
Abstract
The bushmeat crisis is often portrayed as a serious threat to primates, especially slow-reproducing ape species. And yet, most bushmeat markets in Central and West Africa are dominated by ungulates, rodents, and carnivores. While markets are a measure of hunters’ off-take, they may not adequately capture information on species consumed in households or traded locally. Comparing changes in prey availability at markets, households, and other trading points can assist efforts to assess the magnitude of faunal extraction. Overall, subsistence needs of local human populations combined with the transnational demand for bushmeat results in the amplification of hunter catchments. What remains unclear is how the availability of nonhuman primates is reconciled with cultural preferences and market values to determine what appears in cooking pots. This study examines this question using market (n = 157 days) and hunter preference surveys (n = 290), in addition to semi-structured interviews with market women (n = 10) and hunters (n = 210) in the rural town of Bayanga, Central African Republic (2008–2009) located within the Dzanga-Sangha Protected Areas (APDS). Integrating multiple approaches, we were able to reconcile and contextualize discrepancies between market data, hunter surveys, and interviews with market women and community members. We found that the sale and consumption of primate meat at APDS was more likely related to a decline in preferred ungulate species along with an increase in the availability of firearms. These results suggest nuanced avenues for increasing the protection of vulnerable and endangered primate species with the introduction of well-priced, alternative protein sources.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
Most human populations in the Congo Basin rely on wildlife for protein (Bennett et al. 2007; Bowen-Jones and Pendry 1999; Eves and Bakaar 2001; Fa et al. 2002a, 2002b). Such use and extraction of wildlife has become popularly referred to as the bushmeat trade, which is defined as the commercial hunting and sale of wildlife (BCTF 2000). It is a trade that is often characterized as a “crisis” because of its illegal and unsustainable characteristics. Throughout the Congo Basin, burgeoning economies of wildlife, increased human migration, changing technology, and the presence of conservation and development projects threaten the viability of wildlife populations and the human communities dependent upon them (Barnes 2002; Fa and Brown 2009; Linder and Oates 2011; Jost Robinson et al. 2011; Milner-Gulland and Bennett 2003). Milner-Gulland and Bennett (2003) note that bushmeat is consumed on a “massive scale” and that this high level of consumption is directly linked to both global and local economies. At the local level, consumption is based on the intersections of local ecology, subsistence practices, economics, and preferences. Preferences for bushmeat are culturally based and shaped by various factors including tradition, taste preferences, education level, and migration status (Daspit 2011; East et al. 2005; Fa et al. 2003; Schenck et al. 2006).
The continued and increased consumption of bushmeat has been found to be related to both wildlife abundance and preferences for wild meat alongside the high costs of protein alternatives and the low availability and productivity of domestic livestock in tropical forests (Fa and Brown 2009; Wilkie and Godoy 2001; Wilkie et al. 2005). More recently, research has demonstrated the fundamental importance of wild game to human nutrition and health in poor communities throughout the tropics (Blaney et al. 2009; Golden et al. 2011; Fa et al. 2015b). Studies have also explored ways to substitute local diets with other forms of protein; however, this may harm rather than help local inhabitants who depend on the bushmeat trade as a source of income where few alternatives exist (Milner-Gulland and Bennett 2003). The cultural significance of food, specifically bushmeat, is particularly important for human communities in the Congo Basin (de Garine 1997; de Garine and Pagezy 1990; de Merode et al. 2004; Motte-Florac et al. 1993). The importance of wild meat in the diets of local human populations results in greater challenges for conservation and development programs that overlook the cultural significance of bushmeat (Fa et al. 2015b).
In these cases, it may be neither feasible nor appropriate to provide communities with protein alternatives (Robinson and Bennett 2000). In a large-scale analysis , Fa et al. (2015a) recently determined that in some cases, it might be sustainable for smaller, low-density human communities to subsist on faster-reproducing species of game meat. Yet, before we can adequately address the issues of sustainability (Weinbaum et al. 2013), we must develop a better understanding of the complex nature of the relationships that exist between humans and wildlife, including nonhuman primates.
