
Abstract
Members of the Archamoebae comprise free-living and endobiotic amoeboid flagellates, amoeboflagellates, and amoebae, with distinctive hyaline cytoplasm and bulging pseudopodia. They live in anoxic or microoxic habitats and are anaerobes, lacking typical mitochondria, as well as Golgi stacks, plastids, and normal peroxisomal microbodies. They have a distinctive flagellar apparatus present in all flagellated members of the group. Life cycles of individual species can include flagellates, amoebae of various sizes, and cysts. In recent years, the group has been divided into five separate families, Mastigamoebidae, Entamoebidae, Pelomyxidae, Tricholimacidae, and Rhizomastixidae, whose interrelationships have not been completely resolved. Here, we clarify the composition of these groups and the circumscription of genera in the Archamoebae.
Access provided by CONRICYT-eBooks. Download reference work entry PDF
Similar content being viewed by others

Keywords
Summary Classification
-
●Archamoebae
-
●●Pelobiontida
-
●●●Mastigamoebina
-
●●●●Mastigamoebidae (Mastigamoeba, Iodamoeba, Endolimax)
-
●●●●Rhizomastixidae (Rhizomastix)
-
●●●Pelomyxina
-
●●●●Pelomyxidae (Pelomyxa, Mastigella)
-
●●Entamoebida
-
●●●Entamoebidae (Entamoeba)
[Archamoebae incertae sedis: Mastigina, Tricholimacidae (Tricholimax), Endamoeba]
Introduction
General Characteristics
The Archamoebae is a group of amoebae, amoeboid flagellates, and amoeboflagellates (i.e., organisms with both flagellates and amoebae in their life cycle), with distinctive hyaline cytoplasm and bulging pseudopodia (Fig. 1). They comprise three lineages that have collectively been referred to as pelobionts: mastigamoebids, pelomyxids, and Tricholimax, as well as Rhizomastix and entamoebids. All Archamoebae are anaerobic (some more aerotolerant than others) and lack normal mitochondria (which are reduced to remnants), Golgi stacks, plastids, and normal peroxisomal microbodies. Many contain endosymbiotic bacteria and archaea. Flagellated taxa have a distinctive cytoskeleton.

General appearances of genera in the Archamoebae. (a) In the dominant flagellated trophic stage, Mastigamoeba has a microtubular cone that connects the flagellar base to the nucleus. (b) Mastigamoeba also typically has amoeboid stages, which may be multinucleate. (c) Iodamoeba contains small aflagellate amoebae with hyaline lobopodia. (d) Endolimax forms small aflagellate amoebae with variable pseudopodia similar to those in Mastigamoeba. (e) Mastigina contains amoeboid flagellates with a limax appearance and no lateral pseudopodia. (f) Rhizomastix contains small amoeboid flagellates that have a microtubular rhizostyle connecting the flagellar base to the nucleus. (g) Entamoeba contains amoebae with hyaline lobopodia and granular cytoplasm. (h) Endamoeba includes amoebae with a characteristic pattern of nuclear chromatin and no strong distinction between hyaline and granular areas of cytoplasm. (i) Tricholimax includes a single species of multinucleate amoeboid flagellate with a short, nonfunctional flagellum. (j) Mastigella contains amoeboid flagellates where the flagellar base is not connected to the nucleus by the cone of microtubules; the dominant trophic stage may be an aflagellate amoeba. (k) Pelomyxa contains large amoebae with multiple nuclei and very short, nonfunctional flagella. Scale bar = 20 μm
Mastigamoebids currently include Mastigamoeba, Endolimax, and Iodamoeba: these include mostly free-living amoeboflagellates as well as endobiotic amoebae; many have distinctively hyaline cytoplasm and move with pseudopodia when attached to a substrate. In flagellate mastigamoebids, the flagellum is functional and has a conventional 9 + 2 microtubular arrangement, but there is a distinctively “languid” or slow flagellar beat arising from the lack of outer dynein arms in the flagellar axoneme. The base of the flagellum gives rise to a cone of microtubules that connect to the nucleus. Molecular phylogenies divide mastigamoebids into two clades, “A,” containing large species with a broad flagellar apparatus (e.g., Mastigamoeba balamuthi), and “B,” containing small species with a narrow flagellar apparatus and trailing pseudopodia (e.g., Mastigamoeba simplex) as well as Endolimax and Iodamoeba (Ptáčková et al. 2013; Pánek et al. 2016). Members of Endolimax and Iodamoeba have entirely lost the flagellar apparatus and were historically classified within entamoebids. They have only recently been transferred to mastigamoebids on the basis of molecular-phylogenetic analyses (Cavalier-Smith et al. 2004; Ptáčková et al. 2013; Stensvold et al. 2012; Poulsen and Stensvold 2016). Historically, the flagellated Mastigella has usually been thought of as a mastigamoebid, but it is more closely affiliated with Pelomyxa (Zadrobílková et al. 2015).
Pelomyxids include members of Mastigella and Pelomyxa. Mastigella includes amoebae and flagellated amoebae where the base of the flagellum gives rise to a microtubular cone that is not connected to the nucleus. Pelomyxa includes large (up to several millimeters) amoebae, with nonfunctional, short flagella that have a disordered microtubular arrangement and a microtubular cone that is unconnected to the nucleus or nuclei. Recent phylogenetic analyses have shown that Mastigella is closely related to Pelomyxa (Zadrobílková et al. 2015).
Rhizomastix was recently placed in the Archamoebae (Čepička 2011; Ptáčková et al. 2013). It includes amoeboflagellates with a single anterior flagellum, which live endobiotically in the intestines of insects and amphibians or are free-living in freshwater sediments. In Rhizomastix the cone of microtubules arising from the base of the flagellum has been modified into a tapering microtubular tube, the “rhizostyle.”
Entamoebids are aflagellated, mostly endobiotic amoebae that were, until relatively recently, classified among other amoebae and taxonomically separated from the pelobionts. They currently formally include the genus Entamoeba (historically also Endamoeba, Endolimax, and Iodamoeba) and have in the past also included several genera of currently uncertain phylogenetic position, such as Schizamoeba, Hydramoeba, and Malpighamoeba. Their morphological similarities to pelobionts were recognized several decades ago (Cavalier-Smith 1983, 1987a, b) and since confirmed with molecular phylogenetics. Most entamoebids are probably harmless commensals of the digestive tract in invertebrates or vertebrates, including humans (e.g., Entamoeba moshkovskii; Heredia et al. 2012); a recent description reports a free-living or commensal marine species (Shiratori and Ishida 2016). The most important is the parasitic Entamoeba histolytica, which causes the dangerous amebic dysentery of humans.
Mastigina is a poorly known genus, with few sightings and no molecular data, currently classified as incertae sedis (Pánek et al. 2016) though likely to be a member of the mastigamoebids. It has many similarities to Mastigamoeba but has a limax body shape where pseudopodia emerge only at the anterior or posterior ends. Its identity has historically been confused with that of Tricholimax (see section “Systematics and Taxonomy”, below).
Tricholimax hylae, a large multinucleate amoeba with a short, nonfunctional flagellum, is endobiotic in the hindgut of frog tadpoles. The phylogenetic placement of Tricholimax is unknown, in the absence of molecular data, so it is classified as Archamoebae incertae sedis (Pánek et al. 2016); nonetheless, it shows considerable similarity to Mastigella and Pelomyxa. Its identity has historically been confused with that of Mastigina.
Endamoeba is a poorly known genus; it includes aflagellate Archamoebae found in insects. Its morphological similarity to Entamoeba suggests it is likely to be a member of Entamoebidae, but it is currently classified as Archamoebae incertae sedis in the absence of molecular data (Pánek et al. 2016).
History of Knowledge and Literature
Taxonomic History of the Archamoebae as a Group
The name Archamoebae was introduced and used by Cavalier-Smith (1983, 1987a, b) and Cavalier-Smith et al. (2004) to group the pelomyxids, entamoebae, and mastigamoebids. The grouping of entamoebae, pelomyxids, and mastigamoebids was later supported by molecular phylogenetic analyses with complex evolutionary models (Cavalier-Smith et al. 2004; Edgcomb et al. 2002; Kudryavtsev et al. 2005; Milyutina et al. 2001; Nikolaev et al. 2006; Pánek et al. 2016; Ptáčková et al. 2013; Stensvold et al. 2012). The taxonomic concept of Archamoebae has been used at ranks of infraphylum and class (Cavalier-Smith 1998, 2013; Cavalier-Smith et al. 2004) and has been compositionally unstable (Cavalier-Smith 1991, 1997; Cavalier-Smith and Chao 1995). It is currently ranked as a class, Archamoebae Cavalier-Smith, 1983 (Pánek et al. 2016).
In its recent incarnations (e.g., Cavalier-Smith 2013; Ptáčková et al. 2013; Zadrobílková et al. 2015, 2016), Archamoebae has included four main clades, the entamoebae, pelomyxids, mastigamoebids, and Rhizomastix, with Tricholimax sometimes treated as a fifth clade (Cavalier-Smith 2013) or regarded as incertae sedis but probably part of the pelomyxids, as discussed above (Frolov 2011; Pánek et al. 2016; Ptáčková et al. 2013; Zadrobílková et al. 2016). Relationships between clades have recently been resolved using multigene phylogenetics, dividing the group clearly into entamoebids (Entamoeba) and pelobionts (pelomyxids; mastigamoebids and Rhizomastix), and the high-level taxonomy has been updated to reflect this, as presented below here (Pánek et al. 2016).
Flagellated mastigamoebids and pelomyxids have historically been considered together as “pelobionts.” The Order Pelobiontida was originally introduced to include only the genus Pelomyxa (Page 1976, 1987) and has occasionally been used at other ranks, e.g., Class Pelobionta (Krylov et al. 1980). Its trophic form being a large amoeba, Pelomyxa was until recently more usually classified with lobose amoebae (e.g., Bovee 1972; Bütschli 1880; Chatton 1925, 1953; Page 1976; Reichenow 1952; Siemensma 1987) but with increasing recognition that it was distinct from other large amoebae (Whatley and Chapman-Andresen 1990). Griffin (1988) revised the Order Pelobiontida to include Mastigamoeba, Mastigella, Mastigina, and Dinamoeba, on the basis of the ultrastructural evidence for flagella in Pelomyxa (Griffin 1979, 1988). Cavalier-Smith (1987a, b) created a different concept of pelobionts consisting of pelomyxids and entamoebids. However, this composition of the pelobionts has only rarely been used (e.g., Cavalier-Smith et al. 2004). The term “pelobiont” has since been used to encompass mastigamoebids and pelomyxids to the exclusion of entamoebae (Pánek et al. 2016).
The name Entamoebae was created to group aflagellate Archamoebae (Entamoeba, Endamoeba, Endolimax, and Iodamoeba). However, Endolimax and Iodamoeba have been removed from entamoebids, Endamoeba is currently regarded as Archamoebae incertae sedis, and the loss of the flagellum has occurred at least twice independently in Archamoebae. Thus, the name Entamoebae is confusing and should ideally only be used in the future with specific clarification as to its composition, noting that this is different from that of recent years.
History of Genera in the Archamoebae
Mastigamoeba was the first genus created to house species with a flagellum and an amoeboid body, with a hyaline cytoplasm unlike that of other superficially similar taxa such as the cercomonads (Kent 1880; Klebs 1892; Schulze 1875b; Stokes 1886, 1888, 1890). Frenzel (1897) then created Mastigella as a vehicle for species with similar characteristics but no (direct) connection between the nucleus and the flagellum and Mastigina for flagellated species that had a limax shape and few lateral pseudopodia. These three genera have historically been described collectively as “mastigamoebids” (i.e., hyaline amoeboid flagellates), though the composition of this informal group has recently changed (see “General Characteristics”, above). Subsequent revision of Mastigina from Goldschmidt (1907a) added an apical, spherical nucleus as a distinguishing criterion, as well as fountain-flow cytoplasm, though this is not present in all described species of Mastigina (Frenzel 1897) and is present in species of other genera that had already been described at that time, e.g., Pelomyxa palustris, Mastigamoeba aspera, and Tricholimax hylae (Frenzel 1897; Greeff 1874; Leidy 1879; Schulze 1875a, b). This had the effect of narrowing the circumscription of Mastigamoeba to include only hyaline amoeboid flagellates with a connection between the flagellum and the nucleus, with lateral pseudopodia, and with elongated nuclei. Tricholimax hylae (the sole species of Tricholimax) was then also treated as belonging to Mastigina (Brugerolle 1982; Brugerolle and Patterson 2000; Collin 1913; Goldschmidt 1907a; Wickerham and Page 1970), leading to some confusion about the distinguishing criteria of Mastigina (Frolov 2011).
Through the twentieth century, over 200 nominal species of Mastigamoeba and Mastigella were created on the basis of shape and size, pseudopodial form, and contractile vacuole number and location (e.g., Goldschmidt 1907a, Lemmermann 1914; see list of names in Ptáčková et al. 2013). Early studies of life cycles (Goldschmidt 1907a, b) and more recent culture-based studies (Bernard et al. 2000; Chystjakova et al. 2012; Ptáčková et al. 2013; Simpson et al. 1997; Walker et al. 2001; Zadrobílková et al. 2015) have shown that a single individual over time can express a very wide range of size, mode of amoeboid movement, gliding, swimming, and/or pseudopodial form, meaning that many previously described species are difficult to recognize with any confidence. However, recent molecular phylogenetic work has demonstrated that the ephemeral characteristics previously used to distinguish species can be used successfully as taxonomic characters, given a sufficiently detailed description (Ptáčková et al. 2013; Zadrobílková et al. 2015, 2016). Molecular phylogenetics has also shown that there are two main clades within Mastigamoeba, which have different ultrastructural characteristics (Pánek et al. 2016; see below).
What are now the type species of Endolimax and Iodamoeba were described in 1917 and 1912, respectively, as two Entamoeba species from humans (originally Entamoeba nana and Entamoeba buetschlii, respectively). Endolimax was created in 1917 (Kuenen and Swellengrebel 1917), and Iodamoeba in 1919 (Dobell 1919). Species have subsequently been assigned to each genus mostly based on morphology, with some debate about the degree of host specificity leading to fluctuating numbers of nominal species. Recent molecular phylogenetic studies suggest that the diversity encompassed by these descriptions is highly inconsistent from one nominal species to the next. Both genera are in need of revision (Stensvold et al. 2012; Poulsen and Stensvold 2016).
Species description in Pelomyxa has followed a similar pattern to that of Mastigamoeba. The amoeba now known as Pelomyxa palustris was first found by Greeff in 1870 and was named Pelobius (a name already occupied by an insect); it was subsequently redescribed in more detail under its present name, Pelomyxa palustris (Greeff 1874). Greeff recognized that this “new, large freshwater rhizopod” was distinctively different from members of the genus Amoeba and emphasized the great morphological variability of the species. Leidy (1879) was also aware of this variability and suggested that the different forms of Pelomyxa might all be shown later to be different stages in the life cycle of the same species. During the twentieth century, various authors assigned numerous species to Pelomyxa (e.g., Penard 1902), but the prevailing view by the end of the twentieth century was that there was one or a few highly polymorphic species (Whatley and Chapman-Andresen 1990). Subsequently, as studies have employed light and electron microscopy and molecular phylogenetics, the number of species has begun to increase again (Berdieva et al. 2015; Chystjakova and Frolov 2011; Chystjakova et al. 2014; Frolov et al. 2004, 2005a, b, 2006, 2011; Ptáčková et al. 2013). The concept of Pelomyxa as a flagellated amoeba is relatively recent (Griffin 1979, 1988), but its similarities to mastigamoebids have long been noted (Bütschli 1880; Goldschmidt 1907a; Kudo 1939; Page 1970; Penard 1936; Schulze 1875b).
The genus Endamoeba was created in 1879 by Leidy for his newly described species, Endamoeba blattae, from cockroaches. Without noticing this, Casagrandi and Barbagallo (1895) created the genus Entamoeba and transferred into it the human parasitic species known previously as Amoeba coli. Since the names Entamoeba and Endamoeba are very similar, they were often confused, leading to their homonymization and the formal suppression of Entamoeba in 1928. The human pathogen Entamoeba histolytica can be, therefore, found under name Endamoeba histolytica in the older literature. This concept was, however, challenged by many authors (e.g., Kirby 1945) who argued that E. blattae and E. coli were not congeneric. The Entamoeba/Endamoeba problem was settled in 1954 when the International Commission on Zoological Nomenclature refuted the homonymization. Since that time, Entamoeba and Endamoeba have universally been considered distinct genera. The phylogenetic placement of Endamoeba remains unknown: although it is assumed to be related to Entamoeba, the cases of Endolimax and Iodamoeba now being placed in Mastigamoebidae suggest that Endamoeba’s position should be regarded as incertae sedis for now.
Since Entamoeba histolytica is an important human parasite, it is by far the best-known member of the Archamoebae. It was first reported by Lösch in 1875 and has since been intensively studied (see Martínez-Palomo (1993) and Wenyon (1926) for pertinent citations). The research into cell biology and biochemistry of Entamoeba was greatly facilitated when methods of its axenic culture became available (Diamond 1961). Entamoeba histolytica was one of the first putatively amitochondriate eukaryotes whose mitochondrial derivative (mitosome in this case) was discovered (Tovar et al. 1999). The genome sequence of E. histolytica was published more than 10 years ago (Loftus et al. 2005). Genome sequences of several other Entamoeba species are currently available (http://amoebadb.org/amoeba/). Several hundred papers on E. histolytica have been published annually during the last few years.
