
Abstract
Skilled, fluent reading involves mastery of multiple linguistic (e.g., phonological, semantic, morphological, syntactic) and related cognitive processes (e.g., processing speed, attention, working memory). Development of these processes is highly predictive of fluent reading. Among these, impairments in phonological processes such as phonological awareness and decoding (learning of spelling-to-sound correspondences) in combination with processing speed impairments are often implicated as the primary underlying deficits in developmental dyslexia. Here, we review relevant behavioral research on aspects of fluent reading development (phonology, semantics, processing speed) in skilled and disordered reading. We describe recent research in mapping the development of neural systems underlying these reading-related capacities, and their relevant gene-brain-behavior underpinnings. We consider the impact of cross-linguistic and/or multilingual experience on the development of the brain’s reading circuitry. By incorporating behavioral, neural, and genetic research, across different populations (skilled and disordered reading, monolingual and multilingual reading), and across languages, these multiple sources of converging evidence inform the development of fluent reading and contribute to the goal of developing a comprehensive neurobiological model of reading.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Reading fluency
- Neural systems
- Gene-brain-behavior
- Cross-language reading
- Bilingual reading
- Reading disorders
- Phonology
- fMRI
- fNIRS
- Neuroimaging
1 Introduction
Fluent reading is a product of skilled decoding and a complex process involving multiple underlying linguistic and cognitive components. The bulk of reading research, both on typical reading development and on disordered/delayed reading development, has primarily addressed phonological processing (including phonological awareness and phonological working memory) and naming speed (Arnell, Joanisse, Klein, Busseri, & Tannock, 2009; Swanson, Trainin, Necoechea, & Hammill, 2003). Children with developmental dyslexia can often be characterized by a discrete deficit in either phonological processing or processing speed, or a combination of both (McCardle, Scarborough, & Catts, 2001). Although phonology and processing speed appear to be most compromised in reading disabled cohorts, all aspects of language, including orthography, morphology, syntax and semantics are crucial for fluent reading. Aspects of cognition such as attention and memory are likewise crucial components of the development of skilled reading (Pugh et al., 2013). Early development of these capacities is linked to reading mastery and fluency in important ways. In this chapter, we describe our laboratory’s recent research in mapping the neural systems underlying these reading-related capacities and our advances in developing a neurobiological model of reading. A neurobiological model of how multiple linguistic and cognitive systems give rise to reading crucially must look beyond monolingual English reading development in order to account for both universal and language-specific reading phenomena. An adequate model must consider cross-linguistic differences, as well as bilingual and multilingual experiences, and their impact on the development of the brain’s reading circuitry. In our goal to develop a comprehensive model, we begin by reviewing relevant behavioral research on component processing in fluent reading, with specific focus on phonology, semantics, and naming speed. We then review current neurobiological findings relevant to gene-brain-behavior underpinnings of skilled and disabled reading in contrastive languages as well as monolingual versus bilingual populations, with an emphasis on neuroimaging research that permit such insights. We will finally conclude with further directions and new research questions in the cognitive neuroscience of reading.
2 Phonology
Understanding the role of phonology in reading is crucial if we hope to interpret the neural correlates of phonological processing in the brain’s reading circuit. Phonology has received much attention in reading research because of its importance in the very early stages of learning to read. Children’s awareness of and ability to manipulate the sound units in their native language, termed phonological awareness, is a strong predictor of later reading development (Bowey, Cain, & Ryan, 1992; Foy & Mann, 2006; Goswami & Bryant, 1990; Hatcher & Hulme, 1999; Hoien, Lundberg, Stanovich, & Bjaalid, 1995; Hulme et al., 2002, Hulme, Caravolas, Málkova, & Brigstocke, 2005; Melby-Lervåg, Lyster, & Hulme, 2012; Muter, Hulme, Snowling, & Taylor, 1997; Nation & Hulme, 1997; Nation & Snowling, 2004; Preston et al., 2010; Pugh et al., 2013; Wagner & Torgesen, 1987; Weber-Fox & Neville, 1999, 2001). A child’s phonological awareness at the point in development when literacy is emerging (ages 4–6) predicts later literacy outcomes (Goswami & Bryant, 1990; Hulme et al., 2002; Wagner & Torgesen, 1987; Wagner et al., 1997; Ziegler & Goswami, 2005).
Good phonological awareness allows the subsequent understanding of the alphabetic principle in the beginning reader (Liberman, Shankweiler, & Liberman, 1989). Understanding that phonemes can correspond to letters is an important step for learning to read in alphabetic systems. The learning of spelling-to-sound correspondences allows the young reader to successfully decode an unfamiliar word by “sounding out” each letter (Share, 1995). When a child is faced with the task of recognizing a printed word, s/he can assemble a phonological representation by mapping from the orthographic units to their corresponding phonological forms, often referred to as the phonological route, (e.g., Frost, 1998). Indeed, there is evidence that all young readers access phonological representations in association with print for all languages (Georgiou, Torppa, Manolitsis, Lyytinen, & Parrila, 2012; Goswami, 2008; Panah & Padakannaya, 2008). With increased experience older, skilled, readers may come to rely to an increasing degree on direct associations between orthography and semantics (Share, 1995), the so-called lexical route. The phonological route remains relevant in reading frequent words as reading fluency develops. Indeed, there is evidence that phonological activation is both early and automatic in skilled reading (see Frost, 1998 for a review), and various experimental paradigms such as backward masking and priming, indicate that lexical access continues to be constrained by phonology (Lukatela & Turvey, 1994; Perfetti & Bell, 1991). In short, there is good evidence that phonology is still playing an important but more automated (faster) role – and importantly, if the phonological mapping is never mastered, reading will continue to be slow and labored (Frost, 1998; Pugh et al., 2013).
We must also consider differences among written languages. Languages vary in the degree to which spelling-to-sound mappings are multivalent; this has been referred to as orthographic depth (Frost, Katz, & Bentin, 1987). For example, languages such as Spanish, Italian and German have nearly one-to-one spelling-to-sound correspondences, termed consistent or transparent orthographies (Katz & Feldman, 1983). Many alphabetic languages have a transparent orthography, whereas English orthography is considered an opaque orthography with many inconsistencies in spelling-to-sound mappings (e.g., the letter ‘c’ can correspond to both the phoneme /s/ and /k/ as in ‘circus’). Irrespective of such cross-linguistic variation in orthographic depth, phonology is a key component of learning to read in both transparent and opaque orthographies (Caravolas, Lervag, Defior, Seidlova Malkova, & Hulme, 2013; Cossu, Shankweiler, Liberman, Katz, & Tola, 1988; Katz & Frost, 1992). Moreover, this general principle holds true across different writing systems, for example, alphabetic languages such as English and logographic languages such as Chinese (Lo, Hue, & Tsai, 2007; Wang, Yang, & Cheng, 2009).
A set of contemporary computational models of reading, known collectively as “triangle models of reading”, postulate that reading primarily consists of processes distributed over three levels of linguistic representation: orthography, phonology and semantics (Harm & Seidenberg, 2004). In order to decode a written word and access its corresponding meaning, orthographic networks activate corresponding phonologic and semantic networks. Skilled reading involves both the direct orthography-semantic connections as well as the orthography-phonology-semantics connections, and while the division of labour among these pathways changes over development all connections remain crucial to skilled word identification (Harm & Seidenberg, 1999, 2004).
There is considerable agreement that phonological processing deficits have a central role in reading impairments (Bradley & Bryant, 1983; Goswami, 2003; Goswami, Gerson, & Astruc, 2010; Landi, Mencl, Frost, Sandak, & Pugh, 2010; Lavidor, Johnston, & Snowling, 2006; Pugh et al., 2001; Torgesen, Wagner, & Rashotte, 1994; Wolf & Bowers, 1999). However, there is also evidence that children with reading impairments show deficits not limited to phonology (Ackerman & Dykman, 1993; Wolf & Bowers, 1999). Minimally, the processes contributing to successful reading include (1) attention to visual letters and visual processes involved in feature detection, and letter and letter-pattern identification, (2) the integration of this visual information with orthographic representations, (3) integration of orthographic representations with phonological representations, and (4) speed of activation of semantic and lexical information (Grainger & Holcomb, 2009; Seidenberg, Bruck, Fornarolo, & Backman, 1985; Wolf & Bowers, 1999). Theoretically, and practically, any of these component processes can contribute to reading impairments.
Thus, processes underlying rapid word recognition can show impairments that are not entirely reducible to phonological skills such as phonological awareness, indicating that indeed other aspects of language and cognition contribute to the development of reading. For instance, Wolf and Bowers (1999) offer a double-deficit hypothesis for reading impairment that implicates both phonological and naming speed as dissociable sources of reading difficulty. Here, phonological processing represents one side of the deficit, whereas naming speed represents another side.
3 Naming Speed
Although phonological processing deficits have received the lion’s share of research focus in reading sciences, fast, automatic item retrieval and naming are also key components of skilled and fluent reading (Wolf & Bowers, 1999). Naming speed is not entirely dissociable from phonological processing because the phonological representation of a given word is continuously accessed during reading. In order for a child to successful read a printed word, s/he must recognize the target word and retrieve it from her lexicon to access meaning. Moreover, this process progressively becomes automated as a child progresses from a young beginning reader to an older more skilled reader. Skilled and fluent reading involves additional processing beyond the routinization of spelling-to-sound correspondence learning. Strong vocabulary knowledge is related to word reading mastery (Anderson & Freebody, 1983; Berends & Reitsma, 2006; Muter, Hulme, Snowling, & Stevenson, 2004; Oullette, 2006; Roth, Speece, & Cooper, 2002; Share & Leikin, 2004; Snow, Tabors, Nicholson, & Kurland, 1995). Familiarity with words may permit the child to access the lexical entry with greater weighting towards orthographic-to-semantic connections. Thus, semantic knowledge provides an important link between decoding a word and reading comprehension. For example, Berends and Reitsma (2006) observed that practice with printed words, with specific instructions focusing on the semantic characteristics of the word, promotes reading acquisition. Moreover, the size of a child’s vocabulary is related to the ability to understand printed words (Anderson & Freebody, 1983; Proctor, August, Carlo, & Snow, 2006; Rolla San Francisco, Mo, Carlo, August, & Snow, 2006). The importance of semantic knowledge for skilled reading is well expressed by Perfetti and Hart (2002) and Perfetti (2007) in the Lexical Quality Hypothesis in which word knowledge is composed of multiple dimensions (phonology, orthography, semantics) and coherent representations of all these dimensions contribute to skilled reading.
