
Abstract
The northern Bering Sea in the Pacific Arctic is seasonally ice covered, and includes winter polynyas (open areas in ice-covered seas). Overall, this marine system climatically in the winter is little different from the ice-covered Arctic Ocean proper. The macrofauna living in the sediments in this region provide prey for sentinel benthivores in the Arctic, including walrus, gray whales, bearded seals, and diving sea ducks. We have been evaluating the area south of St. Lawrence Island (SLI) for the last 25 years (1990–2015) and this chapter provides a synthesis and overview of ecosystem dynamics in this region seasonally. We initiated our studies during the spring-summer sea ice transition to open water period (May–August) for a decade from 1988 to 1998, but had the opportunity from 1999 to 2007 to use icebreakers for research cruises in the late winter to early spring transition period (March–June), as well as for late winter observations (March–April) from 2008 to 2010. In 2010 we facilitated the development of the Distributed Biological Observatory (DBO), a national and international network of coordinated cruises at five latitudinally arrayed transect lines from south of SLI (DBO1) north to Barrow, Alaska (DBO5). DBO1 includes five time series stations that have been sampled over two decades. The opportunity to sample in different seasons has allowed us to evaluate the phenology of physical, chemical, and biological patterns using an ecosystem approach to understand the transitions. The overall picture that emerges from these seasonal observations is of a biologically active system with strong atmospheric and hydrographic controls that predetermine nutrient distributions and water column mixing through brine rejection. These processes ultimately influence the intensity of seasonal productivity and associated activities of biological components from ice algae to zooplankton to the benthos, and to apex predators that are emblematic features of this subpolar ecosystem that is a sentinel for Arctic-subarctic interactions.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
Polynya is a Russian-origin word that is used to describe ice-free areas in otherwise ice-covered seas. Polynyas are a common feature in both Arctic and Antarctic waters, even at temperatures well below freezing, when prevailing winds blow sea ice away from islands or shorelines (latent heat polynyas), or where warmer waters are upwelled to the surface (sensible heat polynyas). Where these features are predictable from year to year, polynyas often attract higher trophic organisms such as diving ducks and marine mammals that require open water refuges for feeding and rest in frozen sea habitats (Stirling 1980; Gilchrist and Robertson 2000).
The year 2015 marks more than a quarter century since the first internationally coordinated effort to study polynyas, the International Arctic Polynya Program (IAPP; Mikkelsen 2015), undertaken by the Arctic Ocean Sciences Board, a non-governmental international science cooperation entity that has now been merged into the International Arctic Science Committee. Interest in the systematic study of polynyas was inspired in part by the potential for understanding the impacts of future climate change in the polar regions by studying persistent areas where sea ice was not present seasonally (Smith and Barber 2007).
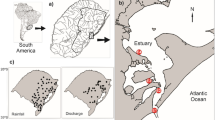
The polynya that persists in the winter to the south of Saint Lawrence Island (SLIP) in the Bering Sea was one of the first candidate polynyas that was targeted for study in the IAPP (Alexander and Muench 1988), and was supported by the US National Science Foundation funding initiated in 1990 to the two authors of this chapter (Fig. 1a, b; Grebmeier and Cooper 1995). Although other studies of polynyas in succeeding years, in the Northeast Water (Greenland), in the North Water (Baffin Bay), the Laptev Sea flaw lead system, and the Cape Bathurst region of the Canadian Arctic have been more intensive and more widely supported on an international scale, the initial study of the SLIP, which was summarized in a special issue of the Journal of Geophysical Research on leads and polynyas (Grebmeier and Cooper 1995), provided an initial understanding of the biological functioning of this shallow continental shelf winter polynya and its influence on benthic biology of the northern Bering Sea. The physics of the SLIP have also been broadly studied, particularly its importance as an area of sea ice production that influences seasonal ice cover in the Bering Sea (e.g., Pease 1987; Drucker et al. 2003; Danielson et al. 2006).

