
Abstract
Iron is essential for algal growth, metabolism and function. Numerous proteins and enzymes require iron, including those involved in photosynthesis, respiration, as well as nitrogen assimilation and fixation. A variety of sources and sinks of iron in marine and freshwater ecosystems lead to a wide range of dissolved iron concentrations, from iron-limiting to iron-replete, for microalgal growth. Decades of physiological and molecular research, in combination with recent genomic advances, have made way for breakthroughs in our understanding of the critical roles iron plays in microalgal metabolism and how iron is acquired in aqueous environments. Herein, we review and integrate these studies to compare and contrast the iron requirements, acquisition abilities and strategies, as well as storage capacities of microalgae, with a particular emphasis on diatoms, green algae and cyanobacteria. These include the ability to perform substitution or the permanent replacement of iron-requiring proteins for non-iron containing functional equivalents, the presence of a high-affinity uptake system and the capacity to store iron in excess of cellular demand. We also include a discussion of iron-related topics of current significance, including iron-light co-limitation, effects of iron limitation on cellular elemental composition, large-scale iron fertilization experiments, and climate change effects on iron bioavailability. Lastly, a brief overview of some common laboratory and field techniques employed to study microalgal iron physiology is provided.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
FormalPara PrefaceNo other trace element has received more attention in relation to influences on microalgal physiology than iron (Fe). At the time of writing, a Web of Knowledge search using keywords “iron” and “phytoplankton” listed approximately 6800 references since the last edition of this textbook in 1974. To summarize all of these studies would be next to impossible. Rather, our goal in this chapter is to provide a comprehensive overview of key findings related to the acquisition and function of Fe within phytoplankton cells, emphasizing what we now know about how Fe is obtained, the important physiological roles of Fe and the ways in which microalgae have evolved to cope with widespread Fe limitation in aquatic environments. In addition, we have attempted to integrate a new wealth of information obtained through genomic approaches that are now common practice in physiological research with phytoplankton. Where possible, we provide reference to extensive literature reviews that will ultimately offer a more in-depth discussion on specific Fe-related topics. Although we have made efforts to include information on all phytoplankton functional groups, including some discussion of cyanobacteria, our expertise lie in diatom physiology, so there is undoubtedly a preferential focus on this group of microalgae. That being said, the vast majority of microalgal physiology studies within the scientific literature relating to transport and physiological adaptations to Fe limitation are focused on diatoms (and to some extent, freshwater green algae), as this group appears to be most affected by low Fe concentrations in the ocean and often exhibits the largest physiological response to both natural and anthropogenic Fe enrichments.
1 Iron Sources and Distributions in Aquatic Environments
A review on Fe physiology of phytoplankton would not be complete without first describing the sources of Fe and how Fe is distributed throughout aquatic environments. Varying Fe concentrations in aquatic systems through space and time have largely shaped the evolutionary trajectories of many phytoplankton groups (Falkowski et al. 2004). Given that phytoplankton communities in an estimated 30–40 % of the oceans are chronically deprived of Fe (Moore et al. 2002), extensive efforts have been made to obtain high-resolution measurements of Fe concentrations in the sea. As such, we will primarily focus on Fe distributions in seawater, although there is growing evidence that Fe may also be an important regulator of phytoplankton growth in freshwater environments (Twiss et al. 2000; Sterner et al. 2004; Havens et al. 2012; Shaked and Lis 2012).
Iron exists in seawater in two oxidation states: soluble Fe (II) and sparingly soluble Fe (III), which has had a profound influence on iron concentrations in the ocean over geologic times. Since the first appearance of oxygen-evolving photosynthetic organisms ca. 3 billion years ago, alternating episodes of anoxic and oxic conditions in the oceans are evident throughout Earth’s geological record with oxic conditions only prevailing since the start of the Phanerozoic era (~540 mya). Increased oxygen concentrations in seawater resulted in a massive decline in soluble Fe concentrations through the formation of insoluble Fe oxides that are rapidly precipitated and removed from the water column (Anbar and Knoll 2002). The end result is that Fe requirements in microalgae remain quite high due to evolutionary constraints relative to the supply, both of which being constrained by the chemistry of Fe. Dissolved Fe in contemporary oceans is typically present at concentrations of 0.02–1 nanomolar (10−9) concentration range. In large areas of the ocean, complexation and speciation of this Fe results in bioavailable concentrations that are far below those needed to support maximum growth rates of many microalgal species (Wells et al. 1995) and have been influential in setting the maximum growth rates in many species (Sunda and Huntsman 1997, 2015).
Restricted availability of Fe to microalgae is largely a consequence of its complex chemistry. Iron is present in the ocean as inorganic soluble and insoluble chemical species, dissolved organic complexes, colloidal material, mineral particles and, of course, as components of living cells. The most stable form of Fe within oxygenated seawater is the oxidized ferric (Fe(III)) state. Dissolved forms of Fe(III) primarily exist as organic complexes with a very minimal amount existing as dissolved inorganic hydrolysis species (Rue and Bruland 1995). It is believed that these dissolved forms supply a significant proportion of the Fe flux to the cell. Ingestion of bacteria or colloidal (i.e. insoluble) forms may serve as other pathways for obtaining Fe for many groups of phytoplankton with the notable exception of diatoms, which unlike most microalgal groups are incapable of phagotrophy (Maranger et al. 1998; Nishioka and Takeda 2000). Most oceanic Fe-containing colloids are believed to consist of Fe in an organic matrix. Inorganic Fe oxyhydroxide mineral colloids appear to be rare in the open sea (Wells and Goldberg 1994). Indirectly, colloidal Fe may be accessible to phytoplankton by thermal, photochemical and possibly biological reduction (Hutchins 1995).
Iron inputs vary among regions and their proximity to land. Sources of Fe to marine euphotic zone waters include terrestrial run-off, atmospheric dust (dry deposition) and precipitation (wet deposition), upwelling of deep waters, and anthropogenic input. Such sources are crucial in determining Fe availability due to its high reactivity and low solubility (Johnson et al. 1997). Throughout the water column, Fe often exhibits a nutrient-like profile as it is significantly involved with the internal cycling of biologically derived particulate material, although in regions with high aeolian input Fe concentrations may be elevated at the surface (Bruland et al. 1994) (Fig. 1). More often, concentrations tend to be lowest in the euphotic zone due to rapid assimilation by phytoplankton and/or adsorption onto biogenic particles and then increase in the subsurface waters as sinking particles succumb to decomposition and dissolution. The residence time of Fe in seawater is relatively short (200–500 years) compared to other elements, eventually being removed from the water column through biological uptake and scavenging onto sinking particles (Bruland et al. 1994; Johnson et al. 1997).

Distributions of Fe in the world ocean. (a) Mean profiles of dissolved iron in the North Atlantic, North Pacific, and Southern Ocean averaged over the depth intervals: 0–100 m, 100–250 m, 250–500 m, 500–1000 m, 1000–1500 m, 1500–2000 m, 2000–3000 m, 3000–4000 m, and 4000–5000 m, (b) Observed Fe concentrations from depths >1000 m in the eastern subtropical Pacific Ocean (20–50°N) plotted as a function of distance from the continental land mass and (c) Modeled estimates of the two most important sources of Fe to the world ocean: mineral dust (24 × 109 mol Fe) and shallow (<502 m) sediments (21 × 109 mol Fe) (Figures are reproduced from Moore and Braucher (2008). Panel c is courtesy of Bruland et al. (2014))
Iron is the fourth most abundant element in the Earth’s crust. Thus, the main inputs of new Fe to the ocean are from continental sources and, therefore, regions in close proximity to land or those that experience a high degree of upwelling of deep, nutrient rich waters tend to have higher Fe concentrations compared to open ocean regions that are far away from land (Fig. 1). Sources of Fe to surface waters vary depending on geographic location. Dominant inputs of Fe to coastal regions are river runoff and benthic inputs. Rivers are a primary source for most of the major ions in seawater, as well as trace metals such as Fe. Although Fe concentrations in rivers may be quite high, most of this Fe is likely removed by flocculation of strongly associated Fe-humic substances and may not make its way out of the estuary (Boyle et al. 1976). However, there are some measurements that suggest that riverine Fe inputs to the ocean can be quite high, constituting a significant portion of new Fe inputs to the adjacent open ocean (Wetz et al. 2006; Klunder et al. 2012).
Benthic inputs of Fe to the overlying water column are primarily from two different sources, the input of continental derived material that is released into the dissolved phase and biogenic material that is exported from the surface layer. For example, off the Oregon coast, the dominant form of Fe to surface waters is a combination of reduced Fe(II) from shelf sediments and sediment that is resuspended through upwelling (Johnson et al. 1999; Chase et al. 2007; Bruland et al. 2008; Lohan and Bruland 2008). The addition of dissolved Fe(II) to oxic seawater will quickly result in its oxidation to less-soluble Fe(III) with subsequent formation of colloidal and particular oxyhydroxide forms that are not readily biologically available. However the complexation of dissolved Fe(III) by strong Fe-binding ligands can maintain Fe(III) in the dissolved phase (see further details on siderophores below). Moore and Braucher (2008) modeled the global input of Fe from sediments and have shown the sedimentary sources along continental margins have a strong impact on open-ocean Fe concentrations, particularly in the Arctic and North Pacific and are equivalent to global estimates of Fe input from atmospheric dust (Fig. 1).
The primary source of Fe to surface waters in remote open ocean regions is through atmospheric deposition of dust from arid regions and anthropogenic emissions from the continents. Atmospheric inputs into the ocean are often spatially and temporally patchy with regions of particularly high atmospheric Fe input within the tropical and North Atlantic, Bay of Bengal, the Arabian Sea, and waters surrounding Australia and the southeastern continental margins of South America. Mineral aerosols consist of dust particles that are lifted into the atmosphere when high winds occur over arid erodible surfaces. These particles may be transported long distances before they are deposited into the ocean via settling, turbulent deposition and precipitation processes (Mahowald et al. 2005). The solubility of Fe in these various atmospheric forms depends on the source, with soluble Fe fractions ranging from 0.01 to 80 % (Mahowald et al. 2005).
Hydrothermal vents are considered another significant source of Fe to the ocean. In addition to emitting sulfur, high levels of trace metals, including Fe, are emitted by vents. Previously, vent-derived Fe was thought to be of limited importance due to rapid oxidation and precipitation around the vents. Yet it is now believed that hydrothermal vent inputs of Fe may in fact be long-lasting, contributing to the dissolved pool of Fe and eventually making their way to sunlit surface waters where they fuel primary productivity (Saito et al. 2013; Conway and John 2014). Stabilization of dissolved and particulate Fe in hydrothermal plumes is likely achieved through association with organic ligands. Modeling and observation-based studies have recently suggested hydrothermal vents may contribute substantially to overall Fe inputs into the ocean, likely being important in buffering the ocean’s dissolved Fe inventory against shorter-term fluctuations in dust deposition. Recent estimates suggest hydrothermal input could provide 9–22 % of the Fe budget in the global deep ocean and upwards of 45–50 % in the tropical Pacific (Tagliabue et al. 2010).
In polar regions, glaciers may also be an appreciable source of Fe to the ocean. Glaciers can be considered as frozen rivers, and when they reach the ocean, they discharge in the form of icebergs and water. Icebergs often will transport significant amounts of continental debris that will be released into the ocean as they melt. In addition, sea ice can act as a cap on the ocean, which accumulates atmospheric sources of Fe and releases the Fe as it melts. Sea ice is believed to be an important mechanism for Fe delivery in high-latitude regions of the Southern and Arctic Oceans. The amount of bioavailable Fe supplied to the Southern Ocean by both aeolian dust and icebergs have been estimated to be similar (Raiswell et al. 2008). Free-drifting icebergs around Antarctica are often “hotspots” of enhanced biological production and transport of organic carbon to the deep sea (Smith et al. 2007). This is likely a consequence of melting ice being a source of Fe that stimulates phytoplankton growth, although estimates on the contributions of Fe by icebergs to such regions are variable (Lin et al. 2011). The importance of melting ice as a source of Fe will likely increase as global temperatures continue to rise.
As in seawater, the Fe speciation in freshwater systems is highly affected by chemical variables (e.g. pH and O2 concentrations), Fe inputs and removal processes, as well as internal recycling. In natural conditions, Fe is supplied from the products of weathered rocks and soil around the watersheds and is controlled by a number of factors including geological processes, soil composition, environmental temperature, precipitation and hydrology (Harris 1992). In addition to these sources, anthropogenic influences such as wastewater and storm-discharge are a major supply of Fe to freshwater lakes. Iron in freshwater environments is commonly orders of magnitude higher than that of seawater, although dissolved Fe concentrations have been suggested to influence rates of NO3 − drawdown and Chl-a concentrations in some lakes (Havens et al. 2012). Iron may also play a regulatory role in the phytoplankton community where cyanobacteria tend be favored over green algae under high Fe conditions (Morton and Lee 1974; Pollingher et al. 1995; Hyenstrand et al. 2000). Similarly, Fe has been suggested to influence diatom composition in streams across the continental USA. Passy (2010) identified a relationship between stream diatom richness and wetland cover, where wetlands were suggested to provide an essential source of bioavailable, dissolved organic-bound Fe to stream ecosystems. More commonly, high Fe concentrations in freshwater systems, particularly rivers and streams, are a problem due to increased loads of Fe resulting from human activities such as mining of Fe enriched ores, intensified forestry, peat production and agricultural drainage.
2 Cellular Iron Requirements and Associated Physiological Processes
The transition metal iron exists in the environment in two oxidation states: Fe(III), the thermodynamically stable state in the presence of dioxygen and the reduced form Fe(II). Its facile redox chemistry has made Fe an essential metal for virtually all organisms, but it is particularly important for oxygenic photoautotrophs due to its essential role in photosynthetic and respiratory electron transport. Iron present in haeme or in the form of Fe-sulfur clusters occurs in a variety of metalloproteins that are involved in a myriad of metabolic pathways (Table 1).
Iron is particularly important in oxygenic photosynthesis. It is present in the two photosynthetic reaction centers (photosystems I and II, which contain respectively 12 and 2–3 iron atoms each) and the cytochrome b6f complex (5 Fe), which is involved in ATP synthesis (Raven 1988; Strzepek and Harrison 2004). It also occurs in two smaller proteins within the photosynthetic electron transport chain: cytochrome c 6 (1 Fe) and ferredoxin (2 Fe) (Raven 1990). Based on iron use calculations (Raven 1988, 1990) and empirical data (Strzepek and Harrison 2004), 50–90 % of the metabolic Fe within phytoplankton occurs within the photosynthetic apparatus.
Phytoplankton experiencing Fe stress have reduced photosynthetic pigments and pigment binding proteins when compared to Fe-replete conditions (Greene et al. 1992; Sunda and Huntsman 1995; Geider and LaRoche 1994). Known as chlorosis, reduced chlorophyll content along with a decline in photosynthetic efficiency and reduced growth rates are the most noticeable symptom of Fe-deficiency in algae, (reviewed by Behrenfeld and Milligan 2013). Protein (Glover 1977; Rueter and Ades 1987; Doucette and Harrison 1991), lipid and carbohydrate (Milligan and Harrison 2000; van Oijen et al. 2004) content are also observed to vary with Fe nutritional state. A reduction in chlorophyll synthesis is likely the result of both a decrease in the number of photosynthetic units due to insufficient iron for their synthesis (Raven 1990; Sunda and Huntsman 1997) and a lower abundance of Fe-containing enzymes involved in chlorophyll synthesis (Behrenfeld and Milligan 2013). Due to iron’s scarce supply to the oceans, microalgae have evolved unique strategies to reduce their photosynthetic Fe demand, a number of which are described below and have also recently been reviewed by Behrenfeld and Milligan (2013).
Iron also plays a large role in the reduction of oxidized chemical species of nitrogen, another essential element for cell growth. Phytoplankton require Fe for the assimilation of inorganic N species such as nitrate (NO3), nitrite (NO2) and nitrogen gas (N2) into ammonium (NH4). The assimilatory enzymes nitrate reductase (NR) and nitrite reductase (NiR) contain Fe (1 Fe: 1NR and 5–6 Fe: 1NiR). Under Fe limitation in the diatom Conticribra (Thalassiosira) weissflogii Footnote 1, the activity of NiR decreased by 50-fold and NO2 was excreted because NO2 assimilation was the rate-limiting step in the NO3 assimilation pathway (Milligan and Harrison 2000). Based on biochemical calculations, cells growing on NO3 should require 1.6 times more Fe than cells supplied with NH4 due to the Fe demand associated with NO3 reduction (Raven 1988; Morel et al. 1991). Experimental evidence supports this prediction since iron use efficiencies were, on average, 1.7 times higher for cells grown on NH4 than those grown on NO3 (Maldonado and Price 1996). Indirect support linking NH4 use to a reduced iron requirement for growth was also provided for the cyanobacterium, Synechococcus, where half saturation constants of Fe for growth (the Fe concentration that decreases growth rate by half) were lower for NH4 then NO3- grown cells (Kudo and Harrison 1997). However, neither field nor laboratory experiments have observed significantly faster growth rates of Fe-limited eukaryotic phytoplankton when grown on NH4 instead of NO3 (Price et al. 1991, 1994; Maldonado and Price 1996). The lack of a N-dependent effect may, in part, be due to the profound influence of Fe limitation on photosynthesis. Conversely, Fe-stressed, low-light, NO3-grown Emiliania huxleyi, a coccolithophore, grew significantly faster than Fe-stressed NH4-grown cells (Muggli and Harrison 1996b). The difference in growth rates was attributed to the reduction in cell volume by NO3-grown cells (see discussion below about Fe effects on cell size). Similarly, studies have shown that Fe-limited, NO3-grown diatoms grew about 25 % faster than Fe-limited, NH4-grown cells despite the higher iron use efficiency for cells grown on NH4 (Price 2005). The higher growth for the iron limited NO3-grown cells was due to a higher cellular Fe uptake rate, perhaps linked to increased rates of iron reduction at the cell surface by inducible transplasmalemma NO3 reductases (Maldonado and Price 2000).
The ability to fix nitrogen also imparts a high Fe demand on diazotrophs, many of which are cyanobacteria (e.g. Trichodesmium) (Kustka et al. 2003; Berman-Frank et al. 2007). This is because the enzyme that catalyzes the conversion of N2 gas to biologically available ammonium requires a large amount of Fe (38 Fe per enzyme). In addition N2 fixation requires a large amount of ATP and reducing equivalents (NADPH) (16 and 8 mol, respectively, per N2 molecule) which ultimately must be produced directly or indirectly from photosynthesis, further increasing the iron demand for the process (Kustka et al. 2003). Because of this increased Fe demand, the rate of N fixation by diazotrophs and thus the supply of fixed nitrogen to support phytoplankton growth, has been shown to be Fe-limited in many regions of the ocean, even when the growth rate of non-diazotrophs may not be directly limited by Fe (Moore et al. 2009; Sohm et al. 2011).
Other examples of cellular processes known to rely on Fe-containing enzymes and proteins include cellular respiration, vitamin synthesis, dehydrogenases and oxygenase reactions involved in fatty acid metabolism and detoxification of reactive oxygen species (e.g., hydrogen peroxide and super oxide radicals) (Table 1). Superoxide detoxification is catalyzed by superoxide dismutases (SODs) which catalyze the conversion of superoxide radicals to molecular oxygen and hydrogen peroxide. Four major groups of SODs are known and are distinguished by their metal co-factors: Fe, Mn, Cu/Zn, and Ni. The Fe/Mn family includes both single metal-binding and the rare cambialistic SODs which can bind either Mn or Fe. The Fe and Mn single metal-binding SODs are very similar but may be distinguished by two critical residues involved in the binding of the metal ion (Wolfe-Simon et al. 2005). Although many microalgae contain the Fe-SOD variants, interestingly, others completely lack Fe-SODs or down-regulate them when grown under low Fe conditions, which may thus contribute to lowering their cellular Fe demands (Allen et al. 2008; Marchetti et al. 2012).
The cellular Fe requirement is often expressed by the intracellular Fe content (or quota; Q) of a microalgal cell relative to its cellular C (or P) content. The cellular quota may be given as a ratio (Fe:C/P), which allows for a measure of Fe demand normalized to biomass and provides a means of comparison among different phytoplankton species of varying sizes and shapes. Recently, Quigg et al. (2003, 2011) suggested microalgal groups that inherited their plastids by endosymbiotic events from the green lineage tended to have higher trace metal:P ratios than those that derived their plastids through the red lineage. This suggested that this may be the result of the lineages originating in contrasting environmental conditions; the green lineage dominating under more reduced ocean conditions during earlier geologic times and the red lineage dominating later under more oxic conditions that are more similar to present day marine environments. To examine if this is also true specifically for Fe, we compiled published Fe:C (and Fe:P) ratios measured in various phytoplankton isolates and grouped them according to taxa. Comparing Fe quotas in phytoplankton grown under Fe-replete conditions is confounded by luxury consumption of Fe in certain groups of microalgae in excess of the cell quotas needed to support maximum growth rates. This is especially true for diatoms where large but variable levels of luxury consumption are observed (see further discussion below). Therefore, we limited our data set to only those Fe:C ratios obtained when cells were grown under conditions of inorganic Fe concentrations that supported maximum or near maximum (μ/μmax >75 %) growth rates, but were below levels that would result in a large amount of luxury Fe uptake.
Cyanobacteria, especially N2-fixing species, and microalgae from the red primary plastid lineage (red algae) have relatively higher Fe requirements than the green algae and secondary red plastid lineage groups (e.g. diatoms, dinoflagellates and haptophytes); the latter groups appear to have somewhat similar Fe requirements (Fig. 2). Although our analysis does not necessarily support a clear distinction between red and green plastid lineages, it does suggest that certain phyla have reduced their Fe requirements more effectively than others. Members of two taxa in particular, the diatoms and haptophytes, possess the lowest Fe:C ratios, which may explain their persistence (along with picocyanobacteria that primarily benefit from their small size) in a number of low-Fe environments.

Box and whisker plots of Fe requirements (presented in the form of Fe:C ratios) for cyanobacteria and microalgae grouped by phyla. Eukaryotic groups are separated by the green plastid lineage (green) and the red plastid lineage (red). All quotas were obtained from cultures grown in medium that enabled maximum or near maximum (μ/μmax >75 %) growth rates and the Fe concentrations were below levels in which significant amounts of luxury uptake of Fe would occur ([Fe´] <0.25 nM) (Data are compiled from Sunda and Huntsman (1995), Maldonado and Price (1996), Ho et al. (2003), Marchetti et al. (2006), Lane et al. (2009), Quigg et al. (2011) and Strzepek et al. (2011))
2.1 Acclimation and Adaptation to Iron Limitation
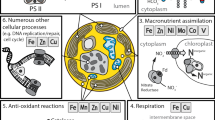
There is considerable pressure on microalgae to evolve mechanisms to reduce their Fe requirements in order to reside in Fe-limited regions. Phytoplankton isolated from Fe-limited oceanic regions typically have lower Fe contents and higher Fe use efficiencies (the amount of C fixed per mole of cellular Fe per day) compared to their coastal congeners. There have been numerous investigations into the mechanisms by which oceanic phytoplankton, particularly diatoms, have acclimate and adapted to the low Fe availability in their environment. One of the most important evolutionary responses to low Fe in the open ocean is a substantial reduction in the cellular Fe requirement for growth (Sunda and Huntsman 1995). Given iron’s involvement in many metabolic pathways, there are numerous strategies invoked by both eukaryotic (Fig. 3) and prokaryotic (Fig. 4) phytoplankton to adapt to low iron availability. These include reducing cell size, using intracellular Fe pools more efficiently, minimizing the use of metabolic pathways (or protein complexes) that require large amounts of Fe and replacing Fe-containing proteins with non-Fe containing ones that are more or less functionally equivalent.