As noted, retrieval speed can impact reading skill with measures such as Rapid Automatized Naming (RAN) shown to be a contributing factor to fluent word reading (Wolf & Bowers, 1999). Deficits in this capacity are characteristic of reading disability (Bowers & Swanson, 1991). Specifically, RAN tasks measure the speed and accuracy of naming an array of familiar stimuli such as digits, letters, or colors (Denckla & Rudel, 1974). Scores on such tests are consistently correlated with reading ability (Swanson et al., 2003). There is indication that RAN is at least partially dissociable from phonological ability. Indeed, RAN and phonological ability account for independent variance in reading achievement (Bowers & Swanson, 1991; Wolf et al., 2002). RAN scores significantly correlates with reading ability even when phonological awareness ability has been removed (Wolf et al., 2002). RAN is dependent on a wide range of cognitive skills as both speed and accuracy can be influenced by multiple different cognitive processes (Arnell et al., 2009). Certainly attentional processes strongly influence RAN performance, and indeed, children with attention deficit/hyperactivity disorder (ADHD) show reduced performance on RAN measures (Waber, Wolff, Forbes, & Weiler, 2000). RAN tasks require a participant to sustain focused attention over time and exhibit inhibition/suppression of previous and upcoming responses while the current response is planned or executed (Wolf & Bowers, 1999). These cognitive processes support fluent reading and have corresponding patterns of neural activation, which we discuss in detail below.
Decades of behavioral research have contributed a rich literature and an understanding of how aspects of linguistic knowledge (i.e. phonology, semantics) and developing cognitive abilities (i.e. rapid naming) contribute to successful reading acquisition. These components of the reading system are also candidates for reading disorders. Theories of reading disorders present deficits in single components (e.g. phonology) or combinations of these components (e.g. phonology and naming speed or working memory) as the underlying causes of reading disorders. Indeed, there is a strong body of scientific work that implicates various linguistic and cognitive skills in reading disorder.
However, behavioral results alone cannot fully reveal the extent to which typical reading acquisition is supported by each factor (phonological, semantic, cognitive, etc.,), the discrete versus overlapping contribution of each factor, and the static versus dynamic significance of each factor throughout the child’s development. The use of functional neuroimaging technologies for developmental research of reading can adjudicate between these remaining considerations. For example, functional magnetic resonance imaging (fMRI) can reveal what brain regions support different aspects of reading, changes across development, and how these brain regions differ as a function of cross-linguistic variation, the child’s experience (i.e. monolingual versus bilingual), or in atypical or delayed reading.
4 Reading and the Brain
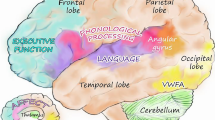
Skilled reading involves mapping orthographic information onto phonological and semantic representations, the lion’s share of which is performed by a reading network that can broadly be divided into three sub-systems: the ventral (occipitotemporal), dorsal (temporoparietal) and an anterior area centered around the left inferior frontal gyrus (LIFG). Visual word input is relayed to the left fusiform gyrus, occipito-temporal region also referred to as the “visual word form area”; VWFA, Brodmann’s area (BA) 37 (McCandliss, Cohen, & Dehaene, 2003; Pugh, Mencl, Jenner, et al., 2000; Schlaggar & McCandliss, 2007). This region functions as a pre-semantic area and appears to be linguistically-tuned and largely dedicated to processing orthographic information (Sandak, Mencl, Frost, & Pugh, 2004). This functional specificity of the VWFA appears to be late developing and directly related to acquisition of literacy (Dehaene et al., 2010; Shaywitz et al., 2002). Developing reading ability is accompanied by a decreased response to faces in the left hemisphere’s VWFA (Dehaene et al., 2010); rather the VWFA begins to respond to orthographic stimuli (Baker et al., 2007; Cohen et al., 2000; Shaywitz et al., 2002). Dehaene et al. (2010) contrasted neural response to spoken and written language, visual faces, and non-face, non-orthographic visual stimuli among adults with various levels of literacy. Neural responses to non-orthographic stimuli in the VWFA decreased with reading performance. This comparison of literate and illiterate adults revealed a significant decrease in neural response to face stimuli, but not to non-face/non-orthographic stimuli in the VWFA among literate versus illiterate adults. Reading acquisition may, to some degree, influence the lateralization of orthographic processing to the left hemisphere (VWFA) and face processing to homologous regions in the right hemisphere (FFA). The implication here is that experience of learning to read changes the brain and reveals to us the plasticity of the brain’s reading circuitry.
Visual input to the VWFA extends into the ventral system including the middle and inferior temporal gyri (MTG, ITG) where word information appears to be semantically processed (Fiebach, Friederici, Muller, & von Cramon, 2002; Price, 2012). The dorsal temporoparietal system includes the inferior parietal lobule (IPL), with the angular gyrus, which is involved in lexical-semantic processing (Seghier, Fagan, & Price, 2010), and the supramarginal gyrus (BA 39 and 40), and has an important role in converting orthography into phonology (Bookheimer, Zeffiro, Blaxton, Gaillard, & Theodore, 1995; Moore & Price, 1999). The dorsal system also includes “classic” language architecture such as the superior temporal gyrus (STG, BA 21/22/42), which is known to be important in phonological processing (e.g., (Petitto et al., 2000; Zatorre & Belin, 2001).
The anterior reading system includes the left inferior frontal gyrus (LIFG), which includes pars triangularis, pars opercularis (Broca’s area, BA 44/45) and pars orbitalis (BA 47), which is involved in syntax, morphology, semantics and phonology, including the search and retrieval of information about the meanings of words (Bookheimer, 2002; Caplan, 2001; Price, 2000, 2010, 2012; Sabb, Bilder, Chou, & Bookheimer, 2007). The more posterior portions of the LIFG show greater activation during tasks that involve sublexical phonological coding, phonological memory, and syntactic processing (Pugh, Mencl, Jenner et al., 2000). However, the more anterior portions of the LIFG show greater activation during tasks that involve semantic retrieval (Poldrack et al., 1999).
Regions within these systems, such as the supramarginal gyrus, LIFG, STG and VWFA, tend to activate for both words and pseudowords, but show greater activation for pseudowords versus familiar words (Bookheimer et al., 1995; Graves, Desai, Humphries, Seidenberg, & Binder, 2010; Heim et al., 2005; Shaywitz et al., 2002). When reading words, the left inferior, frontopolar, and dorsolateral prefrontal cortex (DLPFC) are recruited during the retrieval of semantic content of words. These areas show greater activation as compared with nonsense words, which show significantly less activation (Goldberg, Perfetti, Fiez, & Schneider, 2007). Words that have irregular or inconsistent mappings between spelling and sounds also show greater neural activation as compared with regularly-spelled words during oral naming (Fiebach et al., 2002; Fiez & Petersen, 1998). More, a word’s semantic characteristics influence the phonological processing involved in word naming (Frost et al., 2005). Although naming is primarily driven by phonological assembly from the orthographic input, there is a trade-off between phonology and semantics, such that when phonological processing is slower and less accurate due to inconsistent spelling-to-sound mapping, semantics has a stronger impact on naming performance (Strain, Patterson, & Seidenberg, 1995, 2002). Neuroimaging studies have also demonstrated this tradeoff on activation with greater activation of the inferior frontal gyrus or the middle temporal and angular gyri, depending on the respective impact of phonology or semantics (Frost et al., 2005).
As children become skilled readers, several models suggest a relatively greater reliance on orthographic-to-semantic coding indicating that although phonological knowledge is important for reading, semantic knowledge becomes increasing important for later reading skill (Berends & Reitsma, 2006; Hoover & Gough, 1990; Snowling, 2004). For example, training in semantics for reading is only beneficial at later stages of reading, but not at early stages of reading (Berends & Reitsma, 2006). This relative shift from phonological to semantic weighting over the course of reading development has been argued to be associated with a shift in the recruitment of brain regions classically associated with aspects of language function (Jasińska & Petitto, 2013b; Turkeltaub, Gareau, Flowers, Zeffiro, & Eden, 2003). Jasińska and Petitto (2014) examined the development of neural systems for reading using functional Near Infrared Spectroscopy (fNIRS) in younger (ages 6–8) and older (ages 8–10) children using different word conditions that exploit differences in reading strategy. Children read regularly-spelled words with a one-to-one correspondence between letters and sounds (e.g., hill) and irregularly spelled words without a direct one-to-one correspondence (e.g., circus). Younger readers showed similar neural recruitment of the left STG for both irregular and regular words, however, older readers showed robust neural recruitment of the LIFG for irregular relative to regular words. The STG is known to be important in phonological processing, (e.g., Petitto et al., 2000; Zatorre & Belin, 2001) whereas the LIFG typically participates in syntax, morphology, semantics and phonology, including the search and retrieval of information about the meanings of words and is sensitive to the spelling-to-sound regularity/consistency) and frequency of words (Caplan, 2001; Fiez, Balota, Raichle, & Petersen, 1999; Foundas, Eure, Luevano, & Weinberger, 1998; Kovelman, Baker, & Petitto, 2008b; Price, 2010). Younger readers show greater neural activation in phonological processing tissue at an age when children rely heavily on phonology for reading (Jasińska & Petitto, 2014). The study therefore shows a focal brain correlate of a developmental reading milestone. Older readers also show activation in the STG that is consistent with their reliance on phonology during reading. However older readers also show greater neural activation in tissue classically associated with lexical access at an age when children begin to utilize larger grain sizes in processing words, including whole word units (Jasińska & Petitto, 2014). Moreover, this neural activation is greater for irregularly-spelled words, which have a greater reliance on the orthographic-to-semantic component versus the orthographic-to-phonological-to-semantic component.