Regional market surveys have been used as a rapid assessment tool to measure faunal extraction and changes in relative proportion of wildlife to inform regional management plans (Fa et al. 2000, 2015a). Increases in the relative frequency of primates for sale at markets may signal an increase in human disturbance and reduction in preferred ungulate species rather than a preference for primates . Areas that have large human populations and road networks have fewer elephants, large duikers, buffalos, and red river hogs (Blake et al. 2008; Laurance et al. 2006; Wilkie et al. 2000). Additionally, an influx of guns and hunters’ adoption of them as faster, more accurate hunting technology, increases the threats to primates, especially arboreal species (Kümpel et al. 2008; Noss 1998). Moreover, increases in the representation of fast-reproducing species in wildlife markets have been seen as indicators of overexploitation of primary prey species (Cowlishaw et al. 2005; Dupain et al. 2012). However, market surveys only capture a fraction of what is hunted, consumed, or traded outside of the formal market context. It is thus essential that we more holistically address “what is on the menu,” or in the cooking pots, of local communities from multiple methodological perspectives. This will help to deepen our understanding of the dynamic relationships between humans and prey species and the relative vulnerability of primate populations. These relationships will have ecological and economic consequences beyond declining primate diversity and food insecurity for populations in primate habitat countries. Research demonstrates that the loss of primates and other frugivorous prey species (e.g., Artiodactyls and rodents) from ecosystems has knock-on effects across trophic levels (Abernathy et al. 2013; Henschel et al. 2011; Klailova et al. 2013).
Initially, the bushmeat crisis was portrayed as a primate problem—especially that of slow-reproducing great ape species. Primatologists and biologists owe a great debt of gratitude to the remarkable photographic work of Karl Amman and other conservation researchers who were able to draw initial international attention to multifarious issues surrounding the hunting, trade, and consumption of nonhuman primates and other wild game (BCTF 2000; Bowen-Jones and Pendry 1999; Peterson 2003). In fact, regional research has suggested that in many Congo Basin forests, primates are not the most preferred source of meat (Crookes et al. 2005; Fa et al. 2000, 2002a, b, 2015a; Njiforti 1996). Regardless of preference, nonhuman primates are nonetheless hunted (Abernathy et al. 2013; Fa and Brown 2009; Jost Robinson 2012), especially wherever gun hunters are common. Game depletion and the relative percentages of ungulates and primates sold in markets are highly correlated with human population density and anthropogenic activity (Fa et al. 2015a). Given their larger body size and slow rates of reproduction, we know that many primate populations cannot tolerate even modest levels of hunting.
Studies emphasizing the unsustainable hunting of nonhuman primates and other wildlife for trade and consumption in local and international markets are not novel. However, the ways in which we study, engage with, and understand the interconnected issues of conservation, poverty, and food security require us to incorporate additional methods into our approach. Here, we provide a case study that demonstrates the utility of assessing natural resource use (e.g., hunting) and primate conservation within theoretical and methodological frameworks of ethnoprimatology and multispecies ethnography (Fuentes 2012; Malone et al. 2014; Jost Robinson and Remis 2014). To better determine the degree to which monkeys are on the menu, we address the consumptive use of primates within a larger socio-ecological setting within the Dzanga-Sangha Protected Areas (APDS), Central African Republic (CAR) . We do this by investigating the ecological patterns of forest presence, and off-take as they intersect with economic and cultural patterns of commerce and consumption of monkeys and other nonhuman primate species across three contexts: the forest, the market, and the cooking pot.
Methods
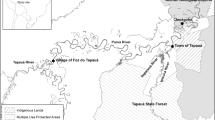
The data presented in this study were collected in the APDS, CAR located in the southwestern part of the country (2°13′26N, 16°11′26E, Fig. 1), from January 2008 to August 2009. Conservation and development activities in the southwestern CAR began in the mid-1980s (Carroll 1986) with the APDS officially designated as a park (1200 km2) and multiple-use reserve (3360 km2) in 1990. Within the APDS park sectors , only research and tourism are permitted. Cable snares are prohibited, but were commonly used through the 1980s and 1990s (see Noss 1998). In the reserve, gathering of plant and animal products, traditional spear, cross-bow and net hunts as well as regulated shotgun hunting during daylight hours are permitted.

Dzanga Sangha Protected Areas (APDS), Central African Republic (CAR)
The APDS was selectively logged at low intensity in the 1970s and again between 2002 and 2005. Human in-migration to APDS (Blom et al. 2004), greater access to arms, and a reduction in preferred ungulate prey are related to the increased hunting and trade of primates (Daspit 2011; Fa et al. 2005; Jost Robinson et al. 2011). Rainfall in the APDS averages 1400 mm per year. The forests can be broadly classified as mixed secondary forest (low canopy forest with dense understory) although mixed primary forest (high canopy forest with a relatively open understory) is also common (Remis 2000).