Major forms of Fe in seawater and depiction of a eukaryotic microalgal cell. (a) Bioavailable Fe primarily exists as a dissolved ferric Fe (Fe(III)) ion or tightly bound to organic ligands (L). Ferrous Fe (Fe(II)) is supplied primarily either through wet deposition and/or photoreduction of ferric Fe. Due to the high oxygen content and short residence time, concentrations of free ferrous Fe are extremely low in seawater. Much of the Fe in oxic seawater will eventually form colloids that precipitate out of the water column. The bioavailability of Fe oxyhydroxides and colloidal Fe is not well known, but assumed to be minimal. (b) Fe uptake may occur through a number of mechanisms. Only Fe transport pathways into the cells are illustrated. In general, Fe bound to organic ligands must be reduced before taken up by the cell. Many microalgae have a high-affinity Fe uptake mechanism that couples the Fe reductase with a multicopper-oxidase and Fe permease. Ferrous Fe may also be taken up through non-Fe specific divalent transporters. There is no illustration for proteins involved in intracellular Fe tracking (which are listed in Table 3). The bulk of the Fe demand in eukaryotic microalgae is used within the photosynthetic reaction centers and for the assimilation of NO3 (also shown). Iron containing proteins are indicated in red. Some non-Fe containing proteins that may substitute when Fe levels are low are indicated in purple. Low-Fe induced proteins and processes are indicated in orange, which include an increased dependence on reduced forms of nitrogen (e.g. NH4 and urea) as well as an increased ratio of PSII: PSI reaction centers. In some (but not all) microalgae, the presence of rhodopsins may compensate for a decreased production of ATP from photosynthesis in Fe-limited cells. The localization of rhodopsins to the vacuolar membrane is speculative. Similarly, some microalgae contain ferritins for Fe storage that are located in the plastid. Full protein names are provided in Table 1 (Fe-related proteins) and Table 3 (Fe transport and associated proteins). Others: AMT NH4 transporter, ATP ATP-synthase, FLD flavodoxin, GS glutamine synthetase, NiRT nitrite transporter, NRT nitrate transporter, PCY plastocyanin, RHO rhodopsin

Major forms of Fe in seawater and depiction of a cyanobacteria cell (a) See legend in Fig. 3. (b) Putative Fe transport pathways in cyanobacteria, as predicted (by genomic analyses, protein determination and physiological studies) for model organisms: Pathway I: Schizokinen transport by Anabaena sp., pathway II: reductive transport of organic and inorganic Fe(III) complexes (Synechocystis, PCC6803) and pathway III: Fe(II) transport in various cyanobacteria. Predicted locations and common substrates for each transporter are shown. The bulk of the Fe demand in cyanobacteria is used within the photosynthetic reaction centers and for the assimilation of N (not shown). Iron containing proteins are indicated in red. Some non-Fe containing proteins that may substitute when Fe levels are low are indicated in purple. Low-Fe induced proteins and processes are indicated in orange, which include an increased ratio of PSII: PSI reaction centers. Most cyanobacteria contain ferritins for Fe storage most commonly in the form of bacterioferritin or DPS ferritin. Full protein names are provided in Table 1 (Fe-related proteins) and Table 3 (Fe transport and associated proteins). Others: Fld flavodoxin, isiA Fe-starved induced protein, PC plastocyanin
A reduction in cell size often occurs when microalgae are acclimated to low light or various nutrient limitations, including Fe limitation (Sunda and Huntsman 1997; Marchetti and Cassar 2009). Nutrient requirements for growth per cell decrease as a function of the cube of the cell radius (r3), whereas nutrient uptake decreases as a function of the available membrane area (r2), and the diffusion-limited rate as a function of the radius (r) (Morel et al. 1991). Decreasing cell size would increase the surface area-to-volume ratio and maximize membrane transporters and subsequently uptake rates relative to Fe requirements. A decrease in cell size also decreases the diffusion boundary layer thickness, improving nutrient uptake kinetics. Such a physiological acclimation is observed in microalgae in both field and laboratory conditions. Many Fe-limited regions are populated primarily by small pico- and nanophytoplankton (<5 μm in diameter), suggesting that this small size-fraction has an advantage in coping with low [Fe] as compared to larger phytoplankton species (e.g. diatoms). This is due to the inherent overall lower Fe requirements for small phytoplankton (on a per cell basis) as well as their more effective uptake capabilities on a cell surface area basis. As a response to Fe limitation, many microalgae show a 20–50 % decrease in mean cell volume per cell (Sunda and Huntsman 1995; Muggli et al. 1996; Marchetti and Harrison 2007). This reduction would represent an increase in surface area-to-volume ratio of 8–26 %. Similarly, even cyanobacteria that benefit from being small, have been shown to decrease their size in response to Fe limitation (Sherman and Sherman 1983; Wilhelm 1995). In addition to a reduction in total cell volume, a change in shape from spherical to elongated results in an increase in the SA:V ratio (Marchetti and Cassar 2009). However, there are likely to be constraints on the morphological changes that particular groups of microalgae may undergo, particularly groups such as diatoms with rigid cell walls. Similarly, although usually rare, the persistence of large cells in Fe-limited regions suggests there are benefits to large cell size, possibly in relation to nutrient (and Fe) storage capacities, periodic vertical migration and resistance to grazing (Smetacek et al. 2004).
Microalgae may also adjust their photosystems in response to Fe limitation. There is a preferential down-regulation of PSI relative to PSII (and Cyt b6f) in prokaryotic and eukaryotic phytoplankton under Fe limitation of growth rate (Greene et al. 1991; Moseley et al. 2002; Strzepek and Harrison 2004; Behrenfeld and Milligan 2013). Under Fe deficiency, cyanobacteria may decrease their PSI:PSII ratio from 4:1 to 1:1 (Straus 2004), while the low-Fe adapted oceanic diatom, Thalassiosira oceanica has been shown to have a constitutively low PSI:PSII ratio of 1:10 (Strzepek and Harrison 2004). Ratios of iron rich cytochrome b 6/f complex (5 Fe) to low iron PSII (2–3 Fe) are also reduced in this species (Strzepek and Harrison 2004). The decreased abundance of PSI (12 Fe) and cytochrome b6f complex, however, may have resulted in a diminished ability by T. oceanica to acclimate to fluctuations in photosynthetically active radiation (see section of Fe-light interactions). A recent analysis of the T. oceanica genome and associated transcriptome under Fe-limitation demonstrated that this diatom possesses the potential to differentially adjust many of its iron-containing proteins and protein complexes (including PS1:PSII ratios) in response to Fe availability (Lommer et al. 2012). Major reductions in the PSI:PSII ratio, however may increase the cell’s dependency on alternative electron pathways such as the plastoquinol oxidase (PTOX) pathway, which is present in both prokaryotic and eukaryotic phytoplankton (reviewed by Zehr and Kudela 2009; Behrenfeld and Milligan 2013). The PTOX pathway provides an electron shunt after PSII, enabling cells to bypass PSI and thereby produce ATP without the production of reducing equivalents (NADPH). Because the PTOX pathway has a low Fe requirement (2 Fe molecules per monomer), activation of this shunt under low Fe conditions would substantially reduce a cell’s Fe demand while still being able to generate ATP, however, an alternative source of reductant must be present for carbon fixation to occur.
Cyanobacteria and red algae also express an induced protein (isiA) under Fe stress, which can become the most abundant chl-binding protein in the cell (Ryan-Keogh et al. 2012). The precise roles of isiA remain elusive. Despite strong sequence similarity between IsiA and the gene encoding the PSII core CP43 protein, IsiA is not thought to be a CP43 replacement nor antenna complex for PSII. Rather, during Fe stress, IsiA forms superstructures around PSI trimers and monomers, resulting in a significant increase in cross-sectional absorption area of PSI (σPSI) (Bibby et al. 2001; Boekema et al. 2001). A continued up-regulation in isiA without any further increase in the σPSI led Ryan-Koegh et al. (2012) to speculate that this protein may also be involved in other secondary roles for coping with Fe limitation.
Many of the Fe-requiring proteins in microalgae may be reduced or substituted for non-Fe containing equivalents under low Fe conditions. Some of the protein substitutions represent acclimation strategies where the Fe-containing protein is preferred when cells are growing in Fe-replete conditions. Other protein substitutions represent evolutionary adaptations where species have adapted to chronically low [Fe] by replacing Fe-containing proteins with proteins that contain no metal cofactor or that use other redox active trace metals that are more readily available in the environment . The best example of Fe metalloprotein substitution is flavodoxin for ferredoxin (La Roche et al. 1993). Ferredoxins are Fe-sulfur cluster containing proteins which mediate electron transfer in a range of metabolic reactions. Flavodoxin, on the other hand, does not contain Fe but instead utilizes a single molecule of riboflavin 5′-phosphate as a cofactor. Although flavodoxin does not have as low a redox potential as ferredoxin, the expression of flavodoxin at suboptimal [Fe] would partially alleviate Fe stress. Flavodoxins are commonly found in cyanobacteria, although not all species are found to contain them. For example, Anabaena ATCC 29211 lacks the potential to synthesize flavodoxin, and simply decreases the content of ferredoxin at very low Fe concentrations (Sandmann et al. 1990). The diatom Phaeodactylum tricornutum increases the expression of flavodoxin 25–50-fold under Fe limitation (Allen et al. 2008). The flavodoxin to ferredoxin ratio has been considered to be an in situ marker for Fe stress in phytoplankton (Doucette et al. 1996; La Roche et al. 1996). However, it should be used with caution given that some microalgae have several copies of flavodoxin, with some gene variants not regulated by Fe availability (Whitney et al. 2011) whereas other phytoplankton groups may express flavodoxin constitutively (Pankowski and McMinn 2009). In T. oceanica there has been a permanent transfer of the ferredoxin gene from the chloroplast genome to the nuclear genome (Lommer et al. 2010). This relocation is thought to allow T. oceanica to better coordinate gene expression for its low-Fe response.
Another well-characterized substitution is that of the copper-containing electron transport protein plastocyanin for the Fe-containing cytochrome c6 in photosynthetic electron transport. Where plastocyanin is more commonly used in green algae (Hill and Merchant 1995), in haptophytes, primarily cytochrome c6 is found whereas in diatoms and cyanobacteria either transport protein may be present, and is largely dependent on growth conditions and biogeography (Sandmann et al. 1983). For example, while some oceanic diatoms have replaced cytochrome c6 with plastocyanin, many coastal diatoms continue to use cytochrome c6. In the oceanic diatom T. oceanica, the constitutive use of plastocyanin is believed to reduce the cellular Fe demand by as much as 10 % (Peers and Price 2006).
3 Iron Acquisition Mechanisms
Cellular transport of an essential element refers to the ability of a cell to transfer that element across the plasma membrane, from the outside of the cell to the inside. In general, the word transport and uptake are used interchangeably. Fe acquisition refers to the ability of microalgae to access Fe within a variety of complex chemical species (both inorganic and organic) before it is transported across the plasma membrane. In contrast, assimilation of an element refers to the intracellular incorporation of that element into cellular biomolecules.
Ions of essential elements enter the cell by moving across the cell’s diffusive boundary layer toward the cell surface, then passing through the cell wall (if present) and plasmalemma into the cytoplasm. The rate of diffusion through the surface boundary layer is inversely related to the cell diameter. Hence, large cells are much more prone to suffer from diffusion limitation of uptake of iron, or other nutrients, than smaller cells. The cell wall is not a barrier to ion entry. In contrast, the plasma membrane—which consists of polar lipid bilayers interspersed with proteins—does not allow the free diffusion of charged ions, polar hydrophilic molecules or large neutral molecules. These species must be transported across the plasma membrane via facilitated diffusion (also called passive transport) or active transport. Some of the common characteristics of facilitated diffusion and active transport include: (a) facilitation by a transmembrane transport protein (either a carrier protein or a channel protein), (b) unidirectional and often ion-specific transport, (c) Michaelis-Menten saturation kinetics, and (d) competitive and non-competitive inhibition of ion transport.
The greatest difference between facilitated diffusion and active transport is that in the latter, the ion is transported against an electrochemical gradient, and thus energy input is required. In both, facilitated diffusion and active transport, the substrate first binds to the receptor sites on the outer surface of the transport protein. These carrier proteins (also called permeases or transporters) then undergo a series of conformational changes to transfer the bound solute across the membrane. In contrast, channel proteins are only involved in facilitated diffusion. Channel proteins form an aqueous pore in the membrane through which a specific solute passes, and thus these proteins interact only weakly with the solute.
In phytoplankton, as in all other organisms, Fe uptake is an active transport process. The mechanisms of Fe transport in phytoplankton are extremely diverse and complex, and point to unique Fe uptake mechanisms acquired via horizontal gene transfer (Morrissey and Bowler 2012). We will describe general mechanisms involved in Fe transport by discussing prokaryotic and eukaryotic phytoplankton separately. So far, the vast majority of work on phytoplankton has focused on Fe acquisition and transport, and not on Fe assimilation. The following sections thus focus on these two topics.
There are physiological and molecular aspects of Fe acquisition and transport. From a physiological perspective, the following questions can be addressed: (1) What are the Fe species (oxidation state and chemical complexes) that are bioavailable for uptake by phytoplankton? (2) What are the Fe species that bind to receptor sites on the transporters? (3) What is the Fe species that is transported into the cell? (4) If Fe transport involves various proteins, what are their activities and functions? (5) What is the limiting step in Fe acquisition? and, (6) Does Fe uptake follow Michaelis-Menten kinetics?
3.1 Physiological Aspects of Iron Transport
The vast majority of physiological Fe transport studies in phytoplankton have used marine diatoms as model organisms. However, we expect that most of the findings apply to many other microalgal taxa, and freshwater algae. There is evidence for two Fe uptake systems, a low-affinity and a high-affinity transport system. The low-affinity Fe transport mechanism is utilized when inorganic Fe species are available for uptake, and cells are Fe sufficient. The high-affinity Fe transport system is operational when the cells are Fe-stressed (but before their growth rate is actually reduced due to low Fe level) or growth limited by Fe and where the concentrations of dissolved inorganic Fe species are extremely low, but there is a larger pool of organically bound Fe (e.g., in the nM range). In the low-affinity system, Fe uptake is a function of the labile dissolved inorganic Fe(III) concentration (Fe´) in the bulk medium (Hudson and Morel 1990), while in the high-affinity system, Fe uptake is best predicted by the concentration of organically bound Fe (Maldonado and Price 2001; Strzepek et al. 2011). These two Fe transport systems have been reconciled in a general kinetic model for Fe acquisition by marine phytoplankton (Shaked et al. 2005), where Fe(II) is an obligate intermediate in both systems. The presence of a low- and a high-affinity transport system is often termed biphasic uptake kinetics for short-term uptake rates as a function of nutrient concentration (best fit using a double rectangular hyperbola equation). Biphasic uptake kinetics have been shown for Zn in the haptophyte Emiliania huxleyi (Sunda and Huntsman 1992) and Cu in diatoms (Guo et al. 2010). For Fe, such data are still rare, though hints of biphasic uptake kinetics have been shown in the Southern Ocean prymnesiophyte, Phaeocystis antarctica (Strzepek et al. 2011). These two Fe acquisition mechanisms are believed to share some components/proteins, which are likely to be up-regulated under Fe limiting conditions (see below).
3.1.1 Low-Affinity Acquisition Mechanisms for Inorganic Iron
Before describing Fe transport mechanisms in depth, it is essential to understand how Fe uptake is controlled by the Fe conditions in the growth medium (or the environment), as well as by intracellular Fe levels. Most studies investigating Fe physiology use a chemically well-defined medium such as Aquil (Price et al. 1988/89; Sunda et al. 2005). In this medium, a large excess of aminocarboxylate chelating agents are added to buffer a (nearly) constant concentration of dissolved inorganic ferric iron hydrolysis species ([Fe´]), which in turn controls cellular iron uptake rates, intracellular iron levels and specific growth rate in iron limited algal cultures (Hudson and Morel 1990; Sunda and Huntsman 1995). A typical medium might have 100 μM ethylenediaminetetraacetic acid (EDTA) and nM to μM concentrations of Fe, depending on what degree of Fe limitation or sufficiency needs to be imparted to the phytoplankton (see further discussion below). The Feʹ concentration is maintained by steady state dissociation and formation reactions of the organically bound Fe, and thus is dependent on the total iron concentration [FeT], the chelator concentration and the affinity of the ligand for Fe(III). Photochemistry might also induce an increase in the Feʹ pool, due to photo-redox cycling of the organically complexed Fe (Sunda and Huntsman 2003).
When the concentrations of all the nutrients are in excess, except for Fe, the concentration of labile inorganic Fe [Feʹ] determines microalgal growth rates. Following the Monod equation (Monod 1942), the specific growth rate is determined by the labile dissolved inorganic Fe concentrations ([Feʹ]), as well as the half-saturation constant for Fe for growth (Kμ), and the maximum specific growth rate (μmax) so that:
The specific growth rate can also be related to intracellular Fe content (Harrison and Morel 1986), using the Droop equation (Droop 1970). In this case, the maximum growth rate is only achieved when the microalgae are able to fulfill their cellular Fe demand:
where QFe min is the minimal intracellular Fe content needed to allow any growth, and QFe is the intracellular Fe level. Here μ’max refers to the ‘impossible’ growth rate at infinite quota, but in general μ’max/μmax ~1 and μmax is achieved when the optimal cellular Fe content (QFe max) is reached (Harrison and Morel 1986).
We can also write this equation in terms of QFe max, which is the optimal cellular Fe content when μmax is achieved, thus
When phytoplankton are growing under steady-state conditions (or exponential growth), and the rate of growth is limited by the Fe supply, the growth rate is directly proportional to the steady-state Fe uptake and inversely proportional to the cellular Fe content, so that
The steady-state Fe uptake rates also follow Michaelis-Menten kinetics and can be described as:
where KμQ stands for half-saturation constant for steady-state Fe uptake. Equations 1, 2 and 4 are related to each other quantitatively according to \( \mu ={\rho}^{\mathrm{ss}}/{\mathrm{Q}}_{\mathrm{Fe}} \) (Eq. 3). We can solve for ρss and take the logarithm of both sides of the equation so that log \( {\rho}^{ss}= \log \mu + \log {\mathrm{Q}}_{\mathrm{Fe}} \). This implies that the log of the steady-state Fe uptake rates is the sum of logarithm of μ and the logarithm of QFe (Morel 1987). A graphic representation of this is given in Fig. 5.

(a) Steady-state Fe uptake rates, (b) Fe quotas and (c) growth rates as a function of inorganic Fe concentration ([Fe´]; 10−12 M) for the coccolithophore, Emiliania huxleyi (Sunda and Huntsman 1995). These log-log graphs show how the half-saturation constant for growth, steady-state and short-term Fe uptake rates are related to each other, so that the half-saturation constant for growth is much lower than those for steady-state and short-term Fe uptake rates (Morel 1987). The short-term Fe uptake rates for Fe-limited and Fe-sufficient Emiliania huxleyi are computed, based on our knowledge in other marine phytoplankton species (Harrison and Morel 1986). Our calculated K values are: for growth: Kμ = 1.46 pM (Monod equation); for Fe quota: KQFe = 26 pM (Droop equation); for steady-state Fe uptake rates: Kρ ss = 29 pM [ρss = ρss max * [Fe´]/(Kρ ss + [Fe´])]; and for short-term Fe uptake rates: Kρ = 100 pM (Michaelis-Menten kinetics). See text for equations
The net result is that the half-saturation constant for steady-state Fe uptake rates is higher than that for growth, and that the quantitative relationship between KμQ and Kμ is related to the ability of the cells to modify their cellular Fe quotas (Harrison and Morel 1986; Morel 1987), such that:
For a phytoplankton cell growing under low Fe, a practical implication of Eq. 3, is that fast growth rates can be maintained by decreasing the QFe (Harrison and Morel 1986; Morel 1987), even though at low [Feʹ] ρss will decrease according to Eq. 4.
Short-term Fe uptake rates reflect the rate of Fe uptake under non steady-state conditions. For example, a culture can be grown under a specific Fe limiting concentration, and can be examined for its ability to take up Fe at various [Fe]. Normally, these short-term Fe uptake experiments should not last more than the time it takes a cell to synthesize new proteins. Typically, approximately <6 h is the recommended time for laboratory Fe uptake experiments, but this depends on the microalgal growth rate (i.e. should be increased for slower growing algae). For exponentially growing cells, when the [Fe] in the short-term Fe uptake experiments is equal to the [Fe] in the growth medium, the short-term Fe uptake rates coincide with the steady-state Fe uptake rates since they are determined by the concentration of Feʹ (see Eq. 9 below). However, in the presence of higher [Fe] (but not saturating) in the uptake medium, the short-term Fe uptake rates will be faster than the steady-state Fe uptake rates due to the higher Feʹ in the uptake medium compared to the growth medium.
In general, inorganic Fe acquisition (low-affinity Fe uptake system) is observed when concentrations of Feʹ are in the nanomolar range, and a typical oceanic species is either Fe sufficient or slightly Fe-stressed. Under this condition, short-term Fe transport in phytoplankton follows typical Michaelis-Menten uptake kinetics, where the rate of Fe uptake is determined by the maximum rate of uptake (ρmax), the concentration of labile dissolved inorganic Fe species ([Feʹ]), as well as the half-saturation constant for Fe uptake (Kρ) (Hudson and Morel 1990).
The half-saturation constant for short-term Fe uptake (Kρ) is species specific and its inverse (1/Kρ) provides a measure of the affinity of transporter for Fe. The Kρ for short-term Fe uptake is less plastic than ρmax, which may vary significantly, depending on the number of Fe transporters at the cell surface (Harrison and Morel 1986, 1990). Fe-limited phytoplankton have been observed to increase ρmax values by up to 20-fold, presumably by increasing the number of Fe transporters on the outer cell membrane (Harrison and Morel 1986). This can be graphically represented by two kinetic curves (ρlow and ρhigh) that have a similar Kρ but different ρmax, so that ρmax low and ρmax high represent the ρmax for the Fe-sufficient and Fe-limited phytoplankton, respectively (Morel 1987).
When the [Fe] in the uptake experiment is equal to that in the growth medium, the short-term Fe uptake rates are equal to the steady-state Fe uptake rates. Therefore, for a Fe- sufficient culture, the short-term and steady-state Fe uptake rate should be the same at high [Fe]. Similarly, for a Fe-limited cell, the short-term and steady-state Fe uptake rate should be identical at low [Fe]. As a result, the half-saturation constant for short-term Fe uptake is higher than for steady-state uptake by a factor of ρmax high/ρmax low, so that
We can thus compare half-saturation constants for growth, short-term Fe uptake and steady-state Fe uptake (Fig. 5). As predicted by Morel (1987), Fig. 5 shows how the half-saturation constants for growth, short-term Fe uptake and steady-state Fe uptake vary in magnitude for E. huxleyi (Kρ = 0.1, KμQ = 0.029, and Kμ = 0.0014 nM Feʹ; using data from Sunda and Huntsman 1995), so that
Thus, the ratio of the half-saturation constants for growth and short-term Fe uptake rates is directly determined by the lower and upper limits of the Fe intracellular levels and of the maximum uptake rates, according to Eq. 7 (Morel 1987):
We compiled data for a wide variety of phytoplankton to compare half-saturation constants for growth, short-term Fe uptake and steady-state Fe uptake in microalgae, as previously done by Morel (1987). As predicted by Morel (1987), Table 2 shows how the half-saturation constants for growth, short-term Fe uptake and steady-state Fe uptake vary by orders of magnitude (for cultures Kρ = 3.7, KμQ = 0.37, and Kμ = 0.041 nM Feʹ; for field studies Kρ = 2.96 vs. Kμ = 0.00032 nM Feʹ). The order-of-magnitude difference among these half-saturation constants (Kρ vs. KμQ vs. Kμ) is related to the plasticity of the cells to regulate intracellular Fe levels (lower under Fe limitation), as well as the number of Fe transporters at the cell surface (higher under Fe limitation) under various Fe conditions.
These half-saturation constants shown in Table 2 can also be used to define the Fe-limited condition of a phytoplankton culture or an in situ population (Morel et al. 1991). Under Fe sufficiency, when [Feʹ] > KμQ, near maximum growth rates are observed, and neither the number of Fe transporters are maximized, nor the QFe is minimized. Under Fe stress, when Kμ < [Feʹ] < KμQ, near maximum growth rates are observed, but the number of Fe transporters is maximized and the QFe is minimized. Under Fe limitation, when [Feʹ] < Kμ, the Fe uptake rate is too slow to fulfill the Fe requirement despite maximizing the number of Fe transporters and minimizing QFe, thus cellular growth rate decreases.
3.2 Physical, Chemical and Biological Factors Controlling Fe Transport
Physical factors that may influence rates of Fe uptake include light, temperature and diffusion constraints. Light availability has been observed to affect Fe uptake rates by either generating changes in aquatic Fe chemistry (i.e. changes in [Feʹ] due to photo-redox cycling of iron; Hudson and Morel 1990; Barbeau et al. 2003; Sunda and Huntsman 2003, 2011; Maldonado et al. 2005; Fujii et al. 2011), and/or influencing the energy supply to the cells (ATP and/or NADPH) for Fe acquisition (Strzepek et al. 2011). In addition, a physiological interaction between light and Fe availability during growth has been shown to affect steady-state Fe transport rates (see biological factors below). The temperature effects on Fe uptake are associated with the typical microalgal temperature coefficient (Q10 value is ~2), which means that, in general, the rate of Fe uptake will be two times faster for every 10 °C increase in temperature, as found in the coastal diatom Thalassiosira pseudonana (Sunda and Huntsman 2011). Low temperature may also enhance Fe uptake by increasing the residence time of photo-chemically produced Feʹ in the presence of photolabile ferric chelates (Sunda and Huntsman 2011).
The effects of diffusion limitation of Fe uptake have been discussed in detail in Morel et al. (1991). In essence, theoretical calculations suggest that even with a very low Fe demand for growth, a phytoplankton cell with a radius ≥10 μm, and dividing once a day, would be diffusion limited for Feʹ uptake in the open ocean. This is due to the relationship between Fe demand of a cell and its Fe uptake rate. The former is proportional to the cellular volume and specific growth rate, while the latter is a function of the number of Fe transporters in the outer cell membrane, and the rate at which Fe diffuses to the cell surface. At a given Fe transporter density on the cell membrane (mol μm−2), a smaller cell will achieved faster Fe uptake rates per unit of cell volume (mol Fe L−1 cell volume h−1) than a larger cell, and thus would be better able to fulfill its Fe demand for growth. To enhance Fe transport, a larger cell (or one grown at low [Fe]) may increase the number of Fe transporters at the cell surface. However, eventually, the maximum rate of Fe uptake will be limited, not by the number of Fe transporters, but instead by the rate at which Feʹ diffuses to the cell surface, and this effect will be more pronounced for a bigger cell. Indeed, a large cell (r ≥ 30 μm; Sunda and Huntsman 1997) requires such a large number of Fe transporters at the cell surface to fulfill its Fe demand, that even if it only divides once a day, its rate of Fe uptake is diffusion limited.
The chemical factors that affect Fe uptake include: the total Fe concentration in the environment, the oxidation state of Fe (Fe(II) vs. Fe(III)), its chemical speciation (dissolved inorganic species versus organic Fe complexes), the concentration and binding strength of ligands in the environment, as well as pH effect on Fe complexation (Schenck et al. 1988; Sunda and Huntsman 2003; Shi et al. 2010). In essence, strong Fe binding ligands in seawater (or the medium) directly compete with cell surface proteins involved in Fe transport for dissolved inorganic Fe.
The biological factors that influence Fe uptake rates are cellular Fe demand (i.e. the minimum intracellular Fe needed for maximum growth rate), the Fe limited condition of the cells, and the cell population density. The Fe-limited condition of the cells (i.e. the extent to which the cellular Fe content falls short of the minimum intracellular Fe needed for maximum growth rate, and thus, their relative growth rates (μ/μmax) is below 1) exerts a negative feedback regulation of iron transport systems. In addition, Fe transport has been shown to be affected by the physiological condition of the cells especially with regards to Cu (Peers et al. 2005; Maldonado et al. 2006). Extensive discussion of these chemical and biological factors follows below.