(a) Study region in the northern Bering Sea south of St. Lawrence Island, USA, with general current patterns by water type. (b) Example of the standard macrofaunal distribution of three benthic community station cluster groups sampled during process cruises in the study region: northern (triangles), eastern (circles), western (squares)
Our initial biologically oriented study (Grebmeier and Cooper 1995) was followed up by a number of other efforts that provide a fuller context of the importance of this polynya within the northern Bering Sea ecosystem (e.g., Cooper et al. 2002, 2009, 2012, 2013; Clement et al. 2004; Lovvorn et al. 2005, 2009, 2014; Grebmeier 2012; Grebmeier et al. 2006a, b; 2015; Grebmeier and Barry 2007; Jay et al. 2014). Data collection has continued over this past quarter century on a number of follow-on projects in the northern Bering Sea, and we use this opportunity to synthesize these findings from over the years with new insights that are developing from the internationally coordinated Distributed Biological Observatory (DBO) project that is annually sampling at biologically productive regions of the northern Bering Sea (including within the SLIP region), and adjoining Chukchi Sea.
The Northern Bering Sea, roughly from St. Matthew Island north, is a distinct ecosystem and functions differently than the pelagic system to the south. This difference is particularly striking in the winter, when walruses, ice seals, and spectacled eiders congregate in large numbers to take advantage of abundant food supplies on the seafloor, and also in some cases from under ice biota such as arctic cod and euphausiids. Productivity in the water column is low due to light limitations, but west-to-east decreases in chlorophyll and nutrients are already present as they are later in the seasonal cycle when massive sea ice edge blooms occur (Cooper et al 2012, 2013).
Our observations indicate the importance of the rich benthic communities south of St. Lawrence Island to top apex predators during winter foraging (Grebmeier 2012) and have contributed to understanding the seasonal cycle of this Arctic-subarctic system that has a winter polynya (Grebmeier and Barry 2007). The benthic macrofauna has historically high biomass levels on the western side of the system, dominated by tellinid, nuculid, and nuculanid bivalves (Grebmeier and Cooper 1995; Simpkins et al. 2003; Grebmeier et al. 2006a, 2015; Grebmeier 2012). By comparison, the eastern section has low infaunal biomass dominated by tunicates, amphipods, polychaetes, and various bivalves. We characterized both macrofaunal and epifaunal habitats and evaluated infaunal to epibenthic relationships, associated abiotic influences such as sediment types, and overlying water column hydrographic measurements. Particularly near Saint Lawrence and Saint Matthew Islands and along the northwest Alaska coast, distinct and diverse epifaunal communities are present that reflect system energy and geomorphology. For example, sand dollars are locally abundant inshore of the boundary between Bering Shelf Water (BSW) south of St. Lawrence Island and Alaska Coastal Water (ACW) (Fig. 1a). Large portions of the continental shelf of the Bering Sea are dominated by brittle stars (Ophiuroidea), including on the outer shelf southwest of Saint Lawrence Island where fine silt and clay sediments occur. By contrast, in sandier sediments to the southeast of Saint Lawrence Island, a wider range of mobile and sessile epifauna (e.g., crabs, gastropods, and tunicates) are present (Lovvorn et al. 2005). These patterns of macrofaunal and epibenthic community separation suggest a potential link between infaunal and epifaunal communities through trophic interactions or the influence of environmental parameters on both communities at similar scales. Patterns in hydrodynamics and/or sea ice and therefore carbon supply are potential driving factors.
Sediment oxygen demand (SOD) has also been determined at multiple sites during many of the cruises as an indicator of carbon supply to the benthos (Grebmeier and Cooper 1995; Cooper et al. 2012, 2013). Focused experiments on temperature controls on SOD in 2007 and 2009 indicated that SOD increased with both temperature and carbon supply (Bailey et al. 2009; Cooper et al. 2013). Nutrient exchanges between the sediments and the water column varied and suggested that interactions with benthic fauna affect rates of exchange, as is the case in other areas in the Pacific Arctic shelf region (Mathis et al. 2014). Sediment cores for these temperature-controlled experiments were collected solely from time series stations maintained since 1990 that had the best time series record for sediment respiration in this region of the Bering Sea. The object of the experiments was to test for changing environmental impacts that may be driving the long-term declines in benthic communities and populations that have now been documented over our full sampling record (Grebmeier et al. 2006b; Grebmeier 2012).