Although much attention has been devoted to mapping the language and reading circuitry of the human brain, multiple additional regions of the brain support reading. Aspects of cognition such as working memory and attention are critical components of reading and are supported by regions in the frontal lobe such as the dorsolateral prefrontal cortex (DLPFC, BA 9/46) (Balconi, 2013; Petrides, 2005). Extensive neural maturation of the frontal lobes takes place between the ages of 3 and 6 years (Courchesne et al., 2000; Huttenlocher & Dabholkar, 1997; Lenneberg, 1967). Children begin formal reading instruction in school precisely at this age. Frontal cortex maturation, including the DLPFC, is particularly protracted as compared to other cortical regions (Cone, Burman, Bitan, Bolger, & Booth, 2008; Shaw et al., 2008). The left temporoparietal areas (STG, IPL), which represent key regions in the reading circuitry, mature earlier in comparison with frontal regions such as the LIFG, or the DLPFC (Cone et al., 2008; Shaw et al., 2008). The maturation of the frontal cortex and the cognitive skills that are supported by this brain region, in addition to other brain areas supporting language and reading, contribute to reading development (Monzalvo & Dehaene-Lambertz, 2013).
5 Reading Disability in the Brain
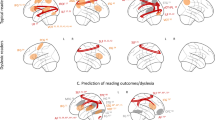
Behavioral studies have revealed differences in phonological awareness and RAN between children who are typical readers and children who show reading disabilities and/or delays (RD). There are corresponding functional differences between typical and atypical readers. A number of functional neuroimaging studies demonstrate disruptions to the left hemisphere posterior reading circuitry including both dorsal and ventral systems. There is a relative under-activation of these neural systems while children with reading impairments are processing print stimuli, including both words and pseudowords (Brunswick, McCrory, Price, Frith, & Frith, 1999; Paulesu et al., 2001; Pugh, Mencl, Jenner et al., 2000, Pugh et al., 2001, 2013; Salmelin, Service, Kiesila, Uutela, & Salonen, 1996; Shaywitz et al., 1996, 1998, 2002). A key distinction between typical readers and children with reading impairment is not limited to differential activation of brain regions that comprise the reading circuit. Rather, the connections between the different sub-components of the reading circuitry vary in typical and impaired reading. The entire reading system must function cooperatively to give rise to accurate and fluent reading. For example, children who show reading impairments show weaker correlations between the angular gyrus and other sites involved in reading as compared with typical readers (Horwitz, Rumsey, & Donohue, 1998; Pugh, Mencl, Shaywitz et al., 2000). This diminished connectivity appears to be related with diminished ability to assemble phonological units from orthographic form or perhaps to integrate this information with semantic knowledge.
As a compensatory mechanism to the hypo-activation of the left hemisphere reading circuitry, children with reading impairments show greater activation of the right hemisphere while reading. The disruptions to the left hemisphere circuitry and corresponding additional right hemisphere activation has been observed across languages that vary in orthographic depth and writing systems, including Finnish, German, French, Italian and Chinese (Paulesu et al., 2001; Salmelin et al., 1996; Shaywitz et al., 2002; Siok, Perfetti, Jin, & Tan, 2004). Such cross-linguistic research indicates that reading impairments arise in global phonological processes underlying orthography-to-phonology mapping common to all languages. When this capacity breaks down in reading impairment, there is often additional recruitment of right hemisphere brain regions that support compensatory processing particularly when task demands are high. There is evidence that children with reading impairment show greater engagement of prefrontal dorsolateral sites as compared with typical readers (Brunswick et al., 1999; Salmelin et al., 1996). Children with reading impairments may be showing an overreliance on covert pronunciation (articulatory recoding) in recruiting frontal regions to compensate for poor phonological decoding (Frost et al., 2008). Additional right hemisphere activation may reflect increased visual processing of the orthographic form in lieu of adequate phonological processing, or more generally, may reflect increased effort during reading that requires additional brain regions for support.
Compared with typical readers, children with reading disabilities show reduced grey matter volume in several brain regions including left occipitotemporal and temporoparietal areas, the fusiform gyrus as well as the cerebellum (Hoeft et al., 2007; Kronbichler et al., 2008; Raschle, Chang, & Gaab, 2011). Grey matter volume in these brain regions is positively related with pre-reading and reading skills, including phonological processing and rapid automatized naming (Kronbichler et al., 2008; Pernet, Andersson, Paulesu, & Demonet, 2009). White matter differences are also observed between typical readers and children with reading disabilities (Klingberg et al., 2000; Silani et al., 2005; Steinbrink et al., 2008). White matter organization appears weaker (via diffusion tensor imaging; DTI) in LIFG, left temporoparietal and posterior reading areas (Klingberg et al., 2000; Rimrodt, Peterson, Denckla, Kaufmann, & Cutting, 2010). Atypical functional activation for reading among children with reading disabilities may be associated with structural brain differences (Clark et al., 2014; Hoeft et al., 2007).
The functional and structural brain characteristics of children with reading disabilities in comparison to children with typical reading skills are well characterized, with an increasing number of studies converging on the neural basis of skilled and impaired reading. More recently we have begun to examine neurochemistry in the typical versus atypical reading brain (Pugh et al., 2014). We sampled individual differences in neurochemistry in children early in reading development (age 7) using magnetic resonance spectroscopy (MRS). Concentrations of choline and glutamate neurometabolites measured at a midline occipital region the brain were negatively correlated with reading skills and measures of phonology and vocabulary. The structural white matter anomalies that are widely observed in children with reading disabilities may be related to choline and glutamate levels in the brain. Moreover, elevated glutamate levels in the brain may also be related to hyperexcitability and consequently affect the coherence of neural networks that support learning to read. The role of neurometabolites in the formation of a stable reading circuitry is just beginning to be investigated, but points to new research directions in reading disability, and the possibility of neuropharmacological treatments. Thus a growing body of evidence reveals structural, functional, and neurochemical signatures for RD. With clear indications that RD is heritable (Castles, Bates, Coltheart, Luciano, & Martin, 2006; Dennis et al., 2009; Fisher & Francks, 2006; Grigorenko, 2001, 2007; Landi et al., 2013), work across these levels of analysis will yield better neurophenotypes and better causal accounts going forward.
Although one objective has been to establish the brain bases of skilled and impaired reading, the major question that still remains is, of course, one concerning the neural plasticity of these brain bases. Are the neural differences between skilled and impaired readers modifiable with good evidence-based treatments? Are these circuits fundamentally disrupted or just poorly organized? Treatments for reading disorders (once a diagnosis has been confirmed) typically take place in schools or special education intervention programs and can result in reading gain depending on the type, intensity and length of treatment (Gabrieli, 2009). If the intervention is more intensive (daily instruction and practice for weeks or months, for example), occurs in smaller groups (e.g. one-to-one teacher-student ratio), incorporates phonological decoding and awareness training, and is begun earlier in younger children, then gains in reading fluency may be seen by ~50 % of students for years after returning to the regular curriculum (Gabrieli, 2009). Treatment for reading disorders result in functional changes in the brain’s activation patterns, resulting in increased activation in classic left hemisphere temporoparietal reading networks and some reductions in frontal areas that typically show hyperactivation in reading disorders (Aylward et al., 2003; Eden et al., 2004; Gabrieli, 2009; Temple et al., 2003).
6 Reading Across Languages
The functional neural anatomy underlying reading has been studied across a variety of languages, both alphabetic and non-alphabetic (Chee, O’Craven, Bergida, Rosen, & Savoy, 1999; Fiebach et al., 2002; Jasińska & Petitto, 2014; Kuo et al., 2004; Paulesu et al., 2000; Salmelin et al., 1996). Neuroimaging studies have revealed activation in left hemisphere brain regions (occipitotemporal, temporoparietal, and inferior frontal networks) while reading in English (e.g., Pugh, Sandak, Frost, Moore, & Mencl, 2005), which are also activated while reading in other languages. For example, brain regions that are involved in phonological processing in English are also active in Chinese in tasks such as homophone judgment (Kuo et al., 2004). Thus, it appears at a global level, varied language and writing system recruit the same brain regions for reading. However, language differences in the orthography-phonology relationship that is crucial at the onset of literacy, as well as other aspects of language structure (for example, morphologically-rich, highly inflected languages with lax word order such as Russian versus analytical languages with stricter word order such as English) have been shown to produce some differences in the degree and extent of activation, with limited differences in location. For example, Kovelman, Baker, and Petitto (2008a) observed difference in neural activation of the LIFG for English sentences varying in syntactic complexity, but not for Spanish sentences. The precise manipulation that resulted in different neural activation in the English contrast, but not the Spanish contrast was depended on differences in the relevance of word order versus morphological inflection between the two languages: English favoring word order, and Spanish favoring inflectional morphology (Kovelman et al. 2008a). Irrespective of these language-specific differences, any comprehensive models of reading should incorporate the cognitive and linguistic processes common to all languages as a set of reading universals (Frost, 2012).
The conception of reading universals posits that disorders in reading should be similarly realized across languages. Differences in the contribution of phonological or semantic processing to reading disorders may exist across languages that differ with respect to these aspects of linguistic knowledge. However, an overall consistent pattern of deficits in phonology, semantics, RAN and other cognitive capacities emerges in cross-linguistic comparisons of reading disorders, with finer features of the reading disorder exhibiting sensitivities to language specifics (Pugh, 2006). Evidence for universals in reading disorders comes from studies of languages that differ in their orthographic-to-phonological mapping (transparent versus opaque orthographies), and therefore are theoretical candidates for revealing reading disorders that vary with orthographic depth. Ziegler, Perry, Ma-Wyatt, Ladner, and Schulte-Korne (2003) compared reading disorder in German (regular orthographic-to-phonological mapping) and English (irregular orthographic-to-phonological mapping), yet their results clearly showed the similarities between children with reading disorders were far greater than their differences across these two languages (Ziegler et al., 2003). Reading disorders across languages manifest as a failures to develop the functional specialization of the VWFA (Paulesu et al., 2001; Shaywitz et al., 2002). Beyond behavioral and neural similarities in reading disorders across languages, the heritability of reading disorder is strong evidence that there is an underlying biological contribution to reading disorder, which is not (and ought not) to be sensitive to cross-linguistic variation (Grigorenko, 2001).
7 Reading in the Bilingual Brain
How do a bilingual child’s two languages impact the brain’s neural circuitry for reading? Though much more is now known about the process by which the monolingual brain reads (Frost et al., 2009; McCandliss et al., 2003), the process by which the bilingual brain reads in each of their two languages remains comparatively less understood (Berens, Kovelman, & Petitto, 2013; Jasińska & Petitto, 2013b, 2014; Kovelman et al. 2008a, 2008b; Lafrance & Gottardo, 2005; Lesaux & Siegel, 2003; Lipka & Siegel, 2007). The bilingual brain must be able to process orthographic, phonological and semantic information across two languages while reading. Moreover, the relationship between orthography, phonology and semantics in each of a bilingual’s two languages can vary, further complicating reading in two languages.