Line Transect Surveys
The ecological data reported here are a subset of a larger, longitudinal ecological monitoring dataset collected by Remis (1997–2005) and Jost Robinson (2008–2009). Remis and Jost Robinson have worked to maintain overlapping members of the field teams to increase accuracy and comparability of datasets. Here, we report on the 2002 (n = 100 km) and 2009 (n = 135.4 km) datasets. We focus solely on direct observations of monkey species on census walks on straight-line transects. Following patterns documented at other sites (i.e., Croes et al. 2006), our data indicate that changing antipredator tactics among cercopithecoid primates in APDS affect the accuracy of auditory detection on transects (Jost Robinson 2012; Remis and Jost Robinson 2012).
Using line transect surveys (Plumptre 2000; Plumptre and Cox 2006), we monitored large mammal species including diurnal primates (Cercopithecus pogonias, C. nictitans, C. cephus, C. neglectus, Lophocebus albigena, Cercocebus agilis, Colobus guereza, Procolobus badius, Pan troglodytes, and Gorilla gorilla gorilla), Elephants (Loxodonta africana), Suids (Potamochoerus porcus), and duikers (Philantomba monticola, Cephalophus callipygus, C. dorsalis, C. leucogaster, C. nigrifrons, C. sylvicultor) at intervals from July to August 1997 and August 2008 to June 2009.
At each of our research sites, we established four to six 5-km line transects, perpendicular to drainage patterns, using an abandoned primary logging road as a baseline. At each site, we located at least two transects within 500 m, of and parallel to, a secondary logging road, and two transects more than a kilometer from the nearest secondary road. Following methods described in White and Edwards (2000) teams of at least two trained observers walked transects between 0700 and 1300 h at a pace of 1 km/h. On the transects, we documented direct observations and calls of large mammals, noting species, age/sex (if possible), location on transect, and perpendicular distance from transect and animal-observer distance (Fashing and Cords 2000; Marshall et al. 2008). During data collection, we also recorded all indications of human activity on transects, including logging and hunting signs (indication of logging trails, stump cuts, snares, or gun shots) and other human traces.
Hunter Off-Take and Interview Data
Ethnographic semi-structured interviews (n = 210) with hunters from APDS included information on hunting methods, preference and returns, as well as their knowledge of hunted wildlife species. Given the potentially sensitive nature of hunting-related topics, we used snowball sampling to opportunistically select individuals who were willing to participate in this study (Bernard 2002; Trotter and Schensul 1998) in order to minimize the risk to human participants. Additionally, 34 weeks of surveys with 15 local hunters (cable = 5, registered firearm = 5, artisanal firearm = 5) yielded data for 793 hunting trips. Surveys included data on munitions taken, and animals captured by age (adult/juvenile) and sex class.
Market Data and Interview Methods
The primary market and ethnographic data were collected from the central town of Bayanga, in addition to four other satellite villages and forest camps. We emphasize and present the data for Bayanga only, as during our study period (2008–2009) this was the location of the single central marketplace for bushmeat and other wild and domesticated foodstuffs, including beef (sold by the Kg) and whole chickens (see Table 1 for wildlife species). Data collected in 2008 are compared to a 2006 market study using identical methodologies (Hodgkinson 2009). At other villages within APDS, the sale of wild meat occurred from homes, on roadsides, or across international borders. Additionally, ethnographic interviews (informal and semi-structured) were conducted with the ten primary bushmeat sellers in Bayanga’s marketplace in order to understand their bushmeat commerce from multiple socioeconomic perspectives.
Results
The Forest
Of the eight nonhuman primate species present in APDS , three species (C. guereza, P. badius, C. galeritus) were not detected on transects during either sampling year. Direct observations of monkeys species on transects have declined over time at APDS between 2002 (Avg = 0.73, n = 73) and 2009 (Avg = 0.26, n = 40). For those species detected (see Fig. 2), there were discernible declines in rates of observation on transects between years.

Differences in monkey observation encounter rates over time in APDS, 2002 vs. 2009
In addition to ecological transect data, we include results related to hunter off-take rates and hunting technology within the context of “forest” datasets, as these are a reflection of prey availability and the ways in which hunters interact with primates. Our datasets document a large increase in guns relative to other forms of hunting since 2006 (Daspit 2011; Jost Robinson 2012). Jost Robinson (2012) interviewed 91 self-identified gun hunters in/from Bayanga, confirming that a large percentage of guns in the region are undocumented or artisanal.