The genus Rhizomastix was described in 1911 by Alexeieff for his species Rhizomastix gracilis from the intestines of an axolotl. Rhizomastix gracilis was soon found also in larvae of crane flies (Mackinnon 1913), and its morphology was studied in detail by Ludwig (1946). Although a few additional Rhizomastix species have been described from insects and amphibians (see Čepička (2011) and Zadrobílková et al. (2016) for pertinent references), the phylogenetic position of this genus remained unclear until the recent molecular phylogenetic study of Ptáčková et al. (2013), which showed that Rhizomastix belongs to Archamoebae. The diversity of the genus has recently been studied using cultures and molecular phylogenetics (Zadrobílková et al. 2016; Pánek et al. 2016).
Practical Importance
Endobiotic Taxa
Most described endobiotic Archamoebae are poorly known, and their relationships with their hosts have not been determined. Nevertheless, most species are probably harmless commensals.
The situation is much clearer in case of the few human symbionts. Several species have been reported from the intestine of humans (Entamoeba histolytica, E. coli, E. dispar, E. polecki, E. hartmanni, E. moshkovskii, Endolimax nana, and Iodamoeba buetschlii) and oral cavity (Entamoeba gingivalis). Although most of them are considered non-pathogenic, Entamoeba histolytica is an important human pathogen, which is responsible for 500 million new cases and 100,000 deaths annually. The disease caused by E. histolytica is called amebiasis or amebic dysentery and is distributed worldwide, though most cases are reported in developing countries. Amebiasis is also commonly diagnosed in travelers returning from tropical countries. The infection by E. histolytica usually occurs by ingestion of cysts with food or water. The primary site of infection is the large intestine where the amoebae may invade the mucosa and cause colitis that itself may be lethal. Occasionally, the amoebae spread via blood to various internal organs (usually the liver), and extraintestinal amebiasis (with significant mortality) develops. For more information on amebiasis, see Martínez-Palomo (1993) and Haque et al. (2003).
Entamoeba dispar is morphologically indistinguishable from E. histolytica and was historically considered its non-pathogenic variant. It has been, however, shown that the two organisms represent separate species (see Diamond and Clark 1993). The pathogenity of E. moshkovskii is currently under debate (Heredia et al. 2012). Entamoeba invadens is known to cause severe reptilian amebiasis (see Reavill and Schmidt 2010).
Endolimax has been considered to be pathogenic by some authors, possibly causing diarrhea, intestinal inflammation, polyarthritis, or urticaria, but evidence for this is inconclusive (Poulsem and Stensvold 2016).
Free-Living Taxa
Pelobionts and rhizomastixids are of unknown practical importance. While they have a worldwide distribution, they are not known to contribute to any specific agricultural or pathogenic processes. They have historically been regarded as extremely “primitive” or deep-branching eukaryotes but are no longer regarded as such (see “Evolutionary History”, below).
Habitats and Ecology
Occurrence/Distribution in Nature
Free-living pelobionts and rhizomastixids are found in the upper layers of mud or sand, mostly in freshwater rivers, lakes, bogs, or pits, and usually in stagnant or near-stagnant water, which creates a low-oxygen environment. The ideal place to find them is among undisturbed algal growth at the water-sediment interface, in shallow low-oxygen ponds (about 10–20% atmospheric oxygen) with relatively low pH (Bernard et al. 2000; Chystjakova et al. 2012; Frolov 2011; Ptáčková et al. 2013; Whatley and Chapman-Andresen 1990). Several papers have described them from sewage treatment works (e.g., Lackey 1923, 1932). Some taxa have been also described from low-oxygen intertidal marine sediments (Bernard et al. 2000; Bovee and Sawyer 1979; Larsen and Patterson 1990; Lee and Patterson 2000; Page 1983; Simpson et al. 1997; Shiratori and Ishida 2016; Zadrobílková et al. 2015).
Pelobionts have been described from all over the world (e.g., Larsen and Patterson 1990; Lee and Patterson 2000). The majority of reports in the literature are from temperate Europe and North America, due to the intensity of study in these locations in the last 200 years.
Endobiotic Archamoebae are widely distributed among various vertebrates and invertebrates. However, host specificity of particular species is only poorly understood and has been partially elucidated only in human parasites. For example, humans seem to be the primary host of Entamoeba histolytica though it has also been isolated from nonhuman primates and dogs. Similarly, Entamoeba coli, another species from the intestines of humans, can infect also nonhuman primates, dogs, and marsupials (see Thompson and Smith 2011). Entamoeba moshkovskii is most probably both endobiotic and free-living (Heredia et al. 2012; our observations).
Characterization and Recognition: Light-Microscopical Features
Recognizing Archamoebae
Most of the nominal species among Archamoebae have been described from a few cells seen on a single occasion (e.g., Calaway and Lackey 1962; Lackey 1923; Larsen and Patterson 1990; Penard 1902, 1909; Stokes 1886, 1888, 1890; Skvortzkov and Noda 1976). Many species can exist as flagellates, uninucleate amoebae, multinucleate amoebae, and cysts, a situation referred to as polymorphism. Also, individual amoebae and flagellates change their appearance and are therefore pleomorphic. As most well-studied pelobionts are both polymorphic and pleomorphic, it would be tempting to assume that only static taxonomic characters that transcend any ephemeral changes can be used to distinguish species, such as the spines on the outside of Mastigina setosa (Goldschmidt 1907a), the extranucleolar “dot” in Mastigamoeba punctachora (Walker et al. 2001), or the doubled nucleus in Mastigamoeba schizophrenia (Simpson et al. 1997). But recent molecular work combined with observations of cultures (Ptáčková et al. 2013; Zadrobílková et al. 2015, 2016) confirms that – while transcendent characters make distinctions easier – ephemeral characters such as pseudopodial shape do correlate with molecular phylogenetic differences. Under these circumstances, it is important to not only observe variation in size and shape, flagellar length, speed of swimming, cytoplasmic movement, or separation into inner and outer layers but also to observe what percentage of the time characters are expressed.
General Appearance of the Cell
Archamoebae are chiefly recognized by being obviously amoeboid or amoeboid flagellates but are differentiable from other, similar taxa (e.g., myxomycete or protostelid swarmers, breviates, cercozoans, lobose amoebae) by having hyaline (clear) cytoplasm, which gives rise to “eruptive” or “bulging” hemispherical pseudopodia, usually at the anterior end of a moving cell (Fig. 1). The range of pseudopodial shapes reported in the literature also includes other morphologies – lateral or trailing pseudopodia may be rarely fine and filose or, more often, tapering finger shaped, broadly conical, or broadly lobate (e.g., Chystjakova et al. 2012; Frolov 2011; Frolov et al. 2004, 2006; Ptáčková et al. 2013; Zadrobílková et al. 2015, 2016). A uroid is frequently present at the posterior end of the cell; though in some taxa, particularly Mastigamoeba simplex and other members of the “Mastigamoebidae B” clade, and some Mastigina species, trailing pseudopodia are present instead (e.g., Goldschmidt 1907a; Pánek et al. 2016; Walker et al. 2001). Rhizomastix displays various pseudopodial shapes (Čepička 2011; Ptáčková et al. 2013; Zadrobílková et al. 2016). Pelomyxa has larger cells, and short flagella, but has many of the amoeboid characteristics seen in Mastigamoeba, as do members of Mastigella (Zadrobílková et al. 2015). Cells of Entamoeba spp. move by eruptive, clear lobopodia, though trailing filaments may be rarely formed. Although Endolimax is superficially similar to Entamoeba, its cells often form short tapering pseudopodia, similar to those seen in some species of Mastigamoeba (see Figs. 1, 3, and 4).
In some archamoebae, the cytoplasm is arranged in a distinct separation between “endoplasm” (interior of the cell body, where organelles are located) and the “ectoplasm” – a clear peripheral layer of cytoplasm, directly underneath the plasma membrane, which remains distinct from the agglomeration of nuclei, vacuoles, and endomembrane system in the center of the cell. This is the layer from which eruptive pseudopodia form, into which the central cell contents then roll. The differentiation of ectoplasm has been described in mastigamoebids, including Mastigamoeba scholaia (Klug 1936), Mastigella nitens (Penard 1909), and Mastigina setosa (Goldschmidt 1907a; Skibbe and Zölffel 1991).
The ectoplasm may also be involved in “fountain flow” cytoplasmic streaming, as the outer part of the cytoplasm that runs backward, relative to the forward motion of the middle of the cell. Cytoplasmic streaming has mostly been described in Pelomyxa (Blochmann 1894; Hollande 1945; Rhumbler 1905; inter alia) but has also been reported in Mastigamoeba aspera (Chystjakova et al. 2012; Schulze 1875b), Tricholimax hylae (Becker 1928; Brugerolle 1982; Collin 1913; Frenzel 1897), and Mastigina setosa (Goldschmidt 1907a; Skibbe and Zölffel 1991).
Polymorphism and Life Cycle
Polymorphism is well-documented and a defining feature in pelobiont Archamoebae, as well as being necessary for completion of the life cycle in parasitic entamoebae. Life cycles have also been studied in several species. However, the actual cytological processes accompanying changes from one form to another remain mostly unknown, except in Entamoeba (Frolov 2011).
Recent descriptions of members of Mastigamoeba, Rhizomastix, and Mastigella have usually employed laboratory culture conditions that do not permit a study of the life cycle under normal environmental conditions. The general picture is that mastigamoebid Archamoebae can produce some or all of the following stages: amoeboid flagellates, aflagellate amoebae, large multinucleate amoebae, and cysts. All of these forms have been documented in detailed studies of Mastigamoeba balamuthi (Chavez et al. 1986; Pánek et al. 2016) and M. schizophrenia (Simpson et al. 1997), while flagellates and amoebae have been documented in M. aspera (Chystjakova et al. 2012), M. simplex (Bernard et al. 2000), M. punctachora (Bernard et al. 2000), M. lenta, M. abducta, M. guttula, M. errans, and Rhizomastix libera (Ptáčková et al. 2013); Mastigella erinacea and Mastigella rubiformis (Zadrobílková et al. 2015); flagellates and cysts in Mastigella nitens (Frolov 2011), Rhizomastix bicoronata, R. tipulae, and R. elongata (Zadrobílková et al. 2016); and flagellates, amoebae, and cysts in R. vacuolata (Zadrobílková et al. 2016).
Earlier studies of members of Mastigamoeba and Mastigella (e.g., Frenzel 1897; Goldschmidt 1907a; Schulze 1875b) employed microcosms rather than typical modern laboratory culture conditions, mimicking normal environmental conditions and allowing some observations of succession of polymorphism, from which life cycle details could be inferred. Flagellates reproducing by mitosis, aflagellate amoebae, and large, multinucleate division stages giving rise to uninucleate cells were documented in Mastigella vitrea and Mastigina setosa (Goldschmidt 1907a, b).
A series of studies of Pelomyxa species has also employed microcosms as a culture method, permitting extensive observation of the life cycle of natural populations (Whatley and Chapman-Andresen 1990; Frolov 2011). Pelomyxa palustris progresses in spring from cysts containing four nuclei, to small binucleate amoebae (100 μm long), then to larger multinucleate amoebae with or without endosymbionts, and then in summer to large elongate multinucleate amoebae (up to 5 mm long) with 30–60 nuclei, endosymbionts, flagella, and a posterior uroid. In autumn, they progress to large, spherical amoeboid cells with bacteria around the thousands of nuclei; these giant cells then divide by plasmotomy to form cysts or small amoebae over winter (Frolov 2011; Frolov et al. 2007; Schirch 1914; Whatley and Chapman-Andresen 1990). Other life-cycle descriptions exist for P. corona (Frolov et al. 2004), P. gruberi (Frolov et al. 2006), P. binucleata (Frolov et al. 2005a; Frolov 2011), P. flava (Frolov et al. 2011), and P. paradoxa (Chystjakova et al. 2014).
Cysts
The structure of the cyst has been best characterized in Entamoeba invadens (see Samuelson and Robbins 2011). The cyst wall is composed mainly from chitin and various glycoproteins. Cysts of particular Entamoeba species are rounded and differ in the number of nuclei present, e.g., the uninucleate E. polecki; E. histolytica, E. hartmanni, and E. marina with four nuclei; and E. coli and E. muris with up to eight nuclei (recently summarized in Shiratori and Ishida 2016). During the encystation of Entamoeba trophozoites, so-called chromatoid bodies are formed in the cytoplasm. These are elongated bars with rounded ends and are formed by ribosomes arranged in helices. The chromatoid bodies disappear from the older cysts (Rosenbaum and Wittner 1970; Schuster 1990).
Apart from Entamoeba spp., cysts with various numbers of nuclei have been documented in some other members of Archamoebae: uninucleate cysts in Mastigamoeba balamuthi (Chavez et al. 1986), M. punctachora (Bernard et al. 2000), M. schizophrenia (Simpson et al. 1997), and Iodamoeba buetschlii, binucleate cysts in Rhizomastix spp. (Čepička 2011; Zadrobílková et al. 2016), and four-nucleated cysts in Endolimax spp. and Pelomyxa palustris (Frolov 2011; Frolov et al. 2007; Griffin 1988; Ptáčková et al. 2013; Whatley and Chapman-Andresen 1990). The cysts of Endamoeba are multinucleate. Tricholimax varies between two and four nuclei according to the age of the cyst (Collin 1913). The composition of the cyst wall of non-Entamoeba Archamoebae is still unknown. In Pelomyxa, the cyst has a central vacuole containing endosymbionts, and the cyst wall has three layers. The laminated structure of cyst walls would be better revealed by cryofixation techniques, which have not yet been employed, than by chemical fixation, which has been used hitherto except on cysts of Rhizomastix (Zadrobílková et al. 2016). The cyst of Iodamoeba buetschlii contains a characteristic glycogen-containing inclusion in the cytoplasm (see Fig. 5).
Ultrastructure
Flagellar Cytoskeleton
Flagella in Archamoebae appear to have degenerated in several groups independently, either to structures with dysfunctional axonemes or to the complete absence of a flagellar apparatus (discussed in Zadrobílková et al. 2015). Members of Entamoeba, Endamoeba, Endolimax, and Iodamoeba have no flagellar apparatus (El-Hashimi and Pitman 1970; Morris 1936; Rosenbaum and Wittner 1970; Zaman et al. 1998, 2000) and are not referred to further in the sections below.
Flagellated Archamoebae have a relatively simple microtubular cytoskeleton (see Fig. 2). Among the flagellated taxa that have been studied by TEM (see Table 1), the flagellar apparatus consists of one or more “monokinetids” – single, flagellated basal bodies – giving rise proximally to a cone or cylinder of microtubules, as well as (in most taxa) a lateral root of microtubules with a bilaminar “root sheet” at the origin of the root on its distal face. Members of Mastigamoeba, Mastigella, Mastigina, Tricholimax, and Rhizomastix typically have only one flagellum per cell, though the type species of Mastigella, M. polymastix, has up to four flagella per cell (Frenzel 1897) and Tricholimax has multiple nuclei, each with its own attached (internal) kinetid (Becker 1925; Brugerolle 1982, 1991; Collin 1913). The flagellar apparatus in Mastigamoeba varies between a wide distribution of microtubules, seen in Mastigamoebidae “A” (as defined in Ptáčková et al. 2013, Pánek et al. 2016; see also Table 1 and Figs. 2a and 3), and a very narrow single layer of microtubules, seen in members of Mastigamoebidae “B” (as defined in Ptáčková et al. 2013; Pánek et al. 2016; see also Table 1 and Figs. 2b and 3).

Flagellar apparatus ultrastructure defines the identities of genera of flagellated Archamoebae. The features common to all flagellated Archamoebae include a single basal body (Bb), from which arises a flagellum (Fl) and a cone (MC) or rhizostyle (Rh) of microtubules, and (in most taxa) a transitional cylinder (TC). In most cells, the flagellar apparatus is usually directed perpendicularly to the cell membrane (CM), but the cone of microtubules can sometimes run parallel to it. (a) Mastigamoeba has a microtubular cone (MC) that connects to the nucleus; a microtubular root (MR) also arises from the basal body, immediately proximal to a bilaminate root sheet (RS); typically there is only one monokinetid flagellum per cell. This shows a representative flagellar apparatus from the group “Mastigamoebidae A” where the cone of microtubules (MC) arises laterally from both the sides and the base of the basal body; the flagellar transition zone is long. Fl flagellar axoneme, TC transition zone cylinder, Bb basal body. RS bilaminar root sheet, MR microtubular root. (b) Mastigamoeba showing a representative of group “B” where the cone of microtubules (MC) arises longitudinally from near the base of the basal body, and the flagellar transition zone (TZ) is short and contains no extra elements. (c) Mastigella has a microtubular cone that does not connect to the nucleus. There may be up to four long flagella per cell in some species. (d) Pelomyxa has numerous nonfunctional, short, monokinetid flagella (nFl) with “n + n” arrangement of microtubules. There is no obvious microtubular root or root sheet. (e) Tricholimax has a flagellar apparatus similar to that of Mastigamoeba, but it has nonfunctional “n + n” flagella (nFl), and it frequently has several nonflagellated monokinetids connected to nuclei in the cytoplasm. (f) Rhizomastix has a rhizostyle (Rh) – a tapering bundle of microtubules that extends from the base of the basal body, to wrap around the nucleus. Its flagellum has two vanes, which may contribute to its fast, jerky swimming movement. Scale bar = 250 nm (Figures (a) and (c) redrawn with modifications from Walker et al. 2001, and Figure (b) reproduced from Walker et al. 2001 with permission from Elsevier)

Mastigamoeba. All images are differential interference contrast light microscopy (DIC), unless otherwise specified. (a–o): Representatives of Mastigamoebidae A. (a–c) Mastigamoeba punctachora, G = extranucleolar granule. (c) Bright field microscopy with protargol staining; B = basal body, immediately adjacent to the stained nucleus. (d) Mastigamoeba balamuthi swimming cell, phase contrast LM. (e–i) Mastigamoeba errans. In e, gliding flagellate cell; f–i, aflagellate cells. (j–m) Mastigamoeba sp. ex Pelomyxa belevskii. F = flagellum. In j, bright field light microscopy. (n) M. punctachora transmission electron microscopy. Double-membraned, mitochondrion-related organelle (Mi). (o) M. punctachora TEM. Nucleus (N) surrounded by microtubules of the cone (MC) cut in transverse section. (p–aa): Representatives of Mastigamoebidae B. (p–s) Mastigamoeba simplex. In p–r, gliding flagellates; in s, aflagellate cell. (t–w) Mastigamoeba scholaia. In t, u, gliding cells; in v, gliding aflagellate cell; in w, aflagellate cell. N = neck. (x–aa) Mastigamoeba guttula. In x–z, flagellates; in aa, aflagellate cell. Scale bars in a–c, e–i, p–aa =10 μm; in d = 20 μm, in j–m = 50 μm, in n, o = 200 nm (Figures a–c, e–m, and p–aa all reproduced from Ptáčková et al. 2013 with permission from Elsevier)
Flagella are usually positioned at the anterior end of the cell and used in movement, though particularly amoeboid cells in Mastigella may move predominantly by pseudopodia, with the poorly functional flagellum pointing in a different direction (Frenzel 1897; Frolov 2011; Goldschmidt 1907a; Walker et al. 2001, Zadrobílková et al. 2015). The flagellum of Mastigella is generally slower beating and less motile than that seen in Mastigamoeba and may be an intermediate stage that gave rise to the extreme flagellar reduction seen in Pelomyxa (discussed in Zadrobílková et al. 2015).