In the 1980s and 1990s a series of studies investigating the kinetics of Fe transport in marine phytoplankton provided the foundation of what we know about Fe transport in these organisms (e.g., Anderson and Morel 1982; Harrison and Morel 1986, 1990), and established the Feʹ model (Hudson and Morel 1990; Sunda and Huntsman 1995, 1997). In essence, the Feʹ model predicts a dependency of Fe uptake rates on the concentrations of labile dissolved inorganic Fe species (Feʹ), which consist of a mixture of dissolved iron hydrolysis species (Fe(OH)2 +, Fe(OH)3, and Fe(OH)4 -), whose composition is highly dependent on pH (Sunda and Huntsman 2003). These studies also established that Fe uptake occurs via specialized active transport proteins on the plasmalemma (Hudson and Morel 1990, 1993). Indeed, binding of Fe to the transport proteins must first occur, followed by internalization. Thus, these Fe transporters behave as surface ligands with a very high affinity for reaction with Feʹ (kL = 2 × 106 M−1 s−1, Hudson and Morel 1990). Iron uptake by these transporters is regulated by a series of reactions:
Based on the limited data available, iron uptake by the iron transport system follows typical Michaelis-Menten uptake kinetics, where the rate of Fe uptake is determined by the maximum rate of uptake (ρmax), the concentration of labile dissolved inorganic Fe species ([Feʹ]), as well as the half-saturation constant for Fe uptake (Kρ) (Harrison and Morel 1986, 1990):
Indeed [Feʹ] is the total concentration of labile dissolved inorganic Fe species whose ‘effective’ reaction rate with the uptake L determines uptake. As a result, Kρ is determined by the rates of metal-ligand binding (kL) and dissociation (k-L), as well as the rate of Fe internalization (kin), such that Kρ = (k-L + kin)/kL. Hudson and Morel (1990) demonstrated that Fe transport was under kinetic control, meaning that the rate of internalization of the Fe bound to the cell surface ligands is much faster than its rate of dissociation from the surface ligands, thus the rate of dissociation can be ignored and Kρ = kin/kL. As discussed above, the maximum rate of Fe uptake (ρmax) is highly affected by the number of transport ligands (LT) at the cell surface, which can increase by more than 20-fold when cells are experiencing Fe limitation (Harrison and Morel 1986). The expected increase in Fe transport due to higher density of Fe transporters (up to 20-fold) is far greater than that expected from a decrease in cell size (~2 fold, see section above).
When phytoplankton are Fe-limited, their maximum short-term Fe uptake rates (in the presence of non-limiting [Feʹ]) are determined by the number of Fe transporters at the cell surface and the rate of internalization, so that
In addition, Kρ = kin/kL, and under Fe-limiting conditions [Feʹ] <<< Kρ, so the upper limit on their steady-state Fe uptake rates can be simplified from the Michaelis-Menten equation above (Eq. 9) to
Thus, under steady-state Fe limiting conditions, the growth rate is defined as:
In summary, the Feʹ model predicts that Fe uptake rates are dependent on dissolved inorganic Fe(III) species (Feʹ), and that these Feʹ species (be it Fe(II)’ or Fe(III)’) bind the Fe surface transporters before the Fe is internalized. This Feʹ model has been extremely useful in the last 25 years for laboratory trace metal algal physiology and toxicity studies.
3.2.1 High-Affinity Acquisition Mechanisms for Organically Bound Iron
The development of extremely sensitive analytical techniques in the 1990s allowed measurements of organic complexation of Fe in the sea. The finding that the vast majority of dissolved Fe (>99.9 %) is bound to very strong organic complexes (Gledhill and van den Berg 1994; Rue and Bruland 1995; for a review see Gledhill and Buck 2012) led to the reexamination of the Feʹ model because the calculated Feʹ in the open ocean, in the absence of photochemical reactions, was shown to be too low to support sufficient Feʹ to meet cellular requirements of open ocean phytoplankton (Rue and Bruland 1995). At the same time that Fe speciation in the ocean was being unraveled, laboratory experiments were also revealing new insights into Fe transport in phytoplankton (e.g. Allnutt and Bonner 1987a, b; Jones et al. 1987; Soria-Dengg and Horstmann 1995; Hutchins et al. 1999b; Maldonado and Price 2000, 2001).
Physiological studies of Fe nutrition in phytoplankton were conducted using model siderophores, such as desferrioxamine B (DFB), to induce severe Fe limiting conditions in the growth medium. Siderophores (from the Greek “iron carriers”) are some of the highest affinity ferric chelators known in nature (Neilands 1995). These molecules have a moderately low molecular mass (usually <1000), and are secreted mainly by Fe-limited bacteria, fungi and grasses to scavenge Fe from the environment. Depending on the Fe(III) binding group, siderophores can be classified as hydroxamates, catecholates or mixed-ligand types (containing another Fe binding group such as α-hydroxy-carboxylate, in addition to hydroxamate or catecholate ligand groups). Once the siderophore complexes a Fe(III) ion, the ferrisiderophore is transported into the cell via specific membrane bound siderophore transporters. In the early 1990s, eukaryotic phytoplankton had not been shown to be able to internalize ferrisiderophores. Therefore, siderophores were added to the medium to achieve extreme in situ Fe-limiting conditions, where Feʹ is practically undetectable.
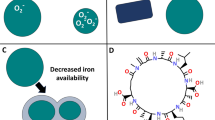
Surprisingly, these physiological studies showed that Fe-limited phytoplankton could access Fe from these strong organic complexes, using an enzymatic reductive mechanism at the cell surface (Allnutt and Bonner 1987a, b; Soria-Dengg and Horstmann 1995; Maldonado and Price 2000, 2001; Weger et al. 2002; Matz et al. 2006). The ability of the phytoplankton to access these strong organic ligands was induced under Fe limitation (Allnutt and Bonner 1987a; Maldonado and Price 1999, 2001; Weger et al. 2002; Strzepek et al. 2011), suggesting that this was a high-affinity Fe transport system similar to that of other well studied eukaryotes (i.e. the yeast Saccharomyces cerevisiae) (for a review, see Van Ho et al. 2002). It is now evident that the high-affinity Fe transport system in phytoplankton involves the activity of Fe permeases in the outer cell membrane, as well as ferric reductases and multi-Cu containing ferroxidases (Fig. 3). Indeed, physiological evidence for the existence of a Fe reductive pathway in microalgae is widespread (see Table A1 in Shaked and Lis 2012).
The ferric reductases are transmembrane proteins in the plasmalemma that transfer an electron from cytosolic NAD(P)H to the iron complexes outside of the cell, including Fe strongly bound in organic chelates. Since most organic Fe chelators have a much higher affinity for Fe(III) than Fe(II), Fe reduction results in a dissociation of the Fe(II) from the ligand, and thus an increase of inorganic Fe(II) at the cell surface. The rates of Fe reduction of the organic complexes are inversely related to the stability of the Fe(III) coordination complex (Maldonado and Price 2001). However, once the Fe is reduced, the fraction of this Fe taken up into the cell depends critically on the relative concentration (and affinities) of the free organic ligands in solution and the free Fe transporters at the cell surface (see the Fe(II)s model below).
Once the Fe(III) is reduced to Fe(II) and dissociates from the organic complex, the free Fe(II) then binds with a receptor site on an iron membrane transport-complex (which consists of a multi-Cu containing oxidase and the permease). The Fe(II) is subsequently (and likely rapidly) oxidized to Fe(III) by multi-Cu oxidases associated with the permease, followed by internalization of the Fe(III) by the permease. These reduction and oxidation steps facilitate the transfer of iron from ligands in the external medium to the receptor site on the iron membrane transport-complex. They also impart specificity and selectivity to the Fe transport system, which is crucial in the case of essential trace elements. Physiological studies with Fe-limited diatoms have measured comparable rates of Fe(III) reduction of organically bound Fe and Fe(II) oxidation (Herbik et al. 2002a, b; Maldonado et al. 2006). The coupling between these oxidation and reduction rates may ensure that, just after the Fe(III) is reduced by the reductase, the Fe(II) is rapidly oxidized by the putative multi-Cu oxidase, before it is internalized by the permease as Fe(III). The proximity of these putative reductases, oxidases and permeases at the cell surface of microalgae may contribute to an efficient cascade of redox reactions. Close proximity of the reductases and oxidases may also allow the formation of a ternary complex, ferric reductase-Fe(III)siderophore-putative Fe(II) oxidase, which may facilitate the reductive dissociation of Fe from very strong organic complexes (Boukhalfa and Crumbliss 2002).
The occurrence of the reductive Fe uptake pathway to acquire Fe from strong organic Fe complexes is widespread in freshwater and marine microalgae (see Table A1 in Shaked and Lis 2012). In many instances, physiological data have been complemented with genomic and proteomic data. One field study also measured Fe reduction rates of organically bound Fe by in situ phytoplankton (Maldonado and Price 1999). Most recently, data from the Global Ocean Survey (GOS) metagenomes have shown that ferric reductases are characteristic of marine eukaryotic phytoplankton Fe uptake systems (Desai et al. 2012). Physiological evidence for the multi-Cu containing oxidases is still limited to a few species, including Chlamydomonas reinhardtii, Thalassiosira pseudonana and T. oceanica (Herbik et al. 2002a, b; La Fontaine et al. 2002; Peers et al. 2005; Maldonado et al. 2006). Molecular and genomic evidence for the occurrence of multi-Cu-containing oxidases is starting to emerge (Maldonado et al. 2006; Kustka et al. 2007; Paz et al. 2007a; Guo et al. 2015).
Recently, a limited set of studies has demonstrated that Fe reduction is also a necessary step for the acquisition of inorganic Fe in the presence of weak organic ligands (such as EDTA), where the dissolved inorganic Fe(III) pool is significant (Shaked et al. 2005); in this case the ferric reductase reduces Fe(III)’ to Fe(II)’. As discussed above in the “inorganic Fe acquisition section”, Fe uptake in this case is still a function of the concentrations of labile dissolved inorganic Fe(III), because the production of Fe(II)’ depends on concentration of Fe(III)’, and can be predicted with the Feʹ model (or Fe(III)’ model). However, these studies have concluded that Fe(II) is an obligate intermediate in phytoplankton Fe acquisition, regardless of whether the cells are acquiring Fe(III) from inorganic or organic Fe(III) complexes (Shaked et al. 2005). Yet, it is important to emphasize that the high-affinity Fe permeases ferry Fe(III) across the plasma membrane, and not Fe(II). Thus, if Fe(II) is an obligate intermediate for Fe acquisition of inorganic Fe (at non-saturating Feʹ concentrations), this Fe(II) is either oxidized by a multi-Cu-containing oxidase-Fe permease complex, like that of the high-affinity Fe transport system for organically bound Fe, or alternatively, is directly internalized as Fe(II) via lower affinity divalent transporters such as ZIP and NRAMPS (see below). Further research is needed to elucidate the mechanism of uptake of Fe(II) after the enzymatic reduction of Fe complexes.
Based on the new findings that organically complexed Fe is bioavailable via a reductive mechanism at the cell surface, and that inorganic Fe acquisition also requires a reductive step, Shaked et al. (2005) proposed a new conceptual model for Fe transport by phytoplankton, “the Fe(II)s model” (Fig. 6). The name of this model was chosen to emphasize that reduction of Fe at the cell surface is an obligate intermediate in Fe acquisition. Unfortunately, to the non-specialist, this name gives the false impression that Fe(II) is the Fe species that is internalized by the phytoplankton.

Cartoon schematic of the most common Fe transport kinetic models in microalgae: Fe´ model (Hudson and Morel 1990) and the Fe(II)s model (Shaked et al. 2005). These kinetic models are for a medium with an excess of a photolabile organic ligands, inorganic Fe(III)’, and inorganic Fe(II)’. Fe(III)’ is more chemically stable than Fe(II)’ and is the dominant species for Fe transport. The concentration of Fe(III)’ is maintained at equilibrium by dissociation (rate constant k d ) and complexation (rate constant k f ) reactions with the organic ligands, as well as photoreduction of Fe(III) bound to photolabile organic ligands (rate constant k hv ) to Fe(II)’, which quickly oxidizes to Fe(III)’ (rate constant k ox ) at seawater pH (~8.1). In the Fe´ model, Fe(III)’ are the species reacting with the Fe transporter. In the Fe(II)s model, both the Fe(III)’ and the organic Fe have to be enzymatically reduced to Fe(II) before Fe is transported into the cell through the Fe(III) transporter complex. This complex is hypothesized to contain a multi-Cu containing oxidase and an Fe permease. Fe(II)’ may also be directly internalized, but these divalent transporters are low affinity and are not illustrated here. For details of the Fe(II)s model and the Fe´ model, see Shaked et al. (2005), and Hudson and Morel (1990), respectively. Most model studies have used eukaryotes, though some prokaryotes have proven to have similar kinetics
The Fe(II)s model builds on the previous kinetic model of Fe uptake (Hudson and Morel 1990), “the Feʹ model”, which predicted Fe uptake rates based on the concentration of labile dissolved inorganic Fe(III) in the bulk medium. The Feʹ model is valid for well-buffered Fe media, in the presence of excess concentrations of aminocarboxylate chelating agents. However, the Feʹ model is incomplete because it is unable to predict rates of Fe uptake when the ratio of [Fe(III)’] to organically bound iron is extremely low; as observed in the presence of very strong organic Fe complexes, such as DFB. The Fe(II)s model is meant for calculating rates of Fe uptake when the concentrations of Fe(III)’ in the media or seawater are non-saturating. This model reconciles the Feʹ model with the new data on the extracellular, biologically mediated Fe reduction of, not only, strong organic Fe complexes, but also weak organic complexes and inorganic Fe species (Shaked et al. 2005). The Fe(II)s model is not mechanistic, but is based on the premise that the Fe acquisition mechanism in marine eukaryotic phytoplankton is similar to that of yeast and involves Fe reductases at the cell surface, followed by the activity of a Fe transport complex, which contains a multi-Cu-containing Fe oxidase and a permease. It aims to predict Fe uptake rates based only on reduction rates of Fe(III), either bound within organic or inorganic complexes. Specifically, the Fe(II)s model predicts Fe uptake based on the concentration of Fe(II) at the cell surface, which is determined by (a) reduction of organically bound Fe (FeY) at the cell surface, (b) reduction of Fe(III)’ at the cell surface, and (c) the competition for free or inorganic Fe (assigned a nomenclature of Fe(II)s, but includes inorganic and free Fe(II) or Fe(III) species) between the cell surface Fe transport complexes and the free organic ligand in the medium (Z). [Note: in cases where only one ligand is present in the medium, Y = Z]. Thus, only three parameters need to be specified: (a) cellular FeY reduction constant (kFeY red; L cell−1 h−1), (b) cellular Fe(III)’ reduction constant (kFe(III)’ red; L cell−1 h−1;), and (c) the ratio (kz/kup; M−1) of effective formation constant of Fe(II)s complexation by excess ligand Z (kz; M−1 h−1) and the rate constant of Fe(II)s complexation by the membrane Fe transport complex under non-saturating [Fe(III)’](kup; h−1).
In the presence of weak organic Fe complexes, such as EDTA (i.e. [EDTA] = 5–100 μM & Fe at ~100 nM), Feʹ is relatively high and reduction of Fe(III)’ dominates. Since EDTA cannot compete effectively with the cell Fe transport complex for the Fe(II)s, Fe uptake rate is simply a function of kFe(III)’ red and the [Fe(III)’], so that Fe uptake (ρFe; mol Fe cell−1 h−1) is defined as
In the presence of very strong organic ligands, such as FeDFB, [Fe(III)’] is practically non-existent, thus reduction of FeY dominates, and competition for Fe(II)s between the cell surface Fe transport complexes and the free organic ligand in the medium (Z) has to be incorporated into the equation (note that Z can be DFB or some other organic ligand), so that
The Fe(II)s model is able to reconcile the Fe(III)’ model (because in the presence of weak organic Fe complexes, the inorganic Fe(III) is the substrate for cellular surface Fe reduction and uptake) with the laboratory and field data showing Fe uptake from strong Fe organic complexes, such as siderophores. This is a kinetic model for Fe acquisition under non-saturating Fe concentrations, in which the activities of the Fe reductases and the Fe-transport complexes (multi-Cu oxidases and Fe permeases) increase proportionally to the concentrations of Fe(III) and Fe(II), respectively. These non-saturating conditions are prominent in the open ocean, as well as in laboratory experiments under Fe limiting conditions. Processes able to increase [Feʹ] in surface oceanic waters, such as photoreduction of organic Fe complexes (Barbeau et al. 2001), photodissolution of Fe oxides (Waite and Morel 1984) or superoxides (reviewed by Rose 2012) would potentially increase Fe(III)’ for cellular reduction at the cell surface. Shaked et al. (2005) estimated that in the open ocean, the Fe needed for phytoplankton growth is supplied by cellular reduction of mainly FeY (2/3rds) and some Fe(III)’ (1/3rd).
3.3 Molecular Aspects of Fe Transport
From a molecular perspective, the following questions can be addressed: (1) Is Fe uptake achieved by a single Fe transporter or a Fe transport complex (i.e. a transport system involving multiple proteins)? (2) What are the expression patterns of the proteins involved in Fe acquisition? (3) How are these proteins arranged in the plasmalemma? and, (4) How many transmembrane domains do these transporters have?
3.3.1 Eukaryotic Microalgae
Though we are starting to learn more about molecular aspects of Fe transport in marine eukaryotic microalgae (reviewed by Blaby-Haas and Merchant 2012) so far, the best characterized Fe transporters in microalgae are those of the freshwater green alga Chlamydomonas reinhardtii (reviewed by Merchant et al. 2006). Fe(III) transport occurs through FTR (Fe TRansporter) (Table 3). These FTRs are localized to the plasma membrane and form a complex with multi-Cu oxidases (referred to as FOXs, FET3Ps, or MUCOXs depending on the organisms). In addition, these Fe transport complexes are associated with ferric reductases (or FREs).
Three types of ferric reductases are involved with Fe(III) reduction in eukaryotes: NAD(P)H oxidases (NOX; cytochrome b558), cytochrome b5 reductases, and cytochrome b561. In C. reinhardtii, FRE1 (a NOX enzyme) is localized in the plasma membrane and is highly induced under Fe deficiency; thus, this Fe reductase is hypothesized to be involved in Fe acquisition (Allen et al. 2007). In diatoms, the following Fe reductases have been identified: T. pseudonana (TpFRE1 and TpFRE2, Kustka et al. 2007), Phaeodactylum tricornutum (PtFRE1-4, Allen et al. 2008), and T. oceanica (ToFRE1, Lommer et al. 2012). TpFRE1 is similar to cytochrome b5 reductase, while PtFRE3 and PtFRE4 are similar to cytochrome b561 (Allen et al. 2008), and TpFRE2, PtFRE1 and PtFRE2 to NOX proteins (Blaby-Haas and Merchant 2012). The transcript abundance of many of these diatoms’ FREs increases under Fe limitation (Kustka et al. 2007; Allen et al. 2008), thus Fe reduction is believe to be an integral component of the high-affinity Fe transport system. In addition, Fe reduction is also a required step for Fe uptake of inorganic Fe into the cell (Shaked et al. 2005), as well as mobilization through intracellular membranes, such as into the chloroplast or out of the vacuole (reviewed by Nouet et al. 2011).
Putative multi-Cu oxidases have been identified in many genomes, including FOX in C. reinhardtii, Chlorella variabilis, Coccomyxa sp. C-169, and Volvox carteri; FET3P in Fragilariopsis cylindrus and Thalassiosira pseudonana; and MUCOX2 in Thalassiosira oceanica (Blaby-Haas and Merchant 2012). As with the ferric reductases, the transcript abundance of these genes is enhanced in Fe-limited cultures of C. reinhardtii (La Fontaine et al. 2002; Allen et al. 2007) and T. oceanica (Maldonado et al. 2006). Surprisingly, gene homologs to these multi-Cu containing ferroxidases have not been found in P. tricornutum (Allen et al. 2008), nor is its expression always enhanced in Fe-limited T. pseudonana (see Maldonado et al. 2006; Kustka et al. 2007). Multi-Cu oxidases contain four Cu atoms, and are divided into two types: FET3p-type and ceruloplasmin. While the multi-Cu oxidases in green algae are similar to the second type, the ones from diatoms seem more comparable to FET3p. Interestingly, the MCO of the pelagophyte Aureococcus anophagefferens is different from these two types, and Cyanidioschyzon merolae is lacking a MCO (Blaby-Haas and Merchant 2012). A physiological interaction between Fe and Cu has been shown in eukaryotic algae (Herbik et al. 2002a, b; La Fontaine et al. 2002; Peers et al. 2005; Maldonado et al. 2006; Annett et al. 2008; Guo et al. 2010, 2012), as well as cyanobacteria (Nicolaisen et al. 2010). For example, when diatoms are co-limited by Cu and Fe, the oxidation rates of Fe are slower, resulting in slower Fe uptake (Maldonado et al. 2006).
In yeast, internalization of Fe by the multi-Cu oxidase-Fe permease complex has been proposed to occur by a Fe-channeling mechanism which is activated when the multi-Cu oxidase oxidizes Fe(II) to Fe(III) and transfers it to the Fe permease (Stearman et al. 1996). In addition, the maturation of the FET3 and FTR proteins is coupled in the secretory pathway, so that only the complex containing both proteins is present in the plasma membrane (Stearman et al. 1996). FTR1 homologs are common in microalgal genomes, except in prasinophytes and some chromalveolates (Blaby-Haas and Merchant 2012). Fe deficiency induces FTR1 genes in T. pseudonana (Kustka et al. 2007) and C. reinhardtii (La Fontaine et al. 2002; Allen et al. 2007). When neither FOX1 nor FTR1 are present, as in prasinophytes or P. tricornutum, direct uptake of organic Fe complexes might occur (Blaby-Haas and Merchant 2012). Indeed, a gene encoding a putative siderophore binding protein was recently identified in P. tricornutum (Allen et al. 2008).
In general, algal high-affinity Fe transporters are structurally divergent from those well characterized in Saccharomyces cerevisiae. Novel proteins have been hypothesized to be involved in Fe transport, such as FEA1- and FEA2-like protein in C. reinhardtii (Allen et al. 2007), FEA1-like protein in Micromonas pusilla and Ostreococcus lucimarinus and “Iron-starved-Induced Proteins” (ISIPs) in diatoms (Lommer et al. 2012). Since M. pusilla and O. lucimarinus lack typical, high-affinity Fe transport FTR1 homologs, these FEA1 proteins may facilitate Fe-specific transport in these organisms (Blaby-Haas and Merchant 2012).
In addition to Fe(III) transporters, Fe(II) transporters such as those belonging to the ZIP (reviewed by Gaither and Eide 2001) and NRAMP (reviewed by Nevo and Nelson 2006) families are found in microalgae (Table 3). An important difference between the Fe(III) transporters discussed above and the ZIP and NRAMP Fe(II) transporters is their specificity. In essence, ZIP and NRAMP proteins may transport Fe(II) in addition to other divalent metals. Thus, these Fe(II) transporters have been hypothesized to be: (a) low(er)-affinity plasma membrane Fe permeases, or (b) proteins involved in the trafficking of intracellular Fe to various cellular compartments. The ZIP family was named after the first discovery of a Zn transporter (Zrt1p and Zrt2p; Zhao and Eide 1996a, b) in S. cerevisiae and the Fe transporter IRT1 in Arabidopsis roots (Eide et al. 1996). ZIP transporters may be localized to plasma and vacuolar membranes or the secretory system. ZIP metal transporters are believed to be passive, mediating metal transport via concentration gradients (Lin et al. 2010). Within the ZIP family, there are four groups of transporters, depending on their phylogenetic relationships: Subfamilies I (with mainly fungal and plant proteins) and II (with plant and animal proteins), GufA (with prokaryotic and eukaryotic proteins) and LZT (with human LIV-1 Zn transporter as the founder) (Gaither and Eide 2001). ZIP transporters are not metal specific but are able to transport a variety of divalent cations, such as those of Fe, Cu, Zn, and Mn. In C. reinhardtii, there are 13 ZIP family members, including IRT1 and IRT2 (from the GufA family), both of which are up-regulated under Fe deficiency (Allen et al. 2007). In addition, IRT1 and IRT2 are induced under Zn and Cu limitation (Castruita et al. 2011), respectively. Since IRT2 is also a potential target for the Cu responsive transcription factor CRR1, IRT2 has been hypothesized to be an intracellular Fe(II) transporter delivering Fe to cytochrome c6, or an alternative plasma membrane Fe transporter when Cu is limiting and the high affinity Fe transporter-complex is not functional (Blaby-Haas and Merchant 2012). The diatoms P. tricornutum (Allen et al. 2008) and T. pseudonana (Kustka et al. 2007) also have ZIP homologs. As in C. reinhardtii, the gene transcript of a ZRT, IRT-like protein, belonging to the ZIP family, was enriched under Fe deficiency in P. tricornutum (Allen et al. 2008). In T. pseudonana, five genes encoding ZIP-like transporters have been identified, and the expression of some of these genes are enhanced under Fe deficiency or repressed after a Cu addition to a low Cu culture (Guo et al. 2015).
Another family of metal permeases able to transport Fe(II) is the NRAMP family (reviewed by Cellier et al. 2001). This family is named after its first identified member, a Natural-Resistance-Associated Macro-Phage protein 1. Most NRAMP permeases are able to transport divalent cations, mainly Fe and Mn, into the cytoplasm. As with ZIP proteins, NRAMPs may be localized to the secretory system, plasma and vacuolar membranes. NRAMP proteins rely on a proton gradient for metal transport. Phylogenetically, the NRAMP family is divided into four subfamilies: prokaryotic MntH group A, B, and C, as well as a eukaryotic group (Cellier et al. 2001). Within the eukaryotic group, three clusters are found: subgroup I (with NRAMPs from major eukaryotic lineages), II (NRAMPs from plants, prasinophytes and C. merolae), and III (fungal proteins). Most microalgal genomes to date contain one NRAMP homolog (Blaby-Haas and Merchant 2012), except for P. tricornutum, which lacks an NRAMP homolog (Kustka et al. 2007). The Chlamydomonas genome encodes four NRAMPs: NRAMP1, NRAMP2, RET1 and NRAMP4, although some of these proteins do not have metal transport activity (Blaby-Haas and Merchant 2012). An ortholog of NRAMP4 is highly up-regulated in Fe-limited T. pseudonana, however whether this permease is located to the plasma or vacuolar membrane remains to be established (Kustka et al. 2007). In a more recent study, the expression of TpNRAMP was up-regulated not only by Fe limitation, but also by low Cu (Guo et al. 2015). Once Fe enters the cell, it must be taken to its sites of incorporation or storage (for a review, see Nouet et al. 2011).