The implications of these declines are that it may be affecting apex predators such as walruses, bearded seals, and several species of diving ducks, including spectacled eiders, who dive to the shallow seafloor (40–60 m) to forage on the abundant clams and other benthic organisms. Moreover, our observations indicate that spectacled eiders can be excluded from optimal foraging areas by the position and extent of seasonal sea ice (Lovvorn et al. 2009, 2014). The benthic biomass declined over the past several decades (Grebmeier et al. 2006b; Grebmeier 2012), and also may bring additional changes to the ecosystem, including the movement northward of industrial fishing operations as water temperature warms, which will have further negative consequences for the ecosystem.
Another key higher trophic organism to consider are the walruses that use the region south of St. Lawrence Island in late winter for rest and feeding (Jay et al. 2012, 2014). Feeding patterns reflect benthic biomass and seafloor community structure. Pacific walruses forage on the seafloor, primarily for benthic invertebrates, and the wide continental shelves of the Chukchi and Bering Seas provide extensive areas of high benthic biomass as prey for these upper trophic organisms. Resource selection, for food or habitat, has a direct impact on walrus survival capabilities and partitioning of populations. The heterogeneity or patchiness of prey and/or habitat resources can occur over multiple temporal and spatial scales. Individual species adapt to the changing environment and in turn this can influence population structure and demographic trends. Both spectacled eider and walrus satellite tracking data can be matched with benthic prey data and have been used in habitat selection analyses by the predators (Lovvorn et al. 2014; Jay et al. 2014; Sexson et al. 2016).
Synthesis Results and Discussion
Overview for Synthesis
Water column temperature, salinity, chlorophyll a content, nutrients, macro- and epifaunal collections, and sediment parameters constitute much of the data collected at stations in the SLIP region over the multiple cruises (see methods in Grebmeier and Cooper 1995; Grebmeier 2012; Grebmeier et al. 2015; Cooper et al. 2012, 2013). Specific to the seasonal summaries below, the benthic sampling of bottom sediments and associated fauna at stations from 1990 to present included four replicates of a 0.1 m2 van Veen grab for macrofaunal collections at each station (Grebmeier and Cooper 1995). Organisms were sieved from each of these grabs through 1 mm screens, preserved, and returned to the laboratory for species identification and determinations of biomass. Additional sediment grabs were also undertaken at each station for surface sediment analysis, including determinations of sediment chlorophyll a content, total organic carbon and nitrogen, and other sediment chemical parameters, when practical.
Summer Sampling in SLIP (July–September 1990–2015)
The first series of samples in the SLIP occurred during the summer period (June–Sept 1990–1999, n = 5 cruises), with an additional 15 cruises using a Canadian icebreaker that sampled a subset of five time-series sites in the western high benthic biomass region offshore of St. Lawrence Island (SLI) (2000–2015; see Grebmeier et al. 2015 for specific details on cruises and methods). A number of prior publications highlighted the spatial variability in water masses south of SLI, with nutrient-rich Anadyr Water (AW; salinity >32.5) occurring in the western region of the study area as well as in the northern area just south of SLI, Alaska Coastal Water (ACW; salinity <31.8) in the eastern region, and Bering Shelf Water (BSW; salinity = 31.8–32.5) occurring between AW and ACW (Grebmeier and Cooper 1995; Cooper et al. 2012, 2013). Most of the water column chlorophyll a descends to the benthos after the spring bloom, so by midsummer, water column stratification has sealed off the cold bottom waters (<−1 °C) remaining near bottom from the winter polynya formation period. This is a function of sea ice formation; when cold, dense saline water is injected into the water as sea ice forms, and by summer these cold bottom waters are separated from surface waters that warm to 4–8 °C. The bottom sediments have higher silt and clay content and sediment chlorophyll a content in the western region compared to the eastern, less nutrient-rich area (Grebmeier and Cooper 1995; Grebmeier 2012). Benthic macrofaunal biomass is highest in the western region, dominated by bivalves and polychaetes, with lower biomass of similar fauna, along with amphipods, in the eastern region (Grebmeier and Cooper 1995; Simpkins et al. 2003; Grebmeier 2012). The northern sector, just south of SLI, is dominated by amphipods and bivalves.