Giant amoeboid cells of Pelomyxa have numerous separate monokinetids, bearing (usually) short, nonfunctional flagella with a disorganized axoneme. Typically, a microtubular cone is formed, but this varies between a dense cone and a single layer of microtubules, with or without a lateral root (Fig. 2d; see Table 1 for further details). The number of monokinetids varies between ca. 50 and ca. 700 in P. palustris (Griffin 1988). Likewise, their density varies more than tenfold (Griffin 1988), and their abundance and distribution over the cell’s sides or uroid also varies (e.g., compare P. flava, P. paradoxa, and P. stagnalis; Chystjakova and Frolov 2011; Chystjakova et al. 2014; Frolov et al. 2011). Whether basal bodies form flagella or merely “buds” can vary widely through the life cycle (e.g., P. paradoxa vs. P. gruberi; Chystjakova et al. 2014; Frolov et al. 2006). Flagella are typically not found on anterior pseudopodia in Pelomyxa (Frolov 2011) and have not been seen at all in P. secunda (Berdieva et al. 2015).
For further details of the flagellar cytoskeleton, see Fig. 2 and Table 1.
Nuclei and Cell Division
Numbers of nuclei are one of the main features that distinguish “pelomyxid” Archamoebae from “mastigamoebid” or “entamoebid” Archamoebae. In many pelomyxids, the usual state is cells with tens or hundreds of nuclei, whereas in other Archamoebae, the usual state is one or few nuclei, though stages with tens of nuclei may exist.
A uninucleate flagellate is the main trophic form in Mastigamoeba, some members of Mastigella, Mastigina, and Rhizomastix. Reproduction of uninucleate flagellates by mitosis has been documented only in Mastigella and Mastigina (Goldschmidt 1907a, b). The nuclei of Mastigamoeba schizophrenia appear in “pairs” of two adpressed nuclei, but the extent of differentiation within these pairs is unknown (Simpson et al. 1997). Some “flagellate” taxa may typically have more than one nucleus, such as Tricholimax hylae, which has 1–6 (Becker 1925; Brugerolle 1982, 1991; Collin 1913; Frenzel 1897), and members of Mastigella (Zadrobílková et al. 2015).
As discussed above, a few “flagellate” species also have distinct, large amoeba stages with more than one nucleus: Mastigamoeba schizophrenia has 2–10 (i.e., 1–5 pairs; Simpson et al. 1997); M. punctachora has 1–8 (Bernard et al. 2000); and M. aspera has two or more nuclei in its amoeba form, which would previously have been described as a different species, Dinamoeba mirabilis (see Chystjakova et al. 2012). In at least one such species, Mastigamoeba balamuthi, the multinucleate amoebae (with on average 15 nuclei but up to 46), are the dominant trophic stage, reproducing while multinucleate by mitosis and subsequent plasmotomy (i.e., cytokinesis not preceded by mitosis), which results in large and asymmetric numbers of nuclei in daughter cells.
“Budding” plasmotomy, from multinucleate amoebae to uninucleate flagellates, only happens rarely and only under specific trophic conditions (Chavez et al. 1986). Production of small uninucleate flagellates, from a large, rounded amoeboid cell that is filled with smaller cells, has also been described from Mastigina setosa and Mastigella vitrea (Goldschmidt 1907a, b); whether this is a form of plasmotomy or more akin to sporogony is unclear. It resembles the processes described in Pelomyxa palustris (Frolov 2011; Whatley and Chapman-Andresen 1990).
In Pelomyxa, the main described trophic “adult” form is a large, multinucleate amoeba, which also has flagella (see above). This “flagellated” trophic form, typically a cylindrical amoeba, may in some cases later transform into an immotile, rounded amoeboid form with hundreds of nuclei (e.g., P. corona, P. gruberi; Frolov 2011; Frolov et al. 2004, 2006). It is unclear whether this rounded form is homologous to the large multinucleate amoebae seen in Mastigamoeba schizophrenia (Simpson et al. 1997) and M. balamuthi (Pánek et al. 2016) or to the large rounded forms that divide by plasmotomy in Mastigella vitrea and Mastigina setosa (Goldschmidt 1907a), as they are only rarely seen and have not been studied by electron microscopy. Species of Pelomyxa can be divided into taxa that have hundreds of nuclei in the adult motile form (e.g., P. palustris, P. corona, P. prima: Frolov 2011; Whatley and Chapman-Andresen 1990) and taxa with tens (e.g., P. belevskii, P. stagnalis; Frolov 2011; Ptáčková et al. 2013) or one or two nuclei in mature cells (e.g., P. binucleata, P. flava, P. paradoxa, P. schiedti; Chystjakova et al. 2014; Frolov 2011; Frolov et al. 2005a; Zadrobílková et al. 2015). Division from multinucleate adult cells to smaller juvenile cells is typically by unequal plasmotomy (Frolov 2011; Ptáčková et al. 2013; Schulze 1875a; Whatley and Chapman-Andresen 1990).
The trophozoites of Entamoeba spp. are uninucleate, and no multinucleate trophic forms are usually created. Mitosis of Entamoeba histolytica was studied, for example, by Solis and Barrios (1991) and by Chávez-Munguía et al. (2006).
Nuclear Chromatin
Patterns of nuclear chromatin appear to distinguish different species in Archamoebae, though there is not a clear taxonomic pattern above species level. Chromatin patterns can be a peripheral arrangement of small clumps of chromatin around the edge of the nucleus or a single large nucleolus or a few chromatin clumps joined together in the center of the nucleus (described further in Frolov 2011). Peripheral arrangements are common in Entamoeba, pelomyxids, and Rhizomastix (Čepička 2011; Chystjakova et al. 2014; Frolov 2011; Frolov et al. 2004, 2005a, b, 2006; Ptáčková et al. 2013; Zadrobílková et al. 2015, 2016) and are also present in “Mastigina trichophora,” Mastigella eilhardi, and Mastigella nitens (Frolov 2011; Penard 1909; Zadrobílková et al. 2015).
Nuclear chromatin is used as a diagnostic feature in Entamoeba species, with E. histolytica, E. dispar, and E. marina showing a fine peripheral ring with a small central dot, E. hartmanni showing a large central nucleolus and thick peripheral clumps of chromatin, and E. coli showing peripheral clumps only (Shiratori and Ishida 2016). The majority of members of Mastigamoeba have large nucleoli, as do Tricholimax hylae, Pelomyxa palustris, P. gruberi, P. stagnalis, and Rhizomastix species. In both Iodamoeba and Endamoeba, the nucleolus is large and central, and there is no peripheral ring of chromatin (see section “Systematics and Taxonomy”, below).
There are cajal-body-like stacked inclusions in the central nucleolus in Pelomyxa stagnalis (Chystjakova and Frolov 2011; Ptáčková et al. 2013), and P. belevskii has a loose stacked arrangement of “vermiform bodies” of chromatin around the edge of the nucleus, similar in size to the inclusion in P. stagnalis (Ptáčková et al. 2013).
Cell Coverings
As reviewed in Frolov (2011), most pelobionts are covered in a thin layer of amorphous or filamentous organic material. The extracellular layer may be irregularly, thinly distributed over the cell surface (e.g., Brugerolle 1982; Chavez et al. 1986; Simpson et al. 1997; Walker et al. 2001), often being thickest at the posterior end of the cell, particularly in pelomyxids (Frolov 2011). Finely filamentous cell coverings may be present, with filaments apparently perpendicular to the cell surface (e.g., Pelomyxa palustris, P. gruberi, Mastigella nitens: Frolov 2011) or parallel to the surface in the case of very thick coverings, which can reach up to 1 μm thickness (e.g., P. flava, Mastigamoeba aspera; Chystjakova et al. 2012; Frolov 2011; Frolov et al. 2011). These fine extracellular layers are rarely preserved well with chemical fixation for electron microscopy; cryofixation would be appropriate for further study.
Numerous species in pelobionts have been described with spines on the outside of the cell, with varying degrees of packing, from the dense, highly refringent layer of very tightly packed spines in Mastigina chlamys (Frenzel 1897; Seckt 1922), through regularly arranged but obviously discrete spines in Mastigina setosa (Goldschmidt 1907a; Skibbe and Zölffel 1991), to irregular and loosely packed spicules in Mastigamoeba trichophora (Lauterborn 1901), Mastigamoeba pilosa (Schoudeten 1907), and Mastigina spicata (Penard 1909). Frolov (2011) shows electron micrographs of “Mastigina trichophora” where hollow spicules are each placed in “boat-shaped” scales, which are packed in a tight layer over the extracellular glycocalyx. It is unclear whether these spines are made only of organic material (Frolov 2011) or are mineralized as well, as seen in the euglenid Trachelomonas (Conforti et al. 1994; Preisig 1994).
Mitochondrial Remnants
Like other anaerobic groups in the eukaryotes, Archamoebae are “secondarily amitochondriate.” In almost all cases, such anaerobic eukaryotes retain reduced mitochondrion-related organelles (MROs) which have usually lost DNA, genome maintenance proteins, and a typical ATP-producing electron transport chain. They often still produce ATP by partial anaerobic oxidation of pyruvate (hydrogenosomes), but some have lost energy metabolic function entirely (mitosomes), preserving only a few other functions (such as aspects of mitochondrial type iron-sulfur cluster assembly machinery) and acquiring the ability to import ATP (Chan et al. 2005; Maguire and Richards 2014).
Both kinds of double-membrane-bound MROs have been described from Archamoebae (Barberà et al. 2007; Hampl and Simpson 2008). Mitochondrial cpn60 function has been demonstrated in Entamoeba histolytica, and localized, to the mitosomes in E. histolytica (Chan et al. 2005; Clark and Roger 1995; Leon-Avila and Tovar 2004; Tovar et al. 1999), and to hydrogenosome-like organelles in Mastigamoeba balamuthi (Gill et al. 2007; Hampl and Simpson 2008). Iron-sulfur cluster assembly proteins are closely related to those of epsilon proteobacteria both in Mastigamoeba balamuthi (Gill et al. 2007) and Entamoeba histolytica (Maralikova et al. 2010; van der Giezen et al. 2005), implying a single lateral transfer event where mitochondrial Fe:S assembly appears to have been lost entirely and replaced with an Fe:S assembly system that is not homologous to the standard mitochondrial system and is now cytosolic (Nývltová et al. 2013). The acquisition of alternatives to normal mitochondrial pyruvate dehydrogenase may have happened more than once, with M balamuthi and E. histolytica each possessing copies of pyruvate ferredoxin/oxidoreductase (Embley and Martin 2006; Gill et al. 2007); but M. balamuthi also possesses a pyruvate formate lyase that was laterally transferred from a firmicute (Stairs et al. 2011). Mitosomes are extremely abundant in E. histolytica implying that some functions are yet to be uncovered in detail (Aguilera et al. 2008) – an example being the synthesis of sulfur compounds required for encystation (Mi-ichi et al. 2015). The MROs of Archamoebae are discussed in the context of other MROs by Maguire and Richards (2014).
Ultrastructural reports also exist for double-membrane-bound organelles in Mastigamoeba simplex, Mastigamoeba punctachora, and Mastigella commutans (Walker et al. 2001), Mastigella rubiformis, M. ineffigiata (Zadrobílková et al. 2015), Endolimax piscium (Constenla et al. 2013), Rhizomastix libera (Ptáčková et al. 2013), R. elongata (Zadrobílková et al. 2016), and Pelomyxa palustris (Seravin and Goodkov 1987c). As previously discussed (Zadrobílková et al. 2015), there is a notable lack of reports of MROs in EM studies of Pelomyxa. This may be because of the difficulty of serially sectioning a large amoeba, a misidentification of MROs as symbionts, or, alternatively, the real absence of MROs in all examined species of Pelomyxa except P. palustris.
Golgi
Canonical Golgi dictyosomes have not been reported in the Archamoebae so far, though related elements of the endomembrane system have been shown to be functionally present in Entamoeba histolytica (Bredeston et al. 2005) and Mastigamoeba balamuthi (Dacks et al. 2004), with all “core” functions being present (Mowbrey and Dacks 2009). However, methods for detecting endomembrane system function and Golgi dictyosomes are diverse, and many eukaryotes do not possess canonical Golgi stacks that are easy to recognize by transmission electron microscopy. Tantalizing reports exist suggesting that Entamoeba may possess Golgi stacks that are only visible with the application of cryofixation techniques (Chavez-Munguia et al. 2000; Ghosh et al. 2000). A similar situation may hold in pelobionts: a few reports exist of Golgi-like stacks of membranes, but in each case, these may be poorly fixed endoplasmic reticulum, and further ultrastructural work, using different techniques, is required. The multi-membrane organelle seen in Rhizomastix libera (Ptáčková et al. 2013) is positioned close to the flagellar base, as per the Golgi dictyosomes of other superficially similar amoeboid taxa with flagella (e.g., Heiss et al. 2013; Walker et al. 2003), but the published preservation of Rhizomastix is not sufficiently good for any clear identification of the organelle, and it is not seen in R. elongata under better fixation conditions (Zadrobílková et al. 2016). Similar, though bigger and more organized stacked structures have been reported from Pelomyxa palustris (Seravin and Goodkov 1987b) and P. corona (Frolov et al. 2004) as well as reticulate multimembranous organelles from P. flava (Frolov et al. 2011). These have subsequently been interpreted as endoplasmic reticulum (Frolov 2011), as have reports of stacked structures in Mastigamoeba schizophrenia (Simpson et al. 1997), M. punctachora (Walker et al. 2001), and Rhizomastix elongata (Zadrobílková et al. 2016).
Peroxisomes
While Archamoebae have traditionally been considered to lack peroxisomes (Žárský and Tachezy 2015), an unpublished thesis reports peroxisomal proteins in both Mastigamoeba balamuthi and Entamoeba histolytica (Žárský 2012), which would be consistent with secondary loss of peroxisomal function in Archamoebae.
Symbionts
The best-known symbionts of Archamoebae are the three morphologies of prokaryotes that surround the nuclei of Pelomyxa species (Berdieva et al. 2015; Chystjakova et al. 2014; Frolov 2011; Frolov et al. 2006; Whatley 1976; Whatley and Chapman-Andresen 1990). There is one large bacterium with a distinctive axial cleft and up to two smaller methanogenic archaea, one of which is known as Methanobacterium formicicum DSM3637 (Frolov et al. 2004, 2005a, b, 2006, 2011; Griffin 1988; van Bruggen et al. 1988; Whatley 1976). Methanobacterium formicicum DSM3637 was isolated from P. palustris (van Bruggen et al. 1983, 1988), and a draft of its genome has been published (Gutierrez 2012). Endobiotic prokaryotes also exist in Mastigella species (Frolov 2011; van Bruggen et al. 1985; Walker et al. 2001; Zadrobílková et al. 2015) and Rhizomastix libera (Ptáčková et al. 2013). The organic layer covering Mastigamoeba aspera also contains numerous small, rod-shaped prokaryotic ectobionts of uncertain identity (Chystjakova et al. 2012; Goldschmidt 1907a; Kudo 1950; Lemmermann 1914; Page 1970; Penard 1902, 1909, 1936; Schulze 1875b; Siemensma 1987).
The physiological role played by these endo- and ecto-symbionts has not been clearly demonstrated, but numerous cases exist of symbiosis between methanogenic archaea (which presumably derive hydrogen from within their hosts) and anaerobic eukaryotes with mitochondria that have been reduced to hydrogenosomes (Embley et al. 1995; Fenchel and Finlay 1995; Martin and Müller 1998; van Bruggen et al. 1983; van Hoek et al. 2000).
Endobiotic mastigamoebids have also been described as emerging from Pelomyxa (Greeff 1874; Hollande 1945; Whatley and Chapman-Andresen 1990); recent observations including sequence data confirm that this is not a life-cycle stage of Pelomyxa (Ptáčková et al. 2013).
Systematics and Taxonomy
-
Class Archamoebae Cavalier-Smith, 1983
Anaerobic/microaerophilic Amoebozoa with reduced mitochondria. May exist as amoebae, amoeboflagellates, or cysts. Ancestrally with a single apical flagellum, arising from a single basal body that gives rise proximally to a microtubular cone and laterally to a microtubular root arising immediately proximal to a double-layered “root sheet” sitting perpendicular to the axis of the basal body. Secondarily aflagellate or multiflagellate, with disordered flagellar axoneme, or with no flagellar apparatus. Amoeboid movement with eruptive lobopodia. Free-living or endobiotic.