Prior to the 2000s, firearms were rare in APDS. During this earlier time, hunters relied on illegal cable snares and other traditional methods (i.e., nets and spears). Cable snares, unlike firearms, do not allow hunters to choose their prey, as catches are opportunistic. Thus, mostly ground-dwelling species, both protected and not, are trapped within snares. Moreover, the use of snares reduced the importance of arboreal primate species as a prey base for hunters. Some hunters may still rely on illegal cable snares and/or nets, given their low cost. However, firearms, both registered and artisanal (i.e., illegal) have become the prominent hunting tool in the APDS. The number of firearms has increased along with continued human in-migration to the area, as well as increased overall circulation with civil conflict in neighboring countries. During a short period of time, registered guns in the APDS increased from 8 in 2006 to 53 in 2009 (Hodgkinson 2009; Nabane pers. comm.); of course this number does not account for illegal, artisanal guns, which are also present and increasing in numbers. As such, hunters target-specific prey species, catch more individuals per hunting trip, and have increased their economic returns.
The increase in the numbers of guns in the region is likely responsible for the tenfold increase in quantities of primates hunted since 1994 (see Noss 1995). Using data on rates of prey off-take, we estimate that 90 gun hunters were able to hunt roughly 10,473 cercopithecoid primates during the 2008–2009 field season. The percentage representation of each species in hunter off-take is represented in Fig. 3. C. nictitans (blue) and L. albigena (red) were the two most common species in hunter off-take, followed by C. cephus (green) and Cercocebus galeritus (purple).

Percentage representation of monkeys in hunter catchments, 2008–2009
The Market
Between 2006 and 2008, market data from Bayanga showed a slight increase (6.62 %) in the annual estimated number of carcasses for all wildlife species (Daspit 2011). Broadly, researchers examined the proportions of species types in the marketplace, including ungulates, primates, and other species (including rodents and carnivores), in order to determine the approximate health of prey populations in the region (Cowlishaw et al. 2005; Fa et al. 2000; Noss 1998). In 2009, ungulates were the most prevalent species in the market (primarily species of blue and red duikers), representing 80 % of all carcasses available, followed by primates (19 %) and other species (1 %), including rodents, tortoises, birds, and small-bodied carnivores. There were some notable changes in the species composition across years, however, which mirror trends observed in other West and Central African markets where declines in ungulate populations from overexploitation are buffered by an increase in the hunting of nonhuman primates and rodents (Cowlishaw et al. 2005; Fa et al. 2000, 2005). In this particular market in Bayanga, there was a 9 % decrease in ungulates species available in the market place coupled with a 9 % increase in primate off-take between 2006 and 2008 (see Daspit 2011; Hodgkinson 2009). Figure 4 compares the percentage of primate species present in hunter off-take with those documented in market profiles. When we compare the proportional representation of what comes out of the forest with what appears in the market we see some interesting differences.

Comparison of percentage representation of monkeys in hunter catchment vs. market availability
C. nictitans and L. albigena were still the two most common species. However, the third most common species in the market in 2009 was C. pogonias. This species, which is taboo among many ethnic groups, was the fifth most common in hunter catchment but the third most common in the market. We saw a similar trend for the black and white colobus (orange), also a taboo species. C. pogonias and C. guereza are taboo species for some local ethnic groups as they are thought to cause bodily harm or be lethal to pregnant women, nursing mothers, and infants. Both C. pogonias and C. guereza were more common in the marketplace than expected given their representation hunter catchments.
For the case of C. pogonias, ethnographic interviews with hunters and market women suggest that this is possibly related to taboos regarding C. pogonias as well as gendered differences in commerce and economic practices. Because hunters were less often able to target their preferred prey species (i.e., duikers, Philantomba monticola and Cephalophus spp.) and many aspects of traditional food culture have eroded, all species are hunted regardless of taboos. Hunters noted that they often sold taboo species to market women and did not keep them for home consumption, thereby exercising choice in what was kept for domestic use and what was sold. We should note that C. pogonias sold for more money than that of the smaller C. cephus monkeys. However, hunters would occasionally sell C. pogonias to the local market because they did not want to consume taboo species rather than for its potential higher profit. This was a rare instance we noted where profit was not the primary motivation for species sales.
However, many market women did not appear to have the luxury to exercise choice in the same way that hunters do, as they needed to have something to sell each day. The more senior market women in Bayanga, who occupied the tables within the interior of the marketplace, would not sell monkeys but preferred species of duikers (Philantomba monticola and Cephalophus spp.). In interviews with Daspit, these market women indicated their preference for purchasing red duikers (Cephalophus spp.) followed by blue duikers (Philontomba monticola) from hunters, explaining that these are what local women preferred to purchase for their daily meals. As such, offering cuts of blue and red duikers for sale would bette r contribute to women’s market profits. Overall, information gathered from hunters, market women, and consumers further highlights the strategic aversion to the hunting and consumption of monkeys when other preferred species are available.
