolreprod51.1.1
Wolkowicz MJ, Shetty J, Westbrook A et al (2003) Equatorial segment protein defines a discrete acrosomal subcompartment persisting throughout acrosomal biogenesis. Biol Reprod 69:735–745. doi:10.1095/biolreprod.103.016675
Xiao N, Kam C, Shen C et al (2009) PICK1 deficiency causes male infertility in mice by disrupting acrosome formation. J Clin Invest 119:802–812. doi:10.1172/JCI36230
Xu X, Toselli PA, Russell LD, Seldin DC (1999) Globozoospermia in mice lacking the casein kinase II α′ catalytic subunit. Nat Genet 23:118–121. doi:10.1038/12729
Yamagata K, Murayama K, Okabe M et al (1998) Acrosin accelerates the dispersal of sperm acrosomal proteins during acrosome reaction. J Biol Chem 273:10470–10474. doi:10.1074/jbc.273.17.10470
Yao R, Ito C, Natsume Y et al (2002) Lack of acrosome formation in mice lacking a Golgi protein, GOPC. Proc Natl Acad Sci 99:11211–11216. doi:10.1073/pnas.162027899
Yu Y, Xu W, Yi Y-J et al (2006) The extracellular protein coat of the inner acrosomal membrane is involved in zona pellucida binding and penetration during fertilization: characterization of its most prominent polypeptide (IAM38). Dev Biol 290:32–43. doi:10.1016/j.ydbio.2005.11.003
Zheng X, Geiger M, Ecke S et al (1994) Inhibition of acrosin by protein C inhibitor and localization of protein C inhibitor to spermatozoa. Am J Physiol 267:C466–C472
Zhou C, Kang W, Baba T (2012) Functional characterization of double-knockout mouse sperm lacking SPAM1 and ACR or SPAM1 and PRSS21 in fertilization. J Reprod Dev 58:330–337. doi:10.1262/jrd.2011-006
Acknowledgments
This work was supported in part by grants R21-ES024527 and P30-ES013508 from the National Institute of Environmental Health Sciences to GLG, and Chenery and Rashkind Research Grants from Randolph-Macon College to JAF.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Foster, J.A., Gerton, G.L. (2016). The Acrosomal Matrix. In: Buffone, M. (eds) Sperm Acrosome Biogenesis and Function During Fertilization. Advances in Anatomy, Embryology and Cell Biology, vol 220. Springer, Cham. https://doi.org/10.1007/978-3-319-30567-7_2
Download citation
DOI: https://doi.org/10.1007/978-3-319-30567-7_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-30565-3
Online ISBN: 978-3-319-30567-7
eBook Packages: MedicineMedicine (R0)