-
Order Pelobiontida Page, 1976
For the current composition, see Pánek et al. (2016). The clade including mastigamoebids, pelomyxids, and rhizomastixids; using the branch-based definition introduced and discussed by Pánek et al. (2016), the clade consisting of Mastigella eilhardii Bürger 1905 and all organisms that share a more recent common ancestor with M. eilhardii than with Entamoeba histolytica Schaudinn 1903.
-
Suborder Mastigamoebina Frenzel, 1897
Rank changed from order to suborder by Pánek et al. (2016). The clade including Mastigamoebidae and Rhizomastixidae; using the node-based definition introduced and discussed by Pánek et al. (2016), the least-inclusive clade consisting of Mastigamoeba balamuthi (Chávez et al. 1986) Simpson et al. 1997, Mastigamoeba abducta Ptáčková et al. 2013, and Rhizomastix libera Ptáčková et al. 2013.
-
Family Mastigamoebidae Goldschmidt, 1907
Archamoebae with trophozoites which are uninucleate to multinucleate, with single motile anterior flagellum associated with microtubular cone, or aflagellate. Amoebae flattened, amoeboid movement slow, typically with multiple pseudopodia. Free-living or endobiotic. Current composition as per Ptáčková et al. (2013).
-
Mastigamoeba Schulze, 1875 (Fig. 3)
Mastigamoeba contains amoeboid cells where the flagellated basal body and the anterior nucleus are immediately adjacent to each other during movement. The basal body and nucleus are joined by a cone of microtubules.
Circumscription
Archamoebae with a uniflagellated trophic stage, in which the nucleus and flagellum are connected by a cone of microtubules that arises from the base and sides of the single (flagellated) basal body; a cylinder is present in the transition zone of the flagellum. A single root of microtubules arises from the side of the basal body, and the root has a bilaminar “root sheet” on its anterior edge (Fig. 2a, b). Basal bodies usually have nine triplets of microtubules, but M. schizophrenia has nine doublets (Simpson et al. 1997). The flagellum has a conventional eukaryotic “9 + 2” arrangement of microtubules but lacks outer dynein arms, giving rise to a distinctively languid flagellar beat. The flagellates may, at least in some species, transform to amoebae with one, few, or many nuclei. Both flagellate and amoeboid forms may transform into cysts. Nuclei are usually single, but are paired in M. schizophrenia, and have vesicular nucleoli. In M. punctachora, the nucleus contains a small extranucleolar “dot” of chromatin (Bernard et al. 2000). The outside of the cell is usually naked, but in M. aspera, there are ectobiotic bacteria (Chystjakova et al. 2012) and spined species have been described. Cells have been found in soils and freshwater and marine habitats. Phylogenetic analyses divide this genus into two separate groups (as defined in Ptáčková et al. 2013; Pánek et al. 2016): “A” with a broad multilayered microtubular cone radiating laterally from the sides of the basal body, a long flagellar transition zone, and, in some taxa, a transitional column or spiral, microtubules arising from the base of the basal body or an MTOC immediately proximal to the basal body (Fig. 2a; Table 1) and “B” with a single-layered microtubular cone extending posteriorly from sides the basal body, and a short transition zone, and none of the extra features seen in some members of “A (Fig. 2b; Table 1).” There is little morphological variation at the light microscopical level in “B” (Fig. 3).
Type Species
Mastigamoeba aspera Schulze, 1875
(See Chystjakova et al. 2012 for a recent description and discussion of synonymy with Dinamoeba; discussed further in Ptáčková et al. 2013)
Remarks
Based on recent phylogenetic analyses (Stensvold et al. 2012; Ptáčková et al. 2013), the flagellum-lacking entamoebid genera Endolimax and Iodamoeba form an internal branch of Mastigamoeba, making the latter genus paraphyletic. Recent analyses also divide Mastigamoeba into “A” and “B” clades, as discussed above. The full scope and character of Mastigamoeba is uncertain, as recent phylogenetic analyses do not include most of the previously described taxa in Mastigamoeba, and we lack electron microscopical data for many taxa, as well as molecular data on the phylogenetic position of M. aspera, the type species. The degree of overlap between Mastigamoeba, Mastigina, and Tricholimax has long been unclear, leading to confused and inconsistent taxonomy of some species (Frenzel 1897; Frolov 2011; Goldschmidt 1907a; Lemmermann 1914). Other genera that fall within the circumscription of Mastigamoeba include Dinamoeba Leidy, 1874 (D. mirabilis has long been regarded as a synonym of M. aspera – as most recently discussed in Chystjakova et al. 2012; Ptáčková et al. 2013) and Phreatamoeba Chávez et al. 1986 (transferred to Mastigamoeba by Simpson et al. 1997).
-
Endolimax Kuenen and Swellengrebel, 1917 (Fig. 4)
Fig. 4 
Endolimax . (a) Endolimax nana live trophozoite (amoeba) showing characteristic finger-shaped pseudopodia and a single nucleus with a vesicular nucleolus. Differential interference contrast light microscopy. (b–d) E. nana prepared slides, stained with iron hematoxylin, bright field light microscopy: (b) trophozoite, showing a large spherical nucleolus in the nucleus. (c, d) Cysts with four nuclei. (e–h) Transmission electron micrographs of Endolimax piscium from Constenla et al. 2013: (e) Whole cell, showing nucleus (N) with central nucleolus (No), and mitochondrion-related organelles (Mi, arrowheads). (f–h) Mitochondrion-related organelles (Mi), with double membranes. Scale bars in a, c, d = 5 μm; in b = 10 μm, in e = 500 nm, in f, g, h = 50 nm (Micrographs in e, f, g, h reprinted from Journal of Fish Diseases 37, Constenla et al., Endolimax piscium sp. nov. (Amoebozoa), causative agent of systemic granulomatous disease of cultured sole, Solea senegalensis Kaup, pp. 229–240, Copyright (2013), with permission from John Wiley & Sons)

Endolimax contains aflagellate uninucleate amoebae reminiscent of Entamoeba. All are intestinal symbionts of various insects and vertebrates including humans.
Circumscription
Trophozoites are aflagellate amoebae measuring up to 20 μm. Actively moving cells form a single broad, hyaline, and eruptive lobopodium. Morphologically, Endolimax closely resembles Entamoeba, except that the nucleus of Endolimax does not contain peripheral heterochromatin (Figs. 4 and 9). There is a single large central nucleolus in the nucleus. Nucleoli in nuclei of cysts are often elongate. Cysts are rounded or elongate, with one to eight nuclei (usually four; rarely many). Approximately 20 Endolimax species have been described, all being commensals of the lower intestine of various animals (e.g., E. nana from humans, E. caviae from rodents, E. gregariniformis from birds, E. clevelandi from reptiles, E. ranarum from amphibians, E. leptocoridis, and E. termitis from insects). Endolimax piscium was reported from lesion in the muscle tissue of a fish (Costenla et al. 2013).
Type Species
Endolimax nana (Wenyon and O’Connor 1917) Kuenen and Swellengrebel, 1917 (basionym Entamoeba nana Wenyon and O’Connor 1917).
Remarks
Because it is aflagellate, Endolimax was traditionally considered a member of Entamoebidae. Based on SSU rRNA phylogeny, Cavalier-Smith et al. (2004) removed Endolimax from Entamoebidae and created the family Endolimacidae for it. Since Endolimax forms an internal branch of Mastigamoeba, Ptáčková et al. (2013) transferred it to Mastigamoebidae. The genus as a whole is relatively understudied (Poulsen and Stensvold 2016).
-
Iodamoeba Dobell, 1919 (Fig. 5)
Fig. 5 
Iodamoeba (a) Trophozoite of Iodamoeba buetschlii showing nucleus with central nucleolus and granules of chromatin; prepared slide, stained with iron-hematoxylin. (b) Cyst of I. buetschlii with a single vacuole; prepared slide, stained with iron-hematoxylin. (c) Transmission electron micrograph of cyst, showing the structure of nucleolar material (No) and chromatin in the nucleus (N). Scale bar in a = 5 μm, in b = 10 μm, in c = 1 μm (Micrograph in c reprinted from Parasitology Research 84, Zaman et al., Ultrastructure of the Iodamoeba bütschlii cyst, pp. 421–422, Copyright (1998), with permission from Springer)

Iodamoeba contains aflagellate uninucleate amoebae reminiscent of Entamoeba and Endolimax that are intestinal commensals of vertebrates.
Circumscription
Trophozoites are aflagellate amoebae measuring up to 20 μm. The cells move slowly by hyaline lobopodia. The nucleus contains a single large nucleolus surrounded by globules, though these do not lie directly beneath the nuclear membrane as in Entamoeba. Cysts are often irregularly shaped and possess a single nucleus with an eccentric nucleolus. Typically, a large glycogen granule is present in the mature cyst. Four Iodamoeba species have been described from lower intestines of mammals (e.g., I. buetschlii from humans) and reptiles (I. testudinis).
Type Species
Iodamoeba buetschlii (Prowazek 1912) Dobell, 1919 (basionym Entamoeba buetschlii Prowazek 1912).
Remarks
Because it is aflagellate, Iodamoeba was traditionally considered a member of Entamoebidae. On the basis of recent molecular phylogenetic work (Stensvold et al. 2012; Ptáčková et al. 2013), it was recently removed from Entamoebidae and transferred to Mastigamoebidae together with Endolimax (Ptáčková et al. 2013).
-
Family Rhizomastixidae Ptáčková et al. 2013
Amoeboflagellate Archamoebae. Trophozoites with single anterior flagellum. Microtubular cone modified into the “rhizostyle.” Amoeboid movement slow. See below and Ptáčková et al. (2013), for discussion of the nomen nudum Rhizomastigidae.
-
Rhizomastix Alexeieff, 1911 (Fig. 6)
Fig. 6 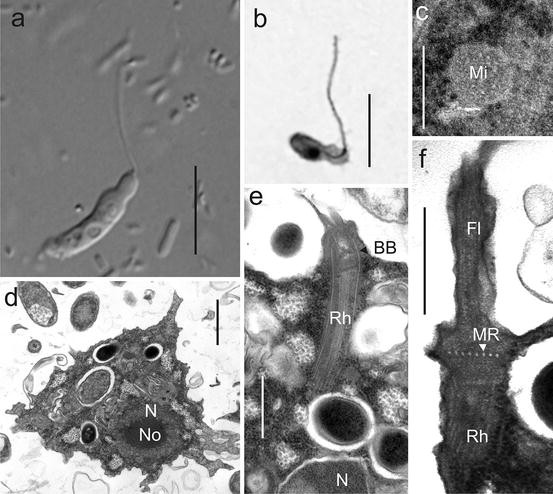
Rhizomastix. (a–f) Rhizomastix libera. (a) Light micrograph of flagellate showing central, rounded nucleus and nucleolus and posterior food vacuoles (DIC). (b) Protargol-stained cell showing the rhizostyle connecting the anterior flagellum to the central nucleus. (c–f) Transmission electron microscopy. (c) Mitochondrion-related organelle (Mi) with double membrane. (d) Whole cell, showing large nucleus (N) and nucleolus (No) and food vacuoles. (e, f) Details of the flagellar apparatus showing the flagellum (Fl), basal body (BB) with proximally extending microtubular rhizostyle (Rh), and laterally extending microtubular root (MR). N nucleus. Scale bar in a, b = 10 μm, in c, f = 200 nm, in d = 1 μm, and in e = 500 nm (Figures c, d, e, and f reprinted from Protist 164, E. Ptáčková et al., Evolution of Archamoebae: Morphological and Molecular Evidence for Pelobionts Including Rhizomastix, Entamoeba, Iodamoeba, and Endolimax, Pp. 380–410, Copyright (2013), with permission from Elsevier)

Rhizomastix contains amoeboid uniflagellate or biflagellate species whose microtubular cone has been modified into a tube that extends through the cell. Most species are endobiotic, but two (and possibly a third) free-living species have been described as well.
Circumscription
Archamoebae with a uniflagellate (biflagellate in R. biflagellata) trophic stage. A single-layered cylindrical bundle of microtubules, the “rhizostyle,” extends from the bottom edge of the basal body through the cell. A second microtubular element, arising from the base of the basal body and running parallel to the rhizostyle, may also be present. A single-layered root of microtubules arises from the side of the basal body (Fig. 2f). Fine structure of the basal body is unknown; a cylinder is present in the transition zone of the flagellum. The flagellum has a conventional eukaryotic “9 + 2” arrangement of microtubules, but it is unclear whether it lacks outer dynein arms. Flagellar movement is relatively fast in comparison with Mastigamoeba. The nucleus is similar to that of Entamoeba, having a single central (large) nucleolus and peripheral heterochromatin granules at least in some species. The flagellates may transform to uninucleate amoebae. Binucleate cysts have been reported in some species. Sixteen Rhizomastix species have been described. Most of them are intestinal commensals of vertebrates and insects (e.g., R. gracilis, R. biflagellata, and R. bicoronata), three are described as free-living (R. libera, R. borealis, R. varia); and some are of uncertain status (R. elongata) (Ptáčková et al. 2013; Zadrobílková et al. 2016).
Type Species
Rhizomastix gracilis Alexeieff, 1911.
Remarks
The genus Rhizomastix was recently transferred into the Archamoebae and a new family Rhizomastixidae established to accommodate it (Ptáčková et al. 2013). Rhizomastix was classified with Mastigamoeba and Mastigella by Kudo (1939, 1977); Čepička (2011) suggested it might be related to pelobionts; its position was clarified on the basis of molecular and ultrastructural data (Ptáčková et al. 2013; Zadrobílková et al. 2016). The name Rhizomastigidae has historically been used for today’s Mastigamoebidae (e.g., Bütschli 1880, 1884; Calkins 1901; Lepşi 1965; Reichenow 1952). The name was created by Bütschli (1884) as Rhizomastigina and later standardized to Rhizomastigidae by Calkins (1901); however, as it was not based on and often did not include Rhizomastix, Rhizomastigidae is regarded by some as a nomen nudum (Loeblich and Tappan 1961). The composition of Rhizomastigidae has typically been very confused (e.g., Cavalier-Smith and Scoble 2013), leading Ptáčková et al. (2013) to create Rhizomastixidae as the family containing Rhizomastix.
Suborder Pelomyxina
Starobogatov, 1980
See Pánek et al. (2016) for current composition. The clade containing Pelomyxa and Mastigella; using the branch-based definition introduced and discussed in Pánek et al. (2016), the clade containing Pelomyxa palustris Greeff, 1874, and all organisms sharing a more recent common ancestor with P. palustris than with Mastigamoeba balamuthi (Chávez et al. 1986) Simpson et al. 1997.
-
Family Pelomyxidae Schulze, 1877
Anaerobic or microaerophilic flagellated amoebae with slow-beating monokinetid or immobile polykinetids. See Zadrobílková et al. 2015 for current composition.
-
Pelomyxa Greeff, 1874 (Fig. 7)
Fig. 7 
Pelomyxa. (a–g) Pelomyxa schiedti, strain SKADARSKE in (a, b), strain TIWI in (c), strain WACT07 in (d–g). (a–c) Differential interference contrast showing cells filled with granules and endosymbionts, distinctive nuclear structure (in b), leading pseudopodium (in a, arrow upper right), a bulbous, villous uroid-like area (in a, arrow lower right), and multiple immobile poorly visible flagella (in c). (d–g) Transmission electron microscopy; (d) section through the cell showing the amoeboid body, single nucleus, and endosymbionts. (e) Transverse section of the flagellum with aberrant arrangement of microtubules. (f) Longitudinal section of the flagellar apparatus. (g) Detail of the nucleus, showing peripheral chromatin and small nucleolus. Scale bars in a–c = 20 μm; d = 5 μm; e = 200 nm; f and g = 1 μm (Figures a–g reprinted from Protist 164, E. Ptáčková et al., Evolution of Archamoebae: Morphological and Molecular Evidence for Pelobionts Including Rhizomastix, Entamoeba, Iodamoeba, and Endolimax, pp. 380–410, Copyright (2013), with permission from Elsevier)

Pelomyxa was originally described as a large multinucleate amoeba, with a division of the cytoplasm into an inner layer containing organelles displaying fountain-flow movement and a clear hyaline outer layer from which pseudopodia can “roll” out and with a posterior uroid attaching the amoeba to the substrate. Later reports extended the description to refer to prokaryotes that coexist endosymbiotically in the cell (van Bruggen et al. 1988) and to non-motile flagella (Frolov 2011; Griffin 1979, 1988; Seravin and Goodkov 1987a).
Circumscription
Pelomyxa is distinguished by the form most usually encountered: a large multinucleate amoeba, with many inactive flagella. It includes polymorphic species, with a large multinucleate amoeboid feeding form up to 5000 μm long, with poorly motile or non-motile flagella that insert into a clear “hyaline” outer cytoplasmic layer, which encloses an inner layer filled with organelles, with endosymbiotic bacteria, and sometimes with refringent cytoplasmic granules (sand); movement is directed by fountain-flow cytoplasm and an anterior monopodium. The posterior uroid is active in food uptake. Cysts about 100 μm in diameter are known from at least one species. Pelomyxa species can be divided into two groups on the basis of the organization of the flagellar apparatus: the first group is characterized by a long basal body and very numerous radiating microtubules, while the second is characterized by a short basal body associated with a very reduced number of radiating microtubules. There is at least one intermediate state between these groups (Chystjakova et al. 2014). Cells have many flagella, each arising from a single basal body; each basal body gives rise proximally to a cone of microtubules, which has not been shown to extend to the nucleus (though several species do have microtubules surrounding the nucleus). The transition zone of each flagellum contains a transitional column. Flagella are with or (in most cases) without a 9 + 2 arrangement of microtubules in flagellar axoneme; the pattern of microtubules may be unstable in individuals in some species, particularly in the central pair or group of axonemal microtubules. The flagella insert into the hyaline outer cytoplasmic layer, which is devoid of organelles (ER, nuclei, vacuoles). Nuclei show distinctive, diagnostic patterns of chromatin condensation and may be surrounded by endosymbiotic bacteria. Morphological and molecular data suggest that Pelomyxa may have evolved from Mastigella by nuclear and flagellar multiplication (Zadrobílková et al. 2015).