Early life bilingual exposure is associated with a bilingual phonological processing advantage (Bialystok, Majumder, & Martin, 2003; Kovelman et al. 2008a; Petitto & Holowka, 2002; Petitto et al., 2012). Bilingual infants demonstrate greater and longer neural sensitivity to universal phonetic distinctions when monolingual infants can no longer make such discriminations. Early bilingual exposure may provide a linguistic “Perceptual Wedge” that extends infants’ sensitivity to universal phonetic contrast and may later aid language and reading development in childhood (Jasińska & Petitto, 2011; Petitto et al., 2012). Indeed, bilingual school-aged children outperform their monolingual peers on measures of phonological awareness (Bialystok et al., 2003; Bruck & Genesee, 1995; Eviatar & Ibrahim, 2000; Jasińska & Petitto, 2014; Kovelman et al. 2008a; Petitto et al., 2001, 2012; Rubin & Turner, 1989). Children educated in bilingual English-Spanish schools from monolingual English-speaking homes outperform children educated in monolingual English schools on a complex phonological awareness task requiring children to break apart a word into individual phonemes (Kovelman et al. 2008a). Bilingual children exposed to two languages from birth show better performance on measures of phonological awareness (phoneme deletion and phoneme segmentation tasks) and word reading tasks when compared with their monolingual peers (Jasińska & Petitto, 2013a). Moreover, English monolingual children who had been exposed to their new language for the first time when they entered a French language program in school also showed higher performance on measures of phonological awareness and word reading in English relative to their monolingual peers who were attending monolingual English schools (Jasińska & Petitto, 2013a, Revise and Resubmit) . Collectively, these studies provide compelling evidence that bilinguals can have a linguistic – specifically phonological, and reading advantage. As an aside, there are still some inconsistencies in the literature specifically concerning differences in vocabulary size between monolingual and bilingual children. Differences emerge between monolinguals’ vocabulary size when compared with bilinguals’ vocabulary size in only one of their two languages, which can be misleadingly interpreted as a bilingual disadvantage. Yet, similarities are found when monolinguals’ vocabulary size is compared with bilinguals’ vocabulary size in both of their two languages (Petitto & Holowka, 2002; Poulin-Dubois, Bialystok, Blaye, Polonia, & Yott, 2013).
A growing body of research shows that bilingual experience in early life has the potential to yield changes in the patterns of neural activation in these language and reading related neural regions. Using fMRI, differential recruitment of language-related brain regions such as the LIFG has been previously observed for bilingual adults as compared with monolingual adults while reading sentences (Kovelman et al. 2008b). Bilingual adults were found to recruit a greater extent and variability of the LIFG while completing a syntactic sentence judgment task relative to monolinguals. The greater recruitment of neural resources supporting language processing in the bilingual brain showed that early exposure to two languages modifies the neural activation of networks that underlie language processing, and reveals the brain’s “neural signature” of bilingualism (Kovelman et al. 2008b). This “neural signature” is also found in the developing brain, indicating that early bilingual exposure can modify language-related brain regions before they have fully matured. Using fNIRS neuroimaging, both bilingual children and adults were found to show greater neural activation in classic language brain areas, including the bilateral superior temporal gyrus and the inferior frontal gyrus during a sentence reading task (Jasińska & Petitto, 2013b) and a word reading task (Jasińska & Petitto, 2014). More recently, we have observed greater functional connectivity between left and right hemisphere language areas in bilingual children as compared to monolingual children while reading (Jasinska & Petitto, submitted). Bilingualism may provide enrichment for the developing brain realized as greater connectivity. That bilingual readers show a greater extent and variability of neural activation in classic language (LIFG, STG, IPL) areas as well as their right hemisphere homologues and higher cognitive (DLPFC, RLPFC) brain areas and increased functional connectivity suggests that bilingualism may lead to enhanced linguistic and cognitive processing and reveals the extent of neural architecture underlying language and reading that can be modified through early life language experience.
Research on the neural basis of healthy and typical bilingual language and reading development continues to amass evidence of a bilingual “neural signature”, that is, a greater extent and variability in the patterns of neural activation that support language and reading in the bilingual brain (Jasińska & Petitto, 2013b, 2014; Kovelman et al. 2008b, Kovelman, Shalinsky, Berens, & Petitto, 2008; Petitto et al., 2012). Bilingual exposure also has the potential to yield structural changes in the brain (Mechelli et al., 2004). These structural and functional neural changes occur as a function of early life bilingual experience; moreover, such neural changes are at least partially modulated by the age of bilingual exposure (Jasińska & Petitto, 2013b).
Yet, we have virtually no knowledge of whether bilingual exposure affects reading disorders, and if so, how? Would reading disorders present in one or both of a bilingual’s two languages? If the reading disorder does present in both languages, would language-specific differences emerge? These questions remain unanswered; yet pursing this line of research has the potential to reveal new information about reading disorders in a bilingual population. Indeed, the majority of the world speaks multiple languages, and thus the focus on reading disorders in predominantly English-speaking monolinguals is under representative of the full scope of reading disorders.
One exciting hypothesis for reading disorders emerges from research on bilingual language and reading processing. Bilinguals show distinctive neural activation changes in phonological processing, which are apparent even in the young bilingual infant (Petitto et al., 2012; Werker, 2012). This pattern of neural activation in the young bilingual infant may underlie the phonological advantages for reading in school-aged bilingual children (Berens et al., 2013; Bialystok et al., 2003; Jasińska & Petitto, Revise and Resubmit; Kang, 2010; Kovelman et al. 2008a; Kuo & Anderson, 2012; Laurent & Martinot, 2009a, 2009b). Given the widely observed phonological processing deficits in reading disorders, it begs us to consider if bilingual advantages in phonological processing can ameliorate some of these deficits in reading disorders.
The hypothesis that early life bilingual exposure may provide an advantage for reading disorders through enhanced phonological processing is not without merit. Phonologically-based interventions for reading disorder show promising results: children show gains in reading fluency after undergoing a year-long treatment where training and practice in mapping letters to sounds was the focus (Shaywitz et al., 2004). A child who grows up bilingual has two phonological systems, and has greater experience with phonology. The possibility that reading disorders may present less frequently in children who grow up bilingual from birth because of growing up bilingual needs to be formally investigated.
8 Future Directions
Research from our laboratory as well as others converge on a similar set of findings regarding the neural circuitry that gives rise to skilled, fluent reading. Skilled reading involves mapping orthographic information onto phonological and semantic representations, which is performed by left-hemisphere frontal, temporoparietal and occipitotemporal cortical regions. The growing set of scientific findings implicates these brain regions, specifically the supramarginal gyrus, the posterior interior frontal gyrus, the superior temporal gyrus, and the VWFA, in aspects of phonological processing. This suggests that these brain regions are phonologically “tuned” and support this key aspect of reading. This is particularly relevant given that a child’s phonological awareness at times in development when literacy is emerging is predictive of later literacy outcomes. By contrast, the angular gyrus, the inferior and middle temporal gyri, the anterior inferior frontal gyrus and the dorsolateral prefrontal cortex have more of a role in abstract lexico-semantic function.
However, there are variations in this system in instances where a child’s reading skill is delayed and/or disordered. Reading impairment in the brain is marked by consistent neural observations including hypo-activation of the left hemisphere posterior reading circuitry, decreased connectivity between sub-systems of the reading circuit, and additional (compensatory) recruitment of the prefrontal cortex and right hemisphere regions. Specifically, disruptions to the reading circuitry that are characteristic of delayed/disordered reading often correspond to behavioral deficits in phonological processing and decoding. Treatments focusing on phonology have resulted in improvements to reading fluency, and are accompanied by increased activation in left hemisphere posterior reading areas (Eden et al., 2004; Gabrieli, 2009; Shaywitz et al., 2004). More, new directions in neuropharmacological treatment for reading disorder are now being investigated given recent exciting findings from Haskins Laboratories indicating neurometabolites such as glutamate and choline are involved in the development of the brain’s reading circuitry (Pugh et al., 2014).
The majority of research on the reading brain has predominantly focused on a monolingual English-speaking population, however, there are also variations in the brain’s reading circuitry that arise as a consequence of early-life dual-language experience and yield reading advantages. The young, healthy and typical bilingual reader shows greater neural recruitment of brain regions classically associated with language function and reading as well as corresponding regions in the right hemisphere. Early exposure to two languages modifies the neural activation of networks that underlie language, and in turn, reading.
Establishing the neural circuitry of reading in the (predominately) English monolingual brain has yielded highly valuable information that has allowed us to track healthy development and compare this against delayed and/or disordered development. Crucially, this information serves as the foundation to developing targeted intervention for reading. Yet, the majority of the world’s children are growing up bilingual. Therefore, the task ahead is to refine the typical neuro-maturational timetable for the bilingual neural circuit and understand how it differs from bilingual reading impairments. We must also consider the possibility that early bilingualism may ameliorate, or even partially prevent, reading disorders. We offer a novel and bold hypothesis that the rich early-life phonological experience that a bilingual child has as compared to her monolingual peer may promote healthy and typical reading development.
References
Ackerman, P. T., & Dykman, R. A. (1993). Phonological processes, confrontational naming, and immediate memory in dyslexia. Journal of Learning Disabilities, 26, 597–609. doi:10.1177/002221949302600910.
Anderson, R. C., & Freebody, P. (1983). Reading comprehension and the assessment and acquisition of word knowledge. Advances in Reading/Language Research, 2, 231–256.
Arnell, K. M., Joanisse, M. F., Klein, R. M., Busseri, M. A., & Tannock, R. (2009). Decomposing the relation between rapid automatized naming (ran) and reading ability. Canadian Journal of Experimental Psychology/Revue Canadienne de Psychologie Expérimentale, 63(3), 173–184.
Aylward, E. H., Richards, T. L., Berninger, V. W., Nagy, W. E., Field, K. M., Grimme, A. C., … Cramer, S. C. (2003). Instructional treatment associated with changes in brain activation in children with dyslexia. Neurology, 61(2), 212–219.