The Cooking Pot
The final context in which we must address the hunting and consumption of monkeys is in the home. In the APDS, hunters, market women, and consumers, whether they were born within the area or were a recent or longer term migrant to this region, all reported preferences for ungulates . Further, in everyday life, we observed that people preferred to purchase and fill their cooking pots with duiker species rather than primates. This observation was further supported by Hodgkinson’s (2009) surveys, conducted in 2006, which showed that people not only preferred but also consumed ungulate species more than primate species (Kilograms consumed/yr: Primates = 3749 (6 %), Ungulates = 57,037 (90 %)) (Fig. 5).

A young woman prepare a blue duiker (P. monticola) for a traditional meal
Discussion
So how do we reconcile these patterns where primates are hunted although not desired? We found that monkeys have both declined and become increasingly cryptic on transects over time (Remis and Jost Robinson 2012), but we also know that overall off-take increased tenfold over a 15-year period. Guns have likely helped to facilitate greater overall off-take of wildlife in this protected area as evidenced by higher carcass numbers reported in both hunter off-take and formal market surveys. Taboo species of monkeys have also become more frequently hunted, signaling the erosion of particular cultural traditions. However, when hunted, these species are likely to be sold to women who sell their foodstuffs on the outskirts of the central marketplace rather than to the market women who sell daily on tables rented within the marketplace. The more common, yet illegal, practice of night hunting (i.e., “jacklighting”) may temporarily diminish the threat of hunting to monkey species in this region, but as preferred ungulate species decline primates will take their place at the dinner table. This pattern has already been observed at West African sites (Covey and McGraw 2014).
In Bayanga , the formal market data have already begun to signal this important change from the hunting of desired prey species to secondary prey. In just 2 years during our study, small, but meaningful differences were seen in the decline in ungulates coupled with an increase in monkeys available in the marketplace. In addition, our ethnographic data suggest that what local communities were consuming was beginning to change during this time period, apparently out of necessity rather than preference. This was especially evidenced on days observed in 2008 when few to no carcasses made it to the marketplace in Bayanga, leading to a perceived scarcity of wild meat. Both market women and the women who came to the market to purchase their daily stew’s ingredients commented on the difficulty in finding meat in the market and even at individuals’ homes, a pattern told to Daspit as being a more recent phenomenon.
We suggest that a combination of ethnographic and primatological approaches helped us to better understand patterns of primate hunting and consumption (Jost Robinson and Remis 2014). In the APDS, while monkeys have not been traditionally top menu choices for many local residents, they have come to fill people’s cooking pots more frequently over time under specific and changing socio-ecological contexts. While guns were not yet exclusively associated with increased primate off-take during our study period, they allowed hunters unprecedented access to these species compared to previous time periods. We thus posit that future household and market surveys will likely point toward the increasing importance of primates to local diets, mirroring shifts in the 2008 market data as compared to 2006. The expected increase in primates will become a necessity as ungulates become less available due to the increased number of guns in the APDS , as well as, their efficiency in depleting populations of preferred prey species. It is at this point that the bushmeat trade within APDS will shift toward/become primarily a primate crisis. The hunting pressure for nonhuman primates was low during our data collection period, as local hunters preferred to target duiker species at night. Yet, as duiker populations decline, we predict that gun hunters in the APDS will revert to daytime hunting, relying more heavily on arboreal primates. Given that shifts toward primates may not result from traditions or food preferences suggests it might be possible to influence or reduce the consumption of primates if sufficiently low-priced alternatives were available (also Wilkie and Godoy 2001). Schenck and colleagues (2006) in Gabon found no strong preference for bushmeat in paired choice trials, suggesting the potential viability of domesticated alternatives.
Ethnographic understandings of wildlife off-take from multiple perspectives combined with quantitative research in forests and markets provide stronger platforms from which research can inform conservation policy, perhaps providing a silver lining for primate species. Hunters would prefer to hunt ungulates, and although we do not suggest that ungulates should be sacrificed, we do suggest they are a keystone species whose populations may be successfully maintained using active approaches to management in transitioning ecosystems and economies such as in the case of the APDS (Fa et al. 2015a).
To more effectively understand the state of nonhuman primate populations and their future in the Congo Basin, it is necessary to contextualize their numbers and threats in relation to other mammalian species that are important to local and regional diets and material economies. The successful conservation of nonhuman primates cannot be accomplished in isolation of complex, interrelated ecological, economic, and cultural practices. We must draw upon theory in anthropology, ethnoprimatology, and historical ecology to develop integrated conservation efforts that aim to provide best management practices for active management of preferred prey populations under conditions of relatively high human density. Maintenance of these populations will be key to the maintenance of nonhuman primate species in this forest. Decline of critical key prey species will have cascading effects for other wildlife species such as nonhuman primates that may be even less resilient to hunting pressures.