Spring (April–June 1999–2007)
We have had four process cruises in the SLIP region in spring (April–June from 1999 to 2007). These spring cruises required us to sample in the ice-covered seas south of SLI using US Coast Guard (USCG) icebreakers that allowed us to observe the ice edge and spring bloom period as the ice receded (Clement et al. 2004). A large part of the area on the central shelf was occupied by Bering Shelf Water with intermediate salinity (32–32.4). Stations close to the south side of SLI showed salinities of ~33 which may be characteristic for waters advected with the southern branch of the Anadyr current that bifurcates on the west side of SLI (the other branch flows northeast through Anadyr Strait separating the northwest cape of St. Lawrence Island from continental Asia; see Fig. 1a). Higher saline water was found to the southeast of SLI. The high salinity values (~33.5–34) of these waters are likely caused by brine rejection events during ice formation (Clement et al. 2004). A modeling study further elaborated into the processes driving physical and biological water column properties during this time (Clement et al. 2005).
This spring period as ice retreats is the most productive period for both ice algae and large diatoms that dominated the primary production and higher water column chlorophyll occurred in the western region (Cooper et al. 2002, 2012). Zooplankton populations are relatively low (Lovvorn et al. 2005), with high export of primary produced organic carbon directly to the benthos (Cooper et al. 2002; Grebmeier and Dunton 2000; Grebmeier 2012). This high export of labile carbon allows for the buildup of high bivalve and polychaete biomass in the western offshore area south of SLI (Fig. 2a–d). The eastern area, as mentioned previously for the summer period, has smaller benthic biomass of bivalves, amphipods, and polychaetes.

Distribution of macrofaunal biomass (gC/m2) and macrofaunal types, by biomass, in spring 2006 for USCGC Healy cruises (HLY0601; a, b, respectively) and 2007 (HLY0702; c, d, respectively)
Winter (March 2008–2010)
Late winter observations in the northern Bering Sea have been rare, but three USCG icebreaker deployments during March in 2008–2010 have provided new insights on how the ecosystem functions and preconditions the productive spring bloom (Cooper et al. 2013) in comparison with spring studies in 2006 and 2007 (Cooper et al. 2012). Salinity is the dominant physical parameter that determines horizontal density variations in spring, with the warmest water offshore to the southwest, but the rest of the study area is influenced by cold <1 °C water produced as sea ice forms at the surface in a well-mixed water column. While water column chlorophyll concentrations were two orders of magnitude lower in late winter than observed during the spring bloom in May, sea ice algal inventories of chlorophyll were high and corresponded to the distribution of water masses with high nutrient content (Cooper et al. 2013). Brine injection while the late winter SLI polynya is active can mix inorganic nutrients from the bottom upward, and the specific vectors of the prevailing north to easterly winds influence the distribution of high nutrient waters that originate on the northwestern Bering Sea Shelf near and within the Gulf of Anadyr. Thus, conditions such as wind and air temperature in March have an influence on the intensity of the spring bloom, in addition to the actual pattern and timing of sea ice retreat. The underlying benthos continued to show the signature of highest benthic biomass of bivalves and polychaetes in the western region of the SLI region, with lower biomass to the east (Fig. 3a–d). After the spring bloom, water column stratification limits phytoplankton production, although the bottom benthic communities continue to build biomass over the summer based on organic carbon resources that were produced both in the SLI system and advected in from the southern Bering Sea.