Baker, C. I., Liu, J., Wald, L. L., Kwong, K. K., Benner, T., & Kanwisher, N. (2007). Visual word processing and experiential origins of functional selectivity in human extrastriate cortex. PNAS Proceedings of the National Academy of Sciences of the United States of America, 104(21), 9087–9092. http://dx.doi.org.myaccess.library.utoronto.ca/10.1073/pnas.0703300104.
Balconi, M. (2013). Dorsolateral prefrontal cortex, working memory and episodic memory processes: Insight through transcranial magnetic stimulation techniques. Neuroscience Bulletin, 29(3), 381–389. doi:10.1007/s12264-013-1309-z.
Berends, I. E., & Reitsma, P. (2006). Addressing semantics promotes the development of reading fluency. Applied Psycholinguistics, 27, 247–265.
Berens, M. S., Kovelman, I., & Petitto, L. A. (2013). Should bilingual children learn reading in two languages at the same time or in sequence? Bilingual Research Journal, 36(1), 35–60. doi:10.1080/15235882.2013.779618.
Bialystok, E., Majumder, S., & Martin, M. M. (2003). Developing phonological awareness: Is there a bilingual advantage? Applied Psycholinguistics, 24, 27–44. doi:10.1017/S014271640300002X.
Bookheimer, S. (2002). Functional MRI of language: New approaches to understanding the cortical organization of semantic processing. Annual Review of Neuroscience, 25, 151–188. doi:10.1146/annurev.neuro.25.112701.142946.
Bookheimer, S. Y., Zeffiro, T. A., Blaxton, T., Gaillard, W., & Theodore, W. (1995). Regional cerebral blood flow during object naming and word reading. Human Brain Mapping, 3(2), 93–106. doi:10.1002/hbm.460030206.
Bowers, P. G., & Swanson, L. B. (1991). Naming speed deficits in reading-disability – multiple measures of a singular process. Journal of Experimental Child Psychology, 51, 195–219. doi:10.1016/0022-0965(91)90032-n.
Bowey, J. A., Cain, M. T., & Ryan, S. M. (1992). A reading-level design study of phonological skills underlying fourth-grade children’s word reading difficulties. Child Development, 63(4), 999–1011. doi:10.2307/1131249.
Bradley, L., & Bryant, P. E. (1983). Categorizing sounds and learning to read: A causal connection. Nature, 301(5899), 419–421. doi:10.1038/301419a0.
Bruck, M., & Genesee, F. (1995). Phonological awareness in young second language learners. Journal of Child Language, 22(2), 307–324.
Brunswick, N., McCrory, E., Price, C. J., Frith, C. D., & Frith, U. (1999). Explicit and implicit processing of words and pseudowords by adult developmental dyslexics: A search for Wernicke’s Wortschatz? Brain, 122(Pt 10), 1901–1917.
Caplan, D. (2001). Functional neuroimaging studies of syntactic processing. Journal of Psycholinguistic Research, 30, 297–320. doi:10.1023/A:1010495018484.
Caravolas, M., Lervag, A., Defior, S., Seidlova Malkova, G., & Hulme, C. (2013). Different patterns, but equivalent predictors, of growth in reading in consistent and inconsistent orthographies. Psychological Science, 24(8), 1398–1407. doi:10.1177/0956797612473122.
Castles, A., Bates, T., Coltheart, M., Luciano, M., & Martin, N. G. (2006). Cognitive modelling and the behaviour genetics of reading. Journal of Research in Reading, 29, 92–103. doi:10.1111/j.1467-9817.2006.00294.x.
Chee, M. W., O’Craven, K. M., Bergida, R., Rosen, B. R., & Savoy, R. L. (1999). Auditory and visual word processing studied with fMRI. Human Brain Mapping, 7(1), 15–28.
Clark, K. A., Helland, T., Specht, K., Narr, K. L., Manis, F. R., Toga, A. W., et al. (2014). Neuroanatomical precursors of dyslexia identified from pre-reading through to age 11. Brain. doi:10.1093/brain/awu229.
Cohen, L., Dehaene, S., Naccache, L., Lehericy, S., Dehaene-Lambertz, G., Henaff, M. A., et al. (2000). The visual word form area: Spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain, 123(2), 291–307.
Cone, N. E., Burman, D. D., Bitan, T., Bolger, D. J., & Booth, J. R. (2008). Developmental changes in brain regions involved in phonological and orthographic processing during spoken language processing. NeuroImage, 41(2), 623–635. doi:10.1016/j.neuroimage.2008.02.055.
Cossu, G., Shankweiler, D., Liberman, I. Y., Katz, L., & Tola, G. (1988). Awareness of phonological segments and reading-ability in Italian children. Applied Psycholinguistics, 9(1), 1–16. doi:10.1017/S0142716400000424.
Courchesne, E., Chisum, H. J., Townsend, J., Cowles, A., Covington, J., Egaas, B., … Press, G. A. (2000). Normal brain development and aging: Quantitative analysis at in vivo mr imaging in healthy volunteers. Radiology, 216(3), 672–682.
Dehaene, S., Pegado, F., Braga, L. W., Ventura, P., NunesFilho, G., Jobert, A., … Cohen, L. (2010). How learning to read changes the cortical networks for vision and language. Science, 330(6009), 1359–1364.
Denckla, M. B., & Rudel, R. (1974). Rapid “automatized” naming of pictured objects, colors, letters and numbers by normal children. Cortex, 10(2), 186–202.
Dennis, M. Y., Paracchini, S., Scerri, T. S., Prokunina-Olsson, L., Knight, J. C., Wade-Martins, R., … Stubbs, L. (2009). A common variant associated with dyslexia reduces expression of the kiaa0319 gene. PLoS Genetics, 5(3), 1. doi:10.1371/journal.pgen.1000436.
Eden, G. F., Jones, K. M., Cappell, K., Gareau, L., Wood, F. B., Zeffiro, T. A., … Flowers, D. L. (2004). Neural changes following remediation in adult developmental dyslexia. Neuron, 44(3), 411–422. doi:10.1016/j.neuron.2004.10.019.
Eviatar, Z., & Ibrahim, R. (2000). Bilingual is as bilingual does: Metalinguistic abilities of arabic-speaking children. Applied Psycholinguistics, 21(4), 451–471.
Fiebach, C. J., Friederici, A. D., Muller, K., & von Cramon, D. Y. (2002). fMRI evidence for dual routes to the mental lexicon in visual word recognition. Journal of Cognitive Neuroscience, 14(1), 11–23. doi:10.1162/089892902317205285.
Fiez, J. A., Balota, D. A., Raichle, M. E., & Petersen, S. E. (1999). Effects of lexicality, frequency, and spelling-to-sound consistency on the functional anatomy of reading. Neuron, 24(1), 205–218.
Fiez, J. A., & Petersen, S. E. (1998). Neuroimaging studies of word reading. Proceedings of the National Academy of Sciences of the United States of America, 95(3), 914–921.
Fisher, S. E., & Francks, C. (2006). Genes, cognition and dyslexia: Learning to read the genome. Trends in Cognitive Science, 10(6), 250–257. doi:10.1016/j.tics.2006.04.003.
Foundas, A. L. A., Eure, K. F. K., Luevano, L. F. L., & Weinberger, D. R. D. (1998). MRI asymmetries of broca’s area: The pars triangularis and pars opercularis. Brain and Language, 64(3), 282–296.
Foy, J. G., & Mann, V. (2006). Changes in letter sound knowledge are associated with development of phonological awareness in pre-school children. Journal of Research in Reading, 29(2), 143–161. doi:10.1111/j.1467-9817.2006.00279.x.
Frost, R. (1998). Toward a strong phonological theory of visual word recognition: True issues and false trails. Psychological Bulletin, 123(1), 71–99.
Frost, R. (2012). Towards a universal model of reading. Behavioral Brain Science, 35(5), 263–279. doi:10.1017/s0140525x11001841.
Frost, R., Katz, L., & Bentin, S. (1987). Strategies for visual word recognition and orthographical depth: A multilingual comparison. Journal of Experimental Psychology: Human Perception and Performance, 13(1), 104–115.
Frost, S. J., Mencl, W. E., Sandak, R., Moore, D. L., Rueckl, J. G., Katz, L., … Pugh, K. R. (2005). A functional magnetic resonance imaging study of the tradeoff between semantics and phonology in reading aloud. Neuroreport, 16(6), 621–624. doi:10.1097/00001756-200504250-00021.
Frost, S. J., Sandak, R., Mencl, W. E., Landi, N., Moore, D., Porta, G. D., … Pugh, K. R. (2008). Neurobiological and behavioral studies of skilled and impaired word reading. In Single-word reading: Behavioral and biological perspectives (pp. 355–376). Mahwah, NJ: Lawrence Erlbaum Associates Publishers.
Frost, S. J., Sandak, R., Mencl, W. E., Landi, N., Rueckl, J. G., Katz, L., et al. (2009). Mapping the word reading circuitry in skilled and disabled readers. In How children learn to read: Current issues and new directions in the integration of cognition, neurobiology and genetics of reading and dyslexia research and practice (pp. 3–19). New York, NY: Psychology Press.
Gabrieli, J. D. (2009). Dyslexia: A new synergy between education and cognitive neuroscience. Science, 325(5938), 280–283. doi:10.1126/science.1171999.
Georgiou, G. K., Torppa, M., Manolitsis, G., Lyytinen, H., & Parrila, R. (2012). Longitudinal predictors of reading and spelling across languages varying in orthographic consistency. Reading and Writing, 25(2), 321–346. doi:10.1007/s11145-010-9271-x.
Goldberg, R. F., Perfetti, C. A., Fiez, J. A., & Schneider, W. (2007). Selective retrieval of abstract semantic knowledge in left prefrontal cortex. Journal of Neuroscience, 27(14), 3790–3798. doi:10.1523/JNEUROSCI.2381-06.2007.
Goswami, U. (2003). Why theories about developmental dyslexia require developmental designs. Trends in Cognitive Science, 7(12), 534–540.
Goswami, U. (2008). Reading, dyslexia and the brain. Educational Research, 50(2), 135–148. doi:10.1080/00131880802082625.
Goswami, U., & Bryant, P. (1990). Phonological skills and learning to read. Hillsdale, NJ: Lawrence Erlbaum Associates.