References
Abernathy, K. A., Coad, L., Gaylor, G., Lee, M. E., & Maisels, F. (2013). Extent and ecological consequences of hunting in Central African rainforests in the twenty-first century. Philosophical Transactions of the Royal Society B, 368, 1–12.
Barnes, R. F. W. (2002). The bushmeat boom and bust in West and Central Africa. Oryx, 36, 236–242.
BCTF. (2000). BUSHMEAT: A wildlife crisis in West and Central Africa and around the world. Washington, DC: The Bushmeat Crisis Task Force.
Bennett, E. L., Blencowe, E., Brandon, K., Brown, D., Burn, R. W., Cowlishaw, G., et al. (2007). Hunting for consensus: Reconciling bushmeat harvest, conservation, and development policy in West and Central Africa. Conservation Biology, 21, 884–887.
Bernard, H. R. (2002). Research methods in Anthropology: Qualitative and quantitative methods. Walnut Creek, CA: AltaMira Press.
Blake, S., Deem, S. L., Strindberg, S., Maisels, F., Momont, L., Isia, I., et al. (2008). Roadless wilderness area determines forest elephant movements in the Congo Basin. PLoS One, 3, e3546.
Blaney, S., Beaudry, M., & Latham, M. (2009). Contribution of natural resources to nutritional status in a protected area of Gabon. Food and Nutrition Bulletin, 30, 49–62.
Blom, A., van Zalinge, R., Mbea, E., & Heitkönig, I. M. A. (2004). Human impact on wildlife populations within a protected Central African forest. African Journal of Ecology, 42, 23–31.
Bowen-Jones, E., & Pendry, S. (1999). The threat to primates and other mammals from the bushmeat trade in Africa, and how this threat could be diminished. Oryx, 33, 233–246.
Carroll, R. W. (1986). Status of the lowland gorilla and other wildlife in the Dzanga-Sangha region of southwestern Central African Republic. Primate Conservation, 7, 38–44.
Cawthorn, D. M., & Hoffman, L. C. (2015). The bushmeat and food security nexus: A global account of contributions, conundrums and ethical collisions. Food Research International, 76, 906–925.
Covey, R., & McGraw, W. S. (2014). Monkeys in a West African bushmeat market: Implications for cercopithecid conservation in eastern Liberia. Tropical Conservation Science, 7, 115–125.
Cowlishaw, G., Mendelson, S., & Rowcliffe, M. (2005). Structure and operation of a bushmeat commodity chain in southwestern Ghana. Conservation Biology, 19, 139–149.
Crookes, D. J., Ankudey, N., & Milner-Gulland, E. J. (2005). The value of a long-term bushmeat market dataset as an indicator of system dynamics. Environmental Conservation, 32, 333–339.
Croes, B. M., Laurance, W. F., Lahm, S. A., Tchignoumba, L., Alonso, A., Lee, M. E., Campbell, P., and Buij, R. (2006). The Influence of Hunting on Antipredator Behavior in Central African Monkeys and Duikers. Biotropica, 39(2):257–263.
Daspit, L. L. (2011). Market women in a central African forest reserve: Engendering wildlife commerce and conservation. Unpublished PhD dissertation, Purdue University.
de Garine, I. (1997). Food preferences and taste in an African perspective. A word of caution. In H. Macbeth (Ed.), Food preference and taste: Continuity and change (pp. 187–207). Providence, RI: Berghahn Books.
de Garine, I., & Pagezy, H. (1990). Seasonal hunger or ‘craving for meat’? In C. M. Hladik, S. Bahuchet, & I. de Garine (Eds.), Food and nutrition in the African rain forest (pp. 43–44). Paris: UNESCO MAB.
de Merode, E., Homewood, K., & Cowlishaw, G. (2004). The value of bushmeat and other wild foods to rural households living in extreme poverty in Democratic Republic of Congo. Biological Conservation, 118, 573–581.
Dupain, J., Nackoney, J., Vargas, J. M., Johnson, P. J., Farfan, M. A., Bofaso, M., et al. (2012). Bushmeat characteristics vary with catchment condition in a Congo market. Biological Conservation, 146, 32–40.
East, T., Kümpel, N. F., Milner-Gulland, E. J., & Rowcliffe, J. M. (2005). Determinants of urban bushmeat consumption in Río Muni, Equatorial Guinea. Biological Conservation, 126, 206–215.