Distribution of macrofaunal biomass (gC/m2) and macrofaunal types, by biomass, in late winter 2008 for USCGC Healy cruises (HLY0801; a, b, respectively) and 2009 (HLY0901; c, d, respectively)
The Northern Bering Sea: Interannual Variability and Change
In addition to the water column data discussed above, benthic infaunal abundance and biomass data for the area south of SLI were also compiled from the many cruises undertaken over the span of 25 years in the SLIP region (details summarized in Grebmeier et al. 2006b, 2015; Grebmeier 2012). Three distinct community groupings were identified on every cruise by cluster and statistical analyses of macrofaunal abundance and biomass (Fig. 1b; discussed previously). However, we have seen dramatic changes in benthic biomass for the dominant three species of bivalves in the western region that are key to the spectacled eider and walrus prey base (Grebmeier et al. 2006b; Grebmeier 2012; Lovvorn et al. 2014). Specifically, nuculanid bivalves continue to decline in biomass, particularly under the influence of nutrient-rich Anadyr water (Grebmeier 2012). However, smaller nuculid bivalves have increased in abundance closer to SLI, but it is not clear if the spectacled eiders that feed there in the winter can take advantage of this shift in food supplies due to ice conditions (Grebmeier 2012; Lovvorn et al. 2003, 2014).
Time Series Stations Within the “Western” Cluster Group Under Anadyr Water
Time series analyses of five stations in the area of highest biomass show that these locations all cluster in a productive western group (Grebmeier et al. 2006b; Grebmeier 2012). Sampling at these sites indicates that the three most southern sites in this cluster group are changing both in infaunal composition and overall biomass in response to larger scale physical and biological forcing that is acting upon the system (Fig. 4a, b; Grebmeier 2012). Statistical analyses of macrofaunal populations and environmental parameters have identified water column depth and sediment grain size as the primary factors influencing benthic macrofaunal community structure, both for stations that are part of our formal time series (r = 0.619, p ≤ 0.05) and for more spatially extensive cruises. Since depth is not changing over time, but is highly correlated with greater silt and clay (Grebmeier 2012; Grebmeier et al. 2015), the primary factor leading to the changing benthic community composition is likely an increase in the percent composition of the silt and clay fraction. These sediment pattern changes suggest reduced currents that are increasing the silt and clay fraction of sediments and altering benthic community composition. Besides a decline in overall station biomass, we observe a shift in macrofaunal dominance from bivalves to polychaetes in 2009 (Fig. 4a, b-red box) where sediments fined. We are continuing tracking of these five time series sites annually as part of the DBO project (Grebmeier et al. 2010, 2015). These stations serve as the most southern line of the DBO network, which is being developed as an international cooperative approach to detecting change in arctic ecological systems.

Benthic macrofaunal biomass (gC/m2) and dominant fauna (% of total station biomass) in (a) March 2008 (HLY0801) and (b) March 2009 (HLY0901) south of St. Lawrence Island in the northern Bering Sea. The red box surrounds the three time series sites where decadal biomass declines have been observed (Grebmeier 2012) and shows the change in dominance from bivalves to polychaetes that occurred in 2009
Benthivores
Late winter is an important foraging period for benthic-feeding Pacific walrus and diving sea ducks, such as the king, spectacled, and common eider. As might be expected, feeding patterns reflect the underlying benthic biomass and seafloor community structure. Sea ice dynamics in addition to benthic food availability also plays a role in the scale and distributions in particular of spectacled eiders.
In our study we characterized benthic habitats and evaluated macrofaunal benthic community relationships to overlying water column hydrographic measurements, abiotic sediments, and sea ice cover. Sediment organic carbon content indicates a deposition zone to the southwest of SLI (Grebmeier 2012) that coincides with high benthic macrofaunal biomass and areas of walrus predation (Jay et al. 2014). We continue to track the overall system health at select stations. These data indicate that after nearly a 50 % decline in carbon deposition and benthic macrofaunal biomass from the 1990s to early 2000s (Grebmeier et al. 2006b), these parameters have now stabilized at lower carbon deposition and benthic biomass levels (Grebmeier 2012). Despite these changes, the western region south of St. Lawrence Island is still a key area for upper trophic predators.