3.3.2 Prokaryotic Phytoplankton
Because siderophore production is more prominent in prokaryotic organisms than in eukaryotes, since the 1970s many cyanobacteria have been tested for their ability to produce siderophores. Cyanobacteria, both marine and freshwater, are able to produce hydroxamate- (i.e. synechobactin A–C, (Ito and Butler 2005); schizokinen, (Simpson and Neilands 1976)) and catecholate-type siderophores (i.e. anachelin-H, and anachelin-1 and -2 (Beiderbeck et al. 2000)), though the former is more commonly synthesized (reviewed by Gademann and Portmann 2008). Interestingly, some of the marine cyanobacterial siderophores are photolabile, and have a fatty acid tail, which is believed to enhance the affinity of the siderophore for the bacterial membrane (Ito and Butler 2005), as previously shown for marine heterotrophic bacteria (Barbeau et al. 2003; Martinez et al. 2003). These two characteristics might aid in Fe acquisition in the dilute oceanic environment.
Siderophores can be synthesized by the non-ribosomal peptide synthase (NRPSs) pathway or by the NRPS-independent siderophore (NISs) biosynthesis pathway. NRPSs are normally associated with polyketide synthases (PKSs) (Table 3). Several genes coding for NRPSs and PKSs have been found in cyanobacteria genomes (Silva-Stenico et al. 2011), but in some cases the specific siderophore that is produced by these synthases is unknown. Both NRPSs and PKSs genes are more prevalent in filamentous and heterocystous cyanobacteria, but were absent in many unicellular strains, including Synechocystis, Prochlorococcus, and Synechoccocus (Ehrenreich et al. 2005). In general, NRPSs genes seem to be more prominent in cyanobacteria genomes than NISs (Hopkinson and Morel 2009). However, many NRPSs pathways synthesize secondary metabolites, instead of siderophores. Thus, the presence of NRPSs genes only suggests the potential for siderophore synthesis. In contrast, finding genes encoding proteins in NIS pathways are indicative of siderophore biosynthesis (mainly hydroxamates and mixed-hydroxamate ligands). In a study of marine prokaryotic genomes, genes for the NIS siderophore biosynthesis pathway were only found in 15 % of the marine prokaryotic genomes, and were completely absent from the oceanic environmental samples (Hopkinson and Barbeau 2012). The low abundance of siderophore biosynthesis genes in the ocean suggests that many marine cyanobacteria utilize non-siderophore iron uptake systems or may be involved in siderophore Fe piracy by stripping the Fe off ferrisiderophores at the cell surface (via a reductive mechanism or a ternary complex formation). Recent studies on the diversity of iron uptake systems in the ocean examined sequenced genomes of marine microbes (eukaryotes and prokaryotes), as well as metagenomes from GOS (Desai et al. 2012). They demonstrated the presence of some of the components of rhizobactin siderophore synthesis in cyanobacteria and eukaryotic phytoplankton genomes, as well as in the metagenomes (though at much lower abundance). One of the few eukaryotic microalgae previously shown to have a siderophore biosynthesis pathway is the prasinophyte O. lucimarinus (Palenik et al. 2007).
Little is known about how cyanobacteria secrete siderophores to the environment after intracellular synthesis. The only siderophore secretion pathway studied in detail is that of Anabaena sp. PCC 7120. Export through the inner membrane (the plasmalemma) is suggested to be mediated by a transporter from the Resistance, Nodulation and cell Division (RND), the Major Facilitator (MFS) or the ATP-binding cassette (ABC) superfamilies, while transport through the outer membrane is suggested to occur by a TonlC-type protein, as in other bacteria (Bleuel et al. 2005; and reviewed by Miethke and Marahiel 2007). In Anabaena sp. PCC 7120, siderophore schizokinen secretion is mediated by the MFS-type protein SchE in the inner membrane and by the TonlC-type protein HgdD in the outer membrane (Nicolaisen et al. 2010).
Siderophore acquisition has been studied in detail in Escherichia coli and Pseudomonas aeruginosa. Siderophore-specific transporters are found in the outer membrane of Gram-negative bacteria, and are part of the TonB-dependent transporter (TBDT) family. Although TBDTs are known to transport a variety of substrates, their structure consists of a membrane bound 22-stranded β-barrel (poorly conserved), and a N-terminal plug domain (highly conserved) located within the barrel. Molecular dynamic simulations indicated that binding of siderophores, such as ferrichrome, initiates a signaling mechanism that ultimately leads to the TonB-mediated partial or total removal of the core domain from the β-barrel, thus opening up a permeable pore (Faraldo-Gomez and Sansom 2003; Faraldo-Gomez et al. 2003). In many cases, the substrate-binding site in the extracellular pocket of the transporter lacks sequence conservation (Chimento et al. 2005), thus allowing TBDTs to have specificity for a variety of substrates, besides siderophores, such as haeme, vitamin B12 and Fe-citrate (reviewed by Schauer et al. 2008).
Even though many TBDTs have been identified in cyanobacteria (Mirus et al. 2009), their function in Fe transport has only been shown in a few species. For example, a TBDT belonging to Anabaena sp. PCC 7120 has been identified (SchT) for the transport of a specific siderophore, schizokinen (Fig. 4) (Nicolaisen et al. 2008). SchT is encoded by alr0379; it spans across the outer membrane, and it is fueled by a “Ton” system anchored in the periplasmic membrane. Bacterial TonB-dependent transporters consists of an energy transducing unit, TonB, and two stabilizing units, ExbB and ExbD (reviewed by Noinaj et al. 2010). The interaction between TBDT and the “Ton” system is mediated by TonB. The TBDT contains a TonB-box in front of its N-terminal plug domain, which is recognized by TonB. In the absence of a ferrisiderophore, the plug domain is blocked. The interaction between a ferrisiderophore-loaded TBDT and TonB results in a conformational change in the plug domain, allowing passage of the siderophore through the TBDT pore (Fig. 4).
Other components of siderophore-mediated iron transport were recently identified in Anabaena sp. PCC 7120, namely TonB3, and the ExbB3/ExbD3 and the Fhu systems (Stevanovic et al. 2012) (Table 3). The transcript abundance of these genes is enhanced under iron-limiting conditions, as is typical of high-affinity Fe transport systems (Stevanovic et al. 2012). In general, transport of ferrisiderophore across the periplasmic and inner membrane is then accomplished by a three-component system: a periplasmic Fe binding protein (e.g. FhuD/FutA/FecA), a membrane-embedded permease (e.g. FhuB/FutB/FecCD) and an ATP-binding protein (e.g. FhuC/FutC/FecB). The FhuD/B/C-system that allows transport of ferrichrome in E. coli is well established (reviewed by Krewulak and Vogel 2011). So far, in Anabaena sp. PCC7120, five gene clusters similar to Fut-, Fec- and Fhu-system have been annotated, but only the Fhu-cluster has been linked to intracellular ferrisiderophore transport (Stevanovic et al. 2012). The expression of some of these genes in Anabaena sp. is enhanced by not only low Fe, but also by high Cu (Stevanovic et al. 2012).
Genes encoding putative TonB-dependent receptors are found in many freshwater cyanobacterial genomes, but are much less common in marine cyanobacteria, and practically absent in picocyanobacteria (Hopkinson and Morel 2009; Hopkinson and Barbeau 2012). Interestingly, the only TBDT-like gene found in picocyanobacteria in this latter study, is a TBDT-type heme transporter (as well as its associated TonB energy translocation system, ExbB/Exbd) in the genome of Prochlorococcus sp. MIT9202. These results suggest that direct internalization of ferrisiderophores might not be a widespread physiological strategy to acquire Fe in the ocean. However, the most recent genomic study by Desai et al. (2012) found that TBD hydroxymate uptake components were relatively abundant in the metagenomes, in agreement with the abundance of hydroxymate siderophores in the open ocean.
Axenic cultures of cyanobacteria, as well as natural populations of marine Synechococcus have been shown to use a reductive mechanism at the cell surface, similar to that of eukaryotic microalgae, to access the Fe within siderophore complexes (Lis and Shaked 2009; Kranzler et al. 2011). This strategy is advantageous in the ocean because it allows access to Fe from a wide variety of naturally occurring strong organic Fe complexes. One of the pending questions in this research is where does the reduction of Fe bound to strong organic complexes occur in cyanobacteria? Iron reduction is most likely to occur either on the surface of the outer membrane or in the periplasmic space. Studies investigating reduction of Fe in strong organic Fe complexes use the Fe(II) trapping agent ferrozine (FZ). These studies measure Fe(II)FZ in solution, suggesting that Fe(II)FZ3 is produced outside the cell. Since Fe(II)FZ3 is not available for uptake, it is possible that FZ crosses the outer membrane, traps the Fe(II) produced in the periplasmic space and then the resultant Fe(II)FZ3 diffuses back across the outer membrane into the surrounding medium.
Recently, a reductive pathway for the transport of inorganic Fe has also been proposed for the freshwater Synechocystis strain (PCC6803) (Kranzler et al. 2014) (Fig. 4). Inorganic Fe(III) (i.e., Fe(III)’) is first transported across the outer membrane, possibly through a non-specific porin (Fujii et al. 2011). Once in the periplasma, Fe(III) is complexed by the high-affinity Fe binding protein FutA2, thus establishing a chemical gradient for Fe(III) influx through the outer membrane. Subsequently iron in the Fe(III) within the Fe(III)FutA2 protein is reduced to Fe(II) by the integral plasma membrane cytochrome c oxidase (Alternate Respiratory Terminal Oxidase). The reductive step results in the release of Fe(II) from the Fe(II)FutA2 complex and the subsequent Fe(II) transport across the plasma membrane via the Fe(II) transporter FeoB (Kranzler et al. 2014). The Fe(II)-specific transporter FeoB genes have been identified in freshwater and coastal cyanobacteria, but appear to be absent in oceanic picocyanobacteria and eukaryotic microalgae (Desai et al. 2012). One coastal strain of Synechococcus has genes encoding a Fe(II)-specific transporter FeoB (Palenik et al. 2006). This might be an adaptation to the faster redox cycling of Fe in coastal waters due to photo-reduction of Fe(III) in the presence of hydroxycarboxylic acids (Kuma et al. 1992). Other divalent transporters, such as ZIPs and NRAMPs have been identified in marine picocyanobacteria, and thus may mediate their Fe(II) uptake, as previously demonstrated in other organisms. In oceanic metagenomes, Fe(II) transporters are not very abundant, but ZIPs, NRAMPs are more common than FeoB-type.
Even though Fe reduction and a plasma membrane Fe(II) transporter seem to be the dominant Fe transport mechanism in the model freshwater cyanobacterium Synechocystis (PCC6803), the presence of an Fe(III) transport system, such as FutABC is also expected in freshwater and marine cyanobacteria, given that some of these genes have been identified (Katoh et al. 2001a, b). This transport system consists of a soluble Fe(III)-binding periplasmic protein (FutA2), a permease (FutB), a peripheral plasma membrane associated ATPase (FutC), and an intracellular FutA1 (Fig. 4) (Katoh et al. 2001a, b). The intracellular subunits FutA1 and FutC of this Fe(III) transport system are believed to regulate the reductive Fe(II) uptake pathway (Kranzler et al. 2014). Genomic analyses have revealed the presence of Fe(III)-specific ABC transport systems in many marine cyanobacteria, as well as picocyanobacteria (Desai et al. 2012; Hopkinson and Barbeau 2012), suggesting that Fe(III) uptake is very common, in agreement with the thermodynamic stability of Fe(III) and the prevalence of Fe(III) organic complexing ligands in seawater. In addition, in oceanic metagenomes Fe(III)ABC transport systems are much more common than Fe(II) uptake systems (Desai et al. 2012; Hopkinson and Barbeau 2012).
4 Iron Storage and Luxury Uptake
Iron storage is another strategy that enables microalgae to cope with low and intermittent Fe supplies. Because many of the new Fe inputs to the ocean are sporadic, being able to rapidly take up and store Fe from ephemeral inputs would be particularly advantageous. The capacity to take up more Fe beyond than required to satisfy biochemical functions for maximum growth is often termed luxury uptake and likely an important Fe-acquisition strategy of microalgae, in particular diatoms. The Fe storage capacity can be approximated by the ratio between the intracellular Fe requirements when microalgae are grown in excessively high [Fe] and the cellular concentration in which growth starts to decrease due to Fe-limiting conditions. The resulting ratio reflects a phytoplankter’s potential to store intracellular Fe. Among the phytoplankton examined, the general trend is that oceanic microalgae have lower minimum cellular iron requirements than coastal microalgae (Fig. 7) (Sunda and Huntsman 1995; Maldonado and Price 1996; Marchetti et al. 2006). Microalgae, particularly bloom-forming species, may benefit from being able to take up and store large quantities of Fe during periods of surplus Fe availability because these reserves can then be drawn upon as Fe concentrations become exhausted. The relatively high storage capacities observed in many diatom isolates from the Fe-limited regions of the Equatorial Pacific relative to capacities in other coastal and oceanic diatom isolates were suggested to be an evolutionary adaptation for living in low Fe environments with sporadic Fe inputs (Maldonado and Price 1996).

A comparison of Fe quotas ranges (as measured by the Fe:C ratios) in marine microalgae grown under Fe-limited (low range) and Fe-replete (high range) conditions. Shown are the average ranges for oceanic (o) and coastal (c) isolates of the diatom genera Pseudonitzschia and Thalassiosira, several haptophytes (Emiliania huxleyi, Phaeocystis pouchetti and Chrysochromulina polylepis) and the dinoflagellate Prorocentrum minimum (=Prorocentrum cordatum). The dashed horizontal line represents the Fe:C ratio for phytoplankton from low Fe regions (Derived from Sunda et al. 1991) that is commonly used in marine biogeochemical models (e.g. see Moore et al. 2002) (Data are compiled from Sunda and Huntsman (1995), Maldonado and Price (1996), Marchetti et al. (2006) and Lane et al. (2009))
The Fe storage capacities of pennate diatoms belonging to the genus Pseudonitzschia, a cosmopolitan diatom genus found in both coastal and oceanic environments, have also been examined (Marchetti et al. 2006). Members of this genus are near-universal responders to artificial Fe enrichment in Fe-limited regions of the ocean (Marchetti et al. 2008; Trick et al. 2010). In Fe-limited oceanic waters, Pseudonitzschia abundance is usually low compared to other phytoplankton although species richness may be high. Upon Fe-enrichment, Pseudonitzschia numerically dominates the Fe-induced diatom bloom and can make up a significant proportion of the phytoplankton biomass. The potential Fe storage capacities (estimated by dividing the maximum Fe quota by the minimum Fe quota) for the oceanic Pseudonitzschia spp. examined were markedly higher than those calculated for several oceanic Thalassiosira spp. Differences in Fe:C ratios were primarily a consequence of a higher mean maximum iron quota rather than a lower mean minimum quota (Fig. 7), suggesting that Pseudonitzschia spp. have exceptionally high Fe storage capabilities.
A molecular basis for the enhanced Fe storage in Pseudonitzschia was provided with the identification of ferritin (FTN) genes within several pennate diatoms (Marchetti et al. 2009). Due to its facile redox chemistry and tendency to generate toxic hydroxyl radicals via Fenten chemistry, extensive intracellular concentrations of unchelated Fe could be detrimental to cells. Ferritin is a Fe-storage protein used by plants, animals, cyanobacteria and other microorganisms to safely concentrate and store Fe, thereby minimizing potential cell damage from reactive oxygen species and oxidative stress. In cyanobacteria, there are three different types of ferritin family proteins that have been identified, including bacterioferritins, ferritins and DNA-binding ferritins (DPS proteins) (Kranzler et al. 2013). Ferritins and bacterioferritins oxidize intracellular Fe(II) to Fe(III), thereby functioning as ferroxidases while generating hydrogen peroxide. Bacterioferritins differ from ferritins by the presence of a haeme molecule, anchored at the interface between two adjacent protein subunits. In eukaryotic ferritins (24 subunits, 480 kDa), up to 4500 Fe(III) atoms can be stored as an Fe oxide mineral and releases reduced Fe(II) on cellular demand (Liu and Theil 2005). Such storage compounds allow Fe to be readily released and utilized when needed, thus acting as a buffer against Fe deficiency. Although ferritins appear to be ubiquitously present in green algae and cyanobacteria, the discovery of ferritin in diatoms was the first indication of the Fe storage protein in any member of the Stramenopile species with whole genomes sequenced. Thus, the acquisition of a ferritin gene in diatoms, likely resulted from a horizontal gene transfer event. The diatom ferritin genes contain signal and target peptides that predicted the protein to be targeted to the chloroplast to control the supply of iron for synthesis of critical iron containing proteins in the photosynthetic apparatus (e.g., those in PSII, PSI, and the b6/f complex) (Marchetti et al. 2009).
The absence of a ferritin gene in T. pseudonana in combination with the significantly reduced iron storage capacity of T. oceanica compared to Pseudonitzschia led to the speculation that many diatoms do not contain ferritin. Although centric diatoms lacking ferritin can certainly store Fe (Sunda and Huntsman 1995; Maldonado and Price 1996), the mechanism as to how this is achieved is not well understood. For example, in the centric diatoms T. pseudonana and C. weissflogii, Nuester et al. (2012) observed the elemental stoichiometry of cellular Fe to be consistent with a potential vacuolar Fe storage pool mechanism. Recent transcriptomic sequencing of a number of centric and pennate coastal diatoms has now identified the sporadic presence of ferritin-like genes within some centric diatoms (Groussman et al. 2015). In addition, multiple homologs of FTN genes have been identified in Chlamydomonas reinhardtii and several diatom genomes, including Pseudonitzschia multiseries. Whether these homologs perform different functions is unknown. This patchy diversity of ferritins throughout the diatom lineage necessitates a more systematic examination into the differences in the Fe storage capabilities of diatoms that either contain or lack ferritin.
While the results from bulk studies with natural phytoplankton assemblages suggest that some microalgae can accumulate Fe following pulsed inputs, these experiments do not clarify which species are accumulating the Fe, or whether the accumulated Fe is simply adsorbed onto the outside of the cells or actually accumulated internally. Synchrotron x-ray fluorescence microscopy (SXRF) has provided this missing information, as it enables quantitative elemental analysis of individual cells, as well as spatial information about where the elements are located within the cells (Twining et al. 2003; Twining and Baines 2013). During the SOFeX Fe fertilization experiment in the Southern Ocean, diatoms increased their Fe:C ratios nearly fivefold after two Fe additions, while autotrophic flagellated cells increased their Fe quotas by only threefold (Twining et al. 2004). Additionally, two-dimensional element maps showed that the extra Fe was accumulated largely in the regions of the chloroplasts, indicating that uptake was internal and not the result of abiotic precipitation and adsorption. SXRF has also been used to measure Fe accumulation by microalgae in the Equatorial Pacific. Baines et al. (2011) found Fe quotas of Pseudonitzschia-like diatoms to increase 24-fold in 48-h following addition of 2 nM Fe to deckboard incubations. Further, this Fe was largely localized in Fe storage bodies adjacent to the chloroplasts, as predicted for ferritin (Marchetti et al. 2009). Over the final 2 days of the 96-h incubation, cellular Fe contained within the storage bodies dropped exponentially at a rate approximating the growth of the diatom population, while overall cellular Fe quotas decreased more slowly (Twining et al. 2011). These data demonstrate the potential for stored Fe to support subsequent population growth, as well as the unique ability of SXRF to quantify luxury uptake of Fe by phytoplankton in natural systems.
5 Iron and Light Co-limitation
Light and Fe availability interact in a complex and bi-directional manner, and have a profound effect on the ability of phytoplankton to photosynthesize. Indeed, Fe limitation (especially at low light) limits the biosynthesis of iron-containing components of the photosynthetic apparatus and impairs the capacity of the cells to absorb light energy, as well as convert light energy into chemical energy. Some phytoplankton acclimate to limiting Fe levels by lowering the abundance of the Fe-rich PSI relative to PSII, as previously discussed. High light may enhance Fe availability under low Fe concentrations by promoting photochemical reactions that increase the pool of inorganic Fe ([Feʹ]) in the presence of photolabile organic Fe complexes (Anderson and Morel 1982; Sunda and Huntsman 2011). In addition, under low Fe, high light may enhance Fe uptake by providing more photosynthetically derived NAD(P)H, which is needed for the functioning of the membrane reductases involved in Fe uptake (Askwith et al. 1996).
Phytoplankton acclimate to low photon flux by changing the ratio of accessory pigments to chl a, as well as the abundance and stoichiometry of Fe-rich photosynthetic electron carriers (Falkowski et al. 1981). Under low light conditions, phytoplankton may modify their PSUs to enhance light harvesting by increasing their numbers and/or their size. These two physiological strategies alter distinctively their Fe requirements for photosynthesis. A higher number of PSUs will increase their Fe demand, as the photosynthetic electron transport protein and protein complexes associated with PSUs are Fe-rich. In contrast, an increase in the size of PSU involves higher concentrations of chlorophyll and light-harvesting accessory pigments per PSU, and does not necessarily increase Fe demand (Geider et al. 1993).
Support for the first strategy has been predicted theoretically (Raven 1990) and documented empirically by showing higher intracellular Fe:C ratios needed to support a given iron-limited specific growth rate under low light intensity or shortened photoperiod, both in laboratory cultures (Sunda and Huntsman 1997, 2011; Strzepek and Price 2000; Strzepek and Harrison 2004; Finkel et al. 2006; Weng et al. 2007), and field populations (Maldonado et al. 1999; Hopkinson and Barbeau 2008). These higher Fe quotas are allocated towards the synthesis of additional Fe-rich photosynthetic electron transport protein complexes to maintain high rates of C fixation at low light (Sunda and Huntsman 1997). Indeed, in the case of the diatom C. weissflogii, the relative increase in cellular Fe growth requirements under low light matches the predicted relative increase in abundance of PSI relative to PSII during photoacclimation (Strzepek and Harrison 2004). However, the higher Fe quotas observed under low light are partly a consequence of slower growth rates (pss Fe = Fe quota * growth rate, Sunda and Huntsman 1997, 2011). In addition, significantly higher half-saturation constants for Fe (Kμ) have been shown for cyanobacteria and diatoms grown under low light than under high light (Kudo and Harrison 1997; Bucciarelli et al. 2010). The higher Kμ under low light is partly explained by lower [Feʹ] in a medium with photolabile organic Fe complexes (\( {\left[\mathrm{F}\mathrm{e}'\right]}_{\mathrm{HL}}={\left[\mathrm{F}\mathrm{e}'\right]}_{\mathrm{LL}}*1.14 \), Bucciarelli et al. (2010)). The remaining difference between these Kμs (e.g. for T. oceanica KμHL and KμLL = 1.29 and 3.01 pM, respectively; Bucciarelli et al. (2010)) is likely due to higher Fe demand by low-light-adapted phytoplankton.
The second low-light acclimation strategy, increasing the size of PSUs, has been shown in cyanobacteria, the green alga Dunaliella salina, and Southern Ocean diatoms and haptophytes. Southern Ocean phytoplankton are extremely well adapted to limiting Fe levels and cope with low light by an unprecedented change in the size (sixfold larger) of the effective absorption cross section (σPSII) of their PSII reaction centers, without an increase in cellular Fe (Strzepek et al. 2012). In a similar manner, Fe-limited cyanobacteria have fewer Fe-rich PSIs, but promote the expression of IsiA antenna proteins associated with PSI. This increases the PSI absorption cross-section and maintains rapid electron flow through PSI (Chauhan et al. 2011). Likewise, under low Fe, the green alga D. salina induces the expression of chlorophyll a/b-binding proteins associated with PSI (Varsano et al. 2006). However, this physiological photo-acclimation strategy has its limitations, as the size of the PSU has an optimal maximum, beyond which the energy transfer efficiency between the light harvesting pigments and PSUs decreases sharply (Raven 1990).
6 Iron Limitation Effects on Elemental Composition
Phytoplankton are responsible for approximately half of the carbon fixation on Earth. Thus minor alterations in their elemental stoichiometry from the ideal Redfield composition (i.e. 106C:16N:1P) due to Fe limitation could have large influences on global biogeochemical cycles. Diatoms, in particular account for approximately one-half of marine primary productivity (Nelson et al. 1995). One important effect of Fe limitation on the elemental composition of diatoms is a relative increase in their Si content. A study in the California coastal upwelling region by Hutchins and Bruland (1998) noted a two to threefold increase in the silicic acid-to-nitrate (Si[OH]4:NO3) and silicic acid-to-dissolved inorganic carbon (Si[OH]4:DIC) drawdown ratios of diatom-dominated communities in Fe-limited controls relative to Fe-enriched seawater samples. The observed decrease in the dissolved nutrient drawdown ratios upon the alleviation of Fe limitation was primarily due to the rapid increase in NO3 utilization with little or no change in Si(OH)4 assimilation. This lead to the speculation that Fe-limited diatoms utilize more Si per cell or unit biomass relative to Fe-replete diatoms and thus were more heavily silicified (Boyle 1998). At the same time, in addition to studying the response to Fe enrichments in natural assemblages, Takeda (1998) found that Fe-limited Antarctic diatoms had higher Si:N ratios relative to Fe-replete diatoms. In a Nitzschia sp., the increase in the ratio was due to more Si per cell, whereas in Chaetoceros dichaeta less N per cell drove the increase in the ratio. Since then, additional studies have documented changes in the Si:N and Si(OH)4:NO3 uptake/consumption ratios in Fe-limited diatoms or diatom-dominated natural assemblages and have attributed these changes to enhancements in silicification and/or reductions in N and C, or have suggested that no changes in the ratios occur when both new (NO3-based) and regenerated (NH4-based) forms of N are considered (reviewed by Marchetti and Cassar 2009). In addition, coordinated gene expression responses to Si and Fe limitation within T. pseudonana suggest both elements have a similar influence on diatom cell wall processes (Mock et al. 2008).