Type Species
Pelomyxa palustris Greeff, 1874.
Remarks
Pelomyxa has been widely reported (e.g., Greeff 1874; Griffin 1988; Grüber 1885; Schirch 1914; Schulze 1875a; Whatley and Chapman-Andresen 1990; Frolov 2011). Pelomyxa palustris is currently relatively poorly known, and descriptions of it (e.g., Griffin 1988; Whatley and Chapman-Andresen 1990) probably encompass the variation of multiple species (Frolov et al. 2004; Goodkov et al. 2004). On the other hand, very few of the light microscopy-based species descriptions currently available can be differentiated from the life cycle described by Whatley and Chapman-Andresen (1990). All the previous accounts of species probably describe real variation; however it is not currently possible to assign most of this variation to different species in any consistent or clear way, on the basis of the published descriptions. Species are currently distinguished mainly on characteristic patterns of nuclear chromatin, the thickness of the outer cytoplasmic layer, and the flagellar apparatus (Berdieva et al. 2015; Chystjakova and Frolov 2011; Chystjakova et al. 2014; Frolov 2011; Frolov et al. 2005a, b, 2006; Griffin 1979, 1988; Seravin and Goodkov 1987a; Ptáčková et al. 2013; Zadrobílková et al. 2015).
-
Mastigella Frenzel, 1897 (Fig. 8)
Fig. 8 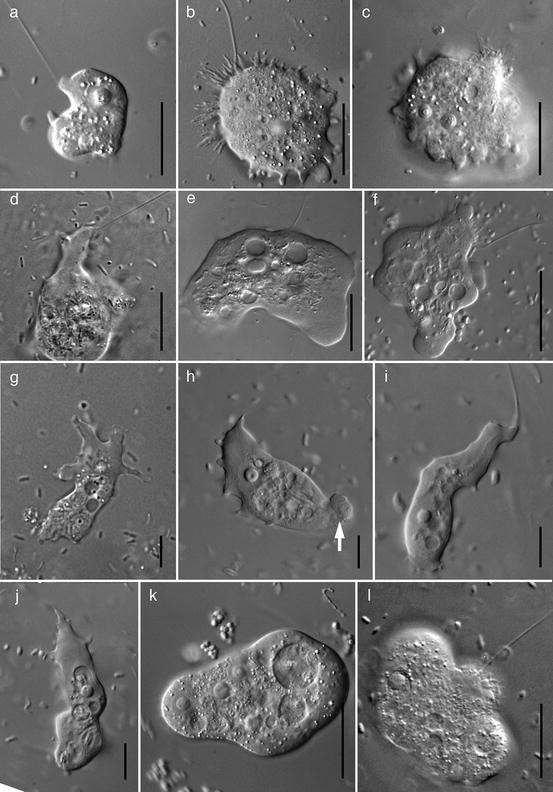
Mastigella. (a–l) Differential interference contrast. (a–c) Mastigella erinacea strain TOLEDO, showing binucleate cells with distinctive “fried-egg” nucleus and granular nucleolus, endosymbiotic bacteria, and highly variable villous, lobate or finger-shaped pseudopodia. (d, e) Mastigella ineffigiata strain OLB6AN, showing “shapeless” morphology and conspicuous endosymbionts. (f) Mastigella rubiformis strain HRAAN, showing cells with hyaline area, distinctive “Pelomyxa-like” nucleus, and prominent endosymbiotic bacteria. (g, h, j) Mastigella eilhardi strain ATCC 50342, showing pseudopodial variation; nucleus with “hollow,” “donut-shaped” nucleolus; and endosymbiotic bacteria, with a posterior uroid (arrow). (i) M. eilhardi strain GO7 showing its characteristic swanlike long “neck” and posterior villous pseudopodia. (k) Mastigella erinacea strain KORISSION, showing binucleate or quadrinucleate cells with distinctive “fried-egg” nucleus with a granular nucleolus and villous pseudopodia from which the flagellum originates in some cells. Scale bars in a–f, i, k, l = 20 μm; g, h, j = 10 μm (Figures a–l reprinted from Protist 164, E. Ptáčková et al., Evolution of Archamoebae: Morphological and Molecular Evidence for Pelobionts Including Rhizomastix, Entamoeba, Iodamoeba, and Endolimax, pp. 380–410, Copyright (2013), with permission from Elsevier)

Mastigella contains amoeboid cells with a flagellated basal body with a cone of microtubules, but there is no connection between the cone and the nucleus. Endosymbiotic prokaryotes are present in several species.
Circumscription
Archamoebae with a uniflagellated trophic stage, in which the nucleus is not connected to a cone of microtubules that arises from the base and sides of the single (flagellated) basal body; a cylinder is present in the transition zone of the flagellum. A single root of microtubules arises from the side of the basal body, and the root has a bilaminar sheet on its anterior edge. Basal bodies have nine triplets of microtubules. The flagellum has a conventional eukaryotic “9 + 2” arrangement of microtubules but lacks outer dynein arms. The flagellum may contribute to cell movement, but its beating is typically slower and less effective than that seen in Mastigamoeba. The flagellates may, at least in some species, transform to amoebae with one, few, or many nuclei. The nuclei may contain a distinctive distribution of chromatin; endosymbiotic bacteria may be present; the outside of the cell may be covered with irregular spines. Both flagellate and amoeboid forms may transform into cysts. Cells are from 3 to 150 μm long with one to four flagella up to 150 μm long. Reported from anoxic or low-oxygen freshwater or marine sediments (Walker et al. 2001; Zadrobílková et al. 2015).
Type Species
Mastigella polymastix Frenzel, 1897.
Remarks
Mastigella was introduced to describe an amoeboid flagellate with multiple long flagella extended from the cell body on small “necks” that wandered over the cell body and were not attached to the nucleus (Frenzel 1897). Subsequently, the concept of the genus changed to one of amoeboid flagellates without a connection between the flagellum and nucleus (Goldschmidt 1907a, b; Lemmermann 1914). Following from Goldschmidt’s (1907a, b) informal group “Mastigamöben,” Mastigamoeba and Mastigella were grouped as the Mastigamoebidae by Chatton (Chatton 1925; Kudo 1939, 1977) and have traditionally been thought of as sister taxa. This was initially supported by phylogenies that included the GenBank sequence AF421219: it had been ascribed to Mastigella commutans but belongs to Mastigamoeba punctachora (Ptáčková et al. 2013), meaning that these phylogenies are silent on the placement of Mastigella. However, cell form and movement, nuclear structure, and the presence of endosymbiotic prokaryotes resembling methanogenic archaea (Frolov 2011; van Bruggen et al. 1985; Walker et al. 2001; Zadrobílková et al. 2015) all suggest a relationship of Mastigella with Pelomyxa. This hypothesis was supported by recent phylogenetic analyses where Pelomyxa forms an internal branch of Mastigella, while Mastigamoeba is phylogenetically more distant (Zadrobílková et al. 2015). Other nominal genera that fall within the circumscription of Mastigella include Limulina Frenzel, 1897; Micromastix Frenzel, 1897 (Frenzel 1897) and Mastigamoebula Fantham, 1919 (Fantham 1919).
-
Order Entamoebida Cavalier-Smith, 1993
Using the branch-based definition introduced by and discussed in Pánek et al. (2016), the clade consisting of Entamoeba histolytica Schaudinn, 1903 and all organisms that share a more recent common ancestor with E. histolytica than with Mastigella eilhardi Bürger, 1905.
-
Family Entamoebidae Chatton, 1925
Aflagellate Archamoebae. Flagellar apparatus completely reduced. Amoeboid movement typically monopodial and relatively fast. See Cavalier-Smith 1993 for current composition.
-
Entamoeba Casagrandi and Barbagallo, 1895 (Fig. 9)
Fig. 9 
Entamoeba. (a) Entamoeba moshkovskii free-living trophozoite amoeba showing an anterior hyaline pseudopodium and central nucleus with the characteristic “dot” nucleolus (DIC). (b) E. moshkovskii resting cell, showing the nucleus with its characteristic “dot” nucleolus and peripheral chromatin (DIC). (c, d) Parasitic Entamoeba histolytica trophozoite (c) and immature cyst (d) with two visible nuclei and chromatoid body (arrow); prepared slide material, stained with iron-hematoxylin to show nuclei with a thick ring of peripheral chromatin and a small nucleolus. (e, f) Parasitic Entamoeba coli trophozoite (e) and cyst (f) with eight nuclei (five visible in the focal plane of the photograph); prepared slide material, stained with iron-hematoxylin to show nuclei with large chromatin granules. (g) Transmission electron micrograph of E. histolytica, showing numerous vacuoles (V) and the characteristic arrangement of nuclear chromatin (Ch) and nucleolus (No) in the nucleus (N). Scale bar in a, c, d, e, f = 10 μm; in b = 5 μm (Figure in g reprinted from Parasitic Protozoa Volume 3, 2nd Edition, J.P. Kreier & J.R. Baker, Chapter 3: Parasitic amebas of the intestinal tract., pp. 65–141, Copyright (1993), with permission from Elsevier)

Entamoeba contains most of the species of aflagellate Archamoebae. They are usually intestinal commensals of various animals, though at least three are likely to be free-living. A few species, including E. histolytica from humans, are pathogenic for their hosts.
Circumscription
Trophozoites are aflagellate, uninucleate amoebae measuring up to 60 μm. The flagellar apparatus of Entamoeba is completely absent, and no cytoplasmic microtubules can be found in nondividing cells. Actively moving cells usually crawl using a single or several eruptive lobopodia; the hyaline lobopodia are clearly distinguishable from the granuloplasm. Uroidal filopodia have been rarely observed (Martínez-Palomo 1993). Nuclear structure is distinctive, with a single small, central nucleolus and conspicuous peripheral granules of heterochromatin (Fig. 9). The number and size of the granules differ between particular Entamoeba species. A cyst stage is documented for many species (see above). The number of nuclei in the cyst is an evolutionarily important feature (Silberman et al. 1999). Cysts have been lost in E. gingivalis, which is found in the oral cavity of humans and transferred between hosts via saliva.
Many Entamoeba species have been described. The vast majority live as intestinal endobionts of various animals (e.g., E. histolytica, E. dispar, and E. coli from humans, E. muris and E. cobayae from rodents, E. gallinarum from birds, E. invadens from reptiles, E. ranarum from amphibians, E. gadi from fish, E. aulastomi from leeches, and E. minchini from insects). Some species are pathogenic, the best known being E. histolytica. Further information on endobiotic and parasitic species is given in the “Practical Importance” section.
Three Entamoeba species may be facultatively free-living. Entamoeba moshkovskii is relatively well known and has been repeatedly isolated from wastewaters, anoxic sediments, as well as from human stool (see Heredia et al. 2012; our observations). The second species, E. ecuadoriensis, has been isolated only once from sewage water (Clark and Diamond 1997). The third, E. marina, was isolated from marine tidal flats (Shiratori and Ishida 2016).
Type Species
Entamoeba coli Grassi, 1879, Casagrandi and Barbagallo 1895 (basionym Amoeba coli Grassi 1879).
Remarks
There was some uncertainty regarding the names of genera Entamoeba and Endamoeba during the first half of the twentieth century (see section “History of Genera in the Archamoebae”). Entamoeba histolytica can be, therefore, found under the name Endamoeba histolytica in the older literature. Since the 1950s, the concept of Entamoeba is stable, and Endamoeba is now considered a separate genus (e.g., Patterson et al. 2000). Entamoeba constitutes the family Entamoebidae. Historically, the other aflagellate Archamoebae, Endolimax and Iodamoeba, were classified within Entamoebidae as well but were recently removed from it (Cavalier-Smith et al. 2004; Ptáčková et al. 2013). Endamoeba is considered here as Archamoebae incertae sedis.
Archamoebae Incertae Sedis
-
Mastigina Frenzel, 1897 (Fig. 10)
Fig. 10 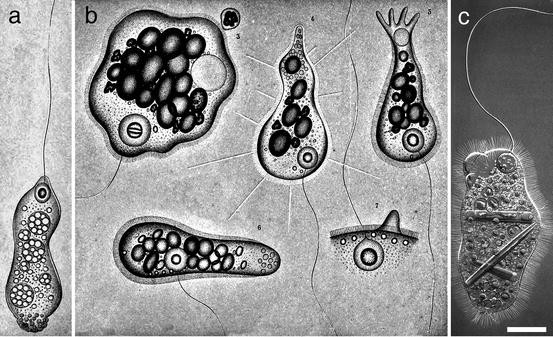
Mastigina. (a) Drawing of one of the two original species of Mastigina, M. paramylon, showing numerous starch granules inside the cell, connection between the long flagellum and the nucleus (with central vesicular nucleolus), and the lack of any lateral pseudopodia. (b) The type species of Mastigina, M. chlamys, showing a dense layer of short, refringent spines and in one case longer spines interspersed with the short ones, a connection between the long flagellum and the nucleus (with central nucleolus), and pseudopodia arising only at the anterior and posterior of the cell, not laterally. (c) Mastigina setosa, showing similar characteristics to M. chlamys but longer, less dense spines. Scale bar in a = 5 μm; in b, c = 20 μm (Drawings in a and b reprinted from Untersuchungen über die mikroskopische Fauna Argentiniens. Erster Teil: Die Protozoen. I und II, Abteilung: die Rhizopoden und Helioamoeben, J. Frenzel, (1897), Erwin Nägele, Stuttgart; with no known copyright restrictions. Drawing in c reprinted from Archiv für Protistenkunde Supplement 1, R. Goldschmidt, Lebensgeschichte der Mastigamöben Mastigella vitrea n.sp. u. Mastigina setosa n.sp., pp. 83–165, Copyright (1907), with permission from Elsevier)

Mastigina contains limax-shape amoeboid cells with a flagellum and no lateral pseudopodia and with a connection between the base of the flagellum and the rounded nucleus. Its microtubular ultrastructure has not been studied by electron microscopy.
Circumscription
Archamoebae with a uniflagellated trophic stage, with limax amoeboid shape and no lateral pseudopodia, and in which the round nucleus is connected to the base of the flagellum, which has the distinctively languid flagellar beat typical of other pelobionts. One species, Mastigina setosa, has fountain-flow cytoplasmic movement (Goldschmidt 1907a). The outside of the cell is covered with closely packed spines in two species, Mastigina chlamys and M. setosa (Frenzel 1897; Goldschmidt 1907a; Skibbe and Zölffel 1991). Reported from anoxic or low-oxygen freshwater sediments.
Type Species
Mastigina chlamys Frenzel, 1897.
Remarks
Mastigina was not clearly distinguished from other pelobiont genera when it was introduced, in the original descriptions of M. chlamys and M. paramylon (Frenzel 1897). Goldschmidt (1907a) defined the genus on the basis of limax body shape with no lateral pseudopodia, round apical nucleus connected closely to the base of the flagellum (as opposed to elongated or drop-shaped, sometimes appearing removed from the base of the flagellum, in Mastigamoeba), characters that are adopted here. Goldschmidt (1907a) also regarded Tricholimax hylae as belonging to Mastigina, leading to later confusion about the characters displayed by Mastigina species: he used fountain-flow cytoplasmic movement as a defining feature of Mastigina, despite it being present only in M. setosa and T. hylae and despite the two species in the original description of Mastigina not displaying it. Ultrastructural studies of Tricholimax hylae have been carried out under the name Mastigina hylae (Brugerolle 1982, 1991), and summaries of the genus have subsequently relied largely on descriptions of T. hylae (Griffin 1988; Brugerolle and Patterson 2000).
Perhaps on the basis that fountain-flow cytoplasm is also present in Mastigamoeba aspera (Chystjakova et al. 2012; Schulze 1875b), Lemmermann (1914) in turn regarded Mastigina as a junior synonym of Mastigamoeba, leading to further confusion about the distribution of fountain-flow cytoplasm and spines in pelobionts. Frolov (2011) redefined Mastigina as containing only taxa with spines on the surface, disregarding the criteria used by Frenzel (1897) and Goldschmidt (1907a). Although 50% of the species currently assigned to Mastigina do have spines, 50% do not, and there are also members of Mastigamoeba and Mastigella with spines.
In the absence of clear ultrastructural and phylogenetic data on any of these species, we see no reason to reject Goldschmidt’s circumscription of the genus.
-
Family Tricholimacidae Cavalier-Smith, 2013
Monotypic family with the characteristics of the sole species. Using the diagnosis of Cavalier-Smith (2013), uniciliate (i.e., uniflagellate) endosymbiotic anaerobes with inactive cilium (i.e., flagellum) with numerous irregularly arranged doublets; no obvious transition zone cylinder; dense microtubule nucleating center attached to the proximal side of the single centriole (with doublets not triplets) nucleates (a) multilayered cone of microtubules that surround nucleus and (b) lateral rhizostyle, a broad band of microtubules with numerous underlying singlet and bundled microtubules.