Eves, H. E. (2001). The bushmeat crisis task force. In The apes: Challenges for the 21st century. Conference Proceedings, Brookfield Zoo, May 10–13, 2000 (pp. 230–231). Brookfield, IL: Chicago Zoological Society.
Eves, H. E., & Bakarr, M. I. (2001). Impacts of bushmeat hunting on wildlife populations in West Africa’s Upper Guinea forest ecosystem. In M. I. Bakarr, G. A. B. daFonseca, R. Mittermeier, A. B. Rylands, & K. W. Painemilla (Eds.), Hunting and bush-meat utilization in the African rain forest (pp. 39–57). Washington, DC: Conservation International.
Fa, J. E., & Brown, D. (2009). Impacts of hunting on mammals in African tropical moist forests: A review and synthesis. Mammal Review, 39, 231–264.
Fa, J. E., Currie, D., & Meeuwig, J. (2003). Bushmeat and food security in the Congo Basin: Linkages between wildlife and people’s future. Environmental Conservation, 30, 71–78.
Fa, J. E., Garcia Yuste, J. E., & Castelo, R. (2000). Bushmeat markets on Bioko Island as a measure of hunting pressure. Conservation Biology, 14, 1602–1613.
Fa, J. E., Juste, J., Burn, R. W., & Broad, G. (2002a). Bushmeat consumption and preferences of two ethnic groups in Bioko Island, West Africa. Human Ecology, 30, 397–416.
Fa, J., Olivero, J., Farlan, M. A., Marquez, A. L., Duarte, J., Nackoney, J., et al. (2015a). Correlates of bushmeat in markets and depletion of wildlife. Conservation Biology, 29, 805–815.
Fa, J. E., Olivero, J., Real, R., Farlan, M. A., Marquez, A. L., Vargas, J. M., et al. (2015b). Disentangling the relative effects of bushmeat availability on human nutrition in central Africa. Scientific Reports, 5, 8168.
Fa, J. E., Peres, C. A., & Meeuwig, J. (2002b). Bushmeat exploitation in tropical forests: An intercontinental comparison. Conservation Biology, 16, 232–237.
Fa, J. E., Ryan, S. F., & Bell, D. J. (2005). Hunting vulnerability, ecological characteristics and harvest rates of bushmeat species in Afrotropical forests. Biological Conservation, 121, 167–176.
Fashing, P., & Cords, M. (2000). Diurnal primate densities and biomass in the Kakamega Forest: An evaluation of census methodology. American Journal of Primatology, 50, 139–152.
Fuentes, A. (2012). Ethnoprimatology and the anthropology of the human-primate interface. Annual Review of Anthropology, 41, 101–117.
Golden, C. D., Fernald, L. C. H., & Brashares, J. S. (2011). Benefits of wildlife consumption to child nutrition in a biodiversity hotspot. Proceedings of the National Academy of Sciences, 108, 19653–19656.
Henschel, P., Hunter, L. T. B., Coad, L., Abernathy, K. A., & Muhlenberg, M. (2011). Leopard prey choice in the Congo Basin rainforest suggests exploitative competition with human bushmeat hunters. Journal of Zoology, 285, 11–20.
Hodgkinson, C. (2009). Tourists, gorillas and guns: Integrating conservation and development in the Central African Republic. Unpublished PhD dissertation, University College, London.
Jost Robinson, C. A. (2012). Beyond hunter and hunted: An integrative anthropology of human-wildlife dynamics and resource use in a Central African Forest. Unpublished PhD dissertation, Purdue University.
Jost Robinson, C. A., Daspit, L. L., & Remis, M. J. (2011). Multi-faceted approaches to understanding changes in wildlife and livelihoods in a protected area: A conservation case study from the Central African Republic. Environmental Conservation, 38, 247–255.
Jost Robinson, C. A., & Remis, M. J. (2014). Entangled realms: Hunters and hunted in the Dzanga Sangha Dense Forest Reserve (RDS), Central African Republic. Anthropology Quarterly, 87, 613–633.
Klailova, M., Casanova, C., Henschel, P., Lee, P., Rovero, F., & Todd, A. (2013). Non-human predator interactions with wild great apes in Africa and the use of camera traps to study their dynamics. Folia Primatologica, 83(3–6), 312–328.
Kümpel, N. F., Milner-Gulland, E. J., Rowcliffe, J. M., & Cowlishaw, G. (2008). Impact of gun-hunting on diurnal primates in continental Equatorial Guinea. International Journal of Primatology, 29, 1065–1082.
Laurance, W. F., Croes, B. M., Tchignoumba, L., Lahm, S. A., Alonso, A., Lee, M. F., et al. (2006). Impacts of roads and hunting on Central African rainforest mammals. Conservation Biology, 20, 1251–1261.