Goswami, U., Gerson, D., & Astruc, L. (2010). Amplitude envelope perception, phonology and prosodic sensitivity in children with developmental dyslexia. Reading and Writing, 23(8), 995–1019. doi:10.1007/s11145-009-9186-6.
Grainger, J., & Holcomb, P. J. (2009). An ERP investigation of orthographic priming with relative-position and absolute-position primes. Brain Research, 1270, 45–53. doi:10.1016/j.brainres.2009.02.080.
Graves, W. W., Desai, R., Humphries, C., Seidenberg, M. S., & Binder, J. R. (2010). Neural systems for reading aloud: A multiparametric approach. Cerebral Cortex, 20(8), 1799–1815. doi:10.1093/cercor/bhp245.
Grigorenko, E. L. (2001). Developmental dyslexia: An update on genes, brains, and environments. Journal of Child Psychology and Psychiatry, 42(1), 91–125. http://dx.doi.org.myaccess.library.utoronto.ca/10.1111/1469-7610.00704.
Grigorenko, E. L. (2007). Triangulating developmental dyslexia: Behavior, brain, and genes. New York: Guilford Press.
Harm, M. W., & Seidenberg, M. S. (1999). Phonology, reading acquisition, and dyslexia: Insights from connectionist models. Psychological Review, 106(3), 491–528.
Harm, M. W., & Seidenberg, M. S. (2004). Computing the meanings of words in reading: Cooperative division of labor between visual and phonological processes. Psychological Review, 111(3), 662–720.
Hatcher, P. J., & Hulme, C. (1999). Phonemes, rhymes, and intelligence as predictors of children’s responsiveness to remedial reading instruction: Evidence from a longitudinal intervention study. Journal of Experimental Child Psychology, 72(2), 130–153.
Heim, S., Alter, K., Ischebeck, A. K., Amunts, K., Eickhoff, S. B., Mohlberg, H., … Friederici, A. D. (2005). Research report: The role of the left brodmann's areas 44 and 45 in reading words and pseudowords. Cognitive Brain Research, 25(3), 982–993. doi:10.1016/j.cogbrainres.2005.09.022.
Hoeft, F., Meyler, A., Hernandez, A., Juel, C., Taylor-Hill, H., Martindale, J. L., … Gabrieli, J. D. (2007). Functional and morphometric brain dissociation between dyslexia and reading ability. Proceedings National Academy Science USA, 104(10), 4234–4239. doi:10.1073/pnas.0609399104.
Hoien, T., Lundberg, I., Stanovich, K. E., & Bjaalid, I.-K. (1995). Components of phonological awareness. Reading and Writing, 7(2), 171–188.
Hoover, W. A., & Gough, P. B. (1990). The simple view of reading. Reading and Writing, 2(2), 127–160. doi:10.1007/Bf00401799.
Horwitz, B., Rumsey, J. M., & Donohue, B. C. (1998). Functional connectivity of the angular gyrus in normal reading and dyslexia. Proceedings of the National Academy of Sciences of the United States of America, 95(15), 8939–8944.
Hulme, C., Caravolas, M., Málkova, G., & Brigstocke, S. (2005). Phoneme isolation ability is not simply a consequence of letter-sound knowledge. Cognition, 97(1), B1–B11. doi:10.1016/j.cognition.2005.01.002.
Hulme, C., Hatcher, P. J., Nation, K., Brown, A., Adams, J., & Stuart, G. (2002). Phoneme awareness is a better predictor of early reading skill than onset-rime awareness. Journal of Experimental Child Psychology, 82(1), 2–28. doi:10.1006/jecp.2002.2670.
Huttenlocher, P. R., & Dabholkar, A. S. (1997). Regional differences in synaptogenesis in human cerebral cortex. Journal of Comparative Neurology, 387(2), 167–178.
Jasińska, K. K., & Petitto, L. A. (Revise and Resubmit). Age of bilingual exposure predicts distinct contributions of phonology and semantics to successful reading development.
Jasińska, K., & Petitto, L. A. (2011). Dual language exposure in infancy can change neural and language processing in the developing brain: An fNIRS investigation. Paper presented at the Society for Neuroscience, Washington, DC.
Jasińska, K., & Petitto, L. A. (2013a). Age of bilingual exposure predicts distinct contributions of phonological and semantic knowledge to successful reading development. Paper presented at the Society for Research in Child Development Seattle, WA.
Jasińska, K. K., & Petitto, L. A. (2013b). How age of bilingual exposure can change the neural systems for language in the developing brain: A functional near infrared spectroscopy investigation of syntactic processing in monolingual and bilingual children. Developmental Cognitive Neuroscience, 6c, 87–101. doi:10.1016/j.dcn.2013.06.005
Jasińska, K. K., & Petitto, L. A. (2014). Development of neural systems for reading in the monolingual and bilingual brain: New insights from functional near infrared spectroscopy neuroimaging. Developmental Neuropsychology, 39(6), 421–439. doi:10.1080/87565641.2014.939180.
Kang, J. Y. (2010). Do bilingual children possess better phonological awareness? Investigation of Korean monolingual and Korean-english bilingual children. Reading and Writing, 25(2), 411–431. doi:10.1007/s11145-010-9277-4.
Katz, L., & Feldman, L. B. (1983). Relation between pronunciation and recognition of printed words in deep and shallow orthographies. Journal of Experimental Psychology: Learning, Memory, and Cognition, 9(1), 157–166. doi:10.1037/0278-7393.9.1.157.
Katz, L., & Frost, R. (1992). The reading process is different for different orthographies: The orthographic depth hypothesis. In R. Frost & L. Katz (Eds.), Orthography, phonology, morphology, and meaning (pp. 67–84). Oxford, UK: North-Holland.
Klingberg, T., Hedehus, M., Temple, E., Salz, T., Gabrieli, J. D., Moseley, M. E., et al. (2000). Microstructure of temporo-parietal white matter as a basis for reading ability: Evidence from diffusion tensor magnetic resonance imaging. Neuron, 25(2), 493–500.
Kovelman, I., Baker, S., & Petitto, L. A. (2008a). Age of first bilingual language exposure as a new window into bilingual reading development. Bilingualism: Language and Cognition, 11(2), 203–223. doi:10.1017/S1366728908003386.
Kovelman, I., Baker, S., & Petitto, L. A. (2008b). Bilingual and monolingual brains compared: A functional magnetic resonance imaging investigation of syntactic processing and a possible “neural signature” of bilingualism. Journal of Cognitive Neuroscience, 20(1), 153–169.
Kovelman, I., Shalinsky, M. H., Berens, M. S., & Petitto, L. A. (2008). Shining new light on the brain’s “bilingual signature”: A functional near infrared spectroscopy investigation of semantic processing. Neuroimage, 39(3), 1457–1471. doi:10.1016/j.neuroimage.2007.10.017
Kronbichler, M., Wimmer, H., Staffen, W., Hutzler, F., Mair, A., & Ladurner, G. (2008). Developmental dyslexia: Gray matter abnormalities in the occipitotemporal cortex. Human Brain Mapping, 29(5), 613–625. doi:10.1002/hbm.20425.
Kuo, L.-J., & Anderson, R. C. (2012). Effects of early bilingualism on learning phonological regularities in a new language. Journal of Experimental Child Psychology, 111(3), 455–467. doi:10.1016/j.jecp.2011.08.013.
Kuo, W. J., Yeh, T. C., Lee, J. R., Chen, L. F., Lee, P. L., Chen, S. S., … Hsieh, J. C. (2004). Orthographic and phonological processing of Chinese characters: An fMRI study. Neuroimage, 21(4), 1721–1731. doi:10.1016/j.neuroimage.2003.12.007.
Lafrance, A., & Gottardo, A. (2005). A longitudinal study of phonological processing skills and reading in bilingual children. Applied Psycholinguistics, 26(4), 559–578. doi:10.1017/S0142716405050307.
Landi, N., Frost, S. J., Mencl, W. E., Preston, J. L., Jacobsen, L. K., Lee, M., … Grigorenko, E. L. (2013). The comt val/met polymorphism is associated with reading-related skills and consistent patterns of functional neural activation. Developmental Science, 16(1), 13–23. doi:10.1111/j.1467-7687.2012.01180.x.
Landi, N., Mencl, W. E., Frost, S. J., Sandak, R., & Pugh, K. R. (2010). An fMRI study of multimodal semantic and phonological processing in reading disabled adolescents. Annals of Dyslexia, 60(1), 102–121. doi:10.1007/s11881-009-0029-6.
Laurent, A., & Martinot, C. (2009a). Bilingualism and phonological awareness: The case of bilingual (French–occitan) children. Reading and Writing, 23(3–4), 435–452. doi:10.1007/s11145-009-9209-3.
Laurent, A., & Martinot, C. (2009b). Bilingualism and phonological segmentation of speech: The case of english-French pre-schoolers. Journal of Early Childhood Literacy, 9(1), 29–49. doi:10.1177/1468798408101102.
Lavidor, M., Johnston, R., & Snowling, M. J. (2006). When phonology fails: Orthographic neighbourhood effects in dyslexia. Brain and Language, 96(3), 318–329.
Lenneberg, E. H. (1967). Biological foundations of language.
Lesaux, N. K., & Siegel, L. S. (2003). The development of reading in children who speak english as a second language. Developmental Psychology, 39(6), 1005–1019. doi:10.1037/0012-1649.39.6.1005.
Liberman, I. Y., Shankweiler, D., & Liberman, A. M. (1989). The alphabetic principle and learning to read. In D. Shankweiler & I. Y. Liberman (Eds.), Phonology and reading disability: Solving the reading puzzle. Ann Arbor, MI: University of Michigan Press.
Lipka, O., & Siegel, L. S. (2007). The development of reading skills in children with english as a second language. Scientific Studies of Reading, 11(2), 105–131. doi:10.1080/10888430709336555.
Lo, M., Hue, C.-W., & Tsai, F.-Z. (2007). Chinese readers’ knowledge of how Chinese orthography represents phonology. Chinese Journal of Psychology, 49(4), 315–334.
Lukatela, G., & Turvey, M. T. (1994). Visual lexical access is initially phonological.1. Evidence from associative priming by words, homophones, and pseudohomophones. Journal of Experimental Psychology General, 123(2), 107–128. doi:10.1037/0096-3445.123.2.107.