Overall Summary
The new insights on how the SLIP ecosystem functions over annual cycles have required seasonal sampling to facilitate understanding the preconditioning required for development of the productive spring bloom. While water column chlorophyll concentrations are two orders of magnitude lower in late winter (March–early April) than observed during the spring bloom in May, sea ice algal inventories of chlorophyll are high and correspond to the distribution of water masses with high nutrient content. Nutrient availability clearly modulates open water and sea ice productivity and the transmission of organic material to the shallow benthos following sea ice retreat. In some areas, significant late winter re-suspension of ammonium also occurs from sediments. Active brine injection can mix these inorganic nutrients, and the specific vectors of the prevailing north to easterly winds influence the distribution of high nutrient waters that originate on the northwestern Bering Sea Shelf near and within the Gulf of Anadyr. Thus conditions such as wind and air temperature in March have an influence on the intensity of the spring bloom, in addition to the actual pattern and timing of sea ice retreat.
Feeding patterns of walruses and spectacled eiders reflect benthic biomass, seafloor community structure, and at least in the case of spectacled eiders that require sufficient open water to dive successfully sea ice dynamics. Kittlitz’s murrelets, which feed on zooplankton, are another late-winter migrant that were observed to take advantage of open water in the polynya south of Saint Lawrence Island. In experimental incubations, high sediment uptake rates occurred with both higher seawater temperatures and increased organic carbon (food supply). Sediment organic carbon content indicates a deposition zone to the southwest of SLI, and matches up well with high sediment oxygen update and benthic infaunal biomass. We continue to track the overall system health at select stations. Times series data indicate a “fining” of sediment (increased ≥5phi size silt and clay particles), indicative of slowing currents and increasing settling of fine particles to the benthos. We are evaluating these sediment grain size trends with both macrofaunal biomass and declined sediment oxygen uptake rates.
The overall picture that emerges from these early seasonal observations is of a biologically active system with strong atmospheric and hydrographic controls. These controls predetermine nutrient distributions and water column mixing through brine rejection and ultimately influence the intensity of seasonal productivity and the subsequent seasonal activities of biological components from ice algae to zooplankton to the benthos, and to apex predators that are emblematic features of this subpolar ecosystem.
References
Alexander V, Muench R (1988) Arctic polynyas. Eos, Trans Am Geophys Union 69:572
Bailey EM, Ceballos MAC, Grebmeier JM, Boynton WR (2009) The effects of temperature and carbon addition on Arctic sediment oxygen and nutrient exchanges. Oral presentation at Estuaries and Coasts in a Changing World: Coastal and Estuarine Research Federation 20th Biennial Conference, November 1-5, 2009, Portland, Oregon
Clement JL, Cooper LW, Grebmeier JM (2004) Late-winter water column and sea ice conditions in the northern Bering Sea. J Geophys Res 109, C03022. doi:10.1029/2003JC002047
Clement JL, Maslowski W, Cooper LW, Grebmeier JM, Walczowski W (2005) Ocean circulation and exchanges through the northern Bering Sea: 1979-2001 model results. Deep-Sea Res II Top Stud Oceanogr 52:3509–3540
Cooper LW, Grebmeier JM, Larsen IL, Egorov VG, Theodorakis C, Kelly HP, Lovvorn JR (2002) Seasonal variation in water column processes and sedimentation of organic materials in the St. Lawrence Island polynya region, Bering Sea. Mar Ecol Prog Ser 226:13–26
Cooper LW, Lalande C, Pirtle-Levy RS, Larsen IL, Grebmeier JM (2009) Seasonal and decadal shifts in particulate organic matter processing and sedimentation in the Bering Strait Shelf Region. Deep-Sea Res II Top Stud Oceanogr 56:1316–1325