Linder, J. M., & Oates, J. F. (2011). Differential impact of bushmeat hunting on monkey species and implications for primate conservation in Korup National Park, Cameroon. Biological Conservation, 144, 738–745.
Malone, N., Fuentes, A., Riley, E. P., Remis, M., Wade, A. H., & Robinson, C. J. (2014). Ethnoprimatology: Critical interdisciplinarity and multispecies approaches in anthropology. Critique of Anthropology, 34, 8–29.
Marshall, A. R., Lovett, J. C., & White, P. C. L. (2008). Selection of line transect methods for estimating density of group-living animals: Lessons from primates. American Journal of Primatology, 70, 452–462.
Milner-Gulland, E. J., & Bennett, E. L. (2003). Wild meat: The bigger picture. Trends in Ecology & Evolution, 18, 351–357.
Motte-Florac, E., Bahuchet, S., & Thomas, J. M. C. (1993). The role of food in the therapeutics of the Aka pygmies of the Central African Republic. In C. M. Hladik et al. (Eds.), Tropical forests, food, and people (pp. 549–560). Paris: UNESCO.
Njiforti, H. L. (1996). Preferences and present demand for bushmeat in north Cameroon: Some implications for wildlife conservation. Environmental Conservation, 23, 149–155.
Noss, A. J. (1995). Duikers, cables, and nets: A cultural ecology of hunting in a Central African forest. Unpublished PhD dissertation, University of Florida.
Noss, A. J. (1998). Cable snares and bushmeat markets in a Central African forest. Environmental Conservation, 25, 228–233.
Peterson, D. (2003). Eating apes. Berkeley, CA: University of California Press.
Plumptre, A. J. (2000). Monitoring mammal populations with line transect techniques in African forest. Journal of Applied Ecology, 37, 356–368.
Plumptre, A. J., & Cox, D. (2006). Counting primates for conservation: Primate surveys in Uganda. Primates, 37, 65–73.
Remis, M. J. (2000). Preliminary assessment of the impacts of human activities on gorillas Gorilla gorilla gorilla and other wildlife at Dzanga-Sangha Reserve, Central African Republic. Oryx, 34, 56–65.
Remis, M. J., & Jost Robinson, C. A. (2012). Reductions in primate abundance and diversity in a multiuse protected area: Synergistic impacts of hunting and logging in a Congo Basin Forest. American Journal of Primatology, 74, 602–612.
Robinson, J. G., & Bennett, E. L. (2000). Hunting for sustainability in the tropics. New York: Columbia University Press.
Schenck, M., Effa, E. N., Starkey, M., Wilkie, D., Abernathy, K., Telfer, P., et al. (2006). Why people eat bushmeat: Results from two-choice, taste tests in Gabon, Central Africa. Human Ecology, 34, 433–445.
Trotter, R. T., & Schensul, J. J. (1998). Methods in applied anthropology. In H. R. Bernard (Ed.), Handbook of methods in cultural anthropology (pp. 691–736). Lanham, MD: AltaMira Press.
Weinbaum, K. Z., Brashares, J. S., Golden, C. D., & Getz, W. M. (2013). Searching for sustainability: Are assessments of wildlife harvests behind the times? Ecology Letters, 16, 99–111.
White, L. J. T., & Edwards, A. (2000). Conservation research in the African rain forests: A technical handbook. New York: Wildlife Conservation Society.
Wilkie, D. S., & Godoy, R. A. (2001). Income and price elasticities of bushmeat demand in lowland Amerindian societies. Conservation Biology, 15, 1–9.
Wilkie, D. S., Shaw, E., Rotberg, F., Morelli, G., & Auzel, P. (2000). Roads, development and conservation in Congo Basin. Conservation Biology, 14, 1614–1622.
Wilkie, D. S., Starkey, M., Abernethy, K., Nstame Effa, E., Telfer, P., & Godoy, R. (2005). Role of prices and wealth in consumer demand for bushmeat in Gabon, Central Africa. Conservation Biology, 19, 268–274.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Robinson, C.J., Daspit, L.L., Remis, M.J. (2016). Monkeys on the Menu? Reconciling Patterns of Primate Hunting and Consumption in a Central African Village. In: Waller, M. (eds) Ethnoprimatology. Developments in Primatology: Progress and Prospects. Springer, Cham. https://doi.org/10.1007/978-3-319-30469-4_3
Download citation
DOI: https://doi.org/10.1007/978-3-319-30469-4_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-30467-0
Online ISBN: 978-3-319-30469-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)