-
Tricholimax Frenzel, 1897 (Fig. 11)
Fig. 11 
Tricholimax hylae. (a–b) Drawings of whole cells showing the microtubular connection between the short, nonfunctional flagellum and the large apical nucleus with its central nucleolus. Accessory nuclei, with attached kinetids, are visible in the cytoplasm in a, along with numerous food vacuoles. (c) Diagram showing the direction of fountain-flow cytoplasmic streaming, where the posterior uroid is not included. (d, e) The microtubular cone and lateral root (which are clearly visible under the light microscope, even without DIC optics). There is some variation in the presence of a single or a double nucleolus. (f) Nuclei dividing, showing a connection between the posterior poles of the nuclei, with anterior kinetids present. (g) Six cyst stages, showing variation in the number of nuclei from two to four, and gradual development of a central vacuole. (h) Transmission electron micrograph showing the flagellum (Fl), single basal body with microtubular root (Rh), and dense cone of microtubules (mt) connecting to the nucleus (N). Scale bar: in a, b, c = 30 μm, in d, e, f = 10 μm, in g = 20 μm, in h = 3 μm (Drawings in a, d, f, and g reprinted from Archives de Zoologie expérimentale et générale 51, B. Collin, Sur un ensemble de protistes parasites des batraciens (Note préliminaire). Pp. 59–76 (1913) with no known copyright restrictions. Drawings in b, c, e reprinted from Journal of Parasitology 11, E. R. Becker, The morphology of Mastigina hylae (Frenzel) from the intestine of the tadpole. Pp. 213–216, Copyright (1925) with permission from Allen Press. Micrograph in h reprinted, from Protistologica 18, G. Brugerolle, Caractères ultrastructuraux d’une mastigamibe: Mastigina hylae (Frenzel), pp. 227–235, Copyright (1982), with permission from Elsevier)

This monotypic genus has been described from the guts of amphibians. Cells have a limax shape with fountain-flow cytoplasmic movement and without lateral pseudopodia, and the anterior nucleus is connected to the short non-motile flagellum, which lacks normal 9 + 2 organization of the microtubular axoneme.
Circumscription
The one species, Tricholimax hylae, exists mostly as flagellates measuring 50–135 μm. Flagellated amoeboid and cyst stages (up to 30 μm in diameter with up to four nuclei) may form. Flagellated cells have one to six nuclei, one of which is connected to the emergent flagellum. In the flagellar apparatus, there is a cone of microtubules arising from base and sides of the single basal body; these enclose the nucleus and connect to the nuclear envelope by microfibrils. A single root of 32 microtubules arises from the side of basal body, and there is a bilaminar sheet associated with the anterior edge. There is an “n + n” arrangement of microtubules in the flagellar axoneme, though no dynein arms are visible. There may be reduced internal basal bodies with cones of microtubules attached to extra internal nuclei (Brugerolle 1982, 1991).
Type Species
Tricholimax hylae Frenzel, 1897.
Remarks
This genus can be distinguished from Mastigina because of the atypical organization of the single non-motile flagellum. It has been widely described (Frenzel 1897; Goldschmidt 1907b; Collin 1913; Lemmermann 1914; Becker 1928; Sassuchin 1928; Ivanic 1936; Chatton 1953; Brugerolle 1982, 1991; Griffin 1988). The occurrence of the “n + n” flagellar organization and fountain-flow cytoplasm both here and in Pelomyxa has resulted in the placement of both genera in the Pelomyxidae, though this circumscription is now widened to also include Mastigella. No molecular phylogenetic information exists for Tricholimax, so its formal placement in the Pelomyxidae remains premature.
-
Endamoeba Leidy, 1879 (Fig. 12)
Fig. 12 
Endamoeba (a–e): Endamoeba blattae, redrawn from Kudo (1960). (a, b) Trophozoites, showing the distinctive arrangement of chromatin in the nucleus and eruptive pseudopodia. Drawn from live material. (c) Nucleus, showing the distinctive ring of chromatin. (d) Multinucleate cyst, as seen when stained with iron-hematoxylin, showing changed nuclear structure, relative to (e) Cyst, showing distinctive ring of chromatin in the nuclei. Scale bar in a, b = 10 μm, in c = 4 μm, in d, e = 5 μm. Pictures redrawn from Kudo (1960)

Poorly known genus containing aflagellate Archamoebae found in insects.
Circumscription
Trophozoites measure up to 120 μm. They are aflagellate and move slowly by one or several lobopodia. Unlike in Entamoeba, there is no marked distinction between hyaloplasm and granuloplasm (Wenyon 1926). The nucleus of Endamoeba has a typical structure dissimilar from that of Entamoeba and other aflagellate Archamoebae (Wenyon 1926; compare Fig. 12 with Figs. 4, 5, and 9). Several species have been described from the hindgut of insects, such as Endamoeba blattae from cockroaches and E. disparata from termites.
Type Species
Endamoeba blattae Bütschli, 1878 Leidy 1879 (basionym Amoeba blattae Bütschli, 1878).
Remarks
Probable member of Entamoebidae. This classification is, however, tentative, since no sequence data from Endamoeba are currently available.
Maintenance and Cultivation
Various free-living Archamoebae from the genera Mastigamoeba, Mastigella, and Rhizomastix can be isolated and maintained in xenic cultures with bacteria. Sonneborn’s Paramecium medium (cereal grass infusion medium, ATCC medium 802; see www.atcc.org) is suitable for freshwater strains (Ptáčková et al. 2013). Seawater 802 medium (ATCC medium 1525; see www.atcc.org) can be used for the cultivation of marine pelobionts (Zadrobílková et al. 2015). The strains grow well at room temperature in sealed 15 ml tubes with 10 ml of medium and transfers every 1–3 weeks or in sealed tissue flasks filled with medium and transfers every 1–2 months. Although the Archamoebae are anaerobic and die when exposed to oxygen for a longer period, no special precautions are necessary, since bacteria present in the culture quickly consume the oxygen. Most strains grow well also under anoxic conditions.
Members of the genera Entamoeba (including the free-living Entamoeba moshkovskii) and Endolimax can be isolated and maintained in xenic cultures as well. Various media developed for cultivation of intestinal flagellates and amoebae, both biphasic (e.g., LE medium, Robinson’s biphasic medium) and monophasic (TYSGM-9 medium, Robinson’s monophasic medium) can be used (see Clark and Diamond 2002). Culture conditions depend on the origin of the organisms – strains from invertebrates and poikilotherm vertebrates are cultivated at room temperature with transfers once a week to once a month; strains from homoiotherms should be cultivated at 37 °C with transfers approximately twice a week.
Only a few species of Entamoeba and Mastigamoeba balamuthi have been successfully axenized. The process of isolation and axenization of Entamoeba histolytica is described in Clark and Diamond (2002). For maintenance, several complex media, such as TYI-S-33, YI-S, and LYI-S-2 (ATCC medium 2154), can be used (for preparation, see Clark and Diamond 2002 and www.atcc.org). Apart from Entamoeba spp., only a single strain of Mastigamoeba balamuthi has been axenized (Chávez et al. 1986). Chávez et al. (1986) described the process of the axenization in a detail. Mastigamoeba balamuthi can be maintained in PYGC (Chávez et al. 1986) or LYI-S-2 (see above) media.
So far, only one successful long-term cultivation attempt of Pelomyxa spp. has been published (Zadrobílková et al. 2015). Cells of Pelomyxa spp. can be maintained alive for several months in hermetically closed flasks filled with fresh water and sediments, or with Lozina-Lozinskii medium supplemented with boiled detritus (see Chystjakova and Frolov 2011; Frolov et al. 2011).
Cultures of Archamoebae can be cryopreserved in the presence of 5% dimethyl sulfoxide using a standard procedure and stored in liquid nitrogen. Detailed protocols for cryopreservation of Entamoeba cultures are described in Diamond (1995) and Samarawickrema et al. (2001).
Evolutionary History
Phylogeny and Evolution
Separately and together, members of the Archamoebae have been considered to be some of the most deep-branching eukaryotes, on the basis of lacking mitochondria and Golgi dictyosomes, and having a “simple” or nonexistent flagellar apparatus. Using the argument that some or all of these traits are primitive, it was suggested that the pelobionts represent a very early stage in the evolution of eukaryotes (Brugerolle 1993; Cavalier-Smith 1983; Griffin 1979, 1988; Margulis 1970; Patterson 1994; Patterson and Sogin 1992; Whatley 1976; Whatley and Chapman-Andresen 1990). This evolutionary significance was in the context of an evolutionary paradigm known as the Archezoa hypothesis (Cavalier-Smith 1983), where “amitochondriate” protists represented basal eukaryotic lineages that had diverged before the acquisition of the mitochondrial endosymbiont and other key eukaryotic innovations (e.g., introns, Golgi dictyosomes, peroxisomes). The Archezoa hypothesis was supported by molecular phylogenetic trees of eukaryotes consisting of a “crown” (animals, plants, fungi, and many algae) and a ladderlike sequential divergence of amitochondriate parasites from the base (Sogin 1991; Sogin and Silberman 1998).
In the late 1990s, developments in phylogenetic methods and the identification of mitochondriate remnant organelles led to the rejection of the Archezoa hypothesis and the crown/base view of eukaryotic relationships (Roger 1999). The development of evolutionary models incorporating heterogeneity among taxa (of evolutionary rate or of base composition) led to the realization that systematic phylogenetic error, such as “long branch attraction,” had had a major effect on the positioning of “archezoan” organisms in the tree (Dacks et al. 2002; Holder and Lewis 2003; Philippe and Germot 2000). The finding of gene sequences of mitochondrial origin (cpn60, HSP70, IscU) and the localization of these gene products (by immuno-microscopy) to double-membrane-bounded organelles (Bui et al. 1996; Clark and Roger 1995; Roger et al. 1996, 1998; Tovar et al. 2003) have led to the identification of mitochondrion-derived organelles (i.e., hydrogenosomes and mitosomes) in almost all of the proposed “amitochondriate” organisms (van der Giezen 2009).
The identification of mitochondrial-remnant organelles (discussed above) and comparisons of ribosomal DNA sequences (Cavalier-Smith 1997; Hinkle et al. 1994; Milyutina et al. 2001; Morin and Mignot 1995; Silberman et al. 1999; Stiller and Hall 1999) and other genes (Arisue et al. 2002; Bapteste et al. 2002; Hannaert et al. 2000) now suggest that the Archamoebae are derived within the mitochondriate taxon Amoebozoa and are not deep-branching eukaryotes (Cavalier-Smith 1997, 1998; Edgcomb et al. 2002; Simpson and Roger 2004).
As discussed above, systematics within the Archamoebae is currently undergoing considerable flux, while many gene and protein sequences are obtained for many more representatives of the group. So far this has clarified that endobiotic taxa are distributed among the free-living taxa, showing that parasitism has clearly evolved multiple times in the Archamoebae (Pánek et al. 2016; Ptáčková et al. 2013; Stensvold et al. 2012; Zadrobílková et al. 2015, 2016). Obtaining multiple sequences from a single taxon can improve the robustness of placement of particular taxa, both at species level and at higher levels – for example the placement of Pelomyxa has long been unstable because the original sequence obtained was highly divergent (Milyutina et al. 2001; Ptáčková et al. 2013). Further sequences from Mastigella, Tricholimax, and Mastigina will be key to resolving relationships among the genera of the Archamoebae; and sequences from Mastigamoeba aspera would permit certainty in the proposed split of Mastigamoeba discussed above (Pánek et al. 2016) (Fig. 13).

Phylogenetic tree of Archamoebae based on SSU rRNA genes, showing generic relationships within families. Mastigamoeba is divided into two main clades, corresponding to “Mastigamoebidae A,” with 100% ML bootstrap support and Bayesian posterior probability of 1, and “Mastigamoebidae B,” with 99% ML bootstrap support and Bayesian posterior probability of 1. Note that the sequence labelled “Mastigella commutans” is thought to be from Mastigamoeba punctachora rather than M. commutans (see Zadrobílková et al. 2015). Support for relationships between families is not recovered from SSU rRNA data alone, requiring a multigene approach (Pánek et al. 2016). Sequences were aligned using the G-INS-I algorithm with default settings, on the MAFFT7 server (http://mafft.cbrc.jp/alignment/server/). The resulting alignment was manually edited to remove ambiguously aligned sites; the final dataset contained 1263 nucleotide positions. A maximum-likelihood phylogenetic tree was constructed in RAxML 7.2.3 (Stamatakis 2006) using the GTRGAMMAI model; the values at the branches represent statistical support in maximum likelihood (RAxML, 1000 pseudoreplicates)/Bayesian posterior probability (MrBayes, GTR + Γ + I + covarion model). Support values below 50/0.90 are not shown or are represented by an asterisk (*)
References
Aguilera, P., Barry, T., & Tovar, J. (2008). Entamoeba histolytica mitosomes: Organelles in search of a function. Experimental Parasitology, 118, 10–16.
Alexeieff, A. (1911). Notes sur les flagellés. Archives de Zoologie expérimentale et générale, 6, 491–527.
Arisue, N., Hashimoto, T., et al. (2002). The phylogenetic position of the pelobiont Mastigamoeba balamuthi based on sequences of rDNA and translation elongation factors EF-1alpha and EF-2. Journal of Eukaryotic Microbiology, 49, 1–10.
Bapteste, E., Brinkmann, H., et al. (2002). The analysis of 100 genes supports the grouping of three highly divergent amobae: Dictyostelium, Entamoeba and Mastigamoeba. Proceedings of the National Academy of Sciences of the USA, 99, 1414–1419.
Barberà, M. J., Ruiz-Trillo, I., et al. (2007). The diversity of mitochondrion-related organelles amongst eukaryotic microbes. In W. F. Martin & M. Müller (Eds.), Origin of mitochondria and hydrogenosomes (pp. 239–275). Berlin: Springer.
Becker, E. R. (1925). The morphology of Mastigina hylae (Frenzel) from the intestine of the tadpole. Journal of Parasitology, 11, 213–216.
Becker, E. R. (1928). Streaming and polarity in Mastigina hylae (Frenzel). Biological Bulletin, 54, 109–117.
Berdieva, M. A., Chystjakova, L. V., Miteva, O. A., Frolov, A. O., & Gudkov, A. V. (2015). Light- and electron-microscopic study of pelobiont Pelomyxa secunda (Gruber, 1884) comb. Nov. (archamoebae, Pelobiontida). Tsitologiya, 57, 62–69.
Bernard, C., Simpson, A. G. B., & Patterson, D. J. (2000). Some free-living flagellates (Protista) from anoxic habitats. Ophelia, 52, 113–142.
Blochmann, F. (1894). Kleine Mitteilungen über Protozoen. Biologisches Zentralblatt, 14, 82–91.
Bovee, E. C. (1972). The lobose amoebas. IV. A key to the order Granulopodida Bovee and Jahn 1966: And descriptions of some new and little-known species in the order. Archiv für Protistenkunde, 114, 371–403.
Bovee, E. C., & Sawyer, T. K. (1979). Marine Flora and Fauna of the Northeastern United States. Protozoa: Sarcodina: Amoebae. Technical Report NMFS Circular 419. U.S. Department of Commerce; National Oceanic and Atmospheric Administration; National Marine Fisheries Service.
Bredeston, L. M., Caffaro, C. E., Samuelson, J., & Hirschberg, C. B. (2005). Golgi and endoplasmic reticulum functions take place in different subcellular compartments of Entamoeba histolytica. Journal of Biological Chemistry, 280, 32168–32176.
Brugerolle, G. (1982). Caractères ultrastructuraux d’une mastigamibe: Mastigina hylae (Frenzel). Protistologica, 18, 227–235.
Brugerolle, G. (1991). Flagellar and cytoskeletal systems in amitochondrial flagellates: Archamoeba, Metamonada and Parabasala. Protoplasma, 164, 70–90.
Brugerolle, G. (1993). Evolution and diversity of amitochondrial zooflagellates. Journal of Eukaryotic Microbiology, 40, 616–618.
Brugerolle, G., & Patterson, D. J. (2000). Order Pelobiontida Page 1976. In J. Lee, G. Leedale, & P. Bradbury (Eds.), An illustrated guide to the protozoa (2nd ed., pp. 1097–1103). Lawrence: Allen Press Inc..
Bui, E. T., Bradley, P. J., & Johnson, P. J. (1996). A common evolutionary origin for mitochondria and hydrogenosomes. Proceedings of the National Academy of Sciences of the USA, 93, 9651–9656.
Bütschli, O. (1880). Protozoa. Erster Band. Erster Abteilung: Sarkodina und Sporozoa. In H. G. Bronn (Ed.), Klassen und Ordnungen des Tier-Reichs, wissenschaftlich dargestellt in Wort und Bild (Vol. 1, Abt. 1, pp. 1–242). Leipzig/Heidelberg: CH Winter Verlag.
Bütschli, O. (1884). Protozoa. Erster Band. Zweiter Abtheilung: Mastigophora. In H. G. Bronn (Ed.), Klassen und Ordnungen des Thier-Reichs, wissenschaftlich dargestellt in Wort und Bild, (Vol. 1, Abt. 1, pp. 617–872). Leipzig/Heidelberg: CH Winter Verlag.
Calaway, W. T., & Lackey, J. B. (1962). Waste treatment protozoa. Flagellata, Florida engineering series (Vol. 3). Gainesville: University of Florida/College of Engineering.
Calkins, G. N. (1901). The protozoa. New York: Columbia Press.
Casagrandi, O., & Barbagallo, P. (1895). Ricerche biologiche e cliniche sull' Amoeba coli (Lösch). (Nota preliminare). Bollettino delle sedute della Accademia Gioenia di Scienze Naturali in Catania. 39: 4.
Cavalier-Smith, T. (1983). A 6-kingdom classification and a unified phylogeny. In W. Schwemmler & H. E. A. Schenk (Eds.), Endocytobiology II. Intracellular space as oligogenetic ecosystem (pp. 1027–1034). Berlin: de Gruyter.
Cavalier-Smith, T. (1987a). The origin of eukaryotic and archaebacterial cells. Annals of the New York Academy of Sciences, 503, 17–54.
Cavalier-Smith, T. (1987b). Eukaryotes with no mitochondria. Nature, 326, 332–333.