Cooper LW, Janout MA, Frey KE, Pirtle-Levy R, Guarinello ML, Grebmeier JM, Lovvorn JR (2012) The relationship between sea ice break-up, water mass variation, chlorophyll biomass, and sedimentation in the northern Bering Sea. Deep Sea Res Part II 65:141–162
Cooper LW, Sexson MG, Grebmeier JM, Gradinger R, Mordy CW, Lovvorn JR (2013) Linkages between sea ice coverage, pelagic-benthic coupling and the distribution of spectacled eiders: observations in March 2008, 2009 and 2010 from the northern Bering Sea. Deep-Sea Res II Top Stud Oceanogr 94:31–43
Danielson S, Aagaard K, Weingartner T, Martin S, Winsor P, Gawarkiewicz G, Quadfasel D (2006) The St. Lawrence polynya and the Bering shelf circulation: New observations and a model comparison. J Geophys Res-Oceans 111(C9): doi 10.1029/2005jc003268
Drucker, R., S. Martin, and R. Moritz. 2003. Observations of ice thickness and frazil ice in the St. Lawrence Island polynya from satellite imagery, upward looking sonar, and salinity/temperature moorings, J. Geophys. Res., Oceans 108(C5), 3149, doi:10.1029/2001jc001213
Gilchrist HG, Robertson GJ (2000) Observations of marine birds and mammals wintering at polynyas and ice edges in the Belcher Islands, Nunavut, Canada. Arctic: 61–68
Grebmeier JM (2012) Biological community shifts in Pacific Arctic and sub-Arctic seas. Ann Rev Mar Sci 4:63–78
Grebmeier JM, Barry JP (2007) Benthic processes in polynyas. In: Smith WO Jr, Barber DG (eds) Polynyas: Windows to the World, vol 74, Oceanography series. Elsevier, Amsterdam, pp 363–390
Grebmeier JM, Cooper LW (1995) Influence of the St. Lawrence Island polynya on the Bering Sea benthos. J Geophys Res 100:4439–4460
Grebmeier JM, Dunton KH (2000) Benthic processes in the northern Bering/Chukchi seas: status and global change. In: Huntington HP (ed) Impacts of Changes in Sea Ice and Other Environmental Parameters in the Arctic. Marine Mammal Commission Workshop, Girdwood, Alaska, 15–17 February 2000, pp 80–93
Grebmeier JM, Cooper LW, Feder HM, Sirenko BI (2006a) Ecosystem dynamics of the Pacific influenced Northern Bering and Chukchi Seas. Prog Oceanogr 71:331–361
Grebmeier JM, Overland JE, Moore SE, Farley EV, Carmack EC, Cooper LW, Frey KE, Helle JH, McLaughlin FA, McNutt L (2006b) A major ecosystem shift in the northern Bering Sea. Science 311:461–1464
Grebmeier JM, Moore SE, Overland JE, Frey KE, Gradinger R (2010) Biological response to recent Pacific Arctic sea ice retreats. Eos, Trans Am Geophys Union 91:161–162
Grebmeier JM, Bluhm BA, Cooper LW, Danielson S, Arrigo KR, Blanchard AL, Clark JT, Day RH, Frey KE, Gradinger RR, Kedra M, Konar B, Kuletz KJ, Lee SH, Lovvorn JR, Norcross BL, Okkonen SR (2015) Ecosystem characteristics and processes facilitating persistent macrobenthic biomass hotspots and associated benthivory in the Pacific Arctic. Prog Oceanogr 136:92–114
Jay CV, Udevitz MS, Kwok R, Fischbach AS, Douglas DC (2012) Divergent movements of walrus and sea ice in the northern Bering Sea. Mar Ecol Prog Ser 407:293–302
Jay CV, Grebmeier JM, Fischbach AS, McDonald TL, Cooper LW, Hornsby F (2014) Pacific Walrus (Odobenus rosmarus divergens) resource selection in the northern Bering Sea. PLoS One. doi:10.1371/journal.pone.0093035
Lovvorn JR, Richman SE, Grebmeier JM, Cooper LW (2003) Diet and body condition of spectacled eiders in pack ice of the Bering Sea. Polar Biol 26:259–267
Lovvorn JR, Cooper LW, Brooks ML, DeRuyck CC, Bump JK, Grebmeier JM (2005) Organic matter pathways to zooplankton and benthos under pack ice in late winter and open water in late summer in the north-central Bering Sea. Mar Ecol Prog Series 291:135–150
Lovvorn JR, Grebmeier JM, Cooper LW, Bump JK, Richman JG (2009) Modeling marine protected areas for threatened eiders in a climatically shifting Bering Sea. Ecol Appl 19:1596–1613
Lovvorn JR, Anderson EM, Rocha AR, Larned WW, Grebmeier JM, Cooper LW, Kolts JM, North CA (2014) Variable wind, pack ice, and prey dispersion affect the long-term adequacy of protected areas for an Arctic sea duck. Ecol Appl 24:396–412