Discrepancies in the causal element influencing the Si:N ratios among these laboratory and field studies may be due to varying levels of Fe deficiency and hence reductions in the specific growth rates, differences in growth conditions, implemented methodology, shifts in microalgal species composition and/or the intrinsic variability among different diatoms (reviewed by Marchetti and Cassar 2009). Despite the variety of possible causes behind changes in the element stoichiometries, the observed outcome of Fe-limited diatoms is generally consistent—an increase in the cellular Si:N ratios, although there are exceptions with certain diatoms under severe Fe limitation (Bucciarelli et al. 2010). This result supports the assertion that Fe-limitation is an important factor in contributing to the preferential export of Si relative to N (Dugdale and Wilkerson 1998; Hutchins and Bruland 1998). However, the specific physiological mechanism driving the changes in elemental stoichiometry of diatoms (i.e., an increase in cellular Si content and/or a decrease in both N and C contents) are important for predicting diatom biogeochemical fluxes to the ocean floor. Cell size and the degree of silicification may affect how effectively a certain diatom species can sequester carbon due to the possibility of relatively larger and thicker-shelled diatoms sinking faster (Smayda 1970; Smetacek 1985, 1999; Raven and Waite 2004; Assmy et al. 2013), being less susceptible to grazing (Smetacek 1999; Hamm et al. 2003; Pondaven et al. 2007) and more effectively preserved in ocean sediments.
Although the C:N ratios of phytoplankton are relatively unaffected by nutrient limitation, including Fe, there are reports of a decrease in N:P (and by inference C:P) ratios in Fe-limited phytoplankton. In the diatom C. weissflogii, this decrease was primarily a result of accumulation of P under low-Fe conditions due to a more rapid decline in specific growth rate than in steady-state uptake rate of phosphorus (Price 2005). Under the most severe Fe-limiting conditions, C. weissflogii contained 1.5 times more P per liter cell volume than when Fe-replete. The trend observed in the laboratory isolate is supported by field observations from Fe-limited natural assemblages dominated by diatoms, where lower N:P and C:P ratios have been observed. Thus, as Fe inputs to Fe-limited regions around the globe vary in space and over geological timescales, considerable changes in the elemental composition of sinking phytoplankton debris would be anticipated.
Iron status may also influence the stoichiometry of other trace elements. For example, as previously discussed, diatoms that contain a multi-copper oxidase as part of their high affinity Fe uptake pathway have an increased demand for copper when this pathway is induced under low Fe-limiting conditions (Peers et al. 2005; Maldonado et al. 2006). Even under Fe-replete conditions, the copper requirements of many oceanic diatoms are higher due to their dependence on the copper-containing electron transport protein plastocyanin, whereas many coastal diatoms use cytochrome c6, which is a Fe-containing protein (Peers and Price 2006).
7 Phytoplankton Response to Iron Fertilization
In vast expanses of our world’s oceans, the biomass of phytoplankton is low even though macronutrient concentrations are quite high. These areas are termed “high nutrient (nitrate), low chlorophyll” or HNLC regions. Until recently, the factors most responsible for minimizing new production in HNLC regions remained unknown, forcing oceanographers to speculate about possible causes. Competing hypotheses that emerged in the late 1980s attempted to explain these paradoxical HNLC regions. As reviewed by Cullen (1991), factors believed to control primary productivity from a “bottom-up” approach included the effects of low temperature (Tilzer et al. 1986), light limitation and/or water column instability and strong turbulence (Chavez et al. 1991; Miller et al. 1991; Mitchell et al. 1991), “shift-up” in NO3 assimilation (Dugdale and Wilkerson 1991) and micronutrient (Fe) limitation (Martin and Fitzwater 1988). “Top-down” or “grazer” control was also suggested as the primary factor regulating phytoplankton biomass in these oceanic regions, as these regions were dominated by small-celled phytoplankton that were rapidly grazed by microzooplankton (McAllister et al. 1960; Banse 1991; Miller et al. 1991). The possibility that the regulation of primary productivity in HNLC regions may be a combination of all these factors rather than a single one was also considered (Harrison et al. 2004).
Martin and Fitzwater (1988) proposed that low Fe concentrations may limit growth of phytoplankton in HNLC regions of the ocean. Although early investigators had proposed the possibility of Fe limitation (Hart 1934), supportive experimental data was difficult to obtain due to the lack of trace metal clean sampling methods that inevitably resulted in Fe contamination. Martin et al. (1989) showed that surface concentrations of dissolved Fe in these HNLC regions were very low (<<1 nM) and dissolved Fe exhibited a nutrient-like profile. Furthermore, bottle experiments consisting of the addition of Fe to the HNLC Subarctic Pacific water samples resulted in an increase in chlorophyll a and a subsequent drawdown of NO3 (Martin and Fitzwater 1988). In particular, the growth rate of large phytoplankton increased, resulting in a species composition shift from cyanobacteria and small flagellates to large diatoms. These experiments provided compelling evidence that Fe may indeed limit phytoplankton growth and validated the “Iron Hypothesis” (Martin 1990). The idea that iron limits the growth of phytoplankton in the vast HNLC regions of the ocean was confirmed convincingly by a series of in-situ iron fertilization experiments conducted in patches of surface seawater tens of km in surface area in major HNLC regions (see discussion below). There are three main regions of the world’s oceans where the phytoplankton have been shown to be consistently or periodically limited by the supply of Fe. These include parts of the Equatorial Pacific, the Subarctic Pacific and most of the Southern Ocean (de Baar et al. 2005; Boyd et al. 2007). There is also compelling evidence for Fe limitation of phytoplankton in coastal upwelling regions (Hutchins et al. 1998; Firme et al. 2003), as well as seasonal Fe limitation primarily of N2 fixation rates in tropical and subtropical regions the Pacific and South Atlantic Oceans receiving low inputs of iron from continental source (Sohm et al. 2011). In addition, iron-light co-limitation has been suggested to limit phytoplankton growth and influence phytoplankton community composition in the deep chlorophyll maximum at the bottom of the euphotic zone in vast regions of thermally stratified subtropical ocean waters, where both light and iron concentrations are very low (Sunda and Huntsman 1997, 2015; Hopkinson and Barbeau 2008).
The confirmation that phytoplankton growth is limited by a trace metal that is very abundant on land has led to the concept of performing mesoscale open ocean Fe enrichment experiments to mitigate increases in atmospheric CO2 caused by the burning of fossil fuels. In addition, since the molar stoichiometry of most phytoplankton is ~105 C:1Fe, compared to ~7C:1N, it is considerably very economical to add Fe to large areas of the ocean rather than N to stimulate algal growth. Though met with considerable debate and skepticism by many scientists (Strong et al. 2009), human-induced Fe enrichment is viewed by some as a possible solution to dealing with climate change and reductions in commercial fish stocks. The unprecedented rate of increase in atmospheric levels of CO2 is considered to be the major contributor to global warming, although methane, nitrous oxides and chlorofluorocarbons also play a role. A number of such Fe enrichment experiments have been performed in the equatorial Pacific (IronEx I and IronEx II), in the Southern Ocean (SOIREE, EisenEx, SOFex, Eifex and Lohafex) and North Pacific (SEEDS I and II and SERIES) (Fig. 8) (Boyd et al. 2007; Smetacek et al. 2012). Each of these experiments demonstrated varying degrees of increased phytoplankton growth in response to Fe additions to surface waters that were dependent on the initial environmental conditions and the initial phytoplankton community composition. With the exception of a few experiments, Fe enrichments resulted in increased diatom growth that subsequently partially or fully depleted macronutrient concentrations leading to bloom termination. The resulting diatom blooms are often large enough to be visible form space (Fig. 8).

(a) Map of global surface NO3 concentrations along with the locations of 12 Fe enrichment experiments (indicated by yellow circles). 1 IronEx-I (1993), 2 IronEx-II (1995), 3 SOIREE (1999), 4 EisenEx (2000), 5 SEEDS-I (2001), 6 SERIES (2002), 7 SOFeX North (2002), 8 SOFeX South (2002), 9 SEEDS-II (2004), 10 EIFEX (2004), 11 SAGE (2004), 12 LOHAFEX (2009). Also indicated is an Fe addition experiment performed by the Haida Salmon Restoration Corporation in 2012 that was not government approved (red circle). NO3 are World Ocean Atlas climatological spring values. (b) SeaWiFs satellite image of the phytoplankton bloom resulting from the SERIES Fe enrichment experiment in the northeast Pacific Ocean (indicated by the number 6 in panel a). The coast of Alaska is shown. Warm colors (reds and yellows) indicate high concentrations of chlorophyll a and thus high phytoplankton biomass; cool colors (blues) indicate low chlorophyll a concentrations. Dark areas over the ocean result from cloud cover. The white-circled region indicates the 700 km2 region of high chlorophyll a concentrations resulting from the addition of Fe. (c) Light microscope image of the oceanic diatom Pseudonitzschia granii; members of this genus are common responders to Fe enrichment in all HNLC regions. Scale bar is 10 μm (Adapted from figures in Behrenfeld et al. (2009), Armbrust (2009) and Morrisey and Bowler (2012). SERIES satellite image courtesy of J. Gower, Orbimage/NASA)
Following the initial Fe enrichment studies performed in each of the well-characterized HNLC regions, subsequent rounds of large-scale in-situ Fe addition experiments have focused on the Southern Ocean, as it is the largest of the HNLC regions containing a vast pool of underutilized macronutrients. The SOFeX experiments added Fe to two separate north and south patches, with the north patch containing low Si(OH)4 concentrations and the south patch containing high Si(OH)4 concentrations (Coale et al. 2004). Although an increase in phytoplankton biomass was observed in both patches, the phytoplankton assemblage differed, as the north patch contained a significantly lower proportion of diatoms due to Si(OH)4 limitation. During Eifex, Fe was added to a vertically coherent, mesoscale eddy of the Antarctic Circumpolar Current (Smetacek et al. 2012). An ensuing large diatom bloom was monitored for 5 weeks and at least half of the phytoplankton bloom biomass sank to a depth of 1000 m with a substantial portion likely reached the sea floor. Similar to natural Fe fertilization observations in the Southern Ocean (Blain et al. 2007; Pollard et al. 2009), Eifex verified a substantial deep carbon export associated with the ensuing Fe-induced diatom bloom.
A molecular basis for the phytoplankton response to Fe enrichment and their mechanism of recovery was achieved through a comparative metatranscriptome analysis of a Fe-limited plankton community in HNLC waters of the NE Pacific Ocean (Marchetti et al. 2012). Consistent with the large-scale Fe enrichment experiments, a large diatom bloom ensued after Fe addition to mesocosms. The immediate diatom response to Fe enrichment was to continue to express genes encoding non-Fe containing proteins rather than carrying out a widespread replacement of these proteins with their Fe-containing counterparts. For example, transcripts associated with the Fe-free electron transfer protein flavodoxin were abundant in all libraries and over-represented after Fe enrichment, suggesting that some or all diatoms continue relying on this protein for photosynthetic electron transfer rather than switching entirely to use the Fe-containing version, ferredoxin. Additionally, diatom transcripts for copper-containing plastocyanin were highly abundant under all conditions whereas very few transcripts for cytochrome c6, the Fe-requiring counterpart, were readily identified. Many oceanic phytoplankton have likely adapted to life under continuous iron-limitation by permanently using the iron-free equivalents of these proteins and may have lost the genes to synthesize ferredoxin and cytochrome c6. Continued dependence on Fe-free proteins may also provide diatoms with a more rapid acclimation to a return to low-Fe conditions.
The addition of Fe to diatoms stimulated what appears to be a system-wide pattern of gene expression in alleviation of chlorosis and preparation for rapid cell division. Specifically, significant increases were observed in transcripts for genes encoding proteins that catalyze the biosynthesis of nucleic acids, amino acids, sugars, chlorophyll (and its precursors, i.e. porphyrin) as well as the prerequisites for long-chain polyamines, which may play an important role in silica deposition in diatom cell wall formation, were observed. Components of the diatom urea cycle were suggested to likely play a significant role in facilitating the Fe response leading to bloom formation following the alleviation of Fe limitation and/or low-Fe-induced NO3 limitation. The ability of diatoms to divert their newly acquired Fe towards nitrate assimilation may underlie why diatoms consistently dominate Fe enrichments in HNLC regions. The distinctiveness of the diatom response was even more apparent when compared to the haptophytes, the other dominant member of the phytoplankton community after Fe enrichment, which displayed a more typical response to Fe enrichment (i.e. increased transcripts of ferredoxin, Fe-SODs and other Fe-requiring protein-encoding genes). In addition, rhodopsin transcripts assigned to diatoms, haptophytes and dinoflagellates were highly abundant in the Fe-limited libraries, and decreased in abundance following Fe enrichment. Rhodopsins are light-driven proton pumps that may be associated with ATP generation (Beja et al. 2000; Fuhrman et al. 2008), which suggested a role for rhodopsins in eukaryotic phytoplankton to deal with low-Fe conditions (Marchetti et al. 2015).
8 Climate Change Effects on Iron-Related Phytoplankton Physiology
As the amount of CO2 in the atmosphere continues to increase, equilibration of higher atmospheric CO2 levels with the surface ocean will acidify ocean waters and decrease their pH, typically referred to as ocean acidification. An extensive amount of research has been conducted examining ocean acidification effects on the calcification processes of marine organisms, including microalgae (e.g. coccolithophores) (for a review, see Doney et al. 2009). As seawater pH is reduced, the most prominent effect is a decrease in the carbonate ion concentration that could impede calcification rates, although the responses of marine organisms appear to be varied. Less is known about other potential effects of ocean acidification and warming on marine ecosystems, including how the speciation and distributions of trace metals such as Fe will be affected (Hoffmann et al. 2012). In addition to changes in carbonate chemistry (i.e. a reduction in carbonate ion concentrations), a decrease in pH will result in a decrease in hydroxide ion concentrations in most natural surface waters. Decreases in pH may have a profound influence on the concentrations and chemical speciation of iron, and hence on iron availability to phytoplankton due to a number of factors: the existence of dissolved inorganic iron (FeIII)’) in seawater as soluble iron hydroxide species, the polymerization and precipitation of iron hydroxides (which removes iron from sea water), and to the fact that many organic complexation reactions are sensitive to changes in pH (Shi et al. 2010). In addition decreasing pH will promote rates of reduction of Fe(III) to soluble Fe(II), and retard the reoxidation of Fe(II) to Fe(III) (Millero et al. 2009).
To some degree, potential increases in Fe solubility along with proposed changes in kinetics could lead to increased iron concentrations and hence in primary productivity in Fe-limited regions in the ocean. However as the vast majority of Fediss is bound to strong organic ligands (Gledhill and van den Berg 1994; Rue and Bruland 1995), the changes in complexation of Fe with these organic ligands at a reduced pH may largely drive how ocean acidification affects Fe availability. The effects of reduced pH on Fe availability to marine phytoplankton when Fe is bound to a variety of organic ligands were recently examined (Shi et al. 2010). In all examined species, which included both coastal and oceanic diatoms as well as the coccolithophore E. huxleyi, Fe uptake rates were related to Feʹ, indicating that the effect of pH is largely due to a change in the chemical speciation of Fe and not a physiological response of the organisms. When Fe was bound to chelators with acidic binding groups that are not protonated in seawater, the extent of decrease in Feʹ with decreasing pH is dependent on the number of protons released upon dissociation of Fe. Likewise, if the uptake of Fe is dependent on enzymatic reduction of the Fe-chelator complex, a decrease in pH could affect rates of the enzymatic reaction if protons are liberated. This was proposed to be the case for a decrease in Fe uptake rates with decreasing pH when Fe is bound to the trihydroxomate siderophore, DFB (Shi et al. 2010). In contrast, there was a negligible effect of pH on Fe uptake when Fe was bound to chelators that are protonated in seawater (e.g. catechols). Thus, in general, it was determined that the availability of Fe to phytoplankton decreases at lower pH and are largely dictated by the acid-base chemistry of the chelating ligands.
Field experiments have shown mixed responses to alteration in CO2 and Fe concentrations within natural phytoplankton assemblages from HNLC regions. In Fe-limited waters within the Gulf of Alaska, a marginal increase in biomass and production was observed at high CO2 and was attributed to the energy savings associated with a down-regulation of the carbon-concentrating mechanisms (CCM) and, in some cases, a reduction of Fe-rich photosynthetic proteins (Hopkinson et al. 2010). Alternatively, under Fe-enriched conditions, increases in growth and biomass at high CO2 compared to low CO2 treatments were suggested to be the result of lower rates of respiration, leading to reductions in carbon loss. Similarly, in coastal waters off Norway, at high CO2 levels, increases in Fediss and Fe(II) concentrations were observed within mesocosm experiments, suggesting increased Fe availability as a result of CO2 enrichment (Breitbarth et al. 2010). In contrast, experiments conducted with natural seawater samples in coastal and oceanic waters in the North Atlantic (Shi et al. 2010) and Fe-limited waters in the Bering Sea (Sugie et al. 2013) inferred decreases in Fe uptake rates and growth of phytoplankton with increasing CO2 levels. In the Bering Sea study, these CO2-mediated influences disappeared under Fe-replete conditions. Such discrepancies could be due to a number of factors, including differences in the Fe concentrations, Fe speciation and the phytoplankton community composition.
Apart from affecting Fe availability, interactive effects of ocean acidification and Fe have been shown to influence other aspects of microalgal physiology and community dynamics. For example, in the coastal diatom Pseudonitzschia pseudodelicatissima, Si:C and Si:N elemental ratios decreased, whereas N:P ratios increased with increasing pCO2 (Sugie and Yoshimura 2013). Thus, in a high CO2 world, these patterns could buffer the changes in elemental ratios that occur when diatoms are Fe-limited, which tend to be in the opposite direction. Within Fe-enriched natural assemblages present in the Weddell Sea, Antarctica, CO2-dependent changes in productivity were accompanied by a taxonomic transition from weakly to heavily silicified diatoms (Hoppe et al. 2013). These transitions did not occur under Fe-limiting conditions, suggesting differences in carbon acquisition strategies and/or pH-mediated cellular physiologies among diatoms that were only evident after the alleviation of Fe stress.
Although our knowledge of ocean acidification effects on cyanobacteria and microalgae has advanced tremendously over the last decade, ocean acidification is only one of multiple factors that will be altered within marine systems as a consequence of elevated atmospheric CO2 levels and resultant climate change. Significant warming of the land and oceans are also projected to occur which will result in changes to water column stratification, salinity and ocean circulation (Doney et al. 2012). In addition, changes in nutrient content and oxygen input to the ocean surface waters are predicted. All of these factors will undoubtedly effect Fe distributions and speciation, perhaps more so than ocean acidification, which will then have downstream biological consequences.
9 Techniques for Studying Iron-Related Microalgal Physiology
9.1 Achieving Iron Limitation in the Laboratory
One of the primary means by which physiologists investigate ways in which microalgae acclimate to varying Fe availabilities is by attaining steady-state Fe-limitation of growth rates in laboratory cultures. Achieving steady-state Fe limitation is now common practice provided that there is access to trace metal clean culturing facilities and implementation of certain culturing protocols (Price et al. 1988/89; Sunda et al. 2005). These protocols include the use of chemical-resistant plastics that contain lower concentrations of trace elements for culturing (such as polycarbonate, low-density polyethylene and teflon), acid-cleaning of all culture tubes and bottles, chelexing of the artificial seawater media—to remove trace metal impurities—and performing all preparation of medium and culture transfers within a trace metal clean laminar flow hood. Either synthetic ocean water (i.e. Aquil or ESAW; Harrison and Berges 2005) or low-Fe natural seawater can be used as growth medium. In most seawater medium recipes, synthetic aminocarboxylate chelating agents, such as EDTA are commonly used to regulate trace metal speciation. Concentrations of EDTA vary among the different types of medium, but typically range between 5 and 100 μM. In the presence of high concentrations of EDTA, virtually all of the iron in the medium is chelated by this ligand, and an insignificant fraction of Fe is bound to strong natural organic ligands (in cases where natural seawater is used); hence binding by the latter are unlikely to have a significant influence on metal speciation. In addition, without EDTA, [Fe]diss exceeding ca. 0.07 nM is expected to precipitate and is mostly unavailable to phytoplankton (Johnson et al. 1997), whereas in the presence of EDTA, Feʹ concentrations can reach up to 0.7 nM before exceeding the empirically observed threshold for precipitation of Fe hydroxides (Sunda and Huntsman 1995). By adjusting the concentrations of Fe and EDTA in the culture medium, the Feʹ concentrations can be lowered to the point where the availability of Fe regulates the growth rate of the phytoplankton. As described previously, EDTA will continuously buffer the concentration of Feʹ until the biomass becomes sufficiently high to increase culture pH (which decreases iron complexation in EDTA buffers in seawater) or the cells take up Feʹ from the medium at a faster rate than it is resupplied from dissociation of Fe-EDTA chelates (which causes Feʹ concentrations to decrease) (Sunda et al. 2005). Thus, to maintain a constant Feʹ concentration in the culture it is essential to not let the culture biomass get too high. The total Fe, Feʹ and Fe(III) concentrations reportedly used in laboratory cultures to achieve Fe replete (e.g. pFe 18 and 19) and Fe-limited (pFe >20.5) growth of microalgae are listed in Fig. 9.

Predicted dissolved inorganic Fe (Fe´), free ferric Fe (Fe(III)) concentrations, and pFe values for growth mediums as a function of total Fe concentrations. The conditional stability constants used to predict Fe´ and Fe(III) were obtained from Sunda et al. (2005), with a 100 μM EDTA metal ion buffer system in seawater at 20 °C, pH of 8.2, and salinity of 36. pFe values are the -log[Fe(III)]. At Fe´ above ~750 pM (black bar), significant iron hydroxide precipitation is likely to occur (Sunda and Huntsman 1995). For simplicity, the Fe´ and Fe(III) are also shown above this maximum
Although useful for laboratory studies, the relevance of using synthetic chelators such as EDTA to buffer trace metals within culture medium to determine the Fe requirements and Fe-related physiology of microalgae has limitations. As previously discussed, the use of large amounts of EDTA results in the bioavailability of Fe to be primarily controlled by concentrations of Feʹ. Yet our current understanding is that the dissolved Fe pool in the ocean is predominantly bound by natural organic ligands and the computed equilibrium Feʹ concentrations are too low to support the observed growth of phytoplankton (Gledhill and van den Berg 1994; Rue and Bruland 1995). This observation suggests that some of these natural organic Fe complexes are bioavailable (e.g. Strzepek et al. 2011). The question can then be posed whether EDTA provides for an accurate representation of these natural organic ligands (Muggli and Harrison 1996a)? Although it is unlikely, it is difficult (but certainly not impossible) to perform Fe physiological investigations without the use of such chelators when growing phytoplankton in natural low-Fe seawater unamended with additional Fe complexes. Therefore, there are only a few studies examining Fe physiology without the use of synthetic chelators such as EDTA (e.g. Timmermans et al. 2001, 2004). Alternatively, natural organic ligands such as DFB can be used (see section below). If the objective is to grow phytoplankton without the use of such chelators, practicing strict trace metal clean procedures, in addition to quantifying and characterizing the naturally present organic and Fe ligand pools are essential.
9.2 Probing the Iron Nutritional Status of Natural Phytoplankton
The physiological state of phytoplankton with regards to Fe nutrition can be assessed by measuring rates of Fe uptake in the presence of DFB or EDTA using Fe radionuclides (i.e. 55Fe or 59Fe). The transport of Fe in the presence of excess DFB at sub-saturating Fe concentrations is indicative of Fe limitation. In contrast, the transport of Fe at saturating concentrations, achieved by nM additions of Fe bound to EDTA (to prevent Fe precipitation), are a measure of the number of Fe transporters at the cell surface. The reason is that at saturating Fe levels, maximum rates of Fe transport are observed and are determined by the number of Fe transporters at the cell surface and the rate of internalization, so that \( {\rho}_{\max }={\mathrm{k}}_{\mathrm{in}\ *}{{\mathrm{L}}_{\mathrm{T}}}^{\max } \).
9.2.1 Determining Sub-saturating Rates of Iron Uptake in the Presence of DFB
The hydroxamate siderophore desferrioxamine B (DFB) has a conditional stability constant for Fe similar to those of in situ strong organic ligands (Rue and Bruland 1995). Most recently, hydroxamate siderophores have been identified and quantified in the Atlantic, and seem to be widely distributed in the ocean (Mawji et al. 2008). Therefore, DFB is good model for the strong naturally occurring organic ligands for Fe in seawater. Many studies (e.g. Hutchins et al. 1999a; Eldridge et al. 2004; Taylor et al. 2013) have used DFB to induce Fe limitation in bottle incubations, by adding high concentrations of DFB that significantly decrease the concentration of inorganic Fe and result in an excess of free DFB. Whether additions of DFB induce Fe limitation depends on the molar ratio of [Fe] to [DFB]. If the concentration of DFB greatly exceeds that of Fe, the free DFB competes directly with the cell surface Fe transport complexes for the biologically generated free Fe, and thus the ability of the cells to take up Fe is hampered, and results in Fe-limited growth. For example, many oceanic and Southern Ocean species are able to grow at maximum rates when Fe and DFB are at equimolar concentrations (e.g. 4 nM each), while coastal species are Fe-limited in these growth conditions. As the concentration of DFB is increased, oceanic and Southern Ocean species start to experience Fe limitation to various degrees. For example, T. oceanica grew at 50 % of μmax in the presence of 4 nM Fe and 40 nM DFB, while to achieve similar reductions in growth rates in Southern Ocean isolates (Phaeocystis antarctica and Proboscia inermis) additions of 400 nM DFB (bound to 4 nM Fe) were needed (Strzepek et al. 2011). In another study, T. oceanica did not achieve Fe limitation at Fe:DFB ratios of 12.9 nM Fe: 1.29 μM DFB, nor at ratios of 40 nM Fe: 4 μM DFB. However at ratios of 120 nM Fe:12 μM DFB, a 50 % reduction in growth rates was observed (Maldonado and Price 2001). In the Arctic, at in situ [Fe]diss = 0.15 nM, Fe limitation was induced with additions of 1 nM DFB (Taylor et al. 2013). In the Weddell Sea, at in situ [Fe]diss = 1.12 nM, Fe limitation was induced with a 10 nM DFB addition (Hoppe et al. 2013). Therefore, it is essential to know the initial concentration of dissolved Fe when conducting DFB addition experiments. As a rule of thumb, a DFB addition at a concentration 10- to 20-fold in excess of that of Fe should be sufficient to induce Fe limitation.