McCandliss, B. D., Cohen, L., & Dehaene, S. (2003). The visual word form area: Expertise for reading in the fusiform gyrus. Trends in Cognitive Science, 7(7), 293–299. doi:10.1016/S1364-6613(03)00134-7.
McCardle, P., Scarborough, H. S., & Catts, H. W. (2001). Predicting, explaining, and preventing children’s reading difficulties. Learning Disabilities Research and Practice, 16(4), 230–239. doi:10.1111/0938-8982.00023.
Mechelli, A., Crinion, J. T., Noppeney, U., O’Doherty, J., Ashburner, J., Frackowiak, R. S., et al. (2004). Neurolinguistics: Structural plasticity in the bilingual brain. Nature, 431(7010), 757. doi:10.1038/431757a.
Melby-Lervåg, M., Lyster, S.-A. H., & Hulme, C. (2012). Phonological skills and their role in learning to read: A meta-analytic review. Psychological Bulletin, 138(2), 322–352. doi:10.1037/a0026744.
Monzalvo, K., & Dehaene-Lambertz, G. (2013). How reading acquisition changes children’s spoken language network. Brain and Language, 127(3), 356–365. doi:10.1016/j.bandl.2013.10.009.
Moore, C. J., & Price, C. J. (1999). Three distinct ventral occipitotemporal regions for reading and object naming. NeuroImage, 10(2), 181–192.
Muter, V., Hulme, C., Snowling, M. J., & Stevenson, J. (2004). Phonemes, rimes, vocabulary, and grammatical skills as foundations of early reading development: Evidence from a longitudinal study. Developmental Psychology, 40(5), 665–681. doi:10.1037/0012-1649.40.5.665.
Muter, V. V., Hulme, C. C., Snowling, M. M., & Taylor, S. S. (1997). Segmentation, not rhyming, predicts early progress in learning to read. Journal of Experimental Child Psychology, 65(3), 370–396.
Nation, K., & Hulme, C. (1997). Phonemic segmentation, not onset-rime segmentation, predicts early reading and spelling skills. Reading Research Quarterly, 32(2), 154–167.
Nation, K., & Snowling, M. J. (2004). Beyond phonological skills: Broader language skills contribute to the development of reading. Journal of Research in Reading, 27(4), 342–356. doi:10.1111/j.1467-9817.2004.00238.x.
Oullette, G. P. (2006). What’s meaning got to do with it: The role of vocabulary in word reading and reading comprehension. Journal of Educational Psychology, 98(3), 554–566.
Panah, M. A., & Padakannaya, P. (2008). Reading acquisition in Persian: Transition from phonological recoding strategy to orthographic strategy. Psychological Studies, 53(2), 122–125.
Paulesu, E., Demonet, J. F., Fazio, F., McCrory, E., Chanoine, V., Brunswick, N., … Frith, U. (2001). Dyslexia: Cultural diversity and biological unity. Science, 291(5511), 2165–2167. doi:10.1126/science.1057179.
Paulesu, E., McCrory, E., Fazio, F., Menoncello, L., Brunswick, N., Cappa, S. F., … Frith, U. (2000). A cultural effect on brain function. Nauret Neuroscience, 3(1), 91–96. doi:10.1038/71163.
Perfetti, C. (2007). Reading ability: Lexical quality to comprehension. Scientific Studies of Reading, 11(4), 357–383.
Perfetti, C. A., & Bell, L. (1991). Phonemic activation during the 1st 40 ms of word identification – evidence from backward-masking and priming. Journal of Memory and Language, 30(4), 473–485. doi:10.1016/0749-596x(91)90017-e.
Perfetti, C., & Hart, L. (2002). The lexical quality hypothesis. In C. Verhoeven, C. Elbro, & P. Reitsma (Eds.), Precursors of functional literacy (pp. 189–213). Amsterdam, The Netherlands/Philadelphia, PA: John Benjamins.
Pernet, C., Andersson, J., Paulesu, E., & Demonet, J. F. (2009). When all hypotheses are right: A multifocal account of dyslexia. Human Brain Mapping, 30(7), 2278–2292. doi:10.1002/hbm.20670.
Petitto, L.-A., Berens, M. S., Kovelman, I., Dubins, M. H., Jasińska, K., & Shalinsky, M. (2012). The “perceptual wedge hypothesis” as the basis for bilingual babies’ phonetic processing advantage: New insights from fNIRS brain imaging. Brain and Language, 121(2), 130–143. doi:10.1016/j.bandl.2011.05.003.
Petitto, L. A., & Holowka, S. (2002). Evaluating attributions of delay and confusion in young bilinguals: Special insights from infants acquiring a signed and a spoken language. Sign Language Studies, 3(1), 4–33.
Petitto, L. A., Katerelos, M., Levy, B. G., Gauna, K., Tetreault, K., & Ferraro, V. (2001). Bilingual signed and spoken language acquisition from birth: Implications for the mechanisms underlying early bilingual language acquisition. Journal of Child Language, 28(2), 453–496.
Petitto, L.-A., Zatorre, R. J., Gauna, K., Nikelski, E. J., Dostie, D., & Evans, A. C. (2000). Speech-like cerebral activity in profoundly deaf people processing signed languages: Implications for the neural basis of human language. Proceedings of the National Academy of Sciences, 97(25), 13961–13966.
Petrides, M. (2005). Lateral prefrontal cortex: Architectonic and functional organization. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 360(1456), 781–795. doi:10.1098/rstb.2005.1631.
Poldrack, R. A., Wagner, A. D., Prull, M. W., Desmond, J. E., Glover, G. H., & Gabrieli, J. D. E. (1999). Functional specialization for semantic and phonological processing in the left inferior prefrontal cortex. NeuroImage, 10(1), 15–35. doi:10.1006/nimg.1999.0441.
Poulin-Dubois, D., Bialystok, E., Blaye, A., Polonia, A., & Yott, J. (2013). Lexical access and vocabulary development in very young bilinguals. International Journal of Billingualism, 17(1), 57–70. doi:10.1177/1367006911431198.
Preston, J. L., Frost, S. J., Mencl, W. E., Fulbright, R. K., Landi, N., Grigorenko, E., … Pugh, K. R. (2010). Early and late talkers: School-age language, literacy and neurolinguistic differences. Brain, 133(8), 2185–2195. doi:10.1093/brain/awq163.
Price, C. J. (2000). The anatomy of language: Contributions from functional neuroimaging. Journal of Anatomy, 197(Pt 3), 335–359.
Price, C. J. (2010). The anatomy of language: A review of 100 fMRI studies published in 2009. Annals of the New York Academy of Sciences, 1191, 62–88. doi:10.1111/j.1749-6632.2010.05444.x.
Price, C. J. (2012). A review and synthesis of the first 20 years of pet and fMRI studies of heard speech, spoken language and reading. NeuroImage, 62(2), 816–847. doi:10.1016/j.neuroimage.2012.04.062.
Proctor, C. P., August, D., Carlo, M. S., & Snow, C. (2006). The intriguing role of Spanish language vocabulary knowledge in predicting english reading comprehension. Journal of Educational Psychology, 98(1), 159–169. doi:10.1037/0022-0663.98.1.159.
Pugh, K. (2006). A neurocognitive overview of reading acquisition and dyslexia across languages. Developmental Science, 9(5), 448–450. doi:10.1111/j.1467-7687.2006.00528.x. discussion 451–443.
Pugh, K. R., Frost, S. J., Rothman, D. L., Hoeft, F., Del Tufo, S. N., Mason, G. F., … Fulbright, R. K. (2014). Glutamate and choline levels predict individual differences in reading ability in emergent readers. Journal of Neuroscience, 34(11), 4082–4089. doi:10.1523/jneurosci.3907-13.2014.
Pugh, K. R., Landi, N., Preston, J. L., Mencl, W. E., Austin, A. C., Sibley, D., … Frost, S. J. (2013). The relationship between phonological and auditory processing and brain organization in beginning readers. Brain and Language, 125(2), 173–183. doi:10.1016/j.bandl.2012.04.004.
Pugh, K. R., Mencl, W. E., Jenner, A. R., Katz, L., Frost, S. J., Lee, J. R., … Shaywitz, B. A. (2000). Functional neuroimaging studies of reading and reading disability (developmental dyslexia). Mental Retardation and Developmental Disabilities Research Reviews, 6(3), 207–213.
Pugh, K. R., Mencl, W. E., Jenner, A. R., Lee, J. R., Katz, L., Frost, S. J., … Shaywitz, B. A. (2001). Neuroimaging studies of reading development and reading disability. Learning Disabilities Research & Practice, 16(4), 240–249. doi:10.1111/0938-8982.00024.
Pugh, K. R., Mencl, W. E., Shaywitz, B. A., Shaywitz, S. E., Fulbright, R. K., Constable, R. T., … Gore, J. C. (2000). The angular gyrus in developmental dyslexia: Task-specific differences in functional connectivity within posterior cortex. Psychological Science, 11(1), 51–56. doi:10.1111/1467-9280.00214.
Pugh, K. R., Sandak, R., Frost, S. J., Moore, D., & Mencl, W. E. (2005). Examining reading development and reading disability in english language learners: Potential contributions from functional neuroimaging. Learning Disabilities Research and Practice, 20(1), 24–30.
Raschle, N. M., Chang, M., & Gaab, N. (2011). Structural brain alterations associated with dyslexia predate reading onset. NeuroImage, 57(3), 742–749. doi:10.1016/j.neuroimage.2010.09.055.
Rimrodt, S. L., Peterson, D. J., Denckla, M. B., Kaufmann, W. E., & Cutting, L. E. (2010). White matter microstructural differences linked to left perisylvian language network in children with dyslexia. Cortex, 46(6), 739–749. doi:10.1016/j.cortex.2009.07.008.
Rolla San Francisco, A., Mo, E., Carlo, M., August, D., & Snow, C. (2006). The influences of language of literacy instruction and vocabulary on the spelling of Spanish-english bilinguals. Reading and Writing, 19(6), 627–642. doi:10.1007/s11145-006-9012-3.
Roth, F. P., Speece, D. L., & Cooper, D. H. (2002). A longitudinal analysis of the connection between oral language and early reading. The Journal of Educational Research, 95(5), 259–272.
Rubin, H., & Turner, A. (1989). Linguistic awareness skills in grade one children in a French immersion setting. Reading and Writing, 1(1), 73–86.