Mathis JT, Grebmeier JM, Hansell DA, Hopcroft RR, Kirchman D, Lee SH, Moran SB (2014) Carbon biogeochemistry of the western Arctic: production, export and ocean acidification. In: Grebmeier JM, Maslowski W (eds) The Pacific Arctic Region: Ecosystem Status and Trends in a Rapidly Changing Environment. Springer, Dordrecht, pp 223–268
Mikkelsen N (2015) History of the Arctic Ocean Sciences Board (AOSB) and Cooperation with other Organizations. In: Rogne O, Rachold V, Hacquebord L, Corell R. (eds) IASC after 25 years, Special Issue of the IASC Bulletin. ISBN 979-3-9813637-7-7. Potsdam, Germany
Pease CH (1987) The size of wind-driven coastal polynyas. J Geophys Res 92(C7):7049–7059
Sexson MG, Petersen MR, Breed GA, Powell AN (2016) Shifts in the distribution of molting Spectacled Eiders (Somateria fischeri) indicate ecosystem change in the Arctic. Condor 118:463–476
Simpkins MA, Hiruki-Raring LS, Sheffield G, Grebmeier JM, Bengston JL (2003) Habitat selection by ice-associated pinnipeds near St. Lawrence Island, Alaska in March 2001. Polar Biol 26:577–586
Smith WO Jr, Barber D (eds) (2007) Polynyas: Windows to the World, vol 74. Elsevier, Amsterdam
Stirling I (1980) The biological importance of polynyas in the Canadian Arctic. Arctic 33:303–315
Acknowledgements
We thank the technical support and graduate students over the years for environmental and biological measurements in the laboratory, most recently Linton Beaven and Stephanie Soques. Eva Bailey and Maria Ceballos kindly assisted in the time series sediment oxygen demand and nutrient exchange experiments in 2009. Alynne Bayard provided GIS mapping imagery for the benthic data. Funding for preparation of this chapter was provided by the US National Science Foundation (currently grant #ARC-1204082), National Oceanic and Atmospheric Administration, and North Pacific Research Board.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Jacqueline M. Grebmeier and Lee W. Cooper
Jacqueline M. Grebmeier and Lee W. Cooper
Jackie Grebmeier and Lee Cooper met at the University of California (UC) Bodega Bay Marine Laboratory in spring 1976 where we studied marine worms and played in the mud. Lee graduated from UC Santa Cruz in 1978 and Jackie graduated from UC Davis in 1977, with a Masters from Stanford University in 1978. In the early 1980s we went to the University of Washington for Masters programs in Botany (Lee) and Marine Affairs/Oceanography (Jackie), then northward to Alaska where we undertook research on Arctic ecosystems (Lee on seagrasses and stable isotopes and Jackie on benthic communities and pelagic-benthic coupling). We both received our Ph.D. degrees in Oceanography from the University of Alaska Fairbanks (Lee in 1986, the year we got married, and Jackie in 1987). We then did postdoctoral work in California, Lee at the University of California, Los Angeles in stable isotope geochemistry and Jackie at the University of Southern California on Arctic benthic systems and sediment radioisotopes. We moved to Tennessee in 1988 where we worked at both Oak Ridge National Laboratory and the University of Tennessee in Knoxville. In 2008 we took up Research Professor positions at the Chesapeake Biological Laboratory at the University of Maryland Center for Environmental Sciences where we are currently located. Besides science, we have managed to raise one daughter, Ruth Cooper, who is a senior at the University of Notre Dame, and one very friendly and tolerant cat named Emily.
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Grebmeier, J.M., Cooper, L.W. (2016). The Saint Lawrence Island Polynya: A 25-Year Evaluation of an Analogue for Climate Change in Polar Regions. In: Glibert, P., Kana, T. (eds) Aquatic Microbial Ecology and Biogeochemistry: A Dual Perspective. Springer, Cham. https://doi.org/10.1007/978-3-319-30259-1_14
Download citation
DOI: https://doi.org/10.1007/978-3-319-30259-1_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-30257-7
Online ISBN: 978-3-319-30259-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)