Cavalier-Smith, T. (1991). Archamoebae: The ancestral eukaryotes? Biosystems, 25, 25–38.
Cavalier-Smith, T. (1997). Amoeboflagellates and mitochondrial cristae in eukaryote evolution: Megasystematics of the new protozoan subkingdoms Eozoa and Neozoa. Archiv für Protistenkunde, 147, 237–258.
Cavalier-Smith, T. (1998). A revised six-kingdom system of life. Biological Reviews, 73, 203–266.
Cavalier-Smith, T. (2013). Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European Journal of Protistology, 49, 115–178.
Cavalier-Smith, T., & Chao, E. E. (1995). The opalozoan Apusomonas is related to the common ancestor of animals, fungi, and Choanoflagellates. Proceedings of the Royal Society of London Series B: Biological Sciences, 261, 1–6.
Cavalier-Smith, T., & Scoble, J. M. (2013). Phylogeny of Heterokonta: Incisomonas marina, a uniciliate gliding opalozoan related to Solenicola (Nanomonadea), and evidence that Actinophryida evolved from raphidophytes. European Journal of Protistology, 49, 328–353.
Cavalier-Smith, T., Chao, E. E. Y., & Oates, B. (2004). Molecular phylogeny of Amoebozoa and the evolutionary significance of the unikont Phalansterium. European Journal of Protistology, 40, 21–48.
Čepička, I. (2011). Rhizomastix biflagellata sp. nov., a new amoeboflagellate of uncertain phylogenetic position isolated from frogs. European Journal of Protistology, 47, 10–15.
Chan, K. W., Slotboom, D. J., et al. (2005). A novel ADP/ATP transporter in the mitosome of the microaerophilic human parasite Entamoeba histolytica. Current Biology, 15, 737–742.
Chatton, E. (1925). Pansporella perplexa. Réflections sur la biologie et la phylogenie des protozoaires. Annales des sciences naturelles, Série, 10(8), 5–84.
Chatton, E. (1953). Classe des Lobosa Leidy, 1879. Ordre des amoebiens nus ou Amoebaea. In P. P. Grassé (Ed.), Traité de Zoologie (pp. 5–91). Paris: Masson et Compagnie.
Chávez, L. A., Balamuth, W., & Gong, T. (1986). A light and electron microscopical study of a new, polymorphic free-living amoeba, Phreatamoeba balamuthi n. g., n. sp. Journal of Protozoology, 33, 397–404.
Chavez-Munguia, B., Espinosa-Cantellano, M., Castanon, G., & Martinez-Palomo, A. (2000). Ultrastructural evidence of smooth endoplasmic reticulum and Golgi-like elements in Entamoeba histolytica and Entamoeba dispar. Archives of Medical Research, 31, S165–S167.
Chávez-Munguía, B., Tsutsumi, V., & Martínez-Palomo, A. (2006). Entamoeba histolytica: Ultrastructure of the chromosomes and the mitotic spindle. Experimental Parasitology, 114, 235–239.
Chystjakova, L. V., & Frolov, A. O. (2011). Light and electron microscopic study of Pelomyxa stagnalis sp. n. (Archamoebae, Pelobiontida). Cell and Tissue Biology, 5, 90–97.
Chystjakova, L. V., Miteva, O. A., & Frolov, A. O. (2012). Morphology of Mastigamoeba aspera Schulze, 1875 (Archamoebae, Pelobiontida). Cell and Tissue Biology, 6, 189–196.
Chystjakova, L. V., Berdieva, M. A., Frolov, A. O., & Goodkov, A. V. (2014). Reisolation and redescription of pelobiont Pelomyxa paradoxa Penard, 1902 (Archamoebae, Pelobiontida). Tsitologiya, 56, 770–778.
Clark, C. G., & Diamond, L. S. (1997). Intraspecific variation and phylogenetic relationships in the genus Entamoeba as revealed by riboprinting. Journal of Eukaryotic Microbiology, 44, 142–154.
Clark, C. G., & Diamond, L. S. (2002). Methods for cultivation of luminal parasitic protists of clinical importance. Clin Microbiol Rev, 15, 329–341.
Clark, C. G., & Roger, A. J. (1995). Direct evidence for secondary loss of mitochondria from Entamoeba histolytica. Proceedings of the National Academy of Sciences of the USA, 92, 6518–6521.
Collin, B. (1913). Sur un ensemble de protistes parasites des batraciens (Note préliminaire). Archives de Zoologie expérimentale et générale, 51, 59–76.
Conforti, V., Walne, P., & Dunlap, J. (1994). Comparative ultrastructure and elemental composition of the envelopes of Trachelomonas and Strombomonas (Euglenophyta). Acta Protozoologica, 33, 71–78.
Constenla, M., Padrós, F., & Palenzuela, O. (2013). Endolimax piscium sp. nov. (Amoebozoa), causative agent of systemic granulomatous disease of cultured sole, Solea senegalensis Kaup. Journal of Fish Diseases, 37, 229–240.
Dacks, J. B., Marinets, A., Doolittle, W. F., Cavalier-Smith, T., & Logsdon, J. M. (2002). Analyses of RNA polymerase II genes from free-living protists: Phylogeny, long branch attraction, and the eukaryotic big bang. Molecular Biology and Evolution, 19, 830–840.
Dacks, J. B., Davis, L. A. M., et al. (2004). Evidence for Golgi bodies in proposed ‘Golgi-lacking’ lineages. Proceedings of the Royal Society of London B: Biological Sciences, 270, S167–S171.
Diamond, L. S. (1961). Axenic cultivation of Entamoeba histolytica. Science, 134, 336–337.
Diamond, L. S., & Clark, C. G. (1993). A redescription of Entamoeba histolytica Schaudin, 1903 (emended Walker, 1911) separating it from Entamoeba dispar Brumpt, 1925. Journal of Eukaryotic Microbiology, 40, 340–344.
Diamond L. S. (1995). Cryopreservation and storage of parasitic protozoa in liquid nitrogen. Journal of Eukaryotic Microbiology, 42, 585–590.
Dobell, C. (1919). The amoebae living in man: A zoological monograph. London: J. Bale and Danielsson.
Edgcomb, V., Simpson, A. G. B., et al. (2002). Pelobionts are degenerate protists: Insights from molecules and morphology. Molecular Biology and Evolution, 19, 978–982.
El-Hashimi, W., & Pitman, F. (1970). Ultrastructure of Entamoeba histolytica trophozoites obtained from the colon and from in vitro cultures. American Journal of Tropical Medicine and Hygiene, 19, 215–226.
Embley, T. M., & Martin, W. (2006). Eukaryotic evolution, changes and challenges. Nature, 440, 623–630.
Embley, T. M., Finlay, B. J., et al. (1995). Multiple origins of anaerobic ciliates with hydrogenosomes within the radiation of aerobic ciliates. Proceedings of the Royal Society of London Series B: Biological Sciences, 262, 87–93.
Fantham, H. B. (1919). Some parasitic protozoa found in South Africa. South African Journal of Science, 16, 185–191.
Fenchel, T., & Finlay, B. J. (1995). Ecology and evolution in anoxic worlds. Oxford: Oxford University Press.
Frenzel, J. (1897). Untersuchungen über die mikroskopische Fauna Argentiniens. Erster Teil: Die Protozoen. I und II, Abteilung: die Rhizopoden und Helioamoeben. Stuttgart: von Erwin Nägele.
Frolov, A. O. (2011). Pelobiontida (Page 1976) Griffin 1988. In O. N. Pugachev (Ed.), Protista, 3. Guide Book on Zoology (pp. 270–307). St Peterburg-Moscow: KMK Scientific Press Ltd..
Frolov, A. O., Chystjakova, L. V., & Goodkov, A. V. (2004). A new pelobiont protist Pelomyxa corona sp. n. (Pelloflagellatea, Pelobiontida). Protistology, 3, 233–241.
Frolov, A. O., Chystjakova, L. V., & Goodkov, A. V. (2005a). Light- and electron-microscopic study of Pelomyxa binucleata (Gruber, 1884) (Peloflagellatea, Pelobiontida). Protistology, 4, 57–73.
Frolov, A. O., Chystjakova, L. V., Malysheva, M. N., & Goodkov, A. V. (2005b). Light and electron microscopic investigation of Pelomyxa prima (Grüber, 1884) (Peloflagellatea, Pelobiontea). Tsitologiya, 47, 89–98.
Frolov, A. O., Goodkov, A. V., Chystjakova, L. V., & Skarlato, S. O. (2006). Structure and development of Pelomyxa gruberi sp. n. (Peloflagellatea, Pelobiontida). Protistology, 4, 227–244.
Frolov, A. O., Chystjakova, L. V., Goodkov, A. V., & Malysheva, M. N. (2007). The morphological study of the cysts Pelomyxa palustris Greeff, 1874. Tsitologiya, 49, 642–651.
Frolov, A. O., Chystjakova, L. V., & Malysheva, M. N. (2011). Light and electron microscopic study of Pelomyxa flava sp. n. (Archamoebae, Pelobiontida). Cell and Tissue Biology, 5, 81–89.
Ghosh, S., Field, J. G., Rogers, R., Hickman, M., & Samuelson, J. (2000). The Entamoeba histolytica mitochondrion-derived organelle (crypton) contains double-stranded DNA and appears to be bound by a double membrane. Infection and Immunity, 68, 4319–4322.
Gill, E. E., Diaz-Triviño, S., et al. (2007). Novel mitochondrion-related organelles in the anaerobic amoeba Mastigamoeba balamuthi. Molecular Microbiology, 66, 1306–1320.
Goldschmidt, R. (1907a). Über die Lebensgeschichte der Mastigella vitrea n. sp. und Mastigina setosa n. sp. Archiv für Protistenkunde Supplement, 1, 83–165.
Goldschmidt, R. (1907b). Über die Lebensgeschichte der Mastigamöben. Sitzungsberichte der Gesellschaft für Morphologie und Physiologie in München, 23, 1–7.
Goodkov, A. V., Chystjakova, L. V., Seravin, L. N., & Frolov, A. O. (2004). The concept of pelobionts (class Peloflagellatea): A brief history and current status. Zoologischesky Zhurnal, 83, 643–654.
Goodkov A. V., & Seravin L. N. (1991). New ideas on the nature of the “giant amoeba” Pelomyxa palustris: the position of this organism in the system of lower eukaryotes (Peloflagellata classis n.). (In Russian) Zoologichesky Zhurnal, 70, 5–16.
Greeff, R. (1874). Pelomyxa palustris (Pelobius), ein amöbenartiger Organismus des süssen Wassers. Archiv für Mikroskopische Anatomie, 10, 51–73.
Griffin, J. L. (1979). Flagellar and other ultrastructure of Pelomyxa palustris, the giant herbivorous amoeboflagellate: More evidence for evolutionary distance from carnivores. Transactions of the American Microscopical Society, 98, 158.
Griffin, J. L. (1988). Fine structure and taxonomic position of the giant amoeboid flagellate Pelomyxa palustris. Journal of Protozoology, 35, 300–315.
Grüber, A. (1885). Studien über Amöben. Zeitschrift für Wissenschaftliche Zoologie, 41, 186–225.
Gutierrez, G. (2012). Draft genome sequence of Methanobacterium formicicum DSM 3637: An Archaebacterium isolated from the methane producer amoeba Pelomyxa palustris. Journal of Bacteriology, 194, 6967.
Hampl, V., & Simpson, A. G. B. (2008). Possible mitochondria-related organelles in poorly-studied “amitochondriate” eukaryotes. In: Tachezy J (ed) hydrogenosomes and mitosomes: Mitochondria of anaerobic eukaryotes. Microbial Monographs, 9, 265–282.
Hannaert, V., Brinkmann, H., et al. (2000). Enolase from Trypanosoma brucei, from the amitochondriate protist Mastigamoeba balamuthi, and from the chloroplast and cytosol of Euglena gracilis: Pieces in the evolutionary puzzle of the eukaryotic glycolytic pathway. Molecular Biology and Evolution, 17, 989–1000.
Haque, R., Huston, C. D. H., Hughes, M., Houpt, E., & Petri Jr., W. A. (2003). Amebiasis. New England Journal of Medicine, 348, 1565–1573.
Heiss, A. A., Walker, G., & Simpson, A. G. B. (2013). The flagellar apparatus of Breviata anathema, a eukaryote without a clear supergroup affinity. European Journal of Protistology, 49, 354–372.
Heredia, R. D., Fonseca, J. A., & López, M. C. (2012). Entamoeba moshkovskii perspectives of a new agent to be considered in the diagnosis of amebiasis. Acta Tropica, 123, 139–145.
Hinkle, G., Leipe, D. D., Nerad, T. A., & Sogin, M. L. (1994). The unusually long small subunit ribosomal RNA of Phreatamoeba balamuthi. Nucleic Acids Research, 22, 465–469.
Holder, M., & Lewis, P. O. (2003). Phylogeny estimation: Traditional and Bayesian approaches. Nature Reviews Genetics, 4, 275–284.
Hollande, A. (1945). Biologie et reproduction des rhizopodes des genres Amoeba et Pelomyxa et cycle évolutif de l’Amoebophilus destructor nov. gen. nov. sp., Chytridinée (?) parasite de Pelomyxa palustris Greeff. Bullétin Biologique de France et Belgique, 79, 31–66.
Ivanic, M. (1936). Zür Kentniss der Entwicklungsgeschichte bei Mastigina hylae Frenzel. Archiv für Protistenkunde, 87, 225–241.
Kent, W. S. (1880). A manual of the infusoria: Including a description of all known flagellate, ciliate, and tentaculiferous protozoa, British and foreign, and an account of the organization and the affinities of the sponges. London: D. Bogue.
Kirby, H. (1945). Entamoeba coli versus Endamoeba coli. Journal of Parasitology, 31, 177–184.
Klebs, G. (1892). Flagellatenstudien, Theil 1. Zeitschrift für Wissenschaftliche Zoologie, 55, 265–445.
Klug, G. (1936). Neue oder wenige bekannte Arten der Gattungen Mastigamoeba, Mastigella, Cercobodo, Tetramitus und Trigonomonas. (Studien über farblose Flagellaten I). Archiv für Protistenkunde, 87, 97–116.
Krylov, M. V., Dobrovol'skij, A. A., et al. (1980). New conceptions of the system of unicellular animals. Trudy Zoologicheskogo Instituta, Leningrad, 94, 122–132.
Kudo, R. R. (1939). Protozoology (2nd ed.). Springfield: Charles C. Thomas.
Kudo, R. R. (1950). A species of Pelomyxa from Illinois. Transactions of the American Microscopical Society, 69, 368–370.
Kudo, R. R. (1960). Protozoology (4th ed.). Springfield: Charles C. Thomas.
Kudo, R. R. (1977). Protozoology (5th ed.). Springfield: Charles C. Thomas.
Kudryavtsev, A., Bernhard, D., Schlegel, M., Chao, E. E. Y., & Cavalier-Smith, T. (2005). 18S ribosomal RNA gene sequences of Cochliopodium (Himatismenida) and the phylogeny of Amoebozoa. Protist, 156, 215–224.
Kuenen, W., & Swellengrebel, N. (1917). Korte beschrijving van enkele minder bekende protozoeo uit denmenschelijken darm. Nederlands Tijdschrift voor Geneeskunde, 1917, 496.
Lackey, J. B. (1923). The fauna of Imhoff tanks. New Jersey Agricultural Experiment Stations Bulletin, 417, 3–39.
Lackey, J. B. (1932). Oxygen deficiency and sewage protozoa: With descriptions of some new species. Biological Bulletin, 63, 287–295.
Larsen, J., & Patterson, D. J. (1990). Some flagellates (Protista) from tropical marine sediments. Journal of Natural History, 24, 801–937.
Lauterborn, R. (1901). Die sapropelische Lebewelt. Zoologischer Anzeiger, 24, 50–55.
Lee, W. J., & Patterson, D. J. (2000). Heterotrophic flagellates (Protista) from marine sediments of Botany Bay, Australia. Journal of Natural History, 34, 483–562.
Leidy, J. (1874). Notice of a remarkable amoeba. Proceedings of the Academy of Natural Sciences of Philadelphia, 1874, 142–143.
Leidy, J. (1879). Freshwater rhizopods of North America. Washington, DC: Government Printing Office.
Lemmermann, E. (1914). Flagellatae 1. In A. Pascher (Ed.), Die Süsswasser-flora Deutschlands, Österreichs und der Schweiz (pp. 1–138). Jena: Gustav Fischer Verlag.
León-Avila, G., & Tovar, J. (2004). Mitosomes of Entamoeba histolytica are abundant mitochondrion-related remnant organelles that lack a detectable organellar genome. Microbiology, 150, 1245–1250.
Lepşi, I. (1965). Protozoologie. Bucureşti: Editura Academiei Republicii Socialiste România.
Loeblich, A. R., & Tappan, H. (1961). Suprageneric classification of the Rhizopodea. Journal of Paleontology, 35, 245–330.
Loftus, B., Anderson, I., et al. (2005). The genome of the protist parasite Entamoeba histolytica. Nature, 433, 865–868.
Ludwig, F. W. (1946). Studies on the protozoan fauna of the larvae of the crane-fly, Tipula abdominalis. I. Flagellates, amoebae, and gregarines. Transactions of the American Microscopical Society, 65, 189–214.
Mackinnon, D. L. (1913). Studies on parasitic protozoa. II. (a) The encystment of Rhizomastix gracilis Alexeieff; (b) Tetratrichomastix parisii n. sub-gen., n. sp. Quarterly Journal of Microscopical Science, 59, 459–470.
Maguire, F., & Richards, T. A. (2014). Organelle evolution: A mosaic of ‘mitochondrial’ functions. Current Biology, 24, R518–R520.