However, Fe uptake in the presence of DFB can also be used to test the Fe-limited conditions of phytoplankton growth. This is due to the fact that phytoplankton are only able to acquire significant amounts of Fe bound within FeDFB when their growth rates are Fe-limited, and thus, have up-regulated their high-affinity Fe transport system (Maldonado and Price 1999; Strzepek et al. 2011). Although the rates of Fe uptake in the presence of FeDFB are orders of magnitude slower than those in the presence of only inorganic Fe or FeEDTA, these FeDFB uptake rates can be used as a proxy for the Fe-limited condition of the phytoplankton (Maldonado and Price 1999). Thus, the rates of 55Fe uptake from 55FeDFB (ρFeDFB) can be determined in order to establish whether plankton in the various treatments or locations are Fe-limited. The addition of Fe and DFB can vary depending on the study. For example, we have used mixtures of 2 nM Fe and 20 nM DFB (100 % of the Fe as 55Fe, Maldonado and Price 1999; Maldonado et al. 2005), or 100 nM Fe and 105 nM DFB (5 % of the Fe as 55Fe, Taylor et al. 2013). Another advantage of using DFB for Fe uptake experiments in situ is that because DFB is a strong Fe-binding ligand (KFe´L cond = 1016.5 M−1) (Rue and Bruland 1995) the addition of FeDFB to seawater does not significantly increase the in situ Feʹ concentration over the course of the 24 h uptake assay. For example, if natural seawater has an [Feʹ] of 1.5 * 10−12 M, and an Fe uptake experiment is performed with additions of 100 nM Fe and 105 nM DFB, the estimated [Feʹ] resulting for the FeDFB addition is ~6.3 * 10−16 M, well below the in situ [Feʹ]. This calculation was performed using \( \left[\mathrm{F}\mathrm{e}'\right]=\left[\mathrm{Fe}\mathrm{DFB}\right]/\left(\left[\mathrm{D}\mathrm{F}\mathrm{B}'\right]\ *{{\mathrm{K}}_{\mathrm{Fe}'\mathrm{L}}}^{\mathrm{cond}}\right) \), where [DFB’] = [DFB]free, and KFeʹL cond = 1016.5 M−1, (Rue and Bruland 1995).
To perform these Fe uptake experiments, before FeDFB is added it to the 1 L incubation bottles, the cold Fe and the 55Fe (specific activity 740–1480 MBq mg−1) must be complexed to DFB in small aliquots of Milli-Q water. All the manipulations must be performed inside a laminar flow hood. The pH of the 55Fe stock solution (~0.11 mM 55Fe in 0.005 M HCl) used to prepare the Fe complexes should be below 3, so that all of the Fe is truly dissolved. The DFB stock solution is prepared in sterile, Milli-Q water, and is kept refrigerated to prevent fungal growth. Complete complexation of the Fe to DFB is ensured by allowing the DFB to react with the Fe for 5–8 h at pH 3.3 (in 18.2 MΩ, MilliQ-H2O with 0.0005 M HCl). At pH > 3, FeDFB is completely coordinated and very stable with respect to dissociation (Monzyk and Crumbliss 1982). Indeed, at pH 3.3 the concentration of DFB is 1013 times higher than that of DFBH+, so most of the DFB is free and available to bind to Fe(III) (Maldonado and Price 2001). After FeDFB complexation has been achieved, the FeDFB can be added to triplicate 1 L polycarbonate bottles of seawater. The bottles are then capped and sealed with Parafilm, and then placed on-deck incubators at natural light levels and temperature. Every 4 h, samples are drawn from the incubation bottles, collected on polycarbonate filters of varying porosity and rinsed with Ti(III) citrate EDTA reagent (Hudson and Morel 1989) or oxalate wash (Tang and Morel 2006) to reductively dissolve iron hydroxides and surface adsorbed iron species. Control samples should be poisoned with glutaraldehyde (0.5 ml of 25 % glutaraldehyde L−1) and then processed in an identical manner to that of the samples. These controls are used to correct for abiotic Fe uptake and 55Fe adsorption by the filters, and normally represent <5 % of the total Fe uptake by the living cells.
Adding nM concentrations of Fe in the FeDFB uptake experiments (or FeEDTA, see below) allows Fe uptake rates to be calculated without a priori knowledge of the in situ Fe concentration, at least in most open ocean surface waters where [Fe]diss ~0.07–0.3 nM (Johnson et al. 1997). Volumetric 55Fe uptake rates (mol L−1 h−1) are then calculated from the linear regression of particulate 55Fe concentration as a function of incubation time. These 55Fe uptake rates are then converted to total Fe uptake rates using the ratio of dissolved concentrations of 55Fe to cold Fe in the experimental Fe uptake bottles. In field Fe uptake studies, the volumetric Fe uptake rates can be normalized to phytoplankton biomass using chlorophyll as a proxy for biomass. These chl a-normalized rates can then be normalized to C using published C to chl-a ratios (Geider 1987). In laboratory Fe uptake experiments, the rates of Fe transport (ρFe) should be normalized to cellular surface area. Reporting Fe uptake rates as mol Fe μm−2 h−1 allows easy comparison between species of different sizes, or between Fe-limited growth treatments of the same species (given that a reduced cell size is a common acclimation to Fe limitation). In addition, Shaked and Lis (2012) suggested normalizing sub-saturating Fe uptake rates to the concentration of dissolved Fe, given that sub-saturating Fe uptake rates by a cell vary with Fe concentration. The resulting parameter is the uptake rate constant (k up in Shaked and Lis 2012; or k in in Lis et al. 2015), and has units of L cell−1 h−1 or L μm−2 h−1, depending on whether the ρFe are, in the first place, expressed as cellular rates or normalized to surface area, respectively. Therefore, k up (or k in) allow easy comparisons among multiple studies that used various sub-saturating dissolved Fe concentrations. Indeed, Lis et al. (2015) compared the uptake rate constants (k in) of many Fe-limited phytoplankton and discovered a tight correlation and proportionality between their k in and their respective surface areas, suggesting a physical constraint on the number of transporters at the cell surface allocated toward Fe uptake, as proposed earlier by Hudson and Morel (1993). Interestingly the k in for Feʹ was 1000-fold higher than the k in for strong organic complexes like FeDFB. Furthermore, many other Fe substrates have k in between the values for Feʹ and FeDFB. This finding suggests that these two substrates (Feʹ and FeDFB) provide an empirical range for the bioavailability of all Fe substrates for uptake by phytoplankton.
Lis et al. (2015) argued that the ~1000-fold difference in the k in between Feʹ and FeDFB is most likely due to large difference (100–1000-fold) between Fe reduction rates of Feʹ versus FeDFB. However, the limited data sets available suggest that biological reduction rates of inorganic and organic Fe complexes do not differ greatly (~2 fold; Weger 1999; Maldonado and Price 2001). So, the question of why there is a 1000-fold difference between the Kin for Feʹ and that of FeDFB, remains unresolved. It is possible that the difference reported in Lis et al. (2015) is an artifact of the way k in was calculated for FeEDTA uptake experiments (ρFe/[Feʹ]) versus FeDFB experiments (ρFe/[FeDFB]).
9.2.2 Determining Saturating Rates of Iron Uptake in the Presence of EDTA
Short-term Fe uptake rates from 55FeEDTA (ρFeEDTA) can be determined to investigate saturating rates of Fe uptake, which are a direct function of the density of plasmalemma-bound Fe transporters, and are faster for Fe-limited phytoplankton (Harrison and Morel 1986). The experiments are performed in an identical manner to those of FeDFB, except that the Fe is added at saturating concentrations and is prechelated to EDTA before it is added to the water as a FeEDTA complex. We have used various concentrations of Fe:EDTA, from 20 nM Fe bound to 500 nM EDTA (5 % of the Fe as 55Fe; Maldonado and Price 1999), to 100 nM Fe bound to 1 μM EDTA (1 % of the Fe as 55Fe; Taylor et al. 2013). The high concentrations of EDTA in the Taylor et al. (2013) experiment were chosen so that most of Fe in the system was bound to EDTA and there was very little competitive binding of Fe by in situ natural ligands or DFB (in the induced Fe-limiting treatment bottles). As a general rule, the lower concentrations of EDTA used in Maldonado and Price (1999) are recommended to minimize EDTA chelation of other essential trace metals such as zinc.
10 Conclusions and Future Directions
Our knowledge of the important roles that iron plays within phytoplankton and the vast number of acquisition strategies invoked to cope with a limited Fe supply has increased substantially since the discovery of widespread Fe limitation in aquatic environments. Over the last century, we have gained an appreciation for the incredible amount of genetic, physiological and species diversity that exists within phytoplankton communities (Andersen 1992). Yet despite this diversity, much of our understanding of microalgal physiology is based on only a handful of model species, with only a subset of these having their full genomes sequenced. Over the next decade, we anticipate a substantial increase in the number of sequenced genomes of phytoplankton, covering all taxonomic groups, both prokaryotic and eukaryotic. This will certainly bring to light the complexity and variety that exists among phytoplankton with respect to the distribution of Fe-requiring proteins, uptake and storage strategies, and ways in which they have evolved to cope with Fe limitation. In addition, the number of studies implementing the ‘omic’ approaches (i.e., genomics, transcriptomics, proteomics and metabolomics) to examine both laboratory isolates and natural assemblages will continue to increase. Their findings will provide new insights into how phytoplankton interact with their biotic and abiotic environments and how Fe-related processes are affected by these interactions. Clearly, we are just at the beginning of a molecular era in phytoplankton research.
Another growing area of research is investigations into the degree to which microalgae experience co-limitation between Fe and other essential elements and compounds required for growth. Although it has been well acknowledged that Fe and light may be co-limiting for phytoplankton growth in their natural environment, other types of co-limitation exist (Saito et al. 2008) and may be widespread. For example, recent studies suggest that in certain HNLC regions, there is co-limitation of microalgal growth by Fe and vitamin B12. Additions of both Fe and vitamin B12 have been shown to either further stimulate the growth of entire phytoplankton communities (Bertrand et al. 2007) or a subset of species, resulting in shifts in community composition when vitamins are added in addition to Fe (Koch et al. 2011). There have also been suggestions of co-limitation of phytoplankton by Fe and other nutrients: N (Behrenfeld et al. 2006; North et al. 2007; Taylor et al. 2013), P (Mills et al. 2004), Si (Marchetti et al. 2010; Brzezinski et al. 2011), as well as other essential trace elements including Cu (Guo et al. 2012), Co (Saito et al. 2005) and Zn (Crawford et al. 2003). Our current understanding of how such co-limitations will impact Fe-related phytoplankton physiology is still limited.
It has been over two decades since John Martin and others provided compelling evidence of widespread Fe limitation in the sea, indicating that Fe regulates the primary productivity of large areas of the ocean and suggesting that it has a profound effect on global climate. He will forever be remembered for proclaiming, “Give me a tanker of Fe and I’ll give you an ice age”. With the growing concerns over climate change, large-scale Fe fertilization continues to be considered as a geo-engineering strategy to reduce atmospheric CO2 concentrations. To be successful, the efficiency in which an increased supply of Fe has the ability to result carbon transport to the deep ocean and thus long term carbon sequestration would need to be well constrained. However, an important lesson from the previous era of large-scale Fe fertilization experiments is that there is extensive biological, chemical and physical variability within the ocean and Fe-limited regions are no exception. With every Fe-stimulated phytoplankton bloom, whether it be by natural or anthropogenic causes, the phytoplankton response and associated carbon export is influenced by the entire plankton community composition, their degree of Fe limitation, the ambient nutrient concentrations as well as a myriad of other variables; all of which can change on very small spatial and short temporal scales making the outcome somewhat unpredictable.
How future climate change will affect Fe-related processes in phytoplankton communities is presently uncertain. As the ocean environment responds to the increasing effects of ocean acidification and warming, there are many unknowns as to how phytoplankton will acclimate or adapt. If ocean acidification affects Fe distributions and speciation as predicted, Fe availability to phytoplankton will be affected. Indirectly, ocean acidification could also influence the distributions of other bioactive metals that interact with Fe nutrition. For example, it is predicted that Cu availability will increase as Fe availability decreases (Millero et al. 2009; Hoffmann et al. 2012). Depending on relative changers in Cu and Fe availability, this increase in Cu availability could be beneficial or detrimental. If the Fe concentration is very low and the Cu concentration is not toxic, higher Cu might enhance the activity of the high-affinity Fe transporters. However, if Fe concentration is very low but Cu concentration is at toxic levels, phytoplankton growth could be impaired. The availability of these metals may also affect the phytoplankton production of the climate-influencing osmolyte, dimethylsulfoniopropionate (DMSP) (Sunda et al. 2002). Recent evidence has also shown ocean acidification slows down N2 fixation in the diazotroph Trichodesmium when they are Fe-limited (Shi et al. 2012). How all of these changes will alter phytoplankton Fe requirements and their elemental compositions as well as important cellular processes such as photosynthesis, respiration and the production of organic compounds such as DMSP is unclear. Most importantly, will all phytoplankton be affected in the same way and how will these changes influence the community composition throughout the different aquatic environments? Resolving the answers to these questions is critical given the significant roles phytoplankton play in aquatic food webs and global biogeochemical cycles.
Notes
- 1.
Wherever possible the currently accepted names for species are used. The name used in the paper cited is also indicated. For details of names see chapter “Systematics, Taxonomy and Species Names: Do They Matter?” of this book (Borowitzka 2016).
References
Allen MD, del Campo JA, Kropat J, Merchant SS (2007) FEA1, FEA2, and FRE1, encoding two homologous secreted proteins and a candidate ferrireductase, are expressed coordinately with FOX1 and FTR1 in iron-deficient Chlamydomonas reinhardtii. Eukaryot Cell 6:1841–1852
Allen A, LaRoche J, Maheswari U, Lommer M, Schauer N, Lopez P, Finazzi G, Fernie A, Bowler C (2008) Whole-cell response of the pennate diatom Phaeodactylum tricornutum to iron starvation. Proc Natl Acad Sci 105:10438–10443
Allnutt FCT, Bonner WD Jr (1987a) Characterization of iron uptake from ferrioxamine B by Chlorella vulgaris. Plant Physiol 85:746–750
Allnutt FCT, Bonner WD (1987b) Evaluation of reductive release as a mechanism for iron uptake from ferrioxamine B by Chlorella vulgaris. Plant Physiol 85:751–756
Anbar AD, Knoll AH (2002) Proterozoic ocean chemistry and evolution: a bioinorganic bridge? Science 297:1137–1142
Andersen RA (1992) Diversity of eukaryotic algae. Biodivers Conserv 1:267–292
Anderson MA, Morel FMM (1982) The influence of aqueous iron chemistry on the uptake of iron by the coastal diatom Thalassiosira weissflogii. Limnol Oceanogr 27:789–813
Annett AL, Lapi S, Ruth TJ, Maldonado MT (2008) The effects of Cu and Fe availability on the growth and Cu: C ratios of marine diatoms. Limnol Oceanogr 53:2451–2461
Armbrust E (2009) The life of diatoms in the world’s oceans. Nature 459:185–192
Askwith CC, de Silva D, Kaplan J (1996) Molecular biology of iron acquisition in Saccharomyces cerevisiae. Mol Microbiol 20:27–34
Assmy P, Smetacek V, Montresor M, Klaas C, Henjes J, Strass VH, Arrieta JM, Bathmann U, Berg GM, Breitbarth E, Cisewski B, Friedrichs L, Fuchs N, Herndl GJ, Jansen S, Kragefsky S, Latasa M, Peeken I, Rottgers R, Scharek R, Schuller SE, Steigenberger S, Webb A, Wolf-Gladrow D (2013) Thick-shelled, grazer-protected diatoms decouple ocean carbon and silicon cycles in the iron-limited Antarctic Circumpolar Current. Proc Natl Acad Sci 110:20633–20638
Baines SB, Twining BS, Vogt S, Balch WM, Fisher NS, Nelson DM (2011) Elemental composition of equatorial Pacific diatoms exposed to additions of silicic acid and iron. Deep Sea Res II 58:512–523
Banse K (1991) Iron availability, nitrate uptake, and exportable new production in the Subarctic Pacific. J Geophys Res Oceans 96:741–748
Barbeau K, Rue EL, Bruland KW, Butler A (2001) Photochemical cycling of iron in the surface ocean mediated by microbial iron(III)-binding ligands. Nature 413:409–413
Barbeau K, Rue EL, Trick CG, Bruland KT, Butler A (2003) Photochemical reactivity of siderophores produced by marine heterotrophic bacteria and cyanobacteria based on characteristic Fe(III) binding groups. Limnol Oceanogr 48:1069–1078
Behrenfeld MJ, Milligan AJ (2013) Photophysiological expressions of iron stress in phytoplankton. Ann Rev Mar Sci 5:217–246
Behrenfeld MJ, Worthington K, Sherrell RM, Chavez FP, Strutton P, McPhaden M, Shea DM (2006) Controls on tropical Pacific Ocean productivity revealed through nutrient stress diagnostics. Nature 442:1025–1028
Behrenfeld MJ, Westberry TK, Boss ES, O’Malley RT, Siegel DA, Wiggert JD, Franz BA, McClain CR, Feldman GC, Doney SC, Moore JK, Dall’Olmo G, Milligan AJ, Lima I, Mahowald N (2009) Satellite-detected fluorescence reveals global physiology of ocean phytoplankton. Biogeosciences 6:779–794
Beiderbeck H, Taraz K, Budzikiewicz H, Walsby AE (2000) Anachelin, the siderophore of the cyanobacterium Anabaena cylindrica CCAP 1403/2A. Zeitschr Naturf C 55:681–687
Beja O, Aravind L, Koonin EV, Suzuki MT, Hadd A, Nguyen LP, Jovanovich S, Gates CM, Feldman RA, Spudich JL, Spudich EN, DeLong EF (2000) Bacterial rhodopsin: evidence for a new type of phototrophy in the sea. Science 289:1902–1906
Berman-Frank I, Quigg A, Finkel ZV, Irwin AJ, Haramaty L (2007) Nitrogen-fixation strategies and Fe requirements in cyanobacteria. Limnol Oceanogr 52:2260–2269
Bertrand EM, Saito MA, Rose JM, Riesselman CR, Lohan MC, Noble AE, Lee PA, DiTullio GR (2007) Vitamin B12 and iron colimitation of phytoplankton growth in the Ross Sea. Limnol Oceanogr 52:1079–1093
Bibby TS, Nield J, Barber J (2001) Iron deficiency induces the formation of an antenna ring around trimeric photosystem I in cyanobacteria. Nature 412:743–745
Blaby-Haas CE, Merchant SS (2012) The ins and outs of algal metal transport. Biochim Biophys Acta-Mol Cell Res 1823:1531–1552
Blain S, Sedwick PN, Griffiths FB, Queguiner B, Bucciarelli E, Fiala M, Pondaven P, Treguer P (2002) Quantification of algal iron requirements in the Subantarctic Southern Ocean (Indian sector). Deep-Sea Res II 49:3255–3273
Blain S, Queguiner B, Armand L, Belviso S, Bombled B, Bopp L, Bowie A, Brunet C, Brussaard C, Carlotti F, Christaki U, Corbiere A, Durand I, Ebersbach F, Fuda JL, Garcia N, Gerringa L, Griffiths B, Guigue C, Guillerm C, Jacquet S, Jeandel C, Laan P, Lefevre D, Lo Monaco C, Malits A, Mosseri J, Obernosterer I, Park YH, Picheral M, Pondaven P, Remenyi T, Sandroni V, Sarthou G, Savoye N, Scouarnec L, Souhaut M, Thuiller D, Timmermans K, Trull T, Uitz J, van Beek P, Veldhuis M, Vincent D, Viollier E, Vong L, Wagener T (2007) Effect of natural iron fertilization on carbon sequestration in the Southern Ocean. Nature 446:1070–U1071
Bleuel C, Grosse C, Taudte N, Scherer J, Wesenberg D, Krauss GJ, Nies DH, Grass G (2005) TolC is involved in enterobactin efflux across the outer membrane of Escherichia coli. J Bacteriol 187:6701–6707
Boekema EJ, Hifney A, Yakushevska AE, Piotrowski M, Keegstra W, Berry S, Michel KP, Pistorius EK, Kruip J (2001) A giant chlorophyll-protein complex induced by iron deficiency in cyanobacteria. Nature 412:745–748
Borowitzka MA (2016) Systematics, taxonomy and species names: do they matter? In: Borowitzka MA, Beardall J, Raven JA (eds) The physiology of microalgae. Springer, Dordrecht, pp 655–681
Boukhalfa H, Crumbliss AL (2002) Chemical aspects of siderophore mediated iron transport. Biometals 15:325–339
Boyd PW, Jickells T, Law CS, Blain S, Boyle EA, Buesseler KO, Coale KH, Cullen JJ, de Baar HJW, Follows M, Harvey M, Lancelot C, Levasseur M, Owens NPJ, Pollard R, Rivkin RB, Sarmiento J, Schoemann V, Smetacek V, Takeda S, Tsuda A, Turner S, Watson AJ (2007) Mesoscale iron enrichment experiments 1993–2005: synthesis and future directions. Science 315:612–617
Boyle E (1998) Pumping iron makes thinner diatoms. Nature 393:733–734
Boyle E, Edmond J, Sholkovitz E (1976) The mechanism of iron removal in estuaries. Geochim Cosmochim Acta 41:1313–1324
Breitbarth E, Bellerby RJ, Neill CC, Ardelan MV, Meyerhöfer M, Zöllner E, Croot PL, Riebesell U (2010) Ocean acidification affects iron speciation during a coastal seawater mesocosm experiment. Biogeosciences 7:1065–1073
Bruland KW, Orians KJ, Cowen JP (1994) Reactive trace metals in the stratified central North Pacific. Geochim Cosmochim Acta 58:3171–3182
Bruland KW, Lohan MC, Aguilar-Islas AM, Smith GJ, Sohst B, Baptista A (2008) Factors influencing the chemistry of the near-field Columbia River plume: nitrate, silicic acid, dissolved Fe, and dissolved Mn. J Geophys Res Oceans 113:C00B02. DOI:10.1029/2007JC004702
Bruland KW, Middag R, Lohan MC (2014) Controls of trace metals in seawater. In: Holland HD, Tureklan KK (eds) Treatise on geochemistry, vol 8, 2nd edn. Elsevier, Oxford, pp 19–51
Brzezinski MA, Baines SB, Balch WM, Beucher C, Chai F, Dugdale RC, Krause JW, Landry MR, Marchi A, Measures C, Nelson DM, Parker A, Poulton A, Selph KE, Strutton P, Taylor AG, Twining BS (2011) Co-limitation of diatoms by iron and silicic acid in the equatorial Pacific. Deep-Sea Res II:493–511
Bucciarelli E, Pondaven P, Sarthou G (2010) Effects of an iron-light co-limitation on the elemental composition (Si, C, N) of the marine diatoms Thalassiosira oceanica and Ditylum brightwellii. Biogeosciences 7:657–669
Castruita M, Casero D, Karpowicz SJ, Kropat J, Vieler A, Hsieh SI, Yan W, Cokus S, Loo JA, Benning C (2011) Systems biology approach in Chlamydomonas reveals connections between copper nutrition and multiple metabolic steps. Plant Cell Online 23:1273–1292
Cellier MFM, Bergevin I, Boyer E, Richer E (2001) Polyphyletic origins of bacterial Nramp transporters. Trends Genet 17:365–370
Chase Z, Strutton PG, Hales B (2007) Iron links river runoff and shelf width to phytoplankton biomass along the U.S. West Coast. Geophys Res Lett 34:L04607. DOI:10.1029/2006GL028069
Chauhan D, Folea IM, Jolley CC, Kouril R, Lubner CE, Lin S, Kolber D, Wolfe-Simon F, Golbeck JH, Boekema EJ (2011) A novel photosynthetic strategy for adaptation to low-iron aquatic environments. Biochemistry 50:686–692
Chavez FP, Buck KR, Coale KH, Martin JH, Ditullio GR, Welschmeyer NA, Jacobson AC, Barber RT (1991) Growth-rates, grazing, sinking, and iron limitation of Equatorial Pacific phytoplankton. Limnol Oceanogr 36:1816–1833