Sabb, F. W., Bilder, R. M., Chou, M., & Bookheimer, S. Y. (2007). Working memory effects on semantic processing: Priming differences in pars orbitalis. NeuroImage, 37(1), 311–322. doi:10.1016/j.neuroimage.2007.04.050.
Salmelin, R., Service, E., Kiesila, P., Uutela, K., & Salonen, O. (1996). Impaired visual word processing in dyslexia revealed with magnetoencephalography. Annals of Neurology, 40(2), 157–162. doi:10.1002/ana.410400206.
Sandak, R., Mencl, W. E., Frost, S. J., & Pugh, K. R. (2004). The neurobiological basis of skilled and impaired reading: Recent findings and new directions. Scientific Studies of Reading, 8(3), 273–292. doi:10.1207/s1532799xssr0803_6.
Schlaggar, B. L., & McCandliss, B. D. (2007). Development of neural systems for reading. Annual Review of Neuroscience, 30, 475–503. doi:10.1146/annurev.neuro.28.061604.135645.
Seghier, M. L., Fagan, E., & Price, C. J. (2010). Functional subdivisions in the left angular gyrus where the semantic system meets and diverges from the default network. Journal of Neuroscience, 30(50), 16809–16817. doi:10.1523/jneurosci.3377-10.2010.
Seidenberg, M. S., Bruck, M., Fornarolo, G., & Backman, J. (1985). Word recognition processes of poor and disabled readers: Do they necessarily differ? Applied Psycholinguistics, 6(2), 161–179. doi:10.1017/S0142716400006093.
Share, D. L. (1995). Phonological recoding and self-teaching: Sine qua non of reading acquisition. Cognition, 55(2), 151–218. discussion 219–126.
Share, D. L., & Leikin, M. (2004). Language impairment at school entry and later reading disability: Connections at lexical versus supralexical levels of reading. Scientific Studies of Reading, 8(1), 87–110.
Shaw, P., Kabani, N. J., Lerch, J. P., Eckstrand, K., Lenroot, R., Gogtay, N., … Wise, S. P. (2008). Neurodevelopmental trajectories of the human cerebral cortex. Journal of Neuroscience, 28(14), 3586–3594. doi:10.1523/jneurosci.5309-07.2008.
Shaywitz, B. A., Shaywitz, S. E., Blachman, B. A., Pugh, K. R., Fulbright, R. K., Skudlarski, P., … Gore, J. C. (2004). Development of left occipitotemporal systems for skilled reading in children after a phonologically- based intervention. Biology Psychiatry, 55(9), 926–933. doi:10.1016/j.biopsych.2003.12.019.
Shaywitz, S. E., Shaywitz, B. A., Pugh, K. R., Fulbright, R. K., Constable, R. T., Mencl, W. E., … Gore, J. C. (1998). Functional disruption in the organization of the brain for reading in dyslexia. Proceedings National Academy Science USA, 95(5), 2636–2641.
Shaywitz, B. A., Shaywitz, S. E., Pugh, K. R., Mencl, W. E., Fulbright, R. K., Skudlarksi, P., … Gore, J. C. (2002). Disruption of posterior brain systems for reading in children with developmental dyslexia. Biology Psychiatry, 52(2), 101–110. doi:10.1016/S0006-3223(02)01365-3.
Shaywitz, B. A., Shaywitz, S. E., Pugh, K. R., Skudlarski, P., Fulbright, R. K., Constable, R., … Gore, J. C. (1996). The functional organization of brain for reading and reading disability (dyslexia. Neuroscientist, 2(4), 245–255. doi:10.1177/107385849600200413.
Silani, G., Frith, U., Demonet, J. F., Fazio, F., Perani, D., Price, C., … Paulesu, E. (2005). Brain abnormalities underlying altered activation in dyslexia: A voxel based morphometry study. Brain, 128(Pt 10), 2453–2461. doi:10.1093/brain/awh579.
Siok, W. T., Perfetti, C. A., Jin, Z., & Tan, L. H. (2004). Biological abnormality of impaired reading is constrained by culture. Nature, 431(7004), 71–76. doi:10.1038/nature02865.
Snow, C. E., Tabors, P. O., Nicholson, P. A., & Kurland, B. F. (1995). Shell: Oral language and early literacy skills in kindergarten and first-grade children. Journal of Research in Childhood Education, 10(1), 37–48. doi:10.1080/02568549509594686.
Snowling, M. J. (2004). Language skills and learning to read. Psychologist, 17, 438–441.
Steinbrink, C., Vogt, K., Kastrup, A., Muller, H. P., Juengling, F. D., Kassubek, J., et al. (2008). The contribution of white and gray matter differences to developmental dyslexia: Insights from DTI and VBM at 3.0 t. Neuropsychologia, 46(13), 3170–3178. doi:10.1016/j.neuropsychologia.2008.07.015.
Strain, E., Patterson, K., & Seidenberg, M. S. (1995). Semantic effects in single-word naming. Journal of Experimental Psychology Learning Memory Cognition, 21(5), 1140–1154.
Strain, E., Patterson, K., & Seidenberg, M. S. (2002). Theories of word naming interact with spelling-sound consistency. Journal of Experimental Psychology Learning Memory Cognition, 28(1), 207–214. discussion 215–220.
Swanson, H. L., Trainin, G., Necoechea, D. M., & Hammill, D. D. (2003). Rapid naming, phonological awareness, and reading: A meta-analysis of the correlation evidence. Review of Educational Research, 73(4), 407–440. doi:10.3102/00346543073004407.
Temple, E., Deutsch, G. K., Poldrack, R. A., Miller, S. L., Tallal, P., Merzenich, M. M., et al. (2003). Neural deficits in children with dyslexia ameliorated by behavioral remediation: Evidence from functional MRI. Proceedings of the National Academy of Sciences, 100(5), 2860–2865. doi:10.1073/pnas.0030098100.
Torgesen, J. K., Wagner, R. K., & Rashotte, C. A. (1994). Longitudinal studies of phonological processing and reading. Journal of Learning Disabilities, 27(5), 276–286. discussion 287–291.
Turkeltaub, P. E., Gareau, L., Flowers, D. L., Zeffiro, T. A., & Eden, G. F. (2003). Development of neural mechanisms for reading. Nature Neuroscience, 6(7), 767–773. doi:10.1038/nn1065.
Waber, D., Wolff, P. H., Forbes, P. W., & Weiler, M. D. (2000). Rapid automatized naming in children referred for evaluation of heterogeneous learning problems: How specific are naming speed deficits to reading disability? Child Neuropsychology, 6(4), 251–261.
Wagner, R. K., & Torgesen, J. K. (1987). The nature of phonological processing and its causal role in the acquisition of reading skills. Psychological Bulletin, 101(2), 192–212. doi:10.1037/0033-2909.101.2.192.
Wagner, R. K., Torgesen, J. K., Rashotte, C. A., Hecht, S. A., Barker, T. A., Burgess, S. R., … Garon, T. (1997). Changing relations between phonological processing abilities and word-level reading as children develop from beginning to skilled readers: A 5-year longitudinal study. Developmental Psychology, 33(3), 468–479. doi:10.1037/0012-1649.33.3.468.
Wang, M., Yang, C., & Cheng, C. (2009). The contributions of phonology, orthography, and morphology in Chinese-english biliteracy acquisition. Applied Psycholinguistics, 30(2), 291–314. doi:10.1017/S0142716409090122.
Weber-Fox, C. M., & Neville, H. J. (1999). Functional neural subsystems are differentially affected by delays in second language immersion: Erp and behavioral evidence in bilinguals. In Second language acquisition and the critical period hypothesis (pp. 23–38). Mahwah, NJ: Lawrence Erlbaum Associates Publishers.
Weber-Fox, C., & Neville, H. J. (2001). Sensitive periods differentiate processing of open- and closed-class words: An ERP study of bilinguals. Journal of Speech, Language, and Hearing Research, 44(6), 1338–1353. doi:10.1044/1092-4388(2001/104).
Werker, J. (2012). Perceptual foundations of bilingual acquisition in infancy. Annals of the New York Academy of Sciences, 1251, 50–61. doi:10.1111/j.1749-6632.2012.06484.x.
Wolf, M., & Bowers, P. G. (1999). The double-deficit hypothesis for the developmental dyslexias. Journal of Educational Psychology, 91(3), 415–438. doi:10.1037/0022-0663.91.3.415.
Wolf, M., O’Rourke, A. G., Gidney, C., Lovett, M., Cirino, P., & Morris, R. (2002). The second deficit: An investigation of the independence of phonological and naming-speed deficits in developmental dyslexia. Reading and Writing, 15(1–2), 43–72.
Zatorre, R. J., & Belin, P. (2001). Spectral and temporal processing in human auditory cortex. Cerebral Cortex, 11(10), 946–953.
Ziegler, J. C., & Goswami, U. (2005). Reading acquisition, developmental dyslexia, and skilled reading across languages: A psycholinguistic grain size theory. Psychological Bulletin, 131(1), 3–29. doi:10.1037/0033-2909.131.1.3.
Ziegler, J. C., Perry, C., Ma-Wyatt, A., Ladner, D., & Schulte-Korne, G. (2003). Developmental dyslexia in different languages: Language-specific or universal? Journal of Experimental Child Psychology, 86(3), 169–193.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Additional information
Note
Portions of this chapter have appeared in:
1. Jasińska, K. & Petitto, L.A. (2014). Development of Neural Systems for Reading in the Monolingual and Bilingual Brain: New Insights from functional Near Infrared Spectroscopy Neuroimaging. Developmental Neuropsychology. 39(6), 421–39. doi: 10.1080/87565641.2014.939180
2. Jasińska, K. & Petitto, L.A. (Revise and Resubmit). Age of Bilingual Exposure Predicts Distinct Contributions of Phonology and Semantics to Successful Reading Development. Manuscript submitted for publication.
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Jasińska, K.K. et al. (2016). Neuroimaging Perspectives on Skilled and Impaired Reading and the Bilingual Experience. In: Khateb, A., Bar-Kochva, I. (eds) Reading Fluency. Literacy Studies, vol 12. Springer, Cham. https://doi.org/10.1007/978-3-319-30478-6_3
Download citation
DOI: https://doi.org/10.1007/978-3-319-30478-6_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-30476-2
Online ISBN: 978-3-319-30478-6
eBook Packages: EducationEducation (R0)