Maralikova, B., Ali, V., et al. (2010). Bacterial-type oxygen detoxification and iron-sulfur cluster assembly in amoebal relict mitochondria. Cellular Microbiology, 12, 331–342.
Margulis, L. (1970). Origin of eukaryotic cells. New Haven: Yale University Press.
Martin, W., & Müller, M. (1998). The hydrogen hypothesis for the first eukaryote. Nature, 392, 37–41.
Martínez-Palomo, A. (1993). Parasitic amebas of the intestinal tract. In J. P. Kreier & J. R. Baker (Eds.), Parasitic Protozoa (Vol. 3, 2nd ed., pp. 65–141). San Diego: Academic Press.
Mi-ichi, F., Miyamoto, T., Takao, S., Jeelani, G., Hashimoto, T., Hara, H., Nozaki, T., & Yoshida, H. (2015). Entamoeba mitosomes play an important role in encystation by association with cholesteryl sulfate synthesis. Proceedings of the National Academy of Sciences of the USA, 112, E2884–E2890.
Milyutina, I. A., Aleshin, V. V., Mikrjukov, K. A., Kedrova, O. S., & Petrov, N. B. (2001). The unusually long small subunit ribosomal RNA gene found in amitochondriate amoeboflagellate Pelomyxa palustris: Its rRNA predicted secondary structure and phylogenetic implication. Gene, 272, 131–139.
Morin, L., & Mignot, J.-P. (1995). Are Archamoebae true archezoa? The phylogenetic position of Pelomyxa sp., as inferred from large subunit ribosomal RNA sequencing. European Journal of Protistology, 31, 160A.
Morris, S. (1936). Studies on Endamoeba blattae (Bütschli). Journal of Morphology, 59, 225–263.
Mowbrey, K., & Dacks, J. B. (2009). Evolution and diversity of the Golgi body. FEBS Letters, 583, 3738–3745.
Nikolaev, S. I., Berney, C., et al. (2006). Phylogenetic position of Multicilia marina and the evolution of Amoebozoa. International Journal of Systematic and Evolutionary Microbiology, 56, 1449–1458.
Nývltová, E., Šuták, R., Harant, K., Šedinová, M., Hrdý, M., Pačes, J., Vlček, Č., & Tachezy, J. (2013). NIF-type iron-sulfur cluster assembly system is duplilcated and distributed in the mitochondria and cytosol of Mastigamoeba balamuthi. Proceedings of the National Academy of Sciences of the USA, 110, 7371–7376.
Page, F. C. (1970). Mastigamoeba aspera from estuarine tidal pools in Maine. Transactions of the American Microscopical Society, 89, 197–200.
Page, F. C. (1976). A revised classification of the Gymnamoebia (Protozoa: Sarcodina). Zoological Journal of the Linnean Society, 58, 61–77.
Page, F. C. (1983). Marine Gymnamoebae. National Environmental Research Council. Lavenham: The Lavenham Press Ltd.
Page, F. C. (1987). The classification of ‘naked’ amoebae (Phylum Rhizopoda). Archiv für Protistenkunde, 133, 199–217.
Pánek, T., Zadrobílková, E., Walker, G., Brown, M. W., Gentekaki, E., Hroudová, M., Kang, S., Roger, A., Tice, A. K., Vlček, Č., & Čepička, I. (2016). First multigene analysis of Archamoebae (Amoebozoa: Conosa) robustly reveals its phylogeny and shows that Entamoebidae represents a deep lineage of the group. Molecular Phylogeny and Evolution, 98, 41–51.
Patterson, D. J. (1994). Protozoa: Evolution and systematics. In K. Hausmann & N. Hülsmann (Eds.), Progress in protozoology. Proceedings of the IX International Congress of Protozoology, Berlin 1993 (pp. 1–14). Stuttgart: Gustav Fischer Verlag.
Patterson, D. J., & Sogin, M. L. (1992). Eukaryote origins and protistan diversity. In H. Hartman & K. Matsuno (Eds.), The origin of prokaryotic and eukaryotic cells (pp. 13–46). Singapore: World Scientific.
Patterson, D. J., Simpson, A. G. B., & Rogerson, A. (2000). Amoebae of uncertain affinities. In J. J. Lee, G. F. Leedale, & P. Bradbury (Eds.), An Illustrated guide to the protozoa (2nd ed., pp. 804–827). Lawrence: Society of Protozoologists.
Penard, E. (1902). Faune rhizopodique du Bassin du Leman. Geneva: Henry Kündig, Librarie de l’Institut.
Penard, E. (1909). Sur quelques mastigamibes des environs de Genève. Révue Suisse de Zoologie, 17, 405–439.
Penard, E. (1936). Rhizopode ou flagellate? Quelques réflexions à propos de la Dinamoeba mirabilis. Bulletin de la Societé Française de Microbiologie de Paris, 5, 154–163.
Philippe, H., & Germot, A. (2000). Phylogeny of eukaryotes based on ribosomal RNA: Long-branch attraction and models of sequence evolution. Molecular Biology and Evolution, 17, 830–834.
Poulsen, C. S., & Stensvold, C. R. (2016). Systematic review on Endolimax nana: A less well studied intestinal ameba. Tropical Parasitology, 6, 8–29.
Preisig, H. R. (1994). Siliceous structures and silicification in flagellated protists. Protoplasma, 181, 29–42.
Ptáčková, E., Kostygov, A. Y., Chystjakova, L. V., Falteisek, L., Frolov, A. O., Patterson, D. J., Walker, G., & Čepička, I. (2013). Evolution of Archamoebae: Morphological and molecular evidence for pelobionts including Rhizomastix, Entamoeba, Iodamoeba and Endolimax. Protist, 164, 380–410.
Reavill, D. R., & Schmidt, R. E. (2010). Urinary tract diseases of reptiles. Journal of Exotic Pet Medicine, 19, 280–289.
Reichenow, E. (1952). Lehrbuch der Protozoenkunde (Begründet von Franz Doflein, Sechste auflage). Zweiter teil: Spezielle naturgeschichte der protozoen (pp. 411–776). Jena: Gustav Fischer.
Rhumbler, L. (1905). Zür theorie der Öberflachenkrafe der Amöben. Zeitschrift für Wissenschaftliche Zoologie, 83, 1–52.
Roger, A. (1999). Reconstructing early events in eukaryotic evolution. American Naturalist, 154, S146–S163.
Roger, A. J., Clark, C. G., & Doolittle, W. F. (1996). A possible mitochondrial gene in the early-branching amitochondriate protist Trichomonas vaginalis. Proceedings of the National Academy of Sciences of the USA, 93, 14618–14622.
Roger, A. J., Svard, S. G., et al. (1998). A mitochondrial-like chaperonin 60 gene in Giardia lamblia: Evidence that diplomonads once harbored an endosymbiont related to the progenitor of mitochondria. Proceedings of the National Academy of Sciences of the USA, 95, 229–234.
Rosenbaum, R. M., & Wittner, M. (1970). Ultrastructure of bacterized and axenic trophozoites of Entamoeba histolytica with particular reference to helical bodies. Journal of Cell Biology, 45, 367–382.
Samarawickrema, N. A., Upcroft, J. A., Thammapalerd, N., & Upcroft, P. (2001). A rapid-cooling method for cryopreserving Entamoeba histolytica. Annals of Tropical Medicine and Parasitology, 95, 853–855.
Samuelson, J., & Robbins, P. (2011). A simple fibril and lectin model for cyst walls of Entamoeba and perhaps Giardia. Trends in Parasitology, 27, 17–22.
Sassuchin, D. (1928). Zür Frage über die ecto- und endoparasitischen Protozoen der Frosch Kaulquappen. Archiv für Protistenkunde, 64, 71.
Schirch, P. (1914). Beitrage zur Kenntniss des Lebenscyclus von Arcella vulgaris Ehrbg. und Pelomyxa palustris Greeff. Archiv für Protistenkunde, 33, 257–273.
Schoudeten, H. (1907). Notes sur quelques Flagellés. Archiv für Protistenkunde, 9, 108–136.
Schulze, F. E. (1875a). Rhizopodenstudien IV. Archiv für Mikroskopische Anatomie und Entwicklungsmechanik, 11, 329–353.
Schulze, F. E. (1875b). Rhizopdenstudien V. Archiv für mikroskopische Anatomie und Entwicklungsmechanik, 11, 583–593.
Schuster, F. L. (1990). Phylum Rhizopoda. In L. Margulis, J. O. Corliss, M. Melkonian, & D. J. Chapman (Eds.), Handbook of Protoctista (pp. 3–18). Boston: Jones and Bartlett.
Seckt, H. (1922). Estudios hidrobiologicos en la Argentina: Flagellatae. Boletin Academia Nacional Ciencias Cordoba, 25, 430–490.
Seravin, L. N., & Goodkov, A. V. (1987a). The flagella of the freshwater amoeba Pelomyxa palustris. Tsitologiya, 29, 721–725.
Seravin, L. N., & Goodkov, A. V. (1987b). Golgi apparatus in the amoeba Pelomyxa palustris. Doklady Akademii Nauk SSSR, 296, 249–250.
Seravin, L. N., & Goodkov, A. V. (1987c). Cytoplasmic microbody-like granules of the amoeba Pelomyxa palustris. Tsitologiya, 29, 600–603.
Seravin, L. N., & Goodkov, A. V. (1987d). Peculiarities of the cytoskeleton organization of the amoeba Pelomyxa palustris. Tsitologiya, 29, 835–837.
Shiratori, T., & Ishida, K.-I. (2016). Entamoeba marina n. sp.; a new species of Entamoeba isolated from tidal flat sediment of Iriomote Island, Okinawa, Japan. Journal of Eukaryotic Microbiology, 63, 280–286.
Siemensma, F. J. (1987). De Nederlandse Naaktamoeben (Rhizopoda, Gymnamoebia). Wetenschappelijke mededelingen Koninklijke Nederlandse Natuurhistorische Vereniging, 181, 1–136.
Silberman, J. D., Clark, C. G., Diamond, L. S., & Sogin, M. L. (1999). Phylogeny of the genera Entamoeba and Endolimax as deduced from small-subunit ribosomal RNA sequences. Molecular Biology and Evolution, 16, 1740–1739.
Simpson, A. G. B., & Roger, A. J. (2004). The real ‘kingdoms’ of eukaryotes. Current Biology, 14, R693–R696.
Simpson, A. G. B., Bernard, C., Fenchel, T., & Patterson, D. J. (1997). The organisation of Mastigamoeba schizophrenia n. sp.: More evidence of ultrastructural idiosyncrasy and simplicity in pelobiont protists. European Journal of Protistology, 33, 87–98.
Skibbe, O., & Zölffel, M. (1991). Der Protisten-Steckbrief: die begeisselte Amöbe Mastigina chlamys. Mikrokosmos, 80, 129–133.
Skvortzkov, B. V., & Noda, M. (1976). Flagellates of clean and polluted waters. V. A short description of colourless flagellates with one swimming flagellum – Genus Mastigella (Frenzel) nob. (Rhizomastigaceae) reported during 1962–1975 in many parts of the world and especially from subtropics of Brasil, South America. In Prof Mitsuko Noda Commemorative Publication on his retirement from the Faculty of Science. Niigata University. Laboratory of Phytotaxo-Morphology, Faculty of Science, Niigata University.
Sogin, M. L. (1991). Early evolution and the origin of eukaryotes. Current Opinion in Genetics and Development, 1, 457–463.
Sogin, M. L., & Silberman, J. D. (1998). Evolution of the protists and protistan parasites from the perspective of molecular systematics. International Journal of Parasitology, 28, 11–20.
Solis, F. J., & Barrios, R. (1991). Entamoeba histolytica: Microtubule movement during mitosis. Experimental Parasitology, 73, 276–284.
Stairs, C. W., Roger, A. J., & Hampl, V. (2011). Eukaryotic pyruvate formate lyase and its activating enzyme were acquired laterally from a firmicute. Molecular Biology and Evolution, 28, 2087–2099.
Stamatakis A. (2006). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics, 22, 2688–90.
Stensvold, C. R., Lebbad, M., & Clark, C. G. (2012). Last of the human protists: The phylogeny and genetic diversity of Iodamoeba. Molecular Biology and Evolution, 29, 39–42.
Stiller, J. W., & Hall, B. D. (1999). Long-branch attraction and the rDNA model of early eukaryotic evolution. Molecular Biology and Evolution, 16, 1270–1279.
Stokes, A. C. (1886). Notices of new fresh-water infusoria. Proceedings of the American Philosophical Society, 23, 562–568.
Stokes, A. C. (1888). A preliminary contribution toward a history of the fresh-water infusoria of the United States. Journal of the Trenton Natural History Society, 1, 73–319.
Stokes, A. C. (1890). Notices of new fresh-water infusoria. Proceedings of the American Philosophical Society, 28, 74–80.
Thompson, R. C. A., & Smith, A. (2011). Zoonotic enteric protozoa. Veterinary Parasitology, 182, 70–78.
Tovar, J., Fischer, A., & Clark, C. G. (1999). The mitosome, a novel organelle related to mitochondria in the amitochondrial parasite Entamoeba histolytica. Molecular Microbiology, 32, 1013–1021.
Tovar, J., León-Avila, G., et al. (2003). Mitochondrial remnant organelles of Giardia function in iron-sulphur protein maturation. Nature, 426, 172–176.
van Bruggen, J. J. A., Stumm, C. K., & Vogels, G. D. (1983). Symbiosis of methanogenic bacteria and sapropelic protozoa. Archives of Microbiology, 136, 89–96.
van Bruggen, J. J. A., Stumm, C. K., Zwart, K. B., & Vogels, G. D. (1985). Endosymbiotic methanogenic bacteria of the sapropelic amoeba Mastigella. FEMS Microbiology and Ecology, 31, 187–192.
van Bruggen, J. J. A., van Rens, G. L. M., Geertman, E. J. M., Zwart, K. B., Stumm, C. K., & Vogels, G. D. (1988). Isolation of a methanogenic endosymbiont of the sapropelic amoeba Pelomyxa palustris Greef. Journal of Protozoology, 35, 20–23.
van der Giezen, M. (2009). Hydrogenosomes and mitosomes: Conservation and evolution of functions. Journal of Eukaryotic Microbiology, 56, 221–231.
van der Giezen, M., Tovar, J., & Clark, C. G. (2005). Mitochondrion-derived organelles in protists and fungi. International Review of Cytology, 244, 177–227.
Van Hoek, A. H. A. M., van Alen, T. A., et al. (2000). Multiple acquisition of methanogenic archaeal symbionts by anaerobic ciliates. Molecular Biology and Evolution, 17, 251–258.
Walker, G., Simpson, A. G. B., Edgcomb, V., Sogin, M., & Patterson, D. J. (2001). Ultrastructural identities of Mastigamoeba punctachora, Mastigamoeba simplex and Mastigella commutans, and assessment of hypotheses of relatedness of the pelobionts (Protista). European Journal of Protistology, 37, 25–49.
Walker, G., Silberman, J. D., et al. (2003). An ultrastructural and molecular study of Hyperamoeba dachnaya, n. sp., and its relationship to the mycetozoan slime moulds. European Journal of Protistology, 39, 319–336.
Wenyon, C. M. (1926). Protozoology. A manual for medical men, veterinarians and zoologists (Vol. 1). Ballière: William Wood and Company.
Whatley, J. M. (1976). Bacteria and nuclei in Pelomyxa palustris: Comments on the theory of serial endosymbiosis. New Phytologist, 76, 111–120.
Whatley, J. M., & Chapman-Andresen, C. (1990). Phylum Karyoblastea. In L. Margulis, J. O. Corliss, M. Melkonian, & D. J. Chapman (Eds.), Handbook of protoctista (pp. 167–185). Boston: Jones and Bartlett.
Wickerham, L. J., & Page, F. C. (1970). Cultivation and lyophilization of Mastigina sp. Journal of Protozoology, 17, 518–520.
Zadrobílková, E., Walker, G., & Čepička, I. (2015). Morphological and molecular evidence support a close relationship between the free-living archamoebae Mastigella and Pelomyxa. Protist, 166, 14–41.
Zadrobílková, E., Smejkalová, P., Walker, G., & Čepička, I. (2016). Morphological and molecular diversity of the neglected genus Rhizomastix Alexeieff, 1911 (Amoebozoa: Archamoebae) with description of five new species. Journal of Eukaryotic Microbiology, 63, 181–197.
Zaman, V., Howe, J., & Ng, M. (1998). Ultrastructure of the nucleus of the Iodamoeba bütschlii cyst. Parasitology Research, 84, 421–422.
Zaman, V., Howe, J., Ng, M., & Goh, T. K. (2000). Ultrastructure of the Endolimax nana cyst. Parasitology Research, 86, 54–56.
Žárský, V. (2012). Protein import into mitochondria and peroxisomes of parasitic protists. Diploma Thesis, Department of Parasitology, Faculty of Science, Charles University, Prague.
Žárský, V., & Tachezy, J. (2015). Evolutionary loss of peroxisomes – Not limited to parasites. Biology Direct, 10, 74.
Acknowledgments
We gratefully acknowledge the input from Professor David J. Patterson on knowledge of free-living taxa and the input on knowledge of parasitic taxa from Professor RNDr. Jaroslav Kulda, who also provided the prepared slide material in the plates. This work was supported by a grant from the Czech Science Foundation (project GA14-14105S).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this entry
Cite this entry
Walker, G., Zadrobílková, E., Čepička, I. (2017). Archamoebae. In: Archibald, J., Simpson, A., Slamovits, C. (eds) Handbook of the Protists. Springer, Cham. https://doi.org/10.1007/978-3-319-28149-0_11
Download citation
DOI: https://doi.org/10.1007/978-3-319-28149-0_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-28147-6
Online ISBN: 978-3-319-28149-0
eBook Packages: Biomedical and Life SciencesReference Module Biomedical and Life Sciences