Chimento DP, Kadner RJ, Wiener MC (2005) Comparative structural analysis of TonB-dependent outer membrane transporters: implications for the transport cycle. Proteins 59:240–251
Coale KH, Fitzwater SE, Gordon RM, Johnson KS, Barber RT (1996) Control of community growth and export production by upwelled iron in the equatorial Pacific Ocean. Nature 379:621–624
Coale KH, Wang X, Tanner SJ, Johnson KS (2003) Phytoplankton growth and biological response to iron and zinc addition in the Ross Sea and Antarctic Circumpolar Current along 170 W. Deep Sea Res II 50:635–653
Coale KH, Johnson KS, Chavez FP, Buesseler KO, Barber RT, Brzezinski MA, Cochlan WP, Millero FJ, Falkowski PG, Bauer JE, Wanninkhof RH, Kudela RM, Altabet MA, Hales BE, Takahashi T, Landry MR, Bidigare RR, Wang XJ, Chase Z, Strutton PG, Friederich GE, Gorbunov MY, Lance VP, Hilting AK, Hiscock MR, Demarest M, Hiscock WT, Sullivan KF, Tanner SJ, Gordon RM, Hunter CN, Elrod VA, Fitzwater SE, Jones JL, Tozzi S, Koblizek M, Roberts AE, Herndon J, Brewster J, Ladizinsky N, Smith G, Cooper D, Timothy D, Brown SL, Selph KE, Sheridan CC, Twining BS, Johnson ZI (2004) Southern ocean iron enrichment experiment: carbon cycling in high- and low-Si waters. Science 304:408–414
Cochlan WP, Bronk DA, Coale KH (2002) Trace metals and nitrogenous nutrition of Antarctic phytoplankton: experimental observations in the Ross Sea. Deep-Sea Res II 49:3365–3390
Conway TM, John SG (2014) Quantification of dissolved iron sources to the North Atlantic Ocean. Nature 511:212–215
Crawford DW, Lipsen MS, Purdie DA, Lohan MC, Statham PJ, Whitney FA, Putland JN, Johnson WK, Sutherland N, Peterson TD, Harrison PJ, Wong CS (2003) Influence of zinc and iron enrichments on phytoplankton growth in the northeastern subarctic Pacific. Limnol Oceanogr 48:1583–1600
Cullen JJ (1991) Hypotheses to explain high-nutrient conditions in the open sea. Limnol Oceanogr 36:1578–1599
Cullen JT, Chase Z, Coale KH, Fitzwater SE, Sherrell RM (2003) Effect of iron limitation on the cadmium to phosphorus ratio of natural phytoplankton assemblages from the Southern Ocean. Limnol Oceanogr 48:1079–1087
Dang TC, Fujii M, Rose AL, Bligh M, Waite TD (2012) Characteristics of the freshwater cyanobacterium Microcystis aeruginosa grown in iron-limited continuous culture. Appl Environ Microbiol 78:1574–1583
de Baar HJW, Boyd PW, Coale KH, Landry MR, Tsuda A, Assmy P, Bakker DCE, Bozec Y, Barber RT, Brzezinski MA, Buesseler KO, Boye M, Croot PL, Gervais F, Gorbunov MY, Harrison PJ, Hiscock WT, Laan P, Lancelot C, Law CS, Levasseur M, Marchetti A, Millero FJ, Nishioka J, Nojiri Y, van Oijen T, Riebesell U, Rijkenberg MJA, Saito H, Takeda S, Timmermans KR, Veldhuis MJW, Waite AM, Wong CS (2005) Synthesis of iron fertilization experiments: from the iron age in the age of enlightenment. J Geophys Res 110:C09S16. doi:10.1029/2004JC002601
Desai DK, Desai FD, LaRoche J (2012) Factors influencing the diversity of iron uptake systems in aquatic microorganisms. Front Microbiol 3:362. doi:10.3389/fmicb.2012.00362
Doney SC, Fabry VJ, Feely RA, Kleypas JA (2009) Ocean acidification: the other CO2 problem. Annu Rev Mar Sci 1:169–192
Doney SC, Ruckelshaus M, Emmett Duffy J, Barry JP, Chan F, English CA, Galindo HM, Grebmeier JM, Hollowed AB, Knowlton N, Polovina J, Rabalais NN, Sydeman WJ, Talley LD (2012) Climate change impacts on marine ecosystems. Annu Rev Mar Sci 4:11–37
Doucette GJ, Harrison PJ (1991) Aspects of iron and nitrogen nutrition in the red tide dinoflagellate Gymnodinium sanguineum. Mar Biol 110:165–173
Doucette GJ, Erdner DL, Peleato ML, Hartman JJ, Anderson DM (1996) Quantitative analysis of iron-stress related proteins in Thalassiosira weissflogii: measurement of flavodoxin and ferredoxin using HPLC. Mar Ecol Prog Ser 130:269–276
Droop MR (1970) Vitamin B12 and marine ecology V. Continuous culture as an approach to nutritional kinetics. Helgol Wiss Meeresun 20:629–636
Dugdale RC, Wilkerson FP (1991) Low specific nitrate uptake rate – a common feature of high-nutrient, low-chlorophyll marine ecosystems. Limnol Oceanogr 36:1678–1688
Dugdale RC, Wilkerson FP (1998) Silicate regulation of new production in the equatorial Pacific upwelling. Nature 391:270–273
Eckhardt U, Buckhout TJ (1998) Iron assimilation in Chlamydomonas reinhardtii involves ferric reduction and is similar to Strategy I higher plants. J Exp Bot 49:1219–1226
Ehrenreich IM, Waterbury JB, Webb EA (2005) Distribution and diversity of natural product genes in marine and freshwater cyanobacterial cultures and genomes. Appl Environ Microbiol 71:7401–7413
Eide D, Broderius M, Fett J, Guerinot ML (1996) A novel iron-regulated metal transporter from plants identified by functional expression in yeast. Proc Natl Acad Sci 93:5624–5628
Eldridge ML, Trick CG, Alm MB, DiTullio GR, Rue EL, Bruland KW, Hutchins DA, Wilhelm SW (2004) Phytoplankton community response to a manipulation of bioavailable iron in HNLC waters of the subtropical Pacific Ocean. Aquat Microb Ecol 35:79–91
Falkowski PG, Owens TG, Ley AC, Mauzerall DC (1981) Effects of growth irradiance levels on the ratio of reaction centers in 2 species of marine phytoplankton. Plant Physiol 68:969–973
Falkowski PG, Katz ME, Knoll AH, Quigg A, Raven JA, Schofield O, Taylor FJR (2004) The evolution of modern eukaryotic phytoplankton. Science 305:354–360
Faraldo-Gomez JD, Sansom MSP (2003) Acquisition of siderophores in gram-negative bacteria. Nat Rev Mol Cell Biol 4:105–116
Faraldo-Gomez JD, Smith GR, Sansom MSP (2003) Molecular dynamics simulations of the bacterial outer membrane protein FhuA: a comparative study of the ferrichrome-free and bound states. Biophys J 85:1406–1420
Finkel ZV, Quigg AS, Raven JA, Reinfelder JR, Schofield OE, Falkowski PG (2006) Irradiance and the elemental stoichiometry of marine phytoplankton. Limnol Oceanogr 51:2690–2701
Firme GF, Rue EL, Weeks DA, Bruland KW, Hutchins DA (2003) Spatial and temporal variability in phytoplankton iron limitation along the California coast and consequences for Si, N, and C biogeochemistry. Global Biogeochem Cycles 17:1016
Fitzwater SE, Coale KH, Gordon RM, Johnson KS, Ondrusek ME (1996) Iron deficiency and phytoplankton growth in the equatorial Pacific. Deep-Sea Res II 43:995–1015
Fuhrman JA, Schwalbach MS, Stingl U (2008) Proteorhodopsins: an array of physiological roles? Nat Rev Microbiol 6:488–494
Fujii M, Dang TC, Rose AL, Omura T, Waite TD (2011) Effect of light on iron uptake by the freshwater cyanobacterium Microcystis aeruginosa. Environ Sci Technol 45:1391–1398
Gademann K, Portmann C (2008) Secondary metabolites from cyanobacteria: complex structures and powerful bioactivities. Curr Org Chem 12:326–341
Gaither LA, Eide DJ (2001) Eukaryotic zinc transporters and their regulation. Biometals 14:251–270
Garcia NS, Sedwick PN, DiTullio GR (2009) Influence of irradiance and iron on the growth of colonial Phaeocystis antarctica: implications for seasonal bloom dynamics in the Ross Sea, Antarctica. Aquat Microb Ecol 57:203–220
Geider RJ (1987) Light and temperature dependence of the carbon to chlorophyll a ratio in microalgae and cyanobacteria: implications for physiology and growth of phytoplankton. New Phytol 106:1–34
Geider RJ, LaRoche J (1994) The role of iron in phytoplankton photosynthesis, and the potential for iron limitation of primary productivity in the sea. Photosynth Res 39:275–301
Geider RJ, La Roche J, Greene R, Olaizola M (1993) Response of the photosynthetic apparatus of Phaeodactylum tricornutum (Bacillariophyceae) to nitrate, phosphate or iron limitation. J Phycol 29:755–766
Gledhill M, Buck KN (2012) The organic complexation of iron in the marine environment: a review. Front Microbiol 3:69. doi:10.3389/fmicb.2012.00069
Gledhill M, van den Berg CMG (1994) Determination of complexation of iron(III) with natural organic complexing ligands in seawater using cathodic stripping voltammetry. Mar Chem 47:41–54
Glover HF (1977) Effects of iron deficiency on the physiology and biochemistry of Isochrysis galbana (Chrysophyceae) and Phaedactylum tricornutum (Bacillariophyceae). J Phycol 13:208–212
Greene RM, Geider RJ, Falkowski PG (1991) Effect of iron limitation on photosynthesis in a marine diatom. Limnol Oceanogr 36:1772–1782
Greene RM, Geider RJ, Kolber Z, Falkowski PG (1992) Iron-induced changes in light harvesting and photochemical energy conversion processes in eukaryotic marine algae. Plant Physiol 100:565–575
Groussman RD, Parker MS, Armbrust EV (2015) Diversity and evolutionary history of iron metabolism genes in diatoms. PLoS ONE 10:e0129081
Guo J, Annett AL, Taylor RL, Lapi S, Ruth TJ, Maldonado MT (2010) Copper uptake kinetics of coastal and oceanic diatoms. J Phycol 46:1218–1228
Guo J, Lapi S, Ruth TJ, Maldonado MT (2012) The effects of iron and copper availability on the copper stoichiometry of marine phytoplankton. J Phycol 48:312–325
Guo J, Green BR, Maldonado MT (2015) Sequence analysis and gene expression of potential components of copper transport and homeostasis in Thalassiosira pseudonana. Protist 166:58–77
Hamm CE, Merkel R, Springer O, Jurkojc P, Maier C, Prechtel K, Smetacek V (2003) Architecture and material properties of diatom shells provide effective mechanical protection. Nature 421:841–843
Harris JE (1992) Weathering of rock, corrosion of stone and rusting of iron. Meccanica 27:233–250
Harrison PJ, Berges JA (2005) Marine culture media. In: Andersen RA (ed) Algal culturing techniques. Elsevier Academic Press, San Diego, pp 21–34
Harrison GI, Morel FMM (1986) Response of the marine diatom Thalassiosira weissflogii to iron stress. Limnol Oceanogr 31:989–997
Harrison PJ, Whitney FA, Tsuda A, Saito H, Tadokoro K (2004) Nutrient and plankton dynamics in the NE and NW gyres of the subarctic Pacific Ocean. J Oceanogr 60:93–117
Hart TJ (1934) On the phytoplankton of the Southwest Atlantic and the Bellingshausen Sea. Discov Rep 8:1–268
Hartnett A, Bottger LH, Matzanke BF, Carrano CJ (2012a) Iron transport and storage in the coccolithophore: Emiliania huxleyi. Metallomics 4:1160–1166
Hartnett A, Bottger LH, Matzanke BF, Carrano CJ (2012b) A multidisciplinary study of iron transport and storage in the marine green alga Tetraselmis suecica. J Inorg Biochem 116:188–194
Havens SM, Hassler CS, North RL, Guildford SJ, Silsbe G, Wilhelm SW, Twiss MR (2012) Iron plays a role in nitrate drawdown by phytoplankton in Lake Erie surface waters as observed in lake-wide assessments. Can J Fish Aquat Sci 69:369–381
Herbik A, Bolling C, Buckhout TJ (2002a) The involvement of a multicopper oxidase in iron uptake by the green algae Chlamydomonas reinhardtii. Plant Physiol 130:2039–2048
Herbik A, Haebel S, Buckhout TJ (2002b) Is a ferroxidase involved in the high-affinity iron uptake in Chlamydomonas reinhardtii. Plant Soil 241:1–9
Hill KL, Merchant S (1995) Coordinate expression of coproporphyrinogen oxidase and cytochrome c6 in the green alga Chlamydomonas reinhardtii in response to changes in copper availability. EMBO J 14:857–865
Ho T-Y, Quigg A, Finkel ZV, Milligan AJ, Wyman K, Falkowski PG, Morel FMM (2003) The elemental composition of some marine phytoplankton. J Phycol 39:1145–1159
Hoffmann LJ, Breitbarth E, Boyd PW, Hunter KA (2012) Influence of ocean warming and acidification on trace metal biogeochemistry. Mar Ecol Prog Ser 470:191–205
Hopkinson B, Barbeau K (2008) Interactive influences of iron and light limitation on phytoplankton at subsurface chlorophyll maxima in the eastern North Pacific. Limnol Oceanogr 53:1303–1318
Hopkinson BM, Barbeau KA (2012) Iron transporters in marine prokaryotic genomes and metagenomes. Environ Microbiol 14:114–128
Hopkinson BM, Morel FMM (2009) The role of siderophores in iron acquisition by photosynthetic marine microorganisms. Biometals 22:659–669
Hopkinson BM, Xu Y, Shi D, McGinn PJ, Morel FMM (2010) The effect of CO2 on the photosynthetic physiology of phytoplankton in the Gulf of Alaska. Limnol Oceanogr 55:2011–2024
Hoppe CJM, Hassler CS, Payne CD, Tortell PD, Rost B, Trimborn S (2013) Iron limitation modulates ocean acidification effects on Southern Ocean phytoplankton communities. PLoS ONE 8:e79890
Hudson RJM, Morel FMM (1989) Distinguishing between extra- and intracellular iron in marine phytoplankton. Limnol Oceanogr 34:111–1120
Hudson RJM, Morel FMM (1990) Iron transport in marine phytoplankton – kinetics of cellular and medium coordination reactions. Limnol Oceanogr 35:1002–1020
Hudson RJM, Morel FMM (1993) Trace-metal transport by marine microorganisms – implications of metal coordination kinetics. Deep-Sea Res I 40:129–150
Hutchins DA (1995) Iron and the marine phytoplankton community. Phycol Res 11:1–49
Hutchins DA, Bruland KW (1998) Iron-limited diatom growth and Si:N uptake ratios in a coastal upwelling regime. Nature 393:561–564
Hutchins DA, DiTullio GR, Zhang Y, Bruland KW (1998) An iron limitation mosaic in the California upwelling regime. Limnol Oceanogr 43:1037–1054
Hutchins DA, Franck VM, Brzezinski MA, Bruland KW (1999a) Inducing phytoplankton iron limitation in iron-replete coastal waters with a strong chelating ligand. Limnol Oceanogr 44:1009–1018
Hutchins DA, Witter AE, Butler A, Luther GW (1999b) Competition among marine phytoplankton for different chelated iron species. Nature 400:858–861
Hutchins DA, Hare CE, Weaver RS, Zhang Y, Firme GF, DiTullio GR, Alm MB, Riseman SF, Maucher JM, Geesey ME (2002) Phytoplankton iron limitation in the Humboldt Current and Peru Upwelling. Limnol Oceanogr 47:997–1011
Hyenstrand P, Rydin E, Gunnerhed M (2000) Response of pelagic cyanobacteria to iron additions – enclosure experiments from Lake Erken. J Plankton Res 22:1113–1126
Ito Y, Butler A (2005) Structure of synechobactins, new siderophores of the marine cyanobacterium Synechococcus sp. PCC 7002. Limnol Oceanogr 50:1918
Johnson KS, Gordon RM, Coale KH (1997) What controls dissolved iron concentrations in the world ocean? Mar Chem 57:137–161
Johnson KS, Chavez FP, Friederich GE (1999) Continental-shelf sediment as a primary source of iron for coastal phytoplankton. Nature 398:697–700
Jones GJ, Palenik BP, Morel FMM (1987) Trace metal reduction by phytoplankton: the role of plasmalemma redox enzymes. J Phycol 23:237–244
Katoh H, Hagino N, Grossman AR, Ogawa T (2001a) Genes essential to iron transport in the cyanobacterium Synechocystis sp strain PCC 6803. J Bacteriol 183:2779–2784
Katoh H, Hagino N, Ogawa T (2001b) Iron-binding activity of FutA1 subunit of an ABC-type iron transporter in the cyanobacterium Synechocystis sp strain PCC 6803. Plant Cell Physiol 42:823–827
Klunder MB, Bauch D, Laan P, de Baar HJW, van Heuven S, Ober S (2012) Dissolved iron in the Arctic shelf seas and surface waters of the central Arctic Ocean: impact of Arctic river water and ice-melt. J Geophys Res 117:C01027
Koch F, Marcoval MA, Panzeca C, Bruland KW, Sanudo-Wilhelmy SA, Gobler CJ (2011) The effect of vitamin B12 on phytoplankton growth and community structure in the Gulf of Alaska. Limnol Oceanogr 56:1023–1034
Kranzler C, Lis H, Shaked Y, Keren N (2011) The role of reduction in iron uptake processes in a unicellular, planktonic cyanobacterium. Environ Microbiol 13:2990–2999
Kranzler C, Rudolf M, Keren N, Schleiff E (2013) Iron in cyanobacteria. Adv Bot Res 65:57–105
Kranzler C, Lis H, Finkel OM, Schmetterer G, Shaked Y, Keren N (2014) Coordinated transporter activity shapes high-affinity iron acquisition in cyanobacteria. ISME J 8:409–417
Krewulak KD, Vogel HJ (2011) TonB or not TonB: is that the question? Biochem Cell Biol 89:87–97
Kudo I, Harrison PJ (1997) Effect of iron nutrition on the marine cyanobacterium Synechococcus grown on different N sources and irradiances. J Phycol 33:232–240
Kudo I, Noiri Y, Nishioka J, Taira Y, Kiyosawa H, Tsuda A (2006) Phytoplankton community response to Fe and temperature gradients in the NE (SERIES) and NW (SEEDS) subarctic Pacific Ocean. Deep Sea Res II 53:2201–2213
Kuma K et al (1992) Photo-reduction of Fe (III) by dissolved organic substances and existence of Fe (II) in seawater during spring blooms. Mar Chem 37:15–27
Kustka AB, Sanudo-Wilhemy SA, Carpenter EJ, Capone D, Burns J, Sunda WG (2003) Iron requirements for dinitrogen- and ammonium-supported growth in cultures of Trichodesmium (IMS 101): comparison with nitrogen fixation rates and iron:carbon ratios of field populations. Limnol Oceanogr 48:1869–1884
Kustka AB, Allen AE, Morel FMM (2007) Sequence analysis and transcriptional regulation of iron acquisition genes in two marine diatoms. J Phycol 43:715–729
La Fontaine S, Quinn JM, Nakamoto SS, Page MD, Gohre V, Moseley JL, Kropat J, Merchant S (2002) Copper-dependent iron assimilation pathway in the model photosynthetic eukaryote Chlamydomonas reinhardtii. Eukaryot Cell 1:736–757
La Roche J, Geider RJ, Graziano LM, Murray H, Lewis K (1993) Induction of specific proteins in eukaryotic algae grown under iron-deficient, phosphorus-deficient, or nitrogen-deficient conditions. J Phycol 29:767–777
La Roche J, Boyd P, McKay R, Geider R (1996) Flavodoxin as an in situ marker for iron stress in phytoplankton. Nature 382:802–805
Lane ES, Semeniuk DM, Strzepek RF, Cullen JT, Maldonado MT (2009) Effects of iron limitation on intracellular cadmium of cultured phytoplankton: implications for surface dissolved cadmium to phosphate ratios. Mar Chem 115:155–162
Lelong A, Bucciarelli E, Hagaret H, Soudant P (2013) Iron and copper limitations differently affect growth rates and photosynthetic and physiological parameters of the marine diatom Pseudo-nitzschia delicatissima. Limnol Oceanogr 58:613–623
Lin W, Chai J, Love J, Fu D (2010) Selective electrodiffusion of zinc ions in a Zrt-, Irt-like protein, ZIPB. J Biol Chem 285:39013–39020
Lin H, Rauschenberg S, Hexel CR, Shaw TJ, Twining BS (2011) Free-drifting icebergs as sources of iron to the Weddell Sea. Deep Sea Res II 58:1392–1406
Lis H, Shaked Y (2009) Probing the bioavailability of organically bound iron: a case study in the Synechococcus-rich waters of the Gulf of Aqaba. Aquat Microb Ecol 56:241–253
Lis H, Shaked Y, Kranzler C, Keren N, Morel FMM (2015) Iron bioavailability to phytoplankton: an empirical approach. ISME J 9:1003–1013
Liu XW, Theil EC (2005) Ferritins: dynamic management of biological iron and oxygen chemistry. Acc Chem Res 38:167–175
Lohan MC, Bruland KW (2008) Elevated Fe(II) and dissolved Fe in hypoxic shelf waters off Oregon and Washington: an enhanced source of iron to coastal upwelling regimes. Environ Sci Technol 42:6462–6468
Lommer M, Roy A, Schilhabel M, Schreiber S, Rosenstiel P, LaRoche J (2010) Recent transfer of an iron-regulated gene from the plastid to the nuclear genome in an oceanic diatom adapted to chronic iron limitation. BMC Genomics 11:718
Lommer M, Specht M, Roy A-S, Kraemer L, Andreson R, Gutowska M, Wolf J, Bergner S, Schilhabel M, Klostermeier U, Beiko R, Rosenstiel P, Hippler M, LaRoche J (2012) Genome and low-iron response of an oceanic diatom adapted to chronic iron limitation. Genome Biol 13:R66
Mahowald NM, Baker AR, Bergametti G, Brooks N, Duce RA, Jickells TD, Kubilay N, Prospero JM, Tegen I (2005) Atmospheric global dust cycle and iron inputs to the ocean. Glob Biogeochem Cycles 19:GB4025
Maldonado MT, Price NM (1996) Influence of N substrate on Fe requirements of marine centric diatoms. Mar Ecol Prog Ser 141:161–172
Maldonado MT, Price NM (1999) Utilization of iron bound to strong organic ligands by plankton communities in the subarctic Pacific Ocean. Deep Sea Res II 46:2447–2473
Maldonado MT, Price NM (2000) Nitrate regulation of Fe reduction and transport by Fe-limited Thalassiosira oceanica. Limnol Oceanogr 45:814–826
Maldonado MT, Price NM (2001) Reduction and transport of organically bound iron by Thalassiosira oceanica (Bacillariophyceae). J Phycol 37:298–309
Maldonado MT, Boyd PW, Harrison PJ, Price NM (1999) Co-limitation of phytoplankton growth by light and Fe during winter in the NE subarctic Pacific Ocean. Deep Sea Res II 46:2475–2485
Maldonado MT, Boyd PW, LaRoche J, Strzepek R, Waite A, Bowie AR, Croot PL, Frew RD, Price NM (2001) Iron uptake and physiological response of phytoplankton during a mesoscale Southern Ocean iron enrichment. Limnol Oceanogr 46:1802–1808
Maldonado MT, Strzepek RF, Sander S, Boyd PW (2005) Acquisition of iron bound to strong organic complexes, with different Fe binding groups and photochemical reactivities, by plankton communities in Fe-limited subantarctic waters. Glob Biogeochem Cycles 19:GB4S23
Maldonado MT, Allen AE, Chong JS, Lin K, Leus D, Karpenko N, Harris SL (2006) Copper-dependent iron transport in coastal and oceanic diatoms. Limnol Oceanogr 51:1729–1743
Maranger R, Bird DF, Price NM (1998) Iron acquisition by photosynthetic marine phytoplankton from ingested bacteria. Nature 396:248–251
Marchetti A, Cassar N (2009) Diatom elemental and morphological changes in response to iron limitation: a brief review with potential paleoceanographic applications. Geobiology 7:419–431
Marchetti A, Harrison PJ (2007) Coupled changes in the cell morphology and the elemental (C, N and Si) composition of the pennate diatom Pseudo-nitzschia due to iron deficiency. Limnol Oceanogr 52:2270–2284
Marchetti A, Maldonado MT, Lane ES, Harrison PJ (2006) Iron requirements of the pennate diatom Pseudo-nitzschia: comparison of oceanic (HNLC) and coastal species. Limnol Oceanogr 51:2092–2101
Marchetti A, Lundholm N, Kotaki Y, Hubbard KA, Harrison PJ, Armbrust EV (2008) Identification and assessment of domoic acid production in oceanic Pseudo-nitzschia (Bacillariophyceae) from iron-limited waters in the NE Subarctic Pacific. J Phycol 44:650–661
Marchetti A, Parker MS, Moccia LP, Lin EO, Arrieta AL, Ribalet F, Murphy MEP, Maldonado MT, Armbrust EV (2009) Ferritin is used for iron storage in bloom-forming marine pennate diatoms. Nature 457:467–470
Marchetti A, Varela DE, Lance VP, Johnson Z, Palmucci M, Giordano M, Armbrust EV (2010) Iron and silicic acid effects on phytoplankton productivity, diversity, and chemical composition in the central equatorial Pacific Ocean. Limnol Oceanogr 55:11–29
Marchetti A, Schruth DM, Durkin CA, Parker MS, Kodner RB, Berthiaume CT, Morales R, Allen AE, Armbrust EV (2012) Comparative metatranscriptomics identifies molecular bases for the physiological responses of phytoplankton to varying iron availability. Proc Natl Acad Sci 109:E317–E325
Marchetti A, Catlett D, Hopkinson BM, Ellis K, Cassar N (2015) Marine diatom proteorhodopsins and their potential role in coping with low iron availability. ISME J. doi:10.1038/ismej.2015.74
Martin JH (1990) Glacial-interglacial CO2 change: the iron hypothesis. Paleoceanography 5:1–13
Martin JH, Fitzwater S (1988) Iron deficiency limits phytoplankton growth in the north-east Pacific subarctic. Nature 331:341–343
Martin JH, Gordon RM, Fitzwater S, Broenkow WW (1989) Vertex – phytoplankton iron studies in the Gulf of Alaska. Deep Sea Res I 36:649–680
Martinez JS, Carter-Franklin JN, Mann EL, Martin JD, Haygood MG, Butler A (2003) Structure and membrane affinity of a suite of amphiphilic siderophores produced by a marine bacterium. Proc Natl Acad Sci 100:3754–3759
Matz CJ, Magnus RS, Walker CN, Fink MB, Treble RG, Weger HG (2006) Differences between two green algae in biological availability of iron bound to strong chelators. Can J Bot 84:400–411
Mawji E, Gledhill M, Milton JA, Tarran GA, Ussher S, Thompson A, Wolff GA, Worsfold PJ, Achterberg EP (2008) Hydroxamate siderophores: occurrence and importance in the Atlantic Ocean. Environ Sci Technol 42:8675–8680
McAllister CD, Parsons TR, Strickland JDH (1960) Primary productivity and fertility at Station P in the north-east Pacific Ocean. ICES J Mar Sci 25:240–259
Merchant SS, Allen MD, Kropat J, Moseley JL, Long JC, Tottey S, Terauchi AM (2006) Between a rock and a hard place: trace element nutrition in Chlamydomonas. Biochim Biophys Acta, Mol Cell Res 1763:578–594
Middlemiss JK, Anderson AM, Stratilo CW, Weger HG (2001) Oxygen consumption associated with ferric reductase activity and iron uptake by iron-limited cells of Chlorella kessleri (Chlorophyceae). J Phycol 37:393–399
Miethke M, Marahiel MA (2007) Siderophore-based iron acquisition and pathogen control. Microbiol Mol Biol Rev 71:413–451
Miller CB, Frost BW, Wheeler PA, Landry MR, Welschmeyer N, Powell TM (1991) Ecological dynamics in the subarctic Pacific, a possibly iron-limited ecosystem. Limnol Oceanogr 36:1600–1615
Millero FJ, Woosley R, DiTrolio BJW (2009) Effect of ocean acidification on the speciation of metals in seawater. Oceanography 22:72–85
Milligan AJ, Harrison PJ (2000) Effects of non-steady-state iron limitation on nitrogen assimilatory enzymes in the marine diatom Thalassiosira weissflogii (Bacillariophyceae). J Phycol 36:78–86
Mills MM, Ridame C, Davey M, La Roche J, Geider RJ (2004) Iron and phosphorus co-limit nitrogen fixation in the eastern tropical North Atlantic. Nature 429:292–294
Mirus O, Strauss S, Nicolaisen K, von Haeseler A, Schleiff E (2009) TonB-dependent transporters and their occurrence in cyanobacteria. BMC Biol 7:68
Mitchell BG, Brody EA, Holm-Hansen O, McClain C, Bishop J (1991) Light limitation of phytoplankton biomass and macronutrient utilization in the Southern Ocean. Limnol Oceanogr 36:1662–1677
Mock T, Samanta MP, Iverson V, Berthiaume C, Robison M, Holtermann K, Durkin C, BonDurant SS, Richmond K, Rodesch M, Kallas T, Huttlin EL, Cerrina F, Sussmann MR, Armbrust EV (2008) Whole-genome expression profiling of the marine diatom Thalassiosira pseudonana identifies genes involved in silicon bioprocesses. Proc Natl Acad Sci U S A 105:1579–1584
Monod J (1942) Recherches sur la croissance des Cultures Bactériennes. Hermann, Paris
Monzyk B, Crumbliss AL (1982) Kinetics and mechanism of the stepwise dissociation of iron(III) from ferrioxamine B in aqueous acid. J Am Chem Soc 104:4921–4929
Moore JK, Braucher O (2008) Sedimentary and mineral dust sources of dissolved iron to the world ocean. Biogeosciences 5:631–656
Moore JK, Doney SC, Glover DM, Fung IY (2002) Iron cycling and nutrient-limitation patterns in surface waters of the World Ocean. Deep Sea Res II 49:463–507
Moore CM, Mills MM, Achterberg EP, Geider RJ, LaRoche J, Lucas MI, McDonagh EL, Pan X, Poulton AJ, Rijkenberg MJA, Suggett DJ, Ussher SJ, Woodward EMS (2009) Large-scale distribution of Atlantic nitrogen fixation controlled by iron availability. Nat Geosci 2:867–871
Morel FMM (1987) Kinetics of nutrient uptake and growth in phytoplankton. J Phycol 23:137–150
Morel FMM, Hudson R, Price N (1991) Limitation of productivity by trace metals in the sea. Limnol Oceanogr 36:1742–1755
Morrissey J, Bowler C (2012) Iron utilization in marine cyanobacteria and eukaryotic algae. Front Microbiol 3:43. doi:10.3389/fmicb.2012.00043
Morton SD, Lee TH (1974) Algal blooms. Possible effects of iron. Environ Sci Technol 8:673–674
Moseley JL, Allinger T, Herzog S, Hoerth P, Wehinger E, Merchant S, Hippler M (2002) Adaptation to Fe-deficiency requires remodeling of the photosynthetic apparatus. EMBO J 21:6709–6720
Muggli DL, Harrison PJ (1996a) EDTA suppresses the growth of oceanic phytoplankton from the Northeast subarctic Pacific. J Exp Mar Biol Ecol 205:221–227
Muggli DL, Harrison PJ (1996b) Effects of nitrogen source on the physiology and metal nutrition of Emiliania huxleyi grown under different iron and light conditions. Mar Ecol Prog Ser 130:255–267
Muggli DL, Lecourt M, Harrison PJ (1996) Effects of iron and nitrogen source on the sinking rate, physiology and metal composition of an oceanic diatom from the subarctic Pacific. Mar Ecol Prog Ser 132:215–227
Neilands JB (1995) Siderophores: structure and function of microbial iron transport compounds. J Biol Chem 270:26723–26726
Nelson DM, Treguer P, Brzezinski MA, Leynaert A, Queguiner B (1995) Production and dissolution of biogenic silica in the ocean: revised global estimates, comparison with regional data and relationships to biogenic sedimentation. Glob Biogeochem Cycles 9:359–372
Nevo Y, Nelson N (2006) The NRAMP family of metal-ion transporters. Biochim Biophys Acta Mol Cell Res 1763:609–620
Nicolaisen K, Moslavac S, Samborski A, Valdebenito M, Hantke K, Maldener I, Muro-Pastor AM, Flores E, Schleiff E (2008) Alr0397 is an outer membrane transporter for the siderophore schizokinen in Anabaena sp. strain PCC 7120. J Bacteriol 190:7500–7507
Nicolaisen K, Hahn A, Valdebenito M, Moslavac S, Samborski A, Maldener I, Wilken C, Valladares A, Flores E, Hantke K (2010) The interplay between siderophore secretion and coupled iron and copper transport in the heterocyst-forming cyanobacterium Anabaena sp. PCC 7120. Biochim Biophys Acta Biomembr 1798:2131–2140
Nishioka J, Takeda S (2000) Change in the concentrations of iron in different size fractions during growth of the oceanic diatom Chaetoceros sp.: importance of small colloidal iron. Mar Biol 137:231–238
Noinaj N, Guillier M, Barnard TJ, Buchanan SK (2010) TonB-dependent transporters: regulation, structure, and function. I Ann Rev Microbiol 64:43–60
North RL, Guildford SJ, Smith REH, Havens SM, Twiss MR (2007) Evidence for phosphorus, nitrogen, and iron colimitation of phytoplankton communities in Lake Erie. Limnol Oceanogr 52:315–328
Nouet C, Motte P, Hanikenne M (2011) Chloroplastic and mitochondrial metal homeostasis. Trends Plant Sci 16:395–404
Nuester J, Vogt S, Twining BS (2012) Localization of iron within centric diatoms of the genus Thalassiosira. J Phycol 48:626–634
Palenik B, Ren Q, Dupont CL, Myers GS, Heidelberg JF, Badger JH, Madupu R, Nelson WC, Brinkac LM, Dodson RJ (2006) Genome sequence of Synechococcus CC9311: insights into adaptation to a coastal environment. Proc Natl Acad Sci 103:13555–13559
Palenik B, Grimwood J, Aerts A, Rouze P, Salamov A, Putnam N, Dupont C, Jorgensen R, Derelle E, Rombauts S, Zhou K, Otillar R, Merchant SS, Podell S, Gaasterland T, Napoli C, Gendler K, Manuell A, Tai V, Vallon O, Piganeau G, Sv J, Heijde M, Jabbari K, Bowler C, Lohr M, Robbens S, Werner G, Dubchak I, Pazour GJ, Ren Q, Paulsen I, Delwiche C, Schmutz J, Rokhsar D, Van de Peer Y, Moreau H, Grigoriev IV (2007) The tiny eukaryote Ostreococcus provides genomic insights into the paradox of plankton speciation. Proc Natl Acad Sci 104:7705–7710
Pankowski A, McMinn A (2009) Iron availability regulates growth, photosynthesis, and production of ferredoxin and flavodoxin in Antarctic sea ice diatoms. Aquat Biol 4:273–288
Passy SI (2010) A distinct latitudinal gradient of diatom diversity is linked to resource supply. Ecology 91:36–41
Paz Y, Katz A, Pick U (2007a) Multicopper ferroxidase involved in iron binding to transferrins in Dunaliella salina plasma membranes. J Biol Chem 282:8658–8666
Paz Y, Shimoni E, Weiss M, Pick U (2007b) Effects of iron deficiency on iron binding and internalization into acidic vacuoles in Dunaliella salina. Plant Physiol 144:1407–1415
Peers G, Price N (2006) Copper-containing plastocyanin used for electron transport by an oceanic diatom. Nature 441:341–344
Peers G, Quesnel SA, Price NM (2005) Copper requirements for iron acquisition and growth of coastal and oceanic diatoms. Limnol Oceanogr 50:1149–1158
Pollard RT, Salter I, Sanders RJ, Lucas MI, Moore CM, Mills RA, Statham PJ, Allen JT, Baker AR, Bakker DCE, Charette MA, Fielding S, Fones GR, French M, Hickman AE, Holland RJ, Hughes JA, Jickells TD, Lampitt RS, Morris PJ, Nedelec FH, Nielsdottir M, Planquette H, Popova EE, Poulton AJ, Read JF, Seeyave S, Smith T, Stinchcombe M, Taylor S, Thomalla S, Venables HJ, Williamson R, Zubkov MV (2009) Southern Ocean deep-water carbon export enhanced by natural iron fertilization. Nature 457:577–580
Pollingher U, Kaplan B, Berman T (1995) The impact of iron and chelators on Lake Kinneret phytoplankton. J Plankton Res 17:1977–1992
Pondaven P, Gallinari M, Chollet S, Bucciarelli E, Sarthou G, Schultes S, Jean F (2007) Grazing-induced changes in cell wall silicification in a marine diatom. Protist 158:21–28
Price NM (2005) The elemental stoichiometry and composition of an iron-limited diatom. Limnol Oceanogr 50:1159–1171
Price NM, Harrison GI, Hering JG, Hudson RJ, Nirel PMV, Palenik B, Morel FMM (1988/89) Preparation and chemistry of the artificial algal culture medium Aquil. Biol Oceanogr 6:443–461
Price NM, Andersen LF, Morel FMM (1991) Iron and nitrogen nutrition of Equatorial Pacific plankton. Deep Sea Res II 38:1361–1378
Price NM, Ahner BA, Morel FMM (1994) The Equatorial Pacific Ocean – grazer-controlled phytoplankton populations in an iron-limited ecosystem. Limnol Oceanogr 39:520–534
Quigg A, Finkel ZV, Irwin AJ, Rosenthal Y, Ho TY, Reinfelder JR, Schofield O, Morel FMM, Falkowski PG (2003) The evolutionary inheritance of elemental stoichiometry in marine phytoplankton. Nature 425:291–294
Quigg A, Irwin AJ, Finkel ZV (2011) Evolutionary inheritance of elemental stoichiometry in phytoplankton. Proc Roy Soc B 278:526–534
Raiswell R, Benning L, Tranter M, Tulaczyk S (2008) Bioavailable iron in the Southern Ocean: the significance of the iceberg conveyor belt. Geochem Trans 9:7
Raven JA (1988) The iron and molybdenum use efficiencies of plant growth with different energy, carbon and nitrogen sources. New Phytol 109:279–287
Raven JA (1990) Predictions of Mn and Fe use efficiencies of phototrophic growth as a function of light availability for growth and of C assimilation pathway. New Phytol 116:1–18
Raven JA, Waite A (2004) The evolution of silicification in diatoms: inescapable sinking and sinking as escape. New Phytol 162:45–65
Rose AL (2012) The influence of extracellular superoxide on iron redox chemistry and bioavailability to aquatic microorganisms. Front Microbiol 3:124. doi:10.3389/fmicb.2012.00124
Rue EL, Bruland KW (1995) Complexation of iron (III) by natural organic ligands in the Central North Pacific as determined by a new competitive ligand equilibration/adsorptive cathodic stripping voltammetric method. Mar Chem 50:117–138
Rueter JG, Ades DR (1987) The role of iron nutrition in photosynthesis and nitrogen assimilation in Scenedesmus quadricauda (Chlorophyceae). J Phycol 23:452–457
Ryan-Keogh TJ, Macey AI, Cockshutt AM, Moore CM, Bibby TS (2012) The cyanobacterial chlorophyll-binding-protein isiA acts to increase the in vivo effective absorption cross-section of PSI under iron limitation. J Phycol 48:145–154
Saito MA, Rocap G, Moffett JW (2005) Production of cobalt binding ligands in a Synechococcus feature at the Costa Rica upwelling dome. Limnol Oceanogr 50:279–290
Saito MA, Goepfert TJ, Ritt JT (2008) Some thoughts on the concept of colimitation: three definitions and the importance of bioavailability. Limnol Oceanogr 53:276–290
Saito MA, Noble AE, Tagliabue A, Goepfert TJ, Lamborg CH, Jenkins WJ (2013) Slow-spreading submarine ridges in the South Atlantic as a significant oceanic iron source. Nat Geosci 6:775–779
Sandmann G, Reck H, Kessler E, Boger P (1983) Distribution of plastocyanin and soluble plastidic cytochrome c in various classes of algae. Arch Microbiol 134:23–27
Sandmann G, Peleato ML, Fillat MF, Lazaro MC, Gomez-Moreno C (1990) Consequences of the iron-dependent formation of ferredoxin and flavodoxin on photosynthesis and nitrogen fixation on Anabaena strains. Photosynth Res 26:119–125
Schauer K, Rodionov DA, de Reuse H (2008) New substrates for TonB-dependent transport: do we only see the “tip of the iceberg”? Trends Biochem Sci 33:330–338
Schenck RC, Tessier A, Campbell PGC (1988) The effect of pH on iron and manganese uptake by a green alga. Limnol Oceanogr 33:538–550
Sedwick PN, Garcia N, Riseman S, Marsay C, DiTullio G (2007) Evidence for high iron requirements of colonial Phaeocystis antarctica at low irradiance. Biogeochemistry 83:83–97
Shaked Y, Lis H (2012) Disassembling iron availability to phytoplankton. Front Microbiol 3:123. doi:10.3389/fmicb.2012.00123
Shaked Y, Kustka A, Morel F (2005) A general kinetic model for iron acquisition by eukaryotic phytoplankton. Limnol Oceanogr 50:872–882
Sherman DM, Sherman LA (1983) Effect of iron deficiency and iron restoration on ultrastructure of Anacystis nidulans. J Bacteriol 156:393–401
Shi D, Xu Y, Hopkinson BM, Morel FMM (2010) Effect of ocean acidification on iron availability to marine phytoplankton. Science 327:676–679
Shi D, Kranz SA, Kim J-M, Morel FMM (2012) Ocean acidification slows nitrogen fixation and growth in the dominant diazotroph Trichodesmium under low-iron conditions. Proc Natl Acad Sci 109:E3094–E3100
Silva-Stenico ME, Silva CS, Lorenzi AS, Shishido TK, Etchegaray A, Lira SP, Moraes LA, Fiore MF (2011) Non-ribosomal peptides produced by Brazilian cyanobacterial isolates with antimicrobial activity. Microb Res 166:161–175
Simpson FB, Neilands JB (1976) Siderochromes in cyanophyceae: isolation and characterization of schizokinen from Anabaena sp. J Phycol 12:44–48
Smayda TJ (1970) The suspension and sinking of phytoplankton in the sea. Oceanogr Mar Biol Ann Rev 8:353–414
Smetacek V (1985) Role of sinking in diatom life history cycles: ecological, evolutionary and geological significance. Mar Biol 84:239–251
Smetacek V (1999) Diatoms and the ocean carbon cycle. Protist 150:25–32
Smetacek V, Assmy P, Henjes J (2004) The role of grazing in structuring Southern Ocean pelagic ecosystems and biogeochemical cycles. Antarct Sci 16:541–558
Smetacek V, Klaas C, Strass VH, Assmy P, Montresor M, Cisewski B, Savoye N, Webb A, d’Ovidio F, Arrieta JM, Bathmann U, Bellerby R, Berg GM, Croot P, Gonzalez S, Henjes J, Herndl GJ, Hoffmann LJ, Leach H, Losch M, Mills MM, Neill C, Peeken I, Rottgers R, Sachs O, Sauter E, Schmidt MM, Schwarz J, Terbruggen A, Wolf-Gladrow D (2012) Deep carbon export from a Southern Ocean iron-fertilized diatom bloom. Nature 487:313–319
Smith K, Robison B, Helly J, Kaufmann R, Ruhl H, Shaw T, Twining B, Vernat M (2007) Free-drifting icebergs: hot spots of chemical and biological enrichment in the Weddell Sea. Science 317:478–483
Sohm JA, Webb EA, Capone DG (2011) Emerging patterns of marine nitrogen fixation. Nat Rev Microbiol 9:499–508
Soria-Dengg S, Horstmann U (1995) Ferroxiamines B and E as iron source for the marine diatom Phaeodactylum tricornutum. Mar Ecol Prog Ser 127:269–277
Stearman R, Yuan DS, Yamaguchi-Iwai Y, Klausner RD, Dancis A (1996) A permease-oxidase complex involved in high-affinity iron uptake in yeast. Science 271:1552–1557
Sterner RW, Smutka TM, McKay RM, Xiaoming Q, Brown ET, Sherrel R (2004) Phosphorus and trace metal limitation of algae and bacteria in Lake Superior. Limnol Oceanogr 49:495–507
Stevanovic M, Hahn A, Nicolaisen K, Mirus O, Schleiff E (2012) The components of the putative iron transport system in the cyanobacterium Anabaena sp. PCC 7120. Environ Microbiol 14:1655–1670
Straus N (2004) Iron deprivation: physiology and gene regulation. In: Bryant D (ed) The molecular biology of cyanobacteria, Book 1. Springer, Dordrecht, pp 731–750
Strong A, Chisholm S, Miller C, Cullen J (2009) Ocean fertilization: time to move on. Nature 461:347–348
Strzepek R, Harrison P (2004) Photosynthetic architecture differs in coastal and oceanic diatoms. Nature 431:689–692
Strzepek RF, Price NM (2000) Influence of irradiance and temperature on the iron content of the marine diatom Thalassiosira weissflogii (Bacillariophyceae). Mar Ecol Prog Ser 206:107–117
Strzepek RF, Maldonado MT, Hunter KAFDR, Boyd PW (2011) Adaptive strategies by Southern Ocean phytoplankton to lessen iron limitation: uptake of organically complexed iron and reduced cellular iron requirements. Limnol Oceanogr 56:1983–2002
Strzepek RF, Hunter KA, Frew RD, Harrison PJ, Boyd PW (2012) Iron-light interactions in Southern Ocean phytoplankton. Limnol Oceanogr 57:1182–1200
Sugie K, Yoshimura T (2013) Effects of pCO2 and iron on the elemental composition and cell geometry of the marine diatom Pseudo-nitzschia pseudodelicatissima (Bacillariophyceae). J Phycol 49:475–488
Sugie K, Endo H, Suzuki K, Nishioka J, Kiyosawa H, Yoshimura T (2013) Synergistic effects of pCO2 and iron availability on nutrient consumption ratio of the Bering Sea phytoplankton community. Biogeosciences 10:6309–6321
Sunda WG, Huntsman SA (1992) Feedback interactions between zinc and phytoplankton in seawater. Limnol Oceanogr 37:25–40
Sunda W, Huntsman SA (1995) Iron uptake and growth limitation in oceanic and coastal phytoplankton. Mar Chemy 50:189–206
Sunda WG, Huntsman SA (1997) Interrelated influence of iron, light and cell size on marine phytoplankton growth. Nature 390:389–392
Sunda W, Huntsman S (2003) Effect of pH, light, and temperature on Fe-EDTA chelation and Fe hydrolysis in seawater. Mar Chem 84:35–47
Sunda WG, Huntsman SA (2011) Interactive effects of light and temperature on iron limitation in a marine diatom: implications for marine productivity and carbon cycling. Limnol Oceanogr 56:1475–1488
Sunda WG, Huntsman SA (2015) High iron requirement for growth, photosynthesis, and low-light acclimation in the coastal cyanobacterium Synechococcus bacillaris. Front Microbiol 6:561
Sunda WG, Swift D, Huntsman S (1991) Low iron requirement for growth in oceanic phytoplankton. Nature 351:55–57
Sunda W, Kieber DJ, Kiene RP, Huntsman S (2002) An antioxidant function for DMSP and DMS in marine algae. Nature 418:317–320
Sunda WG, Price NM, Morel FMM (2005) Trace metal ion buffers and their use in culture studies. In: Anderson RA (ed) Algal culturing techniques. Elsevier Academic Press, London, pp 35–64
Tagliabue A, Bopp L, Dutay J-C, Bowie AR, Chever F, Jean-Baptiste P, Bucciarelli E, Lannuzel D, Remenyi T, Sarthou G, Aumont O, Gehlen M, Jeandel C (2010) Hydrothermal contribution to the oceanic dissolved iron inventory. Nat Geosci 3:252–256
Takeda S (1998) Influence of iron availability on nutrient consumption ratio of diatoms in oceanic waters. Nature 393:774–777
Tang D, Morel FMM (2006) Distinguishing between cellular and Fe-oxide-associated trace elements in phytoplankton. Mar Chem 98:18–30
Taylor RL, Semeniuk DM, Payne CD, Zhou J, Tremblay J-E, Cullen JT, Maldonado MT (2013) Colimitation by light, nitrate, and iron in the Beaufort Sea in late summer. J Geophys Res 118:3260–3277
Tilzer M, Elbrachter M, Gieskes WWC, Beese B (1986) Light-temperature interactions in the control of photosynthesis in Antarctic phytoplankton. Polar Biol 5:105–112
Timmermans KR, Gerringa LJA, de Baar HJW, van der Wagt B, Veldhuis MJW, de Jong JTM, Croot PL, Boye M (2001) Growth rates of large and small Southern Ocean diatoms in relation to availability of iron in natural seawater. Limnol Oceanogr 46:260–266
Timmermans KR, van der Wagt B, de Baar HJW (2004) Growth rates, half-saturation constants, and silicate, nitrate, and phosphate depletion in relation to iron availability of four large, open-ocean diatoms from the Southern Ocean. Limnol Oceanogr 49:2141–2151
Timmermans KR, van der Wagt B, Veldhuis MJW, Maatman A, de Baar HJW (2005) Physiological responses of three species of marine pico-phytoplankton to ammonium, phosphate, iron and light limitation. J Sea Res 53:109–120
Trick CG, Bill BD, Cochlan WP, Wells ML, Trainer VL, Pickell LD (2010) Iron enrichment stimulates toxic diatom production in high-nitrate, low-chlorophyll areas. Proc Natl Acad Sci 107:5887–5892
Twining BS, Baines SB (2013) The trace metal composition of marine phytoplankton. Ann Rev Mar Sci 5:191–215
Twining BS, Baines SB, Fisher NS, Maserr J, Vogt S, Jacobsen C, Tovar-Sanchez A, Sañudo-Wilhelmy SA (2003) Quantifying trace elements in individual aquatic protist cells with a synchrotron X-ray fluorescence microprobe. Anal Chem 75:3806–3816
Twining BS, Baines SB, Fisher NS, Landry MR (2004) Cellular iron contents of plankton during the Southern Ocean Iron Experiment (SOFeX). Deep Sea Res I 51:1827–1850
Twining BS, Baines SB, Bozard JB, Vogt S, Walker EA, Nelson DM (2011) Metal quotas of plankton in the equatorial Pacific Ocean. Deep Sea Res II 58:325–341
Twiss MR, Auclair J-C, Charlton MN (2000) An investigation into iron-stimulated phytoplankton productivity in epipelagic Lake Erie during thermal stratification using trace metal clean techniques. Can J Fish Aquat Sci 57:86–95
Van Ho A, Ward DM, Kaplan J (2002) Transition metal transport in yeast. Annu Rev Microbiol 56:237–261
van Oijen T, van Leeuwe MA, Gieskes WWC, de Baar HJW (2004) Effects of iron limitation on photosynthesis and carbohydrate metabolism in the Antarctic diatom Chaetoceros brevis (Bacillariophyceae). Eur J Phycol 39:161–171
Varsano T, Wolf SG, Pick U (2006) A chlorophyll a/b-binding protein homolog that is induced by iron deficiency is associated with enlarged photosystem I units in the eucaryotic alga Dunaliella salina. J Biol Chem 281:10305–10315
Waite TD, Morel FMM (1984) Photoreductive dissolution of colloidal iron oxides in natural waters. Environ Sci Technol 18:860–868
Weger HG (1999) Ferric and cupric reductase activities in the green alga Chlamydomonas reinhardtii: experiments using iron-limited chemostats. Planta 207:377–384
Weger HG, Middlemiss JK, Petterson CD (2002) Ferric chelate reductase activity as affected by the iron- limited growth rate in four species of unicellular green algae (Chlorophyta). J Phycol 38:513–519
Wells ML, Goldberg ED (1994) The distribution of colloids in the North Atlantic and Southern Oceans. Limnol Oceanogr 39:286–302
Wells ML, Price NM, Bruland KW (1995) Iron chemistry in seawater and its relationship to phytoplankton – a workshop report. Mar Chem 48:157–182
Weng H-X, Sun X-W, Qin Y-C, Chen J-F (2007) Effect of irradiance on Fe and P uptake by Cryptomonas sp. Geochimica 4:008
Wetz MS, Hales B, Chase Z, Wheeler PA, Whitney MM (2006) Riverine input of macronutrients, iron, and organic matter to the coastal ocean off Oregon, USA, during the winter. Limnol Oceanogr 51:2221–2231
Whitney L, Lins J, Hughes M, Wells M, Chappell P, Jenkins B (2011) Characterization of putative iron responsive genes as species-specific indicators of iron stress in Thalassiosiroid diatoms. Front Microbiol 2:234. doi:10.3389/fmicb.2011.00234
Wilhelm SW (1995) Ecology of iron-limited cyanobacteria: a review of physiological responses and implications for aquatic systems. Aquat Microb Ecol 9:295–303
Wolfe-Simon F, Grzebyk D, Schofield O, Falkowski PG (2005) The role and evolution of superoxide dismutases in algae. J Phycol 41:453–465
Zehr JP, Kudela RM (2009) Photosynthesis in the open ocean. Science 326:945–946
Zhao H, Eide D (1996a) The yeast ZRT1 gene encodes the zinc transporter protein of a high-affinity uptake system induced by zinc limitation. Proc Natl Acad Sci 93:2454–2458
Zhao H, Eide D (1996b) The ZRT2 gene encodes the low affinity zinc transporter in Saccharomyces cerevisiae. J Biol Chem 271:23203–23210
Acknowledgements
We thank W.G. Sunda, P.J. Harrison and J.A. Raven for their helpful comments and insights on the manuscript. Supported by NSF-OCE 1334935 (A.M.).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Marchetti, A., Maldonado, M.T. (2016). Iron. In: Borowitzka, M., Beardall, J., Raven, J. (eds) The Physiology of Microalgae. Developments in Applied Phycology, vol 6. Springer, Cham. https://doi.org/10.1007/978-3-319-24945-2_11
Download citation
DOI: https://doi.org/10.1007/978-3-319-24945-2_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-24943-8
Online ISBN: 978-3-319-24945-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)